Embed Size (px)
Citation preview

Spectral reflectance of coral reef bottom-types worldwide and implications
for coral reef remote sensing
Eric J. Hochberga,*, Marlin J. Atkinsona, Serge Andrefouetb
aUniversity of Hawaii, School of Ocean and Earth Science and Technology, Hawaii Institute of Marine Biology, P.O. Box 1346, Kaneohe, HI 96744, USAbUniversity of South Florida, College of Marine Science, Institute for Marine Remote Sensing, St. Petersburg, FL, USA
Received 10 June 2002; received in revised form 11 October 2002; accepted 15 October 2002
Abstract
Coral reef benthic communities are mosaics of individual bottom-types that are distinguished by their taxonomic composition and
functional roles in the ecosystem. Knowledge of community structure is essential to understanding many reef processes. To develop
techniques for identification and mapping of reef bottom-types using remote sensing, we measured 13,100 in situ optical reflectance spectra
(400–700 nm, 1-nm intervals) of 12 basic reef bottom-types in the Atlantic, Pacific, and Indian Oceans: fleshy (1) brown, (2) green, and (3)
red algae; non-fleshy (4) encrusting calcareous and (5) turf algae; (6) bleached, (7) blue, and (8) brown hermatypic coral; (9) soft/gorgonian
coral; (10) seagrass; (11) terrigenous mud; and (12) carbonate sand. Each bottom-type exhibits characteristic spectral reflectance features that
are conservative across biogeographic regions. Most notable are the brightness of carbonate sand and local extrema near 570 nm in blue
(minimum) and brown (maximum) corals. Classification function analyses for the 12 bottom-types achieve mean accuracies of 83%, 76%,
and 71% for full-spectrum data (301-wavelength), 52-wavelength, and 14-wavelength subsets, respectively. The distinguishing spectral
features for the 12 bottom-types exist in well-defined, narrow (10–20 nm) wavelength ranges and are ubiquitous throughout the world. We
reason that spectral reflectance features arise primarily as a result of spectral absorption processes. Radiative transfer modeling shows that in
typically clear coral reef waters, dark substrates such as corals have a depth-of-detection limit on the order of 10–20 m. Our results provide
the foundation for design of a sensor with the purpose of assessing the global status of coral reefs.
Published by Elsevier Science Inc.
Keywords: Coral reef; Spectral reflectance; Remote sensing; Radiative transfer
1. Introduction
Coral reef benthic ecosystems are collections of distinctive
communities, which are distinguished by their characteristic
assemblages of organisms and substrates (Stoddart, 1969). A
benthic community’s structure is defined by its component
set of organisms and substrates; hereafter, we refer to these
fundamental elements of the community as ‘‘bottom-types.’’
Several reef communities may share bottom-types in com-
mon, but the bottom-types’ proportional contributions vary
both between and within communities. From this viewpoint,
all reef communities are simply combinations of some
comprehensive set of bottom-types.
Quantification of benthic community structure is central
to understanding coral reef ecosystem function. Community
structure determines rates of reef metabolism (Kinsey, 1985)
and indicates reef status (Connell, 1997). Different bottom-
types are important in life history strategies of reef-dwelling
organisms, e.g., as recruitment sites for coral larvae (Miller
et al., 2000) and juvenile fish (Light & Jones, 1997), and as
habitat for adult fish (Chabanet et al., 1997). Reef com-
munity structure exhibits tremendous spatial heterogeneity
over scales of centimeters to hundreds of meters and, in
contrast to phytoplankton and macrophyte communities, is
inherently stable on time scales of months to years (Budde-
meier & Smith, 1999). Reports of reef degradation world-
wide (Wilkinson, 2000) are now fueling interest in—and
debate over the causes of—temporal shifts in community
structure on longer time scales.
Conventional methods for determining benthic commun-
ity structure include the use of in situ quadrats, line transects,
and manta tows (Miller & Muller, 1999). These are not
feasible means for accurate determination of bottom-type
spatial distributions over large areas (Ginsburg, 1994).
Digital remote sensing is the most cost-effective approach
for acquiring such data (Mumby, Green, Edwards, & Clark,
0034-4257/03/$ - see front matter. Published by Elsevier Science Inc.
doi:10.1016/S0034-4257(02)00201-8
* Corresponding author. Tel.: +1-808-956-9108; fax: +1-808-956-7112.
E-mail address: [email protected] (E.J. Hochberg).
www.elsevier.com/locate/rse
Remote Sensing of Environment 85 (2003) 159–173

1999), and it is the only available tool to acquire such data on
a global scale. Accepting the need to quantify reef commun-
ity structure through time and over large areas, and accepting
the fact that remote sensing is the most promising tool for the
task, the next obvious step is to develop techniques for using
remote sensing data to detect, discriminate, and quantify the
spatial distributions of reef bottom-types.
Since its inception in the early 1970s, digital remote
sensing of coral reefs has generally followed a sensor-down
approach, in which scene-specific image statistics drive
image classification and interpretation. This approach is
intensely interactive and is largely a product of the fact that
the limited capabilities of available sensors have tended to
generate ambiguous image classification results, requiring
significant human interpretation. In truth, the sensor-down
approach has proven very useful at characterizing reef
geomorphology (reviewed extensively in Green et al.,
1996). With expert knowledge of local reef systems, the
same approach has also been successful for delineation of
whole communities, which are defined a priori to image
classification and, unfortunately, are semi-quantitative at
best (e.g., Andrefouet & Payri, 2001; Mumby, Green, Clark,
& Edwards, 1998). The few attempts to directly detect
fundamental bottom-types—and thus quantify community
structure—have utilized a reef-up approach, in which the
classifier statistics are defined independently of an image
data set (e.g., Hochberg & Atkinson, 2000). In this deter-
ministic approach, the reflectance spectra of bottom-types
are catalogued, and their characteristic features are targeted
through remote sensing. Since all community structures are
defined as subsets of a comprehensive set of bottom-types,
the classifier statistics are applicable across the geographic
area encompassed by the catalog of spectral reflectance data.
Whatever the approach, to be useful in mapping reef
community structure, a remote sensing system must have
the abilities to spatially resolve reef bottom-types and to spec-
trally discriminate between them. Several imaging systems
are currently operational and have specific strategies for coral
reef image acquisition. However, those systems that provide
more spectral information have poor spatial resolution. For
example, SeaWiFS has six narrow wavebands in the visible
spectrum, and MODIS has seven, but these sensors achieve
only 1-km and 250-m pixel resolution, respectively, preclud-
ing their use for spatially discriminating reef bottom-types or
even most reef communities (Andrefouet et al., 2003). Con-
versely, systems with the ability to spatially resolve reef
bottom-types (e.g., Landsat, Ikonos and Quickbird at 30-,
4-, and 2-m pixel resolutions, respectively) have wavebands
that are not optimized for reef bottom-type spectral discrim-
ination (Andrefouet, Muller-Karger, Hochberg, Hu, &
Carder, 2001; Hochberg & Atkinson, 2003).
It is generally agreed that reef bottom-types are spatially
resolvable by (or at least that they cover significant propor-
tions of) the spatial sampling units of current high-resolution
remote sensors (Mumby & Edwards, 2002). Given the 30-
year history of digital coral reef remote sensing, however,
there is surprisingly scant information on spectral reflectance
(R) for reef bottom-types globally. The following is a brief
review of the literature concerning R for coral reef benthos.
Maritorena, Morel, and Gentili (1994) made in vitro
reflectance measurements of carbonate sand, Sargassum
sp., Turbinaria sp., Boodlea sp., Porolithon onkodes, and
Corallinacea sp., but did not examine spectral differences
between the bottom-types. Miyazaki and Harashima (1993)
and Miyazaki, Nakatani, and Harashima (1995) made both
laboratory and in situ measurements of R for nine corals,
rocks, and sand. They visually inspected the spectra and
concluded there were minimal differences between those
three benthic community types. Mazel (1996) examined the
contribution of fluorescence to R, for which he had f 300 in
vivo measurements for 25 coral specimens (10 species).
