Embed Size (px)
Citation preview

Journ
alof
Cell
Scie
nce
SNX9, SNX18 and SNX33 are required for progressionthrough and completion of mitosis
Maggie P. C. Ma and Megan Chircop*Children’s Medical Research Institute, The University of Sydney, 214 Hawkesbury Road, Westmead, NSW 2145, Australia
*Author for correspondence ([email protected])
Accepted 24 May 2012Journal of Cell Science 125, 4372–4382� 2012. Published by The Company of Biologists Ltddoi: 10.1242/jcs.105981
SummaryMitosis involves considerable membrane remodelling and vesicular trafficking to generate two independent cells. Consequently,endocytosis and endocytic proteins are required for efficient mitotic progression and completion. Several endocytic proteins also
participate in mitosis in an endocytosis-independent manner. Here, we report that the sorting nexin 9 (SNX9) subfamily members –SNX9, SNX18 and SNX33 – are required for progression and completion of mitosis. Depletion of any one of these proteins using siRNAinduces multinucleation, an indicator of cytokinesis failure, as well as an accumulation of cytokinetic cells. Time-lapse microscopy on
siRNA-treated cells revealed a role for SNX9 subfamily members in progression through the ingression and abscission stages ofcytokinesis. Depletion of these three proteins disrupted MRLCS19 localization during ingression and recruitment of Rab11-positiverecycling endosomes to the intracellular bridge between nascent daughter cells. SNX9 depletion also disrupted the localization of Golgi
during cytokinesis. Endocytosis of transferrin was blocked during cytokinesis by depletion of the SNX9 subfamily members, suggestingthat these proteins participate in cytokinesis in an endocytosis-dependent manner. In contrast, depletion of SNX9 did not blocktransferrin uptake during metaphase but did delay chromosome alignment and segregation, suggesting that SNX9 plays an additional
non-endocytic role at early mitotic stages. We conclude that SNX9 subfamily members are required for mitosis through bothendocytosis-dependent and -independent processes.
Key words: SNX9, SNX18, SNX33, Mitosis, Vesicle trafficking, Abscission
IntroductionEndocytosis is thought to shut down during mitosis then resume
during the final stage, cytokinesis (Schweitzer et al., 2005). In supportof this idea, several endocytic proteins including disabled-2 (Dab2),dynamin II (dynII), epsin, eps15, a-adaptin and amphiphysin II, areinactivated by Cdk1-mediated phosphorylation during mitosis (Chen
et al., 1999; Chircop et al., 2010; Dephoure et al., 2008; He et al.,2003; Kariya et al., 2000). Furthermore, clathrin, epsin and cyclin-G-associated kinase (GAK) have been shown to play non-endocytic
roles in chromosome alignment and segregation during mitosis (Liuand Zheng, 2009; Royle et al., 2005; Shimizu et al., 2009).
Cytokinesis results in the generation of two independent
daughter cells and can be divided into two stages: (1) membraneingression to generate a cleavage furrow between segregatedchromosomes followed by (2) membrane abscission at a site along
the intracellular bridge (ICB) connecting the two nascent daughtercells (Glotzer, 2005). Vesicles derived from internal organelles,including the Golgi, lysosomes, early endosomes and recyclingendosomes, traffic from both daughter cells to the ICB. These
vesicles accumulate asymmetrically on one side of the centrallylocated midbody ring (MR) and are required for abscission (Gossand Toomre, 2008). However, the role of these vesicles in
abscission is unknown.
During cytokinesis, endocytosis contributes to the generation ofrecycling endosomes that are delivered to the ICB. Depletion or
functional inhibition of dynII and Arf6 results in multinucleatecells, an indicator of cytokinesis failure (Chircop et al., 2011a;Fielding et al., 2005; Joshi et al., 2010). Arf6 depletion blocks
trafficking of Rab11-positive recycling endosomes to the ICB
(Fielding et al., 2005). In contrast, dynII and clathrin localise to themidbody (Chircop et al., 2010; Gromley et al., 2005; Low et al.,
2003; Warner et al., 2006), suggesting that they participate moredirectly in the abscission process. The identification of all
endocytic proteins required for mitosis and their roles during thisprocess are yet to be revealed.
Sorting nexin 9 (SNX9) is one of the major binding partners fordynII in endocytosis (Lundmark and Carlsson, 2004). SNX9 belongs
to the sorting nexin (SNX) superfamily of proteins that consistsof 33 proteins in mammals (Seet and Hong, 2006). All members
contain a SNX–Phox–homology (PX) domain and associate with
phosphotidylinositol-3-monophosphate-enriched elements of theearly endocytic network via their PX domain (Yarar et al., 2008).
The SNXs function in diverse processes, including endocytosis,endosomal sorting and endosomal signalling (Carlton et al., 2005;
Seet and Hong, 2006). Together with SNX18 and SNX33, SNX9belongs to the SNX9 subfamily due to the presence of a Bin–
Amphiphysin–Rvs (BAR) domain that allows dimerization and
contributes to modulation and shaping of membrane curvature(Carlton et al., 2004), as well as a Src-homology 3 (SH3) domain that
allows interaction with a wide range of proline-rich PXXP motifcontaining proteins (Alto et al., 2007; Mayer, 2001). Of the three
SNX9 subfamily members, SNX9 is the best characterized. It is amajor binding partner of a number of proteins involved in clathrin-
mediated endocytosis (CME), such as clathrin, cargo adaptor protein-
2 (AP-2), dynamin, synaptojanin, actin-related protein-2/3 (ARP2/3)complex activator, Wiskott-Aldrich syndrome protein (WASp) and
4372 Research Article

Journ
alof
Cell
Scie
nce
Cdc42-associated kinase (ACK) (Badour et al., 2007; Lundmark and
Carlsson, 2002; Lundmark and Carlsson, 2004; Shin et al., 2008;
Soulet et al., 2005; Yarar et al., 2007; Yeow-Fong et al., 2005). SNX9
is also essential for dorsal-ruffle formation and clathrin-independent,
actin-dependent fluid phase endocytosis (Yarar et al., 2007).
Although the three SNX9 subfamily members share highly similar
structures, they appear to have non-overlapping functions and it
remains unclear if they can form heteromeric complexes (Dislich
et al., 2011; Haberg et al., 2008). SNX18 functions in clathrin-
independent endosomal trafficking that is dependent on AP1 and the
retrograde trafficking protein PACS1 (Haberg et al., 2008). SNX18
also plays a redundant role to SNX9 in CME (Park et al., 2010).
SNX33 has been implicated in dynamin-dependent endocytosis by
modulating amyloid precursor protein (APP) endocytosis and a-
secretase cleavage (Schobel et al., 2008) and the formation of prion
protein (Heiseke et al., 2008). Of the three SNX9 subfamily members,
only SNX33 has been shown to play a role in mitosis, as its
overexpression results in micronucleate cells (Zhang et al., 2009). Its
role in mitosis is thought to be linked to its ability to regulate actin
polymerization via its association with WASp (Zhang et al., 2009).
Thus it is proposed to be a modulator of the actinomyosin II
contractile ring for membrane ingression during cytokinesis.
In this study we aimed to determine the roles of all three SNX9
subfamily members in mitosis. We show that all are required for
successful completion of both the ingression and abscission
stages of cytokinesis and for the delivery of a subset of vesicles to
the ICB during this process. We also show that SNX9 plays an
additional role during metaphase that is not dependent on its
endocytic function.
ResultsDepletion of SNX9, SNX18 or SNX33 causes
multinucleation and delays mitotic exit
SNX33 is required for successful completion of mitosis (Zhang
et al., 2009). Therefore, we aimed to determine whether SNX9
and SNX18 are also required for mitosis by assessing the mitotic
phenotype of HeLa cells depleted of these proteins using siRNA.
Two siRNAs were targeting each SNX9 subfamily member
was assessed. At 72 h post-transfection, immunoblot analyses
revealed satisfactory reduction in the expression level of SNX9,
SNX18 and SNX33 by their specific siRNAs compared to
untransfected and luciferase (Luc) siRNA controls (Fig. 1A–C).
A similar reduction in the protein level of these proteins was also
observed by immunofluorescence microscopy (supplementary
Fig. 1. SNX9, SNX18 or SNX33 depletion
causes cytokinesis failure. HeLa cells were
either untreated (–), or transfected with luciferase
siRNA (Luc) or siRNA targeting SNX9 (A),
SNX18 (B) or SNX33 (C). Two siRNA per
SNX9 subfamily member were assessed. At 72 h
post-transfection, protein lysates (200 mg) were
immunoblotted with anti-SNX9, anti-SNX18 or
anti-SNX33 antibodies. c-tubulin served as a
loading control. Duplicates of each siRNA
treatment were immunoblotted. The separate
sections of blots shown in A–C were all from the
same blot. Cells were transfected as described in
A–C and synchronized at the G2–M boundary
using RO-3306. Upon RO-3306 wash out cells
were released for 6 hours, fixed and
immunostained for a-tubulin and scored for
multinuclei (D) and cytokinesis (E).
(F–H) Rescue experiment. Untreated and SNX-
siRNA-treated HeLa cells were transfected with
either GFP-SNX9, HA-SNX18 or GFP-SNX33.
Prepared protein lysates (200 mg) were
immunoblotted with anti-SNX9 (F), anti-SNX18
(G), anti-SNX33 (H), anti-HA (G) and anti-GFP
(F and H). In parallel, cells were immunostained
for a-tubulin and scored for multinuclei
(F–H, bottom panels). The graphs show the
means 6 s.e.m. from three independent
experiments where n.200 cells were scored for
each sample in each experiment. *P,0.05;
**P,0.01; ***P,0.001 (one-way ANOVA).
SNX9 subfamily is required for mitosis 4373

