Embed Size (px)
Citation preview

T.C.MUSTAFA KEMAL ÜNİVERSİTESİ
FEN-EDEBİYAT FAKÜLTESİBİYOLOJİ BÖLÜMÜ
B. 353 GENETİK I LABORATUVAR KILAVUZU
DROSOPHİLA GENETİĞİ
HAZIRLAYANLARProf. Dr. Mahmut ÇALIŞKAN
Yrd. Doç. Dr. Ayşe YAVUZ KOCAMAN
HATAY-2002

DROSOPHİLA GENETİĞİ
Drosophila melanogaster, genetiğin temel olaylarını uygulamalı olarak göstermek için oldukça uygun bir organizmadır. Genellikle meyve sineği ya da sirke sineği olarak bilinen D. melanogaster Thomas Hunt Morgan'ın çalışmalarıyla 1909 yılından beri genetik çalışmalarında kullanılmaktadır. Günümüzde kalıtımla ilgili bilnenlerin çoğu bu canlılar üzerinde yapılan çalışmalarla kazanılmıştır. Morgan ve öğrencileri Mendel kalıtımının mekanizmasını anlamak ve ilk linkaj (bağlantı) haritasını yapmak için Drosophila'yı kullanmışlardır.
Sitogenetikçiler kromozom morofolojisi ve karyotip çalışmak için; populasyon genetikçileri emrimsel çalışmalar ve seleksiyon deneyleri için Drosophila'nın kullanışlı bir organizma olduğunu bulmuşlardır. Ayrıca D. melanogaster Genetik Mühendisliğinde ve gelişimin moleküler temelinin anlaşılmasında kullanılan ilk ökaryotik organizmalardan biridir. Bu canlının genetik çalışmalarda sıklıkla kullanılmasının nedenlerini şöyle sıralayabiliriz:1. Laboratuvarda yaşatılmaları kolay ve ekonomiktir.2. Uygun büyüklükte olduklarından fazla yer işgal etmezler.3. Generasyon süreleri kısa olup (25°C'de 10 gün) çok sayıda yavru meydana getirirler.4. Kolayca incelenebilen ve ayırt edilebilen dört çift kromozoma sahiptirler (2n=8).5. Larvalarının tükrük bezi hücrelerinde gözlenebilen dev kromozomların bulunuşu sitogenetik çalışmalar için ideal yapılardır.6. Kontrollü eşleştirmeler yapılabilir.7. Çok sayıda genetik varyanta sahip olmaları kalıtımla ilgili problemlerin çözümü için elverişlidir.
Laboratuvar Çalışması için Gerekli Ekipman
Stereo Mikroskop: Drosophila'nın morfolojik özelliklerinin belirlenmesinde, eşey ayırımının yapılmasında vs. gereklidir.
Binoküler mikroskop: Tükrük bezi preparasyonlarında dev kromozomların gözlenmesi çalışmalarında gereklidir.
Anestetizer: Temel olarak küçük bir şişe ve bayıltma işlemi için eter gereklidir.
Morg: Ölü sineklerin içine konulduğu, %70'lik alkol ya da deterjan ile hazırlanan şişelerdir.
Beyaz Fayans: Eterizasyon işlemi uygulanılan sinekler, tasnif işlemleri için beyaz bir zemin üzerine alınarak incelenirler. Bunun için genellikle beyaz fayans kullanılır. Fakat cam levhanın altına beyaz kağıt konularak da aynı işlem yapılabilir.
Fırça: Çalışmalar sırasında sineklerin zarar görmesini engellemek için yumuşak kıllı suluboya fırçaları kullanılır.
İnkübatör (etüv): Sineklerin yetiştirilmesi için 24 °C sabit sıcaklığa ayarlı inkübatör gereklidir.

Genetik Terminoloji
Alel: Bir genin alternatif formlarından biridir.
Diploit: Hücre çekirdeğinde herbir kromozom tipinin iki kopyasının bulunmasıdır. Böyle kromozom takımına diploit ya da 2N denir. Sirke sinekleri diploittir ve 4 çift kromozom taşırlar.
Dominant/resesif: Diploid organizmalarda bir alel fenotipik olarak hem homozigot hem de heterozigot durumlarda ifade edilebiliyorsa dominanttır (baskın). Alel sadece homozigot durumlarda fenotipik özelliğini ortaya koyabiliyorsa resesif (çekinik) denilir.
Fenotip: Organizmanın gözlenebilir özellikleridir. Genotip ile çevrenin etkileşimiyle meydana gelir.
F1: Birinci oğul döl (First filial generation); bir deneysel çaprazlamadan elde edilen yavru döllerdir. F1'in atasoyu (ebeveyn=parent) P olarak gösterilir.
F2: F1 bireylerinin kendi aralarında çaprazlanması sonucu elde edilen yavru döller.
Gen: Gen, klasik anlamda bir kalıtım birimidir ve genellikle genomdaki spesifik bir kromozom üzerinde spesifik bir pozisyonda (lokus) bulunur; pratik olarak organizmanın fenotipi üzerine bir ya da daha fazla spesifik bir etkiye sahip olan bir birimdir; moleküler seviyede bir DNA molekülündeki nükleotidlerin spesifik bir dizilimidir.
Genom mutasyonu: Türü için normalden farklı sayıda kromozomlu bireyler ya da heteroploid hücreler meydana getiren kromozom sayısındaki değişimdir. Böyle bir değişim tüm kromozom takımlarını içerebilir (öploidi) ya da bir ya da daha fazla kromozomda olabilir (anöploidi).
Genotip: Bir hücrenin ya da organizmanın genetik bileşimidir.
Gonozom: Eşey kromozomlarıdır. Drosophila'da X ve Y kromozomları cinsiyeti belirler.
Haploid: N olarak sembolize edilen gametik kromozom sayısı.
Hemizigot: Genin tek bir alel taşıması durumudur. Haploid organizmadaki gen, ya da heterogametik cinsiyetteki cinsiyete bağlı gen olabilir. Örneğin erkek bireylerdeki X'e bağlı genlerin durumuna hemizigot denir. Çünkü bunlar Y kromozomu üzerinde hiçbir karşılık alele sahip değildir.
Heterogametik eşey: Eşey kromozomları bakımından benzer olmayan gametler meydana getiren cinsiyettir (Drosophila ve memelilerin erkek bireyleri X ve Y meydana getirirler).
Heterozigot: Homolog kromozomun karşılıklı lokuslarında farklı alellere sahip olma durumu.
Homogametik eşey: Sadece bir çeşit gamet meydana getiren cinsiyettir (Drosophila ve

memelilerin dişileri sadece X kromozomu taşıyan yumurta meydana getirirler).
Homolog kromozom: Şekil, yapı ve fonksiyon bakımından birbirine benzeyen ve biri anadan diğeri babadan gelen kromozomdur. Homolog kromozomlar genin aynı lineer dizilimini taşırlar ve her bir gen diploid bir organizmada bir çift oluşturur.
Homozigot: Homolog kromozomların karşılıklı lokuslarında benzer alellerin bulunmasıdır.
Kontrol çaprazlama: Bir genotipin homozigot mu yoksa heterozigot mu olduğunu anlamak için resesif homozigot bir genotiple çaprazlanması olayına denir.
Kromozomal mutasyon: Genetik bilgiyi taşıyan kromozomun duplikasyonu, delesyonu gibi birden fazla gen bölgesinin değişime uğraması.
Letal mutasyon: Organizmanın ölümüne neden olan mutasyondur. Dominant letal dominantları öldürürken homozigot letal sadece homozigotları öldürür.
Lokus: (çoğulu: loci) Bir genin kromozom üzerinde yer aldığı alan.
Mendel genetiği: Gregor Johann Mendel'in 1854'te başlayan bezelyelerle yaptığı çaprazlama çalışmalarıyla elde ettiği sonuçlar günümüzde genetiğin temelini oluşturmakta ve Mendel Genetiği olarak bilinmektedir. Mendel genetiğini oluşturan bu kanunları şunlardır:1. Dominantlık (İzotipi) kanunu: Aynı karakter bakımından fark gösteren iki homozigot bireyin eşleşmesi sonucu meydana gelen melez bireylerin genotipleri birbirinin aynıdır, yani bunlar izotoptur. Bu kanuna izotipi kanunu denir.2. Segregasyon (=Ayrılım) kanunu: Gamet oluşumu sırasında bir genin alelleri rastgele ayrılarak oluşan her gametin bunlardan birini alması sağlanır.3. Bağımsız açılım (=Independent assortment) kanunu: Farklı genlere ait alellerin eşey hücrelerinde (gametlerde) bir araya gelmeleri birbirinden bağımsızdır ve rastlantıya bağlıdır. Bu kural eğer genler birbirine bağlı genler değilse geçerlidir.
Mutasyon: Kalıtsal molekülde dölden döle aktarılabilen değişiklikler meydana getiren ani ve tesadüfi değişmelerdir.
Nokta (gen) mutasyon: Sadece bir geni etkileyen mutasyondur.
Otozom: Eşeylikle ilgisi olmayan, vücut kromozomlarıdır (insanda 44, Drosophila'da 6 tanedir).
Poliploidi: Bir bireyin ya da hücrenin ikiden fazla kromozom setine sahip olmasıdır.
Resiprokal çaprazlama: Herhangi bir karakter bakımından farklılık gösteren iki birey arasında yapılan karşılıklı çaprazlamadır. A ♀ x B ♂ ve B ♀ x A ♂ şeklindeki çaprazlamadır.

