Embed Size (px)
Citation preview

Journal of Medical Virology 82:783–790 (2010)
The Entire Core Protein of HCV JFH1 Is Requiredfor Efficient Formation of InfectiousJFH1 Pseudoparticles
Priyanka Shukla,1 Kristina N. Faulk,2 and Suzanne U. Emerson1*1Molecular Hepatitis Section, Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases,National Institutes of Health, Bethesda, Maryland2Hepatitis Viruses Section, Laboratory of Infectious Diseases, National Institute of Allergy and Infectious Diseases,National Institutes of Health, Bethesda, Maryland
The vast majority of hepatitis C virus (HCV)strains cannot be grown in cell culture. Therefore,tests for neutralizing antibodies have reliedheavily on retrovirus pseudoparticles displayingthe envelope glycoproteins of HCV on theirsurface (HCVpp). Unfortunately, the envelopeproteins of some strains, especially of JFH1, didnot efficiently form functional HCVpp. We havemanipulated the length and composition of theHCV core gene in the HCVpp expression vectorsfor three strains of HCV in an attempt to obtainmore efficient production of pseudoparticles. Theresults demonstrated that the truncated coreregion included in the HCV expression plasmidsof the classic pseudoparticle system was optimalfor formation of strain H77pp, suboptimal forstrain J6pp, and insufficient for strain JFH1pp.Efficiency of JFH1pp formation increased 20-foldwhen the truncated core gene was replaced withthe entire core gene. The full core from J6 and HKhad modest effect on the production of infectiousJ6 and HKpp. The data suggested that pairs ofHCV glycoproteins differ inherently in their abilityto associate into functional heterodimers andthat the core protein, provided in cis as thebeginning of the polyprotein product, can insome cases facilitate this process, possiblyby increasing the rate of proper folding ofthe glycoproteins. J. Med. Virol. 82:783–790, 2010. � 2010 Wiley-Liss, Inc.{
KEY WORDS: HCV pseudoparticles; coreenhancement; viral glycopro-tein assembly
INTRODUCTION
Hepatitis C is a serious disease noted for its extremelyhigh rate of chronicity; currently over 170 million peopleworldwide have chronic hepatitis C. In contrast to HIVinfections, which also exhibit a high rate of chronicity,
the immune system remains functional in hepatitis C. Infact, hepatitis C virus (HCV) persists in the presence ofhigh levels of antibodies to the two viral glycoproteins,E1 and E2 that are believed to comprise the viralreceptor [Eckels et al., 1996; Abrignani, 1997; Changet al., 1997]. For a long time, questions of why antibodiesto the glycoproteins did not eliminate the virus weredifficult to study since a cell culture system did notexist and the only animal model, the chimpanzee, wasprohibitively expensive. This changed with the recentdevelopment of the HCVpp and a comparable cellculture (HCVcc) system which provided a practicalmeans to identify and characterize antibodies thatreacted with the E1 and E2 glycoproteins of HCV[Bartosch et al., 2003; Hsu et al., 2003; Lindenbachet al., 2005; Wakita et al., 2005; Zhong et al., 2005].
HCV is an enveloped RNA virus belonging tothe genus Hepacivirus of the family Flaviviridae[Lindenbach and Rice, 2001]. Its positive-sense genomeof 9.6 kb encodes a single polyprotein that is co- and post-translationally processed by cellular and viral proteasesto yield three structural and seven non-structuralproteins [Penin et al., 2004]. The structural proteinsare located at the N-terminus and are translated in thefollowing order: capsid or core (c), E1 and then E2. Coreconsists of 191aa and its C-terminal 20aa serve as asignal sequence for E1 [Lo et al., 1995]. The HCVppsystems developed by Bartosch et al. [2003] and Hsuet al. [2003] are based on pseudoparticles bearing E1 and
Grant sponsor: Intramural Research Program of the NationalInstitutes of Health; Grant sponsor: National Institute of Allergyand Infectious Diseases.
*Correspondence to: Suzanne U. Emerson, MolecularHepatitis Section, LID, NIAID, NIH, Bldg. 50, Rm. 6537, 50 SouthDr., MSC 8009, Bethesda, MD 20892-8009.E-mail: [email protected]
Accepted 23 August 2009
DOI 10.1002/jmv.21660
Published online in Wiley InterScience(www.interscience.wiley.com)
� 2010 WILEY-LISS, INC.{This article is a US Government work and, as such, is inthe public domain in the United States of America.

E2 glycoproteins of HCV. In each case, pseudoparticlesare generated by co-transfecting a plasmid encodingtruncated core and full-length E1/E2 proteins with aplasmid or plasmids encoding a reporter gene (greenfluorescent protein, GFP or luciferase) and a retroviralpackaging system. The assembled pseudoparticlesreleased into the medium are able to infect culturedliver cells by virtue of their E1 and E2 components andthe efficiency of infection can be quantified by the level ofreporter gene expression. Although this system consistsmainly of non-HCV components, its use as a tool to studyneutralization of HCV was validated by the demonstra-tion that antibodies that neutralized HCV in thechimpanzee model neutralized HCVpp [Meunier et al.,2005].
In both HCVpp systems, the sequence encoding the C-terminal 20 or 60aa of core were included to provide thesignal sequence for E1: the C-terminus of E1, in turn,serves as the signal sequence for E2 [Cocquerel et al.,2000]. Therefore, the HCV portion of this system isrelatively easy to manipulate since it contains lessthan 2 kb of HCV genome and is located on a separateplasmid.
In the HCVcc system, autonomously replicatinginfectious virions are produced initially from a full-length recombinant HCV genome and then amplifiedby passage in hepatic cells [Zhong et al., 2005]. As faras is known, the viruses made in cell culture arestructurally equivalent to those in hepatitis C patientsand, therefore, would seem ideal for studying anti-HCV. However, the HCVcc system is much lessversatile than HCVpp for studying anti-E1 or E2because only a limited number of strains can be culturedcurrently. Initially, only the genotype 2a strainJFH1 replicated in cell culture [Wakita et al., 2005]. Alimited number of chimeric viruses expressing theglycoproteins of other strains in the JFH1 backbonehave been developed but each has had singular require-ments for viability and serial passage has usually beenrequired to select adaptive mutations [Lindenbachet al., 2005; Yi et al., 2006, 2007; Zhong et al., 2006;Delgrange et al., 2007; Kaul et al., 2007; Gottwein et al.,2009]. Thus generation of a diverse or expansive setof test viruses would be time consuming and laborintensive.
