Embed Size (px)
Citation preview

J. Anat. (1992) 180, pp. 401-417, with 13 figures Printed in Great Britain
The morphology and innervation of facial vibrissae in thetammar wallaby, Macropus eugenii
L. R. MAR OTTE', F. L. RICE2 AND P. M. E. WAITE3
'Developmental Neurobiology, Research School of Biological Sciences, Australian National University, Canberra, Australia,2Department of Anatomy, Albany Medical College of Union University, Albany, New York, USA, andI School of Anatomy, University of New South Wales, Sydney, Australia
(Accepted 28 January 1992)
ABSTRACT
The morphology of the vibrissal follicles on the mystacial pad of the tammar wallaby is similar to that seenin other species except that the follicles lack a ringwulst or ring sinus. Instead, the mesenchymal sheath isthickened around the central region of the hair shaft. The follicle is innervated by both deep and superficialvibrissal nerves. The deep nerve enters as 4-11 fascicles which can be in close proximity or widelydistributed around the hair. C1 follicles received more myelinated nerve fibres (252+31) than the smaller C4follicles (174+43). The deep vibrissal nerve supplies the thickened mesenchymal sheath, the narrow 'waist'region above and the majority of endings in the inner conical body (ICB), while the superficial nervesprovide a sparse innervation to the ICB and rete ridge. Receptors present in the follicle were of 4 types: (1)Merkel cells, especially numerous in the outer root sheath of the 'waist' region and occasionally in the ICBand rete ridge; (2) and (3) lanceolate and lamellated endings parallel to the hair shaft in both themesenchymal thickening and the 'waist' region where they were particularly dense; (4) free nerve endings inthe mesenchymal thickening, 'waist' region and ICB. No corpuscular, bulbous or Ruffini endings were seen.The innervation of the intervibrissal fur was similar to that described in other species.
INTRODUCTION
Specialised facial hairs, or vibrissae, are a charac-teristic feature of most mammals (Pocock, 1914). Thehairs are commonly found on the mystacial pad,where they are arranged in 5 or more horizontal lines,although they also occur at other sites such as aroundthe eye and ear and at the wrist. The hairs arise froma complex follicle in which the hair and root sheath liein a blood sinus surrounded by a connective tissuecapsule. The morphology and innervation of thevibrissal follicle has been most extensively studied inthe mystacial vibrissae of rodents (Vincent, 1913;Melaragno & Montagna, 1953; Andres, 1966; Patrizi& Munger, 1966; Renehan & Munger, 1986). Eachvibrissa is innervated by a deep vibrissal nerve arisingfrom the infraorbital nerve (Dorfl, 1985) as well as byseveral small superficial nerves (also called conusnerves) which arise from the dermal plexus. Theinnervation is densest for the larger, more caudalvibrissae with about 200 myelinated and 100 un-myelinated fibres in each deep nerve (Waite & Cragg,
1982; Welker & Van der Loos, 1986; Klein et al. 1988;Crissman et al. 1991). These afferents supply a varietyof receptors such as Merkel cells, lanceolate terminalsand free nerve endings (Andres, 1966; Renehan &Munger, 1986).
Vibrissae have attracted attention in recent yearsbecause of their association with cortical barrels(Woolsey & Van der Loos, 1970) and their importancein normal barrel development (Van der Loos &Woolsey, 1973). For barrel formation an importantfactor is peripheral innervation density, with largercaudal whiskers associated with larger cortical barrels(Lee & Woolsey, 1975; Welker & Van der Loos,1986). Barrel-like aggregations are also present in 2Australian marsupials, the brush-tailed possum(Weller, 1972) and the tammar wallaby (Waite et al.1991). However, the aggregations in the wallaby differfrom those in the rodent in that they are apparent onlywith succinic dehydrogenase histochemistry not withthe common Nissl stains. Also, in the wallaby,aggregations are most clearly seen in the immatureanimal and are hard to discern in the adult. It was
Correspondence to Dr P. Waite, School of Anatomy, University of New South Wales, Kensington, Sydney 2033, Australia.
401
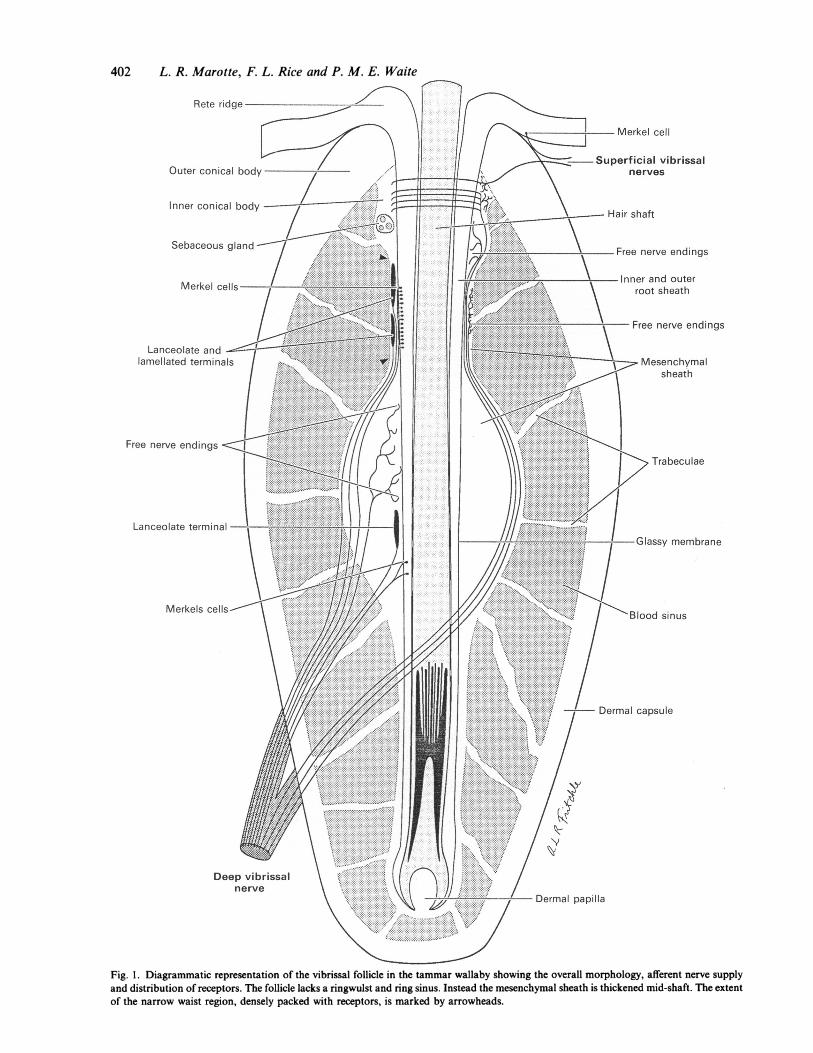
402 L. R. Marotte, F. L. Rice and P. M. E. Waite
Fig. 1. Diagrammatic representation of the vibrissal follicle in the tammar wallaby showing the overall morphology, afferent nerve supplyand distribution of receptors. The follicle lacks a ringwulst and ring sinus. Instead the mesenchymal sheath is thickened mid-shaft. The extentof the narrow waist region, densely packed with receptors, is marked by arrowheads.
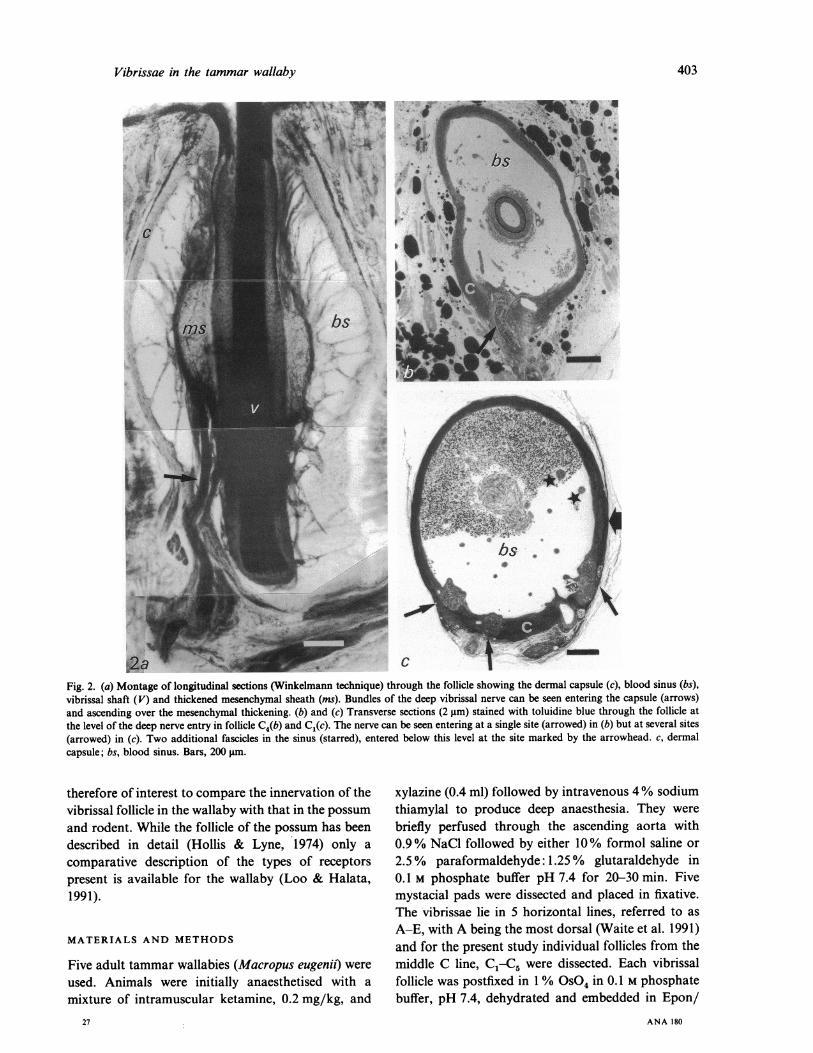
Vibrissae in the tammar wallaby
W.a w >+ c ~ 1 -Fig. 2. (a) Montage of longitudinal sections (Winkelmann technique) through the follicle showing the dermal capsule (c), blood sinus (bs),vibrissal shaft (V) and thickened mesenchymal sheath (ins). Bundles of the deep vibrissal nerve can be seen entering the capsule (arrows)and ascending over the mesenchymal thickening. (b) and (c) Transverse sections (2 im) stained with toluidine blue through the follicle atthe level of the deep nerve entry in follicle C4(b) and Cl(c). The nerve can be seen entering at a single site (arrowed) in (b) but at several sites(arrowed) in (c). Two additional fascicles in the sinus (starred), entered below this level at the site marked by the arrowhead. c, dermalcapsule; bs, blood sinus. Bars, 200 jgm.
