Embed Size (px)
Citation preview

Synthesis and hydrolysis of ATP by the mitochondrial ATP synthase1y2
M. TUENA DE G~MEZ-PUYOU, ORLANDO B. MARTINS, AND A. G~MEZ-PUYOU Instituto de Fisiologia Celular, Universidad Nacional Autbnoma de Mbico, Apartado Postal 70-600, 04510 Mkxico DF,
Mexico
Received December 4 , 1987
TUENA DE G~MEZ-PUYOU, M., MARTINS, 0 . B., and G~MEZ-PUYOU, A. 1988. Synthesis and hydrolysis of ATP by the mitochondrial ATP synthase. Biochem. Cell Biol. 66: 677-682.
A brief summary of the factors that control synthesis and hydrolysis of ATP by the mitochondrial H+-ATP synthase is made. Particular emphasis is placed on the role of the natural ATPase inhibitor protein. It is clear from the existing data obtained with a number of agents that there is no correlation between variations of the rate of ATP hydrolysis and ATP synthesis as driven by respiration. The mechanism by which each condition differentially affects the two activities is not entirely known. For the case of the natural ATPase inhibitor protein, it appears that the protein controls the kinetics of the enzyme. This control seems essential for achieving maximal accumulation of ATP during electron transport in systems that contain relatively high concentrations of ATP.
TUENA DE G~MEZ-PUYOU, M., MARTINS, 0. B., et G~MEZ-PUYOLJ, A. 1988. Synthesis and hydrolysis of ATP by the mitochondrial ATP synthase. Biochem. Cell Biol. 66 : 677-682.
Nous discutons britvement des facteurs qui contr6lent la synthtse et I'hydrolyse de 1'ATP par la H+-ATP synthase mitochondriale. Nous insistons davantage sur le r6le de la protCine inhibitrice naturelle de 1'ATPase. Les donnCes obtenues avec un certain nombre d'agents dtmontrent qu'il n'existe aucune corrklation entre les variations du taux d'hydrolyse de I'ATP et du taux de synthtse de I'ATP occasionnCes par la respiration. Le mtcanisme par lequel chaque condition affecte diffkremment les deux activitts n'est pas entitrement connu. Pour ce qui est de la prottine inhibitrice naturelle de I'ATPase, elle semble contrder la cindtique de I'enzyme. Ce contr6le serait essentiel pour obtenir l'accumulation maximale d'ATP durant le transport des Clectrons dans les systtmes qui renferment des concentrations relativement ClevCes d' ATP.
[Traduit par la revue]
Introduction Electron transfer through the components of the
respiratory chain of the inner membrane of mitochondria results in the vectorial translocation of H + from the matrix space to the outer water space. This creates an electro- chemical H+ gradient that is used to drive the synthesis of ATP from ADP and inorganic phosphate; the electro- chemical H+ gradient may also be utilized for ion translocation and for the transfer of proteins to the mitochondrial matrix. Synthesis of ATP is catalyzed by a multisubunit enzyme known as the H+-ATP synthase. This enzyme traverses the inner membrane of the mitochondria and couples the inward movement of H + with the synthesis of ATP; under some conditions, the
ABBREVIATIONS: DCCD, dicyclohexylcarbodiimide; OSCP, oligomycin sensitivity-confemng protein; K,, Michaelis constant.
'This research was presented at the annual meeting of the CFBS on June 24, 1987, as part of a symposium entitled ATPases. This symposium, chaired by Dr. David H. MacLen- nan, was co-sponsored by the Canadian Biochemical Society and PAABS. The symposium was dedicated to Dr. Jean Mannery-Fisher.
he data of the authors described in this article were obtained through the support of the Consejo Nacional de Ciencia y Tecnologia, Mtxico and the Organization of American States. Pnnted an Canada 1 ImprimC au Canada
enzyme may also establish an electrochemical H+ gradient through the hydrolysis of ATP. The same basic mechanism operates in the ATP synthases of photosyn- thetic membranes and the plasma membrane of bacteria. While excellent reviews have been published in recent years on the structural and catalytic properties of ATP synthases from various sources (Fillingame 1980; Cross 1981; Senior and Wise 1983; Hatefi 1985), the purpose of this article is to describe briefly how the hydrolytic activity of the enzyme relates to the synthesis of ATP.