Holden and LeDrew (1998) made a statistical investigation
on a total of 22 in situ reflectance measurements, representing
bleached and non-bleached Acropora, dead coral rubble, and
algae-covered dead corals. They applied cluster, principal
components and derivative analyses to show that statistically
significant spectral differences could be detected between
healthy Acropora and bleached/dead Acropora. Holden and
LeDrew (1999) measured 133 in situ R’s in Fiji and Sulawesi,
Indonesia for bleached coral, healthy massive corals, healthy
branching corals, dead coral debris, and algae-covered sur-
faces. They performed 181 individual univariate t-tests (one
for each wavelength measured) for each of the classes and
determined that there is no significant spectral difference
between geographic locations within each class. Similar
statistics indicated there is also no spectral difference attrib-
utable to coral morphology, but that there is a difference due
to coral health. Myers, Hardy, Mazel, and Dustan (1999)
reported on five in situ R’s for bleached and pigmented coral
(Montastrea cavernosa), a red crustose alga, a brown algae,
and a red alga (Rhipocephalus phoenix) at Lee Stocking
Island, Bahamas. Their objective was not to discriminate
between the bottom-types but to demonstrate an empirical
relationship between pigmentation and R. In Kaneohe Bay,
Oahu, Hawaii, Hochberg and Atkinson (2000) measured a
total of 247 in situ R’s of three coral species (Montipora
capitata, Porites compressa, Porites lobata), five algal spe-
cies (Dictyosphaeria cavernosa, Gracilaria salicornia, Hal-
imeda sp., Porolithon sp., Sargassum echinocarpum), and
three sand communities (fine-grained carbonate sand, sand
mixed with coral rubble, coral rubble). They used derivative
analysis and the multivariate techniques of stepwise wave-
length selection and linear discriminant analysis to show that
spectral separation of basic bottom-types (coral, algae, and
sand) is possible with as few as four non-contiguous narrow
wavebands. In the lagoon of Rangiroa, French Polynesia,
Clark, Mumby, Chisholm, Jaubert, and Andrefouet (2000)
measured f 1800 in situ R’s for 94 reef targets, including
coral (Porites, Pocillopora, and others), encrusting and turf
algae, and sand. Using derivative analysis, they determined
that it is possible to distinguish between three states of coral
mortality: live coral, coral dead for 6 months, and coral dead
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173160

for >6 months. Joyce and Phinn (2002) analyzed R from
corals with three distinct morphologies (open branching,
closed branching, and table) and determined that the three
are distinguishable by the differing amount of shadow each
morphology generates. Finally, Minghelli-Roman, Chis-
holm, Marchioretti, and Jaubert (2002) measured R for 152
coral colonies in the Red Sea and were able to discriminate
between 14 genera of hard and soft corals using ratios of only
six wavebands. Hedley and Mumby (2002) provide a review
of coral reef R and its biological basis.
These studies represent the greater part of the current state
of knowledge of R for coral reef bottom-types. In their
review, Hedley and Mumby (2002) conclude that ‘‘there is
a great deal of uncertainty and inconsistency in the reported
spectral features that can be employed to discriminate reef
benthos.’’ This is attributable to the fact that the total number
of measurements has been few, methods have been dissimilar
(producing data of varying quality), and objectives have
focused on very localized measurements for a few species.
That is, the database of R for coral reefs is small, and the data
within are nonuniform. As a result, deterministic, reef-up
remote sensing of bottom-types remains impractical, which
in turn precludes quantitative remote sensing of reef com-
munity structure. If remote sensing technology is to be
applied to reef-up assessment of reef community structure,
the first step must be to evaluate the degree to which
fundamental bottom-types can be spectrally discriminated.
Our goals are (1) to characterize spectral reflectance for
reef bottom-types worldwide and (2) to determine spectral
separabilities of the bottom-types. We have used a portable
spectrometer to measure 13,100 in situ R’s at 11 sites in four
major reef biogeographic regions worldwide at water depths
ranging from 0 to 15 m. We assign each measured R to one
of a set of fundamental reef bottom-types, which are defined
primarily by function in the reef system, but also by
taxonomy and by apparent color. We find that each bot-
tom-type exhibits characteristic and distinctive features in R
that exist in well-defined, narrow wavelength ranges and
that are conservative across biogeographic regions. Classi-
fication analysis demonstrates that the bottom-types are
statistically separable and identifiable based on their reflec-
tance spectra. We reason that features in R arise primarily as
a result of spectral absorption processes. Radiative transfer
modeling shows that in typically clear coral reef waters,
dark substrates such as corals have a depth-of-detection
limit on the order of 10–20 m. Our results provide the basis
for quantitative remote sensing of community structure
using consistent criteria for coral reefs worldwide.
2. Methods
2.1. Classification of fundamental bottom-types
Coral reef communities are largely mosaics of coral, va-
rious algae and carbonate sand (Kinsey, 1985), and knowl-
edge of their distributions is fundamental to assessment of
reef status (Connell, 1997; Done, 1992, 1995). Thus, these
three bottom-types form the foundation for our classifica-
tion scheme. There are three basic forms of reef algae: turf
algae, crustose calcareous algae, and fleshy macroalgae
(Berner, 1990). Crustose calcareous algae are important reef
calcifiers (Kinsey, 1985), cementing the products of dis-
integration of various other calcifying reef organisms, thus
creating a harder skin for the reef (Berner, 1990). Turf algae
and fleshy macroalgae are a major source of fixed carbon to
reef primary consumers (Klumpp & McKinnon, 1989).
Because of their importances to different reef processes,
these three algal types form subclasses within the broader
algae class.
While crustose calcareous algae are mainly Rhodophytes
(Corallinaceae), algal turfs are often a mixture of Chlor-
ophytes, Cyanophytes, Phaeophytes, and Rhodophytes
(Berner, 1990) that can mix on spatial scales of < 1 cm,
well beyond the spatial abilities of today’s remote sensors
(smallest footprint f 2 m). However, discounting their
epiphytic communities, fleshy macroalgae—mainly Chlor-
ophytes, Phaeophytes, and Rhodophytes—exist at a spatial
scale (one-tenths to tens of meters) that may be resolvable
by remote sensing. The different fleshy macroalgae taxa are
preferred forage material for different reef consumers
(Glynn, 1990). Furthermore, Chlorophytes, Phaeophytes,
and Rhodophytes possess different photosynthetic accessory
pigments (Kirk, 1994) and thus often exhibit their character-
istic colors of green, brown, and red, respectively. Because
they may be important to understanding energy flow
through the reef system, and because there is potential that
they can be discriminated based on their optical reflectance
spectra, we further separate the fleshy macroalgae subclass
into brown, green, and red divisions.
Soft corals and gorgonians can occupy substantial reef
areas and may compete with scleractinian corals for space
(Bastidas, Benzie, Uthicke, & Fabricius, 2001; Ben-Yosef &
Benayahu, 1999; Fabricius, 1997). Seagrass is essential
habitat in the life histories of many reef species and can
cover extensive back-reef and lagoonal areas (Enrıquez,
Merino, & Iglesias-Prieto, 2002). The spread and deposition
of terrigenous sediments can be deleterious to reefs near
high islands (Watanabe, Nakamura, Samarakoon, Mabuchi,
& Ishibashi, 1993). Because of their respective importances
to coral reef systems, each of these reef bottom-types is
included in the classification scheme.
Examination of the data set has revealed two basic modes
of coral R: one mode is associated with corals that are
visually (to humans) brown, red, orange, yellow, or green,
while the other mode is associated with corals that appear
purple, blue, pink, or gray. These patterns of association
occur across taxonomic lines and in all oceans. Thus, we
divide scleractinian corals into two subclasses, which, for
lack of better terminology, we label ‘‘brown’’ and ‘‘blue.’’
Lastly, the bleached coral subclass is included due to the
prevalence in recent years of reports of coral bleaching
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 161

worldwide (Wilkinson, 2000). Fig. 1 summarizes the overall
classification scheme adopted for this study.
2.2. Spectral measurements and processing
We measured 13,100 R’s at the following sites in the
Atlantic, Indian, and Pacific Oceans (Fig. 2): (1) St. Croix,
USVI; (2) Puerto Rico; (3) Florida Keys; (4) Oahu, Hawaii;
(5) Maui, Hawaii; (6) Rangiroa, French Polynesia; (7)
Moorea, French Polynesia; (8) Palau; (9) Bali, Indonesia;
(10) Mayotte, Comoros; (11) the Waikiki Aquarium (Indo-
Pacific corals grown in aquaria). The 11 sites represent four
major reef biogeographic regions as defined by Veron
(1995): Caribbean, Hawaiian Islands, Central Pacific, and
Indo-west Pacific.
The spectral reflectance R (implicitly a function of wave-
length) of a material is defined as the ratio of the reflected
radiant flux to the incident radiant flux (Morel & Smith,
1993). In our case, R is the fraction of incident light flux that
is reflected by the different bottom-types. We measured and
processed in situ R for visible wavelengths (400–700 nm)
following methods described in Hochberg and Atkinson
(2000). The sampling unit consisted of a 30-m-long fiber
optic cable (400 Am diameter) attached to an Ocean Optics
S2000 portable spectrometer (wavelength range 330–850
nm, with f 0.3-nm sample interval and f 1.3-nm optical
resolution), which in turn was operated by a laptop computer.
