Embed Size (px)
Citation preview

RUDISTS OF TIBET AND THE TARIM BASIN, CHINA: SIGNIFICANCETO REQUIENIIDAE PHYLOGENY
ROBERT W. SCOTT,1 XIAQIAO WAN,2 JINGENG SHA,3 AND SHI-XUAN WEN3
1Department of Geosciences, University of Tulsa, Tulsa, Oklahoma 74104, ,[email protected].; 2China University of Geosciences,Beijing, China; and 3LPS, Nanjing Institute of Geology and Paleontology, Chinese Academy of Sciences, Nanjing
ABSTRACT—Rudists are a principal biotic component of Cretaceous carbonates in Tibet and in the Western TarimBasin. Barremian to Maastrichtian carbonate units are widespread on the northern margin of the Indian Plate andin Tethyan tectonic slices that were welded onto Eurasia in successive stages during the Late Cretaceous andPaleogene. In far northwestern Tibet, Barremian-Cenomanian endemic rudists and cosmopolitan orbitolinidforaminifera occupied isolated carbonate platforms in the eastern Tethys. Rudists, corals, and stromatoporoidscomposed bioherms up to 10 m thick and several kilometers in lateral extent. A unique endemic requieniid rudist,Rutonia, is compared to morphologically similar but older, less derived genera. Associated specimens in thisassemblage are indeterminate requieniid valves, monopleurids, and two genera with three radiolitid species that arere-described and taxonomic positions re-evaluated. In southern Tibet, mainly endemic Campanian-Maastrichtianradiolitid rudists and cosmopolitan larger benthic foraminifera contributed to carbonate shelves on the northernIndian Plate near the Cretaceous equator. In the Western Tarim Basin Cenomanian strata yield Tethyan rudistspecies.
Coiling morphometric analysis using the three-dimensional morphology Raup diagram shows that Requieniidaevalves in contact with the substrate are convergent with the basic gastropod shell. More derived strongly coiled,younger requieniids were adapted to encrusting or semi-infaunal habits. Stratigraphic analysis confirms thatRequieniidae diversity crises coincided with Cretaceous oceanic anoxic events
Two end members of valve geometry each appear to be primitive and derived characters respectively and separatethe family Requieniidae into two clades that are here recognized as two new subfamilies. The end members aredefined by the coiling geometry, whether the spire is close to the plane of commissure or it is translated along thecoiling axis and by myophore structures. The older matheroniform clade has a low spirogyrate LV that is translatedslightly from the commissure along the coiling axis; this group is composed of Matheronia (and its subgenusMonnieria), Hypelasma, Lovetchenia, Rutonia, and Kugleria. Genera in the younger clade have a tall trochospiral LVthat is translated along the coiling axis and consists of Requienia, Toucasia, Pseudotoucasia, Apricardia, Bayleoidea,and Bayleia. Claditics support these relationships.
INTRODUCTION
CRETACEOUS CARBONATE shelves and platforms in the TethysRealm were populated by marine benthic communities
dominated by unconventional rudist bivalves, scleractinia, andvarious types of larger benthic foraminifera (Masse et al.,1995). Rudist distribution and species composition are ratherwell known in the western part of the Tethys Realm of NorthAfrica, southern Europe, Turkey, and the Middle East(Steuber and Loser, 2000). However, rudists in the easternpart of southern Asia are poorly known. In Tibet/Xizang,Cretaceous carbonates are quite thick and widespread andrudists are a principal biotic component. The Chinesegeosciences community has described many rudist species(Qian, 1993; Gou and Shi, 1998; Wen, 1999; Wen et al., 2000).
Douville (1916) first reported Upper Cretaceous rudistsfrom southern Tibet collected during a 1903 British expedition(Fig. 1). The Campanian assemblage consisted of Bournoniahaydeni Douville (1916) and Bournonia tibetica Douville(1916) associated with Biradiolites. The Maastrichtian rudistis Plagioptychus tibeticus Douville (1916) with the largerbenthic foraminifers Orbitoides, Lepidorbitoides and Ompha-locyclus. These assemblages are part of the Campanian-Maastrichtian Zongshan Formation (Willems et al., 1996).
Albian and Cenomanian rudists and orbitolinid foraminif-era are significant components of thick carbonate units in farnorthwestern Tibet in the Ngari, Xizang area (Fig. 1) (Wan etal., 2003). The Albian Lanshang Formation is from 1,000 to3,000 m thick (Fig. 2) and is dated by the foraminifers
Mesorbitolina texana (Roemer, 1849) (senior synonym ofOrbitolina kurdica Henson, 1948, according to Simmons et al.,2000) and Mesorbitolina pervia (Douglas, 1960) together withPraeradiolites hedini Douville (1917), Sphaerulites [formerlyPraeradiolites] biconvexus (Yang et al., 1982), and Praeradio-lites ngariensis Yang et al. (1982). The Cenomanian Gamba-cunkou Formation yields Mesorbitolina aperta (Erman, 1854),Orbitolina concava (Lamarck, 1816), and Conicorbitolinaconica (D’Archiac, 1837) with the rudists Praeradiolites hediniand Rutonia bangonghuensis Yang et al. (1982), and mono-pleurids. Six additional rudist species were created by Gou andShi (1998): list them.
The objectives of this contribution are to demonstrate thegeological importance of Tibetan rudists and to synthesizetheir distribution, biostratigraphy, paleoecology, and phylo-genetic considerations. A modern description of the locallyknown requieniid genus, Rutonia, prompts a morphologicalanalysis and a cladistic hypothesis of the rudist FamilyRequieniidae that suggests the Family Requieniidae be dividedinto two subfamilies. This paper follows phylogenetic researchof Upper Jurassic and Lower Cretaceous species by Masse(1994, 2002) and Gourrat et al. (2003).
MATERIAL
Large collections of rudists and other megafossils as well assamples for microfossil thin sections were collected during the1980 expedition. These materials were distributed between theUniversity Museum at China University of GeosciencesBeijing and the Paleontological Institute in Nanjing. Speci-
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:48:52 1 Cust # 09-137
J. Paleont., 84(3), 2010, pp. 000–000
Copyright ’ 2010, The Paleontological Society
0022-3360/10/0000-0000$03.00
0

mens collected by later expeditions are at the ChengduInstitute of Technology.
Dr. Zetong Nie (personal commun., 2007), who, with Dr.Xiaqiao Wan, participated in the geological expedition toTibet, reviewed for us the rudist occurrences. The rudist-bearing Langshan Formation carbonate is up to 1,000 m thick
and extends for many kilometers across the Ngari region inwestern Tibet (Fig. 1, site 1). In places rudist-coral biohermsup to 10 m thick and several tens of meters wide are encased inmarl. Rudists and corals dominate the bioherms and algae areaccessory biota. At the bioherm margins orbitolinids arecommon and are intermixed with rudist talus debris. Overlyingthe buildups at the base of the marl, orbitolinids andintraclasts of the rudist-coral bioherm are abundant anddiminish up section (Yang et al., 1982, fig. 2).
BIOGEOGRAPHY OF TIBETAN RUDISTS
The Tibetan Plateau today is a complex tectonic provincecomposed of several terranes or blocks sandwiched betweenthe Indian Plate to the south and the Eurasian Plate on thenorth (Fig. 3) (Huang and Chen, 1987; Qian, 1993; Li andXiao, 1995; Wang et al., 1997; Gou and Shi, 1998; Wen, 1999;Wen et al., 2000; Sha et al., 2002; Wan et al., 2003; Sha et al.,2004). North of the Indian Plate in northwestern Tibet theLhasa Block is bounded by the Yarlung-Zangbo and theBangong-Nu River suture zones (Fig. 1). Albian-Cenomaniancarbonates with rudists (Yang et al., 1982) are located at thenorthwest end of the Lhasa Block in the Ngari district (Fig. 1,site 1). During the middle Cretaceous this tectonic slice waspositioned between approximately 10u and 20u north latitudes(Fig. 3; Golonka, 2002). A second tectonic slice, the Qiang-tang block, lies north of the Lhasa Block and south of theEurasian Plate. This latter block hosts Cretaceous carbonatesin the Karakorum and Kunlun mountains and the TarimBasin along its southern margin (Wen, 1999). The Qiangtang
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:19 2 Cust # 09-137
FIGURE 2—Generalized stratigraphic classification of rudist-bearingformations in Tibet (modified from Wen, 1999; Wang et al. 2002).
FIGURE 1—Major sedimentary terranes of the Tibetan-Qinghai-Xizang Plateau in present-day position. Legend: 1(marine shelf facies, 2) deep seafacies, and 3) continental facies (from Wen, 1999). The key rudist localities are 1) Ngari region (Yang et al., 1982): type area of Radiolites bangonghuensis,Praeradiolites biconvexus, and Praeradiolites ngariensis; and 2) Zongshan area with villages of Kampa Dzong and Tuna, (N28u to 28u209 by W88u309 to89u159) (Douville, 1916): type area of Bournonia haydeni and Bournonia tibetica, and Plagioptychus tibeticus.
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010
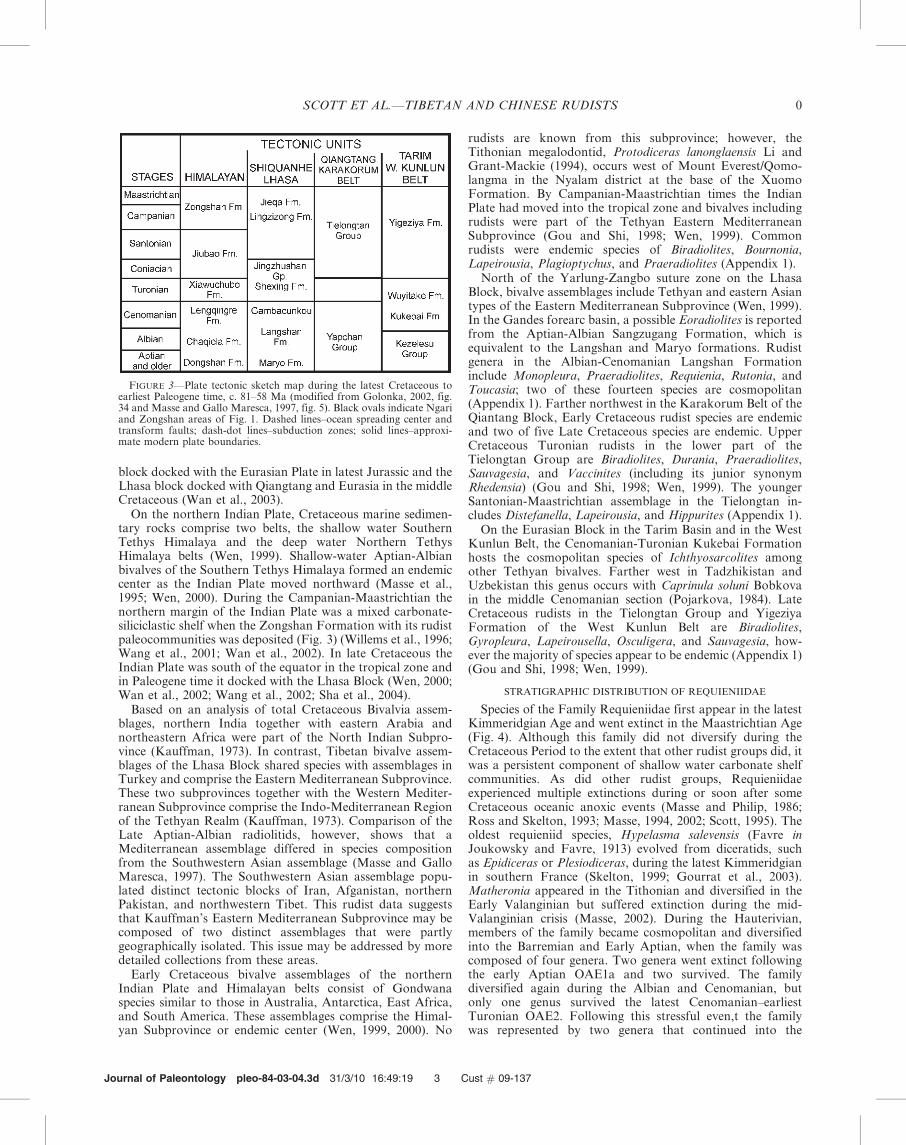
block docked with the Eurasian Plate in latest Jurassic and theLhasa block docked with Qiangtang and Eurasia in the middleCretaceous (Wan et al., 2003).
On the northern Indian Plate, Cretaceous marine sedimen-tary rocks comprise two belts, the shallow water SouthernTethys Himalaya and the deep water Northern TethysHimalaya belts (Wen, 1999). Shallow-water Aptian-Albianbivalves of the Southern Tethys Himalaya formed an endemiccenter as the Indian Plate moved northward (Masse et al.,1995; Wen, 2000). During the Campanian-Maastrichtian thenorthern margin of the Indian Plate was a mixed carbonate-siliciclastic shelf when the Zongshan Formation with its rudistpaleocommunities was deposited (Fig. 3) (Willems et al., 1996;Wang et al., 2001; Wan et al., 2002). In late Cretaceous theIndian Plate was south of the equator in the tropical zone andin Paleogene time it docked with the Lhasa Block (Wen, 2000;Wan et al., 2002; Wang et al., 2002; Sha et al., 2004).
Based on an analysis of total Cretaceous Bivalvia assem-blages, northern India together with eastern Arabia andnortheastern Africa were part of the North Indian Subpro-vince (Kauffman, 1973). In contrast, Tibetan bivalve assem-blages of the Lhasa Block shared species with assemblages inTurkey and comprise the Eastern Mediterranean Subprovince.These two subprovinces together with the Western Mediter-ranean Subprovince comprise the Indo-Mediterranean Regionof the Tethyan Realm (Kauffman, 1973). Comparison of theLate Aptian-Albian radiolitids, however, shows that aMediterranean assemblage differed in species compositionfrom the Southwestern Asian assemblage (Masse and GalloMaresca, 1997). The Southwestern Asian assemblage popu-lated distinct tectonic blocks of Iran, Afganistan, northernPakistan, and northwestern Tibet. This rudist data suggeststhat Kauffman’s Eastern Mediterranean Subprovince may becomposed of two distinct assemblages that were partlygeographically isolated. This issue may be addressed by moredetailed collections from these areas.
Early Cretaceous bivalve assemblages of the northernIndian Plate and Himalayan belts consist of Gondwanaspecies similar to those in Australia, Antarctica, East Africa,and South America. These assemblages comprise the Himal-yan Subprovince or endemic center (Wen, 1999, 2000). No
rudists are known from this subprovince; however, theTithonian megalodontid, Protodiceras lanonglaensis Li andGrant-Mackie (1994), occurs west of Mount Everest/Qomo-langma in the Nyalam district at the base of the XuomoFormation. By Campanian-Maastrichtian times the IndianPlate had moved into the tropical zone and bivalves includingrudists were part of the Tethyan Eastern MediterraneanSubprovince (Gou and Shi, 1998; Wen, 1999). Commonrudists were endemic species of Biradiolites, Bournonia,Lapeirousia, Plagioptychus, and Praeradiolites (Appendix 1).
North of the Yarlung-Zangbo suture zone on the LhasaBlock, bivalve assemblages include Tethyan and eastern Asiantypes of the Eastern Mediterranean Subprovince (Wen, 1999).In the Gandes forearc basin, a possible Eoradiolites is reportedfrom the Aptian-Albian Sangzugang Formation, which isequivalent to the Langshan and Maryo formations. Rudistgenera in the Albian-Cenomanian Langshan Formationinclude Monopleura, Praeradiolites, Requienia, Rutonia, andToucasia; two of these fourteen species are cosmopolitan(Appendix 1). Farther northwest in the Karakorum Belt of theQiantang Block, Early Cretaceous rudist species are endemicand two of five Late Cretaceous species are endemic. UpperCretaceous Turonian rudists in the lower part of theTielongtan Group are Biradiolites, Durania, Praeradiolites,Sauvagesia, and Vaccinites (including its junior synonymRhedensia) (Gou and Shi, 1998; Wen, 1999). The youngerSantonian-Maastrichtian assemblage in the Tielongtan in-cludes Distefanella, Lapeirousia, and Hippurites (Appendix 1).
On the Eurasian Block in the Tarim Basin and in the WestKunlun Belt, the Cenomanian-Turonian Kukebai Formationhosts the cosmopolitan species of Ichthyosarcolites amongother Tethyan bivalves. Farther west in Tadzhikistan andUzbekistan this genus occurs with Caprinula soluni Bobkovain the middle Cenomanian section (Pojarkova, 1984). LateCretaceous rudists in the Tielongtan Group and YigeziyaFormation of the West Kunlun Belt are Biradiolites,Gyropleura, Lapeirousella, Osculigera, and Sauvagesia, how-ever the majority of species appear to be endemic (Appendix 1)(Gou and Shi, 1998; Wen, 1999).