Structure of the ATP synthase The H+-ATP synthase of energy-transducing mem-
branes may be classed among the membrane enzymes in which ATP synthesis and hydrolysis are coupled to the movement of cations across the membrane. However, the H+-ATP synthases are structurally much more complex than the other transport ATPases, in the sense that the former are composed of a significantly greater number of different subunits (Hatefi 1985; Pedersen and Carafoli 1987). It is also noteworthy that the H+-ATP synthases of membranes of mitochondria, chloroplasts, and bacteria are strikingly similar in their basic struc- ture. All of the H+-ATP synthases contain a sector that is embedded in the membrane and that accounts for the transport of H+ across the membrane (Fig. 1). In
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.

678 BIOCHEM. CELL BIOL. VOL. 66. 1988
Subunit b
( DCCD-binding
protein
FIG. 1. Scheme of the structure of the mitochondrial ATP synthase. Fo is a sector of the enzyme that is embedded in the membrane. F, is a component that can be separated from the membrane as a water soluble protein; it is the factor where catalysis takes place.
addition, all ATP synthases contain a moiety known as Fl, which is where catalysis takes place (Fig. 1).
The Fo portion is composed of various different subunits whose number of stoichiometry are not yet clearly defined. In the H+-ATP synthase of Escherichia coli, Fo is formed by three different subunits called a, b, and c. Subunit c is also referred to as the DCCD-binding proteolipid, since it reacts with DCCD yielding a catalytically inserted enzyme. The number of copies of subunit c per complex is six, but a higher number has been described (Sebald et al. 1979; Foster and Filling- ame 1982).
The Ft moiety is very similar in d l ATP synthases. regardless of their biological source. It can readily be separated from the membrane and purified to a high degree of homogeneity {Knowles and Penefsky 1972; Tuena de G6mer-Puyou and G6rnez-Puyou 1977). In the soluble state i t exhibits a high ATPase activity, but under defined conditions it may catalyze the synthesis of enzyme-bound ATP (Sakamoto and Tonomun 1983; Sakamoto 1984; Yoshida 1983; G6mez-Puyou et al. 1986). It is composed of five different subunits in a stoichiometry of a3, P3, ?,a, and E that have molecular weights of 54 000,50 000,31000, 19 000, and 15 000, respectively. Subunits 6 and E appears to be required for the binding of FI to the membrane, and there is good evidence to indicate that subunit p contains the catalytic site for ATP synthesis and hydrolysis. Because there are three p subunits per ATP synthase, the enzyme would possess three catalytic sites.
The mitochondrial ATP synthase contains at least three other proteins. These are F6, and Factor B (McLennan and Tzagoloff 1968; Knowles et al. 1971; Sanadi 1982). These proteins are devoid of catalytic properties and are thought to be involved in the proper attachment of Fl to F,. It is likely that the function of Factor B is to induce a tight coupling of H+ flow to F1 (Sanadi 1982).
Finally, the mitochondria1 ATP synthase possesses a low molecular-weight protein that may be detached from the enzyme (for a recent review, see Schwerzman and Pedersen 1986). It was first isolated by Pullman and Monroy (1963) as a powerful inhibitor of hydrolysis; thus, it was named the natural ATPase inhibitor protein. Proteins with similar properties have been isolated from a wide variety of energy-transducing membranes that possess ATP synthases (Nelson et al. 1972; Cintron and Pedersen 1979; Dreyfus and Satre 1984; Satre et al. 1975; Ebner and Maier 1977). In mitochondria, the inhibitor protein exists in a stoichiometry of 1 : 1 with the ATP synthase, and it is known to bind to one of the 6 subunits (Klein et al. 1980). However, it has been reported that in yeast mitochondria, the binding of the inhibitor protein to the enzyme requires other proteins (Hashimoto et al. 1983). The existing evidence indicates that the inhibitor protein controls the kinetics of the ATP synthase (see Tuena de G6mez-Puyou et al. 1983; also see below).
Synthesis and hydrolysis of ATP in submitochondrial particles
It is difficult to study the structure and mechanism of action of the ATP synthase in whole mitochondria, as the Fl portion and the proteins involved in its attachment to Fn are localized in the inner side of the internal - membrane. Moreover, the kinetics of the enzyme in whole mitochondria are difficult to evaluate, because in this system the synthesis and hydrolysis of ATP involve the movement of ATP, ADP, and phosphate across the membrane through specific carriers (Heldt et al. 1972; Lemasters and Sowers 1979). In fact. the translocator of adenine nucleotides seems to be the rate-limiting step of oxidative phosphorylation (Moreno-Sanchez 1985). Thus, most of the studies on the kinetics and structure of the enzyme have been carried out in submitochondrial particles or in the isolated enzyme and its separate com- ponents (Fo and F Since submitochondrial particles are "inside-out" membrane preparations that possess a full catalytic capacity for hydrolysis and synthesis of ATP, there are no permeability barriers for ATP or ADP.
