Embed Size (px)
Citation preview

Research ArticleThe Neuroprotective Effects of Brazilian Green Propolis onNeurodegenerative Damage in Human Neuronal SH-SY5Y Cells
Junjun Ni1 Zhou Wu12 Jie Meng1 Aiqin Zhu3 Xin Zhong3
Shizheng Wu3 and Hiroshi Nakanishi1
1Department of Aging Science and Pharmacology Faculty of Dental Science Kyushu University Fukuoka 812-8582 Japan2OBT Research Center Faculty of Dental Science Kyushu University Fukuoka 812-8582 Japan3Institute of Geriatrics Qinghai Provincial Hospital Xining 810007 China
Correspondence should be addressed to Zhou Wu zhouwdentkyushu-uacjp
Received 4 July 2016 Revised 19 December 2016 Accepted 28 December 2016 Published 6 February 2017
Academic Editor Byoung-Joon Song
Copyright copy 2017 Junjun Ni et alThis is an open access article distributed under theCreativeCommonsAttribution License whichpermits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Oxidative stress and synapse dysfunction are the major neurodegenerative damage correlated to cognitive impairment inAlzheimerrsquos disease (AD)We have found that Brazilian green propolis (propolis) improves the cognitive functions ofmild cognitiveimpairment patients living at high altitude however mechanism underlying the effects of propolis is unknown In the presentstudy we investigated the effects of propolis on oxidative stress expression of brain-derived neurotrophic factor (BDNF) andactivity-regulated cytoskeleton-associated protein (Arc) the critical factors of synapse efficacy using human neuroblastoma SH-SY5Y cells Pretreatment with propolis significantly ameliorated the hydrogen peroxide- (H
2O2-) induced cytotoxicity in SH-SY5Y
cells Furthermore propolis significantly reduced the H2O2-generated reactive oxygen species (ROS) derived from mitochondria
and 8-oxo-21015840-deoxyguanosine (8-oxo-dG the DNA oxidative damage marker) but significantly reversed the fibrillar 120573-amyloidand IL-1120573-impaired BDNF-induced Arc expression in SH-SY5Y cells Furthermore propolis significantly upregulated BDNFmRNA expression in time- and dose-dependent manners In addition propolis induced Arc mRNA and protein expression viaphosphoinositide-3 kinase (PI3K) These observations strongly suggest that propolis protects from the neurodegenerative damagein neurons through the properties of various antioxidantsThe present study provides a potential molecularmechanism of Braziliangreen propolis in prevention of cognitive impairment in AD as well as aging
1 Introduction
Alzheimerrsquos disease (AD) is the most common form of dem-entia in aging societies worldwide [1] and the number ofAD is growing dramatically [2] Oxidative stress is a majorcomponent of the harmful cascades activated in the develop-ment of aging-related neurodegenerative disorders includingAD [3 4] because the overproduction of reactive oxygenspecies (ROS) causes cell damage through the promotion oflipid peroxidation DNA damage and the regulation of deathproteins [5] Antioxidant therapy therefore is considered asan approach in prevention and clinical management of AD[6] On the other hand dysfunction of hippocampal synapticefficacy is known to be related to cognitive impairment inAD [7 8] as well as in aging [9ndash11] and synaptic efficacy hasprotective effects against AD [12 13]
Activity-regulated cytoskeleton-associated protein (Arc)is a critical immediate-early gene that has been implicated ingenerated stable changes in synaptic efficacy [14 15] as sup-pression of Arc expression impairs the synaptic plasticity andmemory consolidation [11 14 16] As a member of the neuro-trophin family brain-derived neurotrophic factor (BDNF)plays a fundamental role in the synaptic efficacy by inducingArc expression [17 18] and BDNF-induced Arc expressioncontributes to cognitive functions [14 19 20] Indeed fibrillar120573-amyloid the main component of plaques in the brains ofAD patients has been shown to impair the BDNF-inducedArc expression in the cultured cortical neurons even atlow levels [21] therefore interfering with BDNF signalingaffects the downstream neuronal functions that contribute tothe development of AD [22] However mounting evidence
Hindawi Publishing CorporationOxidative Medicine and Cellular LongevityVolume 2017 Article ID 7984327 13 pageshttpdxdoiorg10115520177984327
2 Oxidative Medicine and Cellular Longevity
suggests that neuroinflammation induced by proinflamma-tory cytokines increases the risk of cognitive impairment[23ndash25] As a potent activator for exacerbating neuroinflam-mation interleukin-1120573 (IL-1120573) has been shown to mediatesynaptic efficacy by suppressing BDNF-induced Arc expres-sion [26]
Propolis is a resinous substance produced by honeybeesas a defense against intruders It has relevant therapeuticproperties that have been used since ancient times Thechemical composition of propolis depends on the local floralat the site of collection [27 28] The neuroprotective effectsof propolis and its active components such as baccharin p-Coumaric acid and Artepillin C have been established [29ndash32] In addition Artepillin C amajor component of Brazilianpropolis has been shown to act as a neurotrophic-likefactor for promoting neuron growth factor- (NGF-) inducedneurite outgrowth [33] Our ongoing human research athigh altitude has shown that elderly individuals treated withpropolis score significantly higher on cognitive tests thannontreated subjects (Zhu and Wu unpublished data) and wehypothesized that propolis may have neuroprotective effectson neurons directly However we recently found that propolisprotects against the hypoxia-induced microglia mediatedneuroinflammation [34 35] In the present study we focusedon the effects of propolis on the hydrogen peroxide- (H
2O2-
) induced oxidative stress and expression of BDNF and Arcusing cultured human neuroblastoma SH-SY5Y cells whichare widely used for the study of neurodegenerative damage invitro [36]
2 Material and Methods
21 Reagents Brazilian green propolis ethanol extract(propolis) was purchased from Yamada Apiculture CenterInc (Okayama Japan) Minimal essential medium (MEM)F12 fetal bovine serum (FBS) penicillin-streptomycinand Hoechst 33342 were purchased from Thermo FisherScientific (Waltham MA USA) H
2O2(30) GF 109203X