Much remains to be learned about the humoralresponse to HCV glycoproteins. Given the tremendousdiversity of HCV arising from the six genotypes,hundreds of subtypes, and a plethora of quasi-species,it is clear that a broad spectrum of viruses is required forcomprehensive assays of anti-E1 and E2. For thisreason, the HCVpp system has obvious advantages overthe HCVcc system because, in theory, virtually anyE1E2 glycoprotein pair that functions in vivo would beexpected to function in HCVpp. Although many naturalglycoprotein pairs formed functional pseudoparticles,others such as the glycoprotein pair of JFH1, theprototype virus of the HCVcc system, did not efficientlyassemble into functional HCVpp even though thesesame glycoproteins were incorporated into infectious
virions. The availability of a greater diversity of func-tional HCV glycoprotein pairs could be extremelyvaluable for determining structure–function relation-ships or immune evasion mechanisms.
Therefore, in this report we have attempted to answerthe perplexing question of why JFH1pp were so difficultto produce and whether the results were relevant forproduction of other pp and, therefore, might aid inincreasing the spectrum of glycoproteins available fortesting.
MATERIALS AND METHODS
Cell Culture
HCVpp stocks were produced in HEK-293T cells(ATCC), a human embryonic kidney cell line andHCVpp infections were performed in Huh-7.5 cells, ahuman hepatoma cell line (kind gift from C. Rice). All cellswere propagated in complete Dulbecco’s modified Eagle’smedium with L-glutamine (DMEM; Invitrogen, Carlsbad,CA) supplemented with penicillin/streptomycin (Sigma,St. Louis, MO) and 10% fetal bovine serum (Bio-Whittaker, Walkersville, MD). Cells were grown in100 mm culture dishes (Corning, Lowell, MA) at 378C inthe presence of 5% CO2.
Plasmid Constructs
The three plasmids used to generate HCVpp werefrom the Bartosch system and included the CMV-Gag-pol murine leukemia virus (MLV) packaging con-struct, a plasmid encoding an MLV-based transfervector containing a CMV-GFP internal transcriptionalunit, and the phCMV-7a expression vector encoding60aa from the C-terminus of core and all of E1 and E2from a genotype 1a strain (gift from F.L. Cosset). ThephCMV-7a plasmid was modified by standard cloningmethods to generate all other vectors used. All con-structs were verified by restriction digestion andsequencing. Detailed description of the cloning strategy,plasmids, and primer sequences are available uponrequest. The plasmid lacking E1 and E2 served as anegative control.
Generation of HCV Pseudoparticles
Three million HEK-293T cells were seeded in 100 mmflat culture dishes (Corning) and allowed to adhereovernight. Cells were transfected with LipofectaminePlus reagents (Invitrogen) as per manufacturer’s proto-col. A total of 2.5 mg of plasmid DNA including 0.75 mg ofCMV-Gag-pol packaging construct, 0.75 mg of MLV-GFPplasmid, and 1.0 mg of HCV core E1E2 expression vectorwere diluted in 250ml of serum-free DMEM. Eightmicroliters of Plus reagent was added to the DNAsolution and incubated at room temperature for 15 min.The DNA mix was added drop wise to 250 ml of serum-free DMEM containing 12 ml of Lipofectamine reagentand incubated at room temperature for 15 min. Cellswere washed twice and covered with 2 ml serum-
J. Med. Virol. DOI 10.1002/jmv
784 Shukla et al.
free DMEM. The DNA/Lipofectamine mixture wasadded to the cells and placed at 378C. Three hours later,the transfection mixture was replaced with completeDMEM and cells were cultured for 2 days at 378C.Transfection efficiency was consistently 80–90% basedon visual examination of GFP expression and FACSanalysis of GFP positive HEK-293T cells. Culturesupernatants were harvested and filtered through a0.45mm filter (Millipore, Billerica, MA) to remove cellsand debris. Infection of Huh-7.5 cells was performed onthe same day to avoid freeze-thawing of the virus stocks,and remaining culture supernatants were stored at�808C for subsequent analysis. An E1E2 negativecontrol and untransfected cells were also included ineach experiment: since in all cases, less than 0.04% ofthe control cells scored positive, they are shown only forFigure 1.
Infection of Huh-7.5 Cells With HCVPseudoparticles
Huh-7.5 cells were seeded at a concentration of 40,000cells per well in 24-well plates and cultured overnight.The next day, 200 ml of HCVpp-containing HEK-293Tcell transfection supernatant was mixed with 50 ml of5� Polybrene (Sigma) diluted in serum-free DMEM togive a final concentration of 4 mg/ml. Cell supernatantswere removed and the HCVpp/Polybrene mixturewas added to the cells and incubated for 5 hr at 378Cwith shaking every hour. The infection mixture wasreplaced with 1 ml complete DMEM. Four days later,culture supernatants were removed and the monolayerwas washed once with 300ml of 1� PBS (Invitrogen;pH 7.4), covered with 300ml Trypsin-Versene (Bio-Whittaker), and incubated at 378C for 15 min. Detachedcells were transferred to 1.5 ml Eppendorf tubes,washed with 1 ml PBS, resuspended in 300ml of PBSand placed on ice. For FACS analysis 20,000 cellswere analyzed for GFP expression using FACSscan(BD) with the settings: FSC, E-1 linear, amp gain 4–59;SSC, 360 linear, amp gain 1; FL1, 340 log. FACSanalysis of uninfected cells was performed as a negativecontrol.
Purification of HCVpp
HCVpp were purified from the 293T culture super-natant using 20% sucrose. Briefly, 4 ml of culturesupernatant was layered on 1 ml 20% sucrose andultracentrifuged in a SW55 rotor at 35,000 rpm for 1 hrat 48C. Purified pps were resuspended in 30 ml TN buffer[50 mM Tris (pH 8.0), 10 mM NaCl] and used forimmunoblotting analysis.