STRATIGRAPHIC DISTRIBUTION OF REQUIENIIDAE
Species of the Family Requieniidae first appear in the latestKimmeridgian Age and went extinct in the Maastrichtian Age(Fig. 4). Although this family did not diversify during theCretaceous Period to the extent that other rudist groups did, itwas a persistent component of shallow water carbonate shelfcommunities. As did other rudist groups, Requieniidaeexperienced multiple extinctions during or soon after someCretaceous oceanic anoxic events (Masse and Philip, 1986;Ross and Skelton, 1993; Masse, 1994, 2002; Scott, 1995). Theoldest requieniid species, Hypelasma salevensis (Favre inJoukowsky and Favre, 1913) evolved from diceratids, suchas Epidiceras or Plesiodiceras, during the latest Kimmeridgianin southern France (Skelton, 1999; Gourrat et al., 2003).Matheronia appeared in the Tithonian and diversified in theEarly Valanginian but suffered extinction during the mid-Valanginian crisis (Masse, 2002). During the Hauterivian,members of the family became cosmopolitan and diversifiedinto the Barremian and Early Aptian, when the family wascomposed of four genera. Two genera went extinct followingthe early Aptian OAE1a and two survived. The familydiversified again during the Albian and Cenomanian, butonly one genus survived the latest Cenomanian–earliestTuronian OAE2. Following this stressful even,t the familywas represented by two genera that continued into the
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:19 3 Cust # 09-137
FIGURE 3—Plate tectonic sketch map during the latest Cretaceous toearliest Paleogene time, c. 81–58 Ma (modified from Golonka, 2002, fig.34 and Masse and Gallo Maresca, 1997, fig. 5). Black ovals indicate Ngariand Zongshan areas of Fig. 1. Dashed lines–ocean spreading center andtransform faults; dash-dot lines–subduction zones; solid lines–approxi-mate modern plate boundaries.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

Campanian-Maastrichtian it (Fig. 4). Requieniid speciesdiversity and abundance apparently dwindled through theMaastrichtian and the family did not survive the Cretaceousextinction.
REQUIENIIDAE SHELL GEOMETRY
Shell geometry is genetically programmed for adaptation tospecific substrate relationships (Raup, 1966) as has beendemonstrated for rudists (Skelton, 1991; Masse, 1994, 2002).Raup (1966) has shown that shell geometry can be defined byfour parameters: 1) shape of the generating curve, which ismapped by the commissure outline; 2) the rate at which thewhorl expands during growth; 3) the shift of the commissureaway from the coiling axis; and 4) the rate at which the whorlis displaced along the coiling axis. These parameters weremeasured on caprinids and aid in their taxonomic discrimi-nation and testing of evolutionary hypotheses (Scott andWeaver, 2008; Scott and Weaver, in press).
The Requieniidae are divided into three morphologicgroups by the coiling and expansion rate of the LV-AV andRV-FV and by the inferred orientation to the substrate:matheroniform, requieniform and toucasiform (Masse, 1994,2002). The matheroniform morphotype includes elongateinflated valves that expand in diameter rapidly from theprotoconch, have a low trochospiral form, and have ananterior margin is somewhat flattened; the anterior myo-
phores are rounded swellings on the shell wall and theposterior myophores are platforms. This form was epifaunal,resting on is anterior margin. The requieniform morphotype ischaracterized by a slowly expanding valve with a tightly coiledspire that is translated along the coiling axis and by relativelysymmetrical anterior and posterior margins. The myophoresare either subequal or the posterior is larger. This form wassemi-infaunal with its spire inserted into the sediment. Thetoucasiform morphotype has a rapidly expanding valve with amoderate spiral and with a flattened anterior margin; the valveis greatly compressed anteriorly-posteriorly and is keeled; theposterior myophore is the larger. This form was attached tothe substrate at least in its early growth (Masse, 2002, 2009).
The different coiling geometries of these three morphotypescan be defined by the translation along the coiling axis (T), thewhorl expansion rate (W), and the increasing distance of thecommissure from the coiling axis (D) (Raup, 1966; Raup andStanley, 1971) (Fig. 5). These parameters were estimated bymeasuring selected specimens (Appendix 2) and by comparingtypical specimens of the genera with the diagrams of Raup(1966) and Raup and Stanley (1971). The matheroniformmorphotype is characterized by Matheronia munieri Paquierfrom Barremian limestone in southern France (in Douville,1918, Pl. 1, figs. 8, 9). The LV is coiled approximately onewhorl and translation along the axis is low (T , 1). The whorlexpansion rate is low (W , 10) and the commissure is
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:19 4 Cust # 09-137
FIGURE 4—Stratigraphic ranges of requieniid genera and inferred evolutionary branching.
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010

displaced only slightly along the axis (D , 0.1–0.2) (Fig. 5,MfLV). This morphotype is homeomorphic with manyexogyrid bivalves.
The requieniform morphotype is characterized by Requieniaammonia (Goldfuss) from Barremian limestone in southernFrance (in Douville, 1918, Pl. 1, figs. 6a, 6b; Masse, 1988b, Pl.1, figs. 1–3). The LV is coiled up to three turns and translated(T) moderately. The whorl expansion rate is low (W , 10) andthe displacement is slightly greater than the matheroniform(Fig. 5, RfLV). The toucasiform morphotype is represented byseveral species because the shell form is variable. Thetrochospiral LV has up to three clockwise turns and the lowtrochospiral RV is about two counterclockwise whorls. Thetall spired form (Fig. 5, TfLV1) is represented by Toucasiapatagiata (White, 1884) from the Middle-Upper AlbianEdwards Formation in Texas and the low spired form(Fig. 5, TfLV2) is represented by Toucasia hancockensisWhitney (1952) in Garcıa-Barrera (1995, Pl. 1, figs. 3, 6) fromLower Albian limestones in southwest Mexico and Texas. Therequieniform and toucasiform morphotypes are homeomor-phic with the typical gastropod form as noted earlier byYounge (1967, p. 87). However, shell coiling served verydifferent functions in these groups. The requieniid coilingenabled the bivalve to fix itself in the substrate, in contrast tomobile gastropods in which the coiled valve encased the softparts and positioned them above the large foot.
The diceratids, including Diceras, Heterodiceras, Epidiceras,Plesiodiceras, and Eodiceras, have two inflated, tortedprosogyrous valves. The geometry of both valves is similarto the geometry of the Matheronia LV, except that the coilingaxis of diceratids is at a larger angle to the commissure planethan in requieniids (Gourrat et al., 2003). Thus to deriverequieniids from epidiceratids, the inflation of the RV wasreduced and coiling increased slightly. Requieniids evolvedfrom diceratids and expanded into offshore, mainly shallow,
marine habitats. For example, in Austria Heterodiceras shellsare part of Tithonian subtidal deposits offshore of gravellybeach deposits (Sanders et al., 2007). These shells have beendisturbed by periodic high-energy events and other tapho-nomic processes.
PHYLOGENETIC SPECULATIONS ON REQUIENIIDAE
The Order Hippuritoida Newell (1965) is a clade character-ized by a two-layer shell wall, the outer of which is composedof fibrous prismatic calcite and the inner originally ofaragonite (Skelton, 1978; Skelton and Masse, 2000; Skeltonand Smith, 2000). Dentition consists of two teeth in the RVand one in the LV. Species have two subequal adductormuscles either on the shell wall or on specialized plates. TheHippuritoid clade is divided into two monophyletic cladesbased on which valve is on the substrate. Members of theSuperfamily Requienioidae Douville (1914) are attached to thesubstrate by the LV-AV and the family is composed ofepidiceratids including Epidiceras, Heterodiceras, and Plesio-diceras and the monophyletic Family Requieniidae Douville(1914) (Skelton, 1978, 1999, 2008; Masse, 1994, 2002).However, Malchus (1995) suggested that Requieniidae areparaphyletic because he believed that the genera do not share aderived character. All other rudists comprise the SuperfamilyHippuritoidea Gray (1847), which has the RV-AV on thesubstrate (Skelton, 1978, 2008; Skelton and Masse, 2000).Plesiomorphic characters shared by the epidiceratids andrequieniids are: 1) external ligament in a groove, 2) spirogyrateshell, 3) two teeth in RV and one tooth in LV, 4) subequalmuscle structures, and 5) shell composed of a prismatic outercalcite layer and an inner aragonitic layer (Skelton, 1978, 1999;Masse, 2002). Autapomorphic characters are: 1) highlyinequivalved with smaller RV, 2) LV trochospiral tohelicospiral, 3) coiling axis at small angle to commissuralplane so umbo crosses commissure plane, 4) LV anterior a flat
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:20 5 Cust # 09-137
FIGURE 5—Three-dimensional morphospace of mollusk shell geometry (Raup and Stanley, 1972). Requieniidae morphology compares to thatof Gastropoda.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

surface clinging to substrate (Gourrat et al., 2003). Because ofthese shared characters the Family Requieniidae would seemto be monophyletic.
The Family Requieniidae is composed of twelve genera andsubgenera (Dechaseaux and Perkins, 1969) and two subse-quently created by Yang et al. (1982) and Masse (1988a).Eleven valve characters make up the data matrix used bycladistic anaylsis (Table 1; Appendix 3). Some genera aredescribed less thoroughly than others so that the matrix israther small. To test these phylogenetic hypotheses thePhylogenetic Analysis Using Parsmony-PAUP 4.0 Betasoftware was run (Swofford, 2002) assuming Heterodicerasas the ‘outgroup’ genus. Two consensus trees show similargroups of taxa (Fig. 6A, B). The ‘‘strict consensus tree’’(Fig. 6A) is a very conservative estimate of consensus thatseparates two generic groups (Swofford, 2002; Forey, 2009).The ‘‘majority-rule concensus tree’’ (Fig. 6B) results in similargeneric groups although they are rooted at higher levels thanthe strict consensus tree. These groups are found in more thanhalf of the rival trees. In both trees Apricardia, Requienia,Toucasia, and Pseudotoucasia are closely grouped and Bayleiaand Bayleoidea are in the next closest branch. Bayleoidea,however, requires a thorough re-examination to consider theorigin of the accessory cavity (J.-P. Masse, 2010, personalcommunication). A second group is more loosely allied;Lovetchenia, Hypelasma, Matheronia, and Monnieria areseparated from Kugleria and Rutonia.
The suggested evolutionary relationships and the basiccharacter states delineate two requieniid clades (Fig. 6C).Shared primitive character sets of the Requieniidae are 1)inequivalved shell in which the LV-AV is larger and moretorted than the RV-FV; 2) distinct growth rings; finer growthlines develop in younger genera and radial striae with finegrowth lines are on three genera; 3) RV-FV dentition iscomposed of a larger posterior and a smaller anterior tooth; 3)the posterior muscle attachment is larger than the anterior andattached to various projecting posterior myophore structures;and 4) two-layered wall structure of outer calcite and inneraragonite (Skelton, 1978; Masse, 2002). The LV-AV of speciesof the Requieniidae is prosogyrate coiled clockwise whenviewed from the exterior and is much larger than the RV(Skelton and Smith, 2000; Gourrat et al., 2003). This coilingdirection is opposite to the opisthogyrate counter-clockwiseLV-AV of oysters such as Exogyra, which has been noted bymany rudist specialists (see Malchus, 1995).
Two end members of valve geometry each appear to becharacterized by primitive and derived characters respectivelyand separate the Requieniidae into two clades (Fig. 6C). Theend members are defined by the coiling geometry, whether thespire is close to the plane of commissure or it is translatedalong the coiling axis. The older matheroniform clade has alow spirogyrate LV that is translated slightly from thecommissure along the coiling axis; this group is composed ofMatheronia (and its subgenus Monnieria), Hypelasma, Love-tchenia, Rutonia, and Kugleria. Genera in the younger cladehave a tall trochospiral LV that is translated along the coilingaxis and consists of Requienia, Toucasia, Pseudotoucasia,Apricardia, Bayleoidea, and Bayleia. The requieniform LV ofRequienia is translated more greatly than the toucasiform LV.The geometry of the RV also separates the two clades. In thematheroniform clade the RV is slightly inflated and is coiledwith little translation, whereas in the younger clade ofrequieniform and toucasiform groups the RV is flat to convexwith little or no coiling translation. Growth rings are
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:20 6 Cust # 09-137
TA
BL
E1—
Req
uie
nii
dm
orp
ho
logic
chara
cter
su
sed
incl
ad
isti
can
aly
sis.
See
Ap
pen
dix
2fo
rch
ara
cter
des
crip
tio
ns.
Gen
era
1.
Mic
ro-
stru
ctu
re2
.L
VS
hel
lF
orm
3.
RV
Sh
ell
Fo
rm4
.L
VO
rna
men
t5
.L
VM
yo
ph
ore
6.
RV
My
op
ho
re7
.S
iph
on
al
ba
nd
s8.
Acc
esso
ryca
vit
ies
9.
RV
Po
ster
ior
Den
titi
on
10
.L
VD
enti
tio
n1
1.
Lig
am
ent
Ap
rica
rdia
12
23
01
00
11
0
Req
uie
nia
13
23
11
10
?1
0
To
uca
sia
12
23
00
10
01
0
Pse
ud
oto
uca
sia
12
22
?1
00
??
0
Ba
ley
ia1
22
20
00
10
10
Ba
yle
oid
ea1
22
20
00
12
10
Lo
vetc
hen
ia1
11
20
00
00
10
Hy
pel
asm
a1
11
21
00
00
10
Ku
gle
ria
12
12
?0
00
1?
0
Ma
ther
on
ia1
11
10
00
00
10
Mo
nn
ieri
a1
11
11
00
00
10
Ru
ton
ia1
11
2?
00
00
?0
Ou
tgro
up
Dic
era
s1
00
10
00
00
00
Het
ero
dic
era
s1
00
00
00
00
00
Epi
dic
era
s1
00
10
00
00
00
Ple
siodic
eras
10
01
00
00
00
0
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010
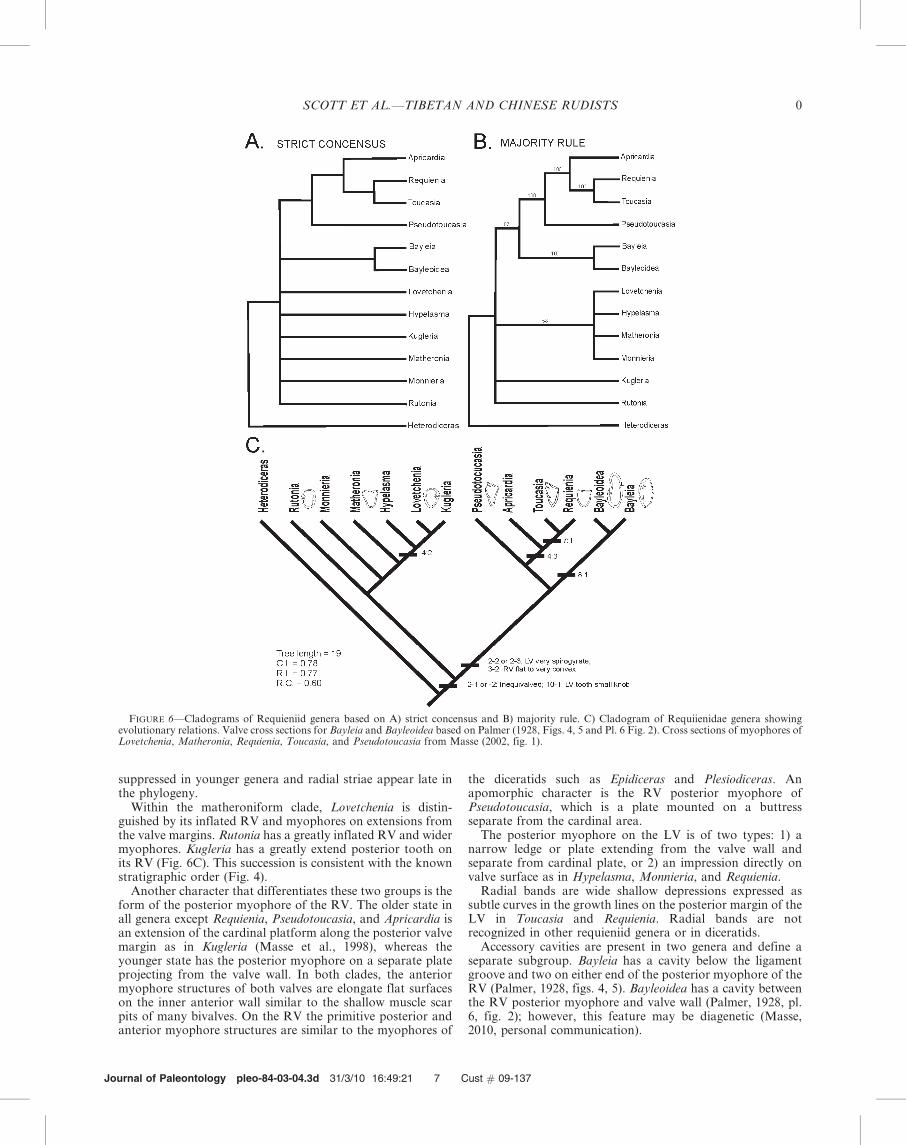
suppressed in younger genera and radial striae appear late inthe phylogeny.
Within the matheroniform clade, Lovetchenia is distin-guished by its inflated RV and myophores on extensions fromthe valve margins. Rutonia has a greatly inflated RV and widermyophores. Kugleria has a greatly extend posterior tooth onits RV (Fig. 6C). This succession is consistent with the knownstratigraphic order (Fig. 4).