Genetik SembollerGenetik deneylerde kullanılan semboller genin normal ya da mutant olmasına göre değişmektedir. Yabanıl tip alel, doğal populasyonlarda en yaygın olarak bulunan, mutasyon meydana gelmeden önceki orijinal aleldir. D. melanogaster'in normal tipini ifade eden yabanıl tip + ile sembolize edilir. + sembolü tüm yabanıl tip genler için geçerlidir. Mutasyonlar için ise mutasyonu tanımlayan adın yazılışından uygun harfler türetilerek sembolize edilir. Örneğin, Drosophila 'da körelmiş kanadı meydana getiren mutasyon için vg (vestigial), kahverengi gözlü mutasyon için se (sepia), tüysüz mutasyon için ise H (hairless) kullanılır.
Resesif mutantlar küçük harfler ile vg yazılırken, dominant mutantlar kullanılan sembolün ilk harfinin büyük olanı ile H yazılır. Diploid organizmalarda homozigotlar sembolü iki kere yazarak vg/vg ya da bazen sembolü tek başına yazarak vg ile gösterilir. Yabanıl tip homozigotlar da +/+ veya sadece + ile belirtilebilir.
Vestigal ve yabanıl tip gibi tek bir gen farklılığına dayanan heterozigotlar +/vg ile gösterilir. Eğer farklı kromozomlarda bulunan iki veya daha fazla gen çifti kaybedilecekse (vg ve H gibi) her iki mutant için homozigot olan sinek vg/vg, H/H veya vg/H olarak gösterilir. İki gen farklılığı için yabanıl tip / mutant heterozigot vg/+, H/+ olarak sembolize edilir.
Yabanıl tip gen ayrıca mutant faktörün sembolü üstüne + yazılarak da belirtilebilir. Örneğin vg+/vg+ ya da +/+ kanat özelliği bakımından homozigot bireyi ifade eder. Benzer şekilde, w/+ ile w/w+ aynı genotipi (heterozigot kırmızı göz) ifade ederler.
Görüldüğü gibi; Drosophila 'da kullanılan genetik sembol sistemi diğer birçok organizmada kullanılandan farklıdır. Bir karakteri belirtmek için aynı harfin büyük ve küçük yazımı kullanılmaz. Örneğin, B ve b farklı genleri gösterir, bir genin alternatif alelleri değildirler. B dominant bir mutasyon (Bar göz) ve onun yabani tip aleli B+'dır. Oysa b resesif mutasyondur (siyah vücut rengi), normal vücut rengi b+ ya da sadece + ile gösterilir.
Bir kural olarak dişi ebeveynin genotipi önce (sola) yazılır sonra çaprazlamayı temsilen X işareti ve sonra da (sağa) erkeğin genotipi yazılır. Eğer çaprazlamadaki genler otozomal ise genetik formül eşey kromozomlarının özel sembolleri olmadan yazılır.
Bir kromozom üzerindeki birkaç gen birlikte kalıtılıyorsa yani bağlantı durumu söz konusu ile, genlerin sembolleri bir homolog kromozom üzerinde birlikte yazılır ve iki yatay çizgi ile diğer homolog üzerindeki genlerden ayrılır:
w m w m w m /
genotipi w (white) ve m (miniature) genlerinin aynı kromozom üzerinde olduğunu ifade eder.
DROSOPHILA BİYOLOJİSİ
Drosophila melanogaster gelişimleri tam metamorfozlu olan holometabol böcekler grubuna girer. Drosophila sineklerinin döllenme ve zigot oluşumunun ardından embriyonik gelişim ve diğer hayat devreleri süre bakımından ortam sıcaklığına bağımlılık gösterir. Optimum büyüme sıcaklığında (25°C) ortalama 10-15 günde ergin duruma geçerler.
Embriyonik gelişim 1 günBirinci larval instar (L1) 1 günİkinci larval instar (L2) 1 günÜçüncü larval instar (L3) 2 gün

Prepupa 4 saatPupa 4.5 günErgin 40-50 gün
Erkekler pupal evreden çıktıktan kısa bir süre sonra eşeysel olarak fertil ve olgun duruma geçerler. Dişilerin olgunluğa erişmesi soylara bağlı olarak 6-12 saat arasında değişir. Dişi bir defa eşeysel olgunluğa eriştiği zaman birçok erkek sinekle tekrar tekrar eşleşebilir. Sperm, dişi tarafından karın kısmında depo edilir ve kısa bir zaman sonra yumurtaları döllemek için kullanılır.
Her ne kadar ergin sineklerin ömrü 25°C'de 40-50 gün arası ise de bireyler 80 güne kadar yaşayabilirler. 10 günlükten daha büyük olmayan genç sinekler çaprazlama çalışmalarında kullanılır. Çünkü sineklerin yaş ile beraber fertiliteleri azalır.
Standart koşullar altında (25°C, % 60 kısmi nem) yumurtadan tam ergin bireyin gelişimi yaklaşık 10 günü alır. Düşük sıcaklıklarda yaşam döngüsü uzar; 23.5 °C'de 14 gün, 18 °C'de 21 gündür. Kültürlerin 18 °C'den daha küçük sıcaklıklarda bulunması metabolizmayı yavaşlatır ve bu da kültürün devamlı olarak yeni besiyerlerine transfer oranını azaltır. Sıcaklığın artmasıyla gelişim hızlanabilir. Fakat 29-30 °C arasında çarpıcı olarak pupal ölüm artışı gözlenir ve birçok soyda 30°C'nin üzerindeki sıcaklıklarda dişiler steril hale gelirler yani yumurta üretemezler. Ayrıca sıcaklık artışı çeşitli bakteri, mantar gibi organizmaların gelişmesine neden olacağından sinek kültürlerinin kullanılamaz hale gelmesine neden olur.
Yumurta ve Embriyolar
Yumurtalar oval şekilli, yaklaşık 0.5 mm uzunluğunda ve 0.2 mm çapındadır. Dorsal tarafları ventral taraflarına göre daha yassıdır. Dorsal tarafın anterior ucunda iki filament bulunur. Bunlar yumurtanın yumuşak besi ortamına batmasını engeller ve oksijen alımını sağlar. Yumurta koruyucu görevi olan koryon zarı ile kaplanmıştır.
Yumurtaları karnında depo eden dişiler şişkin görünürler. Bu durumdaki dişiler erkek içermeyen besi yerine aktarılırlarsa döllenmemiş yumurtaları taşıyan virjin dişiler elde edilir.
Erkeklerin testislerinde, diploid spermatogonialardan haploid spermler üretilir. Spermin boyu 1.7-1.8 µm kadardır.
Kopulasyon sırasında, spermler dişinin uterusuna aktarılır. Sperm burada günlerce canlı kalabilir. Yumurtalar dişinin uterusunda döllenir. Döllenme sırasında tek bir sperm mikropil tarafından yumurtaya girer (monospermi). Dişi, yumurtalarını ya hemen döllenmenin ardından bırakır ya da embriyonik gelişmenin ilk dönemleri uterusta geçer.
Larva
Embriyonik gelişimden yaklaşık 1 gün sonra yumurtadan çıkan larvalar, besiyerinin yüzeyinde bulunurlar. Larval gelişim sırasında bira mayası hücreleri temel besin maddeleridir. 1 gün sonra ilk deri değişimi gerçekleşir. Bu larvanın birinci instar (L1) döneminden ikinci instar (L2) dönemine geçişidir. Larva, 1 gün daha sonra, ikinci deri değişimi ile üçüncü instar döneme (L3) geçer. L3 larva dönemi boyunca 2 gün süresince beslenir ve hızla büyürler. Yaşlı larvalar besi yerini delik deşik ederler ve besi yeri içinde tünel açan işçiler gibidirler. Larvalar kültür şişesinin etrafında ve besi yeri içinde çıplak gözle bile görülebilirler. Besin almalarını sağlayan çengel şeklindeki ağız parçaları önemli

