Embed Size (px)
Citation preview

ST HELENA BAY WATER
QUALITY TRUST:
St Helena Bay
State of the Bay 2012
Prepared by
ANCHORe n v i r o n m e n t a l


St Helena Bay
State of the Bay 2012
Prepared for:
St Helena Bay Water Quality Trust
Andre du Toit PO Box 655 Veldrif 7635
Tel: 022 7832860 Mobile: 083 2511451
Email: [email protected]
Prepared by:
8 Steenberg House, Silverwood Close,
Tokai 7945, South Africa Tel: 021 701 3420, Fax: 0865428711
www.anchorenvironmental.co.za
Authors: K.L. Tunley, B.M. Clark and A. Biccard
September 2012
ANCHORe n v i r o n m e n t a l


Page | i
Executive summary
Introduction
St Helena Bay is situated on the west coast of South Africa and extends from Dwarskersbos in the
north, past the Shelley Point peninsula, to Cape St Martin in the west, encompassing 18 smaller bays
and the estuary of the Berg River. The Bay is positioned in the southern section of the Benguela
Current System, one of four major eastern-boundary current systems which is characterised by the
wind-driven upwelling of cold, nutrient rich water. St Helena Bay is positioned downstream of the
Cape Columbine upwelling cell and is a retention zone for the nutrient rich water that is upwelled in
this cell. The bay is subject to incidence of harmful algal blooms and regular episodes of oxygen
depletion in the coastal waters, which have in the past lead to major mortality events for organisms
such as rock lobsters and fish. The Bay also serves as a major node for industrial and small-scale
fisheries on the west coast, as well as for other industries such as mariculture, ship repair and
shipbuilding.
Regular long-term monitoring in St Helena Bay was initiated by the St Helena Bay Water Quality
Trust in 2001. The monitoring programme focuses on water and sediment quality and biotic indices
of health, and was designed to provide an overview of trends in the health of the bay, and track to
changes that may be caused by human activities. Sediment quality and benthic macrofaunal
communities are monitored approximately every five years as part of this programme. The results
of these assessments are used to make recommendations regarding the sustainable management of
activities in the bay.
This report presents results on sediment characteristics, the levels of organic material in the
sediments, and the abundance and distribution of benthic macrofauna living in the sediments of the
bay in 2012. It is the third assessment conducted as part of the State of the Bay monitoring
program.
Results from the sediment monitoring programme
Sediment and benthic macrofauna samples were collected from 27 sites in St Helena Bay over a 3
day period during the month of April 2012. Samples were collected using a Van Veen grab operated
from a Rigid Inflatable Boat (RIB). Samples were analysed for grain size distribution (percentage
mud, sand and gravel) and organic content (Total Organic Carbon TOC and Total Organic Nitrogen
TON). Assessments of the spatial distribution of different particle size groups and organic content in
2012 were conducted. These were compared with results from 2001 and 2007.
Sediment samples collected in 2012 were comprised predominantly of sand, with a small proportion
of mud and no gravel. Relative to data from 2001 and 2007, the 2012 results indicate a dramatic
reduction in the amount of mud in the sediments and a significant reduction in the coarser gravel
particle fraction in the bay since 2007. This may be associated with an observed increasing intensity
and frequency of large storm events over the period 1996 to 2010.

Page | ii
Data from 2012 also suggest that the spatial distribution of mud in the Bay has also changed
dramatically over time, with areas exhibiting the highest mud content having shifted from the
northerly and southerly reaches of the Bay in 2007 to the south western edge near the Berg River
Estuary mouth in 2012. There have been minor reductions in the levels of organic nitrogen in the
sediments at most sites since 2007, while organic carbon levels decreased in the northern reaches,
increased in the vicinity of the fish factories and underwent minor changes elsewhere in the bay. The
sites in the immediate vicinity of the fish factory outfalls are clearly carbon enriched compared to
other areas in the bay. Effluent from fish factories is thus still making an important contribution to
organic loading in the sediments but this fortunately is localised to the immediate vicinity (within
100m) of the fish factory outfalls.
Carbon: Nitrogen ratios throughout the bay have increased. These changes may be associated with
an increase in denitrification. Denitrification is the breakdown of organic nitrates into nitrogen and
ammonia by denitrifying bacteria. These denitrifying bacteria dominate when oxygen availability is
low and nitrates are available. This suggests that low oxygen conditions may have prevailed within
the benthic environment of the bay since the previous 2007 survey. These conditions may be a result
of variations in phytoplankton productivity associated with fluctuations in upwelling-favourable
winds. These winds reached a peak in 2001 and underwent a decline up until 2006. Changes in the
Southern Oscillation Index (i.e. the oscillation between El Niño and La Niña conditions) may also have
played a role.
Sediment conditions in the vicinity of the fish factory outfalls have deteriorated markedly since
2007. These sites are clearly carbon enriched compared to other areas in the bay, and levels of
enrichment at these sites have increased disproportionately compared to other sites in the bay. This
suggests that the effluent from fish factories is still having a marked effect on the organic
composition of the sediments at a local scale (sites immediately adjacent to fish factory outfalls) but
not at a large scale.
Results from the benthic macrofauna monitoring programme
All benthic macrofauna (organisms >1 mm in size) in sediment samples collected from the bay were
identified to species level, counted, weighed, and assigned to major functional groups (filter feeders,
detritivores, predators, scavengers, and grazers). Statistical analyses were performed on these data
to assess spatial variability in the benthic macrofauna community structure and composition
between sites in 2012, and to assess changes in benthic community structure over time (i.e. in
relation to the 2001 and 2007 surveys). Data from 2012 suggest that the benthic macrofaunal
community in St Helena Bay was dominated in terms of abundance by the polychaete Diopatra
monroi and the amphipod Ampelisca anomala, and in terms of biomass, by the clam Venerupis
corrugata and the polychaete D. monroi. Filter feeders were the dominant functional group in terms
of biomass in 2012, while filter feeders, detritivores and predators all contributed in relatively similar
proportions to total abundance. The remaining groups (scavengers and grazers) contributed
relatively little to either biomass or abundance in 2012.
From a spatial perspective, benthic macrofauna communities at sampling stations in the Berg
estuary were significantly different from those in the rest of the bay. There were also some

Page | iii
significant differences among the sampling stations in the bay, with those from the eastern side of
the bay being distinct from those in the northern and western sectors, and those taken from the
immediate vicinity of the fish factory outfalls being distinct from one another and from other
stations in the bay. Abundance of macrofauna in the stations close to the fish factory outfalls was
on the whole much lower than those in the rest of the bay with the samples taken in the proximity
of the factory in Stompneusbaai being completely devoid of any macrofauna. Given that the
macrofauna on the west coast are typically opportunistic species, able to recover rapidly from
disturbances and that the communities at two stations were in such a poor state, it is evident that
effluent from these particular fish factories are having a negative impact upon the benthic
macrofaunal community at these sites. However, as the results of the 2007 survey indicated, these
impacts are limited to the local area around the fish factories discharge points.
From a temporal perspective, the overall abundance and biomass of macrofauna in the bay
decreased between 2001 and 2007 and then increased between 2007 and 2012. The overall
abundance of macrofauna in the bay reached the highest levels on record in 2012, being higher than
levels recorded in 2001. Changes between 2007 and 2012 were mostly associated with an increase
in the abundance of polychaetes and crustaceans (most notably amphipods) and a change from a
community dominated by detritivores to one with a fairly equal proportion of predators, detritivores
and filter feeders. Diversity values showed an opposite pattern to that of biomass and abundance
with diversity increasing between 2001 and 2007 and then decreasing again by 2012. These
fluctuations in the macrofaunal community are likely to be in response to large scale natural
disturbances, with few pioneering species dominating the benthos in high numbers following
disturbance. Low oxygen events resulting from high levels of productivity are an example of a large
scale natural disturbance that is likely to have resulted in these fluctuations in St Helena Bay.
By contrast, macrofauna abundance at the stations in the estuary decreased between 2007 and
2012 and shifted from one dominated by detritivores in 2007 to one dominated by filter feeders in
2012. The cause of this was not clear though and may be related to changes in freshwater runoff to
the estuary.

Page | iv
TABLE OF CONTENTS
Executive summary .................................................................................................................................. i
TABLE OF CONTENTS ............................................................................................................................. iv
1 INTRODUCTION ............................................................................................................................... 1
2 STATE OF THE BAY MONITORING ................................................................................................... 2
3 SAMPLING METHODS...................................................................................................................... 3
4 SEDIMENT ....................................................................................................................................... 6
4.1 Particle size ............................................................................................................................. 7
4.2 Organic matter ...................................................................................................................... 10
4.2.1 Total Organic Carbon .................................................................................................... 10
4.2.2 Total Organic Nitrogen .................................................................................................. 12
4.2.3 Carbon to nitrogen ratios .............................................................................................. 14
5 BENTHIC MACROFAUNA ............................................................................................................... 16
5.1 Methods ................................................................................................................................ 16
5.1.1 Laboratory Analysis ....................................................................................................... 16
5.1.2 Statistical Analysis ......................................................................................................... 16
5.2 Results ................................................................................................................................... 18
5.2.1 Community structure and composition ........................................................................ 18
5.2.2 Diversity indices ............................................................................................................ 31
6 INTEGRATION OF PHYSICO-CHEMICAL PARAMETERS AND BIOTIC INDICATORS ......................... 33
6.1 Methods ................................................................................................................................ 33
6.2 Results ................................................................................................................................... 33
7 DISCUSSION ................................................................................................................................... 35
7.1 Bay-wide variability ............................................................................................................... 35
7.2 Fish factory sites ................................................................................................................... 38
7.3 Estuarine sites ....................................................................................................................... 38
8 CONCLUSIONS ............................................................................................................................... 40
9 REFERENCES .................................................................................................................................. 41

Page | 1
1 INTRODUCTION
St Helena Bay is situated on the west coast of South Africa and extends from Dwarskersbos in the
north, past the Shelley Point peninsula, to Cape St Martin in the west, encompassing 18 smaller bays
and the estuary of the Berg River. The Bay is positioned in the southern section of the Benguela
Current System (BCS) which extends along the west coast of southern Africa between Cape Agulhas
(South Africa) and the Congo River mouth (Angola). The BCS is one of four major eastern-boundary
current systems which is characterised by the wind-driven upwelling of cold, nutrient rich water
(Shannon & O’Toole 1998). This cold current originates from the South Atlantic Circulation, which
circles just north of the Arctic Circumpolar Current. The naturally cool temperature of the Benguela
current (average temperature 10-14oC) is enhanced by the upwelling of the even cooler nutrient-rich
deep water (Branch 1981). The area experiences strong southerly and south-easterly winds which
are deflected by the Coriolis forces (rotational force of the earth which causes objects in the
southern hemisphere to spin anticlockwise). These prevailing conditions deflect the surface waters
offshore and cause cold, nutrient rich water to upwell against the shore . This water is the nutrient
rich life force of the west coast. Phytoplankton bloom when the nutrients reach the surface waters
where plenty of light is available for photosynthesis, and the phytoplankton is then preyed upon by
zooplankton, which is in turn eaten by filer feeding fish such as anchovy or sardine. This makes the
west coast one of the richest fishing grounds in the world and also attracts large colonies of sea birds
and seals (Branch 1981).
St Helena Bay is positioned downstream of the Cape Columbine upwelling cell and is a retention
zone for the nutrient rich water that is upwelled in this cell (Monteiro and Roychoudhury 2005). St
Helena Bay functions as a productive nursery to early life stages of fish (Bakun 1998, Monteiro and
Roychoudhury 2005), however it is also subject to incidence of harmful algal blooms and regular
episodes of oxygen depletion in the coastal waters, which have in the past lead to major mortality
events for organisms such as rock lobsters and fish (Cockroft et al. 2000, Monteiro & Roychoudhury
2005). A rock lobster reserve has also been declared within the bay in terms of the Marine Living
Resources Act. The Berg River Estuary, which is considered part of the Bay, is one of a few estuaries
draining off the west coast and is ecologically important for both fisheries and birds. Tourism is also
becoming an increasingly important industry supported by the bay given its picturesque and
sheltered nature and the variety of recreational opportunities it offers including sailing, canoeing,
surfing, bathing, diving, kitesurfing, fishing and beach activities.
St Helena Bay was the centre of the pelagic fishing industry when it began in the 1940s and has been
an important part of the West Coast lobster fishing grounds (von Bonde & Merchand 1935,
Hutchings et. al. 2012). The Bay still functions as a major node for industrial and small-scale fisheries
on the west coast. Indeed, the fishing industry is the mainstay of St Helena Bay with the bulk of
South Africa’s fish production and processing being conducted in the St Helena Bay factories. The
factories in the bay produce a variety of products including tinned fish, fresh and frozen hake, fish-
meal and live crayfish for export. Other industries located around the periphery of the bay include
mariculture, ship repair and shipbuilding.

Page | 2
2 STATE OF THE BAY MONITORING
Regular, long-term environmental monitoring is essential to identify impacts and to implement
measures to minimise negative human impacts on the environment (e.g. pollution) and maintain the
beneficial value of an area. This is particularly important to an area such as St Helena Bay, which
serves as a major node for industrial as well as small scale fisheries, while at the same time
supporting important ecological functions and a growing tourism industry. Fish processing and
vessel operations in St Helena Bay have in the past been shown to impact marine biota and water
quality in the Bay, while other industrial, residential and tourism developments also have the
potential to negatively impact on ecosystem health of the Bay. Various methods are available to
monitor the health of the environment, including the measurement of physical parameters (e.g.
water temperature, oxygen levels, and circulation patterns), actual pollutants in the water column,
sediments or tissue of biota (e.g. heavy metals, hydrocarbons, microbiological indicators) and
biological components of the ecosystem (e.g. birds, fish, and invertebrates).
Organic matter is one of the most universal pollutants affecting marine life and if it is allowed to
accumulate to high levels or if it is introduced into the environment at a rate faster than it can be
assimilated, it can lead to significant community disturbance. Organic loading is a particular concern
for St Helena Bay given the number of fish processing facilities operational in the bay and the fact
that the bay is a retention zone. High organic loading typically leads to eutrophication, which may
bring about a number of community responses. These include increased growth rates by primary
producers, disappearance of organisms due to hypoxia or anoxia, changes in community
composition and reduction in the number of species following repeat hypoxia and even complete
disappearance of benthic organisms in severely eutrophic and anoxic sediments (Warwick 1993).
It is important to monitor biological criteria in addition to physico-chemical and ecotoxicological
variables as biological indicators provide a direct measure of the state of the ecosystem. Benthic
macrofauna are the biotic component most frequently monitored to detect changes in the health of
the marine environment. This is primarily because these species are short lived and therefore their
community composition responds noticeably to changes in environment quality over time (Warwick
1993). Given that they are also relatively non-mobile (as compared with fish and birds) they tend to
be directly affected by pollution and they are easy to sample quantitatively (Warwick 1993).
Furthermore they are well-studied scientifically, in comparison to other sediment-dwelling
components (e.g. meiofauna and microfauna) and taxonomic keys are available for most groups. In
addition, community response to a number of anthropogenic influences has been well documented.
The major industries as well as other stakeholder in the St Helena Bay area are represented on the St
Helena Bay Water Quality Trust. The Trust has established a long term monitoring programme,
which monitors water quality, sediment quality and biotic indices of health, to determine the overall
health of the bay and track changes that may be caused by human activities. Sediment quality and
benthic macrofaunal communities are monitored approximately every five years. The results of
these assessments are used to make recommendations regarding the sustainable management of
activities in the bay.
This report presents results on sediment characteristics, the levels of organic material in the
sediments, and the abundance and distribution of benthic macrofauna living in the sediments of the

