Embed Size (px)
Citation preview

Cell Tiss. Res. 191,389~,04 (1978) Cell and Tissue Research �9 by Springer-Verlag 1978
Surface Fine Structure of the Subfornical Organ in the Japanese Quail, Coturnix coturnix japonica *
Yoshio Takei, Kazuhiko Tsuneki, and Hideshi Kobayashi
Misaki Marine Biological Station, University of Tokyo, Misaki, Kanagawa-ken, Japan
Summary. The surface ultrastructure of the subfornical organ (SFO) was investigated in the Japanese quail. The SFO consists of a body and a stalk. The body of the SFO can be divided into rostral and caudal parts. On the rostral part, each ependymal cell possesses a short central solitary cilium; clustered cilia are also occasionally seen. Microvilli are abundant. On the caudal part, cells with a solitary cilium are fewer in number, and clustered cilia are rarely found. Microvilli are not as abundant as on the rostral part. In addition, large bulbous protrusions, tufts of small protrusions, deep funnel-shaped hollows, small pinocytotic invaginations and possible cerebrospinal fluid-contacting axons are sporadically observed on the surface of various regions of the body. Each ependymal cell of the stalk has a wide apical surface. A central solitary cilium, microvilli and other structures are observed more rarely on the stalk than on the body, while clustered cilia are not seen on the stalk. These structures are compared with those of the mammalian SFO and further discussed in relation to the possible dipsogenic receptor function for angiotensin II.
Key words: Subfornical organ - Japanese quail, Coturnix co tu rn i x japon ica -
Ultrastructure - Scanning electron microscopy.
Introduction
The subfornical organ (SFO) is a hemispheric circumventricular protrusion located on the rostral wall of the third ventricle just below the interventricular foramen. For several years, the SFO has attracted attention of physiologists as a dipsogenic receptor site for angiotensin II (AII) in the rat (Simpson and Routtenberg, 1973) and the Japanese quail (Takei, 1977a). Since intracerebro- ventricular injection of AII effects drinking in mammals (see Severs and Summy- Long, 1975) and birds (Takei, 1977b; Schwob and Johnson, 1977), the surface structure of this organ is of particular interest because of its inferred dipsogenic receptor function. So far, ultrastructural studies of the surface of the SFO have been
Send offprint requests to: Yoshio Takei, Misaki Marine Biological Station, University of Tokyo, Misaki, Kanagawa-ken, 238-02 Japan
* The authors would like to express their gratitude to Dr. Ebert A. Ashby for his kindness in reading the manuscript. This work was supported by grants from the Ministry of Education, Japan and from the Ford Foundation to Prof. Hideshi Kobayashi
0302-766X/78/0191/0389/$03.20

390 Y. Takei et al.
c o n c e r n e d exc lus ive ly w i t h m a m m a l s (dog , A n d r e s , 1965; r a b b i t , R u d e r t e t al.,
1968, L e o n h a r d t a n d L i n d e m a n n , 1973; r a t , D e m p s e y , 1968, Ph i l l i p s e t al. , 1974,
D e l l m a n n a n d S i m p s o n , 1975, 1976; m o u s e , S c h i n k o e t al. , 1972; m a n , S c o t t e t al. ,
1972). S tud i e s o f the s u r f ace u l t r a s t r u c t u r e o f t h e a v i a n S F O h a v e n o t b e e n r e p o r t e d p r ev ious ly .
I n t h e p r e s e n t s tudy , t he s u r f a c e u l t r a s t r u c t u r e o f t he S F O in t he J a p a n e s e q u a i l
w a s i n v e s t i g a t e d b y b o t h s c a n n i n g a n d t r a n s m i s s i o n e l e c t r o n m i c r o s c o p y . R e s u l t s
o b t a i n e d a r e c o m p a r e d w i t h t h e s u r f ace s t r u c t u r e s o f t h e m a m m a l i a n S F O a n d
d i s c u s s e d in c o n n e c t i o n w i t h i ts p o s s i b l e f u n c t i o n as a d i p s o g e n i c r e c e p t o r s i te f o r A I I .
Materials and Methods
Thirty male Japanese quail, Coturnix coturnixjaponica Temminck et Schlegel, were purchased from a commercial source at the age of 4 weeks. They were kept individually in small wire cages under a short daily photoperiod (8 L/16D) at 25 + 1 ~ C for at least one week before sacrifice. Pellet food (Nippon Haigo Shiryo, Yokohama) and water were given ad libitum. Each animal weighed about 90 g at the time of sacrifice. For scanning (SEM) and transmission (TEM) electron microscopy, the SFO was prefixed by one of the following three methods:
1. Immersion Fixation
Thirteen birds were used for SEM and four for TEM. Immediately after decapitation, the brain was quickly dissected out and a small piece of tissue containing the SFO was isolated from the brain. The tissue was fixed with either 5 ~ glutaraldehyde in 0.1 M phosphate buffer (pH, 7.4) at 4 ~ C or a modified Karnovsky fixative I (2 ~ paraformaldehyde + 2.5 ~ glutaraldehyde in 0.08 M phosphate buffer, pH, 7.4) for 3 h at 25 ~ C.
2. Systemic Perfusion Fixation
Six birds (2 for SEM and 4 for TEM) were perfused intracardially with 10 ml of a modified Karnovsky fixative II (1 ~ paraformaldehyde + 1.25 ~ glutaraldehyde in 0.08 M phosphate buffer, pH, 7.4). After perfusion, the brain was dissected out and trimmed to a small piece containing the SFO. The tissue was further immersed in the Karnovsky fixative I for 3 h at 25 ~ C.