özelliklerindendir.
Pupa
Üçüncü larval dönemin sonunda, larvalar besi yerinden ayrılırlar ve diğer gelişimlerini tamamlamak için şişenin etrafındaki uygun bir yere tırmanarak pupalaşırlar. Pupalar başlangıçta beyazdır, sonra iki saat içinde koyumsu sarı renk, en sonunda da kahverengileşirler. Pupa evresi hareketsizdir ve beslenme görülmez. Pupal periyodda bir erginin organları ve vücut formuna sahip bir bireyin gelişmesi için gerekli dönüşümler (metamorfoz) gerçekleşir. Bu gelişme 20°C'de 6 gün, 25°C'de 4 günde tamamlanır.
Ergin Sinekler
Pupa formasyonu içinde gelişimin tamamlanması ile ergin sinekler pupa kılıfının anterior ucunu (operkulum) delerek dışarı çıkarlar. Yeni çıkan ergin bireyler önceleri açık renkli ve uzun vücutludur. Fakat daha sonra birkaç saat içinde renkleri koyulaşır. Kanatlar ve vücudun diğer kısımları başlangıçta yumuşaktır. Havadaki oksijene maruz kaldıklarında birkaç saat içinde sertleşir. Böylece başlangıçta kırışık olan kanatlar açılır ve normal ergin birey görünümüne ulaşırlar.
ERGİN SİNEKLERDE EŞEY FARKLILIKLARI
Genetik çaprazlamalar için ergin dişi ve erkek sineklerin birbirlerine karıştırılmadan ayrılması zorunludur. Her ne kadar ergin dişi ve erkekler arasında birçok morfolojik farklılıklar var ise de bunlardan yalnız birkaçı eşey farklılıklarını belirlemek için güvenilirdir.
1. Eşey Tarağı: Ergin sinekler stereo mikroskop altında incelendikleri zaman; erkek bireylerin birinci çift yürüme bacaklarının Tarsus segmentinin bazal tarafında siyah ve kalın bir seri kıldan meydana gelmiş "eşey tarağı" (sex comb) denilen yapılara sahip olduğu gözlenir. Dişilerde bu yapı bulunmaz. Deneysel çaprazlamalar için erkek ve dişi bireyleri seçerken bu özelliğin kullanılması (özellikle pupadan yeni çıkmış sineklerde) güvenilir olmaktadır.
2. Genital Organ ve Abdominal Renklenme: Genital organın dış yapısı her iki cinsiyette birbirinden farklıdır. Fakat bu farklılıklar sadece mikroskopta 25-ya da daha fazla kez büyültme sonucu gözlenebilir. Genel olarak erkeğin genital organı daha koyu yapıya sahiptir.
Dişiler karın bölgesinde 7 abdomen segmentine sahiptir. Erkeklerde ise sadece 5 abdomen segmenti vardır. Erkek sineklerde abdomenin son kısmı siyahtır, çünkü son segmentlerde güçlü bir şekilde pigment birikimi olur ve böylece siyah bir bölge oluşur. Dişi sineklerde ise abdomen segmentleri açık ve koyu bantlar halinde uç kısma kadar uzanır (Bkz. Şekil 1). Bu özellikler, özellikle pupadan yeni çıkmış bireylerin eşey ayırımı için uygun değildir. Çünkü bireylerde pigmentasyon henüz tam olarak gelişmemiştir. Ayrıca bayıltılmış erkek sineklerin abdomenleri uzar ve bu da abdomenin dişilerinkine benzer olarak çizgili görünmesine neden olabilir.

3. Abdomenin Şekli: Erkek sineklerin abdomen ucu küttür oysa dişilerinki uzundur. Ayrıca dişilerin yaşlanması ve devamlı yumurta gelişimi dolayısıyla abdomen geniştir.
4. Vücut Büyüklüğü: Genel olarak dişiler erkeklere göre daha büyüktür. Fakat ergin sineklerin vücut büyüklüğü larval dönem boyunca beslenme durumlarına bağlı olduğundan, güvenilir bir kriter değildir.
5. Kanat Uzunluğu: Dişilerin kanatları erkeklerinkine oranla daha uzundur. Fakat bu özellik de eşey ayırımı için uygun değildir.
GENETİK ÖZELLİKLERİ
Drosophila 'ların somatik doku hücrelerinde diploid sete sahip 4 çift kromozom (2N=2X=8) bulunur. Bu seti oluşturan kromozomlardan 3 çifti otozomal (somatik kromozomlar) 1 çifti de gonozomal (cinsiyet kromozomları) dır. Bunlar X, Y, 2, 3 ve 4 şeklinde numaralandırılmıştır. Dişilerde 2 tane çubuk şeklinde X kromozomu, erkek sineklerde ise 1 X kromozomu ve 1 de ters J şeklinde Y kromozomu bulunur.
Homogametik dişiler (XX) ile heterogametik erkekler (XY) arasındaki eşey kromozomu farklılıklarının özel bir önemi vardır. Erkeklerde X kromozomu sadece 1 tane bulunur ve bu yüzden X üzerindeki genler hemizigot durumda bulunurlar. Bunun sonucu olarak X kromozomu üzerinde meydana gelen mutasyonlar (cinsiyete bağlı genler) dominant olmasına bakılmaksızın erkeklerde fenotipik olarak gözlenir. Yani resesif mutasyonlar erkek bireylerde dominant mutasyonlar kadar fenotipe yansır.
Dev Kromozomlar
Dipter larvalarının tükrük bezleri, barsak, trake, yağ dokusu ve malpighi tübülleri gibi çeşitli somatik doku hücrelerinde bulunan ve aynı organizmanın diğer vücut hücrelerindekine kıyasla yaklaşık 1000 kat daha büyük olan kromozomlara politen kromozomlar denilmektedir. (Bkz. Deney I ve Şekil 5).
Mayozun Genetik Kontrolü
D. melanogaster 'de mayozun çok özel genetik kontrolü dikkat çeker. Dişilerde oogenez sırasında mayozda krosing over düzenli olarak meydana gelir. Bunun aksine erkeklerdeki spermatogenezde mayotik krosing over görülmez ya da çok nadir meydana gelir. Erkek sineklerde krosing over görülmemesinin sebebi kromozom üzerindeki genlerin birbirine çok yakın bulunmasından kaynaklanmaktadır. Bu durum spermatogenez sırasında kiyazma bölgesinin oluşmasını engellemektedir.
SİNEKLERİN LABORATUVARDA YETİŞTİRİLMELERİ
Drosophila kültürleri, laboratuvar koşullarında 24°C sabit sıcaklığa ayarlı havalandırmalı etüvde yetiştirilirler.
Kültür kaplarının büyüklüğü laboratuvar şartlarına ve deneyin amacına göre değişebilir. Fakat, sınıf deneylerinde genellikle 200 ml'lik cam şişeler uygun olmaktadır. Bu