Page | 3
bay in 2012. It is the third assessment conducted as part of the State of the Bay monitoring program
which commenced in 2001. The methods used in this assessment replicated the sampling approach
used in previous State of the Bay surveys as closely as possible, to minimise any variations in
macrofauna abundance or composition that may be introduced from this source.
3 SAMPLING METHODS
Previously (2001 and 2007) sediment samples were collected from 33 sites across several months
period (April-August) throughout the Bay using a van Veen Grab or an Ocean Instruments multicorer.
All samples were analysed for trace metal content, acid volatile sulphides, sediment particle size and
particulate organic carbon and nitrogen. Benthic macrofauna samples were collected at 17 sites
distributed throughout the Bay using a Van Veen grab operated from a Rigid Inflatable Boat (RIB).
The dimensions of the grab used were 0.33 x 0.33 m resulting in a sampled area of ~0.1 m2/grab.
The sediment was washed through a 1 mm mesh sieve. Benthic macrofauna retained on the sieve
was washed into labelled sample bottles, fixed in ~4% formaldehyde solution, and transferred to the
laboratory for processing. One replicate was taken at the majority of sites in the Bay and multiple (3
– 5) replicates were taken at sites in the vicinity of the fish farms. Sites were sampled over three
days spread between April and July of 2007.
In this study (2012) benthic macrofauna and sediment samples were collected from 27 sites
throughout St Helena Bay over a 3 day period during the month of April. Sampling was conducted
using a Van Veen grab operated from a RIB. The dimensions of the grab used were 0.34 x 0.42 m
resulting in a sampled area of ~0.143 m2/grab. A subsample from each grab was collected and used
for sediment texture and organic content analysis and the balance of the sediment was washed
through a 1 mm mesh sieve. Benthic macrofauna retained on the sieve were washed into labelled
sample bottles, fixed in ~4% formaldehyde solution, and transferred to the laboratory for further
processing.
Figure 1: The van Veen Grab used for sediment and benthic macrofauna sample collection in 2012 and the filtering of sediments through a 1mm mesh bag.
Page | 4
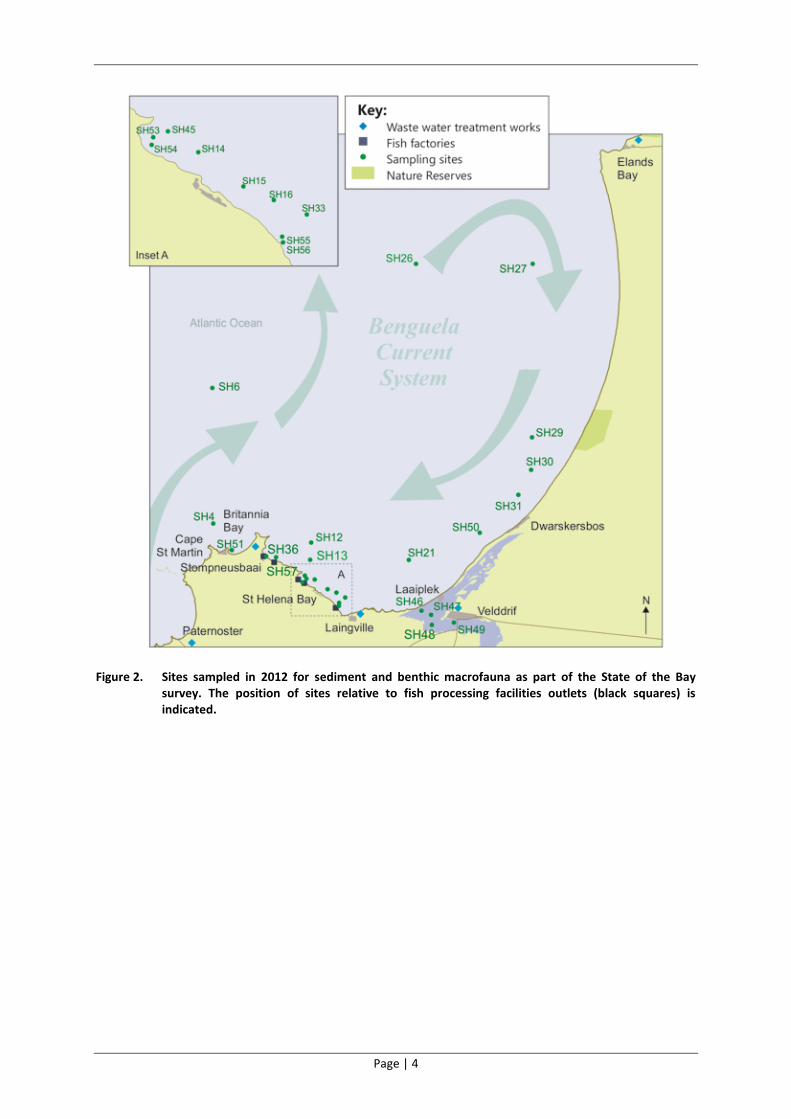
Figure 2. Sites sampled in 2012 for sediment and benthic macrofauna as part of the State of the Bay survey. The position of sites relative to fish processing facilities outlets (black squares) is indicated.
Page | 5
Table 1 Sites sampled for benthic macrofauna and sediment in St Helena Bay during the 2001, 2007 and 2012 State of the Bay surveys.
Lat (S) Lon (E) Site Name
2001 2007 2012
Macrofauna Sediment Macrofauna Sediment Macrofauna and
Sediment
32.6950 17.9250 SH3 X X Not sampled X Not sampled
32.6878 17.9255 SH4 X X Not sampled X X
32.5502 17.9243 SH6 X X X X X
32.6727 18.0237 SH11 X X Not sampled X Not sampled
32.7078 18.0240 SH12 X X X X X
32.7252 18.0233 SH13 X X Not sampled X X
32.7498 18.0280 SH14 X X Not sampled X X
32.7592 18.0403 SH15 X X Not sampled X X
32.7632 18.0492 SH16 X X Not sampled X X
32.7250 18.1238 SH21 X X X X X
32.4245 18.1248 SH26 X X X X X
32.4248 18.2492 SH27 X X Not sampled X X
32.6007 18.2485 SH29 X X X X X
32.6330 18.2480 SH30 X X X X X
32.6587 18.2355 SH31 X X X X X
32.7673 18.0583 SH33 X X Not sampled X X
32.7510 18.0582 SH34 X X Not sampled X Not sampled
32.7257 18.0590 SH35 X X Not sampled X Not sampled
32.7230 17.9888 SH36 X X Not sampled X X
32.7000 17.9500 SH40 Not sampled Not sampled Not sampled X Not sampled
32.7439 18.0191 SH45 Not sampled Not sampled Not sampled X X
32.7766 18.1363 SH46 Not sampled Not sampled X X X
32.7806 18.1460 SH47 Not sampled Not sampled X X X
32.7907 18.1465 SH48 Not sampled Not sampled X X X
32.7881 18.1687 SH49 Not sampled Not sampled X X X
32.6968 18.1960 SH50 X (40) X (40) X X X
32.7180 17.9437 SH51 X (37) X (37) Not sampled X X
32.7169 17.9342 SH52 Not sampled Not sampled Not sampled X Not sampled
32.7457 18.0151 SH53 Not sampled Not sampled X (SH166) X X
32.7478 18.0145 SH54 Not sampled Not sampled X (SH167) X X
32.7736 18.0514 SH55 Not sampled Not sampled X (SH168) X X
32.7753 18.0513 SH56 Not sampled Not sampled X (SH168) X X
32.7233 17.9778 SH57 Not sampled Not sampled X (SH170) X X
Page | 6
4 SEDIMENT
The sediment samples were tested for Total Organic Carbon (TOC) and Total Organic Nitrogen (TON)
by the CSIR using the standard laboratory method CSIR (MALS 3.3) (CSIR 2008). The particle size
composition was determined by Scientific Services using the following sieve mesh sizes: 2000 µm,
1000 µm, 850µm, 710µm, 500µm, 425µm, 300µm, 250µm, 125µm and 63µm.
The assessment of these results considered both the spatial distribution of different particle size
groups and organic content as well as the changes in distribution and the extent of organic loading in
2012 relative to 2001 and 2007. TOC and TON were however not assessed in 2001 and therefore the
temporal assessment of organic loading was only conducted for 2007 and 2012.
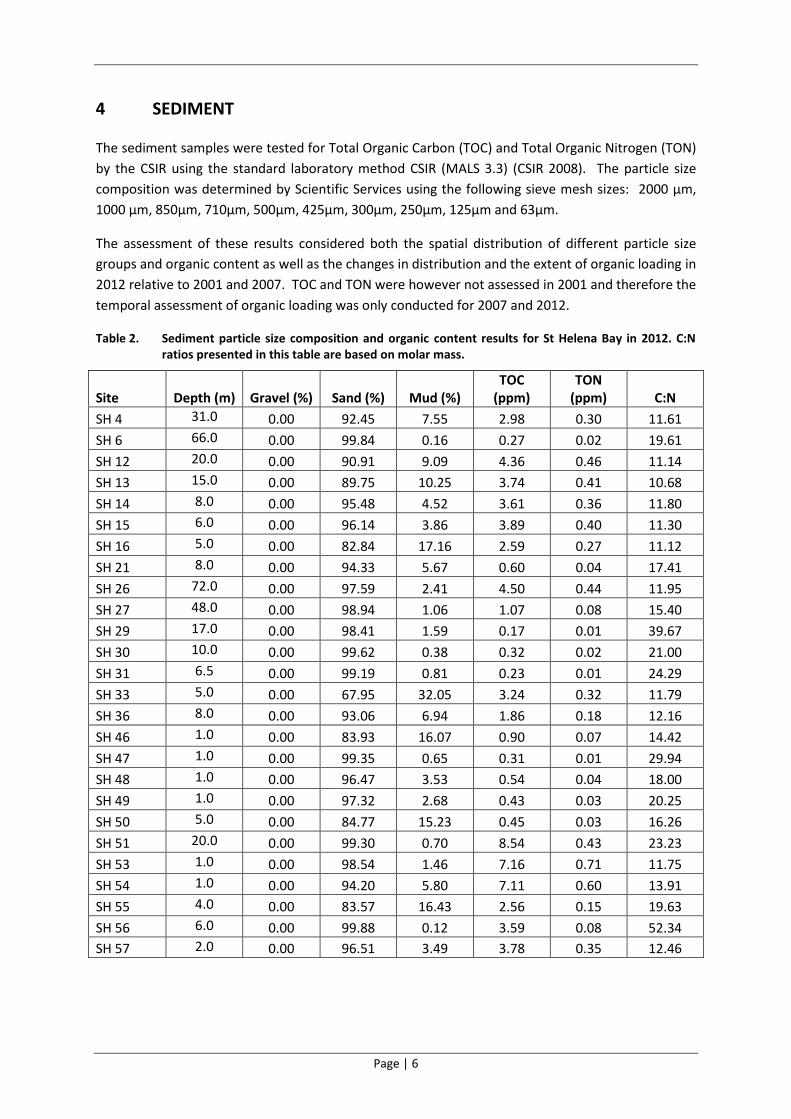
Table 2. Sediment particle size composition and organic content results for St Helena Bay in 2012. C:N ratios presented in this table are based on molar mass.
Site Depth (m) Gravel (%) Sand (%) Mud (%) TOC
(ppm) TON
(ppm) C:N
SH 4 31.0 0.00 92.45 7.55 2.98 0.30 11.61
SH 6 66.0 0.00 99.84 0.16 0.27 0.02 19.61
SH 12 20.0 0.00 90.91 9.09 4.36 0.46 11.14
SH 13 15.0 0.00 89.75 10.25 3.74 0.41 10.68
SH 14 8.0 0.00 95.48 4.52 3.61 0.36 11.80
SH 15 6.0 0.00 96.14 3.86 3.89 0.40 11.30
SH 16 5.0 0.00 82.84 17.16 2.59 0.27 11.12
SH 21 8.0 0.00 94.33 5.67 0.60 0.04 17.41
SH 26 72.0 0.00 97.59 2.41 4.50 0.44 11.95
SH 27 48.0 0.00 98.94 1.06 1.07 0.08 15.40
SH 29 17.0 0.00 98.41 1.59 0.17 0.01 39.67
SH 30 10.0 0.00 99.62 0.38 0.32 0.02 21.00
SH 31 6.5 0.00 99.19 0.81 0.23 0.01 24.29
SH 33 5.0 0.00 67.95 32.05 3.24 0.32 11.79
SH 36 8.0 0.00 93.06 6.94 1.86 0.18 12.16
SH 46 1.0 0.00 83.93 16.07 0.90 0.07 14.42
SH 47 1.0 0.00 99.35 0.65 0.31 0.01 29.94
SH 48 1.0 0.00 96.47 3.53 0.54 0.04 18.00
SH 49 1.0 0.00 97.32 2.68 0.43 0.03 20.25
SH 50 5.0 0.00 84.77 15.23 0.45 0.03 16.26
SH 51 20.0 0.00 99.30 0.70 8.54 0.43 23.23
SH 53 1.0 0.00 98.54 1.46 7.16 0.71 11.75
SH 54 1.0 0.00 94.20 5.80 7.11 0.60 13.91
SH 55 4.0 0.00 83.57 16.43 2.56 0.15 19.63
SH 56 6.0 0.00 99.88 0.12 3.59 0.08 52.34
SH 57 2.0 0.00 96.51 3.49 3.78 0.35 12.46

Page | 7
4.1 Particle size
The sediments sampled at all sites in St Helena Bay in 2012 were comprised predominantly of sand,
with a small proportion of mud and no gravel. The sites situated in the Bay in close proximity to the
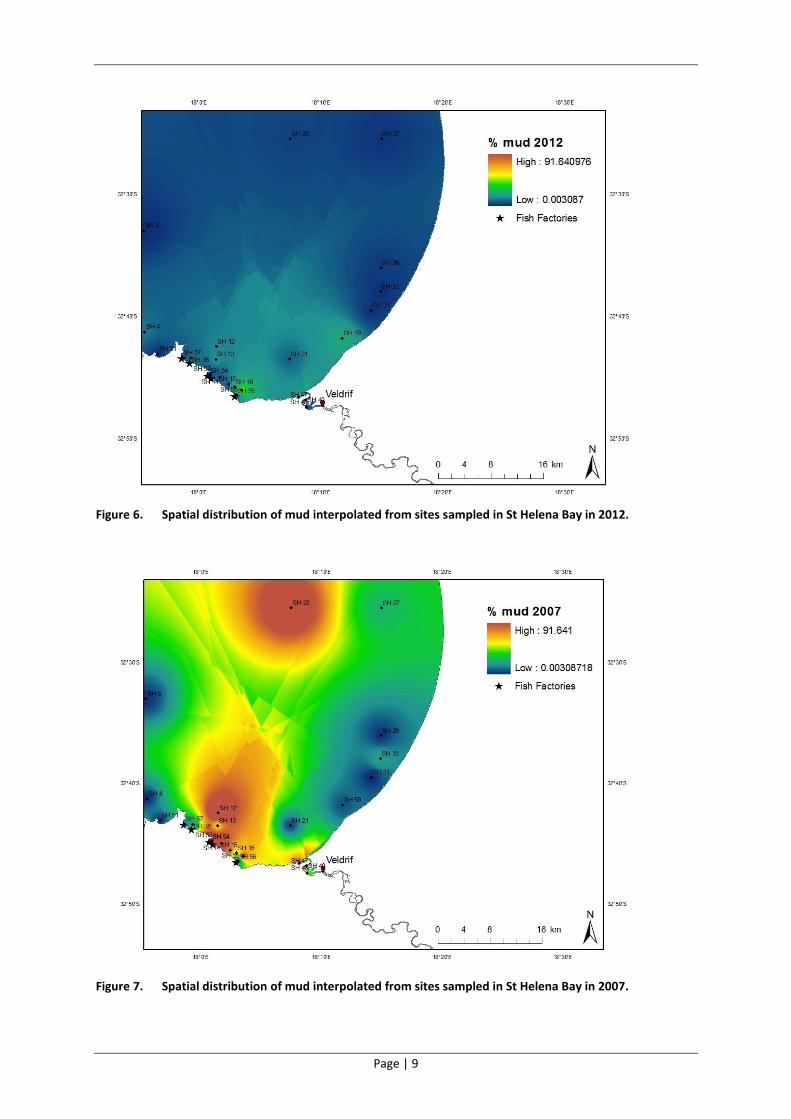
Berg River Estuary mouth had the highest proportion of mud (Figure 3 and Figure 6). Relative to
data from 2001 and 2007, the 2012 results indicate a dramatic reduction in the amount of mud in
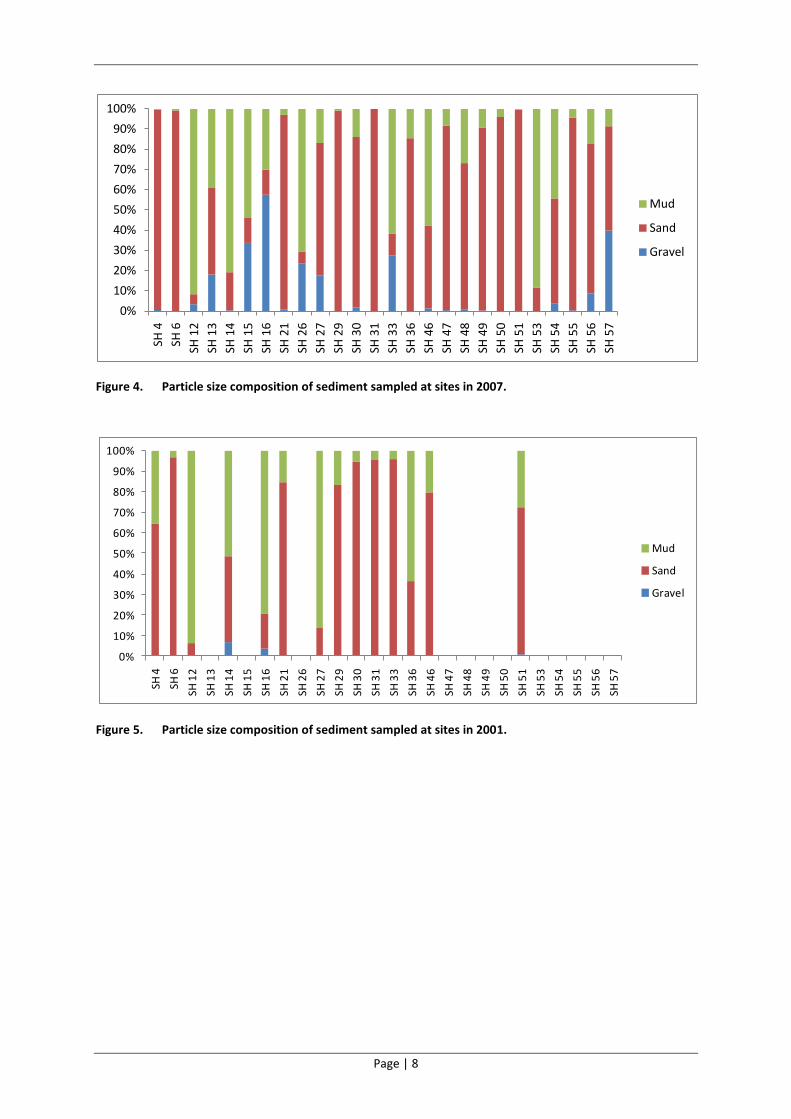
the sediments in St Helena Bay (Figure 3, Figure 4 and Figure 5). In addition, there has been a
significant reduction in the coarser gravel particle fraction in the bay since 2007 (Figure 4), making
the situation in 2012 (Figure 3) more similar to that evident in 2001 (Figure 5). The spatial
distribution of mud in the Bay has also changed dramatically over time, with areas exhibiting the
highest mud content having shifted from the northerly and southerly reaches of the Bay in 2007
(Figure 7) to the south western edge near the Berg River Estuary mouth (Figure 6).
Figure 3: Particle size composition of sediment sampled at sites in 2012.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 2
6
SH 2
7
SH 2
9
SH 3
0
SH 3
1
SH 3
3
SH 3
6
SH 4
6
SH 4
7
SH 4
8
SH 4
9
SH 5
0
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
Mud
Sand
Gravel
Page | 8
Figure 4. Particle size composition of sediment sampled at sites in 2007.
Figure 5. Particle size composition of sediment sampled at sites in 2001.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 2
6
SH 2
7
SH 2
9
SH 3
0
SH 3
1
SH 3
3
SH 3
6
SH 4
6
SH 4
7
SH 4
8
SH 4
9
SH 5
0
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
Mud
Sand
Gravel
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 2
6
SH 2
7
SH 2
9
SH 3
0
SH 3
1
SH 3
3
SH 3
6
SH 4
6
SH 4
7
SH 4
8
SH 4
9
SH 5
0
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
Mud
Sand
Gravel
Page | 9
Figure 6. Spatial distribution of mud interpolated from sites sampled in St Helena Bay in 2012.
Figure 7. Spatial distribution of mud interpolated from sites sampled in St Helena Bay in 2007.
Page | 10
4.2 Organic matter
4.2.1 Total Organic Carbon
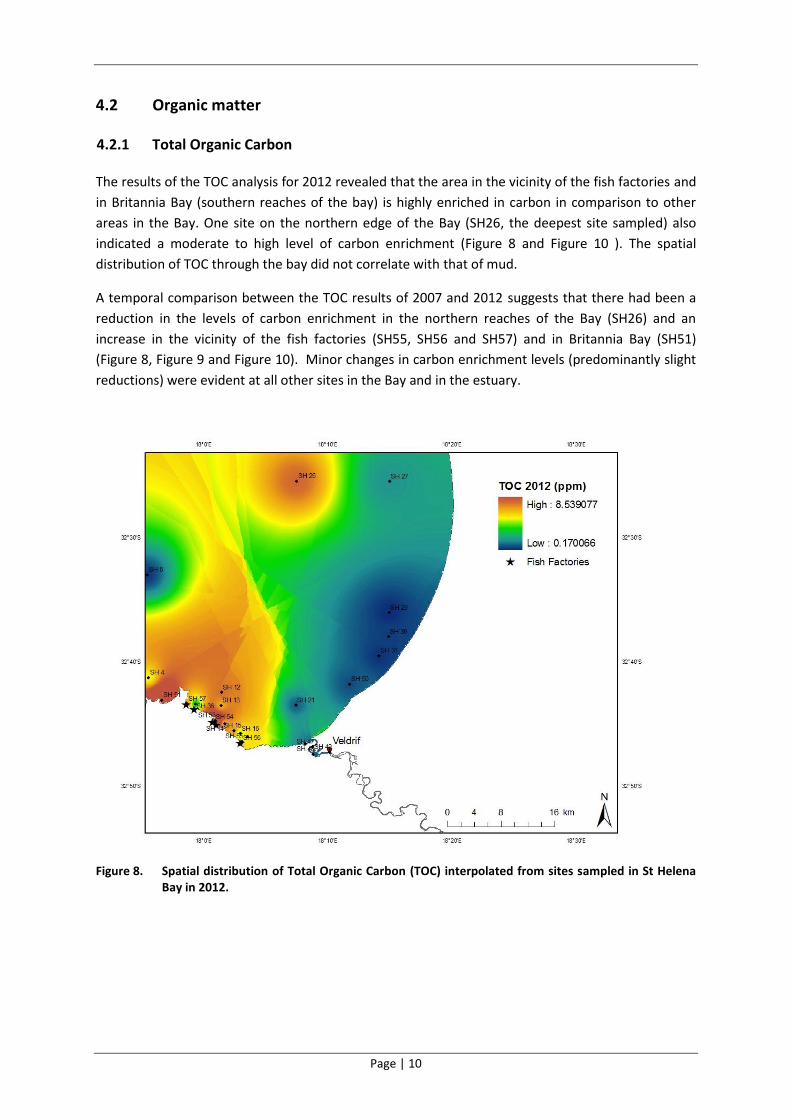
The results of the TOC analysis for 2012 revealed that the area in the vicinity of the fish factories and
in Britannia Bay (southern reaches of the bay) is highly enriched in carbon in comparison to other
areas in the Bay. One site on the northern edge of the Bay (SH26, the deepest site sampled) also
indicated a moderate to high level of carbon enrichment (Figure 8 and Figure 10 ). The spatial
distribution of TOC through the bay did not correlate with that of mud.
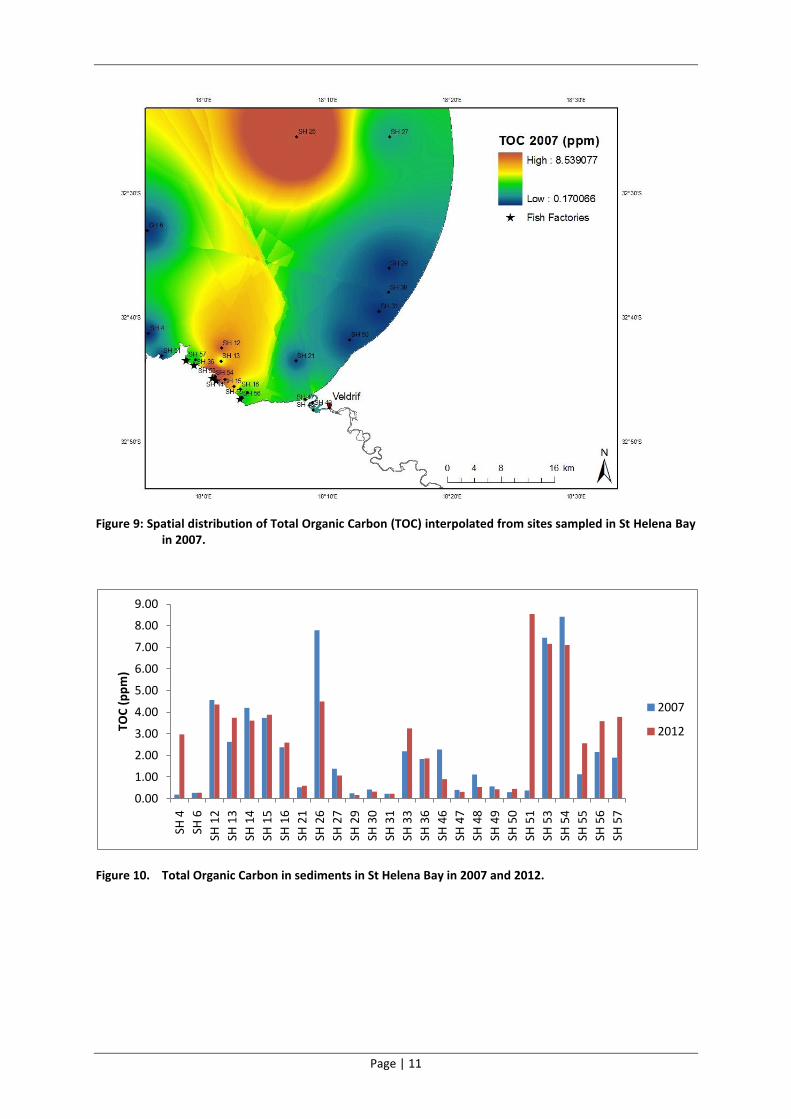
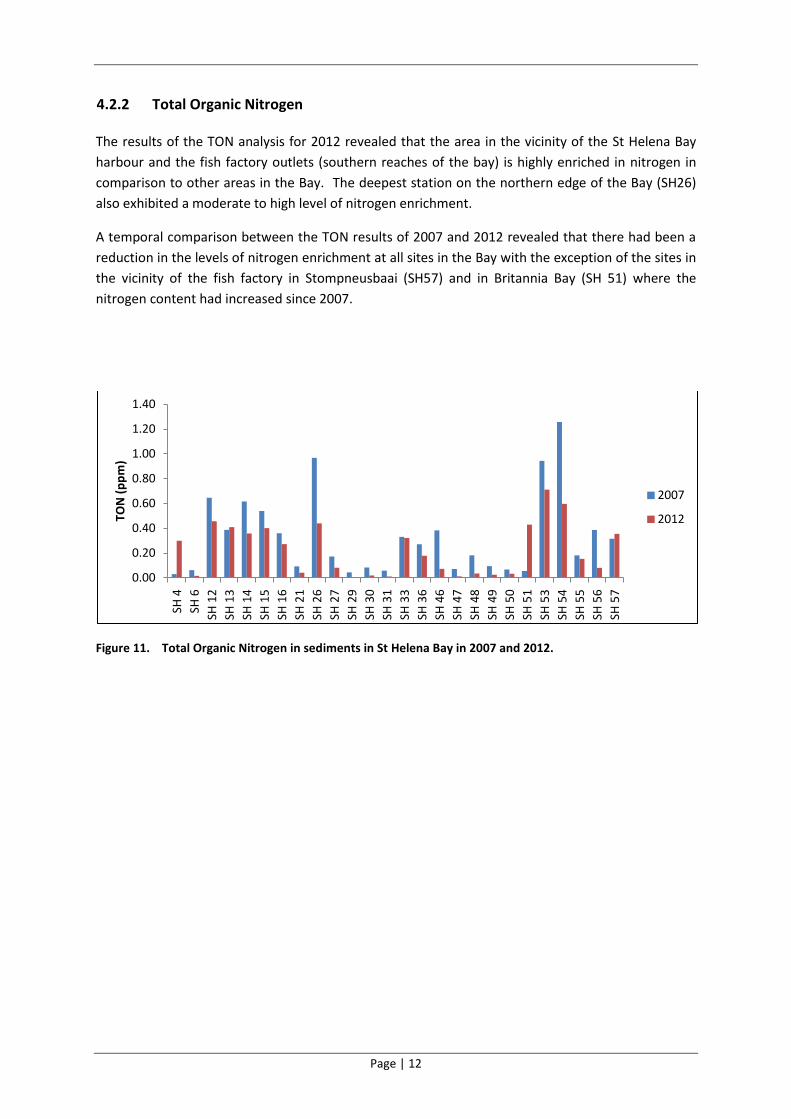
A temporal comparison between the TOC results of 2007 and 2012 suggests that there had been a
reduction in the levels of carbon enrichment in the northern reaches of the Bay (SH26) and an
increase in the vicinity of the fish factories (SH55, SH56 and SH57) and in Britannia Bay (SH51)
(Figure 8, Figure 9 and Figure 10). Minor changes in carbon enrichment levels (predominantly slight
reductions) were evident at all other sites in the Bay and in the estuary.
Figure 8. Spatial distribution of Total Organic Carbon (TOC) interpolated from sites sampled in St Helena Bay in 2012.
Page | 11
Figure 9: Spatial distribution of Total Organic Carbon (TOC) interpolated from sites sampled in St Helena Bay in 2007.
Figure 10. Total Organic Carbon in sediments in St Helena Bay in 2007 and 2012.
0.00
1.00
2.00
3.00
4.00
5.00
6.00
7.00
8.00
9.00
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 2
6
SH 2
7
SH 2
9
SH 3
0
SH 3
1
SH 3
3
SH 3
6
SH 4
6
SH 4
7
SH 4
8
SH 4
9
SH 5
0
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
TOC
(p
pm
)
2007
2012
Page | 12
4.2.2 Total Organic Nitrogen
The results of the TON analysis for 2012 revealed that the area in the vicinity of the St Helena Bay
harbour and the fish factory outlets (southern reaches of the bay) is highly enriched in nitrogen in
comparison to other areas in the Bay. The deepest station on the northern edge of the Bay (SH26)
also exhibited a moderate to high level of nitrogen enrichment.
A temporal comparison between the TON results of 2007 and 2012 revealed that there had been a
reduction in the levels of nitrogen enrichment at all sites in the Bay with the exception of the sites in
the vicinity of the fish factory in Stompneusbaai (SH57) and in Britannia Bay (SH 51) where the
nitrogen content had increased since 2007.
Figure 11. Total Organic Nitrogen in sediments in St Helena Bay in 2007 and 2012.
0.00
0.20
0.40
0.60
0.80
1.00
1.20
1.40
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 2
6
SH 2
7
SH 2
9
SH 3
0
SH 3
1
SH 3
3
SH 3
6
SH 4
6
SH 4
7
SH 4
8
SH 4
9
SH 5
0
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
TON
(p
pm
)
2007
2012