therefore of interest to compare the innervation of thevibrissal follicle in the wallaby with that in the possumand rodent. While the follicle of the possum has beendescribed in detail (Hollis & Lyne, 1974) only acomparative description of the types of receptorspresent is available for the wallaby (Loo & Halata,1991).
MATERIALS AND METHODS
Five adult tammar wallabies (Macropus eugenii) wereused. Animals were initially anaesthetised with amixture of intramuscular ketamine, 0.2 mg/kg, and
27
xylazine (0.4 ml) followed by intravenous 4% sodiumthiamylal to produce deep anaesthesia. They werebriefly perfused through the ascending aorta with0.9% NaCl followed by either 10% formol saline or2.5 % paraformaldehyde: 1.25 % glutaraldehyde in0.1 M phosphate buffer pH 7.4 for 20-30 min. Fivemystacial pads were dissected and placed in fixative.The vibrissae lie in 5 horizontal lines, referred to asA-E, with A being the most dorsal (Waite et al. 1991)and for the present study individual follicles from themiddle C line, C1-C5 were dissected. Each vibrissalfollicle was postfixed in 1 % OS04 in 0.1 M phosphatebuffer, pH 7.4, dehydrated and embedded in Epon/
ANA 180
403

404 L. R. Marotte, F. L. Rice and P. M. E. Waite
Araldite. Transverse and longitudinal sections 2 gmthick were cut through the follicle and stained withtoluidine blue. Counts of the number of myelinatedaxons in the deep nerve were made for vibrissae C1and C4 in 5 mystacial pads from 4 animals. For this,photographs of each of the fascicles of the deepvibrissal nerve were taken at x 400 magnification,immediately after each fascicle entered the capsule. Atthis level axons were cut in cross-section.
For the electron microscopic investigations, goldand silver ultrathin sections were cut transversely andlongitudinally at selected levels in the C1 follicle,stained with uranyl acetate and lead citrate andexamined in a Hitachi H600 microscope.
For Bodian staining, follicles fixed in formol salinewere dehydrated, embedded in wax, sectioned longi-tudinally at 10 or 20 jm and stained (Bodian, 1936).For the Winkelmann reduced silver technique, folliclesfixed in formol saline were cut frozen at 100 gim invarious planes (Winkelmann, 1960).
RESULTS
The vibrissal follicle
As in other mammals, the vibrissal follicle (Figs 1, 2 a)consists of a hair shaft and associated root sheathsurrounded by a blood-filled sinus; the whole complexis enclosed by a thick dermal capsule of connectivetissue. Outside the outer root sheath is a thickenedbasement membrane, the glassy membrane and,surrounding this, a mesenchymal sheath. In contrastto the follicles in most other species, the follicle of thewallaby lacks a subdivision of the blood sinus into aring and cavernous sinus. The wallaby follicle alsolacks a 'ringwulst', the dense collar of connectivetissue which is usually seen around the mesenchymalsheath in the ring sinus. Instead the mesenchymalsheath forms a spindle-shaped thickening around thecentral region of the follicle. Above this the mes-enchymal sheath narrows dramatically before flaringout below the neck of the capsule to form a smallinner conical body (ICB). Trabeculae radiating fromthe mesenchymal sheath, primarily above and belowthe thickened region, traverse the blood sinus. Theupper trabeculae reach the top of the capsule while thelower ones pass from the hair bulb to the base of thecapsule.
Nerve supply
Each follicle receives a deep vibrissal nerve whichenters the capsule in the lower third of the follicle. The
Table 1. Numbers of myelinated axons in the deep vibrissalnerve for vibrissae C1 and C4
Vibrissa C1 Vibrissa C4
Number NumberAnimal of axon Number of axon Numberand side bundles of axons bundles of axons
I R 7 248 4 220L 11 269 4 173
2R 6 2113 L 7 292 5 1854L 8 238 4 116Mean+s.D. 252+31 174+43
/
Fig. 3. Montage of longitudinal sections (Winkelmann) throughthe inner conical body (icb), narrow waist region (w) and thickenedmesenchymal sheath (ms). Note the meandering afferents within themesenchymal thickening and the Merkel endings in the waistregion. Both the deep nerve (arrows) and superficial nerve(arrowheads) can be seen to contribute to the inner conical body.Bar, 100 gim.
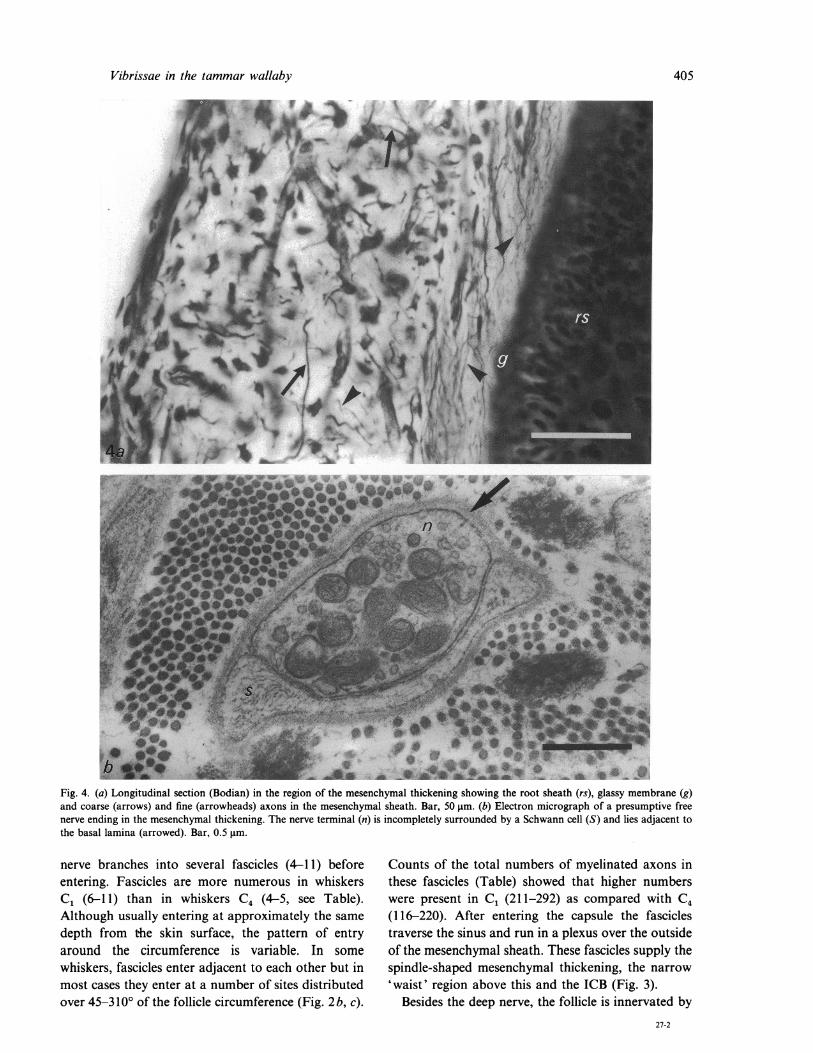
Vibrissae in the tammar wallaby
4
'
a#'~~~~~~~~-.< fw, ~_,t*_ *__
Fig. 4. (a) Longitudinal section (Bodian) in the region of the mesenchymal thickening showing the root sheath (rs), glassy membrane (g)and coarse (arrows) and fine (arrowheads) axons in the mesenchymal sheath. Bar, 50 gm. (b) Electron micrograph of a presumptive freenerve ending in the mesenchymal thickening. The nerve terminal (n) is incompletely surrounded by a Schwann cell (S) and lies adjacent tothe basal lamina (arrowed). Bar, 0.5 pmm.
nerve branches into several fascicles (4-11) beforeentering. Fascicles are more numerous in whiskersC1 (6-11) than in whiskers C4 (4-5, see Table).Although usually entering at approximately the samedepth from the skin surface, the pattern of entryaround the circumference is variable. In somewhiskers, fascicles enter adjacent to each other but inmost cases they enter at a number of sites distributedover 45-3100 of the follicle circumference (Fig. 2 b, c).