3. Cerebroventricular Perfusion Fixation
The ventricles of seven birds were perfused with the Karnovsky fixative I for the SEM study. For the perfusion, birds were fixed in a stereotaxic apparatus after Nembutal anesthesia (4 mg/100 g BW). The frontal and parietal bones were exposed and a part of these bones (8 mm in diameter) was removed carefully with a dental saw. The dura covering the brain was removed using a pair of dental scissors. Care was taken not to break the mid-sagittal cranial sinus. A stainless steel cannula (o.d.=0.5mm) was inserted into the brain and the lower tip was placed near the bottom of the third ventricle. The location of the cannula tip was adjusted by lateral and frontal radiographs. The eannula, a syringe and a connecting polyethylene tube (PE 20), were prefilled with fixative before insertion of the cannula. Perfusion was initiated using an infusion pump (Natsume, Tokyo) at a rate of 0.33 ml/min for 6 min. Immediately after beginning perfusion, an incision was made in the cerebral hemispheres just behind the inserted cannula in order to allow the CSF and the fixative to overflow from the ventricle at the same rate as the infusion. After perfusion, the birds were decapitated and the brain removed and trimmed to a small block. The tissue was further fixed in the Karnovsky fixative I for 3 h at 25 ~ C.
After prefixation by one of the above three methods, the tissue was rinsed gently in 0.1 M phosphate buffer (pH, 7.4) at 4 ~ C. The specimen was then postfixed with 1 ~ osmium tetroxide in 0.1 M phosphate buffer (pH, 7.4) for 2 or 3 h at 4~ In some cases, sucrose (final concentration 0.1 M) was added to the

Surface Fine Structure of the Quail Subfornical Organ 391
rinse solution and the postfixative. The specimen was dehydrated with an ascending series of ethanol concentrations.
For the SEM study, the dehydrated specimen was dried with a critical point drying apparatus (HCP-1, Hitachi) using CO2 as the transient fluid. The walls of ventricle near the SFO were removed carefully under a dissecting microscope in order to expose the SFO. The specimen was mounted on an aluminum stub and coated with gold using an ion coater (IB-3, Eiko Engineering Co. Ltd.). The tissue blocks were viewed with a Hitachi HHS-2R or Hitachi-Akashi HSM-2 scanning electron microscope.
For the TEM study, the dehydrated specimens were embedded in Epon. In most cases, transverse sections were cut. They were stained with uranyl acetate and lead citrate, and observed with a JEOL T-8 transmission electron microscope. Some specimens, used first for the SEM study, were subsequently embedded in Epon and used for the TEM study.
Results
Scanning Electron Microscopy
The SFO of the Japanese quail protrudes into the third ventricle and shows a flask- like appearance (Fig. 1 a). The outstanding feature of the surface is an extreme paucity of cilia compared with the surrounding ventricular walls. The SFO can be divided topographically into two parts, the body and the stalk (Fig. 1 a). The former occupies the ventrorostral part and the latter the dorsocaudal part of the SFO (Fig. 1 b). The body measures about 0.27 m m in length and the stalk 0.23 mm. The width of the body is about 0.19ram at the broadest point and that of the stalk 0.06 mm. The most caudal part of the stalk is covered with the choroid plexus (Fig. 1 a). The following description is based chiefly upon specimens fixed by ventricular perfusion.
1. Body of the SFO
The ventricular surface of the body of the SFO consists of hexagonal ependymal cells of varying sizes. Although the number of cilia on the SFO is generally small compared with the surroundings, at least two types of ependymal cells can be distinguished according to the number of cilia. The first type possesses a cluster of cilia (Fig. 2). The cells of this type are relatively abundant on the rostral and peripheral parts of the body (Fig. 1 a). The number of cilia varies between 17 and 27 (usually 21). The size of each cilium is about 7.3 ~tm in length and 0.18 ~tm in thick- ness. Thin and fairly long microvilli ( length=l .00~tm; thickness=0.07~tm) densely cover the cell surface, especially around the base of cilia (Fig. 2). The cells of the second type lack clustered cilia and are found on nearly the whole surface of the SFO. The cells of this type are clearly distinguishable from each other by distinct cell boundaries (Fig. 3). Some of these cells have a short solitary cilium (length = 0.85-1.88 ~tm; thickness= 0.15-0.19 ~tm) located in the center of the cell. Especially on the rostral part of the body, each cell invariably possesses a solitary cilium (Figs. 3- 6). The cells bearing a solitary cilium decrease in number caudally. In addition, a peripheral long solitary cilium is very rarely found in the rostral part of the body (Fig. 7 a). This cilium can be clearly distinguished from the short central solitary cilium. It cannot be determined whether this latter cell type with a single

392 Y. Takei et al.
Fig. 1. a Scanning electron micrograph of the subfornical organ (SFO) of the Japanese quail viewed at low magnification, cb Caudal part of body, cc clustered cilia observed sporadically on the body of the SFO, ce coagulated erythrocytes, cp choroid plexus, rb rostral part of body, s stalk. Ventricular perfusion. • 250. b Schematic drawing of a mid-sagittal section through the quail brain showing the location of the SFO. A C anterior commissure, OC optic chiasma, PC pallial commissure, POM medial preoptic area, I I I third ventricle
long cilium represents a unique cell type or whether it belongs to the first type of cell with clustered cilia, from which the other cilia have been lost.
Some cells of the second type have few microvilli, while others possesse numerous microvilli. The length of microvilli varies from 0.1 ~tm to 0.9 gin, and the thickness measures about 0.1 gm. A single cell surface exhibits microvilli of almost uniform size. Each cell of the rostral part of the body has a relatively large number of microvilli, while the number of cells without or with few microvilli tends to increase caudally. The ependymal cells of the rostral part are generally smaller than those on the caudal part. For these reasons, the body of the SFO can be divided into rostral and caudal parts according to the size of the ependymal cell and relative abundance of cilia and microvilli, although there is no distinct boundary between these two parts.