Journ
alof
Cell
Scie
nce
material Fig. S1). The siRNAs were highly specific to each SNX
protein as depletion of any one of the three SNX proteins by their
specific siRNA did not affect expression of the other two SNX9
subfamily members (Fig. 1A–C). Depletion of any one of these
proteins caused a significant increase in the percentage of
multinucleate cells, an indicator of cytokinesis failure (Fig. 1D).
Specifically, depletion of SNX9 resulted in a fourfold increase in
multinucleation (untreated: 3.860.9%; Luc siRNA: 3.460.7%;
SNX9-1 siRNA: 11.963.2%; SNX9-2 siRNA: 12.161%;
Fig. 1D), whereas depletion of SNX18 or SNX33 caused a
sevenfold increase in multinucleation (SNX18-1 siRNA:
20.561.7%; SNX18-2 siRNA: 30.460.5%; SNX33-1 siRNA:
17.662.3%; SNX33-2 siRNA: 30.563%; Fig. 1D). At 6 h post-
mitotic entry, there was a significant increase in cells undergoing
cytokinesis following depletion of SNX9 subfamily members
(untreated: 2.060.52%; Luc siRNA: 2.860.4%; SNX9-1 siRNA:
8.263.2%; SNX9-2 siRNA: 10.962.8%; SNX18-1 siRNA:
10.960.9%; SNX18-2 siRNA: 13.661.5%; SNX33-1 siRNA:
15.461.5%; SNX33-2 siRNA: 19.361.5%; Fig. 1E), suggesting
that depletion of any one of these proteins delays mitotic exit.
Since both siRNAs assessed per protein induced similar
phenotypes, SNX9-1, SNX18-1 and SNX33-1 siRNAs were used
in all subsequent experiments. Depletion of all three SNX proteins
produced similar phenotypes in U2OS human osteosarcoma cells
(supplementary material Fig. S2). We next aimed to rescue the
multinucleation phenotype induced by SNX9, SNX18 or SNX33
depletion by overexpressing GFP–SNX9 (Fig. 1F), HA–SNX18
(Fig. 1G) and GFP–SNX33 (Fig. 1H), respectively. Indeed,
ectopic expression of these proteins completely rescued the
multinucleation phenotype (Fig. 1F–H). We conclude that all
three SNX9 subfamily members are required for successful
completion of mitosis.
To further characterise the role of the SNX9 subfamily in
mitosis we carried out time-lapse video microscopy to measure the
time spent by individual cells in mitosis (Fig. 2A). Consistent with
our fixed cell analysis (Fig. 1D,E), an increased number of SNX9-
subfamily-depleted cells failed mitosis and either became
binucleate or did not complete mitosis within the 20 h
experimental period (Fig. 2B). Regardless of whether or not
these depleted cells completed or failed mitosis they spent a longer
period of time in mitosis than cells treated with Luc siRNA
(Fig. 2C,D). Specifically, cells depleted of SNX9, SNX18 and
Fig. 2. SNX9 subfamily proteins are
required for efficient mitotic progression.
(A) Schematic diagram of the phases of
mitosis with a plan of the time-lapse
experiment shown above. Pro, prophase; Ana,
anaphase; Comp, completion; Multi,
multinucleation. (B,C) HeLa cells were
treated with siRNA targeting Luc, SNX9,
SNX18 or SNX33 and visualized by time-
lapse microscopy. (B) The graph shows the
percentage of cells that entered but did not
complete mitosis or were multinucleate by the
end of the 20 h experimental period (n.80
cells per sample). (C) The graph shows the
time each individual cell took to undergo
mitosis (n.80 per sample). Depletion of any
one of the SNX9 subfamily members prolongs
the time cells spend in mitosis. The unbroken
black line represents the median time.
(D) Selected frames from representative time-
lapse movies of Luc-, SNX9-, SNX18- and
SNX33-siRNA-treated cells. Data shown in
B–D are from one representative experiment.
Similar results were obtained in at least two
independent experiments.
Journal of Cell Science 125 (18)4374