amaçla süt ya da ayran şişeleri kullanılabilmektedir. Kültür kabunun renksiz camdan olması, sineklerin gelişimlerinin kolay izlenebilmesi sebebiyle tercih edilmektedir. 200 ml'lik şişeye yaklaşık 40 ml besi yeri konularak ağzı tıpayla kapatılır. Tıpa şişeye yeterli havanın geçişini engellemeyecek bir özellikte olmalıdır. Bu nedenle ya sünger tıpa ya da gazlı bez ile sarılmış pamuk yumağı kullanılır. Şişeler ve tıpalar kullanılmadan önce steril edilirler (kuru hava ile sterilizasyon).
Meyve sinekleri doğada, fermente olan çeşitli bitki materyallerinin üzerinde bulunurlar, fakat doğrudan meyve üzerinden beslenmezler. Mayalar bu sineklerin temel besin maddesini oluştururlar. Bu nedenle maya çoğalmasını sağlayan her türlü ortam onlar için iyi bir besin ortamıdır. İyi bir kültür ortamı, larvanın beslenmesine ve mayanın gelişmesine yeterli olacak miktarda şeker içermeli ve uygun yumuşaklıkta olmalıdır.
Değişik laboratuvarlarda çok çeşitli kültür ortamları hazırlanabilmekte, ya da hazır olarak satın alınabilmektedir. Bizim laboratuvara kullanılan besiyerinin içeriği ve hazırlanışı aşağıda verilmiş olup, hazırlanması kolay ve ucuz olma özelliğine sahiptir.
a) Besiyerinin içeriği:
104 g mısır unu94 g şeker9 g bira mayası6 g agar1020 ml distile su6 ml asit karışımı (7.83 ml Ortofosforik asit + 8.36 ml Propiyonik asit + 1081 ml
distile su)
b) Hazırlanışı:
Mısır unu, toz şeker ve bira mayası tartılarak üzerine 500 ml distile su konur ve iyice karıştırılır. Geri kalan 520 ml distile su içine ise agar tartılarak konur ve tamamen eriyinceye kadar kaynatılır. Bu arada mısır unu, şeker, bira mayası ve sudan oluşan karışımın da pişirilmeye başlanmış olması gerekir. Kaynama durumundaki bu karışımın üzerine, eritilmiş agar üç aşamada dökülür ve devamlı olarak karıştırılır. İyice homojen hale gelince üzerine mantar-bakteri gelişmesini önleyici özelliği olan asit karışımı ilave edilir ve dağılması için kaşık ile karıştırılır. Fakat asit karışımının fazlası gerek maya gerekse sineklerin gelişimini engelleyeceğinden sadece yeterli miktarda konulmalıdır.
Böyle bir ortamda mayalar çok hızlı bir şekilde çoğalırlar ve sinekler için iyi bir besin kaynağı olurlar. Fakat aşırı miktarda maya katılmamasına dikkat edilmelidir. Çünkü maya hücreleri sinekler için gerekli olan O2'yi kullanarak ortamdaki oksijenin azalmasına neden olurlar.
Hazırlanan besi ortamı sıcak iken steril kültür şişelerine yaklaşık 2 cm yükseklikte olacak kadar dökülür ve ağzı tıpa ile kapatılır. Soğurken agar nedeniyle katılaşır. Kullanılana kadar serin ve temiz bir yerde saklanır.
SİNEKLERİ BAYILTMA İŞLEMİ (ETERİZASYON)
Kontrollü çaprazlama deneylerinde kullanılacak olan sineklerin çeşitli fenotipik özelliklerini önceden belirlemek gerekir. Bu nedenle canlı sinekler bayıltma işlemine tabi tutulur. Bu amaçla iki farklı gaz; eter ve CO2 uygundur. Fakat gerekli ekipmanı daha basit

olduğu için sınıf deneylerinde eter kullanmak daha avantajlıdır.Eter kullanımındaki temel prensip sinekleri kısa bir süre için eter buharına maruz
bırakmaktır. Etere maruz bırakma süresi oldukça kısa olmalıdır. Çünkü sinekleri anestezi etmek, kısırlaştırmak ya da ölümlerine neden olmak arasında çok kısa bir süre vardır. Kısırlaşan sinekler döl vermemeleri ile tanınabildikleri halde ölen sinekler kolayca tanınırlar. Çünkü fazla eter buharının etkisi ile ölen sineklerin kanatları thoraxtan ayrılarak 45°'lik bir hal alır. Bacaklar aşırı kasılmadan dolayı gergin uzanırlar ve vücut kıvrılarak hareketsiz kalır.
Eterizasyon işlemi için bayıltma şişesine, etere ve pamuğa ihtiyaç vardır. Kültür şişelerinden bayıltma şişelerine sineklerin aktarılmasında hızlı ve dikkatli olmak gerekir.
(1) Yumak haline getirilmiş pamuk üzerine birkaç damla eter damlatılarak bayıltma şişesinin ağzına kapatılır.
(2) Kültür şişesinde bulunan sineklerin şişenin tabanında toplanması için bir kağıt destesi üzerine vurulur.
(3) Kültür şişesinden pamuk tıpa çıkarılarak ağzı, bayıltma şişesinin ağzı üzerine getirilir. Burada şişelerin durumunu korumasına dikkat edilmelidir. Çünkü meydana gelecek küçük bir açıklıktan sinekler kaçabilirler.
(4) Sineklerin dikey durumda olan şişelerden alttaki şişeye düşmesi için alttaki şişe bir kağıt destesi üzerine hafifçe vurulur. Böylece sinekler bayıltma şişesinin içine düşerler.
(5) Her iki şişe birbirinden ayrılır ve hemen kültür şişesine tıpası tekrar kapatılırken bayıltma şişesine de üzerine eter damlatılmış pamuk tıpa kapatılır.
Yaklaşık 1-2 dakika sonra sinekler eter buharının etkisiyle bayılırlar. Bu anestezik uygulamadan sonra sinekler normal havaya alınır ve stereo mikroskop altında tabanına beyaz kağıt yapıştırılmış bir petri kabı içinde ya da beyaz fayans üzerinde yumuşak bir sulu boya fırçası kullanılarak incelenirler. Sinekler bu şekilde 10-15 dakika kadar kalabilirler. Anestezi etkisinin geçmesiyle hareket etmeye başlayan sinekler eğer gerekirse içine konuldukları petri kabunda tekrar bayıltılabilirler. Ölüm durumunda bazı fenotipik karakterler değişebileceğinden inceleme ve sayım işlemlerine alınmamalıdır. Ölen, işe yaramayan ya da kullanılmayan sinekler morga atılır.
UYARI: Eter ve hava karışımı patlayabilir. Eterin kaynama noktası yaklaşık 35 °C'dir. Eter güçlü bir narkotiktir. Çalışma ortamında yeterince temiz havanın bulunması önemlidir. Eter, zehirli peroksidazın meydana gelmesini engellemek için kahverengi şişelerde saklanmalıdır. Solumamalı ve etrafında ateş yakılmamalıdır.UYARI: Bayıltma işlemi sırasında sinekler doğrudan etere maruz bırakılmamalıdır.
YENİ KÜLTÜRE BAŞLAMA VE TRANSFER İŞLEMLERİ
Yeni bir kültüre başlandığı zaman ne şişe duvarının ne de besiyeri yüzeyinin ıslak olmaması gerekir. Her ne kadar Drosophila 'nın kelime anlamı "nemi seven" olsa da sinekler ıslak kısımlara yapışabilir ve ölebilirler.
Laboratuvarda yetiştirilen Drosophila kültürlerinin en az ayda 1 kez taze besiyeri içeren şişelere aktarılması gerekir. Bunun için öncelikle yeni hazırlanan şişenin üzerine tarih, Drosophila soyu gibi bilgiler yazılarak etiketlendirilir. Aktarma işleminde en basit yol bir şişeden taze besiyeri içeren şişeye sineklerin doğrudan aktarılmasıdır. İlk olarak sinek içeren şişe bir kağıt destesi üzerine vurularak sineklerin şişenin tabanında birikmesi sağlanır. Sonra her iki şişenin tıpası çıkarılır. Sinek içeren şişenin ağız kısmı boş olan şişenin ağzına