Page | 13
Figure 12. Spatial distribution of total organic nitrogen (TON) interpolated from sites sampled in St Helena Bay in 2012.
Figure 13. Spatial distribution of total organic nitrogen (TON) interpolated from sites sampled in St Helena Bay in 2007.
Page | 14
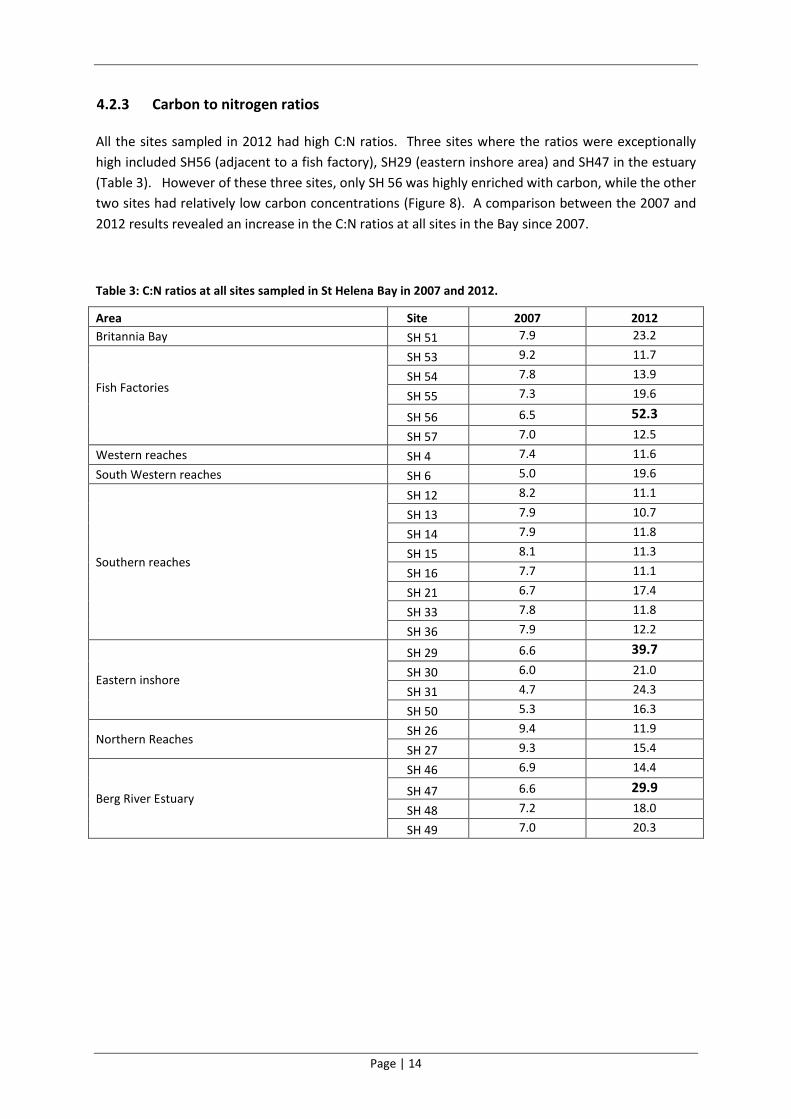
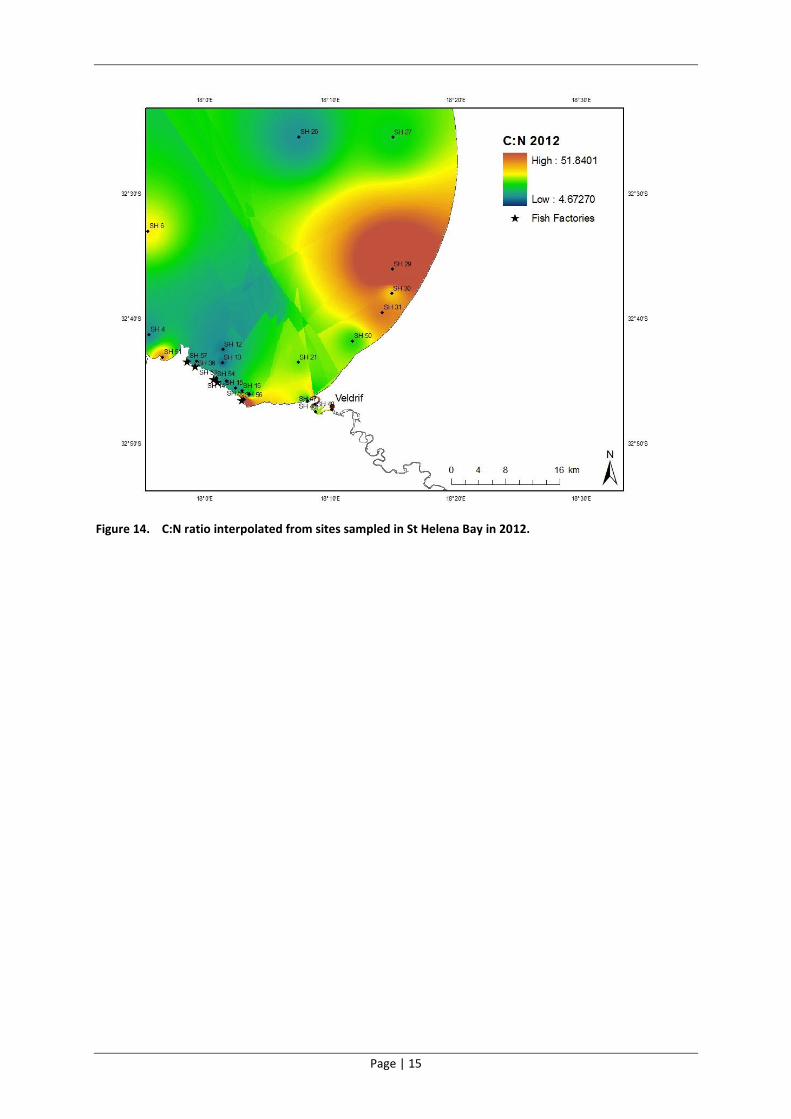
4.2.3 Carbon to nitrogen ratios
All the sites sampled in 2012 had high C:N ratios. Three sites where the ratios were exceptionally
high included SH56 (adjacent to a fish factory), SH29 (eastern inshore area) and SH47 in the estuary
(Table 3). However of these three sites, only SH 56 was highly enriched with carbon, while the other
two sites had relatively low carbon concentrations (Figure 8). A comparison between the 2007 and
2012 results revealed an increase in the C:N ratios at all sites in the Bay since 2007.
Table 3: C:N ratios at all sites sampled in St Helena Bay in 2007 and 2012.
Area Site 2007 2012
Britannia Bay SH 51 7.9 23.2
Fish Factories
SH 53 9.2 11.7
SH 54 7.8 13.9
SH 55 7.3 19.6
SH 56 6.5 52.3
SH 57 7.0 12.5
Western reaches SH 4 7.4 11.6
South Western reaches SH 6 5.0 19.6
Southern reaches
SH 12 8.2 11.1
SH 13 7.9 10.7
SH 14 7.9 11.8
SH 15 8.1 11.3
SH 16 7.7 11.1
SH 21 6.7 17.4
SH 33 7.8 11.8
SH 36 7.9 12.2
Eastern inshore
SH 29 6.6 39.7
SH 30 6.0 21.0
SH 31 4.7 24.3
SH 50 5.3 16.3
Northern Reaches SH 26 9.4 11.9
SH 27 9.3 15.4
Berg River Estuary
SH 46 6.9 14.4
SH 47 6.6 29.9
SH 48 7.2 18.0
SH 49 7.0 20.3
Page | 15
Figure 14. C:N ratio interpolated from sites sampled in St Helena Bay in 2012.

Page | 16
5 BENTHIC MACROFAUNA
5.1 Methods
5.1.1 Laboratory Analysis
In the laboratory, benthic macrofauna samples were washed to remove all traces of formaldehyde,
and transferred to 1% phenoxatol (ethylenglycolmonophenyl-ether). Thereafter all animals
considered alive at the time of collection (i.e. excluding empty shells) were identified to the lowest
taxon possible, blot dried and weighed to the nearest 0.001 g on a precision balance. The reference
collection from the 2007 survey was sourced and compared to that of the 2012 survey to ensure
consistency in the species identification. A majority of the species collected in 2001 were not
identified to a species level and no reference collection was available for this survey.
5.1.2 Statistical Analysis
The principle aim of monitoring the health of an area is to detect the effects of stress, as well as to
monitor recovery after an environmental perturbation. There are numerous indices, based on
benthic invertebrate fauna information, which can be used to reveal conditions and trends in the
state of ecosystems. These indices include those based on community composition, diversity and
species abundance and biomass. Given the complexity inherent in environmental assessment it is
recommended that several indices be used (Salas et al. 2006). The community composition,
diversity, and species abundance and biomass of soft bottom benthic macrofauna samples, collected
in St Helena Bay in 2012, are considered in this report.
The data collected from this survey were used for two purposes
1) to assess spatial variability in the benthic macrofauna community structure and composition
between sites in 2012 and
2) to assess changes in benthic community structure over time (temporal variation) (i.e. in relation
to the 2001 and 2007 surveys).
Both the spatial and temporal assessments are necessary to provide a good indication of the state of
the system.
Community structure and composition
The statistical program PRIMER 6 (Clarke and Warwick 1993) was used to conduct a spatial
assessment of the benthic macrofauna data collected in 2012 and a temporal comparison between
the macro benthic communities sampled in 2007 and 2012. The temporal comparison did not
include the 2001 data set as no reference collection was sourced and consistency in the
identification of species could not be guaranteed. Data were root-root (fourth root) transformed
and converted to a similarity matrix using the Bray-Curtis similarity coefficient. A cluster analysis
was performed in order to find ‘natural groupings’ between samples (sites). The results of the
cluster analysis are displayed on a dendrogram which graphically depicts the similarity among sites