The fiber optic cable tip collected light over a solid angle off 0.1 sr, which at a distance of 10 cm projected to a circular
area of 10 cm2. For each single measurement of R, a diver
pointed the collecting tip of the fiber optic cable at the
desired bottom-type and depressed a button at the end of a
30-m-long trigger cable, prompting the computer to store the
spectrum (in units of digital counts). Immediately thereafter,
the diver pointed the collecting tip at a Spectralon diffuse
reflectance target (same depth as the target bottom-type) and
triggered the storage of its spectrum. In this manner, both
spectra could be acquired within 1–2 s. To maximize the
signal-to-noise ratio, a 10% reflectance target was used for
dark substrates (e.g., corals, algae), and a 99% reflectance
target was used for bright substrates (i.e., carbonate sand). To
ensure a constant ambient light field between the two
measurements, the Spectralon was placed immediately adja-
cent to the target bottom-type, and the diver’s position was
held constant for the 1–2 s required for the measurements.
Measurement depths ranged between 0 and 15 m. For
shallow ( < 5 m) samples, we shaded both target bottom-type
and Spectralon to minimize the influence of wave focusing
(light ‘‘flashes’’). We employed a submersible flashlight
(Underwater Kinetics Sunlight C8) to supplement flux at
red wavelengths for deeper (>5 m) samples.
We corrected all spectra for baseline electrical signal, then
calculated R as the ratio of digital counts measured over the
bottom-type to the digital counts measured over the Spec-
Fig. 1. Classification scheme for reef bottom-types. Coral, algae and sand form the set of reef bottom-types that are of primary interest for assessment of reef
status and are shown in bold. The remaining bottom-types provide insight into various reef processes (see text).
Fig. 2. Worldwide distribution of R sample sites. We measured a total of 13,100 in situ R at 11 sites in four major reef biogeographic regions. Sites included St.
Croix, Puerto Rico, Florida Keys, Oahu, Maui, Rangiroa, Moorea, Palau, Bali, Mayotte and the Waikiki Aquarium (Indo-Pacific corals grown in aquaria).
Represented biogeographic regions were the Caribbean, Hawaiian Islands, Central Pacific and Indo-west Pacific. Gray dots indicate sample sites, and shades
indicate biogeographic regions (after Veron, 1995).
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173162

tralon (corrected to 100% reflectance) for each pair of
measurements. Note that this is the ratio of two radiance
measurements: our calculation of R assumes that both the
bottom-type and Spectralon are diffuse reflectors (see Hoch-
berg & Atkinson, 2000). We linearly interpolated R to 1-nm
intervals over the wavelength range 400–700 nm, then
filtered the result using the Savitsky–Golay method (Savit-
sky & Golay, 1964; Steiner, Termonia, & Deltour, 1972).
2.3. Spectral characterization
We examined spectral shapes by numerically calculating
second-derivative spectra following the Savitsky–Golay
method (Savitsky & Golay, 1964; Steiner et al., 1972) and
identifying the wavelength locations of local maxima
(peaks). Note that derivative analysis merely exaggerates
spectral shapes, highlighting features present in zero-order
Fig. 3. In situ optical reflectance spectra R of coral reef benthic communities. Shaded areas represent following data bounds: light gray = 2.5–97.5%, medium-
gray = 12.5–87.5%, dark gray = 25–75% and black = 37.5–62.5%. Thus, for example, 95% of all spectra lie within the light gray region, and 50% lie within
the dark gray region. White lines shown mean spectra for each bottom-type. R was measured in situ using a diver-operated portable spectrometer. Note scale
difference for carbonate sand.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 163
spectra; derivative analysis does not add information not
already contained in zero-order spectra (Talsky, 1994). For
each bottom-type, we compared spectral shapes between
biogeographic regions by computing the frequencies of
occurrence of the second-derivative peaks. It became appa-
rent that each bottom-type exhibited its own remarkably
consistent features in R across all biogeographic regions.
Therefore, to describe global R for each bottom-type, we
combined spectra from all regions and computed the 2.5, 25,
75, and 97.5 percentile spectra, as well as the overall mean
spectrum.We further computed the frequencies of occurrence
of second-derivative peaks for global R in each bottom-type.
2.4. Spectral separability
To determine the spectral separability of the bottom-
types, we performed a classification analysis following the
partition method (Rencher, 1995): we used half of the R’s in
each class to train linear classification functions (LCFs), and
the other half to test classification accuracy (LCFs are linear
combinations of variables with a different set of variable
coefficients for each bottom-type. The variable coefficients
are calculated considering both the magnitude and shape of
the training spectra. An unknown spectrum is predicted to
belong to the bottom-type for which it has the highest LCF
value). Training and test spectra were chosen using a
pseudo-random number generator. We calculated classifica-
tion rates as the number of individual R’s in the predicted
class divided by the total number of R’s in the actual class,
multiplied by 100. The values in such a classification table
are equivalent to a table of Producer’s Accuracies (Con-
galton, 1991).
Finally, we investigated whether full-resolution spectra
(i.e., 400–700 nm at 1-nm intervals) are necessary for
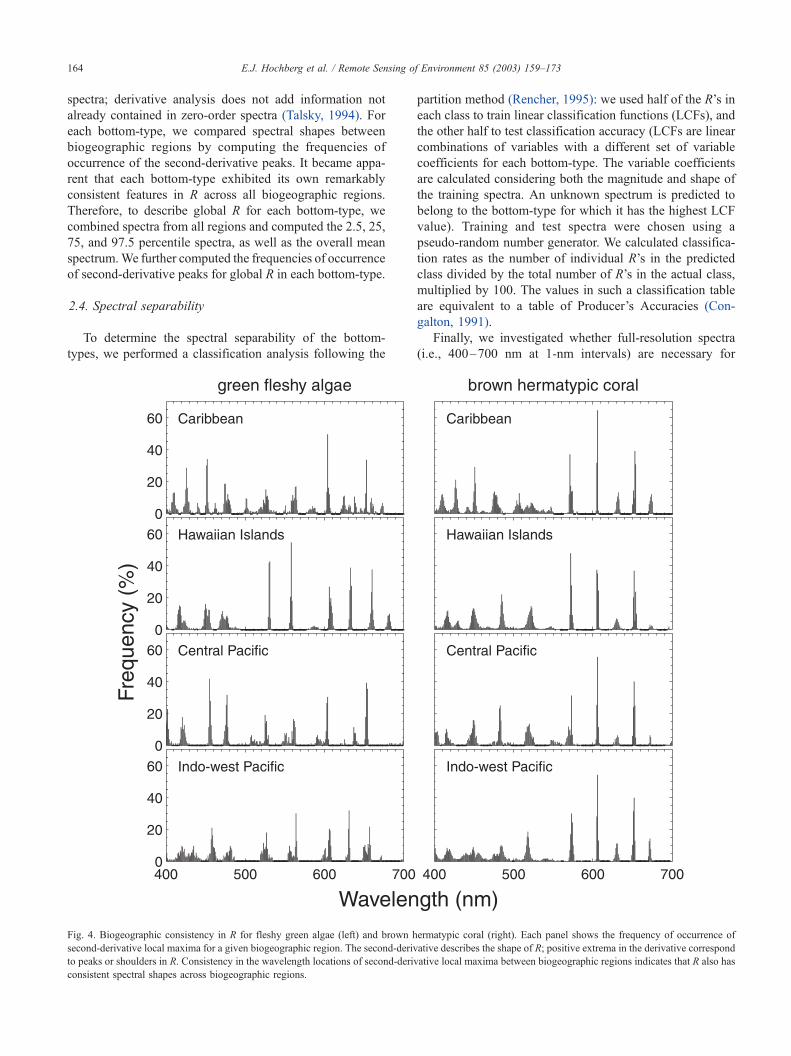
Fig. 4. Biogeographic consistency in R for fleshy green algae (left) and brown hermatypic coral (right). Each panel shows the frequency of occurrence of
second-derivative local maxima for a given biogeographic region. The second-derivative describes the shape of R; positive extrema in the derivative correspond
to peaks or shoulders in R. Consistency in the wavelength locations of second-derivative local maxima between biogeographic regions indicates that R also has
consistent spectral shapes across biogeographic regions.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173164

spectral separation of the bottom-types. We used a multi-
variate stepwise selection procedure (Rencher, 1995) to find
the subset of wavelengths that best separates the 12 classes
(stepwise selection discards those wavelengths that are
statistically redundant for separating the bottom-types).
The selection produced a list of 52 wavelengths, which
appeared to aggregate in clusters of wavelengths. Conse-
quently, we further simplified the wavelength set by finding
the mean wavelength in each cluster, resulting in a 14-
wavelength list. We repeated the classification analysis
using the same training and test R’s, but undersampled to
the 52-wavelength and 14-wavelength sets.