Immunoblotting Analysis of HCVppProtein Expression
Transfected HEK-293T cells were lysed at 48 hr post-transfection with 1� lysis buffer (Promega, Madison,WI) as per manufacturer’s protocol. Lysates and purifiedpp were separated on 4–12% Bis-Tris NuPage gels(Invitrogen) under reducing conditions. After electro-phoresis, proteins were transferred to PVDF mem-branes (Invitrogen), and blots were processed usingSNAP i.d. system (Millipore) as per manufacturer’sprotocol. Briefly, the membrane was blocked for 5 min atroom temperature with 0.5% skim milk in 1� PBS-T(0.1% Tween-20). For detection of JFH1 core protein, amonoclonal antibody which recognizes the N-terminusof JFH1 core (mouse a-core; Anogen, Ontario, Canada)was diluted 1:160 in PBS-T and incubated with the blotat room temperature for 10 min. For detection of JFH1E2, a rabbit antibody to H77 HVR1 [LMF 87, Farci et al.,1996] was used at 1:5,000 dilution and incubated atroom temperature for 10 min. After extensive washingwith PBS-T, the blots were incubated for 10 min at roomtemperature with an HRP-conjugated a-mouse(1:16,000) and a-rabbit (1:30,000) secondary antibodies.After additional washing, blots were developed with
J. Med. Virol. DOI 10.1002/jmv
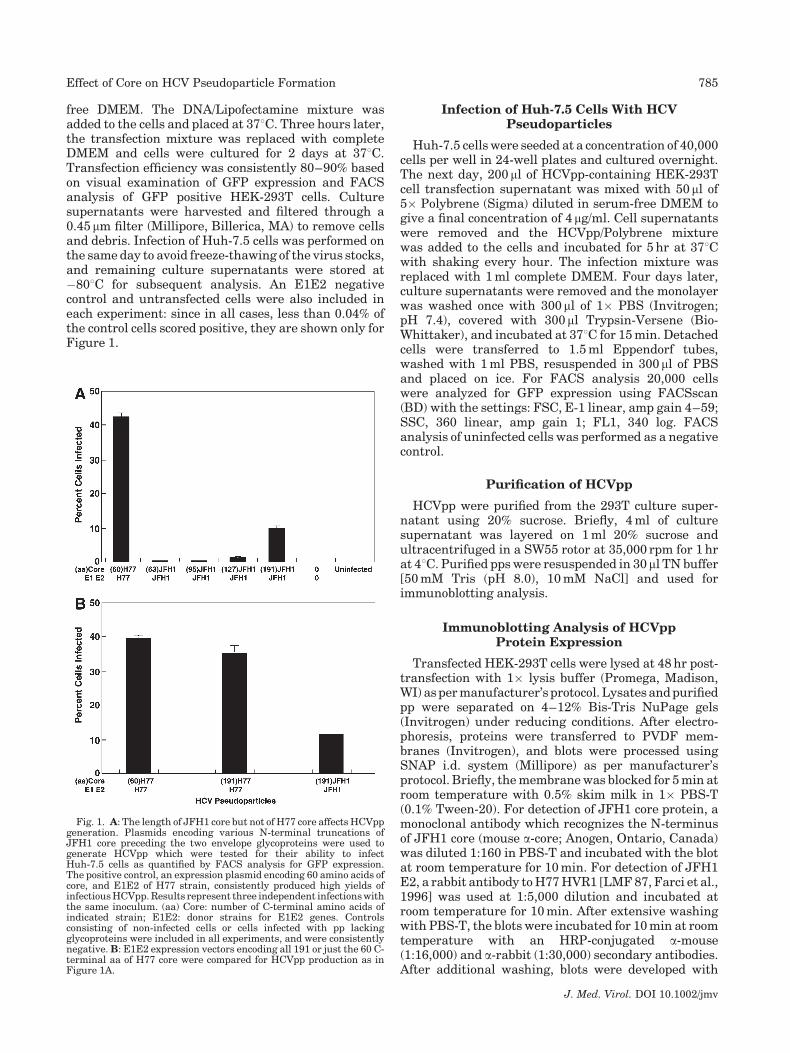
Fig. 1. A: The length of JFH1 core but not of H77 core affects HCVppgeneration. Plasmids encoding various N-terminal truncations ofJFH1 core preceding the two envelope glycoproteins were used togenerate HCVpp which were tested for their ability to infectHuh-7.5 cells as quantified by FACS analysis for GFP expression.The positive control, an expression plasmid encoding 60 amino acids ofcore, and E1E2 of H77 strain, consistently produced high yields ofinfectious HCVpp. Results represent three independent infections withthe same inoculum. (aa) Core: number of C-terminal amino acids ofindicated strain; E1E2: donor strains for E1E2 genes. Controlsconsisting of non-infected cells or cells infected with pp lackingglycoproteins were included in all experiments, and were consistentlynegative. B: E1E2 expression vectors encoding all 191 or just the 60 C-terminal aa of H77 core were compared for HCVpp production as inFigure 1A.
Effect of Core on HCV Pseudoparticle Formation 785
SuperSignal1 West Femto Chemiluminescent Sub-strate (Pierce, Rockford, IL).
RESULTS
Complete Core Gene EnhancesJFH1pp Production
The E1E2 expression plasmids in the pseudoparticleassays developed by Bartosch et al. [2003] and Hsu et al.[2003] encode only the last 60 or 20 C-terminal aminoacids, respectively, of the 191aa core protein. Althoughconstructs expressing similar lengths of truncated coreprotein have been used successfully to produce pseudo-particles representing multiple strains and genotypesof HCV, a JFH1 E1E2 expression plasmid encoding theC-terminal 63 amino acids of core produced negligiblequantities of pp (Fig. 1A). Since Merola et al. [2001] hadreported that the core protein could enhance correctfolding of E1 protein in vitro, vectors containing variablelengths of JFH1 core gene were constructed and testedto determine if they differed in efficiency of JFH1ppproduction. As a positive control, an expression plasmidencoding 60 amino acids of core and E1E2 of H77 strainwas included since it consistently produced high yieldsof infectious pp.