~ubmitochondrial are generally prepared by mechanical disruption of the mitochondria, which in most cases is achieved by sonication. This is followed by fractional centrifugation. The ATPase activity of the resulting preparation may show wide variations; it may range from approximately 0.4 ymol. min- . mg-' to about 10 ymol-min-'.mg-'. The level of the hydrolyt- ic activity largely depends on the composition of the media in which the mitochondria were disrupted, as different conditions may or may not favor the dissocia- tion of the inhibitor protein from the enzyme. Indeed, a correlation has been found between specific activity of hydrolysis and the extent of interaction between- the
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.
3 6 9 12 15 18
Time, min
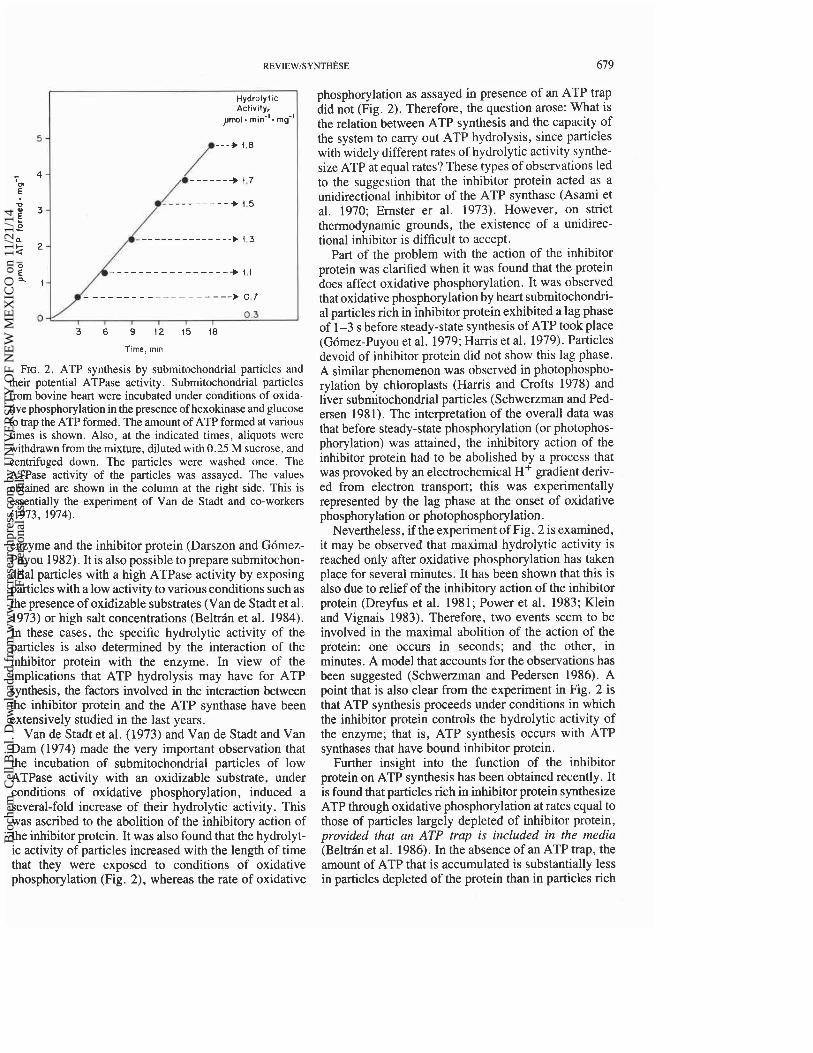
FIG. 2. ATP synthesis by submitochondrial particles and their potential ATPase activity. Submitochondrial particles from bovine heart were incubated under conditions of oxida- tive phosphorylation in the presence of hexokinase and glucose to trap the ATP formed. The amount of ATP formed at various times is shown. Also, at the indicated times, aliquots were withdrawn from the mixture, diluted with 0.25 M sucrose, and centrifuged down. The particles were washed once. The ATPase activity of the particles was assayed. The values obtained are shown in the column at the right side. This is essentially the experiment of Van de Stadt and co-workers (1973, 1974).
enzyme and the inhibitor protein (Darszon and G6mez- Puyou 1982). It is also possible to prepare submitochon- drial particles with a high ATPase activity by exposing particles with a low activity to various conditions such as the presence of oxidizable substrates (Van de Stadt et al. 1973) or high salt concentrations (BeltrBn et al. 1984). In these cases, the specific hydrolytic activity of the particles is also determined by the interaction of the inhibitor protein with the enzyme. In view of the implications that ATP hydrolysis may have for ATP synthesis, the factors involved in the interaction between the inhibitor protein and the ATP synthase have been extensively studied in the last years.