Page | 17
by clustering them in groups. Statistically significant clusters of sites are revealed using a SIMPROF
analysis. These results were plotted geographically using GIS software to reveal any spatial trends in
the sites grouped in accordance with community composition similarity. SIMPER analysis was used
to identify species principally responsible for the clustering of sites. These results were used to
characterise different regions of sites based on the communities present at the sites. It is important
to remember that the community composition is a reflection of not only the physico-chemical health
of the environment but also the ability of communities to recover from disturbance.
A total of eight sites were sampled across all three surveys - 2001, 2007 and 2012. These sites were
all within the bay. No estuary or fish factory sites were sampled in 2001. The abundance and
relative proportions of benthic macrofauna taxonomic groups sampled at these eight sites were
compared.
Diversity Indices
A number of indices (single numbers) can be used as measures of community structure; these
include the total number of individuals (N), total number of species (S), the total biomass (B), and
the species equability or evenness, which is a measure of how evenly individuals are distributed
among different species. Diversity indices provide a measure of diversity, i.e. the way in which the
total number of individuals is divided up among different species. Understanding changes in benthic
diversity is important because increasing levels of environmental stress generally result in a decrease
in diversity.
Two different aspects of community structure contribute to community diversity, namely species
richness and equability (evenness). Species richness refers to the total number of species present
while equability or evenness expresses how evenly the individuals are distributed among different
species. A sample with greater evenness is considered to be more diverse. It is important to note
when interpreting diversity values that predation, competition and disturbance all play a role in
shaping a community. For this reason it is important to consider physical parameters as well as
other biotic indices when drawing a conclusion from a diversity index.
The following measures of diversity were calculated for each sampling location using PRIMER V 6:
The Shannon-Weiner diversity index (H’): H’ = - Σipi(log pi) (1)
where pi is the proportion of the total count arising from the ith species. This is the most
commonly used diversity measure and it incorporates both species richness and equability.
The Pielou’s evenness index (J’): J’ = H’observed / H’max (2)
where H’max is the maximum possible diversity which would be achieved if all species were
equally abundant (= log S). This is the most common expression of equability.
The Margalef’s index (d) of species richness: D = (S-1)/ log N (3)
where S is the total number of species and N is the total number of individuals.
Page | 18
Species richness is often simply referred to as the total number of species (S), but this is dependent
on sample size. The Margalef’s index thus incorporates the total number of individuals (N) and is a
measure of the total number of species present for a given number of individuals.
The diversity (H’) value for each site was plotted geographically and this was used to interpolate
values for the entire system in order to reveal any spatial patterns. Eight sites were sampled in all
three years. The diversity and abundance values for each of these years was compared.
5.2 Results
5.2.1 Community structure and composition
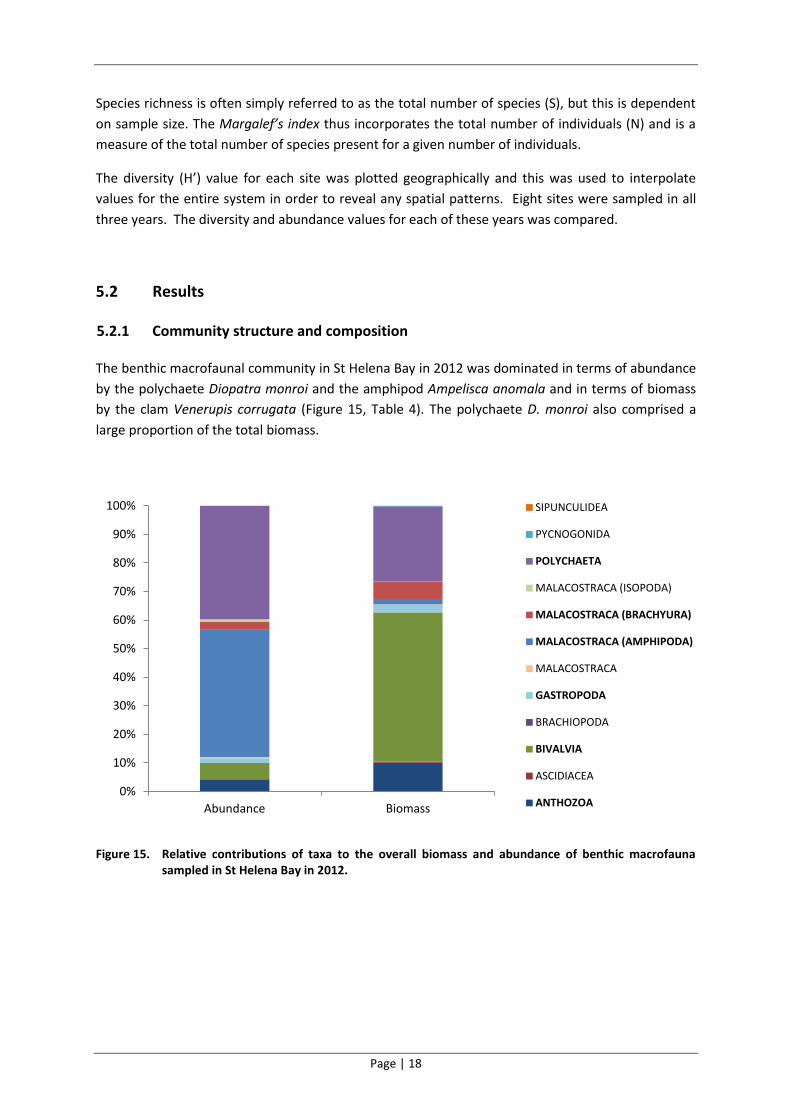
The benthic macrofaunal community in St Helena Bay in 2012 was dominated in terms of abundance
by the polychaete Diopatra monroi and the amphipod Ampelisca anomala and in terms of biomass
by the clam Venerupis corrugata (Figure 15, Table 4). The polychaete D. monroi also comprised a
large proportion of the total biomass.
Figure 15. Relative contributions of taxa to the overall biomass and abundance of benthic macrofauna sampled in St Helena Bay in 2012.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Abundance Biomass
SIPUNCULIDEA
PYCNOGONIDA
POLYCHAETA
MALACOSTRACA (ISOPODA)
MALACOSTRACA (BRACHYURA)
MALACOSTRACA (AMPHIPODA)
MALACOSTRACA
GASTROPODA
BRACHIOPODA
BIVALVIA
ASCIDIACEA
ANTHOZOA
Page | 19
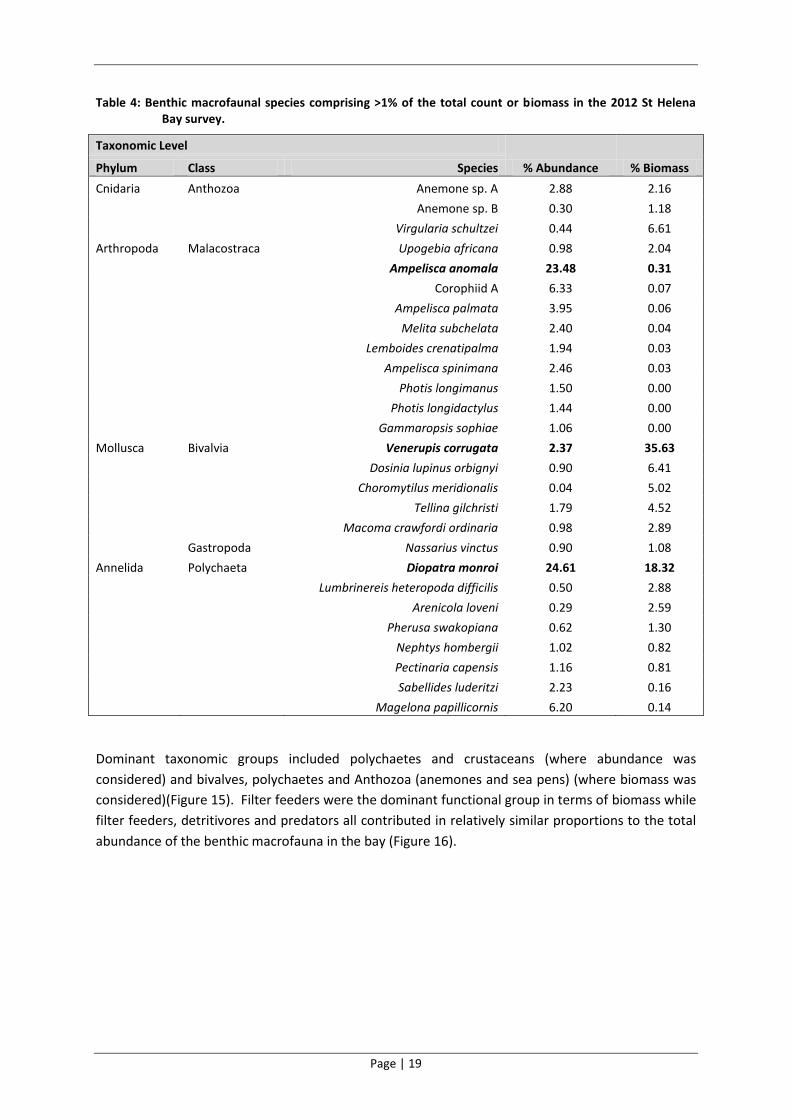
Table 4: Benthic macrofaunal species comprising >1% of the total count or biomass in the 2012 St Helena Bay survey.
Taxonomic Level
% Abundance % Biomass Phylum Class Species
Cnidaria Anthozoa Anemone sp. A 2.88 2.16
Anemone sp. B 0.30 1.18
Virgularia schultzei 0.44 6.61
Arthropoda Malacostraca Upogebia africana 0.98 2.04
Ampelisca anomala 23.48 0.31
Corophiid A 6.33 0.07
Ampelisca palmata 3.95 0.06
Melita subchelata 2.40 0.04
Lemboides crenatipalma 1.94 0.03
Ampelisca spinimana 2.46 0.03
Photis longimanus 1.50 0.00
Photis longidactylus 1.44 0.00
Gammaropsis sophiae 1.06 0.00
Mollusca Bivalvia Venerupis corrugata 2.37 35.63
Dosinia lupinus orbignyi 0.90 6.41
Choromytilus meridionalis 0.04 5.02
Tellina gilchristi 1.79 4.52
Macoma crawfordi ordinaria 0.98 2.89
Gastropoda Nassarius vinctus 0.90 1.08
Annelida Polychaeta Diopatra monroi 24.61 18.32
Lumbrinereis heteropoda difficilis 0.50 2.88
Arenicola loveni 0.29 2.59
Pherusa swakopiana 0.62 1.30
Nephtys hombergii 1.02 0.82
Pectinaria capensis 1.16 0.81
Sabellides luderitzi 2.23 0.16
Magelona papillicornis 6.20 0.14
Dominant taxonomic groups included polychaetes and crustaceans (where abundance was
considered) and bivalves, polychaetes and Anthozoa (anemones and sea pens) (where biomass was
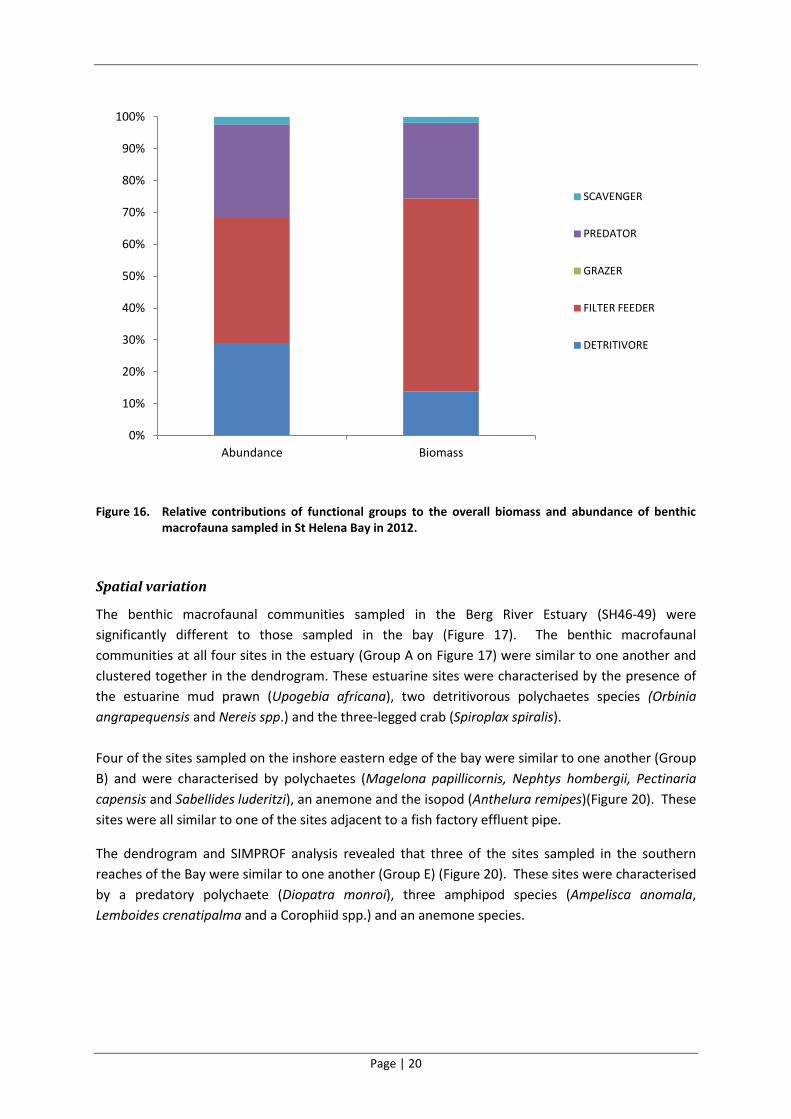
considered)(Figure 15). Filter feeders were the dominant functional group in terms of biomass while
filter feeders, detritivores and predators all contributed in relatively similar proportions to the total
abundance of the benthic macrofauna in the bay (Figure 16).
Page | 20
Figure 16. Relative contributions of functional groups to the overall biomass and abundance of benthic macrofauna sampled in St Helena Bay in 2012.
Spatial variation
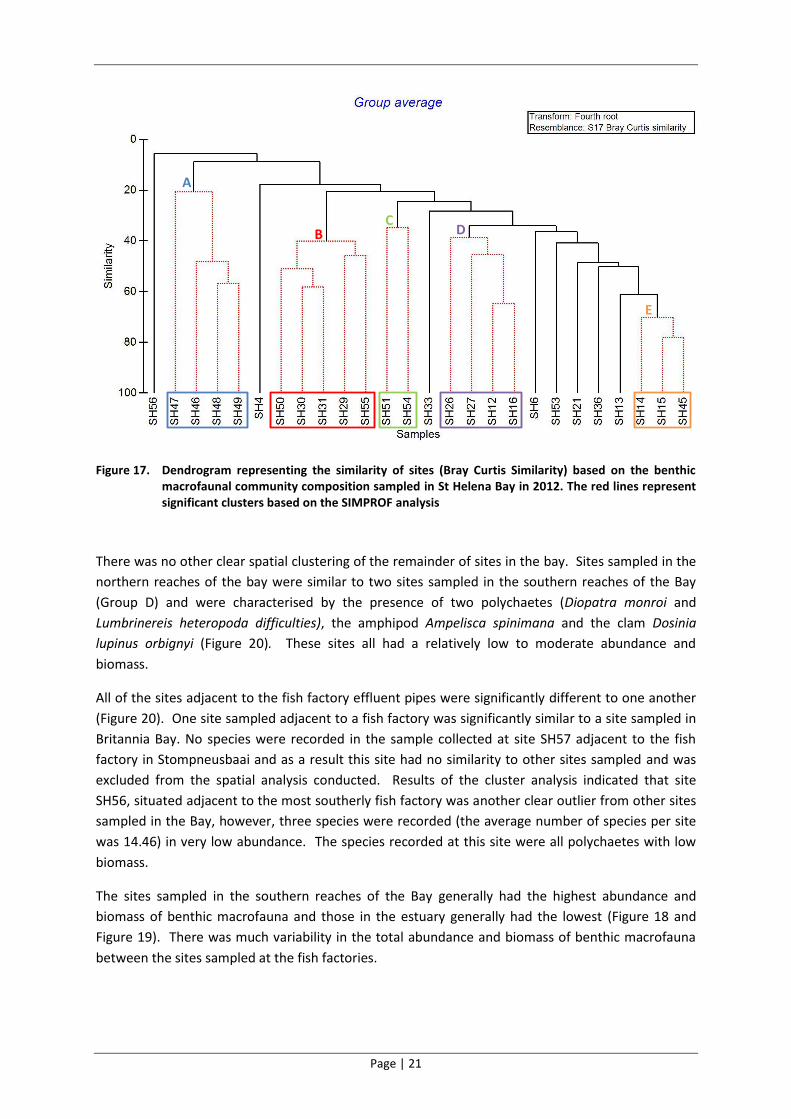
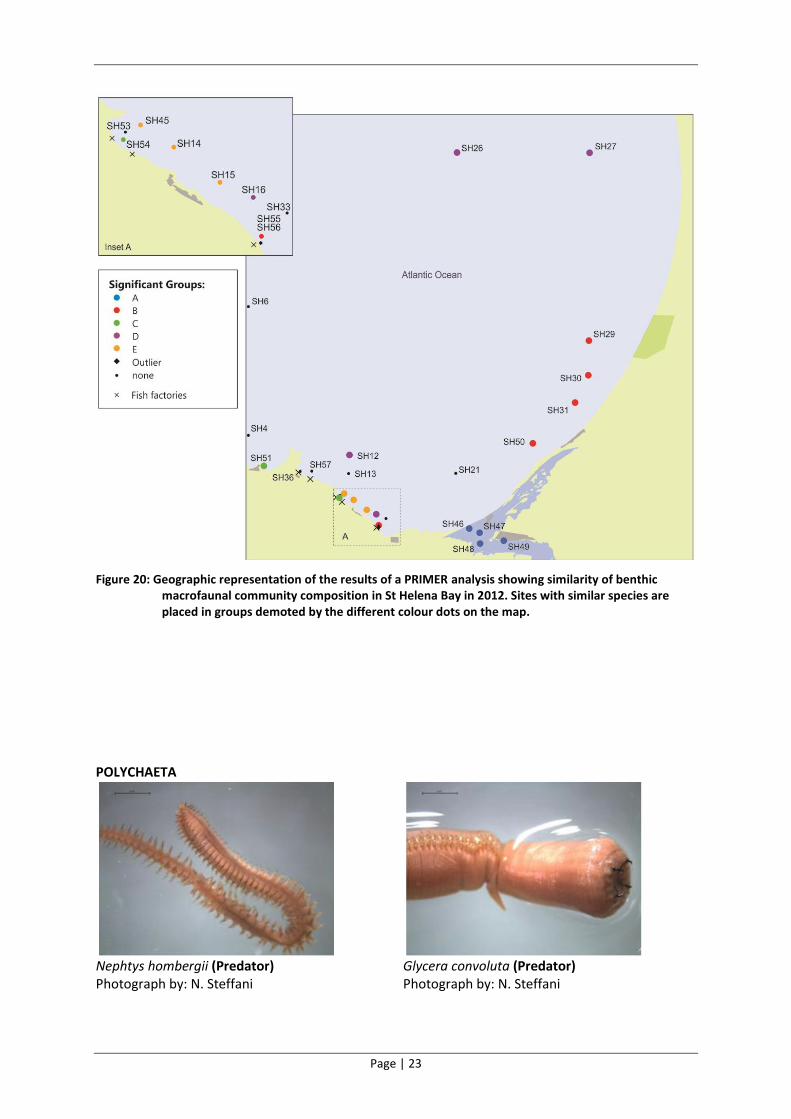
The benthic macrofaunal communities sampled in the Berg River Estuary (SH46-49) were
significantly different to those sampled in the bay (Figure 17). The benthic macrofaunal
communities at all four sites in the estuary (Group A on Figure 17) were similar to one another and
clustered together in the dendrogram. These estuarine sites were characterised by the presence of
the estuarine mud prawn (Upogebia africana), two detritivorous polychaetes species (Orbinia
angrapequensis and Nereis spp.) and the three-legged crab (Spiroplax spiralis).
Four of the sites sampled on the inshore eastern edge of the bay were similar to one another (Group
B) and were characterised by polychaetes (Magelona papillicornis, Nephtys hombergii, Pectinaria
capensis and Sabellides luderitzi), an anemone and the isopod (Anthelura remipes)(Figure 20). These
sites were all similar to one of the sites adjacent to a fish factory effluent pipe.
The dendrogram and SIMPROF analysis revealed that three of the sites sampled in the southern
reaches of the Bay were similar to one another (Group E) (Figure 20). These sites were characterised
by a predatory polychaete (Diopatra monroi), three amphipod species (Ampelisca anomala,
Lemboides crenatipalma and a Corophiid spp.) and an anemone species.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Abundance Biomass
SCAVENGER
PREDATOR
GRAZER
FILTER FEEDER
DETRITIVORE
Page | 21
Figure 17. Dendrogram representing the similarity of sites (Bray Curtis Similarity) based on the benthic macrofaunal community composition sampled in St Helena Bay in 2012. The red lines represent significant clusters based on the SIMPROF analysis
There was no other clear spatial clustering of the remainder of sites in the bay. Sites sampled in the
northern reaches of the bay were similar to two sites sampled in the southern reaches of the Bay
(Group D) and were characterised by the presence of two polychaetes (Diopatra monroi and
Lumbrinereis heteropoda difficulties), the amphipod Ampelisca spinimana and the clam Dosinia
lupinus orbignyi (Figure 20). These sites all had a relatively low to moderate abundance and
biomass.
All of the sites adjacent to the fish factory effluent pipes were significantly different to one another
(Figure 20). One site sampled adjacent to a fish factory was significantly similar to a site sampled in
Britannia Bay. No species were recorded in the sample collected at site SH57 adjacent to the fish
factory in Stompneusbaai and as a result this site had no similarity to other sites sampled and was
excluded from the spatial analysis conducted. Results of the cluster analysis indicated that site
SH56, situated adjacent to the most southerly fish factory was another clear outlier from other sites
sampled in the Bay, however, three species were recorded (the average number of species per site
was 14.46) in very low abundance. The species recorded at this site were all polychaetes with low
biomass.
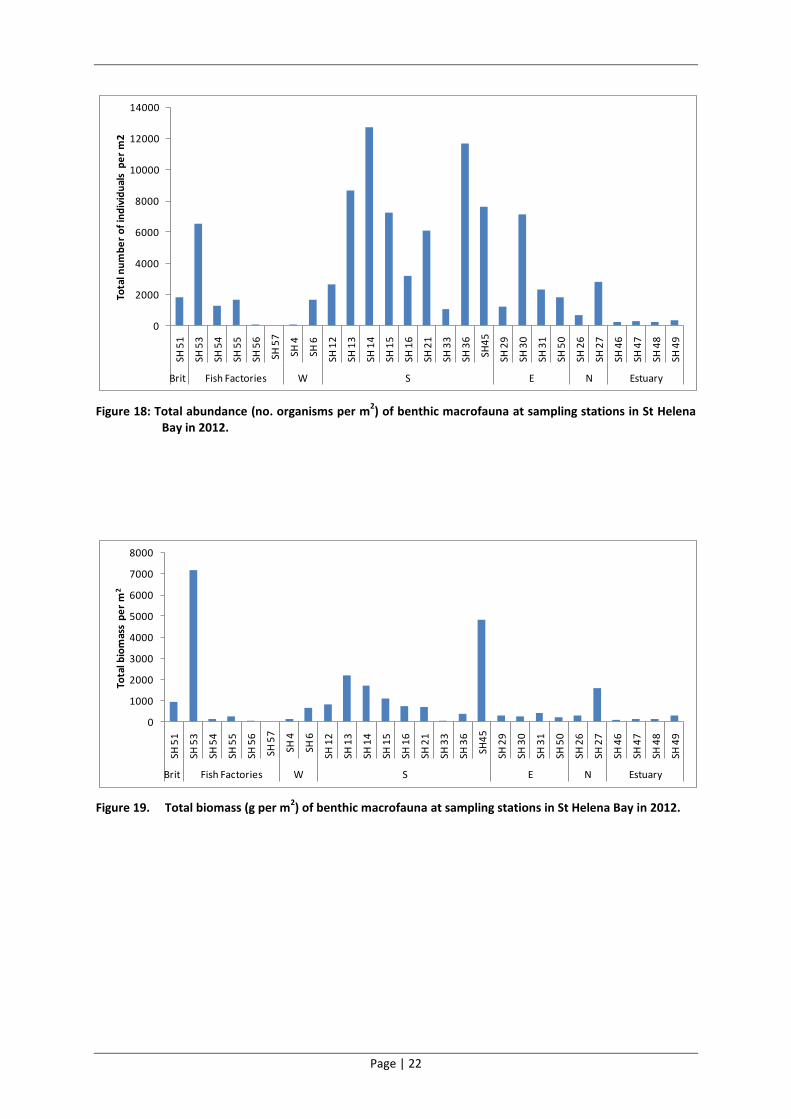
The sites sampled in the southern reaches of the Bay generally had the highest abundance and
biomass of benthic macrofauna and those in the estuary generally had the lowest (Figure 18 and
Figure 19). There was much variability in the total abundance and biomass of benthic macrofauna
between the sites sampled at the fish factories.
A
BC
D
E
Page | 22
Figure 18: Total abundance (no. organisms per m2) of benthic macrofauna at sampling stations in St Helena
Bay in 2012.
Figure 19. Total biomass (g per m2) of benthic macrofauna at sampling stations in St Helena Bay in 2012.
0
2000
4000
6000
8000
10000
12000
14000
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 3
3
SH 3
6
SH4
5
SH 2
9
SH 3
0
SH 3
1
SH 5
0
SH 2
6
SH 2
7
SH 4
6
SH 4
7
SH 4
8
SH 4
9
Brit Fish Factories W S E N Estuary
Tota
l nu
mb
er
of
ind
ivid
ual
s p
er
m2
0
1000
2000
3000
4000
5000
6000
7000
8000
SH 5
1
SH 5
3
SH 5
4
SH 5
5
SH 5
6
SH 5
7
SH 4
SH 6
SH 1
2
SH 1
3
SH 1
4
SH 1
5
SH 1
6
SH 2
1
SH 3
3
SH 3
6
SH4
5
SH 2
9
SH 3
0
SH 3
1
SH 5
0
SH 2
6
SH 2
7
SH 4
6
SH 4
7
SH 4
8
SH 4
9
Brit Fish Factories W S E N Estuary
Tota
l bio
mas
s p
er
m2
Page | 23
Figure 20: Geographic representation of the results of a PRIMER analysis showing similarity of benthic macrofaunal community composition in St Helena Bay in 2012. Sites with similar species are placed in groups demoted by the different colour dots on the map.
POLYCHAETA
Nephtys hombergii (Predator) Photograph by: N. Steffani
Glycera convoluta (Predator) Photograph by: N. Steffani