Another character that differentiates these two groups is theform of the posterior myophore of the RV. The older state inall genera except Requienia, Pseudotoucasia, and Apricardia isan extension of the cardinal platform along the posterior valvemargin as in Kugleria (Masse et al., 1998), whereas theyounger state has the posterior myophore on a separate plateprojecting from the valve wall. In both clades, the anteriormyophore structures of both valves are elongate flat surfaceson the inner anterior wall similar to the shallow muscle scarpits of many bivalves. On the RV the primitive posterior andanterior myophore structures are similar to the myophores of
the diceratids such as Epidiceras and Plesiodiceras. Anapomorphic character is the RV posterior myophore ofPseudotoucasia, which is a plate mounted on a buttressseparate from the cardinal area.
The posterior myophore on the LV is of two types: 1) anarrow ledge or plate extending from the valve wall andseparate from cardinal plate, or 2) an impression directly onvalve surface as in Hypelasma, Monnieria, and Requienia.
Radial bands are wide shallow depressions expressed assubtle curves in the growth lines on the posterior margin of theLV in Toucasia and Requienia. Radial bands are notrecognized in other requieniid genera or in diceratids.
Accessory cavities are present in two genera and define aseparate subgroup. Bayleia has a cavity below the ligamentgroove and two on either end of the posterior myophore of theRV (Palmer, 1928, figs. 4, 5). Bayleoidea has a cavity betweenthe RV posterior myophore and valve wall (Palmer, 1928, pl.6, fig. 2); however, this feature may be diagenetic (Masse,2010, personal communication).
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:21 7 Cust # 09-137
FIGURE 6—Cladograms of Requieniid genera based on A) strict concensus and B) majority rule. C) Cladogram of Requiienidae genera showingevolutionary relations. Valve cross sections for Bayleia and Bayleoidea based on Palmer (1928, Figs. 4, 5 and Pl. 6 Fig. 2). Cross sections of myophores ofLovetchenia, Matheronia, Requienia, Toucasia, and Pseudotoucasia from Masse (2002, fig. 1).
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

Dentition of the RV is composed of two cardinal teeth, theposterior of which is taller than the anterior one and the LVhas one smaller tooth. In most genera the large posterior toothof the RV is arcuate, but in Apricardia and Kugleria it is acurved cone. In Bayleoidea the teeth are subequal knobs.
The ligament is a primitive character of both clades and issituated in a curved groove external to the dentition and onthe posterior margin as it is in the diceratids.
The Tibetan Albian-Cenomanian Rutonia appears to besimilar to older requieniid types and illustrates iterativeevolution. Its morphology is similar to morphologies of LateJurassic-earliest Cretaceous diceratids and the Hauterivian-early Aptian requieniid, Lovetchenia. Both morphotypes haveprosogyrally coiled subequal, inflated valves. The LV wasattached or inserted into the soft substrate. The RV is coiled ina plane with little translation along the coiling axis. Twophylogenetic hypotheses on the origin of Rutonia must betested. First, that it descended from Lovetchenia, although nolate Aptian to early Albian species has yet been found. Masse(2002) proposed that Lovetchenia evolved from Matheronia byexpanding the myophore plates on both valves and inflatingthe RV. The myophore plates of Rutonia are similar in formand structure to those of Lovetchenia. However the coilingplanes of both valves of Lovetchenia are close to thecommissure plane, whereas in Rutonia the spire of the LV isslightly displaced along the coiling axis and the valve is slightlytrochospiral. This hypothesis is consistent with a monophy-letic subfamily Matheroniinae new subfamily herein.
An alternative hypothesis is that Rutonia evolved from oneof the late Albian species of Requienia, Toucasia, orPseduotoucasia. The valves of Requienia are quite asymmetric,the RV-FV being flat operculiform so that the genus is anunlikely ancestor. Toucasia, however, is a candidate ancestorthat ranges from Barremian to Cenomanian and both valvesof some species are trochospirally coiled like Rutonia.However the posterior myophore on the FV of Rutonia is anextension of the cardinal platform and in Toucasia it is aseparate plate extending from the valve wall. Also the largeposterior tooth on the RV-FV of Rutonia is positioned moreposteriorly than it is on Toucasia. This hypothesis implies thatMatheroniinae is polyphyletic.
In the more derived clade of requieniforms and toucasi-forms, the RV of Toucasia and others is flat to opercular andthe RV posterior myophore is Y-shaped (Fig. 6C). InPseudotoucasia the RV posterior myophore has migratedfrom the valve margin to an inner position. Bayleoidea has asingle cavity between the wall and the myophore plate thatmay not be a primary feature. In Apricardia the posteriormyophore is a plate below and separate from the cardinalshelf, and its posterior tooth is enlarged (Mainelli, 1995).Finally Bayleia has developed three small cavities along itsposterior shell margin of the RV. The high spirogyrateRequienia was derived from Hypelasma by the developmentof the posterior myophore plate on the RV. Toucasia andPseudotoucasia then descended from Requienia by the additionof a keeled LV and a posterior myophore plate in the LV.
Palmer (1928, p. 37) suggested that Toucasia was theancestor of Bayleia, which evolved to Bayleoidea. HoweverBayleoidea is older than Bayleia and cannot be the descendentspecies. These three genera have very similar external forms inwhich the larger LV-AV is spirogyrate, with coiling displacedslightly along the coiling axis; its posterior margin is flattened.The much smaller RV-FV is opercular. All three have smallteeth in both valves. The posterior myophore structure onboth valves of each species is a thin plate jutting interiorly
from the valve wall. The anterior muscles were attached to thevalve interior wall. Bayleoidea and Bayleia differ fromToucasia mainly by the presence of one and three posteriorcavities respectively and by the absence of fine radial striaeand radial bands.
SYSTEMATIC PALEONTOLOGY
Order HIPPURITOIDEA Newell, 1965Superfamily REQUIENIOIDAE Douville, 1914
Family REQUIENIIDAE Douville, 1914
This family includes the genera Requienia Matheron 1843,Apricardia Gueranger 1853, Matheronia Munier-Chalmas1873 (including the subgenus Monnieria Paquier, 1898),Toucasia Munier-Chalmas 1873, Bayleia Munier-Chalmas1873, Hypelasma Paquier, 1898, Pseudotoucasia Douville1911, Bayleoidea Palmer 1928, Kugleria Bouman 1938,Rutonia Yang, Nie, Wu, and Liang 1982, and LovetcheniaMasse 1988a. Most species in these genera are stronglyinequivalved, generally the LV umbo intersects the commis-sure plane, and the outer shell layer is thick. The largerprosogyrate left valve (LV) lay upon the substrate as theattached valve (AV); the smaller right valve (RV) is the upper,free valve (FV). Most genera are simply ornamented by faintgrowth lines or winkles; radial striae rare. The externalligament is a shallow arcuate groove dorsal to the cardinalplatform. Dentition in LV-AV is a single small central toothand in RV-FV two unequal teeth, the posterior tooth generallyis the larger of the two. Anterior muscle insertion in LV-AV ison shell wall and in RV-FV it is on the wall or on an extensionof cardinal platform. The posterior muscle insertion in the LV-AV is on the shell wall or a myophore plate. The posteriormuscle insertion in the RV-FV is on a myophore plateextending either from the shell wall at a right angle or from thecardinal platform (Malchus, 1998; Masse et al., 1998; Gourratet al., 2003). Radial bands are weakly developed on posteriorside of LV-AV of some species.
The Requieniidae evolved from diceratids, possibly Plesio-diceras muensteri (Goldfuss, 1840) in the latest Kimmeridgianand diversified moderately during the Cretaceous as did otherrudist families; the family was extinct by the end of theCretaceous (Fig. 4) (Gourrat et al., 2003; Masse, 1994;Skelton, 1978, 1985, 1991). Requiieniid species were locallyabundant as frictional or attached clingers (Skelton, 1991) incaprinid buildups and in shelf biostromes (Scott, 1981).
Subfamily REQUIENIINAE Douville, 1914 new subfamily
Type genus.—Requienia Matheron, 1843 (nominotypicaltaxon).
Diagnosis.—LV requieniform or toucasiform, tall trochos-pire translated along the coiling axis; RV flat to convex withlittle or no coiling translation; the LV myophores are inflatedareas on the shell wall or projecting plates; the RV posteriormyophore plate is separate from the cardinal platform; RVposterior tooth reduced in size in some genera. Growth ringsare suppressed; some genera with radial bands; some withradial striae; some with accessory canals. The subfamilyincludes the genera Requienia, Toucasia, Pseudotoucasia,Apricardia, Bayleoidea, and Bayleia.
Subfamily MATHERONIINAE new subfamily
Type genus.—Matheronia Munier-Chalmas, 1873.Diagnosis.—LV matheroniform, low spirogyre translated
slightly from the commissure along coiling axis; RV slightlyinflated, coiled with little translation along axis; the LVmyophores are expanded plates on the valve wall; on the RV
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:22 8 Cust # 09-137
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010

the posterior myophore plate extends from the cardinalplatform; RV posterior tooth a large arcuate ridge. Thesubfamily includes Matheronia (and its subgenus Monnieria),Hypelasma, Lovetchenia, Rutonia, and Kugleria.
Genus LOVETCHENIA Masse, 1988a
Type species.—Requienia lovcensis Zlatarski, 1886, p. 312,pl. 3.Matheronia lovetchensis (Zlatarski); PAQUIER, 1903, p. 25, pl.
II, fig. 3; pl. III, figs. 1–3.Lovetchenia lovetchensis (Zlatarski); MASSE, 1988a; 1993;
2002.
Diagnosis.—Lovetchenia has a very convex RV-FV that isdistinctly spiral in the plane of commissure. Both myophoresof the RV-FV are interior platforms subparallel with thecommissure plane and extend from the cardinal platform. Onthe LV-AV the posterior myophore is a narrow platformcircumscribed by a distinct bourrelet (Masse, 2002).
Discussion.—Lovetchenia is found in Hauterivian to lowerAptian carbonate strata along the northern margin of theMediterranean Sea (Masse, 1988a).
Genus RUTONIA Yang, Nie, Wu, and Liang, 1982
Type species.—Rutonia bangonghuensis Yang, Nie, Wu, andLiang, 1982, p. 297.
Diagnosis.—Both valves very convex, inflated; beak of LV-AV displaced along coiling axis away from commissure plane.Dentition of RV-FV consists of two teeth, a small, knob-likeanterior tooth and tall arcuate posterior tooth along thepostero-dorsal margin. Posterior muscle of RV-FV inserted onwide plate extending ventrally from cardinal platform. Othermyophores are unknown. Ligament groove narrow alongpostero-dorsal margin.
Original Description (translated).—Character: large inequi-valved shell. Left valve spirally twisted for several whorls;right valve (RV-FV) is free, coiled once, operculate. The rightvalve has two teeth, the anterior one smaller and the posteriorone larger, the anterior tooth only half as long as posteriortooth; both separated by a socket. The LV-AV has nopreserved teeth. The anterior adductor scar in the RV-FV isanterior to the anterior tooth and the posterior muscle scar ison the hinge plate posterior to the tooth. The ventral marginof the right valve is broadly concave, thick shelled with a smallbody cavity.
Original Discussion.—We place this new genus in Requie-niidae because of three main characters: (1) the LV is fixed, theRV is free; (2) the LV is strongly coiled and the RV is cap-shaped; and (3) the RV has two large teeth, the anterior issmall and the posterior is large. The FV-LV is cone-shaped,turbinate, the FV-RV is low cone-shaped and curved; theposterior tooth of the RV is much larger and arched than theanterior tooth; the RV has grooves, which differs from otherRequieniidae genera (translation).
Discussion.—Rutonia differs from Lovetchenia by its some-what smaller posterior myophore plate on the RV-FV and byits more inflated RV-FV. Rutonia differs from Matheronia byits more inflated RV-FV, by its longer posterior tooth and byits longer posterior myophore on the RV-FV. The LV-AV ofMonnieria is more elongate and keeled than that of Rutonia.The LV-AV of Rutonia is tightly coiled with moderatetranslation along the axis and serves to support the animalin the soft substrate. The RV-FV of Rutonia is weakly coiledwith slight translation. Both valves are inflated indicating thatsoft parts were distributed in both valves.
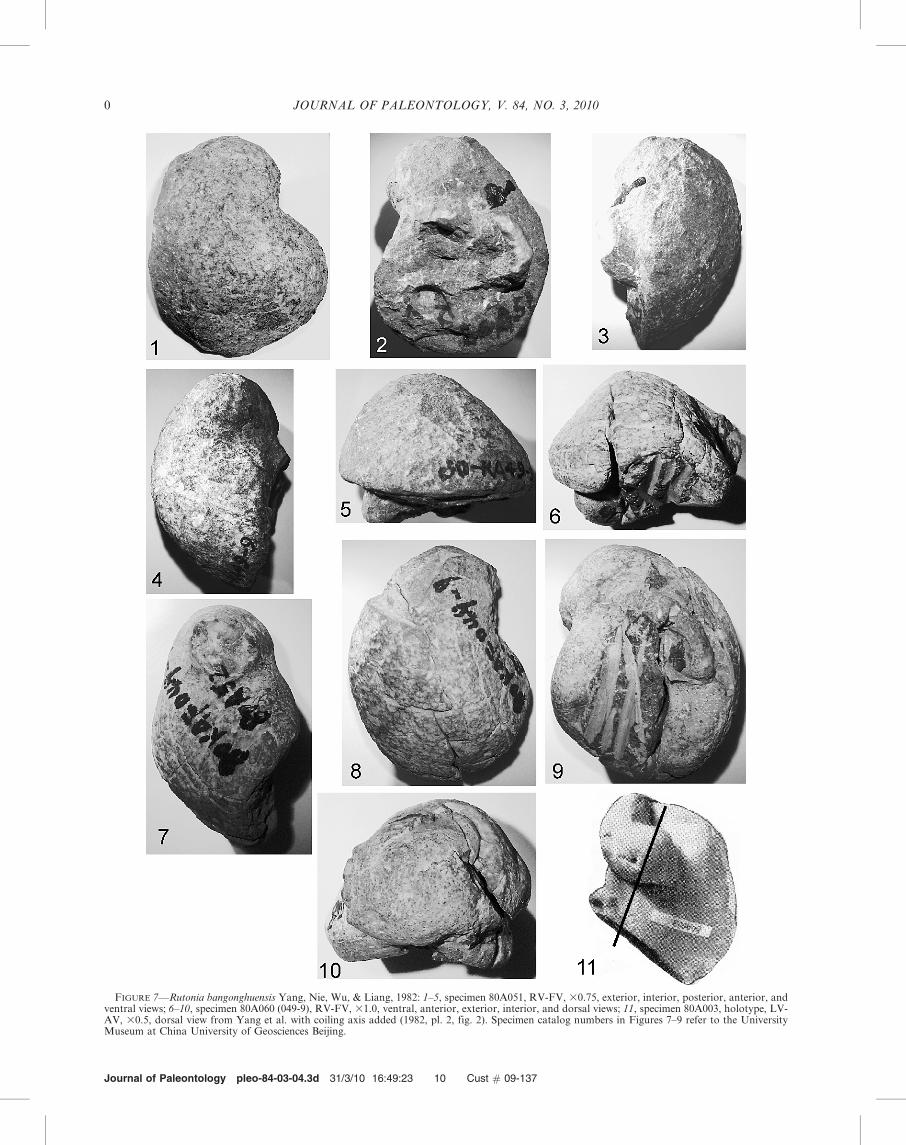
RUTONIA BANGONGHUENSIS Yang, Nie, Wu, and Liang, 1982Figure 7.1–7.11
Rutonia bangonghuensis YANG, NIE, WU, AND LIANG, 1982,pl. 2, figs. 2, 3, 6a, 6b, 7a, 7b.
Type specimens.—Holotype 80A003 LV, location of spec-imen unknown; paratypes 80A058, RV, 80A051, 80A060(052)deposited in the University Museum, China University ofGeosciences, Beijing.
Diagnosis.—Monospecific genus.Original Description (translation).—Shell large, inequi-
valved; LV fixed, coiled, conical to turbinate, curved, changinggreatly in shape. Valve height greater than 100 mm, diameterof the valve up to 70mm. The early growth stage is gentlycoiled and later becomes straight. RV nearly planispiral,coiling for one turn into low, conical shape, cap-like, and hasobvious grooves. The RV has two teeth; the anterior one isabout half the size of the posterior one; the posterior tooth is ahigh, protruding, curved arc; the two teeth are parallel andextend dorsally, separated by a deep triangular pit that reflectsthe tooth of the LV; but the teeth of the LV are not preserved.The shell is thick and the body cavity is shallow (translation).
Description.—LV-AV moderate size, thick, prosogyrateclockwise up to 360u, deeply inflated, beak rounded elongate,expanding slowly into adult valve. Internal features notobserved. RV-FV moderate size, thick, prosogyrate counter-clockwise up to 180u, deeply inflated, beak broadly rounded,oriented posteriorly; commissure outline ovate, slightly extendedpostero-ventrally at postero-ventral corner; commissure margindeeply folded into a trough; umbo greatly arched, posterior flanksteeper than anterior; valve surface with faint concentric growthrings. External ligament groove originates beneath beak andextends posteriorly in a broad arch to end of tooth. Two teeth,anterior tooth conical, posterior tooth tall, thick, long, curvedfrom below beak to posterior margin, surface smooth. Myophoresurfaces gently arched wide, smooth. Measurements of specimen80A051: height 7.5 cm, length 6.1 cm, width 4.0 cm; specimen80A060(052): height 6.3 cm, length 4.8 cm, width 3.4 cm.