Van de Stadt et al. (1973) and Van de Stadt and Van Dam (1974) made the very important observation that the incubation of submitochondrial particles of low ATPase activity with an oxidizable substrate, under conditions of oxidative phosphorylation, induced a several-fold increase of their hydrolytic activity. This was ascribed to the abolition of the inhibitory action of the inhibitor protein. It was also found that the hydrolyt- ic activity of particles increased with the length of time that they were exposed to conditions of oxidative phosphorylation (Fig. 2), whereas the rate of oxidative
phosphorylation as assayed in presence of an ATP trap did not (Fig. 2). Therefore, the question arose: What is the relation between ATP synthesis and the capacity of the system to carry out ATP hydrolysis, since particles with widely different rates of hydrolytic activity synthe- size ATP at equal rates? These types of observations led to the suggestion that the inhibitor protein acted as a unidirectional inhibitor of the ATP synthase (Asami et al. 1970; Ernster er al. 1973). However, on strict thermodynamic grounds, the existence of a unidirec- tional inhibitor is difficult to accept.
Part of the problem with the action of the inhibitor protein was clarified when it was found that the protein does affect oxidative phosphorylation. It was observed that oxidative phosphorylation by heart submitochondri- a1 particles rich in inhibitor protein exhibited a lag phase of 1-3 s before steady-state synthesis of ATP took place (G6mez-Puyou et al. 1979; Harris et a1. 1979). Particles devoid of inhibitor protein did not show this lag phase. A similar phenomenon was observed in photophospho- rylation by chloroplasts (Harris and Crofts 1978) and liver submitochondrial particles (Schwerzman and Ped- ersen 1981). The interpretation of the overall data was that before steady-state phosphorylation (or photophos- phorylation) was attained, the inhibitory action of the inhibitor protein had to be abolished by a process that was provoked by an electrochemical H+ gradient deriv- ed from electron transport; this was experimentally represented by the lag phase at the onset of oxidative phosphorylation or photophosphorylation.
Nevertheless, if the experiment of Fig. 2 is examined, it may be observed that maximal hydrolytic activity is reached only after oxidative phosphorylation has taken place for several minutes. It has been shown that this is also due to relief of the inhibitory action of the inhibitor protein (Dreyfus et al. 1981; Power et al. 1983; Klein and Vignais 1983). Therefore, two events seem to be involved in the maximal abolition of the action of the protein: one occurs in seconds; and the other, in minutes. A model that accounts for the observations has been suggested (Schwerzman and Pedersen 1986). A point that is also clear from the experiment in Fig. 2 is that ATP synthesis proceeds under conditions in which the inhibitor protein controls the hydrolytic activity of the enzyme; that is, ATP synthesis occurs with ATP synthases that have bound inhibitor protein.
Further insight into the function of the inhibitor protein on ATP synthesis has been obtained recently. It is found that particles rich in inhibitor protein synthesize ATP through oxidative phosphorylation at rates equal to those of particles largely depleted of inhibitor protein, provided that an ATP trap is included in the media (BeltrBn et al. 1986). In the absence of an ATP trap, the amount of ATP that is accumulated is substantially less in particles depleted of the protein than in particles rich
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.

680 BIOCHEM. CELL BIOL. VOL. 66. 1988
in the inhibitor protein (Beltrin et al. 1986). This indicates that in the absence of control by the inhibitor protein, part of the ATP that is formed through oxidative phosphorylation is hydrolyzed with a resulting low accumulation of ATP. In light of these findings, the inhibitor protein would appear to be an essential component for attaining maximal conservation of ATP formed from the energy of electron transport. If one considers that mitochondria contain (1-3) X loF3 M ATP (Slater et al. 1973; Letko et al. 1980) under conditions in which steady-state oxidative phosphoryla- tion is taking place, the importance of a mechanism that arrests ATP hydrolysis becomes obvious. As is evident from data in the literature, the function of the inhibitor protein is to modify the kinetics of the ATP synthase so that the rate at which ATP is hydrolyzed is diminished (Tuena de Gornez-Puyou et al. 1983).
Nevertheless, some recent data in the literature indicate that the function of the inhibitor may be more complex. It has been reported by Guerrieri et al. (1987) that particles devoid of inhibitor protein exhibit a rapid flow of H+ through the Fo portion of the ATP synthase; reconstitution of these particles via inhibitor protein significantly diminishes the rate of H+ flow. This suggests that modifications of F1 by its interaction with the protein reflect on the permeability properties of the membrane sector.