Counts of the total numbers of myelinated axons inthese fascicles (Table) showed that higher numberswere present in C1 (211-292) as compared with C4(116-220). After entering the capsule the fasciclestraverse the sinus and run in a plexus over the outsideof the mesenchymal sheath. These fascicles supply thespindle-shaped mesenchymal thickening, the narrow'waist' region above this and the ICB (Fig. 3).
Besides the deep nerve, the follicle is innervated by27-2
405
& ir 1# oh-
4.k., .*2Fz*;;W_zz
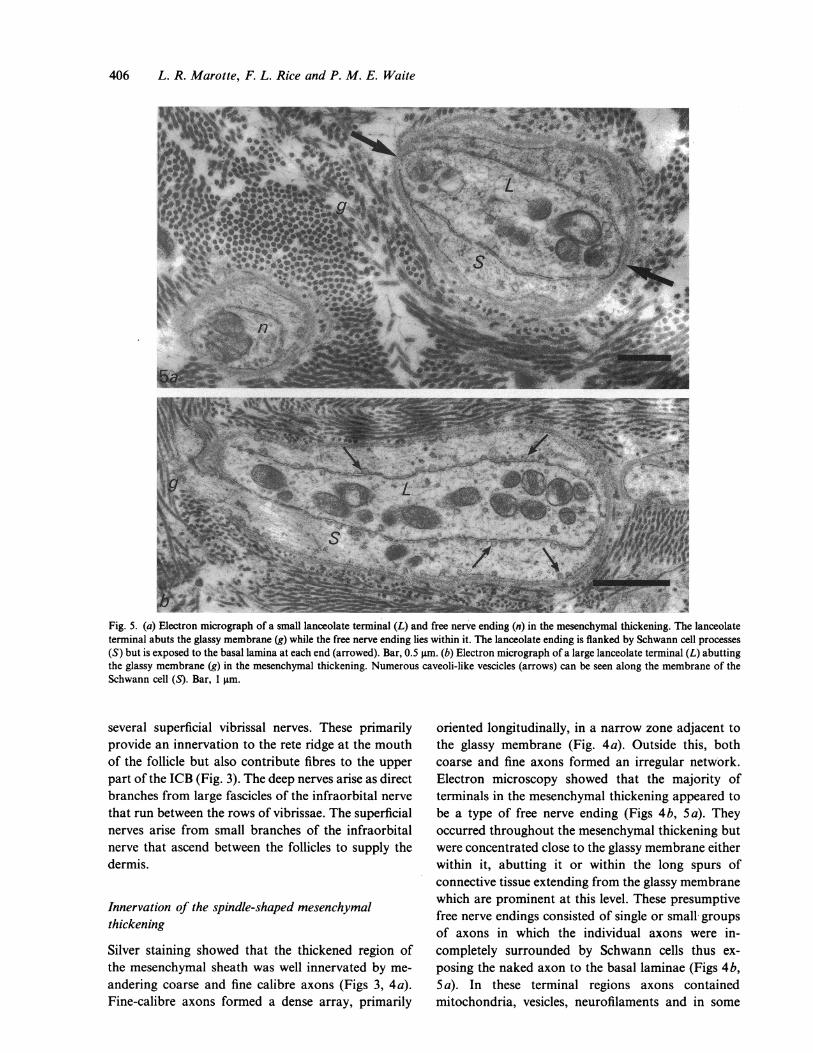
406 L. R. Marotte, F. L. Rice and P. M. E. Waite
S. ~~~~~ ~ ~~ ~ ~~~~ ~ ~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
Fig. 5. (a) Electron micrograph of a small lanceolate terminal (L) and free nerve ending (n) in the mesenchymal thickening. The lanceolateterminal abuts the glassy membrane (g) while the free nerve ending lies within it. The lanceolate ending is flanked by Schwann cell processes(S) but is exposed to the basal lamina at each end (arrowed). Bar, 0.5 gm. (b) Electron micrograph of a large lanceolate terminal (L) abuttingthe glassy membrane (g) in the mesenchymal thickening. Numerous caveoli-like vescicles (arrows) can be seen along the membrane of theSchwann cell (S). Bar, 1 gtm.
several superficial vibrissal nerves. These primarilyprovide an innervation to the rete ridge at the mouthof the follicle but also contribute fibres to the upperpart of the ICB (Fig. 3). The deep nerves arise as directbranches from large fascicles of the infraorbital nervethat run between the rows of vibrissae. The superficialnerves arise from small branches of the infraorbitalnerve that ascend between the follicles to supply thedermis.
Innervation of the spindle-shaped mesenchymalthickening
Silver staining showed that the thickened region ofthe mesenchymal sheath was well innervated by me-andering coarse and fine calibre axons (Figs 3, 4a).Fine-calibre axons formed a dense array, primarily
oriented longitudinally, in a narrow zone adjacent tothe glassy membrane (Fig. 4a). Outside this, bothcoarse and fine axons formed an irregular network.Electron microscopy showed that the majority ofterminals in the mesenchymal thickening appeared tobe a type of free nerve ending (Figs 4b, 5 a). Theyoccurred throughout the mesenchymal thickening butwere concentrated close to the glassy membrane eitherwithin it, abutting it or within the long spurs ofconnective tissue extending from the glassy membranewhich are prominent at this level. These presumptivefree nerve endings consisted of single or small groupsof axons in which the individual axons were in-completely surrounded by Schwann cells thus ex-posing the naked axon to the basal laminae (Figs 4b,5 a). In these terminal regions axons containedmitochondria, vesicles, neurofilaments and in some
Vibrissae in the tammar wallaby
6a _
-.. v- ,N-.
# |;::w _~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~V.
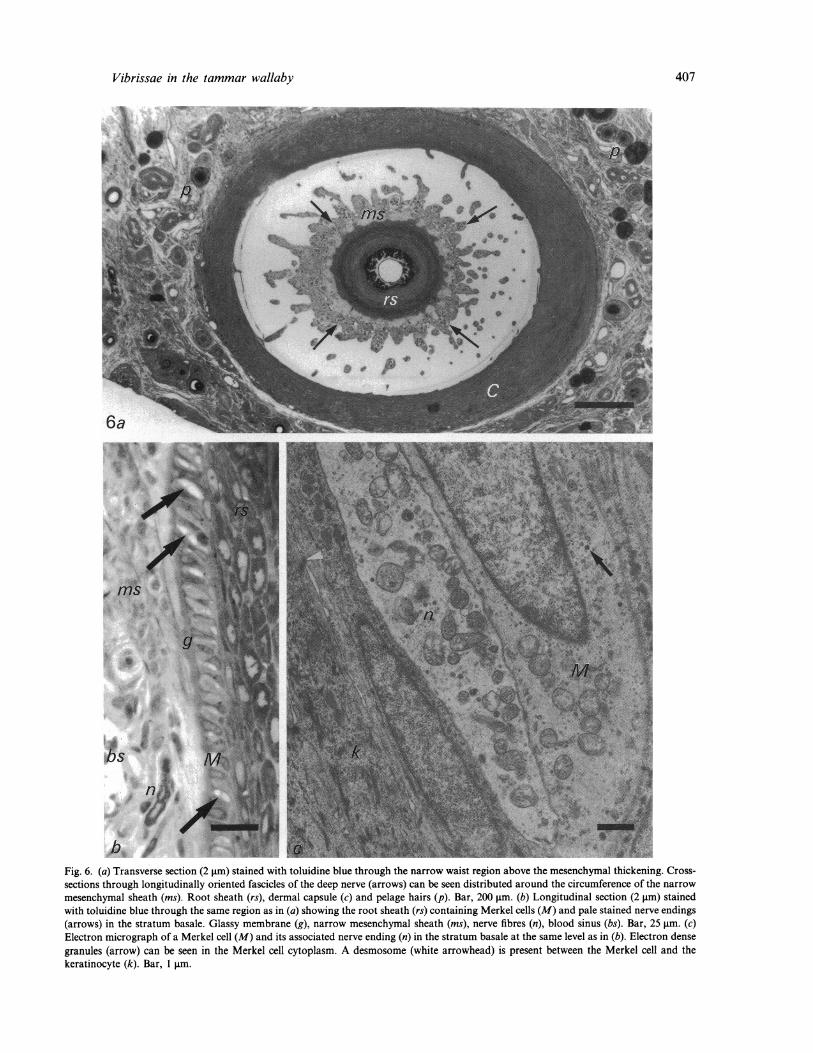
Fig. 6. (a) Transverse section (2 gim) stained with toluidine blue through the narrow waist region above the mesenchymal thickening. Cross-sections through longitudinally oriented fascicles of the deep nerve (arrows) can be seen distributed around the circumference of the narrow
mesenchymal sheath (ms). Root sheath (rs), dermal capsule (c) and pelage hairs (p). Bar, 200 jim. (b) Longitudinal section (2 jm) stainedwith toluidine blue through the same region as in (a) showing the root sheath (rs) containing Merkel cells (M) and pale stained nerve endings(arrows) in the stratum basale. Glassy membrane (g), narrow mesenchymal sheath (ms), nerve fibres (n), blood sinus (bs). Bar, 25 gim. (c)Electron micrograph of a Merkel cell (M) and its associated nerve ending (n) in the stratum basale at the same level as in (b). Electron densegranules (arrow) can be seen in the Merkel cell cytoplasm. A desmosome (white arrowhead) is present between the Merkel cell and thekeratinocyte (k). Bar, 1 pim.