hydrochloride (protein kinase C inhibitor) and ANA-12(TrkB selective antagonist) were purchased fromSelleckchem(Houston TXUSA) Bay11-7082 (a specificNF-120581B inhibitor)U0126 (ERK inhibitors) Carbachol and human BDNF werepurchased from Sigma-Aldrich Inc (St Louis MO USA)Y-27632 dihydrochloride (Rho-associated protein kinaseinhibitor)was purchased fromTOCRIS (Avonmouth BristolUK) Wortmannin (phosphoinositide 3-kinase inhibitor)was purchased from Millipore (California USA) ArtepillinC was purchased from WAKO (Osaka Japan) RNAiso Pluswas purchased fromTakara (Hoto-kuOsaka Japan) Quanti-Tect Reverse Transcription Kit and Rotor-Gene SYBR GreenRT-PCR Kit were purchased from Qiagen (Hilden Ger-many) A Cell-Counting Kit (CCK-8) was purchased fromDojindo (Kumamoto Japan) Human recombinant IL-1120573was purchased from RampD (Minneapolis USA) 120573-Amyloid(1ndash42) was purchased from ANASPEC (California USA)dissolved in endotoxin-free sterile water and incubated at37∘C for 24 h to induce the fibril formation
22 SH-SY5Y Cell Culture Human neuroblastoma SH-SY5Ycells purchased from American Type Culture Collection
(Manassas VA USA) were maintained in a completegrowth medium (MEMF-12 mixture containing 10 fetalbovine serum supplemented with NaHCO
3and 100UmL
penicillin-streptomycin) The cells were cultured at 37∘C ina 5 CO
2humidified incubator
23 Cell Viability Assay SH-SY5Y cells were seeded in 96-well plates (5 times 103 cellswell) overnight After treatment withpropolis fA120573 and IL-1120573 a cell viability assay was performedusing a Cell-Counting Kit (Dojindo) in accordance with thepreviously described method [34] The optical density wasread at a wavelength of 450 nm with a microplate reader Thecell viability was calculated by dividing the optical density ofthe treated group by that of the control group
24 Observations of Morphological Changes The cells wereseeded in 6-well plates (2 times 105 cellsmL) for 24 h and thentreated with H
2O2at a concentration of 100120583M for 24 h
with or without propolis (methanol extraction) The cellularmorphology was observed and photographed using a bright-field microscope (Nikon ECLIPSE Ti-S Japan)
25 Detection of Mitochondrial ROS Mitochondrial ROSwere measured using MitoSOX Red (Invitrogen USA)which is a live-cell permeant that rapidly and selectively tar-gets mitochondria [37] Once in the mitochondria MitoSOXRed reagent is oxidized by superoxide and exhibits red fluo-rescence (with excitation at 510 nm and emission at 580 nm)The cultured SH-SY5Y cells were seeded in 24-well plates (2times 105 cellsmL) and incubated with or without propolis for 1 h(methanol extraction 10 120583gmL) The cells were then furtherexposed to H
2O2(100 120583M) for 1 h After incubation in Hankrsquos
balanced salt solution (HBSS) containing 5mM MitoSOXRed for 10min at 37∘C the cells were washed twice with PBSand thenmounted in a warm buffer for imaging Images werecollected using a fluorescence microscope (Nikon ECLIPSETi-S Japan)
26 Immunofluorescence Imaging Immunofluorescence ima-ging was performed as described previously [34 38] SH-SY5Y cells were seeded in 24-well plates (2 times 105 cellsmL)for 24 h and then treated with H
2O2(100 120583M) for 4 h with
or without pretreatment with propolis (methanol extraction10 120583gmL) for 1 h and fixed with 4 paraformaldehydeAfter washing the cells with PBS twice they were incubatedwith mouse anti-8-oxo-dG (1 500) overnight at 4∘C andthen incubated with anti-mouse Alexa 488 (1 2000 JacksonImmunoresearch Lab Inc) at 4∘C for 2 h After washing byPBS the nucleus was stained by Hoechst 33342 The cellswere mounted in the antifading medium Vecta shield andthe fluorescence images were taken using a confocal laserscanning microscope (CLSM C2si Nikon Japan)
27 Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR) The mRNA isolated from the SH-SY5Y cells at var-ious time points were subjected to a real-time qRT-PCRThe total RNA was extracted using the RNAiso Plus inaccordance with the manufacturerrsquos instructions A total of800 ng of extracted RNA was reverse transcribed to cDNA
Oxidative Medicine and Cellular Longevity 3
using the QuantiTect Reverse Transcription Kit After aninitial denaturation step at 95∘C for 5min temperaturecycling was initiated Each cycle consisted of denaturationat 95∘C for 5 s annealing at 60∘C for 10 s and elongationfor 30 s In total 40 cycles were performed The cDNA wasamplified in duplicate using a Rotor-Gene SYBR Green RT-PCR Kit with a Corbett Rotor-Gene RG-3000A Real-TimePCR System (Sydney Australia) The data were evaluatedusing the RG-3000A software program (version Rotor-Gene6193 Corbett) The sequences of primer pairs were asfollows Arc 51015840-CCACCTGCTTGGACACCTC-31015840 and 51015840-CCGCCCCGAGGAGTTTG-31015840 BDNF 51015840-GGATGAGGA-CCAGAAAGT-31015840 and 51015840-AGCAGAAAGAGAAGAGGAG-31015840 actin 51015840-AGAGCTACGAGCTGCCTGAC-31015840 and 51015840-AGCACTGTGTTGGCGTACAG-31015840
For data normalization an endogenous control (actin)was assessed to control for the cDNA input and the relativeunits were calculated by a comparative Ct method All of thereal-time qRT-PCR experiments were repeated three timesand the results are presented as the means of the ratios plusmnstandard error of the mean
28 Electrophoresis and Immunoblotting SH-SY5Y cells werecultured at a density of 2 times 105 cellsmL Cells were har-vested at each time point with various stimulations andimmunoblotting analyses were conducted Briefly each spec-imen was electrophoresed using 12 SDS-polyacrylamidegels The proteins on the SDS gels were then electrophoret-ically transferred to nitrocellulose membranes Followingblocking the membranes were incubated at 4∘C overnightunder gentle agitation with each primary antibody mouseanti-Arc (1 1000 Abcam Heidelberg Germany) and mouseanti-actin (1 5000 Abcam) After washing the membraneswere incubated with horseradish peroxidase- (HRP-) labeledanti-mouse (1 2000 GE Healthcare Buckinghamshire UK)for 2 h at room temperature Subsequently the membrane-bound HRP-labeled antibodies were detected using anenhanced chemiluminescence detection system (ECK lit GEHealthcare) with an image analyzer (LAS-1000 Fuji PhotoFilm Minato-ku Tokyo Japan)
29 Statistical Analysis The data are represented as themeans plusmn standard error of the mean The statistical analyseswere performed by a one- or two-way analysis of variancewith a post hoc Tukeyrsquos test using the GraphPad Prism soft-ware package (GraphPad Software California USA) A valueof 119901 lt 005was considered to indicate statistical significance
3 Results
31 Effect of Propolis on Cell Viability in SH-SY5Y Cells Wefirst examined the effects of propolis on the viability of SH-SY5Y cells using CCK8 assay kit The mean cell viability wasnot significantly changed after treatment with the ethanolextracts of propolis at final concentrations between 025 and10 120583gmL for 48 h (Figure 1(a)) However themean cell viabil-ity was significantly reduced after pretreatment with propolisat a final concentration of over 25120583gmL (75 of viablecells) We therefore used methanol extracts of propolis at