In addition to cilia and microvilli, other structures are also found on the body of the SFO. Tufts of small protrusions are often encountered, especially on the rostral part of the body (Fig. 6), measuring 1.55 to 1.72 gm in diameter. A funnel-shaped hollow lined with ependymal cells is occasionally observed (Fig. 3). The inner wall of the hollow is covered with small protrusions, cilium-like structures and/or microvilli of varying sizes. Another remarkable feature of the body of the SFO is the
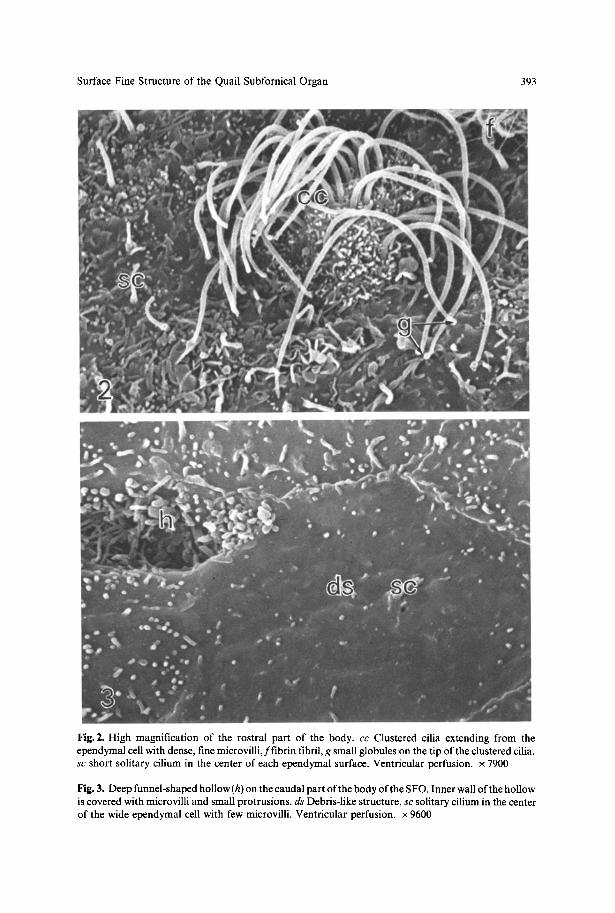
Surface Fine Structure of the Quail Subfornical Organ 393
Fig. 2. High magnification of the rostral part of the body. c c Clustered cilia extending from the ependymal cell with dense, fine microvilli,ffibrin fibril, g small globules on the tip of the clustered cilia, s c short solitary cilium in the center of each ependymal surface. Ventricular perfusion, x 7900
Fig. 3. Deep funnel-shaped hollow (h) on the caudal part of the body of the SFO. Inner wall of the hollow is covered with microvilli and small protrusions, d s Debris-like structure, s c solitary cilium in the center of the wide ependymal cell with few microvilli. Ventricular perfusion, x 9600