Regardless of the extensive studies that have been made on the relation between ATP hydrolysis and synthesis, there are still many important questions to be answered. There is a significant number of reports that show that certain compounds such as organotins (Emanuel et al. 1984; Van der Bend 1985) and adenine nucleotide analogs (Schafer 1982) provide strong inhibi- tion of hydrolytic activity without affecting ATP synthe- sis driven by electron transport to the same extent. To account for these observations, researchers have sug- gested that there are separate sites for ATP synthesis and hydrolysis (Emanuel et al. 1984), a larger affinity for the inhibitor under nonenergized rather than energized conditions (Schafer 1982), different mechanisms for coupling ATP synthesis and hydrolysis, and variations in enzyme kinetics (Matsuno-Yagi and Hatefi 1986). This larger number of different proposals indicates that there is uncertainty as to exactly how ATP synthesis relates to the hydrolytic activity of submitochondrial particles.
Synthesis and hydrolysis of ATP by a soluble F1- inhibitor protein complex
Central to the aforementioned mechanism of action of the inhibitor protein is the F1 complex with inhibitor protein; this possesses catalytic activity. In this respect, data obtained with soluble F1 containing bound inhibitor protein indicate that the complex is indeed catalytically
active. Feinstein and Moudrianakis (1984) isolated a soluble F1 preparation that contained bound inhibitor protein and that had a very low ATPase activity. Under conditions that caused release of the protein, the hydrolytic activity of the preparation increased to values similar to those observed with F1 free of inhibitor protein. The possible existence of catalytic activity in the soluble F1 - inhibitor protein complex was explored. It has been demonstrated that soluble F1 can catalyze the synthesis of ATP from medium phosphate and enzyme- bound ADP, provided that the system is supplemented with M ~ ~ + and an organic cosolvent such as dimethyl sulfoxide, whose function is to alter the structure of water (Sakamoto and Tonomura 1983; Sakarnoto, 1984; Yoshida 1983; G6mez-Puyou et al. 1986). ATP that is formed under these conditions is not released into the medium but remains bound to the enzyme. The amount of ATP formed is approximately 0.4 mol per mol of F1.
With respect to the effect of the inhibitor protein on this catalytic property of F1, it has been found that the F1 -inhibitor protein complex synthesizes ATP to the same extent as F, , and at almost the same rate (G6mez-Puyou et al. 1986). Thus, the complex has this catalytic capacity. Other experiments have shown that this ATP in the F, - inhibitor protein complex could undergo hydrolysis when the complex containing synthesized ATP was transferred to an all-water medium.
These findings are relevant to the mechanism of ATP synthesis. The results have been interpreted by assum- ing that the catalytic site of F1 (as well as that of other energy-transducing ATPases) is hydrophobic. In such an environment, ATP is synthesized without energy. The basis of this postulate is that the equilibrium constant of pyrophosphate hydrolysis is several orders of magnitude greater in all-water media than in media where the structure of the water has been modified by organic cosolvent (de Meis 1984). Also, it has been found that the equilibrium constant of ATP hydrolysis at the catalytic site of F1 approaches unity (Grubmeyer et al. 1982). Therefore, in an all-water medium, the barrier for the synthesis of ATP would be the partitioning of the phosphate between the medium and the catalytic site. The role of the cosolvent would be to favor the partitioning of the medium phosphate into the catalytic site; once phosphate is in the catalytic site, ATP synthesis would occur spontaneously. In fact, it has been reported that the K,,, for phosphate in the spontane- ous synthesis of ATP is higher than 1.00 X lop3 M in an all-water medium, whereas inthe presence of dimethyl sulfoxide it is 1 X M (Sakamoto 1984; G6mez- Puyou et al. 1986). Similar results had been obtained previously for the phosphorylation of the Ca2+-ATPase of sarcoplasmic reticulum by medium phosphate (de Meis et al. 1980).
The overall data obtained about submitochondrial
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.

particles and the soluble enzyme indicate that the soluble and particulate F, - inhibitor protein complex is catalyti- cally active. This may imply that synthesis of ATP during oxidative phosphorylation in complete mito- chondria is catalyzed by an enzyme that has bound inhibitor protein. This is of particular importance, since in intact mitochondria, synthesis of ATP occurs in the matrix space that has a concentration of ATP in the millimolar range. In particles devoid of inhibitor pro- tein, it has been observed that at these concentrations of ATP, net formation of ATP is much lower than in particles controlled by the inhibitor protein (Beltrhn et al. 1986). Therefore, it would seem that the inhibitor protein, through its regulation of the kinetics of the ATP synthase, is an essential component for achieving maximal synthesis of ATP in systems that contain high concentrations of ATP.