concentrations up to 10120583gmL and H2O2at 100120583M concen-
tration [38] to investigate the effects of propolis on the H2O2-
induced cytotoxicity of SH-SY5Y cells in the subsequentexperiments The H
2O2-induced cell death (79 of viable
cells) was significantly restored by pretreatment with propolis(90 of viable cells) (Figure 1(b)) Morphological changesobserved included the degeneration of H
2O2-treated SH-
SY5Y cells which exhibited the disappearance of the neuritesand shrinkage (Figure 1(c))The percentage of viable cells wasalso reduced by H
2O2in a dose-dependent manner (data not
shown) It is noted that the neuritis and shrinkage of cellswere attenuated by pretreatment with propolis (Figure 1(c))These observations strongly demonstrate that pretreatmentwith propolis protects SH-SY5Y cells from H
2O2-induced
cytotoxicity Pretreatment with methanol extracts of propolis(10 120583gmL) for 1 h was set up in the subsequent experiments
32 Effects of Propolis on the H2O2-Induced Oxidative Stress
in SH-SY5Y Cells Oxidative stress is an important inducerof neurotoxicity in AD patients [39] following our previousexperiments we used two approaches to address the effectsof propolis on H
2O2-induced oxidative stress in SH-SY5Y
cells one approach was the use of a MitoSOX Red probe asa marker for mitochondria-derived ROS generation [35] andthe other was immunofluorescence imaging for a biomarkerof oxidation-damaged DNAmarker 8-oxo-dG [40] In com-parison to the untreated cells the expression of MitoSOXRed signals was significantly increased in SH-SY5Y cells afterexposure to H
2O2for 1 h suggesting that the mitochondria
are the early origin of ROS generation during oxidative stressPretreatment with propolis significantly inhibited the H
2O2-
induced mitochondria-derived ROS generation in SH-SY5Ycells (Figure 2(a)) and the mean fluorescent intensity ofMitoSOX Red oxidation was found to significantly increasein comparison to that in the cells that were not exposed toH2O2(493 versus 1 lowastlowastlowast119901 lt 0001 Figure 2(b)) Pretreat-
ment with the propolis methanol extracts for 1 h significantlyreduced the immunofluorescence intensity of MitoSOX Redoxidation in the H
2O2-exposed SH-SY5Y cells (18 versus
493 lowastlowastlowast119901 lt 0001 Figure 2(b)) thus confirming the antiox-idant properties of propolis Immunofluorescence imagingshowed a significant inverse relationship between Hoechstand 8-oxo-dG after exposure of SH-SY5Y cells to H
2O2for
4 h (Figure 2(c)) and the mean fluorescent intensity of 8-oxo-dG was found to significantly increase in comparisonto that in the cells that were not exposed to H
2O2(275
versus 1 lowastlowastlowast119901 lt 0001 Figure 2(d)) It is noted that pre-treatment with the propolis for 2 h significantly reduced theimmunofluorescence intensity of 8-oxo-dG in the H
2O2-
exposed SH-SY5Y cells (136 versus 275 lowastlowastlowast119901 lt 0001Figure 2(d)) These observations demonstrate that propoliscould attenuate oxidative stress in neuronal cells
33 Effect of Propolis on the Fibrillar A120573-Induced Impairmentof BDNF-Induced Arc Expression in SH-SY5Y Cells SolubleA120573 is known to interfere with synaptic efficacy as cognitivedecline precedes the formation of A120573 plaques the hallmarkof AD [41] We therefore examined the effects of propolis on
4 Oxidative Medicine and Cellular Longevity
0 025 05 1 5 10 25
lowastlowastlowast
0
20
40
60
80
100
120
140C
ell v
iabi
lity
()
Propolis concentration (gmL)
(a)
Con
lowastlowastlowast
lowastlowast
H2O2 H2O2 propolis50
60
70
80
90
100
110
Cel
l via
bilit
y (
)
(b)
Con H2O2 H2O2 propolis
(c)
Figure 1 Effects of methanol extracts of propolis on the H2O2-induced toxicity in SH-SY5Y cells (a) Cell viability in SH-SY5Y cells
pretreatment with different concentrations of propolis for 48 hours (b) The effect of pretreatment with methanol extracts of propolis for2 h on H
2O2exposed SH-SY5Y cells Each column and bar represent mean plusmn SEM (119899 = 4 each) An asterisk indicates a statistically significant
difference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001) (c) The morphological changes of SH-SY5Y cells with or withoutpretreatment with propolis (10120583gmL) after expose to H
2O2for 24 h Scale bar = 20 120583m
the impairment of BDNF-induced Arc expression by fA120573 inSH-SY5Y cells because Arc is the critical factor of synapseefficacyThemean cell viability was not significantly changedafter treatment with fA120573 at final concentrations between01 and 5 120583M for 48 h (Figure 3(a)) However significantcytotoxicity was observed in cultures treated with fA120573 over10 120583M (80 of viable cells) Therefore 5 120583M was used asthe nonlethal concentration for SH-SY5Y cells in subsequentexperiments Treatment with BDNF (10 ngmL) for 120mininduced 35-fold Arc expression in SH-SY5Y cells howeverpreexposure to fA120573 (5 120583M) for 6 h suppressed the BDNF-induced Arc expression (Figure 3(b)) Interestingly thisimpairment was significantly reversed following treatmentwith propolis (10 120583gmL) for 120min (Figure 3(b)) Further-more propolis treatment also prevented the fA120573-inducedimpairment of BDNF-induced Arc protein expression (Fig-ures 3(c) and 3(d)) These data indicate that propolis reversesthe fA120573-induced impairment of BDNF-induced Arc mRNAand protein expression
34 Effect of Propolis on the IL-1120573-Induced Impairment ofBDNF-Induced Arc Expression in SH-SY5Y Cells As a potent
activator for exacerbating neuroinflammation IL-1120573 hasrecently been found to suppress BDNF-dependent synapticefficacy [26] We therefore examined the effects of propolison the IL-1120573-induced impairment of BDNF-induced Arcexpression in SH-SY5Y cells We first examined the effectsof IL-1120573 on the cell viability to determine the nonlethalconcentration of IL-1120573 on SH-SY5Y cells The mean cellviability was not significantly changed after treatment withIL-1120573 at final concentrations between 1 and 500 ngmL for48 h (Figure 4(a)) However significant cytotoxicity wasobserved in cultures treated with IL-1120573 over 100 ngmL (90of viable cells) We therefore used 20 ngmL as the nonlethalconcentration for SH-SY5Y cells in subsequent experiments
Treatment with BDNF (10 ngmL) for 120min induced34-fold Arc expression in SH-SY5Y cells however preex-posure to IL-1120573 (20 ngmL) for 6 h significantly suppressedthe BDNF-induced Arc expression (Figure 4(b)) Interest-ingly this impairment was significantly reversed follow-ing treatment with propolis (10mgmL) for 120min (Fig-ure 4(b)) Furthermore propolis treatment also preventedthe IL-1120573-induced impairment of BDNF-inducedArc protein