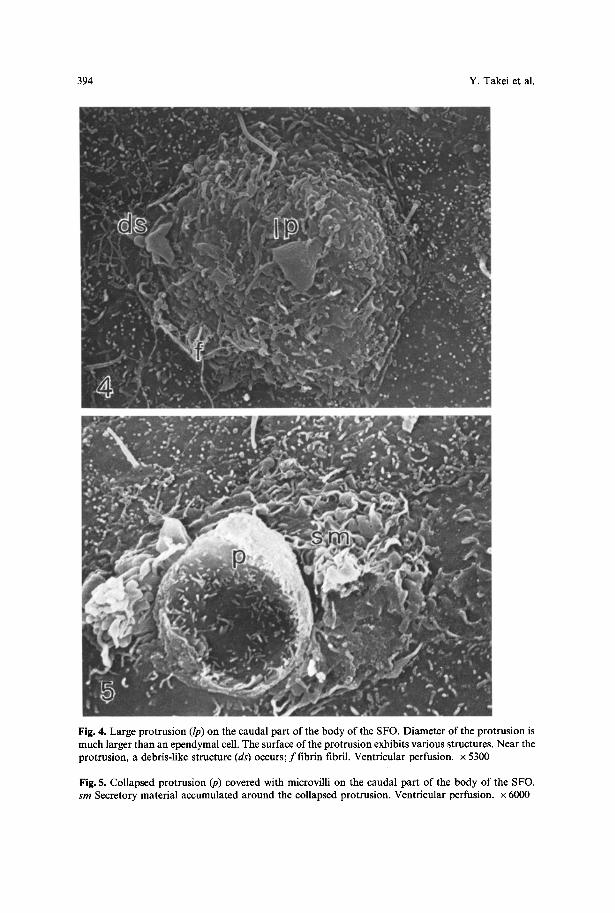
394 Y. Takei et al.
Fig. 4. Large protrusion (lp) on the caudal part of the body of the SFO. Diameter of the protrusion is much larger than an ependymal cell. The surface of the protrusion exhibits various structures. Near the protrusion, a debris-like structure (ds) occurs; f f i b r i n fibril. Ventricular perfusion, x 5300
Fig. 5. Collapsed protrusion (p) covered with microvilli on the caudal part of the body of the SFO. sm Secretory material accumulated around the collapsed protrusion. Ventricular perfusion, x 6000
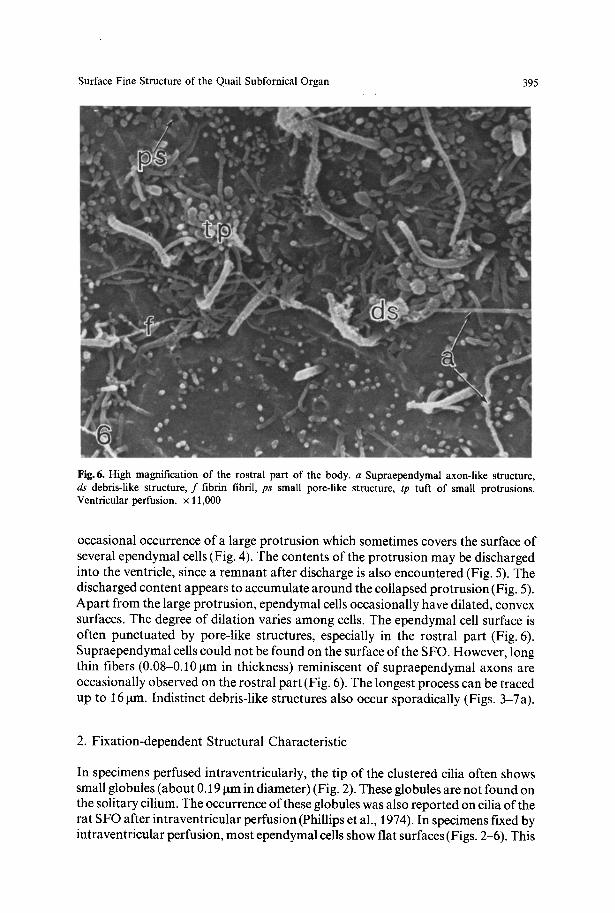
Surface Fine Structure of the Quail Subfornical Organ 395
Fig. 6. High magnification of the rostral part of the body. a Supraependymal axon-like structure, ds debris-like structure, f fibrin fibril, ps small pore-like structure, tp tuft of small protrusions. Ventricular perfusion, x 11,000
occasional occurrence of a large protrusion which sometimes covers the surface of several ependymal cells (Fig. 4). The contents of the protrusion may be discharged into the ventricle, since a remnant after discharge is also encountered (Fig. 5). The discharged content appears to accumulate around the collapsed protrusion (Fig. 5). Apart from the large protrusion, ependymal cells occasionally have dilated, convex surfaces. The degree of dilation varies among cells. The ependymal cell surface is often punctuated by pore-like structures, especially in the rostral part (Fig. 6). Supraependymal cells could not be found on the surface of the SFO. However, long thin fibers (0.08-0.10 gm in thickness) reminiscent of supraependymal axons are occasionally observed on the rostral part (Fig. 6). The longest process can be traced up to 16 gm. Indistinct debris-like structures also occur sporadically (Figs. 3-7a).
2. Fixation-dependent Structural Characteristic
In specimens perfused intraventricularly, the tip of the clustered cilia often shows small globules (about 0.19 gm in diameter) (Fig. 2). These globules are not found on the solitary cilium. The occurrence of these globules was also reported on cilia of the rat SFO after intraventricular perfusion (Phillips et al., 1974). In specimens fixed by intraventricular perfusion, most ependymal cells show flat surfaces (Figs. 2-6). This
396 Y. Takei et al.
is in c o n t r a s t to the convex surfaces obse rved in spec imens fixed by i m m e r s i o n or systemic pe r fu s ion (Figs. 7a a n d b). O n the fiat surface o f the spec imen per fused in t r aven t r i cu la r ly , f ib r in fibri ls f o r m e d by b l o o d c o a g u l a t i o n are f r equen t ly f o u n d (Figs. 2, 4, a n d 6). The e p e n d y m a l surface o f the la tera l b o r d e r o f the S F O somet imes p r o t r u d e s extens ively in to the ventr icle . Th i s fea ture is especial ly p r o m i n e n t in spec imens fixed by systemic pe r fu s ion (Fig. 7b). Smal l b leb- l ike
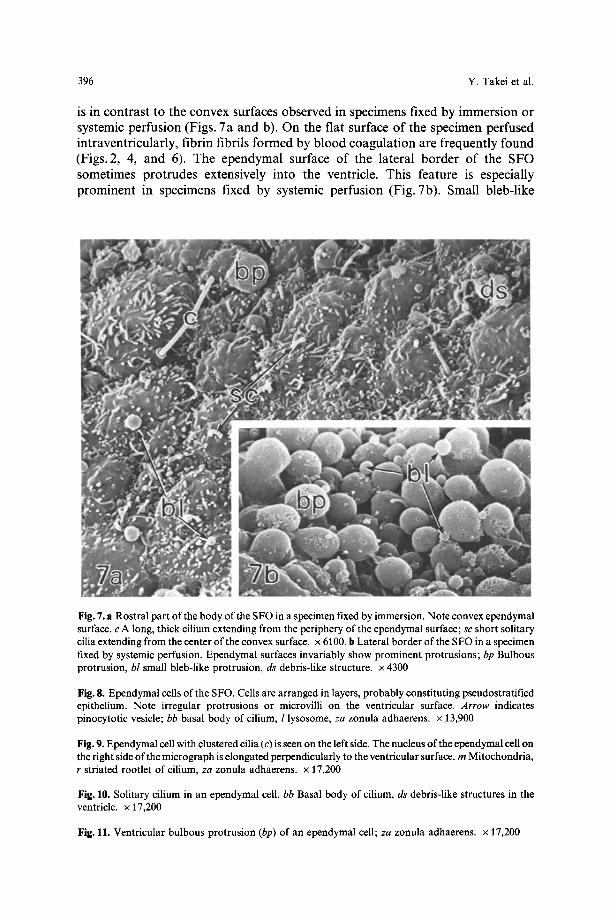
Fig. 7. a Rostral part of the body of the SFO in a specimen fixed by immersion. Note convex ependymal surface, c A long, thick cilium extending from the periphery of the ependymal surface; sc short solitary cilia extending from the center of the convex surface, x 6100. b Lateral border of the SFO in a specimen fixed by systemic perfusion. Ependymal surfaces invariably show prominent protrusions; bp Bulbous protrusion, bl small bleb-like protrusion, ds debris-like structure, x 4300
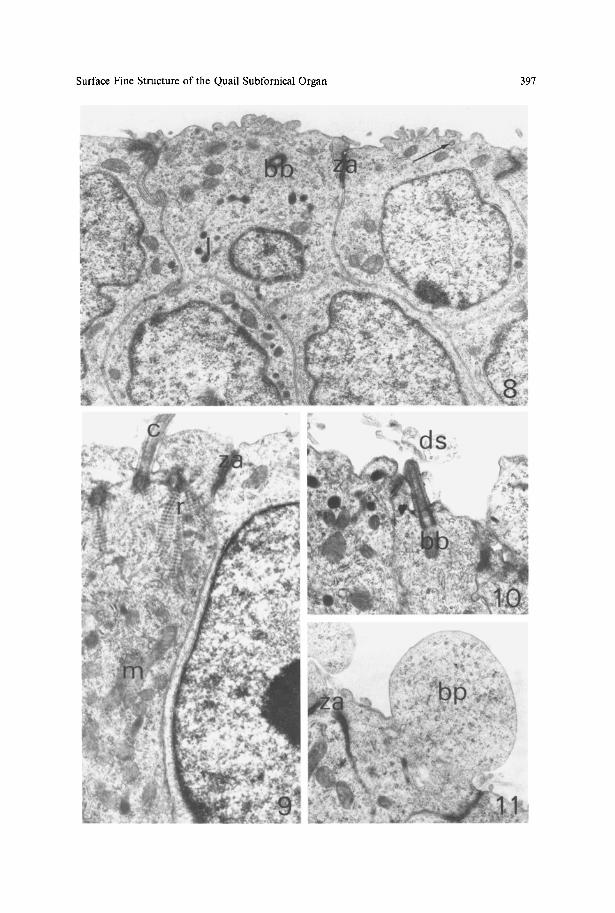
Fig. 8. Ependymal cells of the SFO. Cells are arranged in layers, probably constituting pseudostratified epithelium. Note irregular protrusions or microvilli on the ventricular surface. Arrow indicates pinocytotic vesicle; bb basal body of cilium, l lysosome, za zonula adhaerens, x 13,900