3. Results
There are features in R that are common to all bottom-
types (Fig. 3). Low values at blue and green wavelengths are
largely the result of absorption by photosynthetic and photo-
protective compounds (Bidigare, Ondrusek, Morrow, &
Kiefer, 1990; Dove, Takabayashi, & Hoegh-Guldberg,
1995; Salih, Larkum, Cox, Kuhl, & Hoegh-Guldberg,
2000). Similarly, higher values at red wavelengths indicate
lack of absorption or presence of active fluorescence (Mazel,
1995). Chlorophyll absorption is readily apparent near 675
nm, and the effect of strong near-infrared reflectance is
apparent at 700 nm. Except for carbonate sand, all bottom-
types have low average R, generally falling in the range 0–
30%, and all have either peaks or shoulders near 600 and 650
nm. Finally, all bottom-types exhibit wide variations in the
magnitude of R, while maintaining the same relative shape.
In brown corals, R has the triple-peaked pattern described
by Hochberg and Atkinson (2000), which is marked by a
prominent positive reflectance feature near 570 nm (Fig. 3).
Healthy blue coral R is characterized by an absorption
feature (local minimum) near 580 nm and a plateau between
600 and 650 nm. The presence of fluorescent pigments
(Salih et al., 2000) in the coral tissue is sometimes apparent
as subtle positive features at blue and green wavelengths in
both healthy coral classes (though not apparent in Fig. 3).
The shape of bleached coral R resembles that of carbonate
sand more closely than that of either healthy coral class, but
the magnitude is intermediate between that of the healthy
corals and carbonate sand. Encrusting calcareous and turf
algae have spectral shapes similar to that of red fleshy algae.
These algae are characterized by two broad, positive fea-
tures in R over the ranges 435–490 and 500–565 nm.
Brown fleshy algae R also exhibits the first feature at 435–
490 nm, but lacks the second. Both green fleshy algae and
seagrass have a single broad reflectance feature centered at
550–560 nm. Soft/gorgonian coral R resembles that of
brown coral. Terrigenous mud is characterized by a very
low R, increasing nearly linearly from 1% at 400 nm to 8%
at 700 nm, and exhibiting a minimal chlorophyll absorption
feature near 675 nm. Finally, carbonate sand has a very high
R, with minimum values of 20% at 400 nm and reaching
maximum values of 80% at 700 nm.
In all bottom-types, locations of second-derivative peaks
are consistent across biogeographic regions. Fig. 4 is an
example showing fleshy green algae and brown hermatypic
coral. This consistency in peak location results from each
bottom-type having a suite of pigments that is conservative
throughout the world, and it is spectral absorption by these
pigments that ultimately determines the shape of R. Fig. 5
shows the frequencies of second-derivative peak wave-
lengths for all bottom-types across all biogeographic regions.
Fig. 5. Differences in spectral features between coral reef bottom-types. Each row corresponds to 1 of 12 bottom-types and shows the frequency of occurrence
of second-derivative local maxima across all biogeographic regions. Brighter grays (to white) indicate higher frequencies of occurrence (see color bar at
bottom). In each bottom-type, spectral features occur in tight wavelength bands, indicating worldwide consistency in R. All bottom-types exhibit features near
600 and 650 nm, while most differences between bottom-types occur at wavelengths V 570 nm. Notably, >90% of brown hermatypic corals and soft/gorgonian
corals have a feature near 570 nm, but all algal classes lack this feature.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 165

Notably, brown hermatypic coral and soft/gorgonian coral
each have a strong feature near 570 nm, exhibited by no other
class. Nearly all peaks occur in narrow wavelength ranges,
sometimes as broad as 20 nm, but often on the order of 10 nm.
In the classification analysis, mean accuracy (average of
correct classification rates) was 83% (Table 1A) (we report
mean accuracy, rather than overall accuracy, due to the wide
differences in samples sizes between classes). In all instan-
ces of significant confusion between two bottom-types
(arbitrarily defined as a misclassification rate z 10%), the
confusion was one-way. Soft/gorgonian corals were mis-
classified as brown coral, but the reverse was not true. Most
classification errors occurred within, not between, broader
bottom-types. That is, algae tend to be misclassified as other
algae, and corals tend to be misclassified as other corals.
The class with highest classification error is soft/gorgonian
coral, which tends to be misclassified as brown coral more
often than it is correctly classified. To a much lesser extent
(i.e., < 10%), two-way misclassification occurs between
seagrass and fleshy green algae.
Table 1
Classification rates (%) for 12 basic coral reef bottom-types using in situ R with different waveband sets: full-resolution (301-wavelength at 1-nm intervals), 52-
wavelength, 14-wavelength
Predicted class
Algae Coral Other Sediment
Fleshy Non-fleshy Bleached Blue Brown Soft coral Seagrass Mud Sand
Brown Green Red Coralline Turf
(A) Full-resolution (400–700 nm at 1-nm intervals)
Actual class Algae fleshy brown 92.4 0.0 1.3 2.5 2.4 0.4 0.0 0.0 0.0 0.1 0.7 0.0
green 4.7 81.3 2.2 0.4 4.7 0.0 0.0 0.2 0.0 6.3 0.4 0.0
red 7.4 0.0 85.7 4.4 1.3 0.0 0.0 0.9 0.0 0.2 0.0 0.0
non-fleshy coralline 14.6 0.2 1.7 70.6 9.8 0.0 0.0 0.0 0.0 0.0 3.1 0.0
turf 12.7 0.9 2.3 6.8 73.5 1.1 0.0 0.0 0.0 2.0 0.7 0.0
Coral bleached 3.8 0.0 0.0 0.0 0.0 96.2 0.0 0.0 0.0 0.0 0.0 0.0
blue 0.0 0.4 3.3 0.0 0.0 0.0 72.3 23.1 0.0 0.0 0.8 0.0
brown 0.9 0.0 2.0 0.1 0.3 0.0 0.4 93.4 2.2 0.0 0.5 0.0
Other soft coral 1.1 0.0 0.0 0.0 0.0 0.0 0.0 52.1 43.6 0.0 3.2 0.0
seagrass 0.8 5.3 0.0 0.0 4.6 0.0 0.0 0.0 0.0 88.5 0.8 0.0
Sediment mud 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 100.0 0.0
sand 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 99.7
(B) 52-wavelength
Actual class Algae fleshy brown 71.0 0.0 22.7 3.7 1.9 0.6 0.0 0.0 0.0 0.1 0.0 0.0
green 6.3 75.0 1.3 2.5 2.9 0.2 0.0 0.0 0.0 9.9 2.0 0.0
red 6.7 0.0 85.2 5.4 1.1 0.0 0.0 1.1 0.0 0.0 0.6 0.0
non-fleshy coralline 3.1 0.0 19.3 62.8 12.6 0.0 0.0 0.0 0.0 0.5 1.7 0.0
turf 10.2 0.5 5.6 9.5 69.0 2.5 0.0 0.0 0.0 2.2 0.5 0.0
Coral bleached 0.0 0.0 0.0 0.0 19.2 80.8 0.0 0.0 0.0 0.0 0.0 0.0
blue 0.0 0.0 5.8 0.0 0.0 0.0 69.0 25.2 0.0 0.0 0.0 0.0
brown 0.2 0.0 4.7 0.0 0.3 0.2 0.6 90.7 1.9 0.0 1.2 0.0
Other soft coral 0.0 0.0 0.0 0.0 0.0 0.0 0.0 59.6 38.3 0.0 2.1 0.0
seagrass 0.8 17.6 0.0 0.0 6.9 0.0 0.0 0.0 0.0 74.0 0.8 0.0
Sediment mud 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 99.8 0.0
sand 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 99.7
(C) 14-wavelength
Actual class Algae fleshy brown 53.6 0.0 34.1 5.5 2.5 0.7 0.1 1.8 0.1 1.0 0.4 0.0
green 5.4 72.9 8.8 0.5 1.4 0.0 0.0 0.9 0.2 8.8 1.1 0.0
red 1.1 0.0 85.0 7.8 0.2 1.5 0.0 3.1 0.0 0.2 1.1 0.0
non-fleshy coralline 30.5 0.0 8.1 49.4 10.7 0.2 0.0 0.0 0.0 0.5 0.5 0.0
turf 10.4 0.0 2.2 15.1 59.9 5.7 0.0 1.1 0.0 2.7 3.0 0.0
Coral bleached 3.8 0.0 0.0 0.0 7.7 80.8 0.0 3.8 3.8 0.0 0.0 0.0
blue 0.0 0.0 3.7 0.0 0.0 0.0 69.8 26.0 0.0 0.0 0.4 0.0
brown 0.6 0.0 8.4 0.2 0.0 0.8 1.0 80.8 7.0 0.0 1.0 0.0
Other soft coral 0.0 0.0 0.0 0.0 0.0 0.0 0.0 60.6 39.4 0.0 0.0 0.0
seagrass 1.5 34.4 0.0 0.8 5.3 0.0 0.0 0.0 0.0 58.0 0.0 0.0
Sediment mud 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 99.6 0.0
sand 0.0 0.0 0.0 0.0 0.0 0.6 0.0 0.0 0.0 0.0 0.0 99.4
Classification rates are the number of individual R’s in the predicted class divided by the total number of R’s in the actual class, multiplied by 100. Diagonal
elements (in bold) represent correct classification rates, and off-diagonal elements represent misclassification rates. Misclassification rates z 10% are italicized.