Oxidative Medicine and Cellular Longevity 5
Hoechst MitoSOX Merge
Con
H2O2
H2O2propolis
(a)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
1
2
3
4
5
6
Relat
ive M
itoSO
X flu
ores
cenc
e den
sity
(b)
Hoechst Merge
Con
H2O2
H2O2propolis
8-oxo-dG
(c)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
05
1
15
2
25
3
35
Relat
ive 8
-oxo
-dG
fluo
resc
ence
den
sity
(d)
Figure 2 Effects of methanol extracts of propolis on the H2O2-induced oxidative stress in SH-SY5Y cells (a) Fluorescent images ofMitoSOX
Red signals in SH-SY5Y cells exposed to H2O2for 1 h with or without propolis (10120583gmL) for 2 h Scale bar = 15 120583m (b) The quantitative
analyses of MitoSOX Red signal intensity in (a) (c) Immunofluorescent CLMS images of 8-oxo-dG (red) with Hoechst-stained nuclei (blue)in SH-SY5Y cells exposed to 100 120583M of H
2O2for 4 h with or without pretreatment with propolis (10120583gmL) for 2 h Scale bar = 10 120583m (d)
The quantitative analyses of 8-oxo-dG immunofluorescence signal intensity in (c) The results are expressed as the mean plusmn SEM (119899 = 4 each)and the asterisks indicate a statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
expression (Figures 4(c) and 4(d)) These data indicate thatpropolis reverses the IL-1120573-induced impairment of BDNF-induced Arc mRNA and protein expression
35 Effect of Propolis on BDNF Expression in SH-SY5YCells We next examined the effects of propolis on BDNFexpression in SH-SY5Y cells because BDNF is the criticalneurotrophic factor that is important for synapse efficacyPretreatment with propolis significantly increased the BDNFmRNA expression in SH-SY5Y cells from 60min up to240min even at a low concentration (1120583gmL) (Figure 5(a))Furthermore treatment with propolis for 60min increasedthe BDNF mRNA expression in SH-SY5Y cells in a dose-dependent manner (Figure 5(b)) The propolis-increased
BNDF expression was completely abolished by preincubationwith Wortmannin (200 nM PI-3K inhibitor) but not bytreatment with Y27632 (1 120583M ROCK inhibitor) or GFX(200 nM PKC inhibitor) (Figure 5(c)) BDNFTrkB signalinghas been reported to play an important role in long termpotential to further confirm the BDNF signaling to Arc weuse a Carbachol to stimulate SH-SY5Y and the responses ofCarbachol were examined in the presence of U0126 (ERKinhibitors 10 120583M) or ANA-12 (TrkB selective antagonist100 nM)
1 h of treatment with Carbachol can significantly increasethe protein expression level of Arc However pretreatmentwith U0126 and ANA significantly reduced the Carbachol-increased Arc (Figures 5(d) and 5(e))) Therefore the
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

2 Oxidative Medicine and Cellular Longevity
suggests that neuroinflammation induced by proinflamma-tory cytokines increases the risk of cognitive impairment[23ndash25] As a potent activator for exacerbating neuroinflam-mation interleukin-1120573 (IL-1120573) has been shown to mediatesynaptic efficacy by suppressing BDNF-induced Arc expres-sion [26]
Propolis is a resinous substance produced by honeybeesas a defense against intruders It has relevant therapeuticproperties that have been used since ancient times Thechemical composition of propolis depends on the local floralat the site of collection [27 28] The neuroprotective effectsof propolis and its active components such as baccharin p-Coumaric acid and Artepillin C have been established [29ndash32] In addition Artepillin C amajor component of Brazilianpropolis has been shown to act as a neurotrophic-likefactor for promoting neuron growth factor- (NGF-) inducedneurite outgrowth [33] Our ongoing human research athigh altitude has shown that elderly individuals treated withpropolis score significantly higher on cognitive tests thannontreated subjects (Zhu and Wu unpublished data) and wehypothesized that propolis may have neuroprotective effectson neurons directly However we recently found that propolisprotects against the hypoxia-induced microglia mediatedneuroinflammation [34 35] In the present study we focusedon the effects of propolis on the hydrogen peroxide- (H
2O2-
) induced oxidative stress and expression of BDNF and Arcusing cultured human neuroblastoma SH-SY5Y cells whichare widely used for the study of neurodegenerative damage invitro [36]
2 Material and Methods
21 Reagents Brazilian green propolis ethanol extract(propolis) was purchased from Yamada Apiculture CenterInc (Okayama Japan) Minimal essential medium (MEM)F12 fetal bovine serum (FBS) penicillin-streptomycinand Hoechst 33342 were purchased from Thermo FisherScientific (Waltham MA USA) H
2O2(30) GF 109203X
hydrochloride (protein kinase C inhibitor) and ANA-12(TrkB selective antagonist) were purchased fromSelleckchem(Houston TXUSA) Bay11-7082 (a specificNF-120581B inhibitor)U0126 (ERK inhibitors) Carbachol and human BDNF werepurchased from Sigma-Aldrich Inc (St Louis MO USA)Y-27632 dihydrochloride (Rho-associated protein kinaseinhibitor)was purchased fromTOCRIS (Avonmouth BristolUK) Wortmannin (phosphoinositide 3-kinase inhibitor)was purchased from Millipore (California USA) ArtepillinC was purchased from WAKO (Osaka Japan) RNAiso Pluswas purchased fromTakara (Hoto-kuOsaka Japan) Quanti-Tect Reverse Transcription Kit and Rotor-Gene SYBR GreenRT-PCR Kit were purchased from Qiagen (Hilden Ger-many) A Cell-Counting Kit (CCK-8) was purchased fromDojindo (Kumamoto Japan) Human recombinant IL-1120573was purchased from RampD (Minneapolis USA) 120573-Amyloid(1ndash42) was purchased from ANASPEC (California USA)dissolved in endotoxin-free sterile water and incubated at37∘C for 24 h to induce the fibril formation
22 SH-SY5Y Cell Culture Human neuroblastoma SH-SY5Ycells purchased from American Type Culture Collection
(Manassas VA USA) were maintained in a completegrowth medium (MEMF-12 mixture containing 10 fetalbovine serum supplemented with NaHCO
3and 100UmL
penicillin-streptomycin) The cells were cultured at 37∘C ina 5 CO
2humidified incubator
23 Cell Viability Assay SH-SY5Y cells were seeded in 96-well plates (5 times 103 cellswell) overnight After treatment withpropolis fA120573 and IL-1120573 a cell viability assay was performedusing a Cell-Counting Kit (Dojindo) in accordance with thepreviously described method [34] The optical density wasread at a wavelength of 450 nm with a microplate reader Thecell viability was calculated by dividing the optical density ofthe treated group by that of the control group
24 Observations of Morphological Changes The cells wereseeded in 6-well plates (2 times 105 cellsmL) for 24 h and thentreated with H
2O2at a concentration of 100120583M for 24 h