Fig. 9. Ependymal cell with clustered cilia (c) is seen on the left side. The nucleus of the ependymal cell on the right side of the micrograph is elongated perpendicularly to the ventricular surface, m Mitochondria, r striated rootlet of cilium, za zonula adhaerens, x 17,200
Fig. 10. Solitary cilium in an ependymal cell. bb Basal body of cilium, ds debris-like structures in the ventricle, x 17,200
Fig. 11. Ventricular bulbous protrusion (bp) of an ependymal cell; za zonula adhaerens, x 17,200
Surface Fine Structure of the Quail Subfornical Organ 397

398 Y. Takei et al.
protrusions are also frequently found on the surface of the SFO fixed by immersion or systemic perfusion (Figs. 7a and b). The size of the protrusion varies considerably. It is difficult to distinguish the smallest protrusion from microvilli.
3. Stalk of the SFO
The ependymal cells of the stalk show a uniform surface appearance (Fig. 1 a). Cells with clustered cilia are completely absent in this region. The ventricular surface of the stalk consists only of the second type ofependymal cells. The ventricular surface of each ependymal cell in the stalk is usually wider than that in the body. The numbers of cells with a central solitary cilium and cells with abundant microvilli are much smaller in the stalk than in the body. Other structures observed on the body of the SFO are absent or very rarely found on the surface of the stalk of the SFO.
Transmission Electron Microscopy
The ependymal and subependymal fine structures of the stalk are similar to those of the body. The following description is based on observations on the body unless otherwise stated. The ventricular side of the SFO is lined by ependymal cells (Figs. 8-15). Cilia, basal bodies and striated rootlets are infrequently observed (Figs. 9, 10 and 12). Cilia occasionally project into the ventricle from the shallow apical invagination (Figs. 9 and 12) Several cilia tend to make a cluster on an ependymal cell (Fig. 9). Clustered cilia are not found in the stalk. Ependymal cells with a single cilium occur much more often than those with clustered cilia (Fig. 10). Microvilli are found in relatively large numbers in some ependymal cells, but are few in others. The distribution and shape of microvilli are variable (Fig. 8). Large cytoplasmic bulbous protrusions are occasionally found on the surface (Fig. 11). Similar protrusions are numerous in the peripheral region of the SFO in some birds which were poorly fixed systemically. Large protrusions found in the SEM study on the periphery of the SFO (Fig. 7 b) may correspond to these protrusions. Transient structures between microvilli and cytoplasmic protrusions are present (Fig. 15). These structures may correspond to small bleb-like protrusions in the SEM study after immersion or systemic perfusion (Figs. 7 a and b). Pinocytotic vesicles (Fig. 8) reminiscent of pore-like structures in the SEM study(Fig. 6) are occasionally found. Large hollows are infrequently observed (Fig. 12). Slender electron dense ependymal cells, which are probably degenerating, infrequently face these cavities.
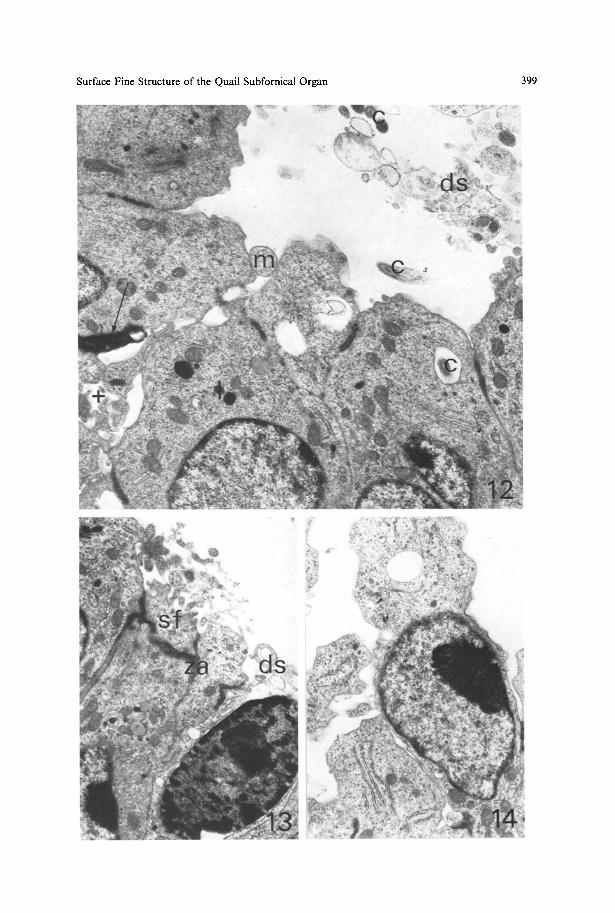
Fig. 12. Large hollow of the ependymal surface. In addition to cilia (c), many debris-like structures (ds) and an intact mitochondrion (m) are seen in the hollow. In the lower left corner, a deep narrow intercellular canal (+) is found, contacting degenerated dark process (arrow); l lysosome, x 13,900
Fig. 13. Superficial flap (sJ) of ependymal cell in contact with the underlying cell by a zonula adhaerens (za). Debris-like structures (ds) seen in the ventricle may originate from the cell situated in the lower right comer, which is apparently degenerating and about to be released into the ventricle. • 10,700
Fig. 14. Ependymal cell in the stalk of the SFO, which may be discharged into the ventricle. Note supranuclear cytoplasmic protrusion containing clear vacuole, x 9200