Discussion.—Rutonia has internal structures that are similarto Lovetchenia. The myophores of the RV-FV are wideplatforms attached to the valve inner wall as in Lovetchenia.The dentition and ligament groove of the RV-FV are also likethat of Lovetchenia. Other features are not yet known. Rutoniaand Lovetchenia differ from Matheronia, which has a flat cap-shaped RV-FV and on the LV-AV the beak is close to thecommissure. The LV-AV of Rutonia differs from that ofLovetchenia; in Rutonia the beak of the LV-AV is displacedalong the coiling axis from the commissure plane, whereas inLovetchenia the beak coils in the commissure plane. The coilingof the LV-AV is closer to that of Toucasia and Pseudotoucasia.
Distribution.—Rutonia bangonghuensis is known only fromthe Ngari district of northwestern Tibet between the Yarlung-Zangbo and the Bangon-Nu River suture zones. It is interbeddedin the Langshan Formation with Upper Albian-Lower Cen-omanian orbitolinids: Orbitolina aperta (Erman) (late Albian-early Cenomanian), Orbitolina concava (Lamarck), (earlyCenomanian), Orbitolina conica (D’Archiac) (early-middle Cen-omanian), and Orbitolina concava var. qatarica Henson (earlyCenomanian) (Yang et al., 1982; Wan et al., 2003).
INDETERMINATE REQUIENIID VALVES
Figure 8.6–8.10
Specimens.—Specimen 80A54 and specimen 80A59 depos-ited in the University Museum, China University of Geosci-ences, Beijing.
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:22 9 Cust # 09-137
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:23 10 Cust # 09-137
FIGURE 7—Rutonia bangonghuensis Yang, Nie, Wu, & Liang, 1982: 1–5, specimen 80A051, RV-FV, 30.75, exterior, interior, posterior, anterior, andventral views; 6–10, specimen 80A060 (049-9), RV-FV, 31.0, ventral, anterior, exterior, interior, and dorsal views; 11, specimen 80A003, holotype, LV-AV, 30.5, dorsal view from Yang et al. with coiling axis added (1982, pl. 2, fig. 2). Specimen catalog numbers in Figures 7–9 refer to the UniversityMuseum at China University of Geosciences Beijing.
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010
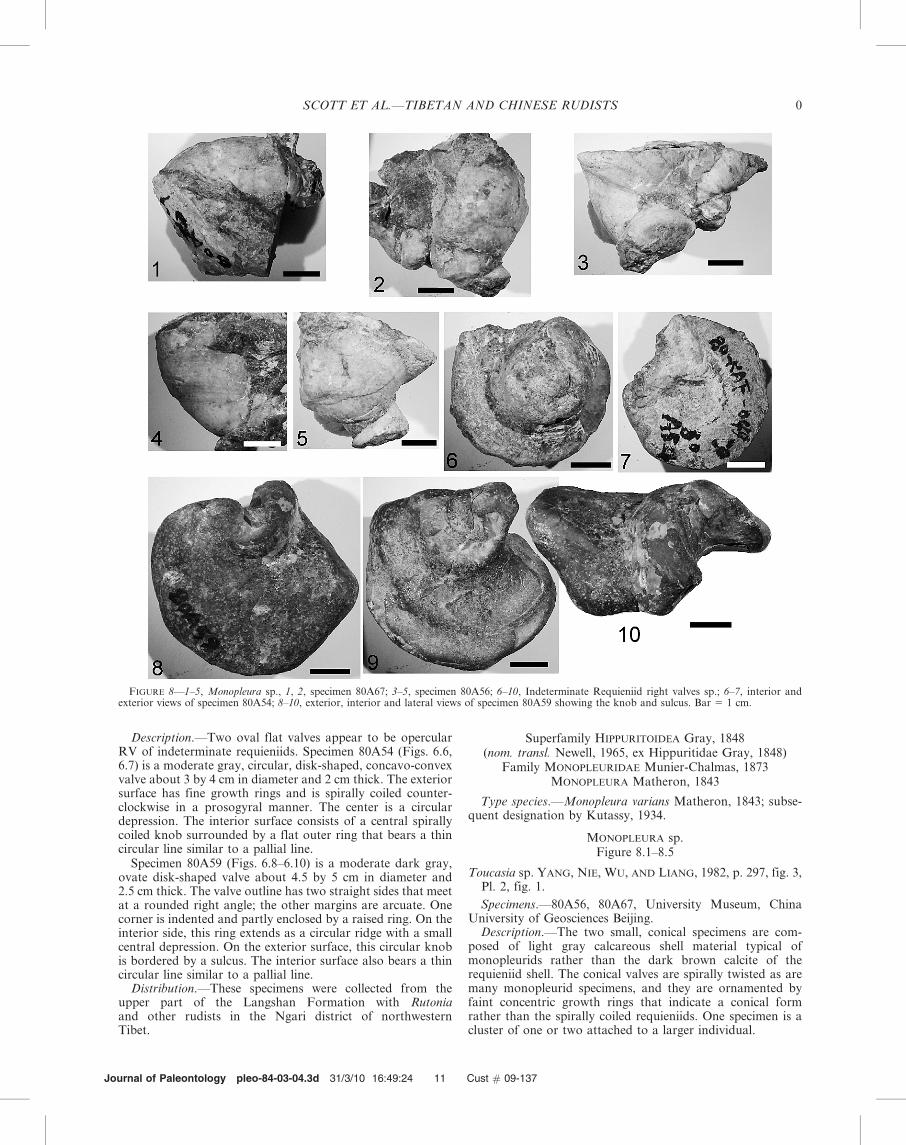
Description.—Two oval flat valves appear to be opercularRV of indeterminate requieniids. Specimen 80A54 (Figs. 6.6,6.7) is a moderate gray, circular, disk-shaped, concavo-convexvalve about 3 by 4 cm in diameter and 2 cm thick. The exteriorsurface has fine growth rings and is spirally coiled counter-clockwise in a prosogyral manner. The center is a circulardepression. The interior surface consists of a central spirallycoiled knob surrounded by a flat outer ring that bears a thincircular line similar to a pallial line.
Specimen 80A59 (Figs. 6.8–6.10) is a moderate dark gray,ovate disk-shaped valve about 4.5 by 5 cm in diameter and2.5 cm thick. The valve outline has two straight sides that meetat a rounded right angle; the other margins are arcuate. Onecorner is indented and partly enclosed by a raised ring. On theinterior side, this ring extends as a circular ridge with a smallcentral depression. On the exterior surface, this circular knobis bordered by a sulcus. The interior surface also bears a thincircular line similar to a pallial line.
Distribution.—These specimens were collected from theupper part of the Langshan Formation with Rutoniaand other rudists in the Ngari district of northwesternTibet.
Superfamily HIPPURITOIDEA Gray, 1848(nom. transl. Newell, 1965, ex Hippuritidae Gray, 1848)
Family MONOPLEURIDAE Munier-Chalmas, 1873MONOPLEURA Matheron, 1843
Type species.—Monopleura varians Matheron, 1843; subse-quent designation by Kutassy, 1934.
MONOPLEURA sp.Figure 8.1–8.5
Toucasia sp. YANG, NIE, WU, AND LIANG, 1982, p. 297, fig. 3,Pl. 2, fig. 1.
Specimens.—80A56, 80A67, University Museum, ChinaUniversity of Geosciences Beijing.
Description.—The two small, conical specimens are com-posed of light gray calcareous shell material typical ofmonopleurids rather than the dark brown calcite of therequieniid shell. The conical valves are spirally twisted as aremany monopleurid specimens, and they are ornamented byfaint concentric growth rings that indicate a conical formrather than the spirally coiled requieniids. One specimen is acluster of one or two attached to a larger individual.
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:24 11 Cust # 09-137
FIGURE 8—1–5, Monopleura sp., 1, 2, specimen 80A67; 3–5, specimen 80A56; 6–10, Indeterminate Requieniid right valves sp.; 6–7, interior andexterior views of specimen 80A54; 8–10, exterior, interior and lateral views of specimen 80A59 showing the knob and sulcus. Bar 5 1 cm.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

Distribution.—The specimens were collected in the Rutogarea with the other rudists in the Langshan Formation(33u289250N, 79u359250E) (Yang et al., 1982).
Family RADIOLITIDAE d’Orbigny, 1848Genus PRAERADIOLITES Douville, 1902
Type species.—Radiolites fleuriaui d’Orbigny, 1842Key generic characters are: 1) ligament ridge, 2) folded or
wavy growth rings on posterior margin, 3) two ventral radialbands, Ab and Pb, with convex up growth laminae separatedby an interband band, Ib, expressed by concave up growthlamellae, and 4) outer calcite shell layer with rectangular cellsand polygonal cells in radial bands (Douville, 1902; Floquet,1998; Masse et al., 2007).
PRAERADIOLITES HEDINI Douville, 1916
Praeradiolites hedini DOUVILLE, 1916, p. 147, pl. 11, fig. 2–6;KUHN, 1932, p. 126; SANCHEZ, 1981, p. 151; YANG et al.,1982, p. 299, pl.1, figs.1, 5a–c, text-fig. 5; MASSE AND
GALLO MARESCA, 1997, p.102, 107; STEUBER, 2000, p. 161.Praeradiolites cf. hedini DOUVILLE, 1916; DOUVILLE, 1926b,
p. 353, text-fig. 8.Eoradiolites hedini (DOUVILLE, 1916); YANIN, 1989, p. 15;
MASSE ET AL., 2007, p. 706
Type specimens.—Ecole des Mine, Lyon.Diagnosis.—Tall slightly tapering RV/AV ornamented by
longitudinal rounded costae; ligament ridge is planar with atee-shaped to Y-shaped tip; Ab and Pb bands broadly roundedseparated by a shallow fold.
Description.—The RV/AV is elongate, cylindrical to slightlyconical, up to 12–15 cm tall and 4–5 cm in diameter; thetransverse section is elliptical; the concentric growth lamellaeof the outer calcite wall are inclined about 30 degrees to thelong axis of the RV and 15–20 mm apart; the outer shell isornamented by rounded longitudinal costae up to 5 mm wide;the inner aragonite shell layer is thin dorsally and laterally,thickening at the ventral Ab and Pb bands. The anterior bandis a higher amplitude fold than the posterior band; they areseparated by a broad shallow fold. The ligament ridge isnarrow, flat dorsally and expands interiorly in a tee- or Y-shape. The LV/FV is unknown.
Discussion.—The planar tee-shaped or Y-shaped ligamentridge and the low rounded longitudinal costae differentiate P.hedini from other species.
Distribution.—This species ranges from the Albian throughthe Cenomanian and is a member of the Southwest Asianfauna (Masse and Gallo Maresca, 1997) in northwestern Indiaand Tibet in the region around Rutog in the LangshanFormation.
Genus EORADIOLITES
Type species.—Eoradiolites davidsoni Adkins, 1928,
Eoradiolites gilgitensis (DOUVILLE, 1926b)Figure 9.1–9.3
Praeradiolites gilgitensis DOUVILLE, 1926b, p. 353, pl. 13, figs.4a, 4b, text-figs. 7, 8; PUDSEY ET AL., 1985, p. 160–161, fig.5.
Eoradiolites gilgitensis DOUVILLE in MATHUR AND VOGEL,1988, p. 698–699, pl. 1, figs. 1–5, pl. 2, figs., 1–5, pl. 3, figs.1–3; provides previous synonomy; MASSE AND GALLO
MARESCA, 1997a, p. 102; STEUBER, 2000, p. 81.Praeradiolites ngariensis YANG, NIE, WU, AND LIANG, 1982,
p. 299, pl. 1, figs. 2, 3; PUDSEY ET AL., 1985, p. 161.
Type specimens.—Praeradiolites gilgitensis types are atEcole des Mines, Lyon; types of Praeradiolites ngariensis80A007, 80A008, are at University Museum, China Universityof Geosciences Beijing.
Diagnosis.—RV/AV subcyclindrical to ovate, tall, orna-mented by evenly spaced, narrow costae, each divided by thinshallow striae; body cavity ovate; ligament ridge changestowards commissure from planar to bulbous; radial bandswide and slightly expressed.
Original Description of P. ngarensis.—Shell cylindrical-conical, with outer shell layer composed of closely spacedfunnel plates, which are longitudinally folded and furrowed;ligament ridge typically bulb-shaped in cross section. Theinner layer is about 1–3 mm thick. In transverse section [of theRV-AV] there is one tooth and two sockets. E and S bands areindistinct on the outer shell (translation).
Description of P. ngarensis.—RV-AV subcyclindrical, tallgradually expanding cone, ornamented by evenly spaced,narrow costae, each divided by thin shallow striae. Internalvalve cross section ovate, longer dimension either dorsal-ventral or anterior-posterior; outer wall dark gray, composedof radial and concentric laminae, one to two times thicker thaninner spar wall. Ligament ridge a wide plate extending fromouter wall layer with a bulbous tip. Central tooth on RV-AV anarrow, curved spar plate projecting posteriorly at about a 45uangle to dorsal margin. RV-AV myophore is a narrow plate.LV-FV cardinal platform a wide horseshoe-shaped plate,anterior and posterior myophore plates wide ovate, embayedaround ligament; distally the anterior bar fades into triangularanterior tooth; posterior tongue-shaped tooth extends frommyophore plate inserted between central tooth of RV-AV andligament ridge; myophore plate has hollow spaces in thickestpart. The ventral exterior margin is eroded so that the Ab andPb bands cannot be seen.
Discussion.—Based on known structures of these twospecies, it is reasonable to place them in synonomy in thegenus Eoradiolites. Praeradiolites gilgitensis clearly possess theanterior and posterior bands (Douville, 1926b, figs. 7, 8)characteristic of Eoradiolites as described by Masse et al.(2007). These bands parallel the valve length and deflect theexterior growth lamellae slightly; in transverse cross sectionsthe concentric lamellae are folded outwards. The microstruc-ture of the outer shell wall is composed of concentric lamellaewith polygonal cells (Pusdey et al., 1985) in contrast to thequadrangular cells of Eoradiolites davidsoni. However Masseet al. (2007) included both microstructures in Eoradiolites. Theouter shell wall of Praeradiolites ngariensis is like that of E.gilgitensis. The radial bands of Praeradiolites ngariensis areincompletely preserved but suggest the out-folded character(Figs. 9.1–9.3) described by Masse et al. (2007). So the latterspecies is tentatively assigned to the genus Eoradiolites. It canbe considered to be a junior synonym of E. gilgitenesis basedon the right valve outline, the configuration of the myocar-dinal structure, and the ornament.
Masse et al., 2007 suggested that Praeradiolites griegsbachibe assigned to the genus Eoradiolites based on the flatposterior band. Eoradiolites griegsbachi is close to E.gilgitensis in its broadly rounded outline (Douville, 1926a)and occurrence in Himalayan Cretaceous strata (Masse andGallo Maresca, 1997). The large myocardinal structures ofthese three sets of specimens are similar to each other and to E.davidsoni, taking into account that the transverse sections ofeach appear to be at different distances below the commissureplane. The ligament ridge of the three Asian taxa projects intothe body cavity and expands towards the commissure as a
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:24 12 Cust # 09-137
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:25 13 Cust # 09-137
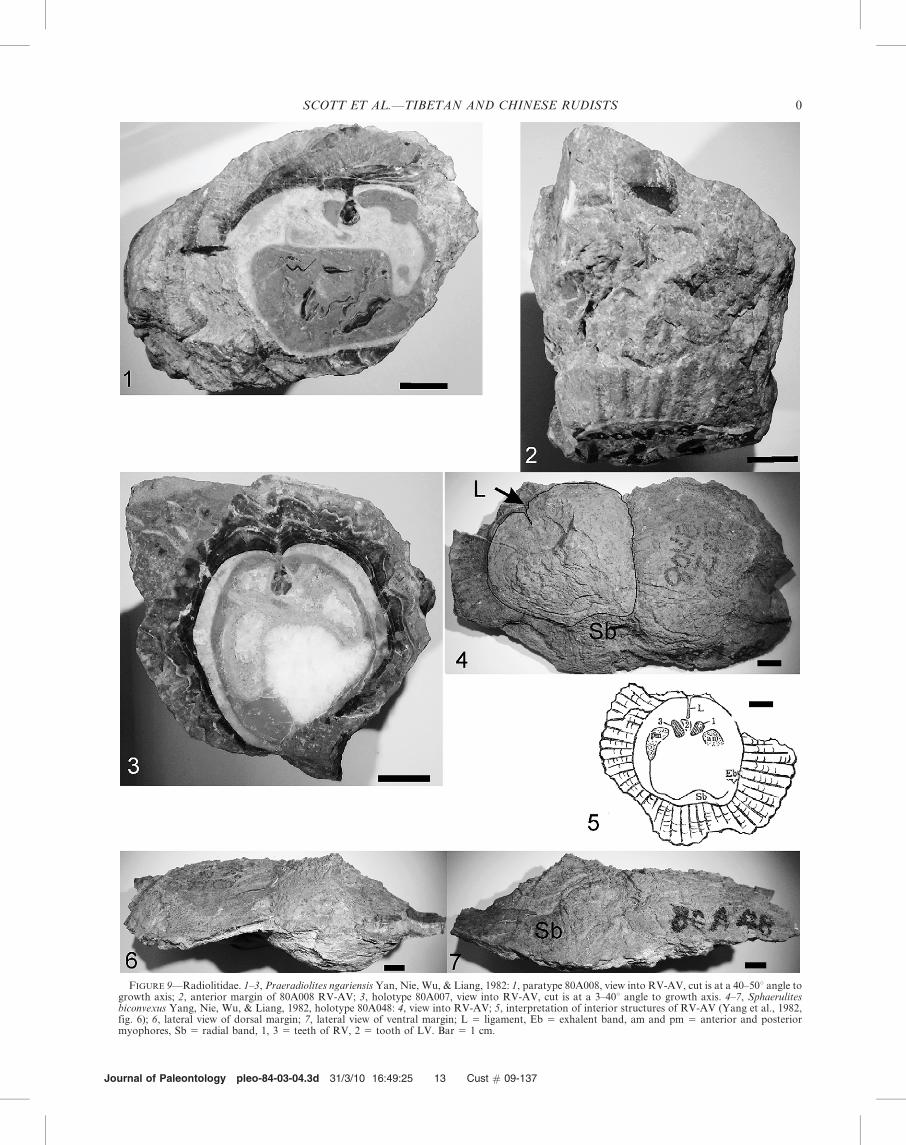
FIGURE 9—Radiolitidae. 1–3, Praeradiolites ngariensis Yan, Nie, Wu, & Liang, 1982: 1, paratype 80A008, view into RV-AV, cut is at a 40–50u angle togrowth axis; 2, anterior margin of 80A008 RV-AV; 3, holotype 80A007, view into RV-AV, cut is at a 3–40u angle to growth axis. 4–7, Sphaerulitesbiconvexus Yang, Nie, Wu, & Liang, 1982, holotype 80A048: 4, view into RV-AV; 5, interpretation of interior structures of RV-AV (Yang et al., 1982,fig. 6); 6, lateral view of dorsal margin; 7, lateral view of ventral margin; L 5 ligament, Eb 5 exhalent band, am and pm 5 anterior and posteriormyophores, Sb 5 radial band, 1, 3 5 teeth of RV, 2 5 tooth of LV. Bar 5 1 cm.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

bulb. The ligament of E. ngariensis is composed of darkcompact outer shell material and not a secondary deposit assuggested by Pudsey et al. (1985; Mathur and Vogel, 1988).However the ligament ridge of E. davidsoni is short andnarrow.