Page | 24
MALACOSTRACA (Amphipoda)
Hippomedon normalis (Scavenger) Photograph by: N. Steffani
Ampelisca brevicornis (Filter feeder) Photograph by: N. Steffani
MALACOSTRACA (Decapoda)
Callichirus kraussi (Detritivore) Photograph by: C. Griffiths
Upogebia africana (Filter feeder) Photograph by: C. Griffiths
GASTROPODA
Nassarius vinctus (Scavenger)
Photograph by: N. Steffani
Venerupis corrugata (Filter feeder)

Page | 25
Temporal variation (2001-2012: Inner bay only)
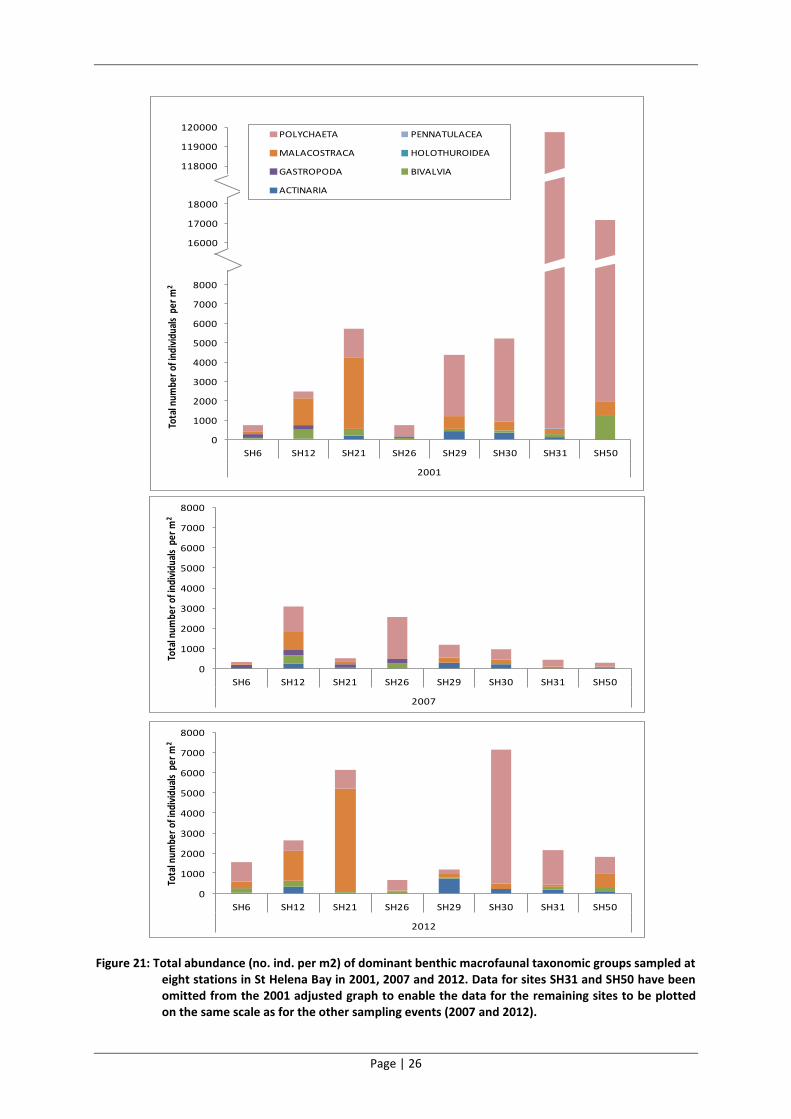
The abundance of macrofauna at the eight sites sampled in 2001, 2007 and 2012 decreased
between 2001 and 2007 and increased again in 2012 (Figure 21).
Polychaetes and malacostraca (most notably the amphipods) dominated at most of these eight sites
in all three sampling years. Furthermore, fluctuations in the abundance of polychaetes and
malacostraca accounted for most of the changes in overall abundance of benthic macrofauna at the
these sites. With the exception of Sites SH31 and SH50, the benthic macrofaunal communities
sampled at these eight sites in 2012 resemble that sampled in 2001 in terms of abundance and
taxonomic dominance.
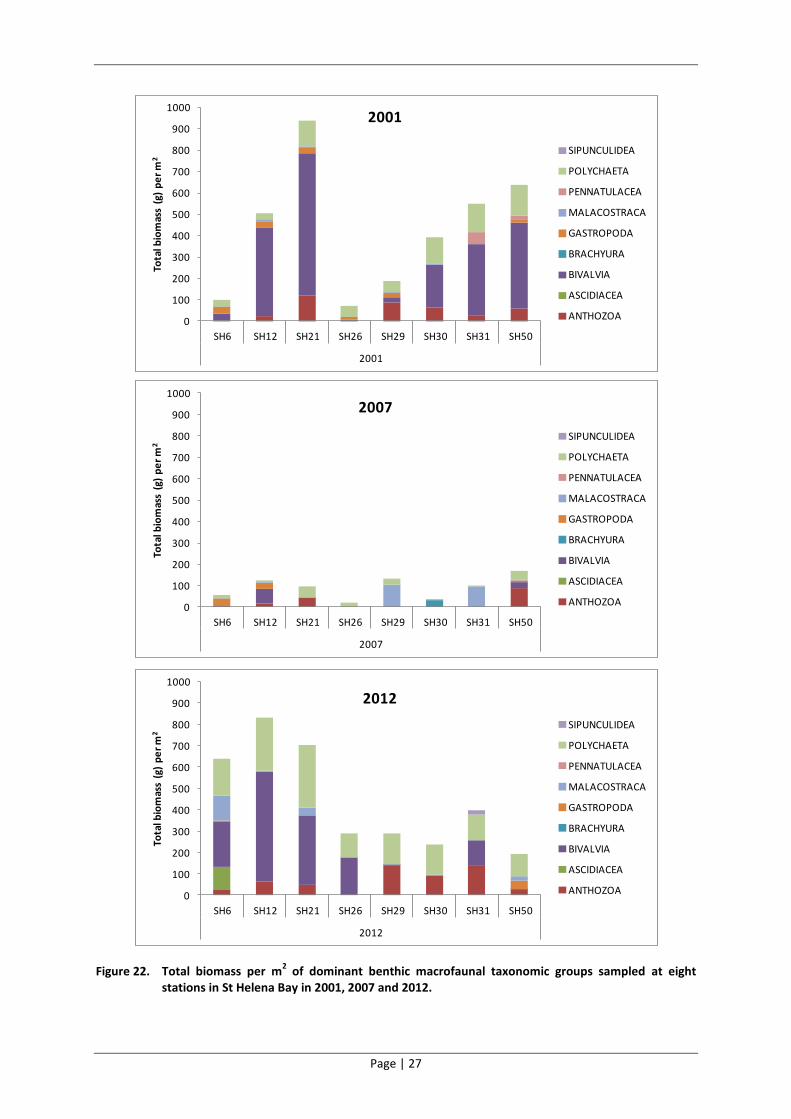
Bivalves, anthozoa (anemones) and polychaetes dominated the benthic macrofaunal communities in
terms of biomass in 2001 (Figure 22). There was a dramatic reduction in the total biomass sampled
at seven of the eight sites between 2001 and 2007 (Site SH29 was the exception). Much of this
reduction was accounted for by bivalves and polychaetes. The biomass then increased between
2007 and 2012 at all sites. Much of this reduction was accounted for by bivalves, anthozoa and
polychaetes. The benthic macrofauna sampled at these eight sites in 2012 resemble that sampled in
2001 in terms of biomass and taxonomic dominance.
Page | 26
Figure 21: Total abundance (no. ind. per m2) of dominant benthic macrofaunal taxonomic groups sampled at eight stations in St Helena Bay in 2001, 2007 and 2012. Data for sites SH31 and SH50 have been omitted from the 2001 adjusted graph to enable the data for the remaining sites to be plotted on the same scale as for the other sampling events (2007 and 2012).
0
1000
2000
3000
4000
5000
6000
7000
8000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2007
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2012
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
9000
10000
11000
12000
13000
14000
15000
16000
17000
18000
19000
20000
21000
22000
23000
24000
25000
26000
27000
28000
29000
30000
31000
32000
33000
34000
35000
36000
37000
38000
39000
40000
41000
42000
43000
44000
45000
46000
47000
48000
49000
50000
51000
52000
53000
54000
55000
56000
57000
58000
59000
60000
61000
62000
63000
64000
65000
66000
67000
68000
69000
70000
71000
72000
73000
74000
75000
76000
77000
78000
79000
80000
81000
82000
83000
84000
85000
86000
87000
88000
89000
90000
91000
92000
93000
94000
95000
96000
97000
98000
99000
100000
101000
102000
103000
104000
105000
106000
107000
108000
109000
110000
111000
112000
113000
114000
115000
116000
117000
118000
119000
120000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l num
ber
of in
divi
dual
s pe
r m
2
0
1000
2000
3000
4000
5000
6000
7000
8000
SH6
SH29
2007
Tota
l num
ber
of in
divi
dual
s pe
r m
2
POLYCHAETA PENNATULACEA
MALACOSTRACA HOLOTHUROIDEA
GASTROPODA BIVALVIA
ACTINARIA
Page | 27
Figure 22. Total biomass per m2 of dominant benthic macrofaunal taxonomic groups sampled at eight
stations in St Helena Bay in 2001, 2007 and 2012.
0
100
200
300
400
500
600
700
800
900
1000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2007
Tota
l bio
mas
s (g
) p
er
m2
SIPUNCULIDEA
POLYCHAETA
PENNATULACEA
MALACOSTRACA
GASTROPODA
BRACHYURA
BIVALVIA
ASCIDIACEA
ANTHOZOA
0
100
200
300
400
500
600
700
800
900
1000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2012
Tota
l bio
mas
s (g
) p
er
m2
SIPUNCULIDEA
POLYCHAETA
PENNATULACEA
MALACOSTRACA
GASTROPODA
BRACHYURA
BIVALVIA
ASCIDIACEA
ANTHOZOA
0
100
200
300
400
500
600
700
800
900
1000
SH6 SH12 SH21 SH26 SH29 SH30 SH31 SH50
2001
Tota
l bio
mas
s (g
) p
er
m2
SIPUNCULIDEA
POLYCHAETA
PENNATULACEA
MALACOSTRACA
GASTROPODA
BRACHYURA
BIVALVIA
ASCIDIACEA
ANTHOZOA
2001
2012
2007
Page | 28
Temporal variation (2007-2012: Inner bay, fish factories and estuary)
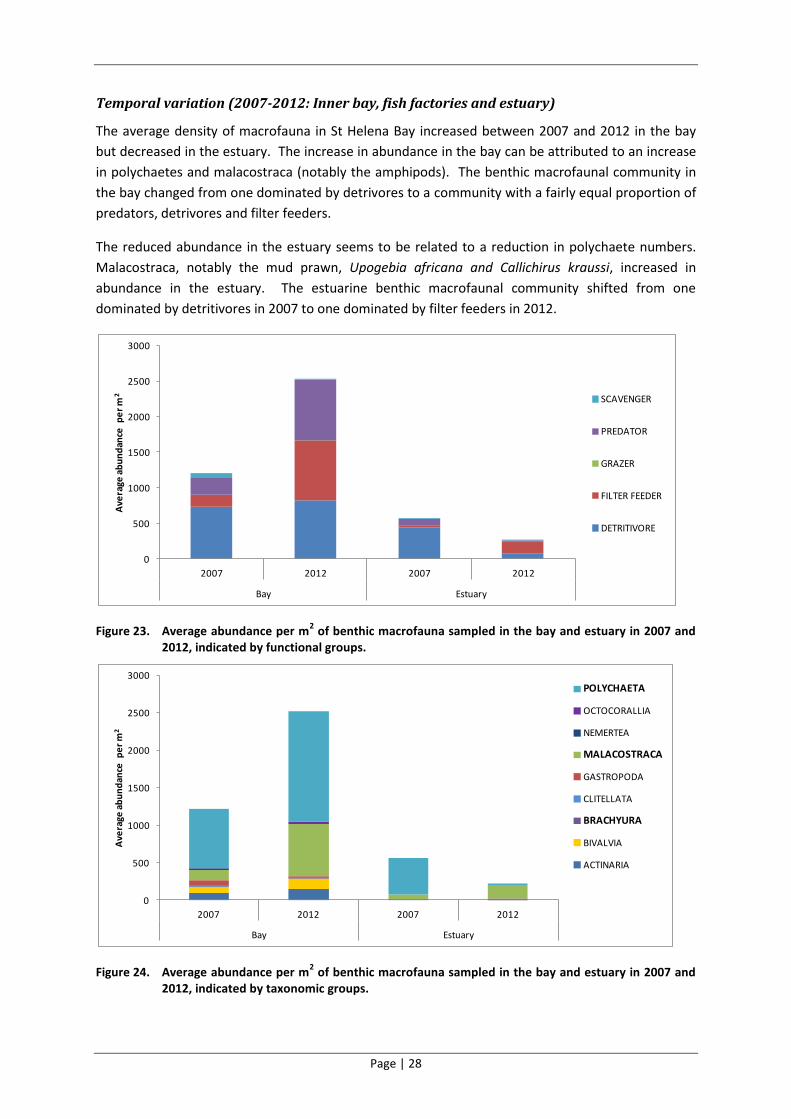
The average density of macrofauna in St Helena Bay increased between 2007 and 2012 in the bay
but decreased in the estuary. The increase in abundance in the bay can be attributed to an increase
in polychaetes and malacostraca (notably the amphipods). The benthic macrofaunal community in
the bay changed from one dominated by detrivores to a community with a fairly equal proportion of
predators, detrivores and filter feeders.
The reduced abundance in the estuary seems to be related to a reduction in polychaete numbers.
Malacostraca, notably the mud prawn, Upogebia africana and Callichirus kraussi, increased in
abundance in the estuary. The estuarine benthic macrofaunal community shifted from one
dominated by detritivores in 2007 to one dominated by filter feeders in 2012.
Figure 23. Average abundance per m2 of benthic macrofauna sampled in the bay and estuary in 2007 and
2012, indicated by functional groups.
Figure 24. Average abundance per m2 of benthic macrofauna sampled in the bay and estuary in 2007 and
2012, indicated by taxonomic groups.
0
500
1000
1500
2000
2500
3000
2007 2012 2007 2012
Bay Estuary
Ave
rage
ab
un
dan
ce p
er
m2
SCAVENGER
PREDATOR
GRAZER
FILTER FEEDER
DETRITIVORE
0
500
1000
1500
2000
2500
3000
2007 2012 2007 2012
Bay Estuary
Ave
rage
ab
un
dan
ce p
er
m2
POLYCHAETA
OCTOCORALLIA
NEMERTEA
MALACOSTRACA
GASTROPODA
CLITELLATA
BRACHYURA
BIVALVIA
ACTINARIA
Page | 29
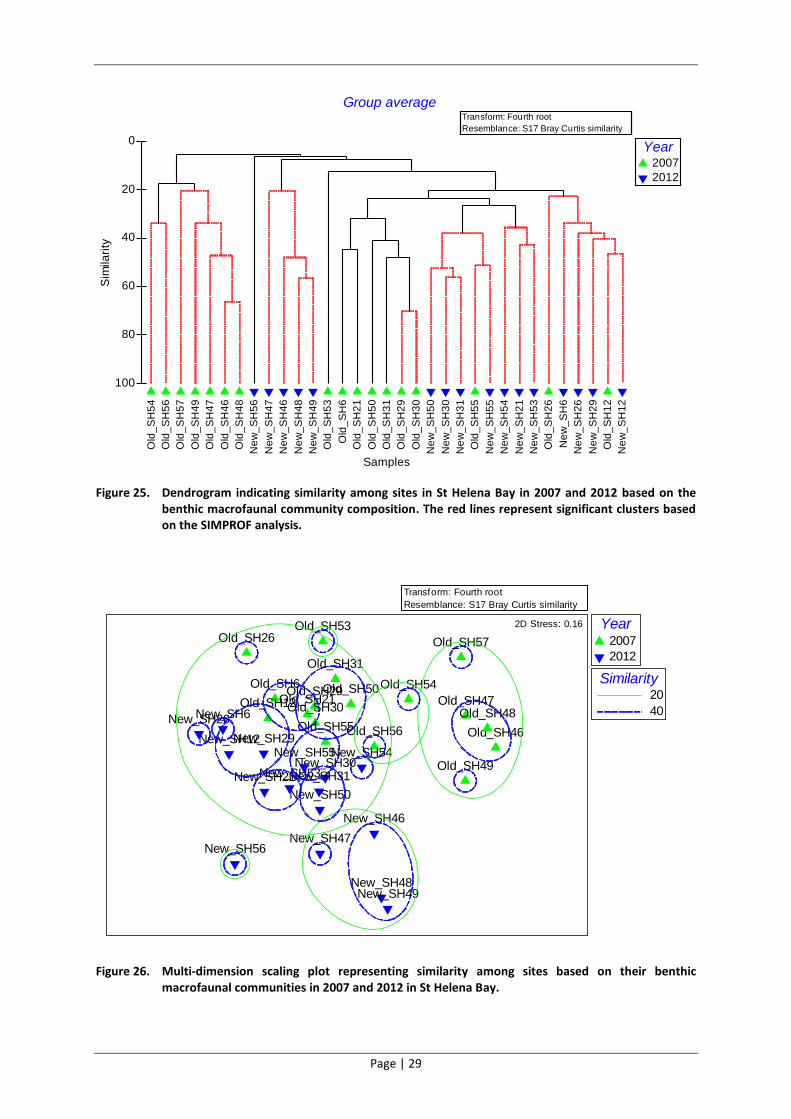
Figure 25. Dendrogram indicating similarity among sites in St Helena Bay in 2007 and 2012 based on the benthic macrofaunal community composition. The red lines represent significant clusters based on the SIMPROF analysis.
Figure 26. Multi-dimension scaling plot representing similarity among sites based on their benthic macrofaunal communities in 2007 and 2012 in St Helena Bay.
Group average
Old
_S
H5
4
Old
_S
H5
6
Old
_S
H5
7
Old
_S
H4
9
Old
_S
H4
7
Old
_S
H4
6
Old
_S
H4
8
Ne
w_
SH
56
Ne
w_
SH
47
Ne
w_
SH
46
Ne
w_
SH
48
Ne
w_
SH
49
Old
_S
H5
3
Old
_S
H6
Old
_S
H2
1
Old
_S
H5
0
Old
_S
H3
1
Old
_S
H2
9
Old
_S
H3
0
Ne
w_
SH
50
Ne
w_
SH
30
Ne
w_
SH
31
Old
_S
H5
5
Ne
w_
SH
55
Ne
w_
SH
54
Ne
w_
SH
21
Ne
w_
SH
53
Old
_S
H2
6
Ne
w_
SH
6
Ne
w_
SH
26
Ne
w_
SH
29
Old
_S
H1
2
Ne
w_
SH
12
Samples
100
80
60
40
20
0
Sim
ila
rity
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
Year2007
2012
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
Year2007
2012
Similarity20
40
Old_SH6
Old_SH12Old_SH21
Old_SH26
Old_SH29
Old_SH30
Old_SH31
Old_SH46
Old_SH47Old_SH48
Old_SH49
Old_SH50
Old_SH53
Old_SH54
Old_SH55Old_SH56
Old_SH57
New_SH6
New_SH12
New_SH21
New_SH26
New_SH29
New_SH30New_SH31
New_SH46
New_SH47
New_SH48New_SH49
New_SH50
New_SH53
New_SH54New_SH55
New_SH56
2D Stress: 0.16
Page | 30
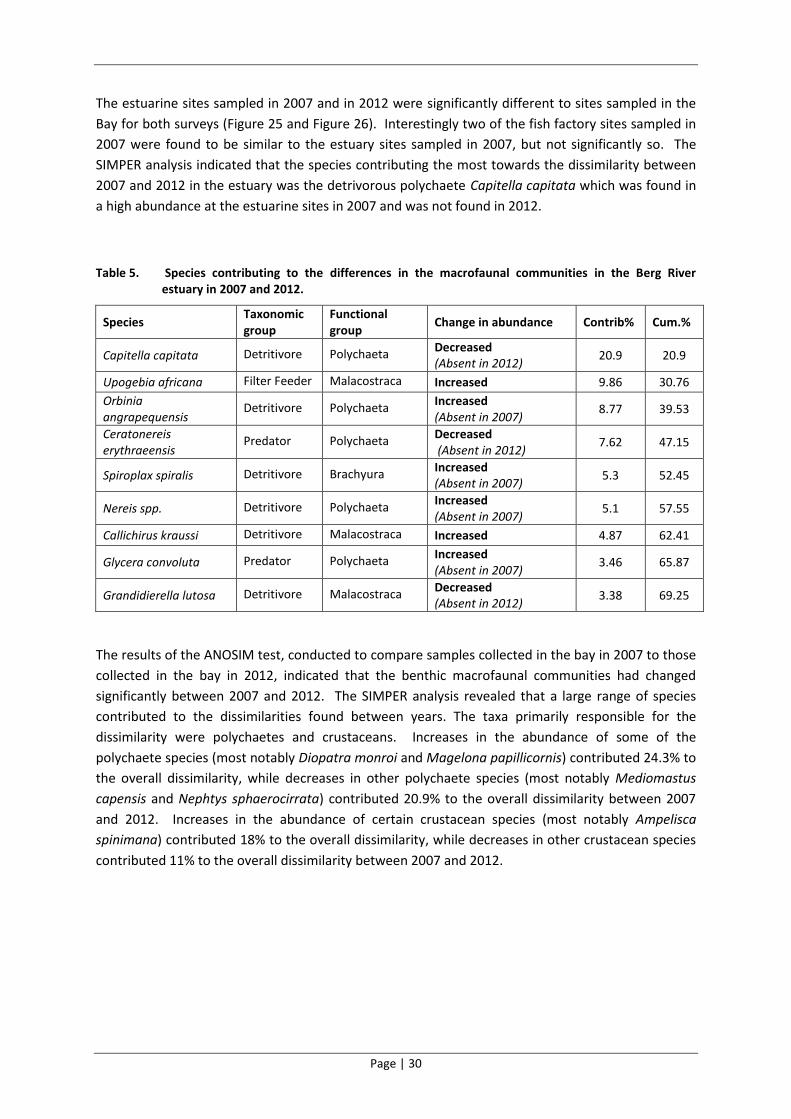
The estuarine sites sampled in 2007 and in 2012 were significantly different to sites sampled in the
Bay for both surveys (Figure 25 and Figure 26). Interestingly two of the fish factory sites sampled in
2007 were found to be similar to the estuary sites sampled in 2007, but not significantly so. The
SIMPER analysis indicated that the species contributing the most towards the dissimilarity between
2007 and 2012 in the estuary was the detrivorous polychaete Capitella capitata which was found in
a high abundance at the estuarine sites in 2007 and was not found in 2012.
Table 5. Species contributing to the differences in the macrofaunal communities in the Berg River estuary in 2007 and 2012.
Species Taxonomic group
Functional group
Change in abundance Contrib% Cum.%
Capitella capitata Detritivore Polychaeta Decreased (Absent in 2012)
20.9 20.9
Upogebia africana Filter Feeder Malacostraca Increased 9.86 30.76
Orbinia angrapequensis
Detritivore Polychaeta Increased (Absent in 2007)
8.77 39.53
Ceratonereis erythraeensis
Predator Polychaeta Decreased (Absent in 2012)
7.62 47.15
Spiroplax spiralis Detritivore Brachyura Increased (Absent in 2007)
5.3 52.45
Nereis spp. Detritivore Polychaeta Increased (Absent in 2007)
5.1 57.55
Callichirus kraussi Detritivore Malacostraca Increased 4.87 62.41
Glycera convoluta Predator Polychaeta Increased (Absent in 2007)
3.46 65.87
Grandidierella lutosa Detritivore Malacostraca Decreased (Absent in 2012)
3.38 69.25
The results of the ANOSIM test, conducted to compare samples collected in the bay in 2007 to those
collected in the bay in 2012, indicated that the benthic macrofaunal communities had changed
significantly between 2007 and 2012. The SIMPER analysis revealed that a large range of species
contributed to the dissimilarities found between years. The taxa primarily responsible for the
dissimilarity were polychaetes and crustaceans. Increases in the abundance of some of the
polychaete species (most notably Diopatra monroi and Magelona papillicornis) contributed 24.3% to
the overall dissimilarity, while decreases in other polychaete species (most notably Mediomastus
capensis and Nephtys sphaerocirrata) contributed 20.9% to the overall dissimilarity between 2007
and 2012. Increases in the abundance of certain crustacean species (most notably Ampelisca
spinimana) contributed 18% to the overall dissimilarity, while decreases in other crustacean species
contributed 11% to the overall dissimilarity between 2007 and 2012.
Page | 31
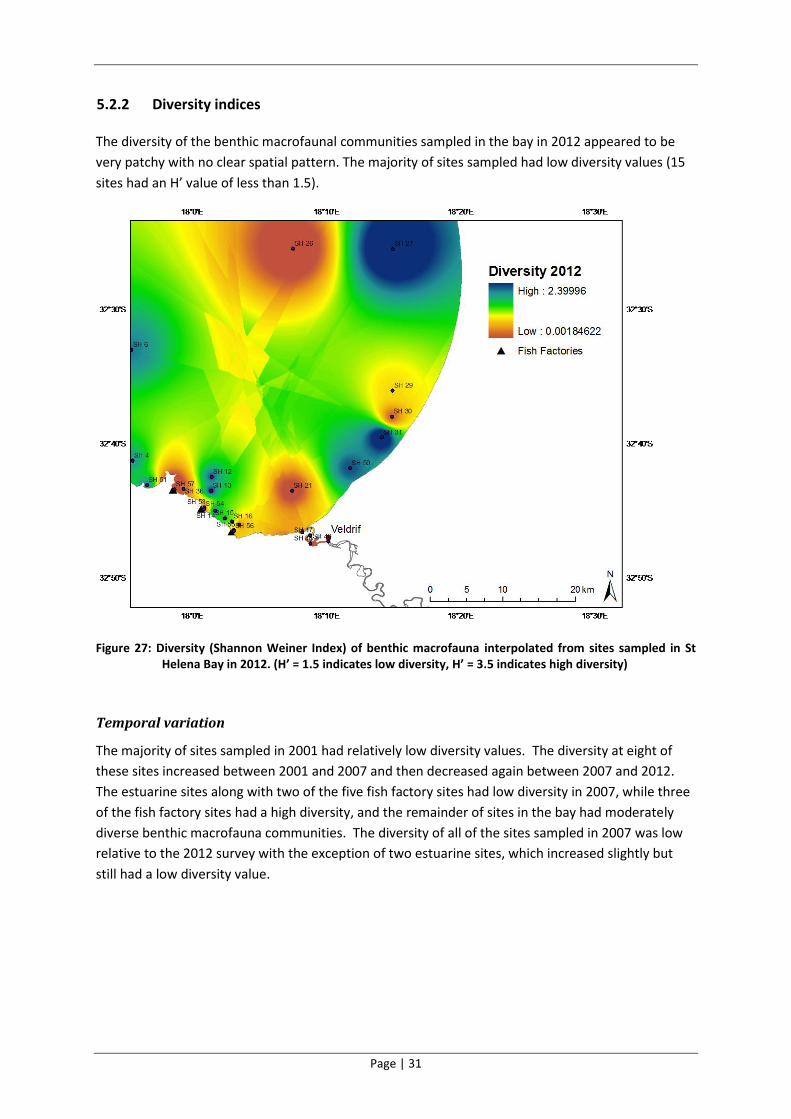
5.2.2 Diversity indices
The diversity of the benthic macrofaunal communities sampled in the bay in 2012 appeared to be
very patchy with no clear spatial pattern. The majority of sites sampled had low diversity values (15
sites had an H’ value of less than 1.5).
Figure 27: Diversity (Shannon Weiner Index) of benthic macrofauna interpolated from sites sampled in St Helena Bay in 2012. (H’ = 1.5 indicates low diversity, H’ = 3.5 indicates high diversity)
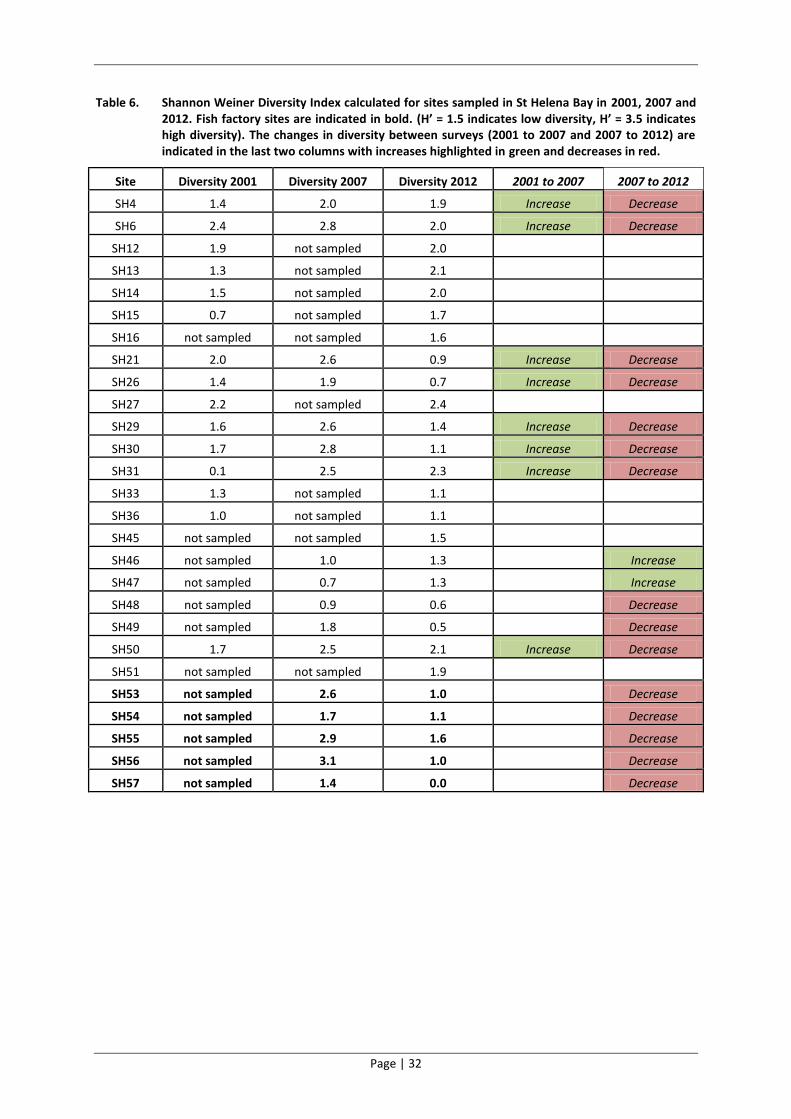
Temporal variation
The majority of sites sampled in 2001 had relatively low diversity values. The diversity at eight of
these sites increased between 2001 and 2007 and then decreased again between 2007 and 2012.
The estuarine sites along with two of the five fish factory sites had low diversity in 2007, while three
of the fish factory sites had a high diversity, and the remainder of sites in the bay had moderately
diverse benthic macrofauna communities. The diversity of all of the sites sampled in 2007 was low
relative to the 2012 survey with the exception of two estuarine sites, which increased slightly but
still had a low diversity value.
Page | 32
Table 6. Shannon Weiner Diversity Index calculated for sites sampled in St Helena Bay in 2001, 2007 and 2012. Fish factory sites are indicated in bold. (H’ = 1.5 indicates low diversity, H’ = 3.5 indicates high diversity). The changes in diversity between surveys (2001 to 2007 and 2007 to 2012) are indicated in the last two columns with increases highlighted in green and decreases in red.
Site Diversity 2001 Diversity 2007 Diversity 2012 2001 to 2007 2007 to 2012
SH4 1.4 2.0 1.9 Increase Decrease
SH6 2.4 2.8 2.0 Increase Decrease
SH12 1.9 not sampled 2.0
SH13 1.3 not sampled 2.1
SH14 1.5 not sampled 2.0
SH15 0.7 not sampled 1.7
SH16 not sampled not sampled 1.6
SH21 2.0 2.6 0.9 Increase Decrease
SH26 1.4 1.9 0.7 Increase Decrease
SH27 2.2 not sampled 2.4
SH29 1.6 2.6 1.4 Increase Decrease
SH30 1.7 2.8 1.1 Increase Decrease
SH31 0.1 2.5 2.3 Increase Decrease
SH33 1.3 not sampled 1.1
SH36 1.0 not sampled 1.1
SH45 not sampled not sampled 1.5
SH46 not sampled 1.0 1.3 Increase
SH47 not sampled 0.7 1.3 Increase
SH48 not sampled 0.9 0.6 Decrease
SH49 not sampled 1.8 0.5 Decrease
SH50 1.7 2.5 2.1 Increase Decrease
SH51 not sampled not sampled 1.9
SH53 not sampled 2.6 1.0 Decrease
SH54 not sampled 1.7 1.1 Decrease
SH55 not sampled 2.9 1.6 Decrease
SH56 not sampled 3.1 1.0 Decrease
SH57 not sampled 1.4 0.0 Decrease