Increasing the size of core to encode all 191aa,rather than just the last 63, increased the yields ofJFH1 infectious pp over 20-fold (Fig. 1A). Shorterregions encoding the last 127 or 95 C-terminal aahad only a threefold or no effect, respectively (Fig. 1A).A C-terminal region of 159aa was approximately twiceas efficient as that with 127aa but was still fourfoldless efficient than the entire 191aa region (data notshown). The results demonstrated the importance ofcore sequence, or at least the NH2-terminal region, forJFH1pp generation. However, note that the yield ofH77pp generated in the presence of partial core was still4 times greater than that of JFH1 with complete core. Asexpected, Western blot analysis of pps did not detect anycore (data not shown).
The stimulation of JFH1 particle production by coreraised the question of whether H77 particle productioncould be increased by lengthening the core gene inthe H77 expression plasmid. E1E2 expression vectorsencoding all 191 amino acids or only the C-terminal60 amino acids of H77 core were compared for theirability to generate infectious H77pp (Fig. 1B). The twovectors produced similar results suggesting that therequirement for the core gene in pp formation differedbetween JFH1 and H77.
Core Protein, Rather Than Core Gene RNA,Enhances JFH1 Particle Production
The enhanced yield of infectious particles followingincorporation of the entire core gene into the plasmidregion directly preceding E1 could reflect a property ofthe core protein itself or, alternatively, it could be RNA-related and reflect greater mRNA stability or trans-latability. If the RNA itself was the critical variable,
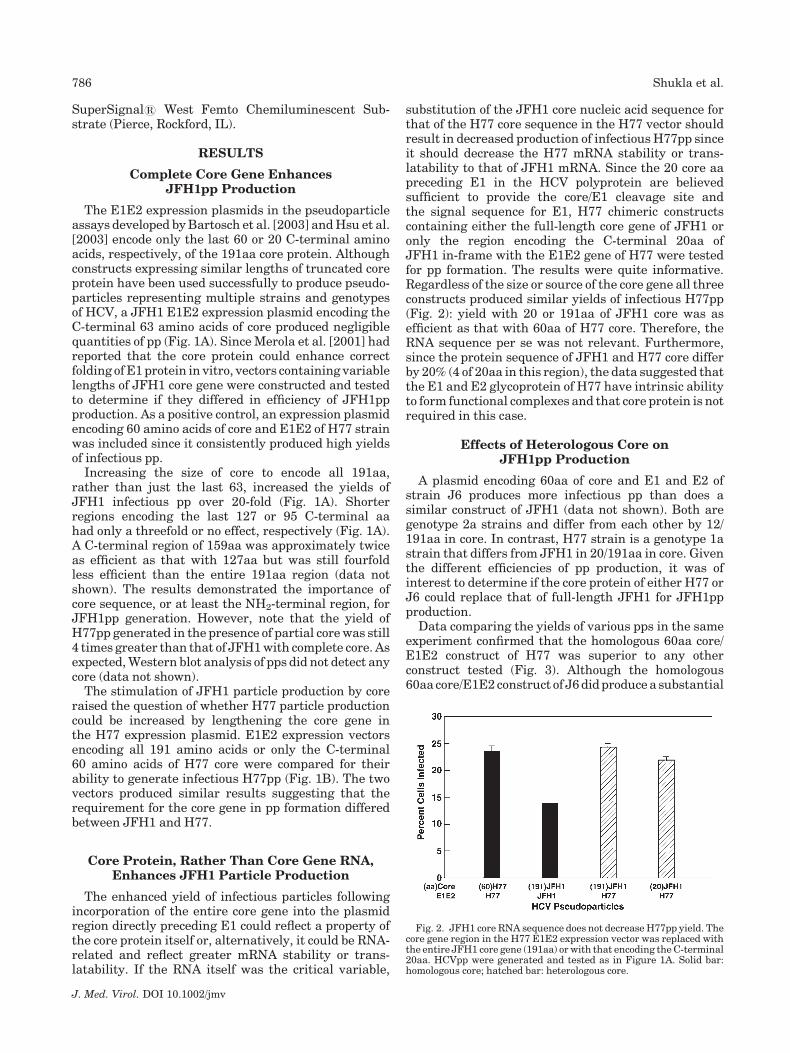
substitution of the JFH1 core nucleic acid sequence forthat of the H77 core sequence in the H77 vector shouldresult in decreased production of infectious H77pp sinceit should decrease the H77 mRNA stability or trans-latability to that of JFH1 mRNA. Since the 20 core aapreceding E1 in the HCV polyprotein are believedsufficient to provide the core/E1 cleavage site andthe signal sequence for E1, H77 chimeric constructscontaining either the full-length core gene of JFH1 oronly the region encoding the C-terminal 20aa ofJFH1 in-frame with the E1E2 gene of H77 were testedfor pp formation. The results were quite informative.Regardless of the size or source of the core gene all threeconstructs produced similar yields of infectious H77pp(Fig. 2): yield with 20 or 191aa of JFH1 core was asefficient as that with 60aa of H77 core. Therefore, theRNA sequence per se was not relevant. Furthermore,since the protein sequence of JFH1 and H77 core differby 20% (4 of 20aa in this region), the data suggested thatthe E1 and E2 glycoprotein of H77 have intrinsic abilityto form functional complexes and that core protein is notrequired in this case.
Effects of Heterologous Core onJFH1pp Production
A plasmid encoding 60aa of core and E1 and E2 ofstrain J6 produces more infectious pp than does asimilar construct of JFH1 (data not shown). Both aregenotype 2a strains and differ from each other by 12/191aa in core. In contrast, H77 strain is a genotype 1astrain that differs from JFH1 in 20/191aa in core. Giventhe different efficiencies of pp production, it was ofinterest to determine if the core protein of either H77 orJ6 could replace that of full-length JFH1 for JFH1ppproduction.