getirilir (ya da tersi uygulanarak, şişeler ters yüz edilir). Tekrar kağıt destesi üzerine vurularak üstteki sineklerin boş şişeye geçmesi sağlanır. Sonra hızla her iki şişenin ağızları tıpaları ile kapatılır.
Çaprazlama deneyleri sırasında ise stereo mikroskop altında eşey tayini yapılan sinekler bayılmış durumdadırlar. Bu haldeki sineklerin yeni besi yerine aktarılması oldukça dikkatli olmayı gerektiren bir işlemdir. Aktarma sırasında anestezi halindeki sineklerin besiyerine ya da diğer yüzeylere yapışmaması gerekir. Bunun için en kolay yol şişeyi eğik tutarak petri kabu içindeki ya da beyaz bir fayans üzerindeki sineklerin şişenin kuru tarafına yumuşak bir fırça yardımıyla aktarılmasıdır. Sinekler hareket edene kadar şişenin yatay konumda kalması gerekir. Anestezinin etkisi geçtikten sonra şişe dik konuma getirilmelidir. Aynı amaç için kullanılan diğer bir yol da Anestezi uygulanmış sineklerin önce boş, kuru ve temiz olan bir şişeye alınıp ayılmalarının beklenmesi ve daha sonra hareketli sineklerin taze besi yeri içeren şişeye aktarılmasıdır.
DENEYSEL ÇAPRAZLAMALAR
Çaprazlamalara başlamadan önce ergin sineklerin eşeysel dimofizme dayanan özellikleri ile mutantların belirgin fenotipik özelliklerinin çok iyi bir şekilde öğrenilmesi gerekmektedir.
1. Virjin Dişilerin Seçilmesi
Çaprazlama deneyleri için henüz hiç döllenmemiş virjin dişilere ihtiyaç duyulur. Virjin dişileri toplamak için tercihen 9-10 günlük, koyu kahverengi pupaları olan kültür seçilir ve bu şişeden tüm erginler uzaklaştırılır. Kültürde tek bir dişi ya da erkek sinek kalmamalıdır. Stok kültüre bağlı olarak 6-8 saat sonra dişiler toplanır. Dişi sinekler pupadan çıktıktan sonra yaklaşık 12 saat çiftleşmezler. Bu süre sonunda sinekler eter ile bayıltılır. Stereo mikroskop altında dişiler ve erkekler iyi bir fırça kullanarak dikkatli bir şekilde ayırt edilir. Erkekler genellikle atılır ya da diğer çaprazlamalarda kullanılmak için uzaklaştırılır. Dişiler hiç erkek içermeyen taze bir besiyerine alınırlar. Burada 2-3 gün içinde eşeysel olgunluğa erişirler.
Virjin dişilerin toplanması Drosophila deneylerinin en kritik kısmını oluşturur. Bu nedenle virjin dişilerin toplanması aşağıda tekrar aşamalar halinde yazılmıştır.
(1) Virjinleri toplamaya başlamadan önce, kültürde tek bir sinek bile kalmamalıdır.(2) Pupadan çıkan ergin sinekler en kısa zamanda şişeden alınarak cinsiyet tayini
yapılmalıdır.(3) Her iki cinsiyet birbirinden çok dikkatli bir şekilde ayırt edilmelidir. Erkeklerdeki
eşey tarağı ve genital bölgenin dış yapısı pupadan yeni çıkmış sineklerin eşey ayırımı için en kullanışlı özelliklerdir.
(4) Yeni kültür şişesine dişileri aktarmadan önce hiç erkek bulunmadığına emin olunmalıdır.
2. Erkeklerin Toplanması
Erkek sinekler herhangi bir zamanda toplanabilirler. Erkeklerin çok genç olması gerekmez. Genellikle 1-2 günlük erkek sinekler toplanır ve besiyerine alınırlar.

3. Bazı MutantlarDeğişik soylardan alınan 2-6 günlük sinekler eter ile bayıltılarak stereo mikroskop altında incelendiğinde, vücut rengi, göz şekli ve rengi, kanat büyüklüğü ve şekli, kanat damarlanma tipi, kılları gibi özellikler bakımından, yabani tip (wild type) Drosophila 'dan farklı morfolojik özelliklere sahip oldukları gözlenir. Bunlar yabani tip Drosophila 'nın mutant tiplerini oluştururlar.
Yabanıl tip terimi, organizma için genotip ve fenotip bakımından standart olarak belirlenen bir soy, gen ya da organizmayı gösterir. Örneğin yabanıl tip Drosophila soyu, gri vücut rengine ve uzun kanatlara sahiptir; göz şekli yuvarlak ve göz rengi ise parlak kırmızıdır. Yabanıl tip genlerin mutasyonal değişimleriyle mutant aleller ve dolayısı ile mutant soylar ortaya çıkar.
Bazı mutant soylar aşağıda listelenmiştir.
Sembol Tanımlama
b black; siyah vücut rengi
B Bar; homozigot B/B ♀ ve hemizigot B/Y ♂: gözler küçülmüş çubuk göze
yaklaşmıştır; B/+ ♀: böbrek şekilli gözler
bw brown; kahverengimsi kırmızı göz rengi
Cy Curly; kanatlar yukarıya doğru kıvrılmıştır.; homozigot hali öldürücüdür.
dm diminutive; kıllar kısa ve incelmiş; dm/dm dişileri kısırdır.
dp dumpy; kanadın posterior ucu kesik
e ebony; siyah vücut rengi; e geninin kromozom üzerinde bulunuş yeri b'den
farklı
ey eyeless; göz, normal gözün yaklaşık olarak 1/4 'ü kadar
f forked; kıllar çatallanmış
H Hairless; postvertikal ve abdominal kısımlarda kılları yoktur; homozigot hali
öldürücüdür.
m miniature; kanatlar kısa; yaklaşık abdomen boyu kadar
Pm Plum; kahverengimsi göz rengi; homozigot hali öldürücü
ry rosy; koyu kırmızı göz rengi
se sepia: kahverengi göz rengi; yaşlandıkça koyulaşır.
sn singed; kıllar kısa ve karışık
vg vestigal; kanatlar körelmiş, küçülmüş
w white; beyaz göz rengi
y yellow; ergin sineklerin vücut rengi sarı

4. Çaprazlama
Farklı mutant formlar arasında çaprazlama yapmak için bir mutant grubundan alınan yaklaşık 3-5 virjin dişi, diğer mutant gruptan alınan aynı sayıdaki erkek sinekle yeni bir kültür ortamında eşleştirilir. Çapraz kurulur kurulmaz kültür şişesinin üzerine çaprazlama çeşidi, kullanılan sineklerin cinsiyeti, sayısı ve tarih gibi bilgiler yazılır (etiketleme). Bu kültür şişeleri 24 °C sabit sıcaklıktaki etüve alınır. Dişiler çiftleşmeden 2 gün sonra yumurta bırakmaya başlarlar. Çaprazlama yapılan kültür şişesinde ortalama 5 gün sonra larvalar ortaya çıkar ve bunlar besin içinde tünel kazan işçiler gibi görünürler. Larvaları koyu renkli ağız parçalarından dolayı ayırt etmek kolaydır. Yaklaşık 7-8 gün sonra pupalar görülmeye başlandığında, kültür şişesinden ebeveynler uzaklaştırılır. Çünkü sayım yapıldığında yeni döl sineklerin ebeveynlerle karışmaması gerekir. Başarılı kültürlerde ilk ergin sinek ortaya çıktıktan sonraki bir hafta içinde yaklaşık 200-300 kadar yavru döl elde edilir.
Resiprokal çaprazlamalar için her iki mutant formdan hem virjin dişiler hem de erkekler seçilir ve bunların zıt kombinasyonlarda iki ayrı kültürü yapılır. Örneğin yabani dişiler ile vestigial erkeklerin çaprazlanmasına karşılık, bunun resiprokali olarak vestigial dişiler yabani erkekler ile çiftleştirilir.
5. Kayıt ve Sayma İşlemleri
Çaprazlama kurulduğu zaman kültür şişesinin üzerindeki etikete yazılan tüm bilgiler deney kayıt formuna da yazılır. Sınıf deneyleri genellikle gruplar halinde yapıldığı için grup numarası da kaydedilmelidir. Çaprazlama için kullanılan mutantların stok kültürlerinden alınan sinekler ebeveyn olarak raporun P1 X P1 kısmında gösterilir. P1 X P1
çaprazlamasından elde edilen ilk döller F1 olarak, sonraki generasyonlar ise F2 ve F3 vs. şeklinde adlandırılır.
Çaprazlamanın olduğu kültür şişesinde sinekler pupadan çıktıktan sonra sayım işlemleri için önce eterle bayıltılmaları gerekir. Daha sonra beyaz zeminde belli bir düzende sayımları yapılır. Hem cinsiyet hem de mutant fenotipleri dikkate alınarak sinekler teker teker ayrılır ve sayımları yapılır.
Sayım işlemleri 8 gün boyunca her gün veya iki günde bir yapılmalıdır. 8 günden daha az sayım ağır gelişme hızına sahip mutant ve eşeydeki sineklerin sayımını engelleyebilirken daha fazla sayım da sonraki generasyonun sineklerini içerebilir. Ayırım ve sayım işlemi yapılan sinekler ile ilgili olarak elde edilen bilgiler hemen kayıt formuna geçirilir ve daha sonra morga atılırlar.
RAPOR YAZMA VE SONUÇLARIN DEĞERLENDİRİLMESİ
Rapor azırlanırken uyulması gereken aşamalar aşağıda verilmiştir:
1. Giriş: Deneyin amacı ve ileri sürülen hipotez açıklanır.2. Materyal ve Yöntem: Deneyde kullanılan malzeme ve deneyin yapılış şekli anlatılır.3. Bulgular: Deney sonucunda elde edilen veriler kayıt formuna kaydedilir.4. Sonuçların değerlendirilmesi: İlk olarak kalıtım mekanizması diagram çizilerek açıklanır. Çaprazlama sonucunda gözlenen ve beklenen değerler arasında farklılıklar meydana gelebilir. χ2 (ki kare) testi kullanılarak beklenen ve gözlenen değerler karşılaştırılır. Böylece öne sürülen hipotezin kabul edilip edilmeyeceğine karar verilir.