407
408 L. R. Marotte, F. L. Rice and P. M. E. Waite
o* 4- *s /tts {
4f e, /{ WII
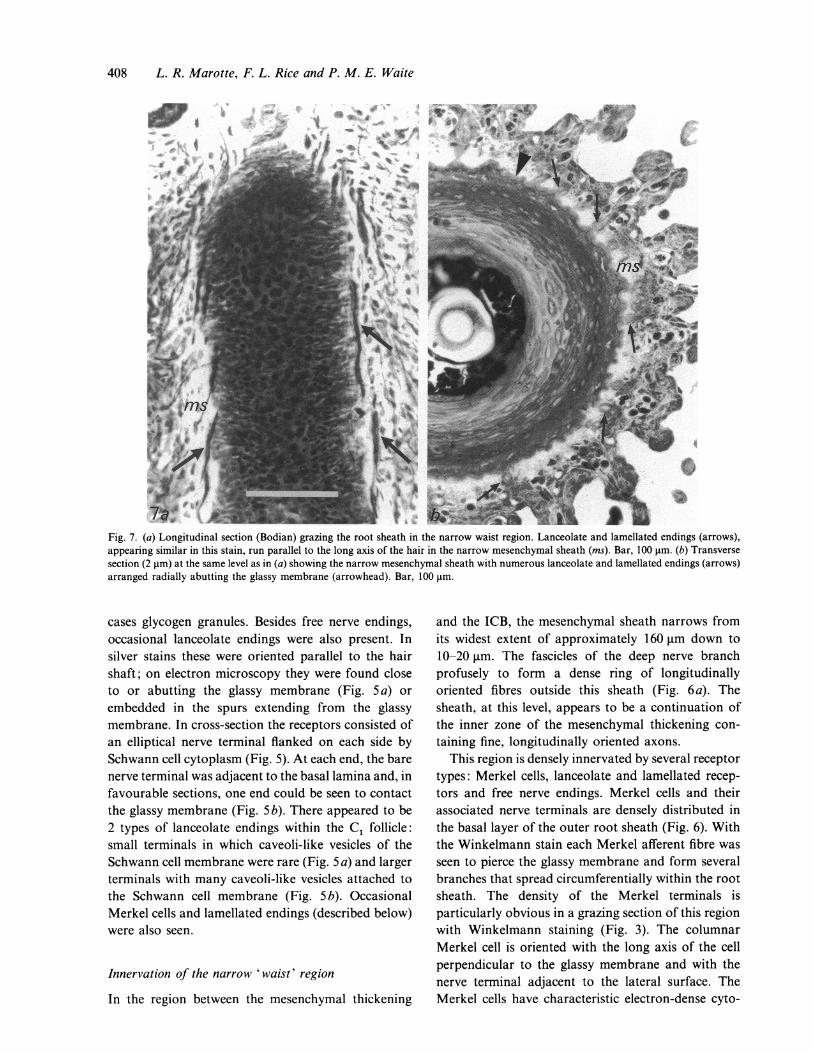
Fig. 7. (a) Longitudinal section (Bodian) grazing the root sheath in the narrow waist region. Lanceolate and lamellated endings (arrows),appearing similar in this stain, run parallel to the long axis of the hair in the narrow mesenchymal sheath (ins). Bar, 100 gim. (b) Transversesection (2 gim) at the same level as in (a) showing the narrow mesenchymal sheath with numerous lanceolate and lamellated endings (arrows)arranged radially abutting the glassy membrane (arrowhead). Bar, 100 gim.
cases glycogen granules. Besides free nerve endings,occasional lanceolate endings were also present. Insilver stains these were oriented parallel to the hairshaft; on electron microscopy they were found closeto or abutting the glassy membrane (Fig. 5 a) orembedded in the spurs extending from the glassymembrane. In cross-section the receptors consisted ofan elliptical nerve terminal flanked on each side bySchwann cell cytoplasm (Fig. 5). At each end, the barenerve terminal was adjacent to the basal lamina and, infavourable sections, one end could be seen to contactthe glassy membrane (Fig. 5b). There appeared to be2 types of lanceolate endings within the C1 follicle:small terminals in which caveoli-like vesicles of theSchwann cell membrane were rare (Fig. 5 a) and largerterminals with many caveoli-like vesicles attached tothe Schwann cell membrane (Fig. 5 b). OccasionalMerkel cells and lamellated endings (described below)were also seen.
Innervation of the narrow 'waist' region
In the region between the mesenchymal thickening
and the ICB, the mesenchymal sheath narrows fromits widest extent of approximately 160 gm down to10-20 gm. The fascicles of the deep nerve branchprofusely to form a dense ring of longitudinallyoriented fibres outside this sheath (Fig. 6a). Thesheath, at this level, appears to be a continuation ofthe inner zone of the mesenchymal thickening con-taining fine, longitudinally oriented axons.
This region is densely innervated by several receptortypes: Merkel cells, lanceolate and lamellated recep-tors and free nerve endings. Merkel cells and theirassociated nerve terminals are densely distributed inthe basal layer of the outer root sheath (Fig. 6). Withthe Winkelmann stain each Merkel afferent fibre wasseen to pierce the glassy membrane and form severalbranches that spread circumferentially within the rootsheath. The density of the Merkel terminals isparticularly obvious in a grazing section of this regionwith Winkelmann staining (Fig. 3). The columnarMerkel cell is oriented with the long axis of the cellperpendicular to the glassy membrane and with thenerve terminal adjacent to the lateral surface. TheMerkel cells have characteristic electron-dense cyto-
..dEwBMW 4

Vibrissae in the tammar wallaby
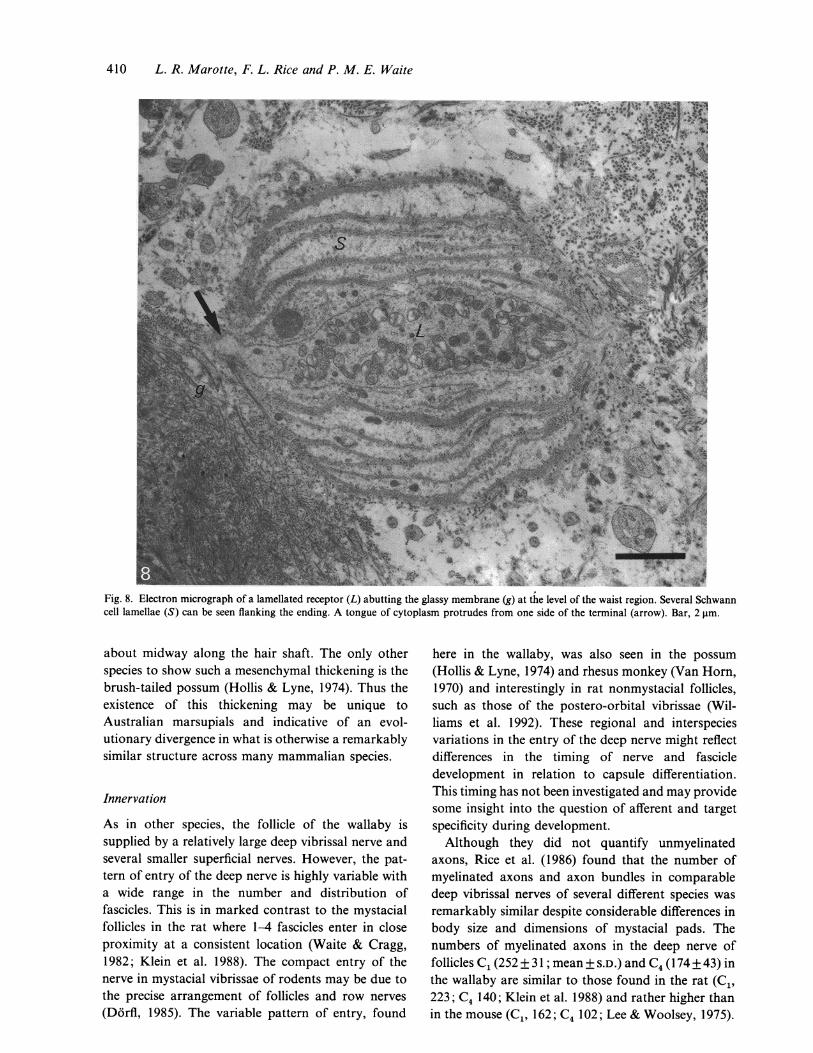
plasmic granules which form desmosomes withkeratinocytes. The nerve terminals contain vesiclesand are rich in mitochondria (Fig. 6). Lanceolate andlamellated receptors can be seen with silver staining aslarge diameter profiles running parallel to the longaxis of the follicle in the mesenchymal sheath (Fig.7a). In transverse semithin sections these receptors aredensely distributed around the circumference of theouter root sheath (Fig. 7b). The outer root sheath andglassy membrane show pronounced ridges projectinginto the mesenchyme and the receptors commonlyabut these projections spanning the mesenchymebetween the glassy membrane and the bundles ofnerve fibres. Lamellated receptors are similar to thelarger lanceolates described above but are flanked by2-11 layers of Schwann cell processes (Fig. 8). Thebasal lamina often extends between the Schwann celllayers and the membranes of these layers commonlyshow numerous caveoli-like vesicles on both theirsurfaces. The terminals are packed with mitochondria.As in the lanceolate endings, the bare edges of thelamellated ending may protrude tongues of cytoplasm(Fig. 8). Occasionally more than 2 protruding tonguesare seen. The free nerve endings at this level were likethose seen in the mesenchymal thickening.