Journ
alof
Cell
Scie
nce
SNX33 spent a median time of 210, 220 and 270 min in mitosis,
respectively, compared to 120 min for Luc siRNA control cells(Fig. 2C,D). Thus, the function of these three proteins is associated
with efficient mitotic progression and completion.
SNX9, 18 and 33 have multiple points of actionthroughout mitosis
To determine the point of action of the three SNX9 subfamilymembers during mitosis, we determined the time siRNA-treated
cells took to progress through the following four mitotic stages:
prophase to anaphase, and anaphase to completion of cytokinesis,
either successfully or unsuccessfully generating a multinucleatecell (Fig. 2A). Neither SNX18 nor SNX33 appear to play a role at
early mitotic stages from prophase to anaphase since cells
depleted of these proteins completed this transition with similarkinetics to Luc siRNA control cells (Fig. 3A). In contrast,
completion of this mitotic transition was delayed in SNX9-
depleted cells, with a median time of 60 min vs 50 min for Luc
siRNA control cells (Fig. 3A). Depletion of all three proteinscaused a significant delay at the anaphase to completion or
multinucleation transition (Fig. 3B). These findings indicate that
all three SNX9 subfamily members are required for efficientprogression through cytokinesis and that SNX9 may have an
Fig. 3. SNX9 subfamily members have multiple points of action
throughout mitosis. (A,B) Those HeLa cells that were analysed in Fig. 2B,C
were further characterized by quantifying the time they took to complete the
following mitotic transitions: prophase–anaphase (A), and anaphase to mitotic
completion or multinucleate formation (B). Pro, prophase; Ana, anaphase;
Comp, completion; Multi, multinucleation; n.80 cells per sample. The
median time per sample is represented by a solid black line. Data shown in
graphs are from one representative experiment. Similar results were obtained
in at least two independent experiments.
Fig. 4. Localization of SNX9, SNX18 and SNX33 in HeLa
cells during mitosis. (A–C) Asynchronously growing HeLa
cells were fixed and stained for SNX9 (A), SNX18 (B), or
SNX33 (C) and co-stained for a-tubulin (red). DNA was
stained with DAPI (blue). Representative microscopy images
illustrating the localization of the three SNX9 subfamily
members at the indicated mitotic stages are shown. Int,
interphase; Promet, prometaphase; Met, metaphase; Ana,
anaphase; Tel, telophase; Cyto, cytokinesis.
(D) Representative microscopy images of cytokinetic HeLa
cells co-stained for the indicated SNX9 subfamily protein
(green) and the midbody ring marker, c-tubulin (red). SNX9
and SNX33 colocalize with c-tubulin.
(E) Representative microscopy images of cytokinetic HeLa
cells ectopically expressing GFP–SNX9 or stained for SNX18
or SNX33 (green) and co-stained with the flanking midbody
ring marker, dynIIS764 (red). SNX9 localizes to the flanking
midbody rings. GFP–SNX9 was ectopically expressed in this
experiment due to antibody cross-reactivity. Scale bars: 10 mm
(A–C); 1 mm (D,E).
SNX9 subfamily is required for mitosis 4375
Journ
alof
Cell
Scie
nce
additional role in ensuring efficient completion of chromosomealignment and segregation in early mitosis.
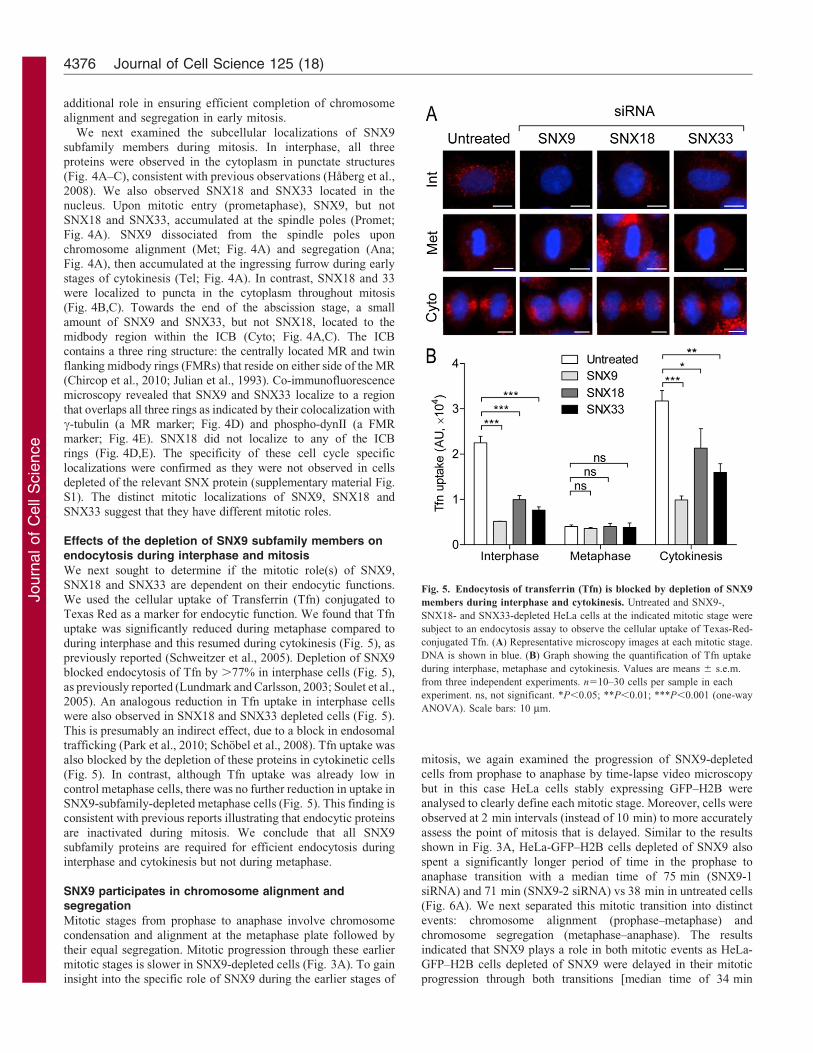
We next examined the subcellular localizations of SNX9subfamily members during mitosis. In interphase, all threeproteins were observed in the cytoplasm in punctate structures
(Fig. 4A–C), consistent with previous observations (Haberg et al.,2008). We also observed SNX18 and SNX33 located in thenucleus. Upon mitotic entry (prometaphase), SNX9, but not
SNX18 and SNX33, accumulated at the spindle poles (Promet;Fig. 4A). SNX9 dissociated from the spindle poles uponchromosome alignment (Met; Fig. 4A) and segregation (Ana;
Fig. 4A), then accumulated at the ingressing furrow during earlystages of cytokinesis (Tel; Fig. 4A). In contrast, SNX18 and 33were localized to puncta in the cytoplasm throughout mitosis(Fig. 4B,C). Towards the end of the abscission stage, a small
amount of SNX9 and SNX33, but not SNX18, located to themidbody region within the ICB (Cyto; Fig. 4A,C). The ICBcontains a three ring structure: the centrally located MR and twin
flanking midbody rings (FMRs) that reside on either side of the MR(Chircop et al., 2010; Julian et al., 1993). Co-immunofluorescencemicroscopy revealed that SNX9 and SNX33 localize to a region
that overlaps all three rings as indicated by their colocalization withc-tubulin (a MR marker; Fig. 4D) and phospho-dynII (a FMRmarker; Fig. 4E). SNX18 did not localize to any of the ICBrings (Fig. 4D,E). The specificity of these cell cycle specific
localizations were confirmed as they were not observed in cellsdepleted of the relevant SNX protein (supplementary material Fig.S1). The distinct mitotic localizations of SNX9, SNX18 and
SNX33 suggest that they have different mitotic roles.
Effects of the depletion of SNX9 subfamily members onendocytosis during interphase and mitosis
We next sought to determine if the mitotic role(s) of SNX9,
SNX18 and SNX33 are dependent on their endocytic functions.We used the cellular uptake of Transferrin (Tfn) conjugated toTexas Red as a marker for endocytic function. We found that Tfnuptake was significantly reduced during metaphase compared to
during interphase and this resumed during cytokinesis (Fig. 5), aspreviously reported (Schweitzer et al., 2005). Depletion of SNX9blocked endocytosis of Tfn by .77% in interphase cells (Fig. 5),
as previously reported (Lundmark and Carlsson, 2003; Soulet et al.,2005). An analogous reduction in Tfn uptake in interphase cellswere also observed in SNX18 and SNX33 depleted cells (Fig. 5).
This is presumably an indirect effect, due to a block in endosomaltrafficking (Park et al., 2010; Schobel et al., 2008). Tfn uptake wasalso blocked by the depletion of these proteins in cytokinetic cells
(Fig. 5). In contrast, although Tfn uptake was already low incontrol metaphase cells, there was no further reduction in uptake inSNX9-subfamily-depleted metaphase cells (Fig. 5). This finding isconsistent with previous reports illustrating that endocytic proteins
are inactivated during mitosis. We conclude that all SNX9subfamily proteins are required for efficient endocytosis duringinterphase and cytokinesis but not during metaphase.
SNX9 participates in chromosome alignment andsegregation
Mitotic stages from prophase to anaphase involve chromosomecondensation and alignment at the metaphase plate followed by
their equal segregation. Mitotic progression through these earliermitotic stages is slower in SNX9-depleted cells (Fig. 3A). To gaininsight into the specific role of SNX9 during the earlier stages of
mitosis, we again examined the progression of SNX9-depleted
cells from prophase to anaphase by time-lapse video microscopy
but in this case HeLa cells stably expressing GFP–H2B were
analysed to clearly define each mitotic stage. Moreover, cells were
observed at 2 min intervals (instead of 10 min) to more accurately
assess the point of mitosis that is delayed. Similar to the results
shown in Fig. 3A, HeLa-GFP–H2B cells depleted of SNX9 also
spent a significantly longer period of time in the prophase to
anaphase transition with a median time of 75 min (SNX9-1
siRNA) and 71 min (SNX9-2 siRNA) vs 38 min in untreated cells
(Fig. 6A). We next separated this mitotic transition into distinct
events: chromosome alignment (prophase–metaphase) and
chromosome segregation (metaphase–anaphase). The results
indicated that SNX9 plays a role in both mitotic events as HeLa-
GFP–H2B cells depleted of SNX9 were delayed in their mitotic
progression through both transitions [median time of 34 min
Fig. 5. Endocytosis of transferrin (Tfn) is blocked by depletion of SNX9
members during interphase and cytokinesis. Untreated and SNX9-,
SNX18- and SNX33-depleted HeLa cells at the indicated mitotic stage were
subject to an endocytosis assay to observe the cellular uptake of Texas-Red-
conjugated Tfn. (A) Representative microscopy images at each mitotic stage.
DNA is shown in blue. (B) Graph showing the quantification of Tfn uptake
during interphase, metaphase and cytokinesis. Values are means 6 s.e.m.
from three independent experiments. n510–30 cells per sample in each
experiment. ns, not significant. *P,0.05; **P,0.01; ***P,0.001 (one-way
ANOVA). Scale bars: 10 mm.
Journal of Cell Science 125 (18)4376