with or without propolis (methanol extraction) The cellularmorphology was observed and photographed using a bright-field microscope (Nikon ECLIPSE Ti-S Japan)
25 Detection of Mitochondrial ROS Mitochondrial ROSwere measured using MitoSOX Red (Invitrogen USA)which is a live-cell permeant that rapidly and selectively tar-gets mitochondria [37] Once in the mitochondria MitoSOXRed reagent is oxidized by superoxide and exhibits red fluo-rescence (with excitation at 510 nm and emission at 580 nm)The cultured SH-SY5Y cells were seeded in 24-well plates (2times 105 cellsmL) and incubated with or without propolis for 1 h(methanol extraction 10 120583gmL) The cells were then furtherexposed to H
2O2(100 120583M) for 1 h After incubation in Hankrsquos
balanced salt solution (HBSS) containing 5mM MitoSOXRed for 10min at 37∘C the cells were washed twice with PBSand thenmounted in a warm buffer for imaging Images werecollected using a fluorescence microscope (Nikon ECLIPSETi-S Japan)
26 Immunofluorescence Imaging Immunofluorescence ima-ging was performed as described previously [34 38] SH-SY5Y cells were seeded in 24-well plates (2 times 105 cellsmL)for 24 h and then treated with H
2O2(100 120583M) for 4 h with
or without pretreatment with propolis (methanol extraction10 120583gmL) for 1 h and fixed with 4 paraformaldehydeAfter washing the cells with PBS twice they were incubatedwith mouse anti-8-oxo-dG (1 500) overnight at 4∘C andthen incubated with anti-mouse Alexa 488 (1 2000 JacksonImmunoresearch Lab Inc) at 4∘C for 2 h After washing byPBS the nucleus was stained by Hoechst 33342 The cellswere mounted in the antifading medium Vecta shield andthe fluorescence images were taken using a confocal laserscanning microscope (CLSM C2si Nikon Japan)
27 Real-Time Quantitative Polymerase Chain Reaction (qRT-PCR) The mRNA isolated from the SH-SY5Y cells at var-ious time points were subjected to a real-time qRT-PCRThe total RNA was extracted using the RNAiso Plus inaccordance with the manufacturerrsquos instructions A total of800 ng of extracted RNA was reverse transcribed to cDNA
Oxidative Medicine and Cellular Longevity 3
using the QuantiTect Reverse Transcription Kit After aninitial denaturation step at 95∘C for 5min temperaturecycling was initiated Each cycle consisted of denaturationat 95∘C for 5 s annealing at 60∘C for 10 s and elongationfor 30 s In total 40 cycles were performed The cDNA wasamplified in duplicate using a Rotor-Gene SYBR Green RT-PCR Kit with a Corbett Rotor-Gene RG-3000A Real-TimePCR System (Sydney Australia) The data were evaluatedusing the RG-3000A software program (version Rotor-Gene6193 Corbett) The sequences of primer pairs were asfollows Arc 51015840-CCACCTGCTTGGACACCTC-31015840 and 51015840-CCGCCCCGAGGAGTTTG-31015840 BDNF 51015840-GGATGAGGA-CCAGAAAGT-31015840 and 51015840-AGCAGAAAGAGAAGAGGAG-31015840 actin 51015840-AGAGCTACGAGCTGCCTGAC-31015840 and 51015840-AGCACTGTGTTGGCGTACAG-31015840
For data normalization an endogenous control (actin)was assessed to control for the cDNA input and the relativeunits were calculated by a comparative Ct method All of thereal-time qRT-PCR experiments were repeated three timesand the results are presented as the means of the ratios plusmnstandard error of the mean
28 Electrophoresis and Immunoblotting SH-SY5Y cells werecultured at a density of 2 times 105 cellsmL Cells were har-vested at each time point with various stimulations andimmunoblotting analyses were conducted Briefly each spec-imen was electrophoresed using 12 SDS-polyacrylamidegels The proteins on the SDS gels were then electrophoret-ically transferred to nitrocellulose membranes Followingblocking the membranes were incubated at 4∘C overnightunder gentle agitation with each primary antibody mouseanti-Arc (1 1000 Abcam Heidelberg Germany) and mouseanti-actin (1 5000 Abcam) After washing the membraneswere incubated with horseradish peroxidase- (HRP-) labeledanti-mouse (1 2000 GE Healthcare Buckinghamshire UK)for 2 h at room temperature Subsequently the membrane-bound HRP-labeled antibodies were detected using anenhanced chemiluminescence detection system (ECK lit GEHealthcare) with an image analyzer (LAS-1000 Fuji PhotoFilm Minato-ku Tokyo Japan)
29 Statistical Analysis The data are represented as themeans plusmn standard error of the mean The statistical analyseswere performed by a one- or two-way analysis of variancewith a post hoc Tukeyrsquos test using the GraphPad Prism soft-ware package (GraphPad Software California USA) A valueof 119901 lt 005was considered to indicate statistical significance
3 Results
31 Effect of Propolis on Cell Viability in SH-SY5Y Cells Wefirst examined the effects of propolis on the viability of SH-SY5Y cells using CCK8 assay kit The mean cell viability wasnot significantly changed after treatment with the ethanolextracts of propolis at final concentrations between 025 and10 120583gmL for 48 h (Figure 1(a)) However themean cell viabil-ity was significantly reduced after pretreatment with propolisat a final concentration of over 25120583gmL (75 of viablecells) We therefore used methanol extracts of propolis at
concentrations up to 10120583gmL and H2O2at 100120583M concen-
tration [38] to investigate the effects of propolis on the H2O2-
induced cytotoxicity of SH-SY5Y cells in the subsequentexperiments The H
2O2-induced cell death (79 of viable
cells) was significantly restored by pretreatment with propolis(90 of viable cells) (Figure 1(b)) Morphological changesobserved included the degeneration of H
2O2-treated SH-
SY5Y cells which exhibited the disappearance of the neuritesand shrinkage (Figure 1(c))The percentage of viable cells wasalso reduced by H
2O2in a dose-dependent manner (data not
shown) It is noted that the neuritis and shrinkage of cellswere attenuated by pretreatment with propolis (Figure 1(c))These observations strongly demonstrate that pretreatmentwith propolis protects SH-SY5Y cells from H
2O2-induced
cytotoxicity Pretreatment with methanol extracts of propolis(10 120583gmL) for 1 h was set up in the subsequent experiments
32 Effects of Propolis on the H2O2-Induced Oxidative Stress
in SH-SY5Y Cells Oxidative stress is an important inducerof neurotoxicity in AD patients [39] following our previousexperiments we used two approaches to address the effectsof propolis on H
2O2-induced oxidative stress in SH-SY5Y
cells one approach was the use of a MitoSOX Red probe asa marker for mitochondria-derived ROS generation [35] andthe other was immunofluorescence imaging for a biomarkerof oxidation-damaged DNAmarker 8-oxo-dG [40] In com-parison to the untreated cells the expression of MitoSOXRed signals was significantly increased in SH-SY5Y cells afterexposure to H
2O2for 1 h suggesting that the mitochondria