Surface Fine Structure of the Quail Subfornical Organ 399
400 Y. Takei et al.
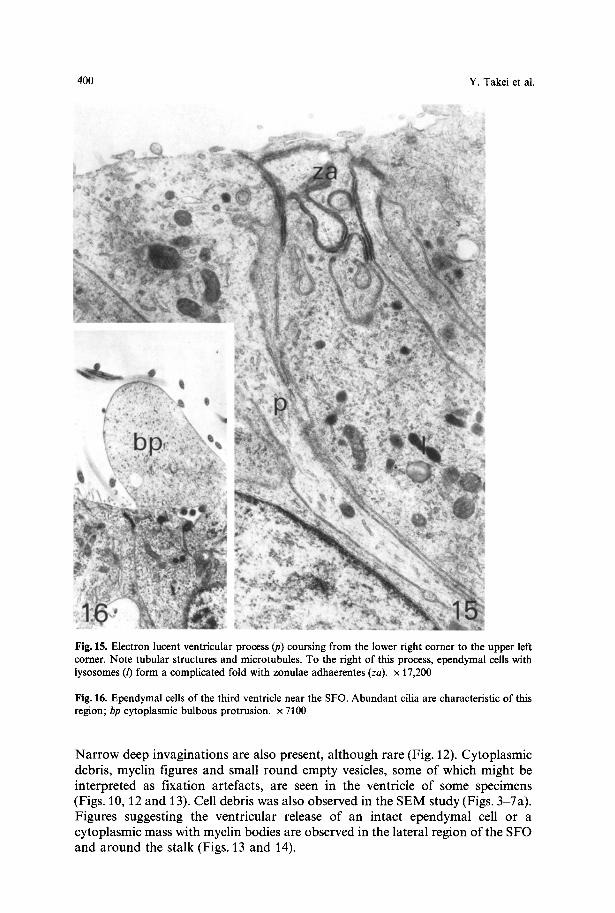
Fig. 15. Electron lucent ventricular process (p) coursing from the lower right corner to the upper left corner. Note tubular structures and microtubules. To the fight of this process, ependymal cells with lysosomes (/) form a complicated fold with zonulae adhaerentes (za). x 17,200
Fig. 16. Ependymal cells of the third ventricle near the SFO. Abundant cilia are characteristic of this region; bp cytoplasmic bulbous protrusion, x 7100
Narrow deep invaginations are also present, although rare (Fig. 12). Cytoplasmic debris, myelin figures and small round empty vesicles, some of which might be interpreted as fixation artefacts, are seen in the ventricle of some specimens (Figs. 10, 12 and 13). Cell debris was also observed in the SEM study (Figs. 3-7a). Figures suggesting the ventricular release of an intact ependymal cell or a cytoplasmic mass with myelin bodies are observed in the lateral region of the SFO and around the stalk (Figs. 13 and 14).
Surface Fine Structure of the Quail Subfornical Organ 401
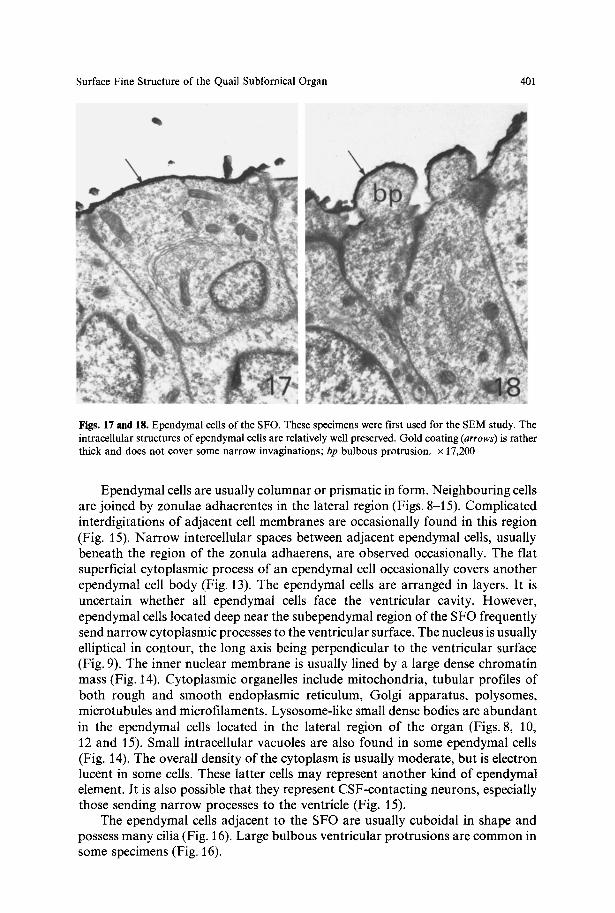
Figs. 17 and 18. Ependymal cells of the SFO. These specimens were first used for the SEM study. The intracellular structures of ependymal cells are relatively well preserved. Gold coating (arrows) is rather thick and does not cover some narrow invaginations; bp bulbous protrusion, x 17,200
Ependymal cells are usually columnar or prismatic in form. Neighbouring cells are joined by zonulae adhaerentes in the lateral region (Figs. 8-15). Complicated interdigitations of adjacent cell membranes are occasionally found in this region (Fig. 15). Narrow intercellular spaces between adjacent ependymal cells, usually beneath the region of the zonula adhaerens, are observed occasionally. The flat superficial cytoplasmic process of an ependymal cell occasionally covers another ependymal cell body (Fig. 13). The ependymal cells are arranged in layers. It is uncertain whether all ependymal cells face the ventricular cavity. However, ependymal cells located deep near the subependymal region of the SFO frequently send narrow cytoplasmic processes to the ventricular surface. The nucleus is usually elliptical in contour, the long axis being perpendicular to the ventricular surface (Fig. 9). The inner nuclear membrane is usually lined by a large dense chromatin mass (Fig. 14). Cytoplasmic organelles include mitochondria, tubular profiles of both rough and smooth endoplasmic reticulum, Golgi apparatus, polysomes, microtubules and microfilaments. Lysosome-like small dense bodies are abundant in the ependymal cells located in the lateral region of the organ (Figs. 8, 10, 12 and 15). Small intracellular vacuoles are also found in some ependymal cells (Fig. 14). The overall density of the cytoplasm is usually moderate, but is electron lucent in some cells. These latter cells may represent another kind of ependymal element. It is also possible that they represent CSF-contacting neurons, especially those sending narrow processes to the ventricle (Fig. 15).