Journ
alof
Cell
Scie
nce
(SNX9-1 siRNA) and 38 min (SNX9-2 siRNA) vs 20 min for
untreated cells during prophase–metaphase (Fig. 6B); median time
of 31 min (SNX9-1 siRNA) and 34 min (SNX9-2 siRNA) vs
22 min for untreated cells during metaphase–anaphase; Fig. 6C].
Again, consistent with our findings in HeLa cells (Fig. 3), depletion
of SNX18 or SNX33 did not affect mitotic progression of HeLa-
GFP–H2B cells through these mitotic phases (Fig. 6A–C).
We next investigated the requirement of SNX9 in regulating
chromosome dynamics. The width of the centre of the metaphase
plate was significantly wider in metaphase HeLa-GFP–H2B cells
that were depleted of SNX9 compared to untreated cells
(Fig. 6D,G). Consistent with the time-lapse data indicating that
SNX18 and SNX33 are not required for progression from prophase
to anaphase, the width of the metaphase plate was not affected in
HeLa-GFP–H2B cells – depleted of these proteins (Fig. 6E–G).
These findings indicate that the metaphase function of SNX9
contributes to chromosome congression and alignment.
Role of SNX9, SNX18 and SNX33 during the ingression and
abscission phases of cytokinesis
Cleavage furrow formation during the ingression phase of
cytokinesis is driven by the activity of the actinomyosin II
contractile ring, which assembles at the cell equator between
segregated chromosomes. Myosin regulatory light chain (MRLC)
is a component of the myosin II filaments, and its phosphorylation
on T18 and S19 drives the assembly of the actinomyosin II
contractile ring (Ikebe et al., 1988) and activity of the myosin II
motor (Komatsu et al., 2000; Yamakita et al., 1994).
Phosphorylation of the major site, S19, allows myosin II to
interact with actin to assemble an actin–myosin II complex and
initiate contraction (Scholey et al., 1980). Given that all three SNX9
subfamily members are required for cytokinesis (Fig. 3B), we
assessed the activity of the myosin II motor using an antibody that
specifically recognises MRLC phosphorylated at S19. MRLCS19
localization was disrupted in SNX9-subfamily-depleted cells
Fig. 6. SNX9 functions during metaphase for chromosome
alignment. (A–C) HeLa-GFP–H2B cells were either
untreated or treated with siRNA targeting Luc, SNX9, SNX18
or SNX33, and visualized by time-lapse microscopy; images
were captured at 2 min intervals for 6 h. The graphs show the
time taken for each individual cell to complete the following
mitotic transitions: prophase to anaphase (A), prophase to
metaphase (B) and metaphase to anaphase (C). Pro, prophase;
Met, metaphase; Ana, anaphase. n56–10 cells. Solid black
line represents the median time. (D–F) Representative
microscopy images of metaphase cells showing the
chromosomes (GFP–H2B) and indicated endogenous SNX9
subfamily members (red) in untreated and SNX-depleted
HeLa-GFP–H2B cells. (G) Graph showing the width of the
metaphase plate, which was quantified by measuring the width
of the centre of the metaphase plate of HeLa-GFP–H2B cells
in metaphase that were treated with the indicated siRNAs.
Values are means 6 s.e.m. from three independent
experiments; n520–30 cells per sample in each experiment.
ns, not significant; ***P,0.001 (one-way ANOVA). Scale
bars: 10 mm.
SNX9 subfamily is required for mitosis 4377

Journ
alof
Cell
Scie
nce
during anaphase and telophase (Fig. 7A). In contrast to control cells
where MRLCS19 was concentrated at the ingressing furrow during
anaphase and telophase, it was diffusely distributed in the
cytoplasm in cells depleted of any one of the SNX9 subfamily
proteins (Fig. 7A). The ratio of the fluorescence intensity of
MRLCS19 staining at the ingressing furrow or cleavage furrow:the
fluorescence intensity of MRLCS19 at the polar region was used to
quantitate this effect (Fig. 7A, right panel). A significant reduction
of .40% in MRLCS19 fluorescence signal at the ingressing furrow
and cleavage furrow was detected (Fig. 7A–C). This was not due to
an overall loss of protein as MRLCS19 levels were not significantly
different in depleted cells compared to untreated controls cells
(supplementary material Fig. S3). These findings indicate that all
SNX9 subfamily members contribute to the efficient accumulation
of active myosin II at the cleavage furrow during ingression.
All three SNX9 members are required for endocytosis (Fig. 5)
and vesicle trafficking is required for the abscission phase of
cytokinesis. Therefore, we next assessed the role of these proteins
in regulating vesicle trafficking from the Golgi, recycling
endosomes (RE), lysosomes and endoplasmic reticulum (ER) to
the ICB. In untreated cells, GM130 (a Golgi marker) was observed
as a network of punctate structures distributed throughout the
cytoplasm in metaphase (Fig. 8A, Met), consistent with the early
stages of Golgi reformation (Sutterlin and Colanzi, 2010). During
cytokinesis, GM130 accumulated in two pools in each of the nascent
daughter cells (Fig. 8A, Cyto), as previously reported (Gaietta et al.,
2006). Depletion of SNX9 resulted in an accumulation of GM130 at
the inner pool, as indicated by an increase in the ratio of the
fluorescence intensity of the inner pool:fluorescence intensity of the
outer pool in each nascent daughter cell (Fig. 8A, bottom panel;Fig. 8B). The overall protein level of GM130 was unaffected by thedepletion of SNX9 during metaphase (supplementary material Fig.
S4A) or cytokinesis (supplementary material Fig. S4B), suggestingthat SNX9 is involved in vesicle trafficking from the Golgi to theICB. Depletion of SNX18 and SNX33 had no effect on thedistribution and the level of GM130 during metaphase and
cytokinesis (Fig. 8A,B; supplementary material Fig. S4).
We next examined the recycling endosomes (RE). The REmarker, Rab11, localized at the spindle poles and the cleavage
furrow during telophase and along the ICB during cytokinesis inuntreated cells (Fig. 8C), as previously observed (Wilson et al.,2005). Depletion of any one of the SNX9 members disruptedRab11 localization at both mitotic stages (Fig. 8C–E) but did not
affect the overall level of Rab11 in telophase (supplementarymaterial Fig. S5A) or cytokinesis (supplementary material Fig.S5B). In contrast, the level and localization of PDI (an ER
marker) and Lamp1 (a lysosome marker) were unaffected by thedepletion of these proteins (supplementary material Figs S6, S7).We conclude that all three SNX9 members are required for
localization of the recycling endosomes to the cleavage furrowand ICB during cytokinesis. SNX9 plays an additional role in theaccumulation of Golgi at the ICB during abscission.
DiscussionIn this study, we confirmed the requirement of SNX33 for mitoticprogression and completion. In addition, our findings show that its
two closest-related SNX members, SNX9 and SNX18, also playimportant roles in mitosis. Collectively, these three SNX9subfamily members are essential for efficient progression andcompletion of cytokinesis. These proteins are also required for
endocytosis during these mitotic phases, suggesting that theycontribute to vesicle trafficking during cytokinesis. Indeed, allthree proteins are required for the correct localization of recycling
endosomes during cytokinesis. SNX9 also contributes to thelocalization of Golgi to the ICB. Our additional major findingindicates that SNX9 is also required for chromosome alignment
and segregation during metaphase and anaphase. SNX9 does notparticipate in endocytosis during metaphase, suggesting that itsrole in this process is independent of its endocytic function. Thesefindings demonstrate that SNX9 subfamily proteins participate in
multiple stages throughout mitosis in both endocytic-dependentand -independent manners.
Only SNX9 was shown to have a role during metaphase. The
increased width of the metaphase plate observed in SNX9-depletedcells supports a role for SNX9 in chromosome alignment.Consistent with this role, it is found to be enriched at the spindlepoles during prometaphase. Clathrin is a major binding partner for
SNX9 in endocytosis (Lundmark and Carlsson, 2003) and is alsorequired for chromosome alignment and segregation (Royle et al.,2005). Like SNX9, its role in this process is independent of its role
in endocytosis (Royle et al., 2005). During metaphase, clathrinlocalizes to the mitotic spindle and spindle poles (Royle et al.,2005). Depletion or mutation of clathrin leads to destabilization of
the kinetochore fibres, which results in a defect in chromosomealignment and delay in chromosome segregation (Royle et al.,2005). This results in persistent activation of the spindle assembly
checkpoint. It will be interesting to determine if SNX9 is alsoinvolved in stabilizing kinetochore fibres and if it co-operates withclathrin at the mitotic spindle for chromosome alignment and
Fig. 7. SNX9 subfamily members regulate active myosin II localization
during anaphase and telophase. (A) Representative microscopy images of
MRLCS19 (red) localization in untreated and SNX9-subfamily-depleted HeLa
cells during anaphase and telophase. Schematic diagrams (right) show the
regions selected for fluorescence intensity quantification in B and C. DNA is
shown in blue. (B,C) Ratio of fluorescence intensity of MRLCS19 at the
ingressing furrow/polar region during anaphase (B) and telophase (C). Values
are means 6 s.e.m. from two independent experiments. n55–10 cells per
sample in each experiment. ns, not significant. *P,0.05; **P,0.01;
***P,0.001 (one-way ANOVA); Scale bars: 10 mm.
Journal of Cell Science 125 (18)4378