Page | 33
6 INTEGRATION OF PHYSICO-CHEMICAL PARAMETERS AND BIOTIC
INDICATORS
6.1 Methods
The aim of this analysis was to determine how the environmental variables (organic content of
sediment, grain size) relate to the observed biological patterns in macrobenthic community
structure. This involved superimposing the concentrations of individual environmental variables
onto biotic multi-dimensional scaling (MDS) plots. An MDS plot is a spatial representation of the
Bray Curtis similarity between sites. MDS plots are constructed using PRIMER V6 from the similarity
matrix in order to graphically view similarities between sample sites. Like the dendrogram, samples
with similar species composition and abundance cluster together, while those that are less similar
are placed further apart. The values of various environmental variables are superimposed on MDS
plots as circles of varying diameter (the larger the circle, the higher the concentration). These are
known as ‘bubble plots’ and they allow one to easily identify the sites at which contaminants are
elevated, as well as to determine if contamination patterns have any correlation to biotic structure.
6.2 Results
The bubble plot based on the 2012 survey results, which incorporated depth values, revealed that
the differences between the benthic macrofaunal communities in the estuary and the bay were
related to some extent to depth in that all estuarine sites were shallower than the Bay. This is to
some extent an artefact of the fact that all sampling sites in the bay were deeper than 4 m and that
all of the estuary sites were 1 m depth or less. It may well be that the difference is actually related
to difference in salinity or some other parameter not measured in this study. The bubble plots with
mud, TOC and TON revealed no clear trend.
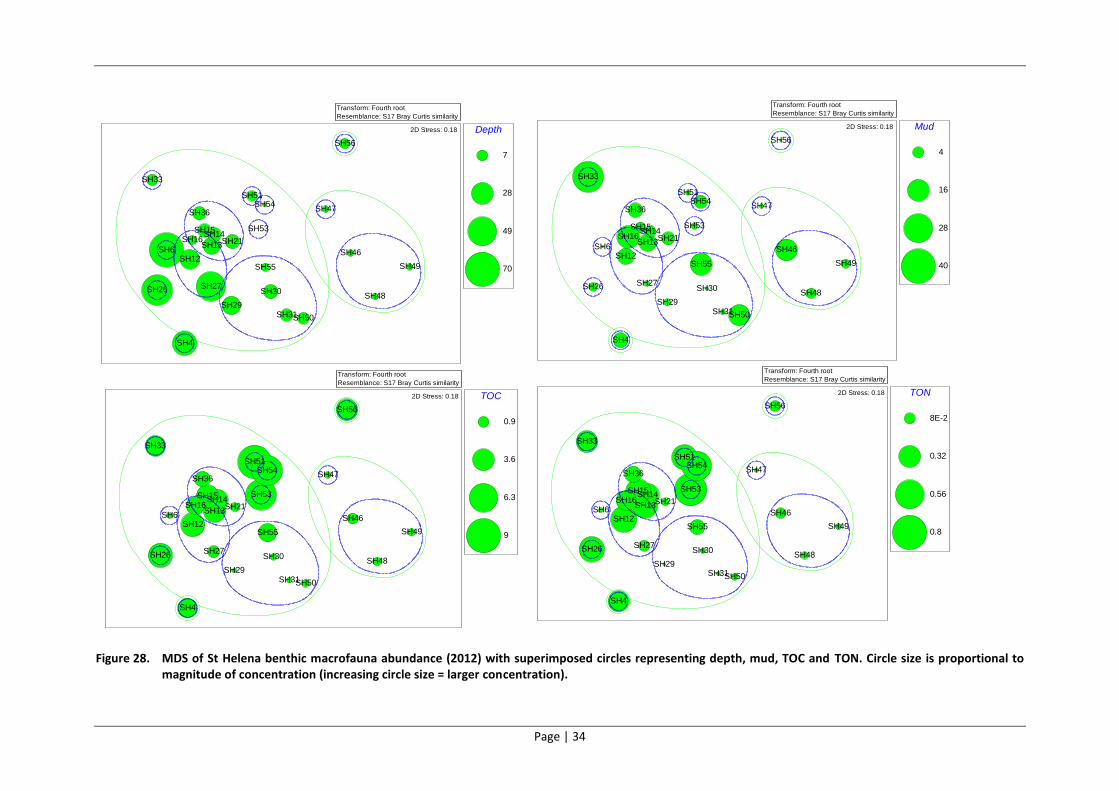
Page | 34
Figure 28. MDS of St Helena benthic macrofauna abundance (2012) with superimposed circles representing depth, mud, TOC and TON. Circle size is proportional to magnitude of concentration (increasing circle size = larger concentration).
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
TON
8E-2
0.32
0.56
0.8
SH4
SH6SH12
SH13
SH14SH15SH16 SH21
SH26 SH27
SH29
SH30
SH31
SH33
SH36
SH46
SH47
SH48
SH49
SH50
SH51
SH53
SH54
SH55
SH56
2D Stress: 0.18
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
Depth
7
28
49
70
SH4
SH6SH12
SH13
SH14SH15
SH16 SH21
SH26 SH27
SH29
SH30
SH31
SH33
SH36
SH46
SH47
SH48
SH49
SH50
SH51
SH53
SH54
SH55
SH56
2D Stress: 0.18
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
Mud
4
16
28
40
SH4
SH6SH12
SH13
SH14SH15
SH16 SH21
SH26 SH27
SH29
SH30
SH31
SH33
SH36
SH46
SH47
SH48
SH49
SH50
SH51
SH53
SH54
SH55
SH56
2D Stress: 0.18
Transform: Fourth root
Resemblance: S17 Bray Curtis similarity
TOC
0.9
3.6
6.3
9
SH4
SH6SH12
SH13
SH14SH15SH16 SH21
SH26 SH27
SH29
SH30
SH31
SH33
SH36
SH46
SH47
SH48
SH49
SH50
SH51
SH53
SH54
SH55
SH56
2D Stress: 0.18

Page | 35
7 DISCUSSION
The monitoring protocol used in the 2012 survey and previous surveys provides snapshots of the
state of physico-chemical environment and the benthic ecology in St Helena Bay since 2001. The
positioning of sites allows for discussion on the state of the bay from three different perspectives:
the state of the benthic ecosystem in the bay as a whole, the intensity and extent of impacts from
point source fish factory effluents, and the state of the lower portion of the Berg River Estuary. The
results of the 2012 survey are discussed in the context of these three different perspectives below.
7.1 Bay-wide variability
Sediments
Particle size composition of the sediments in the marine environment is influenced by the wave
energy, depth and current circulation patterns. Coarser or heavier sand and gravel particles are
found in areas with high wave energy and strong currents as the movement of water in these areas
suspends fine particles (mud and silt) and flushes these out of the area. Alterations to the wave
action and current patterns which reduce or increase the movement of water can result in the
deposition (reduced water movement) or removal (increased water movement) of mud from a
particular area. The 2012 results indicated a dramatic reduction in the mud content of the
sediments in St Helena Bay since 2007 and 2001. This is possibly the result of an increasing intensity
and frequency of large storm events, given that the effects have been experienced at a bay-wide
scale. Indeed, examinations of storm events over the period 1996 to 2010 by Theron et al. (2010)
revealed an increasing trend in the peaks of individual storm events during the stormy winter season
offshore of Cape Town.
St Helena Bay is situated downstream of the Cape Columbine upwelling cell. Nutrients are
transported equatorward from Cape Columbine into St Helena Bay and retained within a clockwise
circulation in the bay (Hutchings et al. 2012). Sun-warming of the surface layers during summer and
autumn results in the stratification of the water column which allows for phytoplankton blooms to
develop as the cells are kept in the euphotic zone and not mixed deeper within the water column
(Bailey 1991). St Helena Bay is an extremely rich zone for plankton due to the enhanced nutrient
availability and a stable euphotic zone, and the bay provides a rich feeding ground for planktivores
and in turn, predators. At a large scale, the mostly likely source of organic matter in St Helena Bay is
from phytoplankton production, the associated detritus that forms from the decay thereof and
faecal pellets. Organic matter enrichment from fish factory effluent is evident at a local scale and
does not appear to impact the bay as a whole (results of 2012 survey, Carter and Steffani 2007).
The results of the 2012 survey indicated that there had been minor reductions in the levels of
organic nitrogen at most sites throughout the bay since 2007, while organic carbon levels decreased
in the northern reaches of the bay, increased in the vicinity of the fish factories and underwent
minor changes elsewhere in the bay. Interestingly the molar carbon:nitrogen ratios increased
dramatically throughout the bay between 2007 and 2012. An increase in the rate of denitrification is
likely to have resulted in the reduction of particulate nitrogen in the sediments and the consequent
increase in the C:N ratios experienced in St Helena Bay since 2012.