The ependymal cells adjacent to the SFO are usually cuboidal in shape and possess many cilia (Fig. 16). Large bulbous ventricular protrusions are common in some specimens (Fig. 16).

402 Y. Takei et al.
The fixation of material first studied with the SEM was satisfactory for TEM observation (Figs. 17 and 18). The gold coating appears as a dense envelope on the ventricular surface, but is absent in the small cytoplasmic invaginations, especially those oblique to the surface. Since this coating is relatively thick, the details of the surface ultrastructure are somewhat indistinct in some specimens.
Discussion
The surface of the quail SFO is covered with relatively few cilia compared with the surrounding ventricular walls as was seen in the mammalian SFO (rabbit, Leonhardt and Lindemann, 1973; rat, Phillips et al., 1974). The organum vasculosum of the lamina terminalis of the Japanese quail (Mikami, 1976) and various mammalian circumventricular organs (Weindl, 1973) also possess a small number of cilia. The paucity of cilia seems to be a common feature of the ependymal cells of circumventricular organs.
Although ependymal cells of the SFO have a small number of cilia in all species so far investigated, there are some differences in the nature and distribution of cilia among species. In the quail SFO, two types of ependymal cells are distinguishable. One type possesses a cluster of cilia and the other a short central solitary cilium. These observations grossly coincide with the report in the rabbit (Leonhardt and Lindemann, 1973) and the rat (Phillips et al., 1974). However, slight differences exist between the Japanese quail and mammals. Although one kind of clustered cilia was found in the quail SFO, two kinds (lined cilia and circular cilia) are distinguishable in the rat. The number ofependymal cells bearing a solitary cilium is much smaller in the rat than in the Japanese quail. The length of clustered cilia in the quail (about 7.3 ~tm) is comparable to that of clustered cilia and a solitary cilium in the rat (8-101ira) and the rabbit (6 Ixm). However, a solitary cilium of the quail (0.85-1.88~tm) is much shorter than the clustered cilia. The physiological significance of relatively abundant short solitary cilium in the quail SFO is unknown.
Recent data show convincingly that the SFO has a dipsogenic receptor function for AII. Direct application of AII to the SFO induces copious drinking almost immediately, and electrolytic lesioning of the SFO greatly attenuates the amount of water intake induced by both intravenous and intracranial injection of AII in the rat (Simpson and Routtenberg, 1973, 1975; Abdelaal et al., 1974) and in the Japanese quail (Takei, 1977a). Intraventricular injection of All also induces drinking in mammals (see Severs and Summy-Long, 1975) and in birds (Takei, 1977b; Schwob and Johnson, 1977). Considering the data mentioned above, a sensory function of the surface structures of the SFO may be surmised. If CSF-contacting neurons (Vigh and Vigh-Teichmann, 1973) exist in the SFO, they are the most convincing morphological candidate for the sensory function of the SFO. However, the presence of such neurons is doubtful in the Japanese quail. Moreover, no authors have so far reported CSF-contacting neurons in the mammalian SFO. Another possible candidate for this function is supraependymal nerve cells as reported in mammals (Dellmann and Simpson, 1975). However, supraependymal nerve cells were not observed in the quail SFO except for possible supraependymal axons.

Surface Fine Structure of the Quail Subfornical Organ 403
Since neurons in direct contact with the CSF were not observed in the quail SFO, intraventricular AII must have reached nerve cells located in the sub- ependymal layer of the quail SFO (Tsuneki et al., 1978). One possible route may be the diffusion of the peptide through intercellular spaces between ependymal cells which were occasionally observed in the present TEM study. Funnel-shaped hollows observed in the present SEM and TEM studies may facilitate the pene- tration of biologically active substance in the CSF to the SFO, since cilia or cilium- like structures were observed on the inner surface. Deep hollows lined with ependymal cells have also been described in the SFO of the dog (Andres, 1965), cat (Pfenninger, 1969), rabbit (Leonhardt and Lindemann, 1973) and rat (Dellmann and Simpson, 1975).
It is also probable that intraventricular Al l is taken up by ependymal cells and conveyed to the nerve cells in the subependymal layer of the quail SFO. In the SEM study, small pore-like structures were often observed on the rostral part of the body of the SFO. Judging from their size and shape, they may correspond to pinocytotic invaginations observed in the TEM study. Therefore, Al l in the CSF may be taken up by ependymal cells via pinocytosis. However, nothing is known concerning the further transport of AII from the ependymal cells to the sensory nerve cells.