Journ
alof
Cell
Scie
nce
segregation, thus providing further mechanistic insight into the
non-endocytic mitotic function(s) of these proteins.
All SNX9 subfamily members participate in cytokinesis. Our
findings suggest that these proteins function in an endocytic-
dependent manner during this mitotic phase. However, they may
also participate in cytokinesis in an endocytic-independent
manner. Consistent with this idea, depletion of any of the three
SNX9 members prevented accumulation of active myosin II at the
cleavage furrow and delayed progression through the ingression
phase. SNX33 has previously been shown to be required for
cytokinesis and its role in this process was hypothesized to be due
to its ability to regulate actin polymerization via interaction with
WASp (Badour et al., 2007; Lundmark and Carlsson, 2003; Zhang
et al., 2009). SNX9 and SNX18 also bind WASp and regulate actin
dynamics (Badour et al., 2007). During clathrin-mediated
endocytosis, SNX9 is thought to recruit N-WASP to the plasma
membrane, which in turns stimulates actin recruitment and
polymerization (Cullen, 2008). Thus, it is possible that SNX9
and its subfamily members may contribute to the recruitment of
WASp and actin polymerization for efficient formation and
activity of the actinomyosin II contractile ring at the ingressing
cleavage furrow. In line with this idea, depletion of any one of the
three SNX9 members resulted in disruption of MRLCS19 cleavage
furrow localization, which is predicted to disrupt contractile ring
function.
The role of SNX9 members during cytokinesis appears to be
dependent on their endocytic activity. Supporting this idea, we
have shown that depletion of any one of these proteins delays or
blocks cytokinesis and also blocks Tfn uptake during this
process. Endocytosis is thought to contribute to the generation
of recycling endosomes that are delivered to the ICB for
abscission. We show that Rab11-positive recycling endosomes do
not accumulate at the cleavage furrow and ICB in cytokinetic
cells depleted of the SNX9 subfamily members. This could be
due to a block in (i) the generation of recycling endosomes, (ii)
endocytic activity at internalized endosomes or (iii) recruitment
of vesicles derived from recycling endosomes to the cleavage
furrow and ICB.
Fig. 8. SNX9 subfamily members regulate membrane
trafficking during cytokinesis. (A,B) Untreated and SNX9-,
SNX18- and SNX33-depleted HeLa cells at the indicated
mitotic phases were stained for the Golgi marker GM130
(red). Representative microscopy images of GM130
localization in untreated and SNX9-subfamily-depleted cells
during metaphase and cytokinesis are shown in A. The graph
in B shows the means 6 s.e.m. (from two independent
experiments) of the amount of GM130 in each sample,
expressed as ratio of fluorescence intensity at the inner pool to
that at the outer pool in each nascent daughter cell as
illustrated in the schematic diagram (bottom panel in A).
n.34 per sample. (C) Representative microscopy images of
Rab11 localization (red) in untreated HeLa cells and in HeLa
cells depleted of SNX9, SNX18 or SNX33 during telophase
and cytokinesis. DNA is shown in blue. (D,E) Graphs show
the localization of Rab11 during telophase (D) and cytokinesis
(E) in cells analysed in C. Values are means 6 s.e.m. from
two independent experiments.; n.20 per sample. ns, not
significant; ***P,0.001 (one-way ANOVA). Scale bars:
10 mm.
SNX9 subfamily is required for mitosis 4379