Data comparing the yields of various pps in the sameexperiment confirmed that the homologous 60aa core/E1E2 construct of H77 was superior to any otherconstruct tested (Fig. 3). Although the homologous60aa core/E1E2 construct of J6 did produce a substantial
J. Med. Virol. DOI 10.1002/jmv
Fig. 2. JFH1 core RNA sequence does not decrease H77pp yield. Thecore gene region in the H77 E1E2 expression vector was replaced withthe entire JFH1 core gene (191aa) or with that encoding the C-terminal20aa. HCVpp were generated and tested as in Figure 1A. Solid bar:homologous core; hatched bar: heterologous core.
786 Shukla et al.
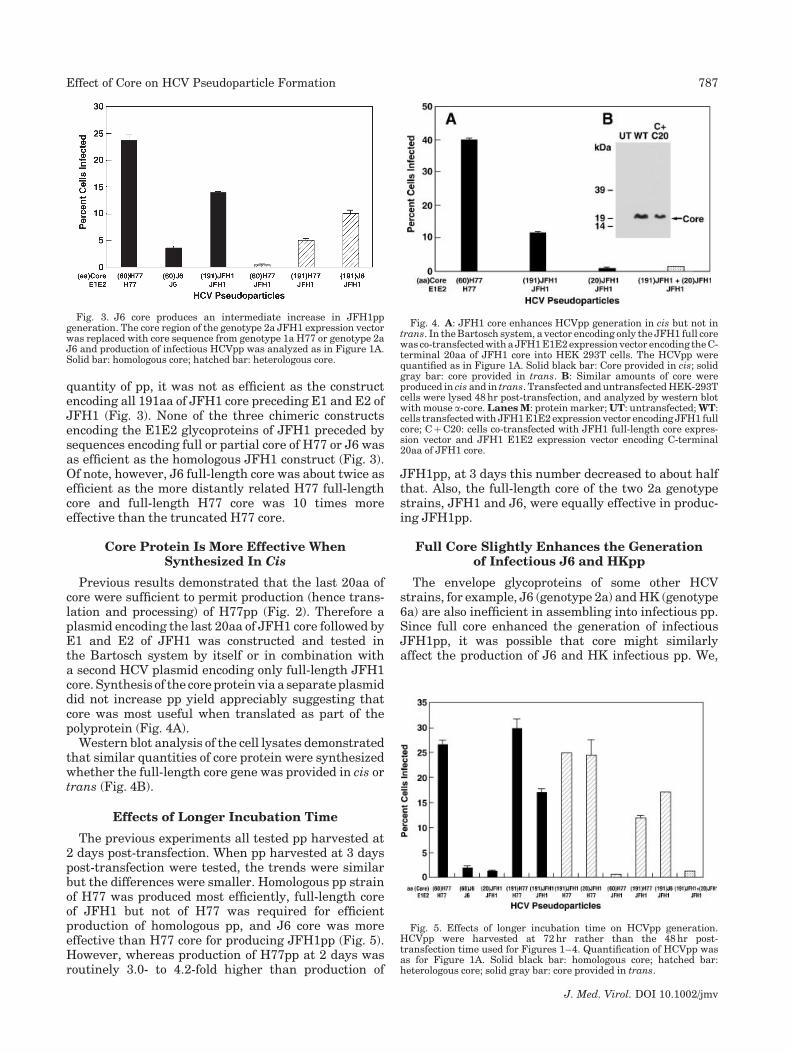
quantity of pp, it was not as efficient as the constructencoding all 191aa of JFH1 core preceding E1 and E2 ofJFH1 (Fig. 3). None of the three chimeric constructsencoding the E1E2 glycoproteins of JFH1 preceded bysequences encoding full or partial core of H77 or J6 wasas efficient as the homologous JFH1 construct (Fig. 3).Of note, however, J6 full-length core was about twice asefficient as the more distantly related H77 full-lengthcore and full-length H77 core was 10 times moreeffective than the truncated H77 core.
Core Protein Is More Effective WhenSynthesized In Cis
Previous results demonstrated that the last 20aa ofcore were sufficient to permit production (hence trans-lation and processing) of H77pp (Fig. 2). Therefore aplasmid encoding the last 20aa of JFH1 core followed byE1 and E2 of JFH1 was constructed and tested inthe Bartosch system by itself or in combination witha second HCV plasmid encoding only full-length JFH1core. Synthesis of the core protein via a separate plasmiddid not increase pp yield appreciably suggesting thatcore was most useful when translated as part of thepolyprotein (Fig. 4A).
Western blot analysis of the cell lysates demonstratedthat similar quantities of core protein were synthesizedwhether the full-length core gene was provided in cis ortrans (Fig. 4B).
Effects of Longer Incubation Time
The previous experiments all tested pp harvested at2 days post-transfection. When pp harvested at 3 dayspost-transfection were tested, the trends were similarbut the differences were smaller. Homologous pp strainof H77 was produced most efficiently, full-length coreof JFH1 but not of H77 was required for efficientproduction of homologous pp, and J6 core was moreeffective than H77 core for producing JFH1pp (Fig. 5).However, whereas production of H77pp at 2 days wasroutinely 3.0- to 4.2-fold higher than production of
JFH1pp, at 3 days this number decreased to about halfthat. Also, the full-length core of the two 2a genotypestrains, JFH1 and J6, were equally effective in produc-ing JFH1pp.
Full Core Slightly Enhances the Generationof Infectious J6 and HKpp
The envelope glycoproteins of some other HCVstrains, for example, J6 (genotype 2a) and HK (genotype6a) are also inefficient in assembling into infectious pp.Since full core enhanced the generation of infectiousJFH1pp, it was possible that core might similarlyaffect the production of J6 and HK infectious pp. We,
J. Med. Virol. DOI 10.1002/jmv
Fig. 3. J6 core produces an intermediate increase in JFH1ppgeneration. The core region of the genotype 2a JFH1 expression vectorwas replaced with core sequence from genotype 1a H77 or genotype 2aJ6 and production of infectious HCVpp was analyzed as in Figure 1A.Solid bar: homologous core; hatched bar: heterologous core.