The cardinal tooth of the RV-AV of P. gilgitensis is long,narrow and hooked; that of ngariensis is also long and narrowand slightly curved, and that of griesbachi is short and narrow.The anterior myophores of the RV of all three species arebroad v-shaped extensions of the inner shell layer. Theposterior myophores of gilgitensis and ngariensis are shortrounded projections from the inner valve wall; greigsbachiappears to have no such projection.
The myocardinal structures of the LV-FV of gilgitensis,ngariensis, and griesgbachi are large arcuate to horseshoe-shaped plates that project into the RV similar to that of E.davidsoni (Masse et al., 2007, fig. 3A2). The anterior teeth ofgilgitensis and ngariensis are large, ovate and the posteriorteeth are rounded. The anterior tooth of griegsbachi is v-shaped and the posterior tooth is rectangular and bifid. TheLV myophores of ngariensis are larger than those ofgilgitentsis and are perforated by large cavities; this could bea function of the position of the transverse cuts.
The four new radiolitid species created by Gou and Shi(1998) possess radial bands like those of Eoradiolites and aredistinguished by features of the ligament ridge and ornament.Praeradiolites gegyainensis Gou and Shi (1998) has a flattenedligament ridge and a thicker inner shell layer than E.ngariensis. Praeradiolites daxungensis Gou and Shi (1998)has a short, quadrate ligament ridge. The ligament ridge ofPraeradiolites bangoinensis Gou and Shi (1998) is described as‘‘…distinct, strong, short (about 5 mm in length), and wedgeshaped.’’ Praeradiolites perbellus Gou and Shi (1998) has avery thin ligament ridge. Praeradiolites coquenensis Gou andShi (1998) has a very short, wide ligament ridge. Serial sectionsthrough successive ontogenetic stages are needed to under-stand the growth stages of these shells. These species may bejunior synonyms of E. gilgitensis. For example the tip of theligament ridge of Praeradiolites ciryi Floquet (1991) variesconsiderably from rounded to blunt to bifurcate depending onits preservation (Floquet, 1998).
Distribution.—The type locality of E. gilgitensis is nearGilgit in the Yasin area of northwestern Pakistan (Douville,1926b, gave these coordinates: 73u209E and 36u209N;locality S2(iii) of Pudsey et al., 1985). The upper Aptian-lower Albian age of the Gilgit, Yasin outcrop is constrainedby Orbitolina (Mesorbitolina) texana (Roemer) (Pudseyet al., 1985). The type locality of E. ngariensis is near Lameila,Ritu County in the Albian Langshan Formation with O.texana, Orbitolina (Mesorbitolina) pervia, Praeradioliteshedini, and Sphaerulites biconvexus. The five Praeradiolitesspecies of Gou and Shi (1998) range throughout the LangshanFormation.
The type locality of S. griesbachi is near Herat, Afganistan(Douville, 1926a) and is not well constrained by age-diagnosticfossils.
Genus SPHAERULITES Lamarck, 1819
Type speciesSphaerulites foliaceus Lamarck, 1819.
SPHAERULITES BICONVEXUS (Yang, Nie, Wu, and Liang, 1982)Figure 9.4–9.7
Praeradiolites biconvexus YANG, NIE, WU, AND LIANG, 1982,p. 299, Pl. 1, figs. 4a–c.
Type specimen.—80A048, University Museum, China Uni-versity of Geosciences Beijing.
Diagnosis.—RV-AV low subconical, wider than high,external prismatic shell layer very wide, flaring; ligament long,narrow; no accessory cavities. Radial bands broadly rounded.
Original Description (translation).—Shell large (diameter ofbody cavity greater than 55 mm), the outer shell layer extends45 mm; total length of the specimen is 55 mm, shell sub-biconvex. The outer shell layer of the FV is about 45 mmthick, marked by closely spaced funnel plates (intersectionangle about 80u between outer and inner shell layers); lamellaeclose to inner layer inclined and become subhorizontaloutwards. The specimen has two major growth rings eachabout 3 mm thick. Ligament ridge narrow, about 8.5 mm long;appearing to join with cardinal tooth, and inserted betweentwo teeth of FV. Muscle insertion plates broad.
Description.—RV/AV low, very wide, anterior-posteriordimension 14.5 cm, dorsal-ventral dimension 8.0 cm; convexbase, commissure surface gently convex; mantle margins verywide from 1.5 to 7.0 cm, thick outer prismatic shell layer. Bodycavity excentrically ovate, anterior-posterior dimension longerthan dorsal-ventral, 6.0 cm by 5.0 cm; anterior margin evenlycurved, antero-ventral margin straight merging into shallowsulcus of E radial band, postero-ventral corner narrow,abrupt, posterior margin straight and inclined posteriorly,dorsal margin broadly curved. Outer wall layer with radial andconcentric prisms, wide anteriorly, extremely extended poste-riorly; inner spar layer very thin. Ligament ridge a thin plate0.85 cm long extending from invagination of interior marginof outer wall layer, radial bands are gentle swellings on innersparry wall layer. Anterior myophore is kidney-shaped shelfon the inner wall. Teeth of RV-AV and LV-FV are ovate withlong direction dorsal-anterior direction.
Discussion.—The morphology of this species meets all thecharacteristics of the genus Sphaerulites. The RV-AV ofSphaerulites Lamarck (1819) is flattened and wider than high;it is characterized by highly foliaceous growth lamellae,ventral bands expressed as undulations or subtle folds in thegrowth lamellae and internally as embayments in the valvemargin, ligament ridge well developed in older forms; LV-FVconvex, rarely planar; cardinal apparatus similar to that ofPraeradiolites (Toucas, 1907; Dechaseaux and Coogan, 1969).
Sphaerulites biconvexus differs from Sphaerulites cantabricusDouville (1889), the Albian species of northern Iberia, by itswider anterior-posterior expansion of the outer prismatic layerand by the absence of accessory cavities. S. biconvexus alsodiffers from the French Cenomanian Sphaerulites foliaceusLamarck (1819) by its thinner inner shell layer, narrowligament, and by the absence of accessory cavities on eitherside of the ligament.
Distribution.—Type locality is in Jiagang, Ritu County andwas collected from the Albian Langshan Formation withMesorbitolina texana, Mesorbitolina pervia, Praeradioliteshedini, and Praeradiolites ngariensis.
CONCLUSIONS
Rudists are a principal biotic component of Cretaceouscarbonates in Tibet and in the Western Tarim Basin.Barremian to Maastrichtian carbonate units are widespreadon the northern margin of the Indian Plate and in Tethyantectonic slices. In northwestern Tibet, Barremian-Cenomanianendemic rudists, corals, stromatoporoids, and cosmopolitanorbitolinid foraminifera built bioherms on carbonate plat-forms in the eastern Tethys. The endemic Requieniid rudistRutonia is compared to morphologically similar but older, less
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:27 14 Cust # 09-137
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010

derived genera. Associated specimens in this assemblage aremonopleurids and two genera with three radiolitid species thatare re-described and taxonomic positions re-evaluated. Insouthern Tibet Campanian-Maastrichtian mainly endemicradiolitid rudists and cosmopolitan larger benthic foraminif-era contributed to carbonate shelves on the northern IndianPlate near the Cretaceous equator. In the Western TarimBasin Cenomanian strata yield Tethyan rudist species.
Three-dimensional morphometric analysis shows that Re-quieniidae valves are convergent with the basic gastropodshell. More derived strongly coiled, younger requieniids wereadapted to encrusting or semi-infaunal habits. Stratigraphicanalysis confirms that Requieniidae diversity crises coincidedwith Cretaceous oceanic anoxic events
Two end members of valve geometry and internal mor-phology appear to be primitive and derived charactersrespectively and separate the Family Requieniidae into twoclades that are here recognized as two new subfamilies. Theend members are defined by the coiling geometry, whether thespire is close to the plane of commissure or it is translatedalong the coiling axis and by myophore structures. The oldermatheroniform clade has a low spirogyrate LV that istranslated slightly from the commissure along the coiling axis;this group is composed of Matheronia (and its subgenusMonnieria), Hypelasma, Lovetchenia, Rutonia, and Kugleria.Genera in the younger clade have a tall trochospiral LV that istranslated along the coiling axis and consists of Requienia,Toucasia, Pseudotoucasia, Apricardia, Bayleoidea, and Bayleia.Claditics support these relationships.
ACKNOWLEDGMENTS
The initial work was done under the auspices of theNational Project 973 (Project NO. 2006CB701403) of China,the National Natural Science Foundation of China (ProjectNO. 40672020). The authors also thank the financial supportof the National Science Foundation of China (40872013). Thefirst author was supported as a visiting professor by ChinaUniversity of Geosciences Beijing (CUGB). The Director ofthe Paleontology Museum of the China University ofGeosciences Beijing permitted access to study the specimensin the museum office. Professor Zetong Nie kindly describedoutcrop conditions and the rudist bioherms. Qiang Ou,CUGB, provided able support for photography. Translationsof Chinese to English were made by Lei Qian Ping and JieWang of the CUGB. Jean-Pierre Masse and Peter W. Skeltonreviewed the range data of Requieniid genera and sharedobservations of requieniid morphology. The bibliography ofrudist taxa by Steuber and Loser (1996) has been extremelyhelpful. Steve Westrop generously ran our data matrix on thePAUP computer program to produce two cladograms.
REFERENCES
ADKINS, W. S. 1930. New rudistids from the Texas and MexicanCretaceous. Bureau of Economic Geology, University of Texas Bulletin3001:77–137.
ASTRE, G. 1954. Radiolitides nord-pyreneens. Societe geologique deFrance, Memoires, 36:1–130.
BOBKOVA, N. N. 1961. Stratigraphy and mollusc lamellibranchiata of theUpper Cretaceous in the Tajik depression. Trudy vsesojuznogonauchno-issledovatel’skogo geologicheskogo Instituta (VSEGEI),54:1–190. (in Russian)
BOUWMAN, L. A. H. 1938. Sur un genre nouveau (Kugleria) de la familiedes requienides pachyodontes. Akademie Nederland Wetenschaft,Proceedings, 41:418–429.
COX, L. R. 1933. The evolutionary history of the rudists with a report ofthe demonstration at the British Museum (Natural History), onSaturday, February 17th, 1933. Proceedings of the Geologist’sAssociation, 44:379–388.
D’ARCHIAC, E. J. A. 1837. Memoire sur la formacion cretace du sud-ouestde la France. Societe geologique de France, Memoire No. 2(7):157–193.
DECHASEAUX, C. AND A. H. COOGAN. 1969. Family Radiolitidae Gray,1848, p. N803–N817. In L. R. Cox et al.. (eds.), Treatise on InvertebratePaleontology. Pt. N. Mollusca 6. Bivalvia, Vol. 2. Geological Society ofAmerican and University of Kansas Press, Lawrence.
DECHASEAUX, C. AND B. F. PERKINS. 1969. Family RequieniidaeDouville, 1914, p. N779–N781. In L. R. Cox et al.. (eds.), Treatise onInvertebrate Paleontology. Pt. N. Mollusca 6. Bivalvia, Vol. 2.Geological Society of American and University of Kansas Press,Lawrence.
DES MOULINS, C. 1826. Essai sur les Spherulites qui existent dans lescollections de MM. F. Jouannet, membre de l’Academie royal desSciences, belle Lettres et Arts de Bordeaux, et Charles Des Moulins.Bulletin d’Histoire naturelle de la Societe Linneenne de Bordeaux,1:148–303.
DOUGLASS, R. C. 1960. The foraminiferal genus Orbitolina in NorthAmerica. U.S. Geological Survey Professional Paper 333, 52 p.
DOUVILLE, H. 1889. Sur quelques rudistes du terrain cretace inferieur desPyrenees. Bulletin de la Societe geologique de France, Series 3, 17:627–655.
DOUVILLE, H. 1902. Classification des Radiolites. Bulletin de la Societegeologique de France, Series 4, 2:461–477.
DOUVILLE, H. 1910. Etudes sur les rudistes de Sicile, d’Algeria, d’Egypt,du Liban et de la Perse. Societe geologique de France, Memoire No. 41,p. 1–84.
DOUVILLE, H. 1911. Observations sur les ostreidea. Origine et classifica-tion. Societe geologique de France, Bulletin, Series 4, 10:635–646.
DOUVILLE, H. 1914. Les Requienides et leur evolution. Bulletin de laSociete geologique de France, Series 4, 14:380–383.
DOUVILLE, H. 1916. Les calcaires a orbitolines et a Radiolites du Thibet.In: S. Hedin (ed.), Southern Tibet. Discoveries in former timescompared with my own researches in 1906–1908, 5:145–147.
DOUVILLE, H. 1917. Les terrains cretaces de l’Asie occidentale. Compterendu sommaire des Seances de la Societe geologique de France, p. 121–122.
DOUVILLE, H. 1918. Le Barremien superieur de Brouzet. Troiseme Partie:Les Rudistes. Societe geologique de France, Memoires No. 52, p. 5–19.
DOUVILLE, H. 1926a. Description de quelques fossiles Cretaces del’Afganistan. Records of the Geological Survey of India, 58, part4:345–348.
DOUVILLE, H. 1926b. Fossiles recueillis par Hayden dans le Kashmir en1906 et les Pamirs en 1914; leur description. Records of the GeologicalSurvey of India, 58, part 4:349–357.
ERMAN, A. 1954. Einige Beobachtungen uber die Kreideformation an derNordkuste van Spanien. Zeitschrift deutsche geologie Gesellschaft,6:596–611.
JOUKOWSKY, E. AND J. FAVRE. 1913. Monographie geologique etpaleontologique du Saleve (Haute Savoie, France). Memoire de laSociete de Physique et d’Histoire naturelle de Geneve, 37:295–523.
FLOQUET, M. 1991. La plate-forme nord-castillane au Cretace superieur.Arriere-pays iberique de la marge passive basco-cantabrique. Sedimen-tation et vie. Memoires Geologiques de l’Universite de Dijon, 14:1–925.
FLOQUET, M. 1998. Praeradiolites ciryi, a polymorphic rudist from upperSantonian and Campanian carbonate formations of the Castilian ramp(Northern Spain). Geobios, Memoir special, 22:111–123.
FOREY, P. 2009. Cladistics for palaeontologists. The PalaeontologicalAssociation Newsletters, 70 April 2009 No. 60-63.
GARCIA-BARRERA, P. 1995. Toucasia hancockensis (Hippuritacea-Requie-niidae) in southwestern Mexico. Revista Mexicana de CienciasGeologicas, 12:191–194.
GOLDFUSS, A. 1837. Petrefacta Germaniae. Arnz, Dusseldorf, Part 2c, lief6, 141–224.
GOLDFUSS, A. 1840. Petrefacta Germaniae. Arnz, Dusseldorf, Part 2d, lief7, 225–312.
GOLOBOFF, P., J. FARRIS, AND K. NIXON. 2008. TNT: a free program forphylogenetic analysis. Cladistics 24:774–786.
GOLONKA, JAN. 2002. Plate-tectonic maps of the Phanerozoic, p. 21–75. InW. Kiessling, E. Flugel, and J. Golonka (eds.), Phanerozoic ReefPatterns. SEPM (Society for Sedimentary Geology) Special Publication72.
GOU, Z. 1987. A Cretaceous bivalve fauna in Gamba area, Tibet.Geological Contributions, Qinghai-Xizang Plateau, 18, p. 146–166.Geological Publishing House, Beijing.