Journ
alof
Cell
Scie
nce
SNX9 and SNX33 may also participate in abscission more
directly. We show that these proteins localise to the FMRs and
MR within the ICB. We have recently shown that dynII locates to
the FMRs (Chircop et al., 2010) and functions here in an
endocytic-independent manner for abscission (Chircop et al.,
2011b). Numerous proteins that locate to the MR are involved in
protein and vesicular recruitment. Some of these include the
endocytic proteins, Arf6, and components of the ESCRT
machinery. Thus, it is possible that SNX9 and SNX33 are
involved in recruiting key proteins and/or vesicles to the
abscission site. Not only did the depletion of all three SNX9
members prevent recruitment of Rab11-positive recycling
endosomes to the ICB, but depletion of SNX9 also caused an
accumulation of Golgi at the edge of the ICB. We hypothesize
that SNX9 is required for the recruitment of Golgi-derived
vesicles to the ICB. This was highly specific as the localization of
vesicles derived from lysosomes or the ER was unaffected. The
exact role of vesicles at the ICB is unclear, however they are
proposed to (i) provide extra total cell surface area, an increase of
at least 25% is required to complete division (Boucrot and
Kirchhausen, 2007), (ii) deliver critical cytokinetic proteins to the
abscission site (Low et al., 2003), and/or (iii) be directly involved
in compound fusion, whereby numerous vesicles fuse with the
plasma membrane during abscission to separate the daughter
cells (Goss and Toomre, 2008; Gromley et al., 2005; Low et al.,
2003; Prekeris and Gould, 2008). The role of SNX9 subfamily
members in this process is critical to understand as cytokinesis
failure leads to aneuploidy.
Overall, our findings revealed that the three SNX9 subfamily
proteins are involved in ensuring efficient progression and
completion of mitosis at several distinct stages. We have shown
that all three SNX9 subfamily members most likely function
during cytokinesis in an endocytic-dependent manner. We have
further revealed a non-endocytic function for SNX9, specifically
in chromosome alignment.
Materials and MethodsCell culture and transfection
HeLa human cervical carcinoma cells and HeLa cells stably expressing GFP–H2Bwere maintained in RPMI 1640 medium. U2OS human osteosarcoma cells weremaintained in DMEM medium. All medium were supplemented with 10% foetalbovine serum (FBS) and cells were grown at 37 C in a humidified 5% CO2
atmosphere. Cells were seeded at 50–60% confluence (16105 cells per 10 cm dish,0.56105 cells per well of a 6-well plate; 0.26105 cells per well of a 12-well plate).For siRNA analyses, cells were transfected with 1000 pmol of siRNA (per 10 cmdish for immunoblotting), 200 pmol of siRNA (per well of a 6-well plate forimmunofluorescence and time-lapse microscopy experiments) or 100 pmol (perwell of a 12-well plate for immunofluorescence and time-lapse microscopyexperiments). For DNA transfections, 1.5 mg of the indicated plasmid DNA wasused in each well of a 6-well plate. In both cases, cells were transfected withLipofectamine 2000 (Invitrogen) according to the manufacturer’s instructions.
Cell synchronization and treatment
HeLa, HeLa-GFP-H2B and U2OS cells grown on glass coverslips weresynchronized at the G2–M border by treatment with the selective Cdk1 small-molecule inhibitor RO-3306 (9 mM) for at least 18 h. Cells were allowed to progressthrough mitosis upon RO-3306 wash out. Following RO-3306 wash out, cells wereincubated at 37 C/5% CO2 for 80 min (metaphase), 2.5 h (cytokinesis) or 6 h(multinucleation and cytokinesis accumulation scoring).
Plasmid constructions
GFP-SNX9 was a gift from Sandra Schmid (Soulet et al., 2005). HA-SNX18 andGFP-SNX33 constructs were kindly provided by Stefan F. Lichtenthaler (Dislichet al., 2011). The siRNA target sequences in the sense orientation for the followingproteins are: SNX9-1: 59-AACCUACUAACACUAAUCGAU-39 (Lundmark andCarlsson, 2004); SNX9-2: 59-AACAGUCGUGCUAGUUCCUCA-39 (Shin et al.,2008); SNX18-1: 59-CGUCAUGGACCUAUUAGCGCUGUAU-39; SNX18-2: 59-
CACCGACGAGAAAGCCUGGAAUU-39 (Qiagen); SNX33-1: 59-CAAGAUCG-CUGAGACAUACUCCA-39; SNX33-2: 59-CACACGGGCCGUACCUAUUGA-A-39; Luciferase: 59-CGUACGCGGAAUACUUCGA-39.
Rescue experiment
HeLa cells were transfected with siRNA targeted the indicated SNX9 subfamilymember using Neon Transfection System as per manufacturer’s instruction(Invitrogen). On the next day, the indicated plasmid construct (GFP-SNX9, HA-SNX18 and GFP-SNX33) were transfected using the Neon Transfection System(Invitrogen) to overexpress the relevant SNX9 member in the appropriate siRNA-depleted cells. For western blot analyses, cells were collected 24 h post-transfection.For multinucleation analyses, cells were synchronized with RO-3306 for 18 hfollowed by RO-3306 wash out for 6 h.
Time-lapse microscopy analyses
Immediately following release into the cell cycle G2–M synchronized HeLa orHeLa-GFP–H2B cells were viewed with an Olympus IX81 inverted microscopeand a time-lapse series was acquired using a fully motorized stage, 106objective,and Metamorph software using the Time-lapse modules as previously described(Chircop et al., 2010; Joshi et al., 2010). Temperature control was achieved usingthe Incubator XL, providing a humidified atmosphere with 5% CO2. Imaging wasperformed for 20 h with a lapse time of 2 min or 10 min as indicated.
Immunofluorescence microscopy
Cells were fixed in ice-cold 100% methanol for 3 min at 220 C and blocked in 3%bovine serum albumin (BSA)/PBS for 45 min prior to incubation with the primaryantibody. For membrane trafficking studies, cells were fixed in ice-cold 4% PFA inPBS for 20 min and then permeabilised with 0.2% Triton X-100 in PBS for 15 minbefore blocking in 3% BSA/PBS for 45 min prior to primary antibody incubation.The following antibodies were used: anti-SH3PX1 (anti-SNX9, NB100-2813,Novus Biological), anti-SNX18 (GTX106319, GeneTex), anti-SH3PX3 (anti-SNX33, H00257364-D01P, Abnova), anti-Lamp1 (ab25630, Abcam), anti-PDI(ab2792, Abcam), anti-Rab11 (610657, BD Transduction), anti-phospho-myosinlight chain S19 (3675S, Cell Signaling), anti-GM130 (610822, BD Transduction),anti-dynI phospho-S778 which detects dynII phospho-S764 (Anggono et al.,2006), anti-a-tubulin (clone DM1A; Sigma) and anti-c-tubulin (GTU88; Sigma).Fluorescein- or Texas-Red-conjugated AffiniPure secondary antibodies (JacksonImmunoResearch Laboratories, Inc.) were then applied. Cell nuclei werecounterstained with DAPI (4969-diamidino-2-phenylindole; Sigma). Cells werewashed three times with PBS between each step except for after blocking. Cellswere viewed and scored with a fluorescence microscope (Olympus IX80).Fluorescence images were captured under an Olympus IX81 inverted microscopeusing 606or 1006oil immersion lenses and deconvolved using AutoDeblur v9.3(AutoQuant Imaging, Watervliet, NY). Integrated fluorescence intensity wasmeasured using Metamorph software (Version 7.7.0.0.).
Immunoblotting
Cellular extracts were prepared as described previously (Fabbro et al., 2004). Inbrief, cells were collected by centrifugation, washed with PBS, then resuspended inice-cold lysis buffer for sonication [25 mM Tris-HCl pH 7.4, 150 mM NaCl,1 mM EDTA, 1 mM EGTA, 1 mM PMSF, 1% Triton X-100, and EDTA-freeComplete protease inhibitor cocktail (Roche)] followed by incubation on ice for30 min. The supernatant was collected following centrifugation at 13,000 rpmfor 30 min at 4 C. Cell lysates (200 mg) were fractionated by SDS-PAGE forimmunoblot analysis with the following antibodies: anti-SNX9 (Santa Cruz), anti-SNX18 (GeneTex), anti-SNX33 [a gift from Stefan F. Lichtenthaler (Dislich et al.,2011)], anti-GFP (Clonetech), anti-HA.11 (Covance) and anti-c-tubulin (Sigma).Antibody bound to the indicated protein was detected by incubation with ahorseradish peroxidase-conjugated secondary antibody (Jackson ImmunoResearchLaboratories, Inc.). Blotted proteins were visualized using the ECL detectionsystem (Pierce).
Endocytosis assay
Quantitative high-throughput receptor-mediated endocytosis (RME) assays wereperformed as previously described (Hill et al., 2009; Odell et al., 2010) usingTransferrin (Tfn) conjugated to Texas Red in untreated and siRNA-transfected HeLacells for 10 min. Cells were then washed with ice-cold PBS and acid washed (0.2 Macetic acid/0.5 M NaCl, pH 2.8) for 15 min on ice. Cells were then fixed with ice-cold 4% PFA in PBS for 15 min and processed for immunofluorescence microscopyanalysis as described above. Cells were imaged on an ImageXpressmicro platereader(Molecular Devices) and Tfn uptake was analysed using MetaXpress software(V3.1.0.79, Molecular Devices) by calculating the cell:vesicle integrated intensity.
AcknowledgementsWe thank Sandra Schmid for providing the GFP-SNX9 constructs andStefan F. Lichtenthaler for providing the anti-SNX33 antibody, as well
Journal of Cell Science 125 (18)4380