Page | 36
Denitrification is the process whereby nitrates (organic nitrogen) are reduced by microbes to form
nitrogen and ammonia. This process occurs in environments where oxygen levels have been
depleted (anoxic or hypoxic) and nitrates are present. In areas where photosynthetic rates are very
high, such as in upwelling systems, a high biological oxygen demand deeper in the water column and
sediments can lead to complete oxygen utilisation. The highly nutrient enriched, stratified and
productive waters of St Helena Bay are conducive to the formation of oxygen deficient conditions
(Bailey 1991) and low oxygen events are common (Chapman and Shannon 1985). Denitrifying
bacteria are likely to dominate in the area when there is an oxygen deficiency as they are able to
substitute the oxygen, required for organic matter degradation, through nitrate reduction and by so
doing, become the most efficient energy extractors (Knowles 1982, Tyrrell and Lucas 2002).
The increased rate of denitrification since 2007 may be a result of increased plankton productivity in
the bay since 2007 and the consequent reduction in oxygen availability in the sediments. Hutchings
et al. (2012) reported decadal scale fluctuations in upwelling-favourable winds observed over the
period 1982 to 2006, with winds reaching a peak in 2001 and subsequently undergoing a decline to
2006. El Niño events have at times been known to suppress upwelling along the coast, while La Niña
increases upwelling intensity (Arntz et al. 2006, Rouault et al. 2010). The 2012 survey was
conducted following several years in which La Niña events dominated while the 2007 survey was
conducted following several years in which El Niño events dominated. This could be an indication
that upwelling-favourable conditions and therefore plankton productivity were higher in years
preceding the 2012 survey compared to years preceding the 2007 survey. However it is important
to note that not all events are experienced by the Benguela system as they are in the Pacific systems
(Arntz et al. 2006).
Figure 29. Time series of the Southern Oscillation Index, indicating El Niño and La Niña events since 1996 and highlighting years in which St Helena Bay has been surveyed. Note the El Niño event of 1997/1998 was not experienced in the Benguela system. (Data source: Arntz et al. 2006 and http://www.bom.gov.au/climate/glossary/soi.shtml).
-30
-20
-10
0
10
20
30
19
96
19
97
19
98
19
99
20
00
20
01
20
02
20
03
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
12
Sou
ther
n O
scill
atio
n In
dex
La Niña
El Niño

Page | 37
Benthic macrofauna
Changes in benthic species composition can be the first indicator of disturbance or habitat
alteration, as certain species are more sensitive (i.e. likely to decrease in abundance in response to
stress) while others are more tolerant of adverse conditions (and may increase in abundance in
response to stress, taking up space or resources vacated by the more sensitive species). Certain
species are able to rapidly invade and colonise disturbed areas as they have high fecundity, rapid
growth and rather short life-cycles (Newell 1998). These species are known as “r-strategists” or
opportunistic species and their presence generally indicates unpredictable short-term variations in
environmental conditions which may result from either natural factors or anthropogenic activities.
In stable environments the community composition is controlled predominantly by biological
interactions rather than by fluctuations in environmental conditions. Species found in these
conditions are known as “K-strategists” and are selected for their competitive ability. K-strategists
are characterised by long-life spans, larger body sizes, delayed reproduction and low death rates.
Intermediate communities with different relative proportions of opportunistic species and K-
strategists are likely to exist between the extremes of stable and unstable environments.
Eastern Boundary Current ecosystems, such as the BCS are generally known to be dominated by
opportunistic benthic species with short life spans and relatively high reproductive rates (r-
strategists), which are adapted to surviving in oxygen deficient conditions (Arntz et al. 2006). Such
opportunistic species should recover rapidly following natural or anthropogenic disturbances. The
2012 survey revealed that the benthic macrofaunal community in St Helena Bay is dominated by the
predatory polychaete Diopatra monroi both in terms of abundance and biomass. The filter feeding
amphipod Ampelisca anomala, also dominated in terms of abundance and the clam, Venerupis
corrugata, in terms of biomass. These species are typical of the continental shelf sediments of the
west coast (Christie 1976, Carter and Steffani 2006, Carter and Steffani 2007). The dominance of
amphipods suggests that the system has a fairly low level of contamination as amphipods have a
lower tolerance than other groups to pollution, particularly organic pollution (Christie and Moldan
1977).
Polychaetes and amphipods, which have dominated the benthic macrofauna in terms of abundance
since 2001, decreased between 2001 and 2007 and then increased between 2007 and 2012.
Similarly bivalves and polychaetes, which have dominated in terms of biomass since 2001,
underwent a reduction between 2001 and 2007 and then an increase between 2007 and 2012.
Diversity values increased between 2001 and 2007 and then decreased between 2007 and 2012.
This suggests that the benthic communities were subjected to a bay-wide variation or disturbance,
potentially of a repetitive nature, and that at lower densities the community is able to support
greater species diversity. As suggested above for the changes in the C:N ratios, the productivity of
phytoplankton and the consequent levels of oxygen availability may have varied in the bay, possibly
due to decadal scale fluctuations in upwelling-favourable winds. The change detected in C:N ratios
between 2007 and 2012 and the known occurrence of denitrification in the BCS, suggests that
oxygen availability in the sediments was higher in 2007 than in 2012. This is likely to have had
consequences for the benthic macrofauna community structure. It may have favoured
environmental conditions that enabled species to share habitat and outcompete those opportunistic
species usually dominating in oxygen deficient conditions.

Page | 38
The sea pen (Virgularia schultzei) is typical of the west coast, but unlike other west coast species, is
relatively slow growing and is thus more sensitive to variation. The sea pen was present at two sites
in 2001 at relatively low densities and was not found again in 2007 and 2012. This suggests that the
benthic ecosystem underwent a disturbance to the state that is usually suited to sea pens.
The results of 2012 survey further emphasized the high natural variability of benthic soft-bottom
communities. The benthic communities in the bay were not spatially grouped and did not clearly
correlate with any of the environmental variables measured (depth, TOC, TON, mud content). It is
likely that the communities in the bay are influenced by complex biological interactions at the
sediment-water interface, various physico-chemical properties and potentially an interplay between
those properties and biological interactions.
7.2 Fish factory sites
Given the known impacts of fish factory discharges on the marine environment (Christie and Moldan
1977) the areas around the fish factory discharge point were prioritised as key sites for monitoring
purposes in the 2007 and 2012 surveys. Results of the 2012 survey revealed that the areas in the
vicinity of the fish factory outfalls were carbon enriched compared to other areas in the bay and that
the levels of enrichment at these sites had increased disproportionately compared to other sites in
the bay. This suggests that the effluent from fish factories is having a marked effect on the organic
composition of the sediments at a local scale (sites within 50m of fish factory outfalls) but not at a
large scale.
The overlapped effects of anthropogenic and natural disturbances on benthic communities present a
challenge in environmental management and in some cases the health of an ecosystem becomes
difficult to evaluate when natural disturbances mask the response of potential ecological indicators.
For this reason it is important to assess the data from specific sites in the context of the larger
system. However, all of the sites sampled adjacent to the fish factory effluent pipes were found to
be significantly different from one another. Two sites sampled in the immediate vicinity of the fish
factory discharge points were in fact outliers and were characterised by depauperate benthic
macrofaunal communities. No benthic macrofauna were found in the sample taken from SH57 and
only three polychaete species, with very low abundances were sampled at SH 56. Given that the
macrofauna on the west coast are typically opportunistic species, able to recover relatively rapidly
from disturbances and that the communities at site SH56 and SH57 were in such a poor state, it is
evident that effluents from these fish factories are having a negative impact upon the benthic
macrofaunal community at these sites. However, as the results of the 2007 survey indicated, these
impacts are limited to the local area around the fish factories discharge points.
7.3 Estuarine sites
The levels of carbon and nitrogen enrichment in the estuary remained relatively low between 2007
and 2012, however the C:N rations increased substantially, possibly as a combined result of
terrigenous nutrient inputs and nutrient inputs from the marine environment. The benthic

Page | 39
macrofaunal communities sampled in the Berg River Estuary were significantly different to those
sampled in the Bay and were characterised by the presence of the estuarine mud prawn (Upogebia
africana), two detritivorous polychaetes species (Orbinia angrapequensis and Nereis spp.) and the
three-legged crab (Spiroplax spiralis). Changes since the 2007 survey include a reduction in the
overall abundance of macrofaunal abundance, particularly polychaetes, an increase in prawn
abundance and a shift in community structure from a system dominated by detritivores to one
dominated by filter feeders.

Page | 40
8 CONCLUSIONS
Both sediment results (C:N ratios) and benthic community results indicate that bay-wide
fluctuations are occurring. These are possibly linked to decadal-scale variations in upwelling
intensity and phytoplankton productivity;
Monitoring in the larger bay area must continue to aid in the interpretation of variability
detected at local scales;
The levels of organic contamination have increased at two of the sites adjacent to fish
factories (fish factory in Stompneusbaai and the most southerly fish factory) as well as in
Britannia Bay since 2007;
The benthic macrofauna sampled at the two heavily contaminated fish factory sites provide
evidence of a negative impact to the marine ecology at a local scale; and
There is evidence of organic enrichment at the site situated in Britannia Bay. Sources of this
enrichment should be investigated.

Page | 41
9 REFERENCES
Arntz, W.E., Gallardo, V.A., Gutierrrez, D., Isla, E., Levin, L.A., Mendo, J., Neira, C., Rowe, G.T.,
Tarazona, J. and M. Wolff. 2006. El Nino and similar perturbation effects on the benthos of
the Humboldt, California, and Benguela Current upwelling ecosystems. Advances in
Geosciences, 6, 243-265.
Bailey, G.W. 1991. Organic carbon flux and development of oxygen deficiency on the modern
Benguela continental shelf south of 22°S: spatial and temporal variability. Modern and
Ancient Continental Shelf Anoxia. Geological Society Special Publication No. 58, 171-183
Bakun, A. 1998. Ocean triads and radical interdecadal stock variability: bane and boon for fishery
management science. In: Pitcher, T.J., Hart, P.J.B., Pauly, D. (Eds.), Reinventing Fisheries
Management. Kluwer Academic Publishers, Dordrecht, Netherlands, pp. 331–358.Bakun
1998,
Branch, G. M., & Branch, M. 1981. The Living Shores of Southern Africa. C. Struik, Cape Town.
Carter, R. and N. Steffani. 2007. State of Saint Helena Bay 2007 Benthic Macrofauna Distributions.
Prepred for CSIR.
Chapman, P. & L.V. Shannon. 1985. The Benguela Ecosystem. 2 Chemistry and related processes.
Oceanography and Marine Biology Annual Review 23, 183-251.
Christie, N.D. 1976. A numerical analysis of the distribution of a shallow sublittoral sand macrofauna
along a transect at Lamberts Bay, South Africa. Trans. R. Soc. S. Afr. 42: 149-Christie 1976
Christie, N.D. & A. Moldan. 1977. Effects of Fish Factory Effluent on the Benthic Macrofauna of
Saldanha Bay. Marine Pollution Bulletin, 8, 2, 41-45.
Cockcroft, A.C., Schoeman, D.S., Pitcher, G.C., Bailey, G.W. & D.C. Van Zyl 2000 -A mass stranding, or
"walkout" of west coast rock lobster Jasus lalandii in Elands Bay, South Africa: causes, results
and implications. In The Biodiversity Crises and Crustacea.Von Kaupel Klein, J.C. and F.R.
Schram (Eds). Crustacean Iss. 11: 362-688.
CSIR 2008. St Helena Bay Water Quality Trust: The Biogeochemical status of surface sediments in St
Helena Bay in 2007. Prepared for the St Helena Bay Water Quality Trust
Hutchings, L., Jarre, A., Lamont, T. & M. Van den Berg. In Press. St Helena Bay, then and now: muted
climate signals, large human impact. African Journal of Marine Science.
Knowles, R. 1982. Denitrification. Microbiological Reviews 46 (1), 43-70
Monteiro, P.M.S. & A.M. Roychoudhury. 2005. Spatial Characteristics of sediment trace metals in an
eastern boundary upwelling retention area (St. Helena Bay, South Africa): A hydrodynamic-
biological pump hypothesis. Estuarine, Coastal and Shelf Science, 65, 123-134.
Rouault, M., Pohl, B. & P. Penven. 2010. Coastal oceanic climate change and variability from 1982 to
2009 around South Africa. African Journal of Marine Science.32(2):237-246
Shannon, L.V. & M.J. O’Toole, 1998. BCLME Thematic Report 2: Integrated overview of the
oceanography and environmental variability of the Benguela Current region. Unpublished BCLME
Report, 58pp
Theron, A. , M.Rossouw , L. Barwell , A.Maherry , G.Diedericks & P. de Wet. 2010. Quantification of
risks to coastal areas and development: wave run-up and erosion. NE20-PA-F
Tyrrell, T. & M. Lucas. 2002. Geochemical evidence of denitrification in the Benguela upwelling
system. Continental Shelf Research, 22, 2497-2511
Warwick, R.M. 1993. Environmental impact studies on marine communities: Pragmatical
considerations. Australian Journal of Ecology 18: 63-80.
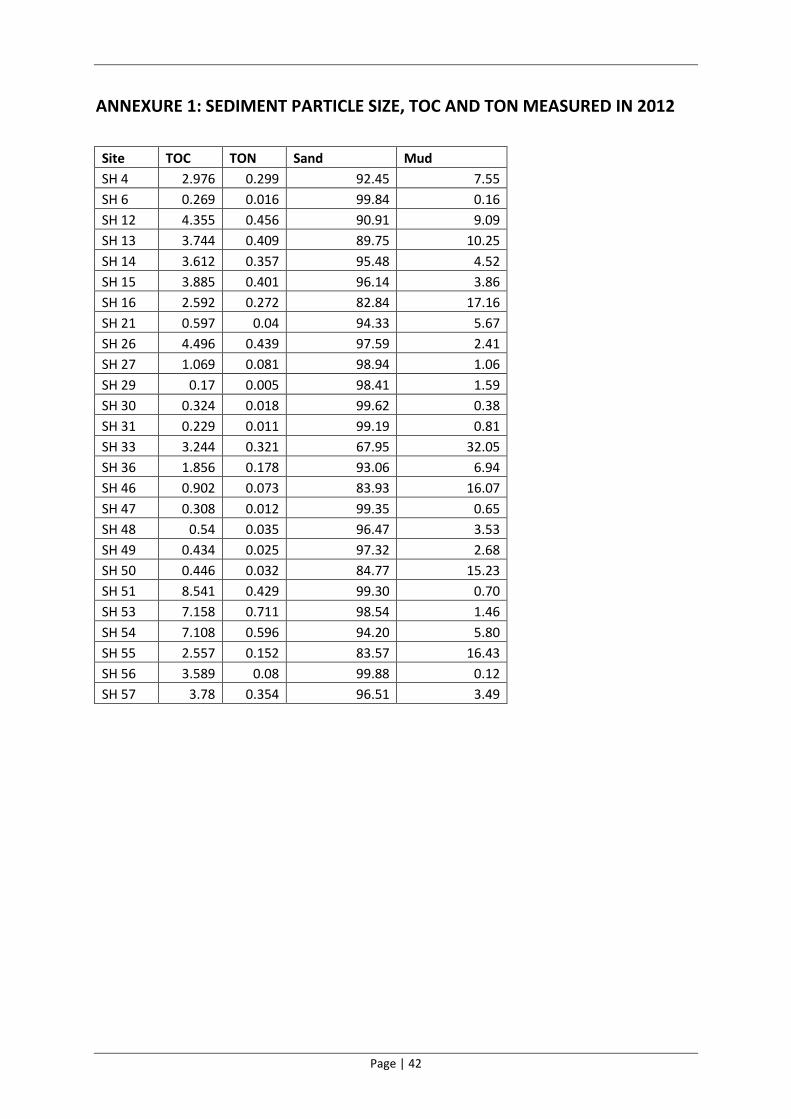
Page | 42
ANNEXURE 1: SEDIMENT PARTICLE SIZE, TOC AND TON MEASURED IN 2012
Site TOC TON Sand Mud
SH 4 2.976 0.299 92.45 7.55
SH 6 0.269 0.016 99.84 0.16
SH 12 4.355 0.456 90.91 9.09
SH 13 3.744 0.409 89.75 10.25
SH 14 3.612 0.357 95.48 4.52
SH 15 3.885 0.401 96.14 3.86
SH 16 2.592 0.272 82.84 17.16
SH 21 0.597 0.04 94.33 5.67
SH 26 4.496 0.439 97.59 2.41
SH 27 1.069 0.081 98.94 1.06
SH 29 0.17 0.005 98.41 1.59
SH 30 0.324 0.018 99.62 0.38
SH 31 0.229 0.011 99.19 0.81
SH 33 3.244 0.321 67.95 32.05
SH 36 1.856 0.178 93.06 6.94
SH 46 0.902 0.073 83.93 16.07
SH 47 0.308 0.012 99.35 0.65
SH 48 0.54 0.035 96.47 3.53
SH 49 0.434 0.025 97.32 2.68
SH 50 0.446 0.032 84.77 15.23
SH 51 8.541 0.429 99.30 0.70
SH 53 7.158 0.711 98.54 1.46
SH 54 7.108 0.596 94.20 5.80
SH 55 2.557 0.152 83.57 16.43
SH 56 3.589 0.08 99.88 0.12
SH 57 3.78 0.354 96.51 3.49