χ2 (ki kare) değerini hesaplamak için; elde edilen değerle (gözlenen) beklenen değer arasındaki farklar bulunur. Farkların kareleri beklenen değerlere bölünür. Elde edilen değerlerin toplamı χ2 (ki kare) değerini verir.
Bu testte, hangi sapma derecesinin rastlantıya bağlı hangisinin ise önemli olduğuna karar vermek için bir ölçüt saptanmıştır. Bu ölçüt olasılık (P) derecesiyle ifade edilir. Elde edilen orana ait P değeri 0.05'e eşit veya daha küçük ise gözlenen ve beklenen değerler arasındaki sapmanın önemli olduğu kabul edilir ve öne sürülen hipotez reddedilir. Eğer, P = 0.01 ise sapma çok önemlidir. Bu durumda, gözlenen ve beklenen arasındaki sapma kesinlikle rastlantıya bağlı değildir ve sonucu etkileyen etkenler vardır. P değeri, 0.05'den büyük ise sapma istatistiki bakımdan önemli değildir ve öne sürülen hipotez kabul edilir. Meydana gelen sapma sadece rastlantıya bağlıdır.
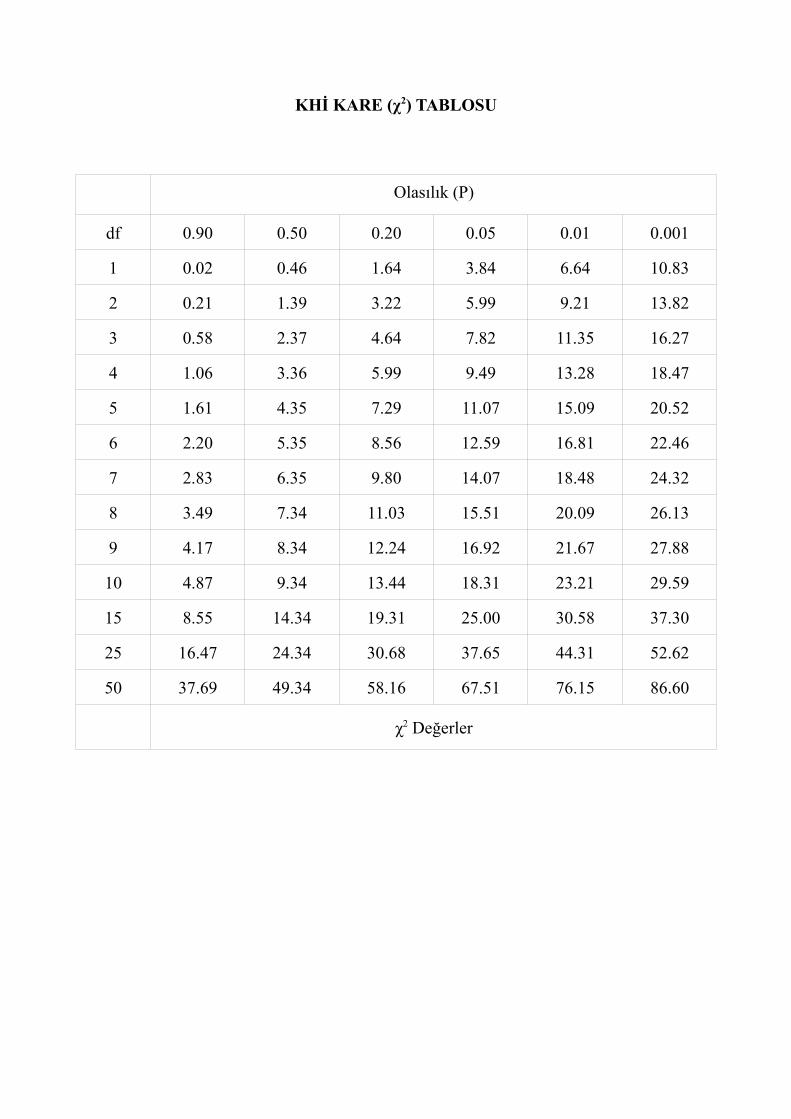
Elde edilen χ2 (ki kare) değerini kullanarak P değerini bulmak için χ2 tabloları düzenlenmiştir. Fakat P değerini belirleyebilmek için önce serbestlik derecesinin (df) hesaplanması gerekir. Serbestlik derecesi N sayıda grubu (farklı fenotipi) kapsayan bir analizde N-1'e eşittir. χ2 (ki kare) tablosu üzerinde, analizdeki serbestlik derecesinin bulunduğu yatay sütunda hesaplanan χ2 değerinin uygun düştüğü dikey sütun belirlenir ve bu bölgenin hangi P değerinin karşılığı olduğu bulunur.
Böyle çalışmalarda öncelikle kritik ki kare değeri belirlenerek hipotezin kabul edilip edilmeyeceğine karar verilebilir. Daha sonra eğer sapma varsa önem seviyesi belirlenir. Kritik χ2 (ki kare) değeri, ki kare tablolarında düşey sütundaki serbestlik derecesinin 0.05 düzeyindeki χ2 değeridir. Eğer elde edilen χ2 (ki kare) değeri, kritik χ2 değerinden büyükse sapma vardır ve hipotez reddedilir. χ2 (ki kare) değeri kritik χ2 değerine eşit veya küçükse hipotez kabul edilir.
Aşağıdaki örneği inceleyiniz.
P: AaBb ♀ x aabb ♂ (A, B; a ve b'ye dominanttır)
G: AB, Ab, aB, ab ab
F1: ¼ AaBb, ¼ Aabb, ¼ aaBb, ¼ aabb
F1 neslinde dört fenotipik sınıfın 1:1:1:1 oranında ortaya çıkmasını bekleriz. Eğer çalışılan toplam birey sayısı 200 ise fenotipik sınıflara göre dağılımının aşağıdaki gibi olması gerekir.Beklenen birey sayısı:AaBb=50 Aabb=50 aaBb=50 aabb=50
Gözlenen birey sayısı ise çalışma sonunda her bir fenotipten sayılan birey sayısıdır.Örneğin:AaBb=44 Aabb=66 aaBb=32 aabb=58 olsun. Gözlenen ve beklenen değerler aşağıdaki tabloya yazılır.
Fenotipler ToplamGözlenen (o) 44 66 32 58 200Beklenen (e) 50 50 50 50 200Sapma (o-e) -6 16 -18 8

Sapma2 (d2) 36 256 324 64Sapma2/beklenen
(d2/e)0.72 5.12 6.48 1.28 χ2=13.6
Bu deneyde farklı fenotipik sınıf sayısı (N) 4 olduğundan dolayı serbestlik derecesi (N-1) 3'tür. χ2 tablosu üzerinde, serbestlik derecesinin 3 olduğu yatay sütunda hesaplanan χ2
değerinin uygun düştüğü dikey sütun belirlendiğinde P değerinin 0.01 ile 0.001 arasında olduğu görülür (P<0.01).
df=3 olduğunda 0.05 düzeyindeki kritik χ2 değeri=7.82'dir. Elde edilen χ2 =13.6 kritik ki kare değerinden büyük olduğundan hipotez reddedilir. Buna göre, beklenen ile gözlenen değerler arasında sapma vardır. Sapmanın önem seviyesi belirlendiğinde ise Px0.01 düzeyinde anlamlı olduğu görülür. Yani meydana gelen sapma rastlantıya bağlı değildir ve sonucu etkileyen önemli sebepler vardır (örneğin genler bağımsız olarak birbirlerinden ayrılmamış olabilirler).
5. Tartışma: Elde edilen sonuçlar benzer çalışmaların sonuçları ile karşılaştırılır. Çaprazlama sonucunda elde edilen verilere göre, beklenen ve gözlenen oranlar arasında Hardy-Weinberg dengesinden sapma varsa sebepleri açıklanır.