Journ
alof
Cell
Scie
nce
as HA-SNX18 and GFP-SNX33 constructs. Scott L. Page is thankedfor his technical assistance. We also thank Phillip J. Robinson, RoseBoutros and Scott L. Page for critic reading of the manuscript.
FundingThis work was supported by grants from the National Health andMedical Research Council of Australia [grant number 477102 toM.C.] and the National Health and Medical Research CouncilBiomedical Career Development Award Fellowship [grant number477104 to M.C.].
Supplementary material available online at
http://jcs.biologists.org/lookup/suppl/doi:10.1242/jcs.105981/-/DC1
ReferencesAlto, N. M., Weflen, A. W., Rardin, M. J., Yarar, D., Lazar, C. S., Tonikian, R.,
Koller, A., Taylor, S. S., Boone, C., Sidhu, S. S. et al. (2007). The type III effectorEspF coordinates membrane trafficking by the spatiotemporal activation of two
eukaryotic signaling pathways. J. Cell Biol. 178, 1265-1278.
Anggono, V., Smillie, K. J., Graham, M. E., Valova, V. A., Cousin, M. A. and
Robinson, P. J. (2006). Syndapin I is the phosphorylation-regulated dynamin Ipartner in synaptic vesicle endocytosis. Nat. Neurosci. 9, 752-760.
Badour, K., McGavin, M. K., Zhang, J., Freeman, S., Vieira, C., Filipp, D., Julius,
M., Mills, G. B. and Siminovitch, K. A. (2007). Interaction of the Wiskott-Aldrichsyndrome protein with sorting nexin 9 is required for CD28 endocytosis and
cosignaling in T cells. Proc. Natl. Acad. Sci. USA 104, 1593-1598.
Boucrot, E. and Kirchhausen, T. (2007). Endosomal recycling controls plasmamembrane area during mitosis. Proc. Natl. Acad. Sci. USA 104, 7939-7944.
Carlton, J., Bujny, M., Peter, B. J., Oorschot, V. M., Rutherford, A., Mellor, H.,
Klumperman, J., McMahon, H. T. and Cullen, P. J. (2004). Sorting nexin-1mediates tubular endosome-to-TGN transport through coincidence sensing of high-curvature membranes and 3-phosphoinositides. Curr. Biol. 14, 1791-1800.
Carlton, J., Bujny, M., Rutherford, A. and Cullen, P. (2005). Sorting nexins –unifying trends and new perspectives. Traffic 6, 75-82.
Chen, H., Slepnev, V. I., Di Fiore, P. P. and De Camilli, P. (1999). The interaction ofepsin and Eps15 with the clathrin adaptor AP-2 is inhibited by mitotic phosphorylationand enhanced by stimulation-dependent dephosphorylation in nerve terminals.J. Biol. Chem. 274, 3257-3260.
Chircop, M., Malladi, C. S., Lian, A. T., Page, S. L., Zavortink, M., Gordon, C. P.,
McCluskey, A. and Robinson, P. J. (2010). Calcineurin activity is required for thecompletion of cytokinesis. Cell. Mol. Life Sci. 67, 3725-3737.
Chircop, M., Perera, S., Mariana, A., Lau, H., Ma, M. P., Gilbert, J., Jones, N. C.,
Gordon, C. P., Young, K. A., Morokoff, A. et al. (2011a). Inhibition of dynamin bydynole 34-2 induces cell death following cytokinesis failure in cancer cells. Mol.
Cancer Ther. 10, 1553-1562.
Chircop, M., Sarcevic, B., Larsen, M. R., Malladi, C. S., Chau, N., Zavortink, M.,
Smith, C. M., Quan, A., Anggono, V., Hains, P. G. et al. (2011b). Phosphorylationof dynamin II at serine-764 is associated with cytokinesis. Biochim. Biophys. Acta
1813, 1689-1699.
Cullen, P. J. (2008). Endosomal sorting and signalling: an emerging role for sortingnexins. Nat. Rev. Mol. Cell Biol. 9, 574-582.
Dephoure, N., Zhou, C., Villen, J., Beausoleil, S. A., Bakalarski, C. E., Elledge, S. J.
and Gygi, S. P. (2008). A quantitative atlas of mitotic phosphorylation. Proc. Natl.
Acad. Sci. USA 105, 10762-10767.
Dislich, B., Than, M. E. and Lichtenthaler, S. F. (2011). Specific amino acids in theBAR domain allow homodimerization and prevent heterodimerization of sortingnexin 33. Biochem. J. 433, 75-83.
Fabbro, M., Savage, K., Hobson, K., Deans, A. J., Powell, S. N., McArthur, G. A.
and Khanna, K. K. (2004). BRCA1-BARD1 complexes are required for p53Ser-15phosphorylation and a G1/S arrest following ionizing radiation-induced DNAdamage. J. Biol. Chem. 279, 31251-31258.
Fielding, A. B., Schonteich, E., Matheson, J., Wilson, G., Yu, X., Hickson, G. R.,
Srivastava, S., Baldwin, S. A., Prekeris, R. and Gould, G. W. (2005). Rab11-FIP3and FIP4 interact with Arf6 and the exocyst to control membrane traffic incytokinesis. EMBO J. 24, 3389-3399.
Gaietta, G. M., Giepmans, B. N., Deerinck, T. J., Smith, W. B., Ngan, L., Llopis, J.,
Adams, S. R., Tsien, R. Y. and Ellisman, M. H. (2006). Golgi twins in late mitosisrevealed by genetically encoded tags for live cell imaging and correlated electronmicroscopy. Proc. Natl. Acad. Sci. USA 103, 17777-17782.
Glotzer, M. (2005). The molecular requirements for cytokinesis. Science 307, 1735-1739.
Goss, J. W. and Toomre, D. K. (2008). Both daughter cells traffic and exocytosemembrane at the cleavage furrow during mammalian cytokinesis. J. Cell Biol. 181,1047-1054.
Gromley, A., Yeaman, C., Rosa, J., Redick, S., Chen, C. T., Mirabelle, S., Guha, M.,
Sillibourne, J. and Doxsey, S. J. (2005). Centriolin anchoring of exocyst andSNARE complexes at the midbody is required for secretory-vesicle-mediatedabscission. Cell 123, 75-87.
Haberg, K., Lundmark, R. and Carlsson, S. R. (2008). SNX18 is an SNX9 paralogthat acts as a membrane tubulator in AP-1-positive endosomal trafficking. J. Cell Sci.
121, 1495-1505.
He, J., Xu, J., Xu, X. X. and Hall, R. A. (2003). Cell cycle-dependent phosphorylationof Disabled-2 by cdc2. Oncogene 22, 4524-4530.
Heiseke, A., Schobel, S., Lichtenthaler, S. F., Vorberg, I., Groschup, M. H.,
Kretzschmar, H., Schatzl, H. M. and Nunziante, M. (2008). The novel sortingnexin SNX33 interferes with cellular PrP formation by modulation of PrP shedding.Traffic 9, 1116-1129.
Hill, T. A., Gordon, C. P., McGeachie, A. B., Venn-Brown, B., Odell, L. R., Chau,
N., Quan, A., Mariana, A., Sakoff, J. A., Chircop, M. et al. (2009). Inhibition ofdynamin mediated endocytosis by the dynoles – synthesis and functional activity of afamily of indoles. J. Med. Chem. 52, 3762-3773.
Ikebe, M., Koretz, J. and Hartshorne, D. J. (1988). Effects of phosphorylation of lightchain residues threonine 18 and serine 19 on the properties and conformation ofsmooth muscle myosin. J. Biol. Chem. 263, 6432-6437.
Joshi, S., Perera, S., Gilbert, J., Smith, C. M., Mariana, A., Gordon, C. P., Sakoff,
J. A., McCluskey, A., Robinson, P. J., Braithwaite, A. W. et al. (2010). Thedynamin inhibitors MiTMAB and OcTMAB induce cytokinesis failure and inhibitcell proliferation in human cancer cells. Mol. Cancer Ther. 9, 1995-2006.
Julian, M., Tollon, Y., Lajoie-Mazenc, I., Moisand, A., Mazarguil, H., Puget, A. and
Wright, M. (1993). gamma-tubulin participates in the formation of the midbodyduring cytokinesis in mammalian cells. J. Cell Sci. 105, 145-156.
Kariya, K., Koyama, S., Nakashima, S., Oshiro, T., Morinaka, K. and Kikuchi,
A. (2000). Regulation of complex formation of POB1/epsin/adaptor protein complex2 by mitotic phosphorylation. J. Biol. Chem. 275, 18399-18406.
Komatsu, S., Yano, T., Shibata, M., Tuft, R. A. and Ikebe, M. (2000). Effects of theregulatory light chain phosphorylation of myosin II on mitosis and cytokinesis ofmammalian cells. J. Biol. Chem. 275, 34512-34520.
Liu, Z. and Zheng, Y. (2009). A requirement for epsin in mitotic membrane and spindleorganization. J. Cell Biol. 186, 473-480.
Low, S. H., Li, X., Miura, M., Kudo, N., Quinones, B. and Weimbs, T. (2003).Syntaxin 2 and endobrevin are required for the terminal step of cytokinesis inmammalian cells. Dev. Cell 4, 753-759.
Lundmark, R. and Carlsson, S. R. (2002). The beta-appendages of the four adaptor-protein (AP) complexes: structure and binding properties, and identification of sortingnexin 9 as an accessory protein to AP-2. Biochem. J. 362, 597-607.
Lundmark, R. and Carlsson, S. R. (2003). Sorting nexin 9 participates in clathrin-mediated endocytosis through interactions with the core components. J. Biol. Chem.
278, 46772-46781.
Lundmark, R. and Carlsson, S. R. (2004). Regulated membrane recruitment ofdynamin-2 mediated by sorting nexin 9. J. Biol. Chem. 279, 42694-42702.
Mayer, B. J. (2001). SH3 domains: complexity in moderation. J. Cell Sci. 114, 1253-1263.
Odell, L. R., Howan, D., Gordon, C. P., Robertson, M. J., Chau, N., Mariana,A., Whiting, A. E., Abagyan, R., Daniel, J. A., Gorgani, N. N. et al. (2010). Thepthaladyns: GTP competitive inhibitors of dynamin I and II GTPase derived fromvirtual screening. J. Med. Chem. 53, 5267-5280.
Park, J., Kim, Y., Lee, S., Park, J. J., Park, Z. Y., Sun, W., Kim, H. and Chang, S.
(2010). SNX18 shares a redundant role with SNX9 and modulates endocytictrafficking at the plasma membrane. J. Cell Sci. 123, 1742-1750.
Prekeris, R. and Gould, G. W. (2008). Breaking up is hard to do - membrane traffic incytokinesis. J. Cell Sci. 121, 1569-1576.
Royle, S. J., Bright, N. A. and Lagnado, L. (2005). Clathrin is required for the functionof the mitotic spindle. Nature 434, 1152-1157.
Schobel, S., Neumann, S., Hertweck, M., Dislich, B., Kuhn, P. H., Kremmer,E., Seed, B., Baumeister, R., Haass, C. and Lichtenthaler, S. F. (2008). A novelsorting nexin modulates endocytic trafficking and alpha-secretase cleavage of theamyloid precursor protein. J. Biol. Chem. 283, 14257-14268.
Scholey, J. M., Taylor, K. A. and Kendrick-Jones, J. (1980). Regulation of non-muscle myosin assembly by calmodulin-dependent light chain kinase. Nature 287,233-235.
Schweitzer, J. K., Burke, E. E., Goodson, H. V. and D’Souza-Schorey, C. (2005).Endocytosis resumes during late mitosis and is required for cytokinesis. J. Biol.
Chem. 280, 41628-41635.
Seet, L. F. and Hong, W. (2006). The Phox (PX) domain proteins and membrane traffic.Biochim. Biophys. Acta 1761, 878-896.
Shimizu, H., Nagamori, I., Yabuta, N. and Nojima, H. (2009). GAK, a regulator ofclathrin-mediated membrane traffic, also controls centrosome integrity and chromosomecongression. J. Cell Sci. 122, 3145-3152.
Shin, N., Ahn, N., Chang-Ileto, B., Park, J., Takei, K., Ahn, S. G., Kim, S. A.,
Di Paolo, G. and Chang, S. (2008). SNX9 regulates tubular invagination of theplasma membrane through interaction with actin cytoskeleton and dynamin 2. J. Cell
Sci. 121, 1252-1263.
Soulet, F., Yarar, D., Leonard, M. and Schmid, S. L. (2005). SNX9 regulates dynaminassembly and is required for efficient clathrin-mediated endocytosis. Mol. Biol. Cell
16, 2058-2067.
Sutterlin, C. and Colanzi, A. (2010). The Golgi and the centrosome: building afunctional partnership. J. Cell Biol. 188, 621-628.
Warner, A. K., Keen, J. H. and Wang, Y. L. (2006). Dynamics of membrane clathrin-coated structures during cytokinesis. Traffic 7, 205-215.
Wilson, G. M., Fielding, A. B., Simon, G. C., Yu, X., Andrews, P. D., Hames, R. S.,
Frey, A. M., Peden, A. A., Gould, G. W. and Prekeris, R. (2005). The FIP3-Rab11
SNX9 subfamily is required for mitosis 4381