Fig. 4. A: JFH1 core enhances HCVpp generation in cis but not intrans. In the Bartosch system, a vector encoding only the JFH1 full corewas co-transfected with a JFH1 E1E2 expression vector encoding the C-terminal 20aa of JFH1 core into HEK 293T cells. The HCVpp werequantified as in Figure 1A. Solid black bar: Core provided in cis; solidgray bar: core provided in trans. B: Similar amounts of core wereproduced in cis and in trans. Transfected and untransfected HEK-293Tcells were lysed 48 hr post-transfection, and analyzed by western blotwith mouse a-core. Lanes M: protein marker; UT: untransfected; WT:cells transfected with JFH1 E1E2 expression vector encoding JFH1 fullcore; CþC20: cells co-transfected with JFH1 full-length core expres-sion vector and JFH1 E1E2 expression vector encoding C-terminal20aa of JFH1 core.
Fig. 5. Effects of longer incubation time on HCVpp generation.HCVpp were harvested at 72 hr rather than the 48 hr post-transfection time used for Figures 1–4. Quantification of HCVpp wasas for Figure 1A. Solid black bar: homologous core; hatched bar:heterologous core; solid gray bar: core provided in trans.
Effect of Core on HCV Pseudoparticle Formation 787
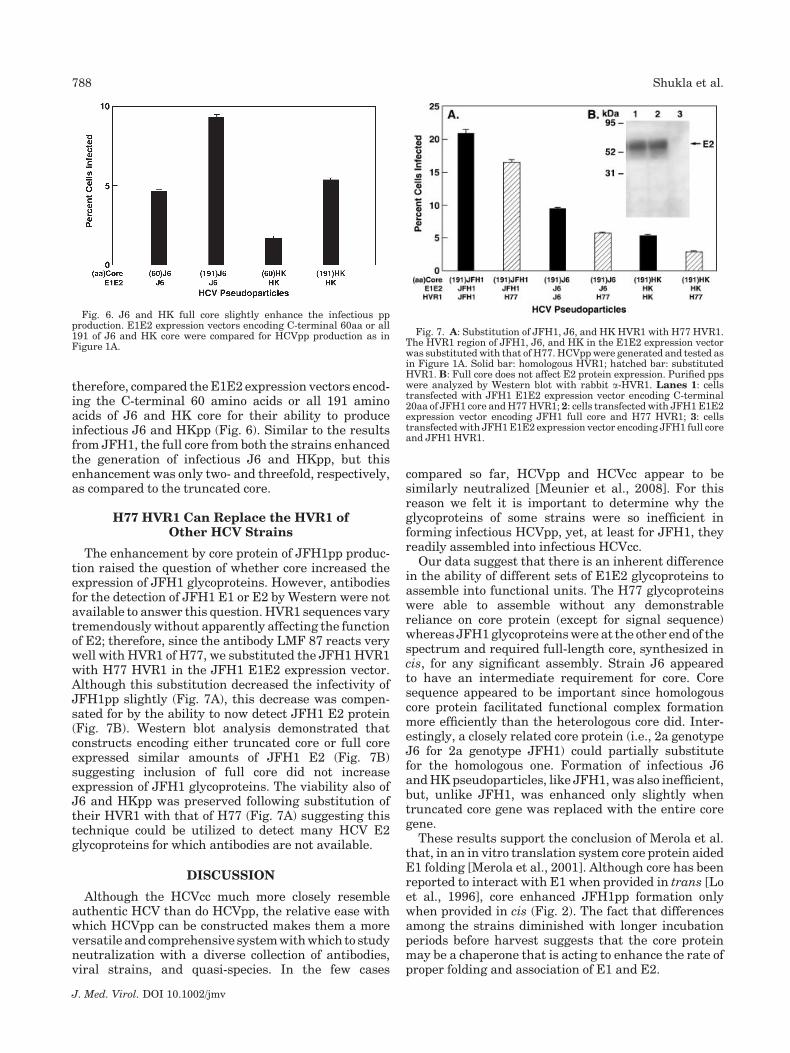
therefore, compared the E1E2 expression vectors encod-ing the C-terminal 60 amino acids or all 191 aminoacids of J6 and HK core for their ability to produceinfectious J6 and HKpp (Fig. 6). Similar to the resultsfrom JFH1, the full core from both the strains enhancedthe generation of infectious J6 and HKpp, but thisenhancement was only two- and threefold, respectively,as compared to the truncated core.
H77 HVR1 Can Replace the HVR1 ofOther HCV Strains
The enhancement by core protein of JFH1pp produc-tion raised the question of whether core increased theexpression of JFH1 glycoproteins. However, antibodiesfor the detection of JFH1 E1 or E2 by Western were notavailable to answer this question. HVR1 sequences varytremendously without apparently affecting the functionof E2; therefore, since the antibody LMF 87 reacts verywell with HVR1 of H77, we substituted the JFH1 HVR1with H77 HVR1 in the JFH1 E1E2 expression vector.Although this substitution decreased the infectivity ofJFH1pp slightly (Fig. 7A), this decrease was compen-sated for by the ability to now detect JFH1 E2 protein(Fig. 7B). Western blot analysis demonstrated thatconstructs encoding either truncated core or full coreexpressed similar amounts of JFH1 E2 (Fig. 7B)suggesting inclusion of full core did not increaseexpression of JFH1 glycoproteins. The viability also ofJ6 and HKpp was preserved following substitution oftheir HVR1 with that of H77 (Fig. 7A) suggesting thistechnique could be utilized to detect many HCV E2glycoproteins for which antibodies are not available.
DISCUSSION
Although the HCVcc much more closely resembleauthentic HCV than do HCVpp, the relative ease withwhich HCVpp can be constructed makes them a moreversatile and comprehensive system with which to studyneutralization with a diverse collection of antibodies,viral strains, and quasi-species. In the few cases
compared so far, HCVpp and HCVcc appear to besimilarly neutralized [Meunier et al., 2008]. For thisreason we felt it is important to determine why theglycoproteins of some strains were so inefficient informing infectious HCVpp, yet, at least for JFH1, theyreadily assembled into infectious HCVcc.