KAYIT FORMU
Adı Soyadı :No/Grubu : Çaprazlama Tarihi:
Deneyin Adı :Deneyin Amacı:
P1 x P1 ♀ x ♂
F1 Genotip ___________________ Fenotip
F1 x F1 ♀ x ♂
F2 Fenotipleri:
Sayım Tarihi
F2 Fenotipleri ve elde edilen sinek sayısı
♀ ♂ ♀ ♂ ♀ ♂ ♀ ♂
1
2
3
4
5
Toplam
Beklenen
χ2
P
KHİ KARE (χ2) TABLOSU
Olasılık (P)
df 0.90 0.50 0.20 0.05 0.01 0.001
1 0.02 0.46 1.64 3.84 6.64 10.83
2 0.21 1.39 3.22 5.99 9.21 13.82
3 0.58 2.37 4.64 7.82 11.35 16.27
4 1.06 3.36 5.99 9.49 13.28 18.47
5 1.61 4.35 7.29 11.07 15.09 20.52
6 2.20 5.35 8.56 12.59 16.81 22.46
7 2.83 6.35 9.80 14.07 18.48 24.32
8 3.49 7.34 11.03 15.51 20.09 26.13
9 4.17 8.34 12.24 16.92 21.67 27.88
10 4.87 9.34 13.44 18.31 23.21 29.59
15 8.55 14.34 19.31 25.00 30.58 37.30
25 16.47 24.34 30.68 37.65 44.31 52.62
50 37.69 49.34 58.16 67.51 76.15 86.60
χ2 Değerler

DENEY I
Deneyin Adı: Monohibrit Çaprazlama
Bir gen çifti bakımından farklılık gösteren bireyler arasındaki çaprazlamaya monohibrit çaprazlama denir. Bu deneyde iki otozomal genin monohibrit çaprazı için yabani tip (normal kanatlı, +) Drosophila ile vestigial (körelmiş kanatlı, vg) Drosophila kullanılacaktır. Vestigial geni 2'nci kromozom üzerinde bulunur ve otozomal mutanttır.
Yöntem:
1) Çaprazlamaya başlamadan önce her iki soya ait ayırt edici fenotipik özellikleri yeniden gözden geçiriniz.2) Virgin dişiler kullanarak resiprokal çaprazlamalar yapınız.
a) ++ ♀ x vgvg ♂ b) vgvg ♀ x ++ ♂Çaprazlama 5 dişi ve 5 erkek sinekle başlatılır. Her kültür şişesinin üzerine
çaprazlama tipi ve tarih yazılır. Bir kültür şişesine yabani tip dişi ve mutant erkek (a), diğer kültür şişesine mutant dişi ve yabani erkek (b) sinekler konulur. Resiprokal çaprazlama denilen bu iki yönlü çaprazlama, çalışılan karakterde cinsiyete bağlı bir durum olup olmadığını ortaya koyar.3) 7 gün sonra ebeveyn sinekler (P1) kültürden uzaklaştırılır. Bu işlem yapıldığında, kültür şişesinde çok sayıda larva ve pupa olması gerekir.4) F1 generasyonunun (birinci yavrudöl kuşağı) sinekleri ortaya çıktığı zaman fenotipleri kontrol edilir.5) Yeni bir kültür şişesinde F1 generasyonundan elde edilen 5 dişi ve 5 erkek sinek çaprazlanır. Etiketlendirilir.6) F1 çaprazlaması kurulduktan 6-7 gün sonra, kültür şişesinde larvalar gözlenmeye başlandığı zaman birinci yavru döller atılır ve 8 gün boyunca sayım yapılır. Elde edilen sonuçlar kayıt formuna kaydedilir.7) Sayım yaptığınız toplam birey sayısına göre, farklı fenotiplere sahip olan sineklerin oranları hesaplanır. Teorik olarak beklenen oranlarla karşılaştırılır. Çaprazlamadaki kalıtım mekanizması Mendel kanunları göz önüne alınarak diagram halinde gösterilir (yani gametleri meydana getirmek için genlerin ayrılımı ve zigotta tekrar birleşmeleri gibi). Ki kare testi kullanılarak sonuçların doğruluğu kontrol edilir. Beklenen oranlarla gözlenen oranlar arasında sapma olup olmadığı, varsa % 5 kriterine göre önem seviyesi belirlenir.

DENEY 2
Deneyin Adı: Test Çaprazlama (Geri Çaprazlama)
Test çaprazlaması, dominant fenotipli fakat genotipi bilinmeyen organizmanın homozigot resesif bireyle çaprazlanarak genotipinin belirlenmesi amacıyla yapılır.
Monohibrit çapraz sonucu elde edilen ve fenotipik olarak benzer olan heterozigot F1
döllerinin genotipik yapısı gösterilir.
Yöntem:
1) Birinci deneyde elde edilen F1 sineklerinden aynı cinsiyettekiler alınır ve vestigial kanatlı karşıt cinsiyettekilerle çaprazlanır. Bu çaprazlamanın resiprokali de yapılabilir.2) Monohibrit çaprazlamada izlenilen aşamalar uygulanır.3) Deney sonunda çaprazlamanın mekanizması ve elde edilen değerler açıklanır.

DENEY 3
Deneyin Adı: Eşeye Bağlı Kalıtım
Mendel'in dominantlık kuralına göre, resesif ve dominant homozigot bireylerin resiprokal çaprazlamalarından elde edilen hibritlerin (melez) aynı genotipi göstereceği önceden bellidir. Fakat erkeklerde eşeye bağlı genlerin hemizigot olması sebebiyle, bu genler resiprokal çaprazlamalarda daha farklı bir durum gösterirler. Bu deneyde Drosophila melanogaster 'de eşeye bağlı kalıtım gözlenmeye çalışılacaktır. Deneyde beyaz gözlü (w) Drosophila soyu kullanılacaktır. Beyaz göz geni, birinci kromozom (X) üzerinde taşınan eşeye bağlı resesif bir gendir.
Yöntem:
1) Normal kırmızı gözlü Drosophila ile beyaz gözlü mutant Drosophila 'lar arasında resiprokal çaprazlamalar yapılır.
I. Çapraz II. Çapraz + w w +
x x + / w /
X kromozomunu; / Y kromozomunu ifade eder.
2. F1 dölleri çıkınca kendi aralarında çaprazlanır ve elde edilen F2 dölleri fenotiplerine ve cinsiyetlerine göre dikkatli bir şekilde sayılır.
3. Eşey kromozomlarının dağılımını göstermek için diagram çizilir, bu iki çaprazın sonuçları değerlendirilir.
4. Bu çaprazlamaların sonuçları ile monohibrit çaprazlama sonuçlarını karşılaştırınız. Benzerlik ya da farklılıklar var mıdır? Neden?

DENEY 4
Deneyin Adı: Dihibrit Çaprazlama
İki gen çifti bakımından farklılık gösteren bireyler arasındaki çaprazlamaya dihibrit çaprazlama denir.
Mendel'in bağımsız dağılım prensibine göre; gamet oluşumu sırasında farklı genlere ait alellerin bir araya gelmeleri birbirinden bağımsızdır ve rastlantıya bağlıdır. Bu kural eğer genler birbirine bağlı genler değilse geçerlidir. Buna göre, dihibrit çaprazlama, ayrı kromozomlarda yer alan iki gen çiftinin döllere aktarılmasında birbirleri ile nasıl bir ilişki içinde olduklarını açıklayan bağımsız dağılım prensibine dayanır.
Herhangi bir genetik çaprazlamanın sonuçlarını önceden tahmin etmek için ilk aşama her bir ebeveynin meydana getirebileceği gametleri belirlemektir. Her gamet, farklı her gene ait sadece bir alel içermelidir. Meydana gelen tüm kombinasyonları gösteren en basit yol dallanma yöntemidir. Homozigot olan lokus sadece bir tip alel meydana getirirken heterozigot olan lokus iki çeşit alel meydana getirir. Örneğin genotipi a+a; bb; c+c; dd olduğu varsayılan bir bireyin meydana getireceği gametleri bir diyagram çizerek gösterelim:
Gamet çeşitleri c+ d a+ b c+ d
___________ a+ b c d a+ b c d
c+ d a b c+ d a b
c d a b c d
Mümkün olan tüm kombinasyonlar formül uygulayarak kontrol edilebilir. Gamet çeşidi sayısı = 2n formülüyle hesaplanabilir. Burada n=heterozigot lokusların (dolayısıyla genlerin) sayısıdır. Yukarıdaki örneğe göre, dört lokus vardır, bunlardan sadece ikisi heterozigottur. Bu nedenle n=2 ve meydana gelecek olan gamet çeşidi sayısı 4'tür.
Yöntem:
1) Virjin dişiler kullanarak aşağıda belirtilen sinekler arasında resiprokal çaprazlamalar kurulur.Beyaz gözlü (w, 1. X kromozomu eşeye bağlı) ve körelmiş kanatlı (vg, 2. kromozom) Drosophila soyları kullanılır. Beyaz gözlü Drosophila kanat özelliği bakımından yabani forma sahip (wvg+) iken körelmiş kanatlı Drosophila göz rengi bakımından yabani forma (w+vg) sahiptir.2) 7 gün sonra ebeveynler (P1) uzaklaştırılır.3) F1 sineklerinin fenotipleri belirlenir.4) F2 neslini gözlemek için F1 dölleri kendi aralarında çaprazlanır. Bu çaprazlamada F1
dişilerinin virjin olması gerekir.5) 7 gün sonra bu çaprazlamanın ebeveynleri (P2) uzaklaştırılır.6) F2 yavruları görülmeye başlandığı zaman inceleme, sayım ve kayıt işlemleri başlatılır. F2
sinekleri fenotipik sınıflara ayrılarak sayılır ve istatistik analizi yapılır.