ASAMI, K., JUNTTI, K., and ERNSTER, L. 1970. Possible regulatory function of a mitochondrial ATPase inhibition in respiratory chain linked phosphorylation. Biochim. Biophys. Acta, 205: 307-31 1.
BELTRAN, C., TUENA DE G~MEZ-PUYOU, M., G ~ M E Z - Puyou, A., and DARSZON, A. 1984. Release of the inhibitory action of the natural ATPase inhibitor protein in mitochondrial ATPase. Eur. J . Biochem. 144: 15 1- 157.
BELTRAN, C., TUENA DE G~MEZ-PUYOU, M., DARSZON, A., and G~MEZ- YOU, A. 1986. Simultaneous synthesis and hydrolysis of ATP regulated by the inhibitor protein. Eur. J. Biochem. 160: 163- 168.
CINTRON, N. W., and PEDERSEN, P. L. 1979. A protein inhibitor of the mitochondrial adenosine triphosphatase complex of rat liver. Purification and characterization. J. Biol. Chem. 254: 3439-3443.
CROSS, R. L. 1981. The mechanism and regulation of ATP synthesis by FI-ATPases. Annu. Rev. Biochem. 50: 681- -. . 114.
DARSZON, A., and G~MEZ-PIJYOU, A. 1982. Extraction of mitochondrial protein-lipid complexes into organic sol- vents: An approach to the study of the interaction of the ATPase inhibitor with the ATPase. Eur. J. Biochem. 121: 427-433.
DE MEIS, L. 1984. Pyrophosphate of high and low energy. Contributions of pH, Caz+ and water to free energy of hydrolysis. J. Biol. Chem. 259: 6090-6097.
DE MEIS, L., MARTINS, 0. B., and ALVES, E. W. 1980. Role of water, hydrogen ion, and temperature on the synthesis of adenosine triphosphate by sarcoplasmic reticulum adeno- sine triphosphatase in the absence of a calcium ion gradient. Biochemistry, 19: 4252-4261.
DREYFUS, G., and SATRE, M. 1984. The subunit E as an ATPase inhibitor of the F1-ATPase in Escherichia coli. Arch. Biochem. Biophys. 229: 212-219.
DREYEUS, G., G~MEZ-PUYOU, A., and TUENA DE G ~ M E Z - PUYOU, M. 1981. Electrochemical gradient induced dis- placement of the natural ATPase inhibitor as detected by antibodies against the inhibitor protein. Biochem. Biophys. Res. Cornrnun. 100: 400-407.
EBNER, E., and MAIER, K. L. 1977. A protein inhibitor of mitochondrial adenosine triphosphatase (F1) from Sacharo- myces cerevisiae. J . Biol. Chem. 252: 671-676.
EMANUEL, E. L., CARVER, M. A., SOLANI, G. C., and GRIFFITHS, D. E. 1984. Differential inhibition of FoFI- ATPase catalyzed reactions in bovine-heart submitochon- drial particles by organotin compounds. Biochim. Biophys. Acta, 766: 209-214.
ERNSTER, L., JUNTTI, K., and ASAMI, K. 1973. Mechanisms of energy conservation in the mitochondrial membrane. J. Bioenerg. 4: 149- 159.
FEINSTEIN, D. L., and MOUDRIANAKIS, E. N. 1984. Response of the adenosine triphosphatase activity of the soluble latent FI enzyme from beef heart mitochondria to changes in ~ g ' + and H+ concentrations. J. Bio. Chem. 259: 4230-4236.
FILLINGAME, R. H. 1980. Biochemistry and genetics of bacterial H+-translocating ATPases. Curr. Top. Bioenerg., 11: 35-106.
FOSTER, D. L., and FILLINGAME, R. H. 1982. Stoichiometry of subunits in the Hf -ATPase complex of Escherichia coli. J. Biol. Chem. 257: 2009-2015.
G~MEZ-PUYOU, A., TUENA DE G~MEZ-PUYOU, M., and ERNSTER, L. 1979. Inactive to active transitions of the mitochondrial ATPase as controlled by the ATPase inhibi- tor. Biochim. Biophys. Acta, 547: 252-257.
G~MEZ-PUYOU, A,, TUENA DE G~MEZ-PUYOU, M., and DE
MEIS, L. 1986. Synthesis of ATP by soluble mitochondrial FI-ATPase and F1-inhibitor complex in the presence of organic solvents. Eur. J. Biochem. 159: 133-140.