Our data suggest that there is an inherent differencein the ability of different sets of E1E2 glycoproteins toassemble into functional units. The H77 glycoproteinswere able to assemble without any demonstrablereliance on core protein (except for signal sequence)whereas JFH1 glycoproteins were at the other end of thespectrum and required full-length core, synthesized incis, for any significant assembly. Strain J6 appearedto have an intermediate requirement for core. Coresequence appeared to be important since homologouscore protein facilitated functional complex formationmore efficiently than the heterologous core did. Inter-estingly, a closely related core protein (i.e., 2a genotypeJ6 for 2a genotype JFH1) could partially substitutefor the homologous one. Formation of infectious J6and HK pseudoparticles, like JFH1, was also inefficient,but, unlike JFH1, was enhanced only slightly whentruncated core gene was replaced with the entire coregene.
These results support the conclusion of Merola et al.that, in an in vitro translation system core protein aidedE1 folding [Merola et al., 2001]. Although core has beenreported to interact with E1 when provided in trans [Loet al., 1996], core enhanced JFH1pp formation onlywhen provided in cis (Fig. 2). The fact that differencesamong the strains diminished with longer incubationperiods before harvest suggests that the core proteinmay be a chaperone that is acting to enhance the rate ofproper folding and association of E1 and E2.
J. Med. Virol. DOI 10.1002/jmv
Fig. 6. J6 and HK full core slightly enhance the infectious ppproduction. E1E2 expression vectors encoding C-terminal 60aa or all191 of J6 and HK core were compared for HCVpp production as inFigure 1A.
Fig. 7. A: Substitution of JFH1, J6, and HK HVR1 with H77 HVR1.The HVR1 region of JFH1, J6, and HK in the E1E2 expression vectorwas substituted with that of H77. HCVpp were generated and tested asin Figure 1A. Solid bar: homologous HVR1; hatched bar: substitutedHVR1. B: Full core does not affect E2 protein expression. Purified ppswere analyzed by Western blot with rabbit a-HVR1. Lanes 1: cellstransfected with JFH1 E1E2 expression vector encoding C-terminal20aa of JFH1 core and H77 HVR1; 2: cells transfected with JFH1 E1E2expression vector encoding JFH1 full core and H77 HVR1; 3: cellstransfected with JFH1 E1E2 expression vector encoding JFH1 full coreand JFH1 HVR1.
788 Shukla et al.

As the name hypervariable region implies, the HVR1region in the E2 protein exhibits a tremendous degree ofsequence variability in clinical specimens and has beenproposed to function as a decoy to focus the immuneresponse to non-critical regions. Indeed, Forns et al.[2000] showed that the HVR1 is not required forinfection of chimpanzees. Therefore, it seemed possiblethat the HVR1 of one strain could be substituted for thatof another strain while maintaining the structure–function relationships of E2. However, substitution ofJFH1, J6, or HK HVR1 with that of H77 HVR1 in therespective E1E2 expression vector resulted in a small,but measurable decrease in the infectivity of the ppsuggesting that HVR1 is not functionally inconsequen-tial but acts in concert with the rest of E2 in the stepsleading to infectious particle assembly or to cell entryand that sequence differences in HVR1 are not allrandom. Indeed, Bartosch et al. [2005] showed that non-conservative substitution of conserved amino acids inHVR1 decreases virus infectivity suggesting that theHVR1 plays an important role in virus infection. Thismay relate to the observation that HVR1 is essentialfor E2 binding to the scavenger receptor class B type I(SR-BI) [Scarselli et al., 2002]. Although HVR1 switch-ing is not without consequence, it should provide a morenatural change than insertion of a foreign tag epitopeand substitution of one HVR1 with another providesa minimally intrusive approach for antibody detection offunctional E2 of various strains. Such HVR1 swaps mayalso provide insight into exactly how HVR1 functions.These findings emphasize how much remains to bediscovered about the way in which HCV glycoproteinsand their different domains interact and promote cellentry.
A number of implications follow from these data. Firstthey suggest that production of HCVpp of perhaps manystrains might benefit from incorporation of full-lengthcore into the glycoprotein-expressing plasmid. Second,attempts to produce recombinant E1E2 complexes,whether for vaccine development or otherwise, mightachieve better success if core is included. Third,generation of HCVcc chimeras might depend on thesequence of core in ways not appreciated to date. Itseems reasonable to speculate that selection of muta-tions in core during adaptation to growth in cell culturemight represent an RNA packaging effect but itcould actually represent an advantage for glycoproteinfunction. Finally, these data suggest that studies ofthe interactions of HCV structural proteins mightprovide different results depending on which strainwas analyzed.
ACKNOWLEDGMENTS
This research was supported by the IntramuralResearch Program of the National Institutes of Health,National Institute of Allergy and Infectious Diseases.We are grateful to F.L. Cosset (Institut National de laSante et de la Recherche Medicale, France) and Charles
Rice (Rockefeller University, New York) for providingessential reagents.
REFERENCES
Abrignani S. 1997. Immune responses throughout hepatitis C virus(HCV) infection: HCV from the immune system point of view.Springer Semin Immunopathol 19:47–55.
Bartosch BJ, Dubuisson J, Cosset FL. 2003. Infectious hepatitis C viruspseudo-particles containing functional E1–E2 protein complexes.J Exp Med 197:633–642.
Bartosch BJ, Verney G, Dreux M, Donot P, Morice Y, Penin F,Pawlotsky JM, Lavillette D, Cosset FL. 2005. An interplay betweenhypervariable region 1 of the hepatitis C virus E2 glycoprotein, thescavenger receptor BI, and high density lipoprotein promotes bothenhancement of infection and protection against neutralizingantibodies. J Virol 282:32357–32369.
Chang KM, Rehermann B, Chisari FV. 1997. Immunopathology ofhepatitis C. Springer Semin Immunopathol 19:57–68.
Cocquerel L, Wychowski C, Minner F, Penin F, Dubuisson J. 2000.Charged residues in the transmembrane domains of hepatitis Cvirus glycoproteins play a major role in the processing, subcellularlocalization, and assembly of these envelope proteins. J Virol74:3623–3633.