Predicted classification rates (i.e., rows) sum to 100% (or near to 100% due to roundoff error).
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173166

The same patterns are present in both the 52-wavelength
and 14-wavelength classification rates (Table 1B and C).
Generally, with fewer wavelengths, misclassification rates
increase. Mean classification accuracy is 76% and 71% for
the 52-wavelength and 14-wavelength cases, respectively.
Table 2 lists the 52- and 14-wavelength sets used in the
subset analyses.
4. Discussion
R as reported here is a combination of the light flux not
absorbed by the bottom-types and the light fluoresced by the
bottom-types. Coral and zooxanthellae (unicellular algae
living endosymbiotically within coral-host tissue) pigments
have been shown to fluoresce (Dove, Hoegh-Guldberg, &
Ranganathan, 2001, Dove et al., 1995; Mazel, 1995, 1996;
Myers et al., 1999; Salih et al., 2000), and our data have
indications of that fluorescence. However, it is largely
spectral absorption by pigments that determines the overall
spectral shape of R. We demonstrate this by performing a
simple modeling exercise: we create a relative absorption
spectrum for brown coral (Fig. 6A) by summing the absorp-
tion spectra of five zooxanthellae pigments (chlorophyll a,
chlorophyll c, h-carotene, diadinoxanthin, peridinin), which
Fig. 6. Model of relative reflectance for brown coral using absorption spectra weighted by relative concentrations reported in literature. See text for details of
calculations. (A) Relative spectral absorptions by the five pigments in the model, and the total relative absorption. chl a= chlorophyll a, chl c= chlorophyll c, h-car =h-carotene, diadin = diadinoxanthin, per = peridinin. (B) Normalized reflectance for modeled and measured coral spectra. (C) Second derivative of (B).
Table 2
The 52- and 14-wavelength subsets used in classification analyses
52-wavelength subset 14-wavelength subset
400, 401, 405, 406, 407, 409, 411, 417,
420, 421, 427, 429, 431, 434, 438, 441,
445, 451, 455, 457, 466, 470, 481, 497,
499, 508, 509, 528, 540, 541, 559, 565,
571, 576, 579, 585, 599, 601, 609, 611,
629, 639, 644, 651, 666, 670, 678, 679,
685, 686, 689, 700
406, 430, 454, 467, 480,
499, 507, 529, 540, 577,
602, 608, 643, 684
The 52-wavelength subset was identified by multivariate stepwise selection
as those wavelengths that, without redundancy, provide greatest separation
of the 12 bottom-types. The 14-wavelength subset was identified inter-
actively as average wavelengths of clusters from 52-wavelength subset.
Wavelengths listed in nanometers.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 167
have been weighted by their relative concentrations in corals,
as reported in the literature (Andersen, Bidigare, Keller, &
Latasa, 1996; Cottone, 1995; Fang, Liao, & Liu, 1995;
Kleppel, Dodge, & Reese, 1989; Myers et al., 1999). We
convert to relative reflectance by taking the base-10 loga-
rithm of the inverse of relative absorption (Talsky, 1994),
then normalizing by the result’s vector length. For compar-
ison, we average 4308 brown coral R’s and normalize that
spectrum by its vector length. The normalized spectra (Fig.
6B) are relative (as opposed to absolute) reflectances and
represent spectral shape independent of magnitude. The
modeled and in situ spectra compare favorably, each show-
ing the same triple-peaked pattern characteristic of brown
corals. Comparison of the second derivatives of the relative
reflectances confirms that both spectra have the same basic
shape (Fig. 6C). Furthermore, the R we have measured for
brown coral (and soft/gorgonian corals) is the inverse pattern
of spectral absorption by peridinin-containing dinoflagel-
lates (Johnsen, Samset, Granskog, & Sakshaug, 1994), the
group to which zooxanthellae belong. Thus, we conclude
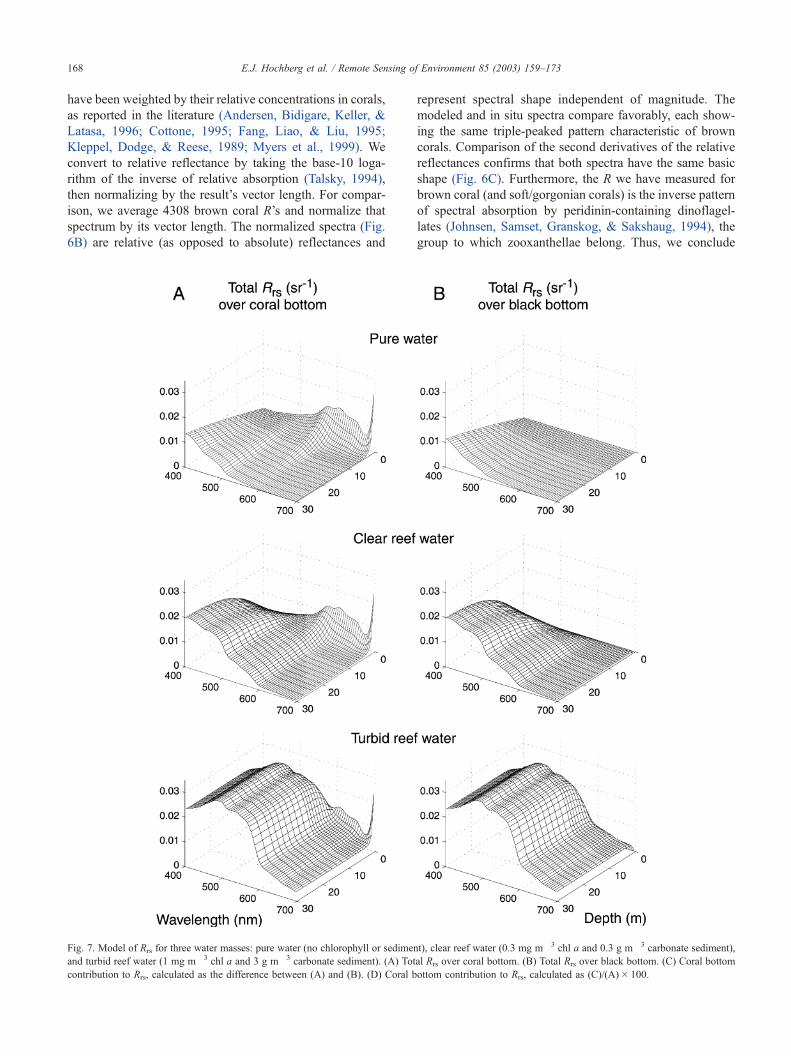
Fig. 7. Model of Rrs for three water masses: pure water (no chlorophyll or sediment), clear reef water (0.3 mg m� 3 chl a and 0.3 g m� 3 carbonate sediment),
and turbid reef water (1 mg m� 3 chl a and 3 g m� 3 carbonate sediment). (A) Total Rrs over coral bottom. (B) Total Rrs over black bottom. (C) Coral bottom
contribution to Rrs, calculated as the difference between (A) and (B). (D) Coral bottom contribution to Rrs, calculated as (C)/(A)� 100.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173168

that, while fluorescence does contribute to R in specific
cases, the shape of R for brown coral is primarily determined
by zooxanthellae pigment absorption.
The concept that absorption is the primary determinant of
R can be extended to other reef bottom-types. For example,
the absorption feature near 580 nm in blue corals indicates
the presence of pocilloporin in the coral host (Dove et al.,
1995). Thus, R in blue corals results from a combination of
absorptions by zooxanthellae and host pigments. Encrusting
calcareous algae, turf algae, and red fleshy algae exhibit
similar R’s because the dominant (or sole) components of
these bottom-types all belong to the taxonomic division
Rhodophyta and therefore share the same suite of absorbing
pigments. Bleached coral has R similar to that of carbonate
sand due to loss of zooxanthellae combined with a decrease
in zooxanthellar and host pigmentation, resulting in optical
exposure of the coral carbonate skeleton (Kleppel et al.,
1989). Finally, R for carbonate sand itself shows the com-
bined effects of absorption by calcium carbonate sand grains
and chlorophyll in benthic microalgae (Roelfsema, Phinn, &
Dennison, 2002).
The fact that absorption by pigments is responsible for
the shape of R in reef bottom-types is expected. The same
principle has been the basis for remote sensing of ocean
Fig. 7 (continued).
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 169

color for more than 20 years (Gordon & Clark, 1980; Morel
& Prieur, 1977). It has been demonstrated that there is a link
between induced fluorescence of reef benthic organisms and
their pigment compositions and concentrations (Hardy,
Hodge, Yungel, & Dodge, 1992; Myers et al., 1999). Based
on those studies and our results, it may now be appropriate
to develop the field of ‘‘coral reef color.’’