are the early origin of ROS generation during oxidative stressPretreatment with propolis significantly inhibited the H
2O2-
induced mitochondria-derived ROS generation in SH-SY5Ycells (Figure 2(a)) and the mean fluorescent intensity ofMitoSOX Red oxidation was found to significantly increasein comparison to that in the cells that were not exposed toH2O2(493 versus 1 lowastlowastlowast119901 lt 0001 Figure 2(b)) Pretreat-
ment with the propolis methanol extracts for 1 h significantlyreduced the immunofluorescence intensity of MitoSOX Redoxidation in the H
2O2-exposed SH-SY5Y cells (18 versus
493 lowastlowastlowast119901 lt 0001 Figure 2(b)) thus confirming the antiox-idant properties of propolis Immunofluorescence imagingshowed a significant inverse relationship between Hoechstand 8-oxo-dG after exposure of SH-SY5Y cells to H
2O2for
4 h (Figure 2(c)) and the mean fluorescent intensity of 8-oxo-dG was found to significantly increase in comparisonto that in the cells that were not exposed to H
2O2(275
versus 1 lowastlowastlowast119901 lt 0001 Figure 2(d)) It is noted that pre-treatment with the propolis for 2 h significantly reduced theimmunofluorescence intensity of 8-oxo-dG in the H
2O2-
exposed SH-SY5Y cells (136 versus 275 lowastlowastlowast119901 lt 0001Figure 2(d)) These observations demonstrate that propoliscould attenuate oxidative stress in neuronal cells
33 Effect of Propolis on the Fibrillar A120573-Induced Impairmentof BDNF-Induced Arc Expression in SH-SY5Y Cells SolubleA120573 is known to interfere with synaptic efficacy as cognitivedecline precedes the formation of A120573 plaques the hallmarkof AD [41] We therefore examined the effects of propolis on
4 Oxidative Medicine and Cellular Longevity
0 025 05 1 5 10 25
lowastlowastlowast
0
20
40
60
80
100
120
140C
ell v
iabi
lity
()
Propolis concentration (gmL)
(a)
Con
lowastlowastlowast
lowastlowast
H2O2 H2O2 propolis50
60
70
80
90
100
110
Cel
l via
bilit
y (
)
(b)
Con H2O2 H2O2 propolis
(c)
Figure 1 Effects of methanol extracts of propolis on the H2O2-induced toxicity in SH-SY5Y cells (a) Cell viability in SH-SY5Y cells
pretreatment with different concentrations of propolis for 48 hours (b) The effect of pretreatment with methanol extracts of propolis for2 h on H
2O2exposed SH-SY5Y cells Each column and bar represent mean plusmn SEM (119899 = 4 each) An asterisk indicates a statistically significant
difference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001) (c) The morphological changes of SH-SY5Y cells with or withoutpretreatment with propolis (10120583gmL) after expose to H
2O2for 24 h Scale bar = 20 120583m
the impairment of BDNF-induced Arc expression by fA120573 inSH-SY5Y cells because Arc is the critical factor of synapseefficacyThemean cell viability was not significantly changedafter treatment with fA120573 at final concentrations between01 and 5 120583M for 48 h (Figure 3(a)) However significantcytotoxicity was observed in cultures treated with fA120573 over10 120583M (80 of viable cells) Therefore 5 120583M was used asthe nonlethal concentration for SH-SY5Y cells in subsequentexperiments Treatment with BDNF (10 ngmL) for 120mininduced 35-fold Arc expression in SH-SY5Y cells howeverpreexposure to fA120573 (5 120583M) for 6 h suppressed the BDNF-induced Arc expression (Figure 3(b)) Interestingly thisimpairment was significantly reversed following treatmentwith propolis (10 120583gmL) for 120min (Figure 3(b)) Further-more propolis treatment also prevented the fA120573-inducedimpairment of BDNF-induced Arc protein expression (Fig-ures 3(c) and 3(d)) These data indicate that propolis reversesthe fA120573-induced impairment of BDNF-induced Arc mRNAand protein expression
34 Effect of Propolis on the IL-1120573-Induced Impairment ofBDNF-Induced Arc Expression in SH-SY5Y Cells As a potent
activator for exacerbating neuroinflammation IL-1120573 hasrecently been found to suppress BDNF-dependent synapticefficacy [26] We therefore examined the effects of propolison the IL-1120573-induced impairment of BDNF-induced Arcexpression in SH-SY5Y cells We first examined the effectsof IL-1120573 on the cell viability to determine the nonlethalconcentration of IL-1120573 on SH-SY5Y cells The mean cellviability was not significantly changed after treatment withIL-1120573 at final concentrations between 1 and 500 ngmL for48 h (Figure 4(a)) However significant cytotoxicity wasobserved in cultures treated with IL-1120573 over 100 ngmL (90of viable cells) We therefore used 20 ngmL as the nonlethalconcentration for SH-SY5Y cells in subsequent experiments
Treatment with BDNF (10 ngmL) for 120min induced34-fold Arc expression in SH-SY5Y cells however preex-posure to IL-1120573 (20 ngmL) for 6 h significantly suppressedthe BDNF-induced Arc expression (Figure 4(b)) Interest-ingly this impairment was significantly reversed follow-ing treatment with propolis (10mgmL) for 120min (Fig-ure 4(b)) Furthermore propolis treatment also preventedthe IL-1120573-induced impairment of BDNF-inducedArc protein
Oxidative Medicine and Cellular Longevity 5
Hoechst MitoSOX Merge
Con
H2O2
H2O2propolis
(a)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
1
2
3
4
5
6
Relat
ive M
itoSO
X flu
ores
cenc
e den
sity
(b)
Hoechst Merge
Con
H2O2
H2O2propolis
8-oxo-dG
(c)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
05
1
15
2
25
3
35
Relat
ive 8
-oxo
-dG
fluo
resc
ence
den
sity
(d)
Figure 2 Effects of methanol extracts of propolis on the H2O2-induced oxidative stress in SH-SY5Y cells (a) Fluorescent images ofMitoSOX
Red signals in SH-SY5Y cells exposed to H2O2for 1 h with or without propolis (10120583gmL) for 2 h Scale bar = 15 120583m (b) The quantitative
analyses of MitoSOX Red signal intensity in (a) (c) Immunofluorescent CLMS images of 8-oxo-dG (red) with Hoechst-stained nuclei (blue)in SH-SY5Y cells exposed to 100 120583M of H
2O2for 4 h with or without pretreatment with propolis (10120583gmL) for 2 h Scale bar = 10 120583m (d)
The quantitative analyses of 8-oxo-dG immunofluorescence signal intensity in (c) The results are expressed as the mean plusmn SEM (119899 = 4 each)and the asterisks indicate a statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
expression (Figures 4(c) and 4(d)) These data indicate thatpropolis reverses the IL-1120573-induced impairment of BDNF-induced Arc mRNA and protein expression
35 Effect of Propolis on BDNF Expression in SH-SY5YCells We next examined the effects of propolis on BDNFexpression in SH-SY5Y cells because BDNF is the criticalneurotrophic factor that is important for synapse efficacyPretreatment with propolis significantly increased the BDNFmRNA expression in SH-SY5Y cells from 60min up to240min even at a low concentration (1120583gmL) (Figure 5(a))Furthermore treatment with propolis for 60min increasedthe BDNF mRNA expression in SH-SY5Y cells in a dose-dependent manner (Figure 5(b)) The propolis-increased