Some structures observed in this study indicate a secretory function of the SFO. The large protrusions on the body of the SFO may discharge their contents into the ventricle, since a remnant after discharge and secretory material were both seen near the protrusion. Leonhardt and Lindemann (1973) also described similar large protrusions in the rabbit SFO. In the latter species, a TEM study showed that these protrusions originate from parenchymal neurons. In the Japanese quail, however, the origin of large protrusions could not be determined in the TEM study. Apart from these large protrusions, bulbous protrusions containing cytoplasmic organelles (Figs. 7 a, 11) may also represent secretion by ependymal cells into the ventricle. The functional significance of these types of ventricular secretion is at present unknown.
References
Abdelaal, A.E., Assaf, S.Y., Kucharczyk, J., Mogenson, GJ.: Effect of ablation of the subfornical organ on water intake elicited by systemically administered angiotensin-II. Canad. J. Physiol. Pharmacol. 52, 1217-1220 (1974)
Andres, K.H.: Der Feinbau des Subfornikalorgans vom Hund. Z. Zellforsch. 68, 445-473 (1965) Dellmann, H.-D., Simpson, J.B.: Comparative ultrastructure and function of the subfornical organ. In:
Brain endocrine interaction II. The ventricular system in neuroendocrine mechanisms (K.M. Knigge, D.E. Scott, H. Kobayashi and S. Ishii, eds.), pp. 166-189. Basel: Karger 1975
Dellmann, H.-D., Simpson, J.B.: Regional differences in the morphology of the rat subfornical organ. Brain Res. 116, 389-400 (1976)
Dempsey, E.W.: Fine-structure of the rat's intercolumnar tubercle and its adjacent ependyma and choroid plexus, with especial reference to the appearance of its sinusoidal vessels in experimental argyria. Exp. Neurol. 22, 568-589 (1968)
Leonhardt, H., Lindemann, B.: Surface morphology of the subfornical organ in the rabbit's brain. Z. Zellforsch. 146, 243-260 (1973)
Mikami, S.: Ultrastructure of the organum vasculosum of the lamina terminalis of the Japanese quail, Coturnix coturnixjaponica. Cell Tiss. Res. 172, 227-243 (1976)

404 Y. Takei et al.
Pfenninger, K.: Subfornikalorgan und Liquor cerebrospinalis. In: Zirkumventrikul~ire Organe und Liquor (G. Sterba, ed.), pp. 103-106. Jena: Fischer 1969
Phillips, M.I., Balhorn, L., Leavitt, M., Hoffman, W.: Scanning electron microscope study of the rat subfornical organ. Brain Res. 80, 95-110 (1974)
Rudert, H., Scbwink, A., Wetzstein, R.: Die Feinstruktur des Subfornikalorgans beim Kaninchen. Z. Zellforsch. 88, 145-179 (1968)
Schinko, I., Rohrschneider, I., Wetzstein, R.: Elektronenmikroskopische Untersuchungen am Sub- fornikalorgan der Maus. Z. Zellforsch. 123, 277-294 (1972)
Schwob, J.E., Johnson, A.K.: Angiotensin-induced dipsogenesis in domestic fowl (Gallus gallus). J. comp. physiol. Psycbol. 91, 182-188 (1977)
Scott, D.E., Paull, W.K., Dudley, G.K.: A comparative scanning electron microscopic analysis of the human cerebral ventricular system. Z. Zellforsch. 132, 203-215 (1972)
Severs, W.B., Summy-Long, J.: The role of angiotensin in thirst. Life Sci. 17, 1513-1526 (1975) Simpson, J.B., Routtenberg, A.: Subfornical organ: Site of drinking elicitation by angiotensin II. Science
151, 1172-1174 (1973) Simpson, J.B., Routtenberg, A.: Subfornical organ lesions reduce intravenous angiotensin-induced
drinking. Brain Res. 88, 154-161 (1975) Takei, Y.: The role of the subfornical organ in drinking induced by angiotensin in the Japanese quail,
Coturnix coturnixjaponica. Cell Tiss. Res. 185, 175-181 (1977a) Takei, Y.: Angiotensin and water intake in the Japanese quail (Coturnix coturnixjaponica). Gen. comp.
Endocrinol. 31, 364-372 (1977b) Tsuneki, K., Takei, Y., Kobayashi, H.: Parenchymal fine structure of the subfornical organ of the
Japanese quail, Coturnix coturnix japonica. Cell Tiss. Res. (in press, 1978) Vigh, B., Vigh-Teichmann, I.: Comparative ultrastructure of the cerebrospinal fluid-contacting
neurons. Int. Rev. Cytol. 35, 189-251 (1973) Weindl, A.: Neuroendocrine aspects of circumventricular organs. In: Frontiers in Neuroendocrinology
(W.F. Ganong, L. Martini, eds.), pp. 3-32. New York: Oxford University Press 1973
Accepted May 9, 1978
Note Added in Proof
After this paper was submitted, the following pertinent paper was published. Their findings are essentially similar to ours. Mikami, S., Asari, M.: Ultrastructure of the subfornical organ of the Japanese quail, Coturnix coturnix. Cell Tiss. Res. 188, 19-33 (1978)



![LQ WK H 3 UR F H V V R I ) H UWLOL]D WLR Q LQ 4 X D LO · quail Coturnix japonica. The fully developed sperm-associated body, which is about 100 µ m long, consisted of two parts;](https://img.pdfslide.net/doc/110x75/5e77d19ce5fef4461e3e0143/lq-wk-h-3-ur-f-h-v-v-r-i-h-uwlold-wlr-q-lq-4-x-d-lo-quail-coturnix-japonica.jpg)






![Heterodermia japonica 15620 - fschumm.bplaced.netfschumm.bplaced.net/Schumm_Flechtenbilder/Heterodermia japonica 15620.pdf934 Heterodermia japonica (Sato) Swinscow & Krog [15620],](https://img.pdfslide.net/doc/110x75/5e0ce4eb0f7a5004ee3e9e97/heterodermia-japonica-15620-japonica-15620pdf934-heterodermia-japonica-sato.jpg)

