We have presented Fig. 4 as a qualitative example show-
ing that the shape of R is geographically invariant. Unfortu-
nately, we cannot statistically explore this issue, since,
despite our large sample size, we do not have sufficient
numbers of measurements for each bottom-type in each
biogeographic region for appropriate multivariate tests (Ren-
cher, 1995). However, the peaks of second-derivative spectra
are consistent between regions (e.g., Fig. 4), which indicates
that the shapes of zero-order R are also consistent (Talsky,
1994). Geographic invariance in R is attributable to the facts
that (1) absorption is a fundamental process that is independ-
ent of biogeography and (2) the pigments responsible for
absorption also exist independently of biogeography. Geo-
graphic invariance of R within bottom-types has a corollary:
differences between bottom-types are also geographically
invariant. This is the biooptical basis for the spectral sepa-
ration achieved here through classification analysis.
Previous studies have demonstrated the feasibility of
using high-resolution remote sensing to assess coral reef
status (e.g., percent of live and dead corals) in localized
settings using extensive ground-truthing and unsupervised
classifications (e.g., Mumby, Chisolm, Clark, Hedley, &
Jaubert, 2001), but generalizations of methods have not
been possible. For example, the results obtained by Mumby
et al. (2001) at Rangiroa (Tuamotu Archipelago, French
Polynesia) are not reproducible at nearby Moorea (Society
Islands, French Polynesia), even using data from the same
hyperspectral imaging system, due to the influence of fleshy
brown algae at Moorea (Andrefouet, unpublished data;
Mumby, personal communication). That R for a given
bottom-type has the same shape regardless of geography
ultimately provides the foundation for development of
global discrimination/classification algorithms.
It is important to note that R is a convolution of
magnitude and relative shape, and both components con-
tribute to spectral separation between the bottom-types. An
example is the comparison between carbonate sand and
bleached coral: both bottom-types exhibit the same relative
spectral shape, because the dominant optically absorbing
component in each is the mineral calcium carbonate. How-
ever, carbonate sand has much higher overall reflectance
because bleached coral (1) actually retains some pigmenta-
tion in the zooxanthellae and coral host and (2) maintains its
three-dimensional structure which creates shadows, while
(3) the benthic microalgal community resident on sand
generates a smaller pigment absorption signature than that
of bleached coral, and (4) sand has a more two-dimensional
structure. More subtly, several of the bottom-types share
many second-derivative features (e.g., fleshy red algae and
non-fleshy turf algae), but do exhibit slight differences in
both the wavelengths at which the features occur (Fig. 5)
and their absolute magnitudes (Fig. 3). The implication is
that discrimination of bottom-types based on these subtle
spectral differences requires relatively high spectral resolu-
tion, with maximum bandwidths on the order of 10–20 nm.
For each of the three different wavelength sets used in the
classification analysis, the greatest occurrence of error is
soft/gorgonian coral misclassified as brown hermatypic
coral. In a region where soft/gorgonian corals are common
(e.g., many sites in the Caribbean), a remote sensing study
that considers only spectral variables may produce signifi-
cant overestimates of hermatypic coral cover. For many of
the world’s reefs, however, soft/gorgonian corals are not a
dominant benthic component and often occupy different reef
zones than hermatypic corals (Fabricius & Alderslade,
2001). Thus, an applied remote sensing study should also
make use of knowledge-based algorithms, employing eco-
logical (i.e., spatial) as well as spectral variables (e.g.,
Andrefouet & Roux, 1998).
Our results represent an upper limit to spectral classifica-
tion accuracy, because in situ R is independent of absorption
and scattering within the atmosphere and water column.
These radiative transfer processes spectrally alter the reef-
surface signal before it is received by the remote sensor, and
continued research is necessary to develop methods for
inverting their effects. One important factor that has yet to
be explored in detail (except seeMaritorena et al., 1994) is the
depth at which the bottom is no longer detectable. Consider-
ing only the water column and seafloor, this depth-of-detec-
tion limit (zlim) is a function of R and of the water column’s
optical properties. Because both R and water column optical
properties are functions of wavelength, so is zlim. We have
explored the behavior of zlim with respect to water column
optical properties using the radiative transfer model Hydro-
light 4.1 (C.D. Mobley and L.K. Sundman, Sequoia Scien-
tific, Redmond, WA, USA). We use a four-component model
(pure water, chlorophyll-bearing particles, colored dissolved
organic matter covarying with chlorophyll, and suspended
calcareous sediment) to simulate remote sensing reflectance
(Rrs, water-leaving radiance divided by downwelling irradi-
ance) for three water masses: pure water (no chlorophyll or
sediment), clear reef water (0.3 mg m� 3 chl a and 0.3 g m� 3
carbonate sediment), and turbid reef water (1 mg m� 3 chl a
and 3 g m� 3 carbonate sediment). We execute the model for
water depths in the range 0–30 m with the bottom boundary
specified as a coral bottom (our average brown coral spec-
trum, Fig. 7A) and as a perfectly absorbing bottom (flat
spectrum of 0% reflectance across all wavelengths, Fig. 7B).
Rrs over a coral bottom is comprised of a bottom component
and a water column component, whileRrs over a black bottom
expresses only the water column component (Maritorena et
al., 1994). By subtracting the latter from the former, we
estimate the contribution of the coral bottom to total Rrs (Fig.
7C), and by dividing this value by totalRrs andmultiplying by
100, we obtain the coral bottom contribution to total Rrs as a
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173170

percent (Fig. 7D). This value represents the relative signal-to-
noise level for the coral bottom, where noise is the water
column contribution to Rrs.
Fig. 8 shows the coral 5% and 10% signal levels as
functions of wavelength for the three water masses. In theory,
these lines are exact; we choose not to draw the 0% signal line
because, in practice, a radiometer pointed downward at a reef
will have difficulty resolving lower signal levels (i.e., sensor
signal-to-noise issues). Thus, these lines approximate func-
tional zlim. It is no surprise that zlim is much shallower for
turbid water than for clear water. It also should be noted that
most reef areas of interest in studies of community structure
lie well within the clear reef water zlim and that reef waters,
especially outer reef slopes, are often more clear than our
clear reef water case. Finally, these results are generally valid
for all the bottom-types except carbonate sand, because all
have roughly the same magnitude R as brown coral.
The wavelength dependence of zlim in clear reef water
indicates that remote sensing of coral reefs is best achieved at
wavelengths shorter than about 580 nm. This of course results
from the rapid rise in attenuation at longer wavelengths due to
absorption by water. There is potential for this effect to
interfere with remote discrimination of reef bottom-types:
the characteristic 570 nm feature in R of brown hermatypic
coral is at the cusp of increasing attenuation, and the attenu-
ation feature tends to produce a strong shoulder in Rrs in this
wavelength region for all water clarities and all bottom
compositions. The strength of the effect and the depth at
which it becomes apparent vary with water clarity. As long as
the bottom is not past zlim, however, there is potential for
inverse radiative transfer modeling to remove the effect, thus
allowing accurate identification of bottom-type.
With regard to the specific classification technique, we
have achieved high classification accuracy rates with simple
LCFs. With further tuning of our classifiers, it is possible to
achieve even higher levels of accuracy. More sophisticated
classifiers such as quadratic classification functions or multi-
variate density estimators will likely increase classification
accuracy of the ‘‘pure’’ bottom-type spectra, and others may
afford the ability to deconvolve spectrally mixed signals.
LCFs with full 1-nm-resolution spectra (i.e., 301 contig-
uous wavelengths) demonstrate high spectral separability of
the classes. Existing satellite sensors, however, do not
approach this degree of spectral resolution, and this fact
limits their abilities to discriminate between the 12 bottom-
types that we have defined. A repeat of our classification
analysis using appropriate sensor wavebands resulted in
mean classification accuracies of 41%, 45%, and 47% for
the satellite sensors SPOT-HRV (Systeme Pour l’Observa-
tion de la Terre—High Resolution Visible), Ikonos and
Landsat-ETM+ (Enhanced Thematic Mapper Plus), respec-
tively. At 78%, only the airborne sensor AVIRIS (Airborne
Visible Infrared Imaging Spectrometer, visible wavebands
only) has a mean classification accuracy commensurate with
that of full-resolution spectra. Fortuitously, in the near future,
satellite sensors will begin to match the spectral capabilities
of AVIRIS.