BNDF expression was completely abolished by preincubationwith Wortmannin (200 nM PI-3K inhibitor) but not bytreatment with Y27632 (1 120583M ROCK inhibitor) or GFX(200 nM PKC inhibitor) (Figure 5(c)) BDNFTrkB signalinghas been reported to play an important role in long termpotential to further confirm the BDNF signaling to Arc weuse a Carbachol to stimulate SH-SY5Y and the responses ofCarbachol were examined in the presence of U0126 (ERKinhibitors 10 120583M) or ANA-12 (TrkB selective antagonist100 nM)
1 h of treatment with Carbachol can significantly increasethe protein expression level of Arc However pretreatmentwith U0126 and ANA significantly reduced the Carbachol-increased Arc (Figures 5(d) and 5(e))) Therefore the
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

Oxidative Medicine and Cellular Longevity 3
using the QuantiTect Reverse Transcription Kit After aninitial denaturation step at 95∘C for 5min temperaturecycling was initiated Each cycle consisted of denaturationat 95∘C for 5 s annealing at 60∘C for 10 s and elongationfor 30 s In total 40 cycles were performed The cDNA wasamplified in duplicate using a Rotor-Gene SYBR Green RT-PCR Kit with a Corbett Rotor-Gene RG-3000A Real-TimePCR System (Sydney Australia) The data were evaluatedusing the RG-3000A software program (version Rotor-Gene6193 Corbett) The sequences of primer pairs were asfollows Arc 51015840-CCACCTGCTTGGACACCTC-31015840 and 51015840-CCGCCCCGAGGAGTTTG-31015840 BDNF 51015840-GGATGAGGA-CCAGAAAGT-31015840 and 51015840-AGCAGAAAGAGAAGAGGAG-31015840 actin 51015840-AGAGCTACGAGCTGCCTGAC-31015840 and 51015840-AGCACTGTGTTGGCGTACAG-31015840
For data normalization an endogenous control (actin)was assessed to control for the cDNA input and the relativeunits were calculated by a comparative Ct method All of thereal-time qRT-PCR experiments were repeated three timesand the results are presented as the means of the ratios plusmnstandard error of the mean
28 Electrophoresis and Immunoblotting SH-SY5Y cells werecultured at a density of 2 times 105 cellsmL Cells were har-vested at each time point with various stimulations andimmunoblotting analyses were conducted Briefly each spec-imen was electrophoresed using 12 SDS-polyacrylamidegels The proteins on the SDS gels were then electrophoret-ically transferred to nitrocellulose membranes Followingblocking the membranes were incubated at 4∘C overnightunder gentle agitation with each primary antibody mouseanti-Arc (1 1000 Abcam Heidelberg Germany) and mouseanti-actin (1 5000 Abcam) After washing the membraneswere incubated with horseradish peroxidase- (HRP-) labeledanti-mouse (1 2000 GE Healthcare Buckinghamshire UK)for 2 h at room temperature Subsequently the membrane-bound HRP-labeled antibodies were detected using anenhanced chemiluminescence detection system (ECK lit GEHealthcare) with an image analyzer (LAS-1000 Fuji PhotoFilm Minato-ku Tokyo Japan)
29 Statistical Analysis The data are represented as themeans plusmn standard error of the mean The statistical analyseswere performed by a one- or two-way analysis of variancewith a post hoc Tukeyrsquos test using the GraphPad Prism soft-ware package (GraphPad Software California USA) A valueof 119901 lt 005was considered to indicate statistical significance
3 Results
31 Effect of Propolis on Cell Viability in SH-SY5Y Cells Wefirst examined the effects of propolis on the viability of SH-SY5Y cells using CCK8 assay kit The mean cell viability wasnot significantly changed after treatment with the ethanolextracts of propolis at final concentrations between 025 and10 120583gmL for 48 h (Figure 1(a)) However themean cell viabil-ity was significantly reduced after pretreatment with propolisat a final concentration of over 25120583gmL (75 of viablecells) We therefore used methanol extracts of propolis at
concentrations up to 10120583gmL and H2O2at 100120583M concen-
tration [38] to investigate the effects of propolis on the H2O2-
induced cytotoxicity of SH-SY5Y cells in the subsequentexperiments The H
2O2-induced cell death (79 of viable
cells) was significantly restored by pretreatment with propolis(90 of viable cells) (Figure 1(b)) Morphological changesobserved included the degeneration of H
2O2-treated SH-
SY5Y cells which exhibited the disappearance of the neuritesand shrinkage (Figure 1(c))The percentage of viable cells wasalso reduced by H
2O2in a dose-dependent manner (data not
shown) It is noted that the neuritis and shrinkage of cellswere attenuated by pretreatment with propolis (Figure 1(c))These observations strongly demonstrate that pretreatmentwith propolis protects SH-SY5Y cells from H
2O2-induced
cytotoxicity Pretreatment with methanol extracts of propolis(10 120583gmL) for 1 h was set up in the subsequent experiments
32 Effects of Propolis on the H2O2-Induced Oxidative Stress
in SH-SY5Y Cells Oxidative stress is an important inducerof neurotoxicity in AD patients [39] following our previousexperiments we used two approaches to address the effectsof propolis on H
2O2-induced oxidative stress in SH-SY5Y
cells one approach was the use of a MitoSOX Red probe asa marker for mitochondria-derived ROS generation [35] andthe other was immunofluorescence imaging for a biomarkerof oxidation-damaged DNAmarker 8-oxo-dG [40] In com-parison to the untreated cells the expression of MitoSOXRed signals was significantly increased in SH-SY5Y cells afterexposure to H
2O2for 1 h suggesting that the mitochondria
are the early origin of ROS generation during oxidative stressPretreatment with propolis significantly inhibited the H
2O2-
induced mitochondria-derived ROS generation in SH-SY5Ycells (Figure 2(a)) and the mean fluorescent intensity ofMitoSOX Red oxidation was found to significantly increasein comparison to that in the cells that were not exposed toH2O2(493 versus 1 lowastlowastlowast119901 lt 0001 Figure 2(b)) Pretreat-
ment with the propolis methanol extracts for 1 h significantlyreduced the immunofluorescence intensity of MitoSOX Redoxidation in the H
2O2-exposed SH-SY5Y cells (18 versus
493 lowastlowastlowast119901 lt 0001 Figure 2(b)) thus confirming the antiox-idant properties of propolis Immunofluorescence imagingshowed a significant inverse relationship between Hoechstand 8-oxo-dG after exposure of SH-SY5Y cells to H
2O2for
4 h (Figure 2(c)) and the mean fluorescent intensity of 8-oxo-dG was found to significantly increase in comparisonto that in the cells that were not exposed to H