Delgrange D, Pillez A, Castelain S, Cocquerel L, Rouille Y, Dubuisson J,Wakita T, Duverlie G, Wychowski C. 2007. Robust production ofinfectious viral particles in Huh-7 cells by introducing mutations inhepatitis C virus structural proteins. J Gen Virol 88:2495–2503.
Eckels D, Flomenberg P, Gill JC. 1996. Hepatitis C virus: Models ofimmunopathogenesis and prophylaxis. Transfusion 36:836–844.
Farci P, Shimoda A, Wong D, Cabezon T, De Gioannis D, Strazzera A,Shimizu Y, Shapiro M, Alter HJ, Purcell RH. 1996. Prevention ofhepatitis C virus infection in chimpanzees by hyperimmune serumagainst the hypervariable region 1 of the envelope 2 protein. ProcNatl Acad Sci USA 93:15394–15399.
Forns X, Thimme R, Govindarajan S, Emerson SU, Purcell RH, ChisariFV, Bukh J. 2000. Hepatitis C virus lacking the hypervariableregion 1 of the second envelope protein is infectious and causesacute resolving or persistent infection in chimpanzees. Proc NatlAcad Sci USA 97:13318–13323.
Gottwein JM, Scheel TK, Jensen TB, Lademann JB, Prentoe JC,Knudsen ML, Hoegh AM, Bukh J. 2009. Development andcharacterization of hepatitis C virus genotype 1–7 cell culturesystems: Role of CD81 and scavenger receptor class B type I andeffect of antiviral drugs. Hepatology 49:364–377.
Hsu M, Zhang J, Flint M, Logvinoff C, Cheng-Mayer C, Rice CM,McKeating JA. 2003. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc NatlAcad Sci USA 100:7271–7276.
Kaul A, Woerz I, Meuleman P, Leroux-Roels G, Bartenschlager R. 2007.Cell culture adaptation of hepatitis C virus and in vivo viability ofan adapted variant. J Virol 81:13168–13179.
Lindenbach BD, Rice CM. 2001. Flaviviridae: The viruses and theirreplication. In: Knipe DM, Howley PM, Griffin DE, Lamb RA,Martin MA, Roizman B, Straus SE, editors. Fields virology, 4thedition. Philadelphia, PA: Lippincott Williams & Wilkins. pp 991–1041.
Lindenbach BD, Evans MJ, Syder AJ, Wolk B, Tellinghuisen TL, LiuCC, Maruyama T, Hynes RO, Burton DR, McKeating JA, Rice CM.2005. Complete replication of hepatitis C virus in cell culture.Science 309:623–626.
Lo S, Masiarz YF, Hwang SB, Lai MM, Ou JH. 1995. Differentialsubcellular localization of hepatitis C virus core gene products.Virology 213:455–461.
Lo SY, Selby MJ, Ou JH. 1996. Interaction between hepatitis C viruscore protein and E1 envelope protein. J Virol 70:5177–5182.
Merola M, Brazzoli M, Cocchiarella F, Heile JM, Helenius A, WeinerAJ, Houghton M, Abrignani S. 2001. Folding of hepatitis C virus E1glycoprotein in a cell-free system. J Virol 75:11205–11217.
Meunier JC, Engle RE, Faulk K, Zhao M, Bartosch B, Alter H, EmersonSU, Cosset FL, Purcell RH, Bukh J. 2005. Evidence for cross-genotype neutralization of hepatitis C virus pseudo-particles andenhancement of infectivity by apolipoprotein C1. Proc Natl Acad SciUSA 102:4560–4565.
Meunier JC, Russell RS, Goossens V, Priem S, Walter ED, Union A,Faulk KN, Bukh J, Emerson SU, Purcell RH. 2008. Isolation and
J. Med. Virol. DOI 10.1002/jmv
Effect of Core on HCV Pseudoparticle Formation 789

characterization of broadly neutralizing human monoclonal anti-bodies to E1 glycoprotein of hepatitis C virus. J Virol 82:966–973.
Penin F, Dubuisson J, Rey FA, Moradpour D, Pawlotsky JM. 2004.Structural biology of hepatitis C virus. Hepatology 39:5–19.
Scarselli E, Ansuini H, Cerino R, Roccasecca RM, Acali S, Filocamo G,Traboni C, Nicosia A, Cortese R, Vitelli A. 2002. The humanscavenger receptor class B type I is a novel candidate receptor forthe hepatitis C virus. EMBO J 21:5017–5025.
Wakita T, Pietschmann T, Kato T, Date T, Miyamoto M, Zhao Z,Murthy K, Habermann A, Krausslich HG, Mizokami M, Bartens-chlager R, Liang TJ. 2005. Production of infectious hepatitis C virusin tissue culture from cloned viral genome. Nat Med 11:791–796.
Yi M, Villanueva RA, Thomas DL, Wakita T, Lemon SM. 2006.Production of infectious genotype 1a hepatitis C virus (Hutchinson
strain) in cultured human hepatoma cells. Proc Natl Acad Sci USA103:2310–2315.
Yi M, Ma Y, Yates J, Lemon SM. 2007. Compensatory mutations in E1,p7, NS2, and NS3 enhance yields of cell culture-infectiousintergenotypic chimeric hepatitis C virus. J Virol 81:629–638.
Zhong J, Gastaminza P, Cheng G, Kapadia S, Kato T, Burton DR,Wieland SF, Uprichard SL, Wakita T, Chisari FV. 2005. Robusthepatitis C virus infection in vitro. Proc Natl Acad Sci USA102:9294–9299.
Zhong J, Gastaminza P, Chung J, Stamataki Z, Isogawa M, Cheng G,McKeating JA, Chisari FV. 2006. Persistent hepatitis C virusinfection in vitro: Coevolution of virus and host. J Virol 80:11082–11093.
J. Med. Virol. DOI 10.1002/jmv
790 Shukla et al.





