Innervation of the inner conical body
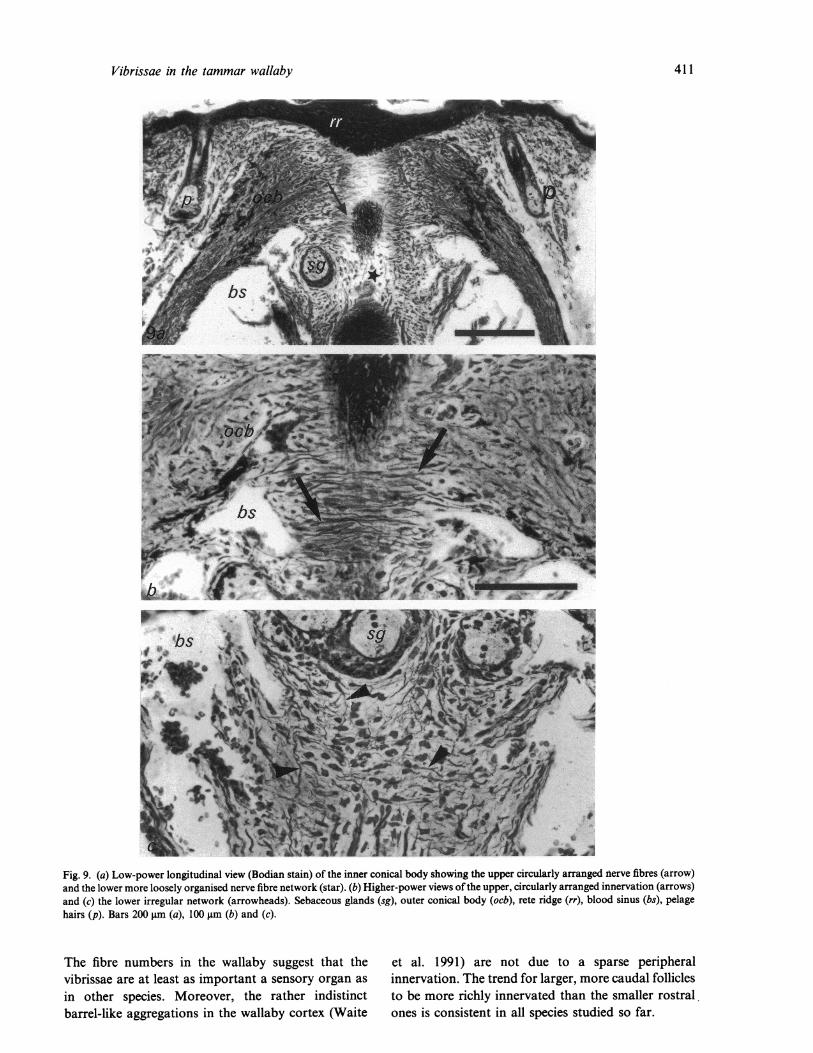
There are 2 patterns of nerve fibre organisation in theICB. In the lower part, below the sebaceous gland,nerve fibres form an irregular network throughout thewidened mesenchymal sheath (Fig. 9a, c). Above this,fine nerve fibres form a circular array around the rootsheath (Fig. 9a, b). By electron microscopy themajority of these were unmyelinated but somemyelinated axons were also present. The majority ofthis innervation ascends from the deep nerve but finenerve fibres of the superficial nerve cross the capsuleto reach the ICB (Fig. 10a, b). Electron microscopyshowed that the vast majority of endings in the ICBwere free nerve endings (Fig. 11) that were fairlysparse in the lower part but more frequent in theupper region of the ICB. Rarely, Merkel cells wereseen in the outer root sheath.
Innervation of the rete ridge
Several superficial nerves enter both the outer conicalbody and pigmented rete ridge at the mouth of thefollicle (Figs 3, 12). Free nerve endings are commonly
present at the outer margins of the rete ridge (Fig. 12).At this site the nerve terminal lies between the Merkelcell and the basal lamina.
Intervibrissal fur
The intervibrissal fur contains many small guard hairsand more numerous, smaller vellus hairs and isinnervated from a plexus of axon bundles in thedermis. The guard hairs are well supplied by a
palisade of small, longitudinally oriented lanceolateendings surrounded by an array of circumferentiallyoriented fibres (Fig. 13 a, b). The innervation may
encircle the accompanying sebaceous gland andoccasionally an adjacent vellus hair. In a few cases, thevellus hairs were supplied by fine, circumferentiallyoriented fibres. Other than this, the silver stains rarelyrevealed any innervation associated with the vellushairs. Occasionally, some axons were observedascending to an epidermal thickening at the mouth ofa guard hair.
DISCUSSION
General morphology of the vibrissalfollicles
The mystacial vibrissal follicle of the tammar wallabyhas many similarities to that described before in othermarsupials as well as rodents, insectivores, lago-morphs, carnivores and primates (Andres, 1966; VanHorn, 1970; Stephens et al. 1973; Hollis & Lyne,1974; Yohro, 1977; Halata & Munger, 1980; Rice etal. 1986). The hair shaft and root sheaths are
surrounded by a thick basement membrane, the glassymembrane, and a mesenchymal sheath (Melaragno &Montagna, 1953). This multilayered structure ispositioned at the core of a blood-filled cavity enclosedwithin a dermal capsule. However, there are 2differences between the follicles of the tammar wallabyand most other species. Firstly, in most species studiedso far, the blood sinus is subdivided into a lowercavernous sinus containing trabeculae and an upper
ring sinus devoid of trabeculae. The ring sinuscontains a ring of tissue, the ringwulst, suspendedaround the mesenchymal sheath. However, the wall-aby lacks any subdivision of the blood sinus.Trabeculae are found throughout the cavernous sinusand the ring sinus and ringwulst are absent. The onlyother animals lacking a ring sinus and ringwulst are
the brush-tailed possum (Hollis & Lyne, 1974),another Australian marsupial, and the rhesus monkey(Van Horn, 1970). The second difference concerns the
seen in this region. Occasional Merkel cells are also
409
thickening or widening of the mesenchymal sheath
410 L. R. Marotte, F. L. Rice and P. M. E. Waite
|_ 2 | z : <' " * trt t tl ~~~~~~~~~~~V4" .ft4 '*
Fig. 8. Electron micrograph of a lamellated receptor (L) abutting the glassy membrane (g) at the level of the waist region. Several Schwanncell lamellae (S) can be seen flanking the ending. A tongue of cytoplasm protrudes from one side of the terminal (arrow). Bar, 2 gm.
about midway along the hair shaft. The only otherspecies to show such a mesenchymal thickening is thebrush-tailed possum (Hollis & Lyne, 1974). Thus theexistence of this thickening may be unique toAustralian marsupials and indicative of an evol-utionary divergence in what is otherwise a remarkablysimilar structure across many mammalian species.
Innervation
As in other species, the follicle of the wallaby issupplied by a relatively large deep vibrissal nerve andseveral smaller superficial nerves. However, the pat-tern of entry of the deep nerve is highly variable witha wide range in the number and distribution offascicles. This is in marked contrast to the mystacialfollicles in the rat where 1-4 fascicles enter in closeproximity at a consistent location (Waite & Cragg,1982; Klein et al. 1988). The compact entry of thenerve in mystacial vibrissae of rodents may be due tothe precise arrangement of follicles and row nerves(Dorfl, 1985). The variable pattern of entry, found
here in the wallaby, was also seen in the possum(Hollis & Lyne, 1974) and rhesus monkey (Van Horn,1970) and interestingly in rat nonmystacial follicles,such as those of the postero-orbital vibrissae (Wil-liams et al. 1992). These regional and interspeciesvariations in the entry of the deep nerve might reflectdifferences in the timing of nerve and fascicledevelopment in relation to capsule differentiation.This timing has not been investigated and may providesome insight into the question of afferent and targetspecificity during development.Although they did not quantify unmyelinated
axons, Rice et al. (1986) found that the number ofmyelinated axons and axon bundles in comparabledeep vibrissal nerves of several different species wasremarkably similar despite considerable differences inbody size and dimensions of mystacial pads. Thenumbers of myelinated axons in the deep nerve offollicles C1 (252 + 31; mean + S.D.) and C4 (174 +43) inthe wallaby are similar to those found in the rat (C1,223; C4 140; Klein et al. 1988) and rather higher thanin the mouse (C1, 162; C4 102; Lee & Woolsey, 1975).
Vibrissae in the tammar wallaby
Fig. 9. (a) Low-power longitudinal view (Bodian stain) of the inner conical body showing the upper circularly arranged nterve fibres (arrow)and the lower more loosely organised nerve fibre network (star). (b) Higher-power views of the upper, circularly arranged innervation (arrows)and (c) the lower irregular network (arrowheads). Sebaceous glands (sg), outer conical body (ocb), rete ridge (rr), blood sinus (bs), pelagehairs (p). Bars 200 gm (a), 100 gm (b) and (c).
The fibre numbers in the wallaby suggest that thevibrissae are at least as important a sensory organ as
in other species. Moreover, the rather indistinctbarrel-like aggregations in the wallaby cortex (Waite
et al. 1991) are not due to a sparse peripheralinnervation. The trend for larger, more caudal folliclesto be more richly innervated than the smaller rostral.ones is consistent in all species studied so far.