GOU, Z. AND H. SHI. 1998. Rudists (Bivalvia) from the Cretaceous ofTibet, China, with descriptions of new species, p. 255–266. In P. A.Johnston and J. W. Haggart (eds.), Bivalves: An Eon of Evolution–Paleobiological Studies Honoring Norman D. Newell. University ofCalgary Press, Calgary.
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:28 15 Cust # 09-137
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

GOURRAT, C., J.-P. MASSE, AND P. W. SKELTON. 2003. Hypelasmasalevensis (Favre, 1913) from the Upper Kimmeridgian of the FrenchJura, and the origin of the rudist Family Requieniidae. GeologicaCroatica, 56:139–148.
GRAS, A. 1854. Catalogue des corps organises fossiles qui se recontrentdans le Departement de l’Isere. Bulletin de la Societe de Statistique, desSciences naturelles et des Arts industrielles du Deoartement de l’Isere,(2), 2:1–54, Grenoble.
GRAY, J. E. 1848. On the arrangement of the Brachiopoda. Annals andMagazine of natural History, 2:435–440.
GUERANGER, E. 1853. Essai d’un repertoire paleontologique de Depart-ment de la Sarthe. Le Mans.
HENSON, F. R. S. 1948. Larger imperforate Foraminifera of south-westAsia. Families Lituolidae, Orbitolinidae and Meandropsinideea. BritishMuseum (Natural History), London, 127 p., 16 pl., 16 figs.
HUANG, J. AND B. CHEN. 1987. The evolution of the Tethys in China andadjacent region. Geological Publishing House, Beijing, 109 p.
HUANG, S., H. SHI, L. SHEN, M. ZHANG, AND W. WU. 2005. Globalcorrelation for strontium isotope curve in the Late Cretaceous of Tibetand dating marine sediments. Science in China Ser. D, Earth Sciences,48:199–209.
KAUFFMAN, E. G. 1973. Cretaceous Bivalvia, p. 353–383. In A. Hallam(ed.), Atlas of Palaeobiogeography. Elsevier Scientific PublishingCompany, Amsterdam.
KUHN, O. 1932. Fossilium Catalogus, I. Animalia, Pars 54, Rudistae.Gustav Feller, Neubrandenburg 200 p.
KUTASSY, A. 1934. Pachydonta mesozoica (Rudistis exlusis). FossiliumCatalogus, I. Animalia, Pars 68, Gustave Feller, Neubrandenburg, 202pp.
LAN, X. AND J. WEI. 1995. Late Cretaceous-Early Tertiary marine bivalvefauna from the western Tarim Basin. Science Press, 212 p., 70 plates.(Chinese with English summary)
LAPEIROUSE, P. DE. 1781. Description de plusieurs nouvelles especesd’Orthoceratites et d’Ostracites. Walther, Erlangen, 1–48 p.
LI, T. AND X. XIOA. 1995. Tectonic evolution and uplift of the Qinghai-Tibet Plateau. Episodes, 18:31–35.
LI, X. AND J. A. GRANT-MACKIE. 1994. New Middle Jurassic-LowerCretaceous bivalves from southern Tibet. Journal of Southeast AsianEarth Sciences, 9:263–276.
MAINELLI, M. 1995. Apricardia manuelae n. sp. in the Lower Turonian ofnortheastern Matese (South Apennines), Italy. Revista Mexicana deCiencias Geologicas, 12:195–200.
MALCHUS, N. 1995. The meaning of ‘‘inversion’’ in chamids and rudists(Bivalvia) reviewed and an unbiased theoretical approach to LateJurassic-Early Cretaceous rudist phylogeny. Revista Mexicana daCiencia Geologicas, 12:211–223.
MALCHUS, N. 1998. Aptian (Lower Cretaceous) rudist bivalves from NESpain: taxonomic problems and preliminary results. Geobios, Memoirespecial, 22:181–191.
MASSE, J.-P. 1976. Les calcaires Urgoniens de Provence, Valanginien-Aptien Inferieur. Stratigraphie, paleontologie, les paleoenvironments etleur evolution. These, Universite d’Aix-Marseille II, U.E.R. desSciences de la Mer et de l’environment, Marseille. 510 p. (C.N.R.S.no. AO 12390).
MASSE, J.-P. 1988a. Importance relative, chronologie et significationphylogenetique des modifications morphologiques et anatomiques chezles Requieniidae (Rudistes) du Cretace inferieur. Serbian GeologicalSociety, 1st International Conference on Rudists, Abstracts, Belgrade, p.15.
MASSE, J.-P. 1988b. Paleontologie, paleobiologie et paleoecologie deRequienia ammonia (Goldfuss) (Rudiste, Requieniidae). Bulletin de laSociete d’Etude Science naturel de Vaucluse, 1988:111–119.
MASSE, J.-P. 1993. Systematique, stratigraphie et paleobiogeographie dugenre Lovetchenia (Requieniidae) du Cretace inferieur Mediterraneen.Geobios, 26:699–708.
MASSE, J.-P. 1994. L’evolution des Requieniidae (rudistes) du Cretaceinferieur: caracteres, signification fonctionnelle, adaptative et relationsavec les modifications des paleoenvironnements. Geobios, 27:321–333.
MASSE, J.-P. 2002. Importance relative, chronologie et significationphylogenetique des modifications morphologiques et anatomiques chezles Requieniideae (Rudistes) du Cretace inferieur, p. 155–171. In M.Sladic-Trifunovic (ed.), Proceedings First International Conference onRudists–Belgrad, 1988–‘‘Rudists.’’ Union of Geological Societies ofYugoslavia, Memorial Publication.
MASSE, J.-P., M. FENERCI-MASSE, L. VILAS, AND C. ARIAS. 2007. LateAptian-Albian primitive Radiolitidae (bivavles, hippuritoidea) fromSpain and SW France. Cretaceous Research, 28:697–718.
MASSE, J.-P., AND M. GALLO MARESCA. 1997. Late Aptian Radiolitidae(rudist bivalves) from the Mediterranean and Southwest Asiatic regions:
taxonomic, biostratigraphic and palaeobiogeographic aspects. Palaeo-geography, Palaeoclimatology, Palaeoecology, 128:101–110.
MASSE, J.-P., M. GALLO MARESCA, AND E. LUPERTO sINNI. 1997. Albianrudist faunas from southern Italy: Taxonomic, biostratigraphic andpalaeobiogeographic aspects. Geobios, 31:47–59.
MASSE, J.-P., C. GOURRAT, D. ORBETTE, AND D. SCHMUCK. 1998.Hauterivian rudist faunas of southern Jura (France). Geobios, Memoirespecial, 22:225–233.
MASSE, J.-P. AND J. PHILIP. 1996. L’Evolution des rudistes au regard desprincipaux evenements geologiques du Cretace. Bulletin des CentresRecherches Exploration-Production Elf-Aquitaine, 10:437–456.
MASSE, J.-P., J. PHILIP, AND G. CAMOIN. 1995. The Cretaceous Tethys, p.215–236. In A. E. M. Nairn, L. E. Ricou, B. Vrielynck, and J. Dercourt(eds.), The Ocean Basins and Margins, Vol. 8: The Tethys Ocean.Plenum Press, New York.
MATHERON, P. 1843. Catalogue methodique et descriptif des corpsorganises fossiles du Departement des Bouches-du-Rhone et lieuxcirconvoisins. Carnaud Fils, Marseille, 1–269 p.
MONTENAT, C., J.-P. MASSE, AND J. PHILIP. (1982) - Le Cretac6 inferieur.Orbitolines et Rudistes d’Afghanistan central. Geol. Medit., Marsellle,9, 2:109–122.
MUNIER-CHALMAS, H. 1873. Prodrome d’une classification des rudistes.Journal de Conchyliologie, 13:71–75.
NEWELL, N. D. 1965. Classification of the Bivalvia: American Museum ofNatural History, Novitates, 2206.
NIXON, K. C., 2002. WinClada ver. 1.00.08. Published by the author,Ithaca, NY, USA.
D’ORBIGNY, A. 1842. Quelques considerations zoologiques et geologiquessur les rudistes. Bulletin de la Societe geologique de France, 17:148–163.
PALMER, R. H. 1928. The rudistids of southern Mexico. OccasionalPapers of the California Academy of Sciences, Vol. 14, p.
PAQUIER, V. 1898. Sur quelques diceratines nouveaux du Tithonique.Societe geologique de France, Bulletin, Series 3, 25:843–851.
PAQUIER, V. 1903. Les rudistes urgoniens. Premiere partie. MemoiresSociete geologique de France, Paleontologie, 29:1–46.
PARONA, C. F. 1921. Fauna del neocretacico della Tripolitania. Memoireper servire alla Descrizione della Carta geologica d’Italia, 8:1–21.
POJARKOVA, Z. N. 1955. Some rudists from Upper Cretaceous formationsin the Zeravsham and Turkestan mountains. Academic ProceedingsLGU no. 189, geological series, Academy of Sciences, 6:27–53. (inRussian)
POJARKOVA, Z. N. 1984. The Cenomanian and Turonian in northeasternAsia. Cretaceous Research, 5:1–14.
POLSAK, A. 1967. Macrofaune cretacee de l’Istrie meridionale (Yougosla-vie). Palaeontologica jugoslavica, 8:1–219.
PUDSEY, C. J., R. SCHROEDER, AND P. W. SKELTON. 1985. Cretaceous(Aptian/Albian) age for island-arc volcanics, Kohistan, N. Pakistan, p.150–168. In V. J. Gupta (ed.), Contributions to Himalayan Geology, v.3, Geology of western Himalayas. Hindustan Publishing Corporation,Delhi.
QIAN, DINGYU. 1993. The distribution and biogeographic provincializa-tion of Cretaceous rudists in Xizang (Tibet). Tibet Geology, 2:19–25.(Chinese with English abstract)
RAUP, D. R. 1966. Geometric analysis of shell coiling: General problems.Journal of Paleontology, 40:1178–1190.
RAUP, D. AND S. M. STANLEY. 1972. Principles of Paleontology. W. H.Freeman and Company, San Francisco.
ROEMER, F. 1849. Texas. Adolph Marcus, Bonn, 464 p.ROSS, D. J. AND P. W. SKELTON. 1993. Rudist formations of the
Cretaceous: a palaeoecological, sedimentological and stratigraphicalreview, p. 73–91. In V. P. Wright, (ed.), Sedimentology Review/1:Blackwell Scientific Publications.
SANCHEZ, V. 1981. Hippuritidae y Radiolitidae (Bivalvia). Catalogo deespecies. Universidad Autonoma de Barcelona, Publicaciones deGeologıa 15, 228 p.
SANDERS, D., M. LUKESCH, M. RASSER AND P. W. SKELTON. 2007. Shellbeds of diceratid rudists ahead of a low-energy gravelly beach(Tithonian, Northern Calcareous Alps, Austria): Palaeoecology andtaphonomy. Austrian Journal of Earth Sciences, 100:186–199.
SCHNARRENBERGER, C. 1901. Ueber die Kreideformation der Monted’Ocre-Kettein den aquilaner Abruzzen. Naturforschung Gesellschaft,Freiburg, Berichte, 11:176–214.
SCOTT, R. W. 1981. Biotic relations in Early Cretaceous coral-algal-rudistreefs, Arizona. Journal of Paleontology, 55:463–478.
SCOTT, R. W. 1995, Global environmental controls on Cretaceous reefalecosystems. Palaeogeography, Palaeoclimatology, Palaeoecology,119(1–2):187–199.
SCOTT, R. W. AND M. WEAVER. 2008, Ontogeny and functionalmorphology of a Lower Cretaceous caprinid, Comanche Shelf, US
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:28 16 Cust # 09-137
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010

Gulf Coast: Eighth International Congress on Rudists, June 23–25,2008, Dokuz Eylul University, Izmir, Turkey, p. 37.
SCOTT, R. W. AND M. WEAVER. In press. Ontogeny and functionalmorphology of a Lower Cretaceous caprinid rudist (Bivalvia, hippur-itoida). Turkish Journal of Earth Sciences.
SHA, J., P. L. SMITH, AND F. T. FURSICH. 2002. Jurassic Ostreoida(Bivalvia) from China (Tanggula Mountains, Qinghai-Xizang Plateau)and their paleobiogeographic content. Journal of Paleontology, 76:431–446.
SHA, J., A. L. A. JOHNSON, AND F. T. FURSICH. 2004. From deep-sea tohigh mountain ranges: palaeogeographic and biotic changes in Hohxil,the source area of the Yangtze River (Tibet Plateau) since the LatePalaeozoic. Neues Jahrbuch fur Geologie und Palaontologie, 233:169–195.
SIMMONS, M. D., J. E. WHITTAKER, AND R. W. JONES. 2000. Orbitolinidsfrom Cretaceous sediments of the Middle East–a revision of the F. R. S.Henson and Associates collection, p. 411–437. In M. B. Hart, M. A.Kaminski, and C. W. Smart (eds.), Proceedings of the FifthInternational Workshop on Agglutinated Foraminifera. GrzybowskiFoundation Special Publication 7.
SKELTON, P. W. 1978. The evolution of functional design in rudists(Hippuritacea) and its taxonomic implications. Philosophical Transac-tions, Royal Society London, B. 284:305–318.
SKELTON, P. W. 1985. Preadaptation and evolutionary innovation inrudist bivalves. Palaeontology, Special Papers, 33:159–173.
SKELTON, P. W. 1991. Morphogenetic versus environmental cues foradaptive radiations, p. 375–386. In N. Schmidt-Kittler and K. Vogel(eds.), Constructional Morphology and Evolution. Springer-Verlag,Berlin.
SKELTON, P. W. 1999. Synoptic guide to Kimmeridgian rudists for theKelheim field visit, p. 83–89. In R. Hofling and T. Steuber (eds.), FifthInternational Congress on Rudists, Abstracts and Field Trip Guides.Erlanger geologische Abhandlungen, Sonderband 3, Erlangen.
SKELTON, P. W. 2008. Proposed revisions of rudist bivalve classification.Eighth International Congress on rudists, Abstracts, Dokuz EylulUniversity, Izmir, Turkey, p. 51.
SKELTON, P. W. AND J.-P. MASSE. 2000. Synoptic Guide to LowerCretaceous Rudist Bivalves of Arabia, p. 89–99. In A. S. Alsharhan andR. W. Scott (eds.), Middle East Models of Jurassic/CretaceousCarbonate Systems. SEPM Special Publication 69.
SKELTON, P. W. AND A. B. SMITH. 2000. A preliminary phylogeny forrudist bivalves: sifting clades from grades, p. 97–127. In E. M. Harper, J.D. Taylor and J. A. Crame (eds.), The evolutionary biology of Bivalvia.Geological Society of London, Special Publication 177.
STEUBER, T., 2000. Rudist_Data_base_web_pages.STEUBER, T. AND H. LOSER. 1996. Jurassic-Cretaceous rudists (Mollusca,
Hippuritacea)–Bibliography 1758–1994. Neue Palaontologische Abhan-dlungen, Band 1, 1–123 p.
STEUBER, T. AND H. LOSER. 2000. Species richness and abundancepatterns of Tethyan Cretaceous rudist bivalves (Mollusca: Hippurita-cea) in the central-eastern Mediterranean and Middle East, analysedfrom a palaeontological data base. Palaeogeography, Palaeoclimatol-ogy, Palaeoecology, 162:75–104.
SWOFFORD, D.L. 2000. PAUP*: Phylogenetic Analysis Using Parsimony(* and Other Methods), Version 4. Sinauer Associates, Sunderland,Massachusetts.
TOUCAS, A. 1907. Etudes sur la classification et l’evolution desRadiolitides. Societe geologique de France, Paleontologie, Memoire,36:1–130.
WAN, X., L. F. JANSA, AND M. SARTI. 2002. Cretaceous and Paleogeneboundary strata in southern Tibet and their implication for the India-Eurasia collision. Lethaia, 35:131–146.
WAN, X., Y. WU, AND G. LI. 2003. Distribution of mid-Cretaceousorbitolinids in Xizang (Tibet) and its paleobiogeographic implications.Acta Geological Sinica, 77:1–8. (In Chinese with English abstract)
WANG, CHENGSHAN, EDMUND, Z. CHANG, AND SHAONAN ZHANG. 1997.Potential oil- and gas-bearing basins of the Qinghai-Tibetan Plateau,China. International Geology Review, 39:876–890.
WANG, C. S., X. HU, L. F. JANSA, X. Q. WAN, AND R. TAO. 2001. TheCenomanian-Turonian anoxic event in southern Tibet. CretaceousResearch, 22:481–490.
WANG, C. S., X. LI, X. HU, AND L. F. JANSA. 2002. Latest marine horizonnorth of Qomolangma (Mt. Everest): implications for closure of Tethysseaway and collision tectonics. Terra Nova, 14:114–120.
WEN, S. 1999. Cretaceous bivalve biogeography in Qinghai-XizangPlateau. Acta Palaeontolgica Sinica, 38:1–30.
WEN, S. 2000. Cretaceous bivalves of Kangpa Group, South Xizang,China and their Biogeography. Acta Palaeontologica Sinica, 39:1–27.