GRUBMEYER, C., CROSS, R. L., and PENEFSKY, H. S. 1982. Mechanism of ATP hydrolysis of beef heart mitochondrial ATPase. J. Biol. Chem. 257: 12 092 - 12 100.
GUERRIERI, F., SCAREO, R., ZANOTTI, F., CHE, Y. W., and PAPA, S. 1987. Regulatory role of the ATPase inhibitor protein on proton conduction by mitochondrial H+-ATPase complex. FEBS Lett. 213: 67-72.
HARRIS, D. A., and CROFTS, A. R. 1978. The initial stages of photophosphorylation studies using excitation. Biochim. Bioohvs. Acta. 502: 87-102. . .
HARRIS, D. A., 'VON TSCHARNER, V., and RADDA, G. K. 1979. The ATPase inhibitor protein in oxidative phosphoryl- ation. The rate limiting factor to phosporylation in submito- chondrial particles. Biochim. Biophys. Acta, 548: 72-84.
HASHIMOTO, T., YOSHIDA, Y., and TAGAWA, K. 1983. Binding properties of an intrinsic ATPase inhibitor and occurrence in yeast mitochondria of a protein factor which stabilizes and facilitates the binding of the inhibitor to FI-Fo ATPase. J. Biochem. (Tokyo), 94: 715-720.
HATEEI, Y. 1985. The mitochondrial electron transport and oxidative phosphorylation. Annu. Rev. Biochem. 54: 1015-1069.
HELDT, H. W., KLINGENBERG, M., and MILOVANCEV, M. 1972. Differences between the ATP/ADP ratios in the mitochondrial matrix and in the extramitochondrial space. Eur. J. Biochem. 30: 434-440.
KLEIN, G., and VIGNAIS, P. V. 1983. Effect of the proton motive force on ATP-linked processes and mobilization of the bound natural ATPase inhibitor in beef heart submito- chondrial particles. J. Bioenerg. Biomembr. 15: 347-362.
KLEIN, G., SATRE, M., DIANOUX, A. C., and VIGNAIS, P. V.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.

682 BIOCHEM. CELL BIOL. VOL. 66. 1988
1980. Radiolabeling of natural adenosine triphosphatase inhibitor with phenyl I4c isothiocyanate and study of its interaction with rnitochondrial adenosine triphosphatase. Localization of inhibitor binding sites and stoichiometry of binding. Biochemistry, 19: 2919-2925.
KNOWLES, A. F., and PENEFSKY, H. S. 1972. The subunit structure of beef mitochondrial adenosine triphosphatase. Isolation procedures. J. Biol. Chem. 247: 6617-6623.
KNOWLES, A. F., GUILLORY, R., and RACKER, E. 1971. Partial resolution of the enzymes catalyzing oxidation phosphorylation. J. Biol. Chem. 246: 2672-2679.
LEMASTERS, J. J. , and SOWERS, A. E. 1979. Phosphate dependence and atractyloside inhibition of mitochondrial oxidative phosphorylation. The ADP-ATP carrier is rate limiting. J. Biol. Chem. 254: 1248-1251.
LETKO, G., KUSTER, U. , DUSLINSKI, J. , and KUNZ, W. 1980. Investigation of the dependence of the intramitochondrial ATP/ADP ratio on the respiration rate. Biochim. Biophys. Acta, 593: 196-203.
MATSUNO-YAGI, A., and HATEFI, Y. 1986. Kinetic modal- ities of ATP synthesis regulation by the mitochondrial respiratory chain. J . Biol. Chem. 261: 14031 - 14038.
MCLENNAN, D. H., and TZAGOLOFF, A. 1968. Studies of the mitochondrial adenosine triphosphatase system IV. Purifi- cation and characterization of the oligomycin. Biochemistry, 7: 1603-1610.
MORENO-SANCHEZ, A. 1985. Contribution of the translocator of adenine nucleotides and ATP synthase to the control of oxidative phosphorylation and arsenylation in liver mito- chondria. J . Biol. Chem. 260: 12 554 - 12 560.
NELSON, N., NELSON, H., and RACKER, E. 1972. Partial resolution of the enzymes catalyzing photophosphorylation. Purification and properties of an inhibitor isolated from chloroplast coupling factor 1. J. Biol. Chem. 247: 7657-7662.
PEDERSEN, P. L., and CARAFOLI, E. 1987. Ion motive ATPases I ubiquity, properties, and significance to cell function. Trends Biochem. Sci. 12: 146- 150.
POWER, R., CROSS, R. L., and HARRIS, D. A. 1983. Interaction of F1-ATPase from heart mitochondria with its naturally occurring inhibitor protein. Studies using ratio- iodinated inhibitor protein. Biochim. Biophys. Acta, 724: 128-141.