DENEY 5
Deneyin Adı: Bağlantılı Gen Dağılımı
Herhangi bir organizmadaki kromozom sayısı bireyin sahip olduğu gen sayısına göre oldukça azdır. Bu durum Drosophila gibi sadece birkaç çift kromozomu olan fakat kalıtsal özellikleri çok fazla olan organizmalarda daha fazla göze çarpar.
Bilindiği gibi kromozomlar bir bütün halinde kalıtılma eğilimindedirler. Genler kromozomlar üzerinde yer aldıklarına göre hücre bölünmesinde kromozomların davranışlarını izlerler. Mayoz sırasında homolog kromozomlar birbirlerinden ayrılırlar, dolayısıyla üzerinde bulunan genler de birbirine bağlı olarak kalıtılırlar. Fakat bu arada homolog kromozomlar arasında gerçekleşen krosing over'dan dolayı genetik yapının değişmesi ve rekombinasyonun meydana gelmesi söz konusudur. Fakat, mayozdaki ayrılım sırasında, belirli bazı aleller, aynı kromozom üzerinde birbirine yakın bulunduğundan, bir bağlantı grubu (linkaj) oluştururlar ve birlikte kalıtılma eğilimi gösterirler. Bir çift homologun ayrılımı diğer çift homologun ayrılımından etkilenmez. Bu nedenle, belli bir kromozom üzerinde bulunan alel çiftleri farklı bir linkaj grubunu taşıyan alel çiftlerinden bağımsız olarak ayrılacaktır.
Drosophila 'da her biri bir grup gen taşıyan 4 çift kromozom olduğuna göre bu genlerin bağlantı durumları da söz konusudur.
Yöntem: Bu deneyde X kromozomu üzerinde yer alan üç resesif mutant alel arasında bağlantı testi yapılacaktır. Bu amaçla yabani Drosophila soyu ile, beyaz gözlü, kanat ve vücut boyu küçülmüş ve çatallı kılları olan (wmf) mutant Drosophila soyu kullanılacaktır.
1) Virjin wmf dişileri, yabani soydan erkek sineklerle çaprazlanır.2) 7 gün sonra ebeveynler uzaklaştırılır.3) F1 dölleri ortaya çıktığı zaman 2 kültür şişesinde F2 çaprazı başlatılır.4) 7 gün sonra F1 sinekleri uzaklaştırılır.5) F2'de ortaya çıkan farklı fenotipler sayılır ve kaydedilir.6) Sonuçlar istatistiki olarak değerlendirilir.

DENEY 6
Deneyin Adı: Drosophila melanogaster 'de Politen Kromozomlar
Politen (Dev) kromozomlar, kromatid yığınlarını içeren özel bir kromozom tipi olup Diptera grubu sineklerin belirli dokularında karakteristiktir. Dev kromozomlar Drosophila larvalarının tükrük bezleri, orta barsak, malpighi tübülleri gibi metabolik aktivitenin yoğun olduğu organlarda gözlenir. Bu dokular, hücre sayısının artışıyla değil hücrenin genişlemesi yoluyla büyürler. Dev hücreler normal hücrelere göre daha fazla genetik materyale ihtiyaç duyarlar ve bu da dev kromozomun oluşumu ve dolayısı ile gen sayısının arttırımı ile büyük miktarlarda ve hızlı mRNA sentezi ile karşılanır. Dev kromozomlar "Endomitoz" denilen özel bir işlem sonucu meydana gelirler. Endomitozda kromozomlar çekirdek zarı parçalanmaksızın çoğalırlar. Böylece kromozom sayısı 2n'in katları şeklinde artarak poliploidi durumu oluştururlar. Kromozomlar ayrılma olmadan tekrar tekrar replike edilerek genişlerler ve ayrıca uzunlukları da artar. Yeni oluşan kromozomlar bir arada kalarak bir ünite (birim) oluştururlar. Bunun nedeni mitotik iğe bağlanma bölgesinin tam fonksiyonunu gerçekleştirememesi ve birçok DNA replikasyonu ile ortaya çıkan kromatidlerin ayrılamamasıdır. Bu politenin ya da çok iplikçikli yapının oluşumunu başlatır.
Dev kromozomlar, hücre bölünmesindeki normal hücrelerin kromozomlarına göre yaklaşık 100 kat daha kalın ve 100 kat daha uzundur. Dev kromozomların sentromerleri kromosenter denilen bölgede bir araya gelirler. Bu durum karşılıklı çekim yoluyla olur.
Dev kromozomlar düzenli ve çok belirgin bantlama modelleri gösterirler. Çeşitli kalınlıkta olan bantlar yüksek oranlarda DNA ve histon içerirler. Bantlar arasındaki alanlar interbant olarak adlandırılırlar ve DNA ile histonu çok az oranlarda içerirler. Bu bantların düzenleniş şekillerinin türe özel ve fenotipik karakterlerle bağlantılı olması bunların kromozom haritalarının hazırlanmasında kullanılmasına nedendir. Enine bantların sayısı D. melanogaster 'de 5000'e kadar ulaşır.
Dev kromozomların belirli bölgeleri özellikle aktif olduklarında tüm benzer DNA molekülleri bu bölgede fırça benzeri loplar oluştururlar. Lopların yan yana takımlar oluşturması "kromozomal puf" olarak adlandırılır. Farklı dokularda ya da aynı dokunun farklı gelişim aşamalarında pufların kromozomlar üzerinde yerleşim alanları farklıdır. Puflar hızlı RNA sentezinin olduğu, aktif genlerin yerleşimini gösteren yerlerdir. Puflar RNA sentezi için yüzeyi maksimum tutmak amacıyla dışa doğru loplar oluşturmuş DNA'nın yüzlerce kopyasını içerir.
Yöntem:
Dev kromozom yapısının mikroskopta iyi incelenmesi için larvaların iyi yetiştirilmiş olması gereklidir. Bu nedenle kültür şişesinde yumurtalar görüldükten sonra en geç 2 gün içinde ebeveynler uzaklaştırılır. Ayrıca, özellikle büyük tükrük bezi kromozomlarının preparasyonu için düşük sıcaklıklarda (16-18 °C) yetiştirilmiş larvalar kullanılır.1) Kültür şişesinde, besiyerinden ayrılarak şişenin duvarlarında yukarıya doğru tırmanmaya başlayan üçüncü instar dönemindeki larvalar; bir fırça yardımıyla üzerine bir damla % 0.7'lik tuz (NaCl) konulmuş bir lam üzerine alınır.2) Disseksiyon iğneleri ya da iki tane ince uçlu pens kullanılarak larvanın ağız parçalarının bulunduğu ön ucu hızlı bir hareketle ayrılır. Geri kalan 2/3'lük kısmı atılır (Bkz. Şekil 5).3) Tükrük bezi kurumadan üzerine aseto karmin (veya aseto orsein) boyası damlatılır.

4) Üç dakika kadar beklenerek kromozomların fikse edilmesi ve boyanmaları sağlanır. Sürenin daha uzun tutulması kromozomları daha kolay kırılır hale getirirken kısa oluşu da kromozom bant yapılarının boyanmamasına neden olur.5) Lam üzeri lamelle kapatılır, preparat bir parça filtre kağıdı ile sarılarak üzerine hafif birkaç darbe ile fiziksel basınç uygulanır. Böylece hücreler parçalanır ve kromozomlar kırılmadan açığa çıkabilir.6) Hazırlanan preparat binoküler mikroskopta incelenerek, dev kromozomlar belirlenir ve çizilir.
Aseto-Karmin Boyasının Hazırlanışı:
(1) 90 ml asetik asit 110 ml saf su ile seyreltilir.(2) Üzerine 1 g karmin boyası konularak orta derecedeki bir alevde 2 saat kaynatılır.(3) Kaynama sırasında buharlaşma önlenir. Daha sonra soğutularak boya filtre edilir.(4) Hazırlanan boya kahverengi şişede ağzı sıkıca kapatılarak saklanır.