411
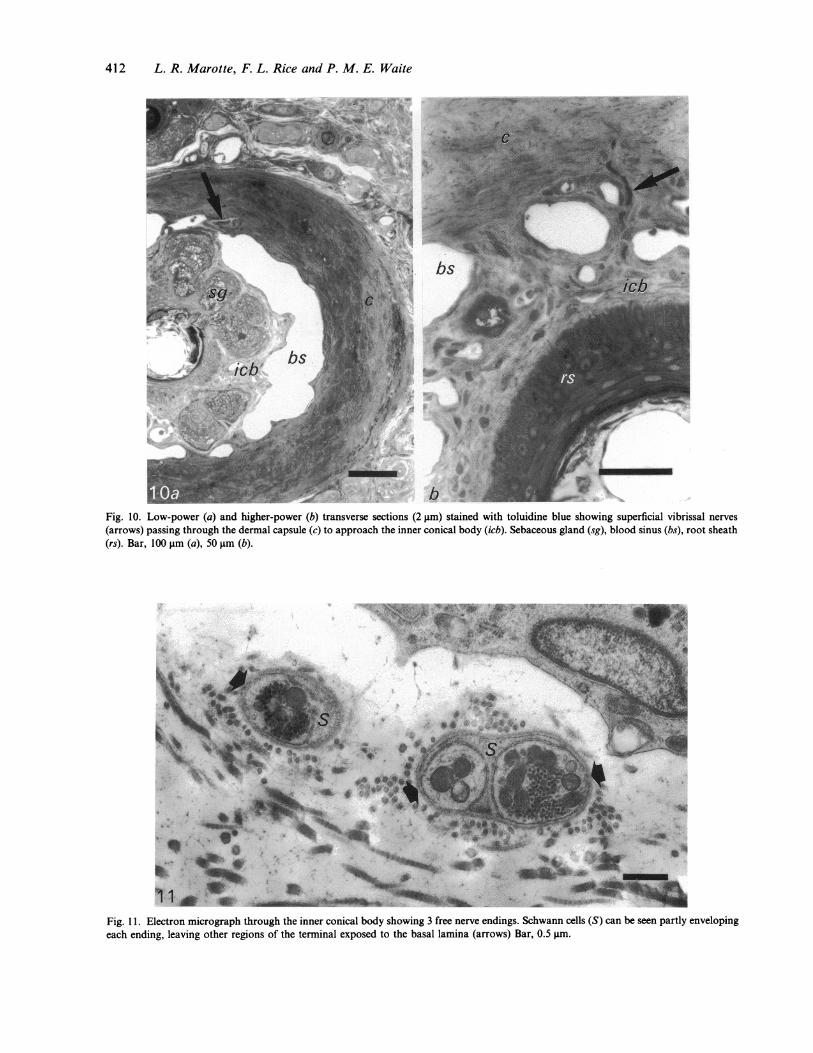
412 L. R. Marotte, F. L. Rice and P. M. E. Waite
Fig. 10. Low-power (a) and higher-power (b) transverse sections (2 gm) stained with toluidine blue showing superficial vibrissal nerves(arrows) passing through the dermal capsule (c) to approach the inner conical body (icb). Sebaceous gland (sg), blood sinus (bs), root sheath(rs). Bar, 100 gm (a), 50 gm (b).
9 1
.' .' ' w
.s. .o a .S...Xs w\ v <*4.s S * .... 4} *_ 1
* i'w Xw r ' v 44 S_ t_ts% | F , ' =| < -W @ _
.. ....^ e £ * s v
1 1 * 'Z' ._
_ _ _.W iL'I .v?4U J~ r_"r -
_~~wt .d__im?t*?b
Fig. 11. Electron micrograph through the inner conical body showing 3 free nerve endings. Schwann cells (S) can be seen partly envelopingeach ending, leaving other regions of the terminal exposed to the basal lamina (arrows) Bar, 0.5 tm.
.,io, *..410.o
,LA (R., '9AiM'4
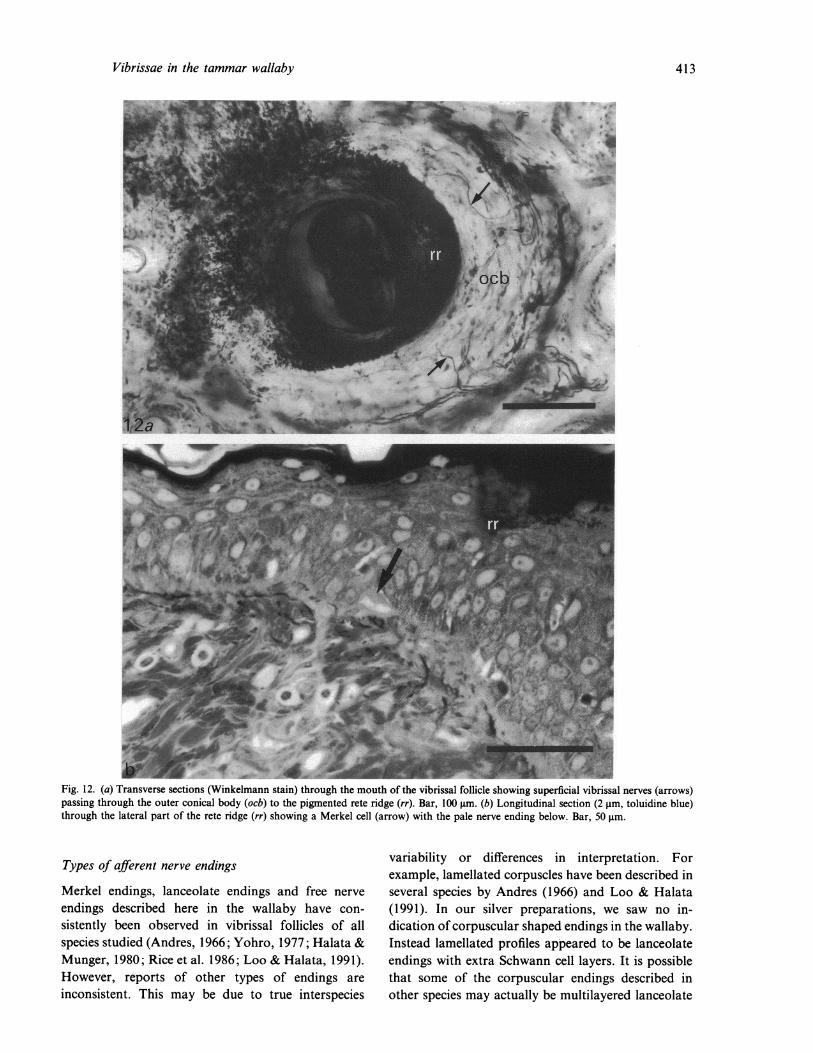
Vibrissae in the tammar wallaby
S
-3uwmlmr
Fig. 12. (a) Transverse sections (Winkelmann stain) through the mouth of the vibrissal follicle showing superficial vibrissal nerves (arrows)passing through the outer conical body (ocb) to the pigmented rete ridge (rr). Bar, 100 gim. (b) Longitudinal section (2 gim, toluidine blue)through the lateral part of the rete ridge (rr) showing a Merkel cell (arrow) with the pale nerve ending below. Bar, 50 gm.
Types of afferent nerve endings
Merkel endings, lanceolate endings and free nerve
endings described here in the wallaby have con-
sistently been observed in vibrissal follicles of allspecies studied (Andres, 1966; Yohro, 1977; Halata &Munger, 1980; Rice et al. 1986; Loo & Halata, 1991).However, reports of other types of endings are
inconsistent. This may be due to true interspecies
variability or differences in interpretation. Forexample, lamellated corpuscles have been described inseveral species by Andres (1966) and Loo & Halata(1991). In our silver preparations, we saw no in-dication of corpuscular shaped endings in the wallaby.Instead lamellated profiles appeared to be lanceolateendings with extra Schwann cell layers. It is possiblethat some of the corpuscular endings described inother species may actually be multilayered lanceolate
413
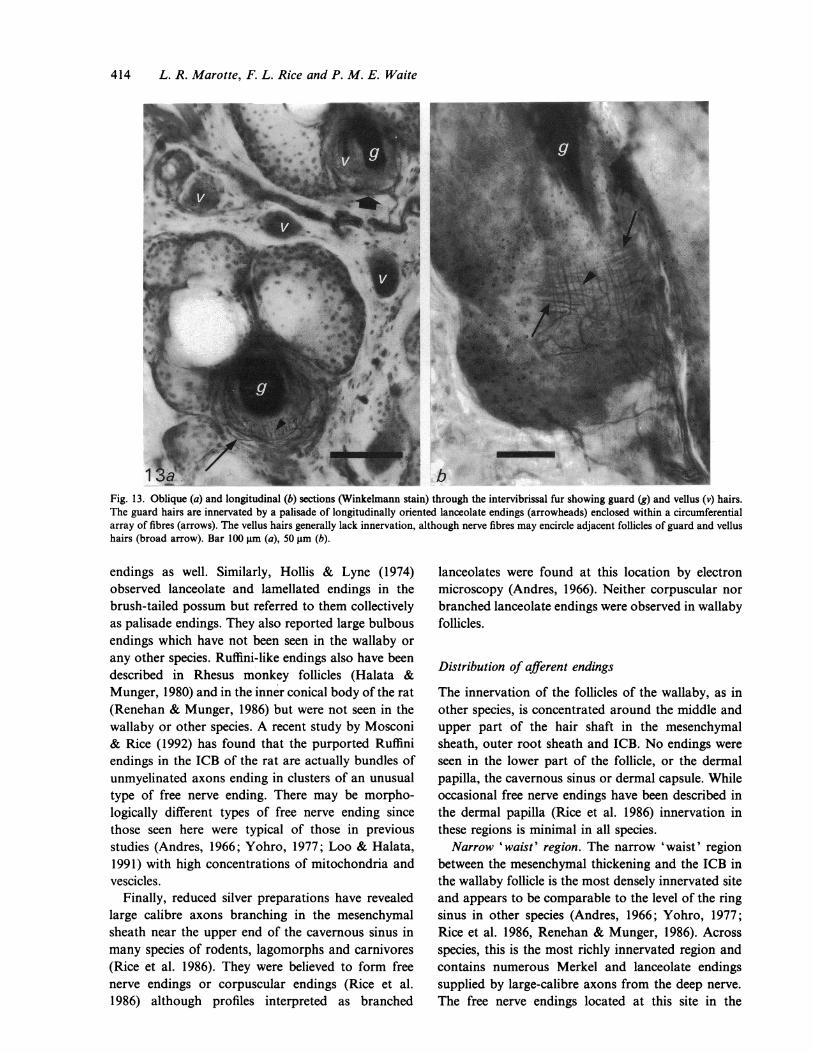
414 L. R. Marotte, F. L. Rice and P. M. E. Waite
Fig. 13. Oblique (a) and longitudinal (b) sections (Winkelmann stain) through the intervibrissal fur showing guard (g) and vellus (v) hairs.The guard hairs are innervated by a palisade of longitudinally oriented lanceolate endings (arrowheads) enclosed within a circumferentialarray of fibres (arrows). The vellus hairs generally lack innervation, although nerve fibres may encircle adjacent follicles of guard and vellushairs (broad arrow). Bar 100 gm (a), 50 gm (b).