Journ
alof
Cell
Scie
nce
protein complex regulates recycling endosome targeting to the cleavage furrow duringlate cytokinesis. Mol. Biol. Cell 16, 849-860.
Yamakita, Y., Yamashiro, S. and Matsumura, F. (1994). In vivo phosphorylation ofregulatory light chain of myosin II during mitosis of cultured cells. J. Cell Biol. 124,129-137.
Yarar, D., Waterman-Storer, C. M. and Schmid, S. L. (2007). SNX9 couples actinassembly to phosphoinositide signals and is required for membrane remodeling duringendocytosis. Dev. Cell 13, 43-56.
Yarar, D., Surka, M. C., Leonard, M. C. and Schmid, S. L. (2008). SNX9 activitiesare regulated by multiple phosphoinositides through both PX and BAR domains.Traffic 9, 133-146.
Yeow-Fong, L., Lim, L. and Manser, E. (2005). SNX9 as an adaptor for linkingsynaptojanin-1 to the Cdc42 effector ACK1. FEBS Lett. 579, 5040-5048.
Zhang, J., Zhang, X., Guo, Y., Xu, L. and Pei, D. (2009). Sorting nexin 33 inducesmammalian cell micronucleated phenotype and actin polymerization by interactingwith Wiskott-Aldrich syndrome protein. J. Biol. Chem. 284, 21659-21669.
Journal of Cell Science 125 (18)4382

Fig. S1. Validating the specificity of SNX9, SNX18 and SNX33 antibodies by immunofluorescence microscopy. HeLa cells depleted of the indicated SNX9 subfamily member were stained with anti-SNX9, anti-SNX18 and anti-SNX33 antibodies (green). No detectable staining was observed in cells at the indicated cell cycle stage illustrating that these antibodies have no detectable non-specific cross-reactivity. DNA shown in blue. Scale bars: 10 mm.

Fig. S2. SNX9, SNX18 and SNX33 cause cytokinesis failure in U2OS cells. (A-C) U2OS cells were either untreated (-), or transfected with luciferase siRNA (Luc) or siRNA targeting SNX9 (A), SNX18 (B) and SNX33 (C). At 72 h post-transfection, protein lysates (200 mg) were immunoblotted with anti-SNX9, anti-SNX18 or anti-SNX33 antibodies respectively. g-tubulin served as a loading control. (D,E) Cells were transfected as described in A-C followed by synchronization at the G2/M boundary using RO-3306. Upon RO-3306 wash out cells were released for 6 h, fixed and immunostained for a-tubulin and scored for multinucleated cells (D) and cytokinetic cells (E). The graphs show the mean ± s.e.m. from three independent experiments where n>200 cells were scored for each sample in each experiment. *P<0.05; **P<0.01; ***P<0.001 (one-way ANOVA).

Fig. S3. Expression level of MRLCS19 in SNX9 subfamily-depleted HeLa cells. Quantification of the total MRLCS19 fluorescence within the whole cell as a measure of its expression level in untreated and siRNA-treated HeLa cells during anaphase (A) and telophase (B). Graphs represent the mean ± s.e.m. from two independent experiments. n=5-10 cells per sample in each experiment. ns, not significant. (one-way ANOVA).

Fig. S4. Effect of depletion of SNX9 subfamily members on the expression level of GM130 in HeLa cells during mitosis. (A,B) Graphs represent the mean fluorescence of GM130 in metaphase (A) or cytokinesis (B). Values represent the means ± s.e.m. from two independent experiments. n>30 per sample. ns, not significant (one-way ANOVA).

Fig. S5. Effect of depletion of SNX9 subfamily members on the expression level of Rab11 in HeLa cells during mitosis. (A,B) Graphs represent the mean fluorescence of Rab11 in telophase (A) or cytokinesis (B). Values represent the means ± s.e.m. from two independent experiments. n>20 per sample. ns, not significant (one-way ANOVA).

Fig. S6. Effect of depletion of SNX9 subfamily members on the distribution of lyosomes during cytokinesis. (A) Representative microscopy images of Lamp1 localization in untreated and SNX9 subfamily-depleted cells during telophase and cytokinesis. DNA is shown in blue. (B,C) The graphs represent the distribution of Lamp1 in HeLa cells as shown in A, expressed as a ratio of fluorescence intensity of the inner pool compared to the outer pool in each nascent daughter cell during telophase (B) and cytokinesis (C). (D,E) Graphs illustrate the overall fluorescence intensity of Lamp1 within the whole cell in HeLa cells depleted of SNX9 subfamily members during telophase (D) and cytokinesis (E). Scale bars: 10 mm. Values represent the mean ± s.e.m. from two independent experiments. n>6 per sample. ns, not significant (one-way ANOVA).
Fig. S7. The expression level and localization of endoplasmic reticulum in SNX9 subfamily-depleted HeLa cells during cytokinesis. (A) Representative microscopy images of PDI localization (red) in untreated HeLa cells and in HeLa cells depleted of the indicated SNX9 subfamily member during telophase and cytokinesis. DNA is shown in blue. (B,C) Graphs show the overall fluorescence intensity of PDI within the whole cell in HeLa cells depleted of SNX9 subfamily members during telophase (B) and cytokinesis (C). Scale bars: 10 mm. Values represent the mean ± s.e.m. from two independent experiments. n>10 per sample. ns, not significant (one-way ANOVA).





