PULLMAN, M. E., and MONROY, G. C. 1963. A naturally occurring inhibitor of rnitochondrial adenosine triphospha- tase. J. Biol. Chem. 238: 3762-3769.
SAKAMOTO, J . 1984. Identification of the nucleotide-binding site for ATP synthesis and hydrolysis in mitochondrial soluble Fl-ATPase. J. Biochem. (Tokyo), 96: 475-48 1.
SAKAMOTO, J., and TONOMURA, Y. 1983. Synthesis of enzyme bound ATP by mitochondrial soluble F1-ATPase in the presence of dimethyl sulfoxide. J. Biochem. (Tokyo), 93: 1601-1614.
SANADI, D. R. 1982. Mitochondria1 coupling factor B. Properties and role in ATP synthesis. Biochim. Biophy s. Acta, 683: 39-56.
SATRE, M., DE JERPHANION, M. B., HUET, J., and VIGNAIS, P. V. 1975. ATPase inhibitor from yeast mitochondria purification and properties. Biochim. Biophys. Acta, 387: 241-255.
SCHAFER, G. 1982. Differentiation of two states of F1-ATPase by nucleotide analogs. FEBS Lett. 139: 271-275.
SCHWERZMAN, K . , and PEDERSEN, P. L. 1981. Proton- adenosine-triphosphatase complex of rat liver mitochon- dria. Effect of energy state on its interaction with the adenosine triphosphatase inhibitor peptide. Biochemistry, 20: 6305-63 11.
1986. Regulation of the mitochondrial ATP synthasel ATPase complex. Arch. Biochem. Biophys. 250: 1-8.
SEBALD, W., GRAF, T. , and LUKINS, H. B. 1979. The dicyclohexylcarbodiimide-binding protein of the mitochon- drial ATPase complex from Neurospora crassa and Sacha- romyces cerevisiae. Eur. J . Biochem. 93: 587-599.
SENIOR, A. E., and WISE, J. G. 1983. The proton ATPase of bacteria and mitochondria. J. Membr. Biol. 73: 105-124.
SLATER, E. C., ROSING, J., and H o t , A. 1973. The phospho- rylation potential generated by respiring mitochondria. Biochim. Biophys. Acta, 292: 534-553.
TUENA DE G~MEZ-PUYOU, M., and G ~ M E Z - h y o u , A. 1977. A simple method of purification of a soluble oligomycin insensitive mitochondria ATPase. Arch. Biochem. Biophys. 182: 82-86.
TUENA DE G~MEZ-PUYOU, M., MULLER, U . , DREYFUS, G., AYALA, G., and G~MEZ-PUYOU, A. 1983. Regulation of the synthesis and hydrolysis of ATP by mitochondrial ATPase. Role of M ~ " . J. Biol. Chem. 258: 13 680 - 13 684.
VAN DER BEND, R. L., DUETZ, W., COLEN, A. M. A. F., VAN DAM, K., and BERDEN, J. A. 1985. Differentialeffects of triphenyltin and 8-azido ATP on the ATP synthesis ATP-P, exchange and ATP hydrolysis in liposomes con- taining ATP synthase and bacteriorhodopsin. Arch. Bio- chem. Biophys. 241: 461-471.
VAN DE STADT, R. J., and VAN DAM, K. 1974. The equilibrium between mitochondrial ATPase (F1) and its natural inhibitor in submitochondrial particles. Biochim. Biophys. Acta, 347: 240-252.
VAN DE STADT, R. J., DE BOER, B. L., and VAN DAM, K. 1973. The interaction between the rnitochondrial ATPase (F1) and the ATPase inhibitor. Biochim. Biophys. Acta, 292: 338-349.
YOSHIDA, M. 1983. The synthesis of enzyme bound ATP by the F1-ATPase from the thermophilic bacterium PS3 in 50% dimethyl sulfoxide. Biochem. Biophys. Res. Comrnun. 114: 907-912.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
ER
SIT
Y O
F N
EW
ME
XIC
O o
n 11
/21/
14Fo
r pe
rson
al u
se o
nly.







![ATP Synthase Subunit a Supports Permeability Transition in ...Mitochondrial ATP synthase, an enzyme that provides cellular energy in the form of ATP, is composed of 17 subunits [1]](https://img.pdfslide.net/doc/110x75/5f101bf57e708231d4477d9e/atp-synthase-subunit-a-supports-permeability-transition-in-mitochondrial-atp.jpg)





