Alternatively, advancing technology has made it feasible
to design a satellite sensor with the purpose of addressing
specific questions in global coral reef science. Such a sensor
would possess particular wavebands explicitly chosen for the
task at hand, and it is important to determine classification
accuracies for these more realistic waveband sets. For
classification of the three more general bottom-types—algae,
coral, and sand—LCFs using four 20-nm-wide wavebands
achieved an overall accuracy of 91% (Hochberg & Atkinson,
2003). Coupling this waveband set with radiative transfer
models indicates that this spectral separation is achievable to
Fig. 8. Functional depth-of-detection limits (zlim) for the three water masses. Below zlim, inverse radiative transfer calculations are unable to reconstruct bottom
reflectance signatures. At a given wavelength, zlim is a function of water clarity and magnitude of bottom reflectance. These cases of zlim are computed for the
brown hermatypic coral bottom in Fig. 7D, but these results are extendable to other bottom-types, because most have R at approximately the same order of
magnitude as coral.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 171

water depths between 5 and 10 m under a clean tropical
atmosphere and case 1 ocean water (Atkinson et al., 2001).
This demonstrates the utility of employing characteristic R as
the basis for space-borne coral reef remote sensing. Such
simplified waveband combinations ease satellite engineering
constraints (i.e., a sensor with fewer wavebands is easier to
design and operate), facilitating lower cost remote sensing
solutions to the problem of global coral reef study and
monitoring.
We have found measurement of R for any bottom-type to
be extremely repeatable. The R’s we have measured are
consistent in shape and magnitude with those reported by
other researchers using a similar methodology: near-simul-
taneous measurement of incident and reflected light fluxes
(Andrefouet et al., 2001; Clark et al., 2000; Hochberg &
Atkinson, 2000; Holden & LeDrew, 1998, 1999; Joyce &
Phinn, 2002; Lubin, Li, Sustan, Mazel, & Stamnes, 2001;
Maritorena et al., 1994; Miyazaki & Harashima, 1993;
Myers et al., 1999). We have shown that basic reef
bottom-types have characteristic R, that within each bot-
tom-type, the shape of R is consistent across biogeographic
regions, and that the bottom-types are spectrally separable
from each other. Our results provide the basis for other
research efforts aimed at detailed ecological interpretation of
global coral reef remote sensing imagery. Given that coral
reefs around the world are believed to be in peril, and that
current in situ sampling methods are incapable of providing
the synoptic information required for assessment of the
world’s coral reef resources, the need for a reef-monitoring
satellite is now greater than ever.
Acknowledgements
We are grateful for field assistance from Shannon
Atkinson, Mark Carmichael, Steve Dollar, Jim Falter, Jim
Fleming, Michael Guidry, Mika Hochberg, Brian Lapointe,
Jennifer Liebeler, Claude Payri, Russell Perkins, Brendalee
Phillips, Michel Pichon, Ann Tarrant, Bernard Thomassin,
and Robert Tomasetti. Logistical support for field operations
was provided by the US NOAA/NOS Biogeography
Program, the US National Park Service, and the US
Geological Survey. Our most effusive gratitude goes to
S.V. Smith for his comments and suggestions. This work
was funded by NASA award numbers NAG5-7513, NAG5-
5276 and NAG5-10908, and NOAA award number NA07-
OA0571. This is SOEST contribution 6053, HIMB
contribution 1147, and IMaRS contribution 0041.
References
Andersen, R., Bidigare, R., Keller, M., & Latasa, M. (1996). A comparison
of HPLC pigment signatures and electron microscopic observations for
oligotrophic waters of the North Atlantic and Pacific Oceans. Deep-Sea
Research II, 43, 517–537.
Andrefouet, S., Muller-Karger, F. E., Hochberg, E. J., Hu, C., & Carder,
K. L. (2001). Change detection in shallow coral reef environments
using Landsat 7 ETM+ data. Remote Sensing of Environment, 79,
150–162.
Andrefouet, S., & Payri, C. (2001). Scaling-up carbon and carbonate me-
tabolism of coral reefs using in-situ data and remote sensing. Coral
Reefs, 19, 259–269.
Andrefouet, S., Robinson, J. A., Hu, C., Feldman, G. C., Salvat, B., Payri,
C., & Muller-Karger, F. E. (2003). Influence of the spatial resolution of
SeaWiFS, Landsat 7, SPOT and International Space Station data on
estimates of landscape parameters of Pacific Ocean atolls. Canadian
Journal of Remote Sensing, 29 (No. 2).
Andrefouet, S., & Roux, L. (1998). Characterisation of ecotones using
membership degrees computed with a fuzzy classifier. International
Journal of Remote Sensing, 19, 3205–3211.
Atkinson, M. J., Lucey, P. G., Taylor, G. J., Porter, J., Dollar, S., & An-
drefouet, S. (2001). CRESPO: Coral Reef Ecosystem Spectro-Photo-
metric Observatory, concept study report to the University Earth
System Science Program, National Aeronautics and Space Administra-
tion. Honolulu: University of Hawaii, 137 pp.
Bastidas, C., Benzie, J. A. H., Uthicke, S., & Fabricius, K. E. (2001).
Genetic differentiation among populations of a broadcast spawning soft
coral, Sinularia flexibilis, on the Great Barrier Reef. Marine Biology,
138, 517–525.
Ben-Yosef, D. Z., & Benayahu, Y. (1999). The gorgonian coral Acabaria
biseralis: life history of a successful colonizer of artificial substrata.
Marine Biology, 135, 473–481.
Berner, T. (1990). Coral-reef algae. In Z. Dubinsky (Ed.), Ecosystems of the
World 25: Coral Reefs ( pp. 253–264). Amsterdam: Elsevier.
Bidigare, R. R., Ondrusek, M. E., Morrow, J. H., & Kiefer, D. A. (1990). In
vivo absorption properties of algal pigments. Proceedings of Ocean
Optics X, SPIE vol. 1302 ( pp. 290–302). Orlando, FL: SPIE.
Buddemeier, R., & Smith, S. V. (1999). Coral adaptation and acclimatiza-
tion: a most ingenious paradox. American Zoologist, 39, 1–9.
Chabanet, P., Ralambondrainy, H., Amanieu, M., Faure, G., & Galzin, R.
(1997). Relationships between coral reef substrata and fish. Coral Reefs,
16, 93–102.
Clark, C. D., Mumby, P. J., Chisholm, J. R. M., Jaubert, J., & Andrefouet,
S. (2000). Spectral discrimination of coral mortality states following a
severe bleaching event. International Journal of Remote Sensing, 21,
2321–2327.
Congalton, R. G. (1991). A review of assessing the accuracy of classifi-
cations of remotely sensed data. Remote Sensing of Environment, 37,
35–46.
Connell, J. H. (1997). Disturbance and recovery of coral assemblages.
Coral Reefs, 16, S101–S113.
Cottone, M. C. (1995). Coral pigments: quantification using HPLC and
detection by remote sensing. Master’s Thesis, Western Washington Uni-
versity.
Done, T. J. (1992). Phase shifts in coral reef communities and their eco-
logical significance. Hydrobiologia, 247, 121–132.
Done, T. J. (1995). Ecological criteria for evaluating coral reefs and their
implications for managers and researchers. Coral Reefs, 14, 183–192.
Dove, S. G., Hoegh-Guldberg, O., & Ranganathan, S. (2001). Major colour
patterns of reef-building corals are due to a family of GFP-like proteins.
Coral Reefs, 19, 197–204.
Dove, S. G., Takabayashi, M., & Hoegh-Guldberg, O. (1995). Isolation and
partial characterization of the pink and blue pigments of Pocilloporid
and Acroporid corals. Biological Bulletin, 189, 288–297.
Enrıquez, S., Merino, M., & Iglesias-Prieto, R. (2002). Variations in the
photosynthetic performance along the leaves of the tropical seagrass
Thalassia testudinum. Marine Biology, 140, 891–900.
Fabricius, K. E. (1997). Soft coral abundance on the central Great Barrier
Reef: effects of Acanthaster planci, space availability, and aspects of the
physical environment. Coral Reefs, 16, 159–167.
Fabricius, K. E., & Alderslade, P. (2001). Soft corals and sea fans: a com-
prehensive guide to the tropical shallow-water genera of the Central-
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173172

West Pacific, the Indian Ocean and the Red Sea. Townsville, Queens-
land, Australia: Australian Institute of Marine Science, 264 pp.
Fang, L.-S., Liao, C.-W., & Liu, M.-C. (1995). Pigment composition in
different-colored scleractinian corals before and during the bleaching
process. Zoological Studies, 34, 10–17.
Ginsburg, R. N. (Ed.) (1994). Proceedings of the Colloquium on Global
Aspects of Coral Reefs: Health, Hazards and History. Miami: Rosenstiel
School of Marine and Atmospheric Sciences, University of Miami,
420 pp.
Glynn, P. W. (1990). Feeding ecology of coral-reef macroconsumers. In: Z.
Dubinsky (Ed.), Ecosystems of the world 25: coral reefs (pp. 365–400).
Amsterdam: Elsevier.
Gordon, H. R., & Clark, D. K. (1980). Atmospheric effects in the remote
sensing of phytoplankton pigments. Boundary-layer Meteorology, 18,
299–313.