Page | 43
ANNEXURE 2: BENTHIC MACROFAUNA SAMPLED IN 2012 (abundance per m2)
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Afrophaxas decipiens 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 6.9 0.0 0.0
Amaryllis macrophthalma 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca anomola 0.0 0.0 0.0 2571.2 4225.2 472.6 0.0 4774.1 0.0 0.0 0.0 27.8 0.0 708.8 8394.7 139.0 0.0 20.8 0.0 0.0 6.9 0.0 34.7 0.0 0.0 0.0 0.0
Ampelisca brevicornis 0.0 69.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca palmata 0.0 20.8 0.0 2800.6 708.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 48.6 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca spinimana 0.0 0.0 840.9 0.0 0.0 0.0 173.7 0.0 13.9 236.3 6.9 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 6.9 0.0 0.0 952.1 0.0 0.0
Amphieteis gunneri 0.0 0.0 0.0 62.5 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Amphilochus neapolitanus 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Anemone 0.0 13.9 312.7 229.3 132.0 104.2 118.1 13.9 0.0 69.5 722.7 145.9 111.2 0.0 97.3 132.0 6.9 0.0 0.0 0.0 0.0 132.0 173.7 27.8 76.4 0.0 0.0
Anemone 2 6.9 0.0 0.0 6.9 0.0 0.0 0.0 13.9 0.0 0.0 27.8 69.5 83.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 62.5 0.0 0.0 0.0 0.0 0.0 0.0
Anemone 3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Anthelura remipes 6.9 0.0 0.0 6.9 55.6 20.8 0.0 48.6 0.0 0.0 208.5 104.2 83.4 0.0 34.7 41.7 0.0 0.0 6.9 0.0 48.6 0.0 0.0 0.0 6.9 0.0 0.0
Arenicola loveni 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 250.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0
Ascidian 0.0 20.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Bathyporeia sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0
Bullia laevissima 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0
Callichirus kraussi 0.0 0.0 0.0 6.9 6.9 41.7 118.1 104.2 0.0 0.0 0.0 6.9 0.0 0.0 0.0 13.9 0.0 166.8 0.0 0.0 0.0 0.0 34.7 0.0 6.9 0.0 0.0
Calyptraea chinensis 0.0 0.0 0.0 465.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Capitella capitata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 514.2 0.0 0.0 0.0
Caprellid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Caprellid 2 0.0 0.0 0.0 0.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0

Page | 44
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Choromytilus meridionalis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 20.8 0.0 0.0 0.0 0.0
Cirolana hirtipes 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Cirolana sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Cirriformia tentaculata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0
Corophiid 1 0.0 90.3 145.9 549.0 1855.5 2293.3 145.9 0.0 20.8 0.0 0.0 0.0 0.0 90.3 76.4 500.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Corophiid 2 0.0 0.0 0.0 0.0 0.0 0.0 27.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0
Crepidula spp. 0.0 0.0 0.0 20.8 0.0 41.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 27.8 0.0 0.0 0.0 0.0 0.0 0.0 118.1 0.0 0.0 0.0 0.0
Cumacea sp. 2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 20.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Diopatra monroi 0.0 889.5 257.1 465.6 3182.8 2911.7 1820.7 840.9 535.1 694.9 62.5 13.9 0.0 0.0 0.0 4725.5 0.0 0.0 0.0 0.0 0.0 347.5 4975.7 625.4 62.5 0.0 0.0
Discinisca tenuis 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dosinia lupinus orbignyi 6.9 34.7 13.9 76.4 13.9 6.9 6.9 0.0 83.4 542.0 6.9 0.0 0.0 0.0 27.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dromidia sp. 0.0 41.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Eunicella papillosa 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Fusciolariidae 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaropsis sophiae 0.0 0.0 361.4 20.8 0.0 0.0 458.7 0.0 0.0 125.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Glycera convoluta 0.0 41.7 0.0 0.0 13.9 6.9 0.0 0.0 0.0 27.8 0.0 6.9 13.9 0.0 0.0 0.0 6.9 6.9 0.0 0.0 0.0 0.0 20.8 6.9 6.9 0.0 0.0
Harmothoe spp. 0.0 0.0 6.9 27.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Hippomedon onconotus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 41.7 0.0 0.0 0.0 0.0 0.0
Hymensoma obiculare 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Iphinae sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 152.9 0.0 97.3 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 48.6 0.0 0.0 0.0 0.0 0.0 0.0
Lemboides crenatipalma 0.0 6.9 83.4 145.9 576.8 632.4 48.6 0.0 0.0 0.0 0.0 0.0 0.0 20.8 13.9 236.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Leucothoe richiardi 0.0 20.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Listriella lindae 0.0 0.0 6.9 0.0 41.7 76.4 20.8 27.8 0.0 6.9 0.0 0.0 0.0 34.7 41.7 27.8 0.0 0.0 0.0 0.0 0.0 0.0 20.8 0.0 13.9 0.0 0.0
Lumbrinereis heteropoda 20.8 6.9 27.8 83.4 41.7 13.9 34.7 27.8 6.9 27.8 55.6 20.8 0.0 0.0 6.9 69.5 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0

Page | 45
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
difficilis
Lysianassa ceratina 0.0 48.6 0.0 13.9 0.0 0.0 0.0 0.0 0.0 20.8 0.0 0.0 0.0 0.0 41.7 0.0 41.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Macoma crawfordi ordinaria 6.9 76.4 0.0 145.9 194.6 145.9 76.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 180.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 27.8 41.7 0.0 0.0
Maera inaequipes 0.0 6.9 0.0 132.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Magelona papillicornis 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 13.9 5017.4 368.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 222.4 0.0 0.0 0.0 20.8 0.0 0.0
Melita sp. 0.0 0.0 0.0 6.9 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0
Melita subchelata 0.0 0.0 27.8 76.4 701.9 83.4 76.4 41.7 0.0 6.9 0.0 0.0 0.0 48.6 1042.4 76.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nassarius vinctus 27.8 27.8 0.0 41.7 90.3 180.7 0.0 0.0 0.0 201.5 0.0 0.0 0.0 0.0 48.6 97.3 0.0 0.0 0.0 0.0 0.0 0.0 104.2 0.0 0.0 0.0 0.0
Nebalia capensis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nematode 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nephtys hombergii 0.0 0.0 6.9 0.0 13.9 0.0 0.0 27.8 0.0 0.0 0.0 333.6 166.8 0.0 20.8 6.9 0.0 13.9 0.0 0.0 125.1 111.2 13.9 13.9 69.5 6.9 0.0
Nephtys sphaerocirrata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 34.7 0.0 0.0 0.0 0.0 111.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 27.8 0.0 0.0
Nereis spp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 6.9 0.0 13.9 0.0 20.8 0.0 0.0 20.8 0.0 0.0 0.0 0.0 0.0
Notomastus latericeus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nucula nucleus 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Orbinia angrapequensis 0.0 0.0 0.0 159.8 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 6.9 27.8 6.9 6.9 0.0 0.0 0.0 0.0 13.9 0.0 0.0
Owenia fusiformis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 180.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Paramoera capensis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 597.6 0.0 0.0 0.0 0.0 0.0
Paraprionospio pinnata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pectinaria capensis 0.0 0.0 0.0 27.8 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 271.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 486.4 0.0 41.7 0.0 208.5 13.9 0.0
Pherusa swakopiana 0.0 6.9 222.4 0.0 0.0 0.0 0.0 0.0 0.0 166.8 76.4 34.7 48.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0
Photis longidactylus 0.0 20.8 0.0 0.0 173.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 785.3 0.0 0.0 0.0 0.0 0.0 0.0 333.6 0.0 0.0 0.0 0.0 0.0
Photis longimanus 0.0 6.9 0.0 6.9 132.0 90.3 0.0 111.2 0.0 0.0 0.0 0.0 0.0 159.8 820.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0

Page | 46
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Pseudopotamilla reniformis 0.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pterygosquilla armata capensis 0.0 6.9 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pycnogonid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Sabellides luderitzi 0.0 0.0 0.0 0.0 20.8 0.0 0.0 0.0 0.0 13.9 13.9 1237.0 646.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 97.3 0.0 0.0
Schistomeringos rudolphii 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Sipunculid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 20.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Sphaerodorum gracile 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0
Sphaeromatidae 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Spiroplax spiralis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 34.7 0.0 13.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Tellimya sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 222.4 0.0 0.0 0.0 0.0 0.0 0.0
Tellina gilchristi 6.9 104.2 312.7 180.7 278.0 0.0 0.0 0.0 6.9 152.9 0.0 0.0 0.0 0.0 6.9 562.9 0.0 0.0 0.0 0.0 0.0 20.8 0.0 0.0 0.0 0.0 0.0
Tellina trilatera 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 104.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Upogebia capensis 0.0 0.0 0.0 0.0 41.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0
Upogebia africana 0.0 0.0 0.0 6.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 0.0 0.0 0.0 152.9 0.0 180.7 291.9 250.2 0.0 0.0 0.0 0.0 0.0 0.0
Urothoe grimaldi 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 34.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 333.6 159.8 0.0 0.0 0.0 0.0 0.0
Venerupis corrugatus 0.0 0.0 0.0 298.8 173.7 83.4 48.6 34.7 0.0 0.0 0.0 0.0 0.0 0.0 6.9 736.6 6.9 0.0 0.0 0.0 0.0 20.8 729.7 20.8 0.0 0.0 0.0
Virgularia schultzei 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 145.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 13.9 0.0 222.4 0.0 0.0 0.0 0.0

Page | 47
ANNEXURE 3: BENTHIC MACROFAUNA SAMPLED IN 2012 (biomass per m2)
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Afrophaxas decipiens 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 26.7 0.0 0.0 2.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0
Amaryllis macrophthalma 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca anomola 0.0 0.0 0.0 10.7 10.9 1.2 0.0 26.6 0.0 0.0 0.0 0.1 0.0 3.1 25.3 0.3 0.0 0.1 0.0 0.0 0.1 0.0 0.1 0.0 0.0 0.0 0.0
Ampelisca brevicornis 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca palmata 0.0 0.0 0.0 10.5 3.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Ampelisca spinimana 0.0 0.0 2.2 0.0 0.0 0.0 0.6 0.0 0.0 0.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 4.1 0.0 0.0
Amphieteis gunneri 0.0 0.0 0.0 9.2 0.0 0.0 1.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Amphilochus neapolitanus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Anemone 0.0 4.8 64.8 36.5 24.0 19.5 50.4 0.3 0.0 2.8 99.0 26.6 8.2 0.0 33.7 30.7 0.1 0.0 0.0 0.0 0.0 25.0 36.9 5.2 80.1 0.0 0.0
Anemone 2 44.6 0.0 0.0 80.5 0.0 0.0 0.0 45.8 0.0 0.0 39.9 31.9 33.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 23.3 0.0 0.0 0.0 0.0 0.0 0.0
Anemone 3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Anthelura remipes 0.3 0.0 0.0 0.1 0.6 0.7 0.0 0.6 0.0 0.0 3.1 2.4 2.2 0.0 0.9 0.8 0.0 0.0 0.1 0.0 1.6 0.0 0.0 0.0 0.3 0.0 0.0
Arenicola loveni 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 630.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 27.3 0.0 0.0
Ascidian 0.0 106.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Bathyporeia sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Bullia laevissima 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 141.7 0.0 0.0 0.0 0.0 0.0 41.4 0.0 0.0 0.0 0.0 0.0 0.0
Callichirus kraussi 0.0 0.0 0.0 0.8 1.2 11.3 41.8 6.9 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.8 0.0 74.7 0.0 0.0 0.0 0.0 0.4 0.0 1.0 0.0 0.0
Calyptraea chinensis 0.0 0.0 0.0 94.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Capitella capitata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.8 0.0 0.0 0.0
Caprellid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Caprellid 2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Choromytilus meridionalis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 839.2 435.0 0.0 0.0 0.0 0.0
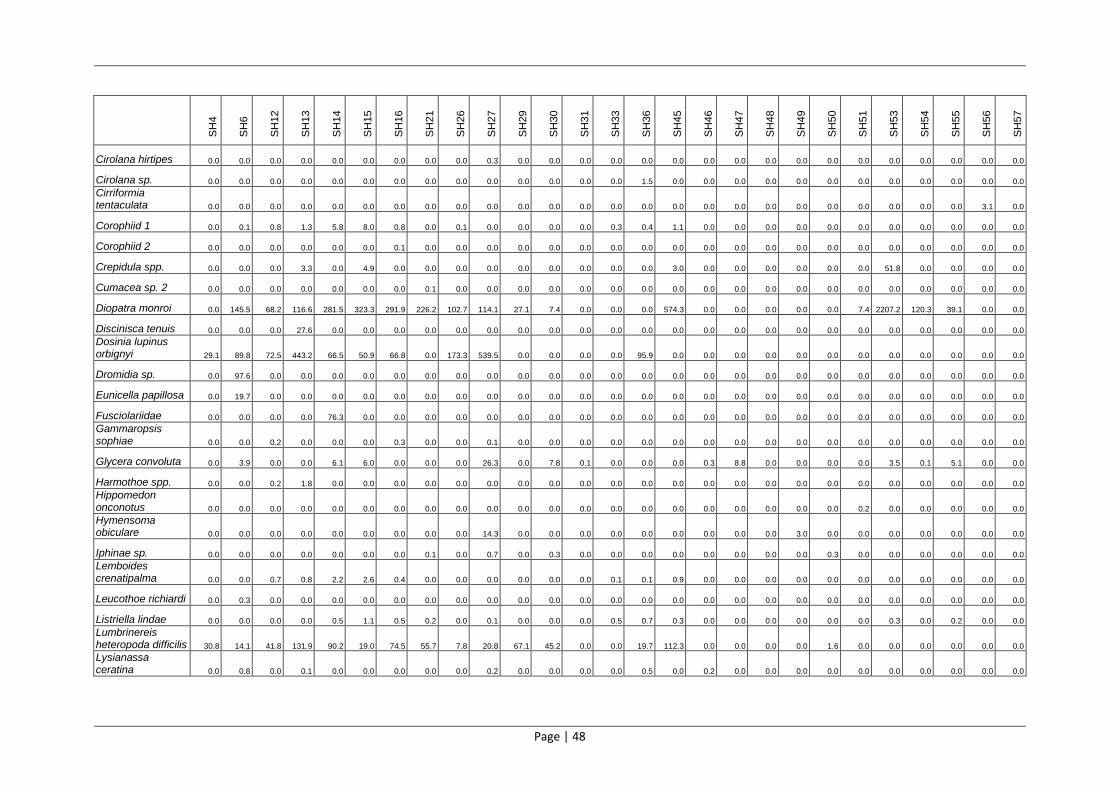
Page | 48
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Cirolana hirtipes 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Cirolana sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Cirriformia tentaculata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.1 0.0
Corophiid 1 0.0 0.1 0.8 1.3 5.8 8.0 0.8 0.0 0.1 0.0 0.0 0.0 0.0 0.3 0.4 1.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Corophiid 2 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Crepidula spp. 0.0 0.0 0.0 3.3 0.0 4.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.0 0.0 0.0 0.0 0.0 0.0 0.0 51.8 0.0 0.0 0.0 0.0
Cumacea sp. 2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Diopatra monroi 0.0 145.5 68.2 116.6 281.5 323.3 291.9 226.2 102.7 114.1 27.1 7.4 0.0 0.0 0.0 574.3 0.0 0.0 0.0 0.0 0.0 7.4 2207.2 120.3 39.1 0.0 0.0
Discinisca tenuis 0.0 0.0 0.0 27.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dosinia lupinus orbignyi 29.1 89.8 72.5 443.2 66.5 50.9 66.8 0.0 173.3 539.5 0.0 0.0 0.0 0.0 95.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Dromidia sp. 0.0 97.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Eunicella papillosa 0.0 19.7 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Fusciolariidae 0.0 0.0 0.0 0.0 76.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Gammaropsis sophiae 0.0 0.0 0.2 0.0 0.0 0.0 0.3 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Glycera convoluta 0.0 3.9 0.0 0.0 6.1 6.0 0.0 0.0 0.0 26.3 0.0 7.8 0.1 0.0 0.0 0.0 0.3 8.8 0.0 0.0 0.0 0.0 3.5 0.1 5.1 0.0 0.0
Harmothoe spp. 0.0 0.0 0.2 1.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Hippomedon onconotus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 0.0
Hymensoma obiculare 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 14.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Iphinae sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.7 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0
Lemboides crenatipalma 0.0 0.0 0.7 0.8 2.2 2.6 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.1 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Leucothoe richiardi 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Listriella lindae 0.0 0.0 0.0 0.0 0.5 1.1 0.5 0.2 0.0 0.1 0.0 0.0 0.0 0.5 0.7 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.2 0.0 0.0
Lumbrinereis heteropoda difficilis 30.8 14.1 41.8 131.9 90.2 19.0 74.5 55.7 7.8 20.8 67.1 45.2 0.0 0.0 19.7 112.3 0.0 0.0 0.0 0.0 1.6 0.0 0.0 0.0 0.0 0.0 0.0
Lysianassa ceratina 0.0 0.8 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 0.5 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0

Page | 49
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Macoma crawfordi ordinaria 1.9 69.8 0.0 199.1 152.3 137.5 94.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 67.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 10.6 1.1 0.0 0.0
Maera inaequipes 0.0 0.2 0.0 2.2 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Magelona papillicornis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 27.3 0.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 6.7 0.0 0.0 0.0 0.4 0.0 0.0
Melita sp. 0.0 0.0 0.0 0.0 0.0 0.1 0.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0
Melita subchelata 0.0 0.0 0.2 0.3 3.2 0.5 0.0 0.2 0.0 0.1 0.0 0.0 0.0 0.4 5.5 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nassarius vinctus 9.5 6.3 0.0 11.0 33.8 74.1 0.0 0.0 0.0 68.9 0.0 0.0 0.0 0.0 9.1 30.4 0.0 0.0 0.0 0.0 0.0 0.0 32.5 0.0 0.0 0.0 0.0
Nebalia capensis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nematode 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nephtys hombergii 0.0 0.0 3.9 0.0 3.2 0.0 0.0 9.7 0.0 0.0 0.0 5.1 9.4 0.0 9.8 3.7 0.0 20.2 0.0 0.0 38.4 64.0 7.7 0.4 33.4 0.1 0.0
Nephtys sphaerocirrata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0
Nereis spp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 1.9 0.0 0.1 0.0 0.1 0.0 0.0 0.6 0.0 0.0 0.0 0.0 0.0
Notomastus latericeus 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Nucula nucleus 0.0 6.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Orbinia angrapequensis 0.0 0.0 0.0 41.5 1.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.1 1.7 0.3 0.1 0.0 0.0 0.0 0.0 0.6 0.0 0.0
Owenia fusiformis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Paramoera capensis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 1.7 0.0 0.0 0.0 0.0 0.0
Paraprionospio pinnata 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pectinaria capensis 0.0 0.0 0.0 4.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 10.6 0.0 0.0 0.0 0.0 0.0 0.0 0.0 58.1 0.0 83.8 0.0 48.3 0.1 0.0
Pherusa swakopiana 0.0 3.7 133.4 0.0 0.0 0.0 0.0 0.0 0.0 27.3 52.1 31.5 78.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 3.5 0.0 0.0
Photis longidactylus 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.5 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0
Photis longimanus 0.0 0.0 0.0 0.0 0.1 0.1 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.1 0.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pseudopotamilla reniformis 0.0 5.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pterygosquilla armata capensis 0.0 17.7 0.0 0.0 0.0 15.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 10.4 12.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Pycnogonid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.4 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0

Page | 50
SH
4
SH
6
SH
12
SH
13
SH
14
SH
15
SH
16
SH
21
SH
26
SH
27
SH
29
SH
30
SH
31
SH
33
SH
36
SH
45
SH
46
SH
47
SH
48
SH
49
SH
50
SH
51
SH
53
SH
54
SH
55
SH
56
SH
57
Sabellides luderitzi 0.0 0.0 0.0 0.0 3.9 0.0 0.0 0.0 0.0 0.1 0.2 15.8 18.3 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.5 0.0 0.0
Schistomeringos rudolphii 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Sipunculid 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 19.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Sphaerodorum gracile 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.3 0.0 0.0 0.0
Sphaeromatidae 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.6 0.0 0.2 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Spiroplax spiralis 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.5 0.0 1.9 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Tellimya sp. 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.2 0.0 0.0 0.0 0.0 0.0 0.0
Tellina gilchristi 6.2 46.1 440.4 125.9 105.3 0.0 0.0 0.0 4.7 120.8 0.0 0.0 0.0 0.0 0.9 286.8 0.0 0.0 0.0 0.0 0.0 11.6 0.0 0.0 0.0 0.0 0.0
Tellina trilatera 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 113.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0
Upogebia capensis 0.0 0.0 0.0 0.0 4.5 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.7 0.0 0.0 0.0 0.0
Upogebia africana 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 1.4 0.0 0.0 0.0 0.0 88.0 0.0 136.5 275.2 16.0 0.0 0.0 0.0 0.0 0.0 0.0
Urothoe grimaldi 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.1 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.9 0.4 0.0 0.0 0.0 0.0 0.0
Venerupis corrugatus 0.0 0.0 0.0 823.0 822.8 439.8 124.0 325.6 0.0 0.0 0.0 0.0 0.0 0.0 18.3 3712.7 5.5 0.0 0.0 0.0 0.0 3.4 2773.4 1.5 0.0 0.0 0.0
Virgularia schultzei 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 0.0 34.5 96.8 0.0 0.0 0.0 0.0 0.0 0.0 0.0 2.8 0.0 1545.4 0.0 0.0 0.0 0.0





