WEN, S., X. LAN, J. CHEN, Z. ZHANG, C. CHEN, AND Z. GU. 1976. FossilLamellibranchs from Mt. Qomolangma region: Report of ScientificExpedition to the Mt. Qomolongma region (1966–1968), Palaeontology.Science Press, Beijing, 3, 152 p. (In Chinese with English abstract)
WEN, S., J. SHA, B. ZHANG, AND B. CAI. 2000. Marine Cretaceous, p.315–327. In Nanjing Institute of Geology and Palaeontology, ChineseAcademy of Sciences (ed.) Stratigraphical Studies in China (1979–1999).Press of University of Science and Technology of China, Heifei. (InChinese)
WHITE, C. A. 1884. On Mesozoic fossils. U.S. Geological Survey Bulletin,4:87–125.
WHITNEY, M. 1952. Some new pelecypoda from the Glen Rose Formationof Texas. Journal of Paleontology, 26:697–707.
WILLEMS, H., Z. ZHOU, B. ZHANG, AND K.-U. GRAFE. 1996. Stratigraphyof the Upper Cretaceous and lower Tertiary strata in the TethyanHimalayas of Tibet (Tingri area, China). Geologische Rundschau,85:723–754.
YANG, Z. 1984. Marine facies Bivalvia, Hippuritoida, Palaeontologicalplates of Northwest region, China (III) (Xinjiang). Geology PublishingHouse, Beijing, p. 150–165.
YANG, Z., Z. NIE, S. WU, AND D. LIANG. 1982. Cretaceous rudists fromNgari, Xizang (Tibet), Autonomous Region, China and their geologicsignificance. Acta Geologica Sinica, 56:293–301. (In Chinese withEnglish abstract)
YANIN, B. T. 1989. The Jurassic and Cretaceous rudists: stratigraphicaland geographical distribution. Moscow, Nauka, 214 p., 16 pls.
YOUNGE, C. M. 1967. Form, habit and evolution in the Chamidae(Bivalvia) with reference to conditions in the rudists (Hippuritacea).Philosophical Transactions Royal Society London, Series B, BiologicalSciences, No. 775, 252:49–105.
ZHANG, K. J. 2000. Cretaceous palaeogeography of Tibet and adjacentareas (China): tectonic implications. Cretaceous Research, 21:23–33.
ZLATARSKI, G. 1886. Geologische Untersuchungen im centralem Balkanund in den angrenzenden Gebieten. Sitzungsb. der KoninlicheAkademie der Wissenshaffen Math.-Naturwissenschaft, Vol. XCIII, IAbth., 1:249–340.
ACCEPTED 18 JANUARY 2010
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:29 17 Cust # 09-137
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0
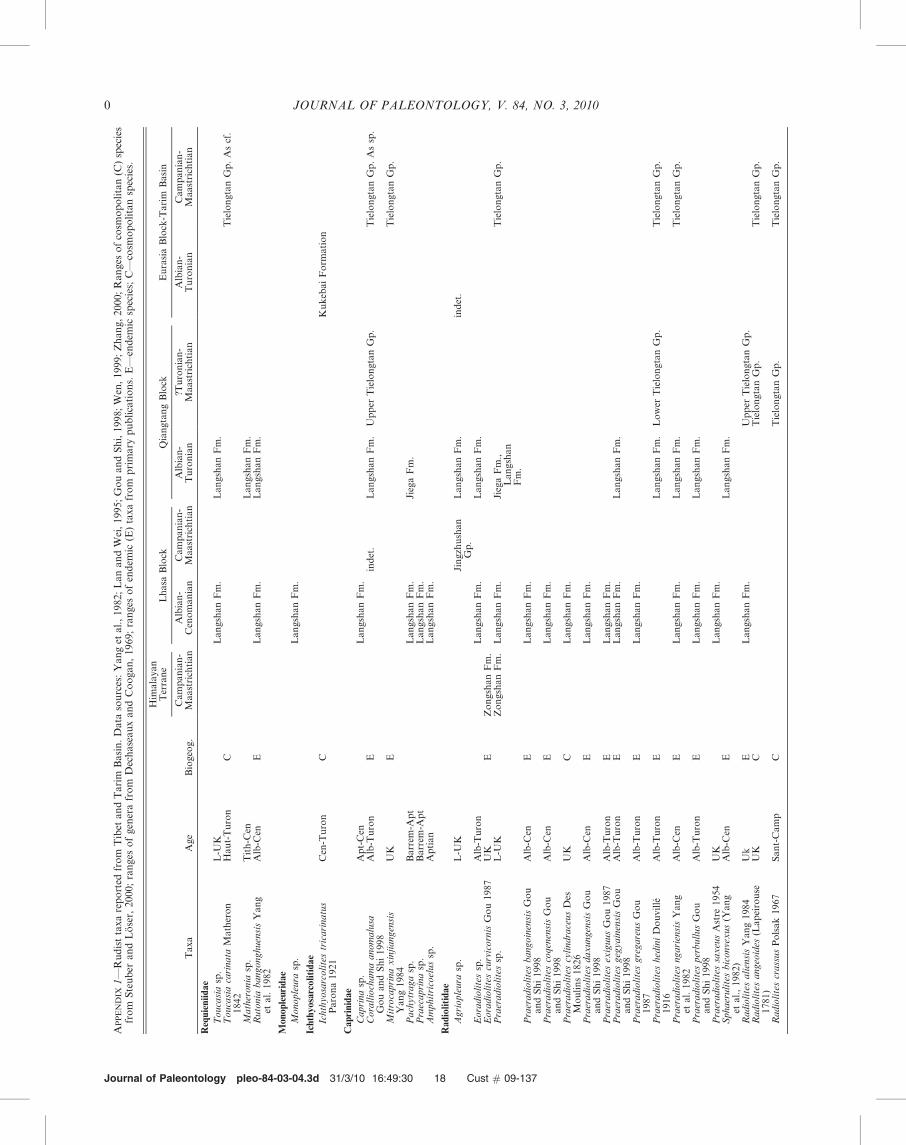
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:30 18 Cust # 09-137
AP
PE
ND
IX1—
Ru
dis
tta
xa
rep
ort
edfr
om
Tib
eta
nd
Ta
rim
Ba
sin
.D
ata
sou
rces
:Y
an
get
al.
,1
98
2;
Lan
an
dW
ei,
19
95
;G
ou
an
dS
hi,
19
98;
Wen
,1
99
9;
Zh
an
g,
20
00
;R
an
ges
of
cosm
op
oli
tan
(C)
spec
ies
fro
mS
teu
ber
an
dL
ose
r,2
00
0;
ran
ges
of
gen
era
fro
mD
ech
ase
au
xa
nd
Co
og
an
,1
96
9;
ran
ges
of
end
emic
(E)
tax
afr
om
pri
ma
ryp
ub
lica
tio
ns.
E—
end
emic
spec
ies;
C—
cosm
op
oli
tan
spec
ies.
Taxa
Age
Bio
geo
g.
Him
ala
yan
Ter
ran
eL
hasa
Blo
ckQ
ian
gta
ng
Blo
ckE
ura
sia
Blo
ck-T
ari
mB
asi
n
Cam
pan
ian
-M
aast
rich
tian
Alb
ian
-C
eno
man
ian
Cam
pan
ian
-M
aast
rich
tian
Alb
ian
-T
uro
nia
n?T
uro
nia
n-
Maast
rich
tian
Alb
ian
-T
uro
nia
nC
am
pan
ian
-M
aast
rich
tian
Req
uie
nii
dae
Touca
sia
sp.
L-U
KL
an
gsh
an
Fm
.L
an
gsh
an
Fm
.T
ouca
sia
cari
nata
Math
ero
n1842
Hau
t-T
uro
nC
Tie
lon
gta
nG
p.
As
cf.
Math
eronia
sp.
Tit
h-C
enL
an
gsh
an
Fm
.R
uto
nia
bangonghuen
sis
Ya
ng
etal.
1982
Alb
-Cen
EL
an
gsh
an
Fm
.L
an
gsh
an
Fm
.
Monople
uri
dae
Monople
ura
sp.
Lan
gsh
an
Fm
.
Ichth
yosa
rcoli
tidae
Ichth
yosa
rcoli
tes
tric
ari
natu
sP
aro
na
1921
Cen
-Tu
ron
CK
uk
ebai
Fo
rmati
on
Capri
nid
ae
Capri
na
sp.
Ap
t-C
enL
an
gsh
an
Fm
.C
ora
llio
cham
aanom
alu
saG
ou
an
dS
hi
19
98
Alb
-Tu
ron
Ein
det
.L
an
gsh
an
Fm
.U
pp
erT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
As
sp.
Mit
roca
pri
na
xin
jiangen
sis
Yan
g1984
UK
ET
ielo
ngta
nG
p.
Pach
ytr
aga
sp.
Barr
em-A
pt
Lan
gsh
an
Fm
.Ji
ega
Fm
.P
raec
apri
na
sp.
Barr
em-A
pt
Lan
gsh
an
Fm
.A
mphit
rico
elus
sp.
Ap
tian
Lan
gsh
an
Fm
.
Radio
liti
dae
Agri
ople
ura
sp.
L-U
KJi
ngzh
ush
an
Gp
.L
an
gsh
an
Fm
.in
det
.
Eora
dio
lite
ssp
.A
lb-T
uro
nL
an
gsh
an
Fm
.L
an
gsh
an
Fm
.E
ora
dio
lite
scu
rvic
orn
isG
ou
19
87
UK
EZ
on
gsh
an
Fm
.P
raer
adio
lite
ssp
.L
-UK
Zo
ngsh
an
Fm
.L
an
gsh
an
Fm
.Ji
ega
Fm
.,L
an
gsh
an
Fm
.
Tie
lon
gta
nG
p.
Pra
eradio
lite
sbangoin
ensi
sG
ou
an
dS
hi
19
98
Alb
-Cen
EL
an
gsh
an
Fm
.
Pra
eradio
lite
sco
qen
ensi
sG
ou
an
dS
hi
19
98
Alb
-Cen
EL
an
gsh
an
Fm
.
Pra
eradio
lite
scy
lindra
ceus
Des
Mo
uli
ns
1826
UK
CL
an
gsh
an
Fm
.
Pra
eradio
lite
sdax
ungen
sis
Go
ua
nd
Sh
i1
99
8A
lb-C
enE
Lan
gsh
an
Fm
.
Pra
eradio
lite
sex
iguus
Go
u1987
Alb
-Tu
ron
EL
an
gsh
an
Fm
.P
raer
adio
lite
sgeg
yain
ensi
sG
ou
an
dS
hi
19
98
Alb
-Tu
ron
EL
an
gsh
an
Fm
.L
an
gsh
an
Fm
.
Pra
eradio
lite
sgre
gare
us
Go
u1987
Alb
-Tu
ron
EL
an
gsh
an
Fm
.
Pra
eradio
lite
shed
ini
Do
uvil
le1916
Alb
-Tu
ron
EL
an
gsh
an
Fm
.L
ow
erT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
Pra
eradio
lite
sngari
ensi
sY
an
get
al.
1982
Alb
-Cen
EL
an
gsh
an
Fm
.L
an
gsh
an
Fm
.T
ielo
ngta
nG
p.
Pra
eradio
lite
sper
bull
us
Go
ua
nd
Sh
i1
99
8A
lb-T
uro
nE
Lan
gsh
an
Fm
.L
an
gsh
an
Fm
.
Pra
eradio
lite
ssa
xeu
sA
stre
1954
UK
Lan
gsh
an
Fm
.S
phaer
uli
tes
bic
onve
xus
(Ya
ng
etal.
,1982)
Alb
-Cen
EL
an
gsh
an
Fm
.
Radio
lite
sali
ensi
sY
an
g1984
Uk
EL
an
gsh
an
Fm
.U
pp
erT
ielo
ngta
nG
p.
Radio
lite
sangeo
ides
(Lap
eiro
use
1781)
UK
CT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
Radio
lite
scr
ass
us
Po
lsak
1967
San
t-C
am
pC
Tie
lon
gta
nG
p.
Tie
lon
gta
nG
p.
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010

Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:33 19 Cust # 09-137
Taxa
Age
Bio
geo
g.
Him
ala
yan
Ter
ran
eL
hasa
Blo
ckQ
ian
gta
ng
Blo
ckE
ura
sia
Blo
ck-T
ari
mB
asi
n
Cam
pan
ian
-M
aast
rich
tian
Alb
ian
-C
eno
man
ian
Cam
pan
ian
-M
aast
rich
tian
Alb
ian
-T
uro
nia
n?T
uro
nia
n-
Maast
rich
tian
Alb
ian
-T
uro
nia
nC
am
pan
ian
-M
aast
rich
tian
Radio
lite
sk
unlu
nen
sis
Ya
ng
19
84
UK
ET
ielo
ngta
nG
p.
Radio
lite
sx
inji
angen
sis
Ya
ng
19
84
UK
ET
ielo
ngta
nG
p.
Bir
adio
lite
ssp
.U
KZ
on
gsh
an
Fm
.L
an
gsh
an
Fm
.Ji
ega
Fm
.L
ow
erT
ielo
ngta
nG
p.
Bir
adio
lite
sbold
juanen
sis
Bo
bk
ova
1961
Maast
EY
igez
iya
Fm
.T
ielo
ngta
nG
p.
Bir
adio
lite
slu
mbri
coid
esD
ou
vil
le1926
UK
CT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
Bir
adio
lite
sm
inor
Po
jark
ova
1955
Ca
mp
EY
igez
iya
Fm
.
Gorj
anovi
cia
acu
tico
stata
Po
lsak
1967
UK
CT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
Med
eell
a?
Sp
.U
KL
an
gsh
an
Fm
.M
ilova
novi
cia
?S
p.
Tu
ron
ian
Tie
lon
gta
nG
p.
Bourn
onia
fourt
aui
Do
uvil
le1910
UK
CB
ourn
onia
hayden
iD
ou
vil
le1916
Cam
p-M
aas
EZ
on
gsh
an
Fm
.L
an
gsh
an
Fm
.Ji
ega
Fm
.T
ielo
ngta
nG
p.
Bourn
onia
tibet
ica
Do
uvil
le1916
Cam
p-M
aas
EZ
on
gsh
an
Fm
.B
ourn
onia
xin
jiangen
sis
Ya
ng
1984
UK
ET
ielo
ngta
nG
p.
Dis
tefa
nel
lasp
.U
KU
pp
erT
ielo
ngta
nG
p.
Sauva
ges
iak
unlu
nen
sis
Ya
ng
19
84
UK
ET
ielo
ngta
nG
p.
Sauva
ges
iate
xana
Ro
emer
1852
Alb
-Cen
CT
ielo
ngta
nG
p.
Sauva
ges
iax
inji
angen
isis
Ya
ng
1984
UK
ET
ielo
ngta
nG
p.
Dura
nia
sp.
L-U
KL
ow
erT
ielo
ngta
nG
p.
Lapei
rousi
ajo
uannet
iD
esM
ou
lin
s1826
UK
CL
an
gsh
an
Fm
.U
pp
erT
ielo
ngta
nG
p.-
sp.
Lapei
rousi
alo
mata
Go
ua
nd
Sh
i1998
Alb
-Tu
ron
EZ
on
gsh
an
Fm
.L
an
gsh
an
Fm
.
Lapei
rouse
lla
qie
maganen
sis
Lan
an
dW
ei1995
Co
n-S
an
tE
Yig
eziy
aF
m.
Osc
uli
ger
aoyta
rensi
sL
an
an
dW
ei1995
Maas
EY
igez
iya
Fm
.
Osc
uli
ger
ayen
gis
are
nsi
sY
an
g1984
UK
ET
ielo
ngta
nG
p.
Pet
kovi
cia
sp.
San
ton
ian
Lan
gsh
an
Fm
.T
ielo
ngta
nG
p.
New
Fam
ily?
Him
erali
tes
sp.
Cen
om
an
ian
La
ng
sha
nF
m.
Hip
puri
tidae
Hip
puri
tes
sp.
UK
Lan
gsh
an
Fm
.U
pp
erT
ielo
ngta
nG
p.
Tie
lon
gta
nG
p.
Rhed
ensi
asp
.T
uro
nia
nL
ow
erT
ielo
ngta
nG
p.
Vacc
init
essp
.U
KL
ow
erT
ielo
ngta
nG
p.
Poly
conit
idae
Gyro
ple
ura
magia
nen
sis
Po
jark
ova
1955
Ca
mp
EL
an
gsh
an
Fm
.as
sp.
Yig
eziy
aF
m.
Gyro
ple
ura
vak
hsc
hen
sis
Bo
bk
ov
a1961
Ca
mp
EY
igez
iya
Fm
.
Gyro
ple
ura
vak
hsc
hen
sis
darw
ase
ana
Bo
bk
ova
1961
Ca
mp
EY
igez
iya
Fm
.
Hori
ople
ura
hayden
iD
ou
vil
le1926
Cam
p-M
aas
EY
igez
iya
Fm
.
Pla
gio
pty
chid
ae
Pla
gio
pty
chus
cf.
tibet
ica
Do
uvil
le1916
Cam
p-M
aas
EZ
on
gsh
an
Fm
.in
det
.