endings as well. Similarly, Hollis & Lyne (1974)observed lanceolate and lamellated endings in thebrush-tailed possum but referred to them collectivelyas palisade endings. They also reported large bulbousendings which have not been seen in the wallaby orany other species. Ruffini-like endings also have beendescribed in Rhesus monkey follicles (Halata &Munger, 1980) and in the inner conical body of the rat(Renehan & Munger, 1986) but were not seen in thewallaby or other species. A recent study by Mosconi& Rice (1992) has found that the purported Ruffiniendings in the ICB of the rat are actually bundles ofunmyelinated axons ending in clusters of an unusualtype of free nerve ending. There may be morpho-logically different types of free nerve ending sincethose seen here were typical of those in previousstudies (Andres, 1966; Yohro, 1977; Loo & Halata,1991) with high concentrations of mitochondria andvescicles.
Finally, reduced silver preparations have revealedlarge calibre axons branching in the mesenchymalsheath near the upper end of the cavernous sinus inmany species of rodents, lagomorphs and carnivores(Rice et al. 1986). They were believed to form freenerve endings or corpuscular endings (Rice et al.1986) although profiles interpreted as branched
lanceolates were found at this location by electronmicroscopy (Andres, 1966). Neither corpuscular norbranched lanceolate endings were observed in wallabyfollicles.
Distribution of afferent endings
The innervation of the follicles of the wallaby, as inother species, is concentrated around the middle andupper part of the hair shaft in the mesenchymalsheath, outer root sheath and ICB. No endings wereseen in the lower part of the follicle, or the dermalpapilla, the cavernous sinus or dermal capsule. Whileoccasional free nerve endings have been described inthe dermal papilla (Rice et al. 1986) innervation inthese regions is minimal in all species.Narrow 'waist' region. The narrow 'waist' region
between the mesenchymal thickening and the ICB inthe wallaby follicle is the most densely innervated siteand appears to be comparable to the level of the ringsinus in other species (Andres, 1966; Yohro, 1977;Rice et al. 1986, Renehan & Munger, 1986). Acrossspecies, this is the most richly innervated region andcontains numerous Merkel and lanceolate endingssupplied by large-calibre axons from the deep nerve.The free nerve endings located at this site in the

Vibrissae in the tammar wallaby
wallaby have not been reported in the ultrastructuralstudies of other species (Andres, 1966; Patrizi &Munger, 1966; Yohro, 1977; Halata & Munger, 1980)but this may have been an oversight.
Merkel cells are densely packed in the stratumbasale of the outer root sheath. As in other species,Merkel cells at this site are elongated with their longaxis acutely angled or perpendicular to the glassymembrane. Merkel cell endings give slowly adaptingtype I responses in both glabrous skin (Iggo, 1974;Johansson & Vallbo, 1983) and hairy skin (Brown &Iggo, 1967) and are probably responsible for the sameresponses in sinus hairs (Gottschaldt et al. 1973;Gottschaldt & Vahle-Hinz, 1981; Lichtenstein et al.1990).Numerous lanceolate and lamellated terminals form
a closely spaced palisade within the mesenchymalsheath. The caveoli-like vescicles attached to theSchwann cell membrane surrounding the axon ter-minal have been seen in other species (Andres, 1966;Hollis & Lyne, 1974; Loo & Halata, 1991) althoughtheir function is unknown. Lanceolate endings arethought to be responsible for rapidly adaptingresponses (Iggo, 1974). Rice et al. (1986) speculatedthat they may detect acceleration of vibrissal deflec-tion, possibly due to the inertia of the ringwulst.However, these nerve endings are clearly well repre-sented in the wallaby in the absence of a ringwulst.
Finally, the function of the free nerve endings invibrissal follicles is unknown. Such receptors arecommonly considered to give high threshold responsesbut in a detailed study of responses from singleganglion cells in the rat, only 1.5% of the responsesfrom the vibrissae were high threshold (Jacquin et al.1986). Their relatively protected location in the folliclemay indicate that they are low threshold mechano-receptors.Mesenchymal thickening. The mesenchymal thick-
ening does not appear to be merely a homologue ofthe ringwulst in New World opossums and eutherians.The ringwulst lacks any innervation whereas themesenchymal thickening is innervated by coarse andfine calibre axons from the deep nerve. These supplythe numerous free nerve endings and the occasionallanceolate endings that were observed in the thick-ening as well as the few Merkel endings in the adjacentouter root sheath. In most other species, a slightthickening of the mesenchymal sheath may be presentin the upper part of the cavernous sinus just below thering sinus and ringwulst (Rice et al. 1986; Rice, Jones& Munger, unpublished). This may be the comparableregion to the thickening in the wallaby but, if so, the
afferents (Rice et al. 1986), no free nerve endings or
Merkel cells have been reported at this site in otherspecies (Andres, 1966; Patrizi & Munger, 1966;Renehan & Munger, 1986). Instead, the majorinnervation at this level in other species appears to bebranched lanceolate-like endings in the mesenchymalsheath (Andres, 1966; Yohro, 1977). Based on theseobservations the mesenchymal thickening in thewallaby, and perhaps the brush-tailed possum,
appears to be a unique or highly modified sensory
specialisation.Inner conical body. The innervation of this region
shows great variability amongst different species. Atone extreme, the ICB is innervated by a densecircumferential array of fibres arising mainly from thesuperficial nerve with only a small contribution fromthe deep nerve. This is seen in many rodents and theBrazilian pygmy opossum (Rice et al. 1986; Rice,Jones & Munger, unpublished). At the other extremein, for example, carnivores and a few rodents the ICBis very sparsely innervated with a few fibres from thedeep nerve (Andres, 1966; Rice et al. 1986; Mosconi& Rice, 1992). The innervation of the ICB inAustralian marsupials has not been described before.Comparatively, the innervation of the wallaby ICBwas moderate and arose primarily from the deepnerve with only a small contribution from thesuperficial nerve. The only other animal in which thedeep nerve provides a major innervation to the ICB isthe western pocket gopher, a North American rodent(Rice, Jones & Munger, unpublished).The arrangement of fibres in the wallaby ICB is also
unusual. In the part of the ICB above the sebaceousgland the axons formed a moderately dense cir-cumferential array like that in the rabbit (Rice et al.1986). Below the gland the organisation is more
haphazard, like that seen in the guinea-pig andVirginian opossum (Rice et al. 1986; Rice, Jones &Munger, unpublished). The endings seen in the ICBwere predominantly free nerve endings with theoccasional Merkel cell. Circularly oriented lanceolatesand Ruffini endings have been described in the ICB ofthe rat (Andres, 1966; Renehan & Munger, 1986) butwere not seen in the present study.The significance of deep versus superficial afferents
to the ICB has not been investigated in any species.Dense innervation of the ICB is generally associatedwith small nocturnal species which show rhythmicwhisking of vibrissae during exploratory behaviour(Rice et al. 1986). This observation led to thehypothesis that dense ICB innervation may play a rolein the sensory feedback for the motor control of
receptor types differ. Although containing many fine
415
whisking. The moderate density of ICB innervation in

416 L. R. Marotte, F. L. Rice and P. M. E. Waite
the wallaby which is crepuscular and does not whiskappears to be an exception. However, an importantfactor in whisking as opposed to nonwhisking speciesmay be the source of ICB innervation since in mostwhisking species the ICB is supplied primarily by thesuperficial nerve, whereas in the wallaby the supply isprimarily from the deep nerve. Arvidsson & Rice(1991) have found that the projections from thesuperficial nerve to the brainstem trigeminal nuclei ofthe rat are different from those of the deep nerve.Thus there may be morphological and functionaldifferences between afferents projecting via these 2nerves.