Green, E. P., Mumby, P. J., Edwards, A. J., & Clark, C. D. (1996). A review
of remote sensing for the assessment and management of tropical coast-
al resources. Coastal Management, 24, 1–40.
Hardy, J. T., Hodge, F. E., Yungel, J. K., & Dodge, R. E. (1992). Remote
detection of coral ’bleaching’ using pulsed-laser fluorescence spectro-
scopy. Marine Ecology Progress Series, 88, 247–255.
Hedley, J. D., & Mumby, P. J. (2002). Biological and remote sensing
perspectives of pigmentation in coral reef organisms. In A. J. Young,
C. M. Young, & L. A. Fuiman (Eds.), Advances in Marine Biology, vol.
43 (pp. 279–317). San Diego: Academic Press.
Hochberg, E. J., & Atkinson, M. J. (2000). Spectral discrimination of coral
reef benthic communities. Coral Reefs, 19, 164–171.
Hochberg, E. J. & Atkinson, M. J. (2003). Capabilities of remote sensors to
classify coral, algae and sand as pure and mixed spectra. Remote Sens-
ing of Environment 85, 174–189 (this issue).
Holden, H., & LeDrew, E. (1998). Spectral discrimination of healthy and
non-healthy corals based on cluster analysis, principal components anal-
ysis, and derivative spectroscopy. Remote Sensing of Environment, 65,
217–224.
Holden, H., & LeDrew, E. (1999). Hyperspectral identification of coral reef
features. International Journal of Remote Sensing, 20, 2545–2563.
Johnsen, G., Samset, O., Granskog, L., & Sakshaug, E. (1994). In vivo
absorption characteristics in 10 classes of bloom-forming phytoplank-
ton: taxonomic characteristics and responses to photoadaptation by
means of discriminant and HPLC analysis. Marine Ecology Progress
Series, 105, 149–157.
Joyce, K. E., & Phinn, S. R. (2002). Bi-directional reflectance of corals.
International Journal of Remote Sensing, 23, 389–394.
Kinsey, D. W. (1985). Metabolism, calcification and carbon production I:
systems level studies. Fifth International Coral Reef Congress. Tahiti,
vol. 4 (pp. 505–526). Moorea, French, Polynesia: Antenne Museum-
EPHE.
Kirk, J. T. O. (1994). Light and photosynthesis in aquatic environments.
Cambridge: Cambridge Univ. Press, 509 pp.
Kleppel, G., Dodge, R., & Reese, C. (1989). Changes in pigmentation
associated with the bleaching of stony corals. Limnology and Ocean-
ography, 34, 1331–1335.
Klumpp, D. W., & McKinnon, A. D. (1989). Temporal and spatial patterns
in primary production of a coral-reef epilithic algal community. Journal
of Experimental Marine Biology and Ecology, 131, 1–22.
Light, P. R., & Jones, G. P. (1997). Habitat preference in newly settled coral
trout (Plectropomus leopardus, Serranidae). Coral Reefs, 16, 117–126.
Lubin, D., Li, W., Dustan, P., Mazel, C. H., & Stamnes, K. (2001). Spectral
signatures of coral reefs: features from space. Remote Sensing of Envi-
ronment, 75, 127–137.
Maritorena, S., Morel, A., & Gentili, B. (1994). Diffuse reflectance of
oceanic shallow waters: influence of water depth and bottom albedo.
Limnology and Oceanography, 39, 1689–1703.
Mazel, C. H. (1995). Spectral measurements of fluorescence emission in
Caribbean cnidarians. Marine Ecology Progress Series, 120, 185–191.
Mazel, C. H. (1996). Coral fluorescence characteristics: excitation-emission
spectra, fluorescence efficiencies, and contribution to apparent reflec-
tance. Proceedings of Ocean Optics XIII-SPIE vol. 2963, 1 (pp. 240–
245). Halifax, Nova Scotia, Canada: SPIE.
Miller, I., & Muller, R. (1999). Validity and reproducibility of benthic cover
estimates made during broadscale surveys of coral reefs by manta tow.
Coral Reefs, 18, 353–356.
Miller, M. W., Weil, E., & Szmant, A. M. (2000). Coral recruitment and
juvenile mortality as structuring factors for reef benthic communities in
Biscayne National Park, USA. Coral Reefs, 19, 115–123.
Minghelli-Roman, A., Chisholm, J. R. M., Marchioretti, M., & Jaubert, J.
M. (2002). Discrimination of coral reflectance spectra in the Red Sea.
Coral Reefs, 21, 307–314.
Miyazaki, T., & Harashima, A. (1993). Measuring the spectral signatures of
coral reefs. Thirteenth Annual International Geoscience and Remote
Sensing Symposium, vol. 2 ( pp. 693–695). Tokyo, Japan: IEEE.
Miyazaki, T., Nakatani, Y., & Harashima, A. (1995). Measuring the coral
reef distribution of Kuroshima Island by satellite remote sensing.
Proceedings of Global Process Monitoring and Remote Sensing of
the Ocean and Sea Ice-SPIE vol. 2586, 1 ( pp. 65–72). Paris, France:
SPIE.
Morel, A., & Prieur, L. (1977). Analysis of variations in ocean color.
Limnology and Oceanography, 22, 709–722.
Morel, A., & Smith, R. C. (1993). Terminology and units in optical ocean-
ography. US JGOFS Planning Report Number 18: Bio-Optics in US
JGOFS: 149–157.
Mumby, P. J., Chisolm, J. R. M., Clark, C. D., Hedley, J. D., & Jaubert, J.
(2001). A bird’s eye view of the health of coral reefs. Nature, 413, 36.
Mumby, P. J., & Edwards, A. J. (2002). Mapping marine environments
with IKONOS imagery: enhanced spatial resolution can deliver greater
thematic accuracy. Remote Sensing of Environment, 82, 248–257.
Mumby, P. J., Green, E. P., Clark, C. D., & Edwards, A. J. (1998). Digital
analysis of multispectral airborne imagery of coral reefs. Coral Reefs,
17, 59–69.
Mumby, P. J., Green, E. P., Edwards, A. J., & Clark, C. D. (1999). The
cost-effectiveness of remote sensing for tropical coastal resources as-
sessment and management. Journal of Environmental Management, 55,
157–166.
Myers, M. R., Hardy, J. T., Mazel, C. H., & Dustan, P. (1999). Optical
spectra and pigmentation of Caribbean reef corals and macroalgae.
Coral Reefs, 18, 179–186.
Rencher, A. C. (1995).Methods of multivariate analysis. New York: Wiley,
627 pp.
Roelfsema, C. M., Phinn, S. R., & Dennison, W. C. (2002). Spatial dis-
tribution of benthic microalgae on coral reefs determined by remote
sensing. Coral Reefs, 21, 264–274.
Salih, A., Larkum, A., Cox, G., Kuhl, M., & Hoegh-Guldberg, O.
(2000). Fluorescent pigments in corals are photoprotective. Nature,
408, 850–853.
Savitsky, A., & Golay, M. J. E. (1964). Smoothing and differentiation of
data by simplified least squares procedures. Analytical Chemistry, 36,
1627–1639.
Steiner, J., Termonia, Y., & Deltour, J. (1972). Comments on smoothing
and differentiation of data by simplified least square procedure. Ana-
lytical Chemistry, 44, 1906–1909.
Stoddart, D. R. (1969). Ecology and morphology of recent coral reefs. Bio-
logical Reviews of the Cambridge Philosophical Society, 44, 433–498.
Talsky, G. (1994). Derivative spectrophotometry: low and high order. New
York: Weinheim, 228 pp.
Veron, J. E. N. (1995). Corals in space and time: the biogeography and
evolution of the Scleractinia. Ithaca: Cornell Univ. Press, 321 pp.
Watanabe, F., Nakamura, K., Samarakoon, L., Mabuchi, Y., & Ishibashi, A.
(1993). A procedure for estimating and monitoring red soil spread on
coral reefs of Okinawa using multitemporal Landsat TM data. 1993
Proceedings of the Thirteenth International Geoscience and Remote
Sensing Symposium, vol 2 (pp. 696–699). Tokyo, Japan: IEEE.
Wilkinson, C. (Ed.) (2000). Status of coral reefs of the world: 2000.
Townsville, Queensland, Australia: Australian Institute of Marine Sci-
ence, 363 pp.
E.J. Hochberg et al. / Remote Sensing of Environment 85 (2003) 159–173 173








![Contents lists available at ScienceDirect Journal of ... 2014 JQSRT.pdfthe spectral reflectance and then calculate the emittance as one minus the reflectance [7,25].TheopticalpropertiesofAg](https://img.pdfslide.net/doc/110x75/5f5255ab5f1b4b113e42d4ec/contents-lists-available-at-sciencedirect-journal-of-2014-jqsrtpdf-the-spectral.jpg)




