Dic
tyopty
chid
ae
Dic
tyopty
chus
per
sica
Co
x1933
Cam
p-M
aast
ER
epo
rted
inL
an
gsh
an
Fm
.-d
ou
btf
ulD
ura
nia
cf.
per
sica
of
Go
uan
dS
hi
1998]
AP
PE
ND
IX1—
Co
nti
nu
ed.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0
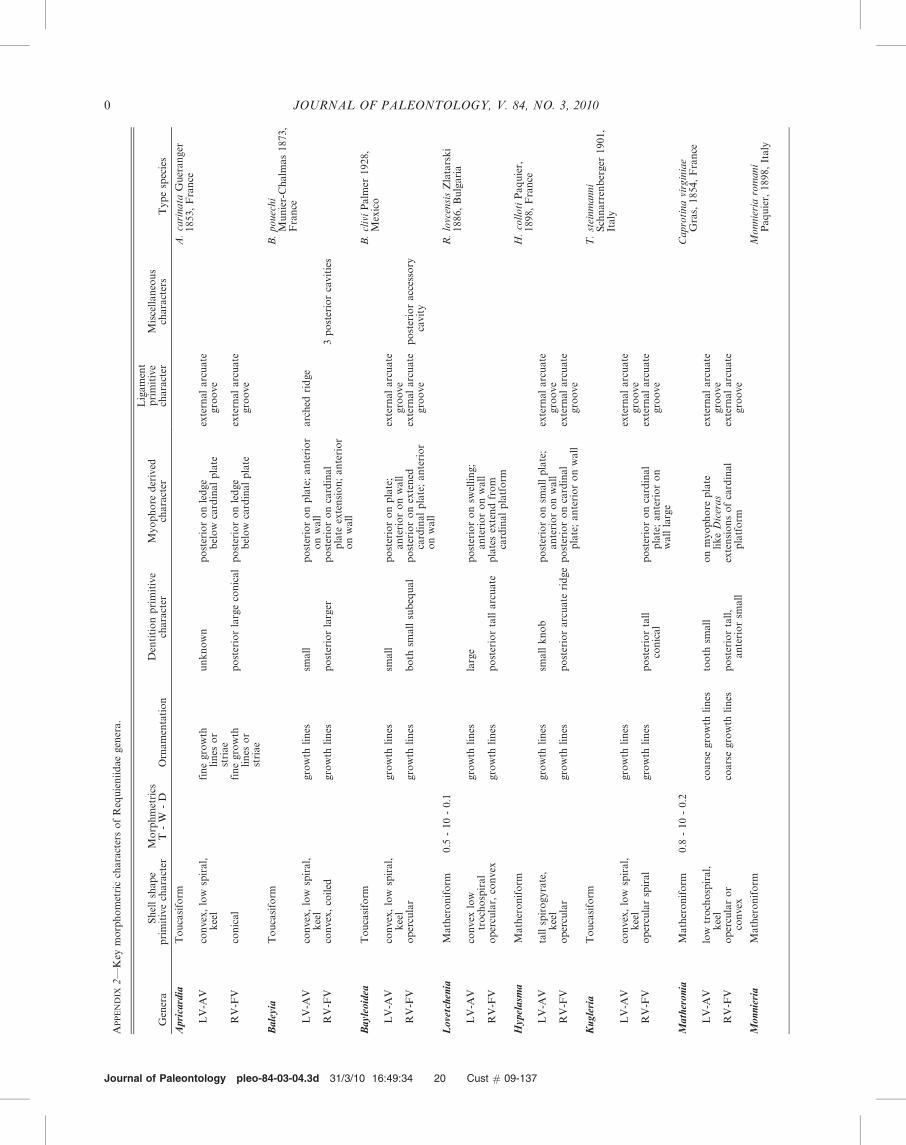
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:34 20 Cust # 09-137
AP
PE
ND
IX2—
Key
mo
rph
om
etri
cch
ara
cter
so
fR
equ
ien
iid
ae
gen
era
.
Gen
era
Sh
ell
shap
ep
rim
itiv
ech
ara
cter
Mo
rph
met
rics
T-
W-
DO
rna
men
tati
on
Den
titi
on
pri
mit
ive
cha
ract
erM
yo
ph
ore
der
ived
cha
ract
er
Lig
am
ent
pri
mit
ive
cha
ract
erM
isce
lla
neo
us
cha
ract
ers
Ty
pe
spec
ies
Ap
rica
rdia
To
uca
sifo
rmA
.ca
rin
ata
Gu
era
ng
er1
85
3,
Fra
nce
LV
-AV
con
vex
,lo
wsp
ira
l,k
eel
fin
eg
row
thli
nes
or
stri
ae
un
kn
ow
np
ost
erio
ro
nle
dge
bel
ow
card
ina
lp
late
exte
rnal
arc
ua
teg
roo
ve
RV
-FV
con
ica
lfi
ne
gro
wth
lin
eso
rst
ria
e
po
ster
ior
larg
eco
nic
al
po
ster
ior
on
led
ge
bel
ow
card
ina
lp
late
exte
rnal
arc
ua
teg
roo
ve
Bal
eyia
To
uca
sifo
rmB
.p
ou
echi
Mu
nie
r-C
ha
lma
s1
87
3,
Fra
nce
LV
-AV
con
vex
,lo
wsp
ira
l,k
eel
gro
wth
lin
essm
all
po
ster
ior
on
pla
te;
an
teri
or
on
wa
lla
rch
edri
dge
RV
-FV
con
vex
,co
iled
gro
wth
lin
esp
ost
erio
rla
rger
po
ster
ior
on
card
inal
pla
teex
ten
sio
n;
an
teri
or
on
wa
ll
3p
ost
erio
rca
vit
ies
Bay
leoid
eaT
ou
casi
form
B.
cliv
iP
alm
er1
92
8,
Mex
ico
LV
-AV
con
vex
,lo
wsp
ira
l,k
eel
gro
wth
lin
essm
all
po
ster
ior
on
pla
te;
an
teri
or
on
wa
llex
tern
al
arc
ua
teg
roo
ve
RV
-FV
op
ercu
lar
gro
wth
lin
esb
oth
small
sub
equ
al
po
ster
ior
on
exte
ned
card
ina
lp
late
;a
nte
rio
ro
nw
all
exte
rnal
arc
ua
teg
roo
ve
po
ster
ior
acc
esso
ryca
vit
y
Lov
etch
enia
Ma
ther
on
ifo
rm0
.5-
10
-0
.1R
.lo
vcen
sis
Zla
tars
ki
18
86,
Bu
lga
ria
LV
-AV
con
vex
low
tro
cho
spir
al
gro
wth
lin
esla
rge
po
ster
ior
on
swel
lin
g;
an
teri
or
on
wa
llR
V-F
Vo
per
cula
r,co
nvex
gro
wth
lin
esp
ost
erio
rta
llarc
uate
pla
tes
exte
nd
fro
mca
rdin
al
pla
tfo
rm
Hy
pela
sma
Ma
ther
on
ifo
rmH
.co
lloti
Pa
qu
ier,
18
98,
Fra
nce
LV
-AV
tall
spir
og
yra
te,
kee
lgro
wth
lin
essm
all
kn
ob
po
ster
ior
on
small
pla
te;
an
teri
or
on
wa
llex
tern
al
arc
ua
teg
roo
ve
RV
-FV
op
ercu
lar
gro
wth
lin
esp
ost
erio
rarc
uate
rid
ge
po
ster
ior
on
card
inal
pla
te;
an
teri
or
on
wa
llex
tern
al
arc
ua
teg
roo
ve
Ku
gle
ria
To
uca
sifo
rmT
.st
ein
ma
nn
iS
chn
arr
enb
erg
er1
90
1,
Ita
ly
LV
-AV
con
vex
,lo
wsp
ira
l,k
eel
gro
wth
lin
esex
tern
al
arc
ua
teg
roo
ve
RV
-FV
op
ercu
lar
spir
al
gro
wth
lin
esp
ost
erio
rta
llco
nic
al
po
ster
ior
on
card
inal
pla
te;
an
teri
or
on
wa
llla
rge
exte
rnal
arc
ua
teg
roo
ve
Ma
ther
on
iaM
ath
ero
nif
orm
0.8
-1
0-
0.2
Ca
pro
tin
avi
rgin
iae
Gra
s,1
85
4,
Fra
nce
LV
-AV
low
tro
cho
spir
al,
kee
lco
ars
eg
row
thli
nes
too
thsm
all
on
my
op
ho
rep
late
lik
eD
icer
as
exte
rnal
arc
ua
teg
roo
ve
RV
-FV
op
ercu
lar
or
con
vex
coars
egro
wth
lin
esp
ost
erio
rta
ll,
an
teri
or
small
exte
nsi
on
so
fca
rdin
al
pla
tfo
rmex
tern
al
arc
ua
teg
roo
ve
Mo
nn
ieri
aM
ath
ero
nif
orm
Mo
nn
ieri
aro
man
iP
aq
uie
r,1
89
8,
Ita
ly
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010
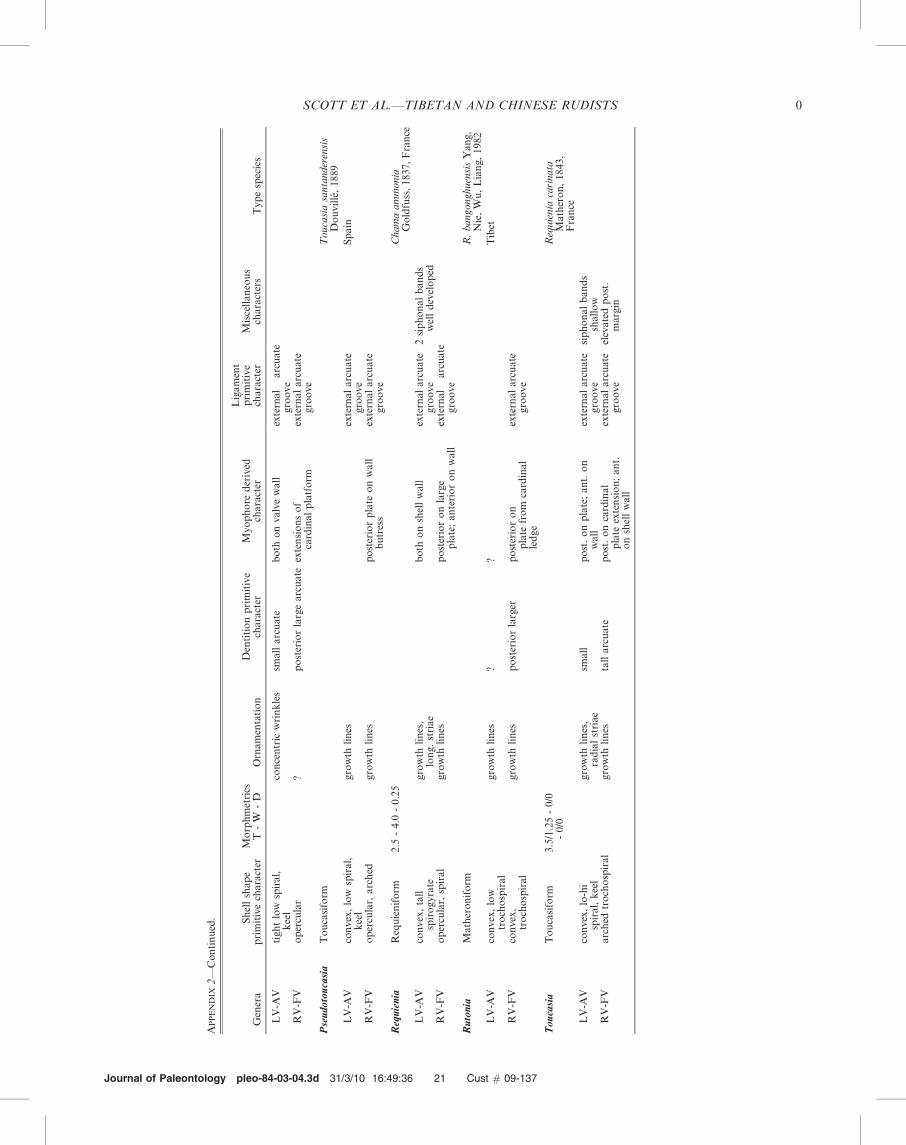
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:36 21 Cust # 09-137
Gen
era
Sh
ell
sha
pe
pri
mit
ive
cha
ract
erM
orp
hm
etri
csT
-W
-D
Orn
am
enta
tio
nD
enti
tio
np
rim
itiv
ech
ara
cter
My
op
ho
red
eriv
edch
ara
cter
Lig
am
ent
pri
mit
ive
cha
ract
erM
isce
lla
neo
us
cha
ract
ers
Ty
pe
spec
ies
LV
-AV
tig
ht
low
spir
al,
kee
lco
nce
ntr
icw
rin
kle
ssm
all
arc
uate
bo
tho
nvalv
ew
all
exte
rnal
arc
uate
gro
ov
eR
V-F
Vo
per
cula
r?
po
ster
ior
larg
earc
uate
exte
nsi
on
so
fca
rdin
al
pla
tfo
rmex
tern
al
arc
ua
teg
roo
ve
Pse
ud
oto
uca
sia
To
uca
sifo
rmT
ouca
sia
santa
nder
ensi
sD
ou
vil
le,
18
89
LV
-AV
con
vex
,lo
wsp
ira
l,k
eel
gro
wth
lin
esex
tern
al
arc
ua
teg
roo
ve
Sp
ain
RV
-FV
op
ercu
lar,
arc
hed
gro
wth
lin
esp
ost
erio
rp
late
on
wa
llb
utr
ess
exte
rna
la
rcu
ate
gro
ov
e
Req
uie
nia
Req
uie
nif
orm
2.5
-4
.0-
0.2
5C
ha
ma
am
mo
nia
Go
ldfu
ss,
18
37,
Fra
nce
LV
-AV
con
vex
,ta
llsp
iro
gy
rate
gro
wth
lin
es,
lon
g.
stri
ae
bo
tho
nsh
ell
wa
llex
tern
al
arc
ua
teg
roo
ve
2si
ph
on
al
ba
nd
sw
ell
dev
elo
ped
RV
-FV
op
ercu
lar,
spir
al
gro
wth
lin
esp
ost
erio
ro
nla
rge
pla
te;
an
teri
or
on
wa
llex
tern
al
arc
ua
teg
roo
ve
Ru
ton
iaM
ath
ero
nif
orm
R.
ba
ng
on
gh
uen
sis
Ya
ng
,N
ie,
Wu
,L
ian
g,
19
82
LV
-AV
con
vex
,lo
wtr
och
osp
ira
lg
row
thli
nes
??
Tib
et
RV
-FV
con
vex
,tr
och
osp
ira
lgro
wth
lin
esp
ost
erio
rla
rger
po
ster
ior
on
pla
tefr
om
card
ina
lle
dg
e
exte
rna
la
rcu
ate
gro
ov
e
Tou
casi
aT
ou
casi
form
3.5
/1.2
5-
0/0
-0
/0R
equie
nia
cari
na
taM
ath
ero
n,
18
43
,F
ran
ce
LV
-AV
con
vex
,lo
-hi
spir
al,
kee
lg
row
thli
nes
,ra
dia
lst
ria
esm
all
po
st.
on
pla
te;
an
t.o
nw
all
exte
rna
la
rcu
ate
gro
ov
esi
ph
on
al
ba
nd
ssh
all
ow
RV
-FV
arc
hed
tro
cho
spir
al
gro
wth
lin
esta
lla
rcu
ate
po
st.
on
card
ina
lp
late
exte
nsi
on
;a
nt.
on
shel
lw
all
exte
rna
la
rcu
ate
gro
ov
eel
eva
ted
po
st.
ma
rgin
AP
PE
ND
IX2—
Co
nti
nu
ed.
SCOTT ET AL.—TIBETAN AND CHINESE RUDISTS 0

APPENDIX 3
REQUIENIIDAE CHARACTER STATES.
1. Shell Microstructure. 0, Single layer aragonite; 1, two layers-calcite &aragonite.
2. Shell Form LV. 0, subequivalved spirogyrate, moderate translationalong coiling axis; 1, inequivalved, low spirogyrate, translated slightlyfrom the commissure along the coiling axis, matheroniform; 2, veryinequivalved, tall trochospiral, translated moderately along thecoiling axis, toucasiform; 3, very inequivalved, tall trochospiral,translated greatly along the coiling axis, requieniform.
3. Shell Form RV. 0, inflated, strongly coiled; 1, slightly inflated and iscoiled with little translation; 2, flat to convex with little or no coilingtranslation.
4. Ornament. 0, coarse growth rings; 1, coarse growth rings and radialstriae; 2, fine growth lines only; 3, fine growth lines and radial striae.
5. LV Posterior Myophore. 0, separate ledge or plate extending fromvalve wall; 1, impressed directly on interior valve surface.
6. RV Myophore. 0, posterior myophore plate extending from thecardinal platform along the posterior valve margin; 1, posteriormyophore is a separate plate attached to and projecting from thevalve wall.
7. Siphonal Bands. 0, absent; 1, wide shallow depressions expressedas subtle curves in the growth lines on the posterior margin of theLV.
8. Accessory Cavity. 0, none; 1, one or more associated with RVposterior myophore.
9. RV Posterior Tooth. 0, a large arcuate ridge; 1, curved cone; 2, smallsubequal.
10. LV Central Tooth. 0, large conical; 1, small knob-like.11. Ligament. 0, long arcuate groove posterior to hinge plate; 1, short
small pit below beak.
Journal of Paleontology pleo-84-03-04.3d 31/3/10 16:49:37 22 Cust # 09-137
0 JOURNAL OF PALEONTOLOGY, V. 84, NO. 3, 2010





