Rete ridge. In the wallaby, as in the majority ofother species studied, the rete ridge is innervated bythe superficial nerve, which supplies endings to Merkelcells in the stratum basale (Halata & Munger, 1980;Rice et al. 1986). However, the density of Merkel cellsin the wallaby rete ridge appeared to be relatively lowand in this respect it was intermediate between well-innervated species like the rat and cat and thoselacking any rete ridge receptors such as the rabbit(Rice et al. 1986).
Intervibrissal hairs. As in most other species, theintervibrissal skin of the wallaby contained bothguard and vellus hairs innervated by the dermalplexus. Comparative studies have revealed consider-able interspecies variation in both the organisationand innervation of the intervibrissal fur (Rice et al.1986; Mosconi & Rice, 1992). The guard hairsof the wallaby were similar to those in most otherspecies with a palisade of small lanceolate endingssurrounded by circumferentially oriented, fine-calibreterminals (Biemsderfer et al. 1978; Rice & Munger,1986; Loo & Halata, 1991). Some axons wereobserved ascending to the mouths of a few guard hairswhere they presumably form Merkel endings. In theRhesus monkey, the circumferential profiles wereidentified by electron microscopy as a mixture of thinlanceolate endings, free nerve endings and Ruffini-likeendings (Biemsderfer et al. 1978). Of these, only thinlanceolate endings were reported in several marsupialspecies (Loo & Halata, 1991).
In most species, vellus hairs are innervated by a fewmeandering axons which form free nerve endings orthe occasional lanceolate ending. Vellus hairs in thewallaby were rarely innervated. However, they werefrequently so close to guard hairs that both types offollicles- were surrounded by the same piloneuralcomplex.The overall innervation of the intervibrissal fur of
the wallaby is relatively moderate and comparable tothat of the rabbit (Rice & Munger, 1986). It has been
suggested that intervibrissal innervation is low inspecies with prominent vibrissa-related barrels in thesomatosensory cortex (Rice & Munger, 1986). As inthe rabbit (Rice et al. 1985), barrels in the wallaby arerelatively indistinct. They are best seen early indevelopment with succinic dehydrogenase reactivityand are never apparent with Nissl staining (Waite etal. 1991).
ACKNOWLEDGEMENTS
The authors would like to thank Ms M. Porter andMs M. Dockum for excellent technical assistance, andMs A. Fritchle for the graphics in Figure 1. Ms A.Devlin provided expert care of the wallaby colony andDr R. Meischke gave veterinary advice.
REFERENCES
ANDRES, KH (1966) Uber die Feinstruktur der Rezeptoren anSinushaaren. Zeitschrift far Zellforschung 75, 339-365.
ARVIDSSON J, RICE FL (1991) Central projections of primarysensory neurons innervating different parts of the vibrissa follicleand intervibrissal skin on the mystacial pad of the rat. Journal ofComparative Neurology 309, 1-16.
BIEMESDERFER D, MUNGER BL, BINCK J, DUBNER R (1978) The pilo-Ruffini complex: a non-sinus hair associated slowly-adaptingmechanoreceptor in primate facial skin, Brain Research 142,197-222.
BODIAN D (1936) New method for staining nerve fibres and nerveendings in mounted paraffin sections. Anatomical Record 65,89-97.
BROWN AG, IGGo A (1967) A quantitative study of cutaneousreceptors and afferent fibres in the cat and rabbit. Journal ofPhysiology 193, 707-733.
CRISSMAN RS, WARDEN RJ, SICILIANO DA, KLEIN BG, RENEHANWE et al. (1991) Numbers of axons innervating mystacial vibrissafollicles in newborn and adult rats. Somatosensory and MotorResearch 8, 103-109.
DORFL J (1985) The innervation of the mystacial region of the whitemouse. A topographical study. Journal ofAnatomy 142, 173-184.
GOTrSCHALDT K-M, IGGo A, YOUNG DW (1973) Functionalcharacteristics of mechanoreceptors in sinus hair follicles of thecat. Journal of Physiology 235, 287-315.
GoTTScHALDT K-M, VAHLE-HINZ C 1981) Merkel cell receptors:structure and transducer function. Science 214, 183-186.
HALATA Z, MUNGER BL (1980) Sensory nerve endings in Rhesusmonkey sinus hair. Journal of Comparative Neurology 192,645-663.
HOLLIs DE, LYNE AG (1974) Innervation of vibrissa follicles in themarsupial Trichosurus vulpecula. Australian Journal of Zoology22, 263-276.
IbGo A (1974) Cutaneous receptors. In The Peripheral NervousSystem (ed. J. I. Hubbard), pp. 347-404. New York: Plenum.
JACQUIN MF, RENEHAN WE, KLEIN BG, MOONEY RD, RHoADESRW (1986) Functional consequences of neonatal infraorbitalnerve section in rat trigeminal ganglion. Journal ofNeuroscience6, 3706-3720.
JOHANSSON RS, VALLBO AK (1983) Tactile sensory coding in theglabrous skin of the human hand. Trends in Neuroscience 6,27-32.
KLEIN HG, RENEHAN WE, JACQUIN MF, RHOADES RW (1988)Anatomical consequences of neonatal infraorbital nerve tran-

Vibrissae in the tammar wallaby 417
section upon the trigeminal ganglion and vibrissa follicle nervesin the adult rat. Journal of Comparative Neurology 268, 469-488.
LEE KJ, WOOLSEY TA (1975) A proportional relationship betweenperipheral innervation density and cortical neuron number in thesomatosensory system of the mouse. Brain Research 99, 349-353.
LICHrENSTEIN SH, CARVELL GE, SIMONS, DJ (1990) Responses ofrat trigeminal ganglion neurons to movements of vibrissae indifferent directions. Somatosensory and Motor Research 7, 47-75.
Loo S-K, HALATA Z (1991) Innervation of hairs in the facial skin ofmarsupial mammals. Journal ofAnatomy 174, 207-219.
MELARAGNO HP, MONTAGNA W (1953) The tactile hair follicles inthe mouse. Anatomical Record 115, 129-149.
MOSCONI TM, RICE FL (1992) Sequential maturation of sensoryendings in the mystacial pad of the ferret. Journal ofComparativeNeurology, in press.
PATRIZI G, MUNGER BL (1966) The ultrastructure and innervationof rat vibrissae. Journal of Comparative Neurology 26, 423-436.
POCOCK RI (1914) On the facial vibrissae ofMammalia. Proceedingsof the Zoological Society 48, 889-912.
RENEHAN WE, MUNGER BL (1986) Degeneration and regenerationof peripheral nerve in the rat trigeminal system. I. Identificationand characterisation of the multiple afferent innervation of themystacial vibrissae. Journal of Comparative Neurology 2A6,129-145.
RICE FL, GOMEZ C, BARsTow C, BURNET A, SANDS P (1985) Acomparative analysis of the development of primary somato-sensory cortex: interspecies similarities during barrel and laminadevelopment. Journal of Comparative Neurology 236, 477-495.
RICE FL, MANCE A, MUNGER BL (1986) A comparative lightmicroscopic analysis of the sensory innervation of the mystacialpad. I. Innervation of vibrissal follicle-sinus complexes. Journalof Comparative Neurology 252, 154-174.
RICE FL, MUNGER BL (1986) A comparative light microscopeanalysis of the sensory innervation of the mystacial pad. II. Thecommon fur between the vibrissae. Journal of ComparativeNeurology 252, 186-205.
STEPHENS RJ, BEEBE IJ, POULTER TC (1973) Innervation of thevibrissae of the California sea lion, Zalophus californianus.Anatomical Record 176, 421-442.
VAN DER Loos H, WOOLSEY TA (1973) Somatosensory cortex:structural alterations following early injury to sense organs.Science 179, 395-398.
VAN HORN RN (1970) Vibrissae structure in the rhesus monkey.Folia Primatologica 13, 241-285.
VINCENT SB (1913) The tactile hair of the white rat. Journal ofComparative Neurology 23, 1-36.
WAITE PME, CRAGG BG (1982) The peripheral and central changesresulting from cutting or crushing the afferent nerve supply to thewhiskers. Proceedings of the Royal Society London B 214,191-211.
WAITE PME, MAROTTE LR, MARK RF (1991) Development ofwhisker representation in the cortex of the tammar wallabyMacropus eugenii. Developmental Brain Research 58, 35-41.
WELKER E, VAN DER LOOS H (1986) Quantitative correlationbetween barrel-field size and the sensory innervation of thewhiskerpad: a comparative study in six strains of mice bred fordifferent patterns of mystacial vibrissae. Journal of Neuroscience6, 3355-3373.
WELLER, WL (1972) Barrels in somatic sensory neocortex of themarsupial Trichosurus vulpecula (Brush-tailed possum). BrainResearch 43, 11-24.
WILLIAMS JB, DE PERMENTEER P, WAITE PME (1992) The rat'spostero-orbital sinus hair: II. Normal morphology and theincrease in peripheral innervation with adjacent nerve section.Journal of Comparative Neurology, in press.
WINKELMANN, RK (1960) Nerve Endings in Normal and PathologicSkins. Springfield, Illinois: C. C. Thomas.
WOOLSEY TA, VAN DER Loos H (1970) The structural organizationof layer IV in the somatosensory region (SI) of mouse cerebralcortex. Brain Research 17, 205-242.
YoHRo Y (1977) Structure of the sinus hair follicle in the big-clawedshrew, Sorex unguiculatus. Journal ofMorphology 153, 333-354.





