2O2(275
versus 1 lowastlowastlowast119901 lt 0001 Figure 2(d)) It is noted that pre-treatment with the propolis for 2 h significantly reduced theimmunofluorescence intensity of 8-oxo-dG in the H
2O2-
exposed SH-SY5Y cells (136 versus 275 lowastlowastlowast119901 lt 0001Figure 2(d)) These observations demonstrate that propoliscould attenuate oxidative stress in neuronal cells
33 Effect of Propolis on the Fibrillar A120573-Induced Impairmentof BDNF-Induced Arc Expression in SH-SY5Y Cells SolubleA120573 is known to interfere with synaptic efficacy as cognitivedecline precedes the formation of A120573 plaques the hallmarkof AD [41] We therefore examined the effects of propolis on
4 Oxidative Medicine and Cellular Longevity
0 025 05 1 5 10 25
lowastlowastlowast
0
20
40
60
80
100
120
140C
ell v
iabi
lity
()
Propolis concentration (gmL)
(a)
Con
lowastlowastlowast
lowastlowast
H2O2 H2O2 propolis50
60
70
80
90
100
110
Cel
l via
bilit
y (
)
(b)
Con H2O2 H2O2 propolis
(c)
Figure 1 Effects of methanol extracts of propolis on the H2O2-induced toxicity in SH-SY5Y cells (a) Cell viability in SH-SY5Y cells
pretreatment with different concentrations of propolis for 48 hours (b) The effect of pretreatment with methanol extracts of propolis for2 h on H
2O2exposed SH-SY5Y cells Each column and bar represent mean plusmn SEM (119899 = 4 each) An asterisk indicates a statistically significant
difference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001) (c) The morphological changes of SH-SY5Y cells with or withoutpretreatment with propolis (10120583gmL) after expose to H
2O2for 24 h Scale bar = 20 120583m
the impairment of BDNF-induced Arc expression by fA120573 inSH-SY5Y cells because Arc is the critical factor of synapseefficacyThemean cell viability was not significantly changedafter treatment with fA120573 at final concentrations between01 and 5 120583M for 48 h (Figure 3(a)) However significantcytotoxicity was observed in cultures treated with fA120573 over10 120583M (80 of viable cells) Therefore 5 120583M was used asthe nonlethal concentration for SH-SY5Y cells in subsequentexperiments Treatment with BDNF (10 ngmL) for 120mininduced 35-fold Arc expression in SH-SY5Y cells howeverpreexposure to fA120573 (5 120583M) for 6 h suppressed the BDNF-induced Arc expression (Figure 3(b)) Interestingly thisimpairment was significantly reversed following treatmentwith propolis (10 120583gmL) for 120min (Figure 3(b)) Further-more propolis treatment also prevented the fA120573-inducedimpairment of BDNF-induced Arc protein expression (Fig-ures 3(c) and 3(d)) These data indicate that propolis reversesthe fA120573-induced impairment of BDNF-induced Arc mRNAand protein expression
34 Effect of Propolis on the IL-1120573-Induced Impairment ofBDNF-Induced Arc Expression in SH-SY5Y Cells As a potent
activator for exacerbating neuroinflammation IL-1120573 hasrecently been found to suppress BDNF-dependent synapticefficacy [26] We therefore examined the effects of propolison the IL-1120573-induced impairment of BDNF-induced Arcexpression in SH-SY5Y cells We first examined the effectsof IL-1120573 on the cell viability to determine the nonlethalconcentration of IL-1120573 on SH-SY5Y cells The mean cellviability was not significantly changed after treatment withIL-1120573 at final concentrations between 1 and 500 ngmL for48 h (Figure 4(a)) However significant cytotoxicity wasobserved in cultures treated with IL-1120573 over 100 ngmL (90of viable cells) We therefore used 20 ngmL as the nonlethalconcentration for SH-SY5Y cells in subsequent experiments
Treatment with BDNF (10 ngmL) for 120min induced34-fold Arc expression in SH-SY5Y cells however preex-posure to IL-1120573 (20 ngmL) for 6 h significantly suppressedthe BDNF-induced Arc expression (Figure 4(b)) Interest-ingly this impairment was significantly reversed follow-ing treatment with propolis (10mgmL) for 120min (Fig-ure 4(b)) Furthermore propolis treatment also preventedthe IL-1120573-induced impairment of BDNF-inducedArc protein
Oxidative Medicine and Cellular Longevity 5
Hoechst MitoSOX Merge
Con
H2O2
H2O2propolis
(a)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
1
2
3
4
5
6
Relat
ive M
itoSO
X flu
ores
cenc
e den
sity
(b)
Hoechst Merge
Con
H2O2
H2O2propolis
8-oxo-dG
(c)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
05
1
15
2
25
3
35
Relat
ive 8
-oxo
-dG
fluo
resc
ence
den
sity
(d)
Figure 2 Effects of methanol extracts of propolis on the H2O2-induced oxidative stress in SH-SY5Y cells (a) Fluorescent images ofMitoSOX
Red signals in SH-SY5Y cells exposed to H2O2for 1 h with or without propolis (10120583gmL) for 2 h Scale bar = 15 120583m (b) The quantitative
analyses of MitoSOX Red signal intensity in (a) (c) Immunofluorescent CLMS images of 8-oxo-dG (red) with Hoechst-stained nuclei (blue)in SH-SY5Y cells exposed to 100 120583M of H
2O2for 4 h with or without pretreatment with propolis (10120583gmL) for 2 h Scale bar = 10 120583m (d)
The quantitative analyses of 8-oxo-dG immunofluorescence signal intensity in (c) The results are expressed as the mean plusmn SEM (119899 = 4 each)and the asterisks indicate a statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
expression (Figures 4(c) and 4(d)) These data indicate thatpropolis reverses the IL-1120573-induced impairment of BDNF-induced Arc mRNA and protein expression
35 Effect of Propolis on BDNF Expression in SH-SY5YCells We next examined the effects of propolis on BDNFexpression in SH-SY5Y cells because BDNF is the criticalneurotrophic factor that is important for synapse efficacyPretreatment with propolis significantly increased the BDNFmRNA expression in SH-SY5Y cells from 60min up to240min even at a low concentration (1120583gmL) (Figure 5(a))Furthermore treatment with propolis for 60min increasedthe BDNF mRNA expression in SH-SY5Y cells in a dose-dependent manner (Figure 5(b)) The propolis-increased
BNDF expression was completely abolished by preincubationwith Wortmannin (200 nM PI-3K inhibitor) but not bytreatment with Y27632 (1 120583M ROCK inhibitor) or GFX(200 nM PKC inhibitor) (Figure 5(c)) BDNFTrkB signalinghas been reported to play an important role in long termpotential to further confirm the BDNF signaling to Arc weuse a Carbachol to stimulate SH-SY5Y and the responses ofCarbachol were examined in the presence of U0126 (ERKinhibitors 10 120583M) or ANA-12 (TrkB selective antagonist100 nM)
1 h of treatment with Carbachol can significantly increasethe protein expression level of Arc However pretreatmentwith U0126 and ANA significantly reduced the Carbachol-increased Arc (Figures 5(d) and 5(e))) Therefore the
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
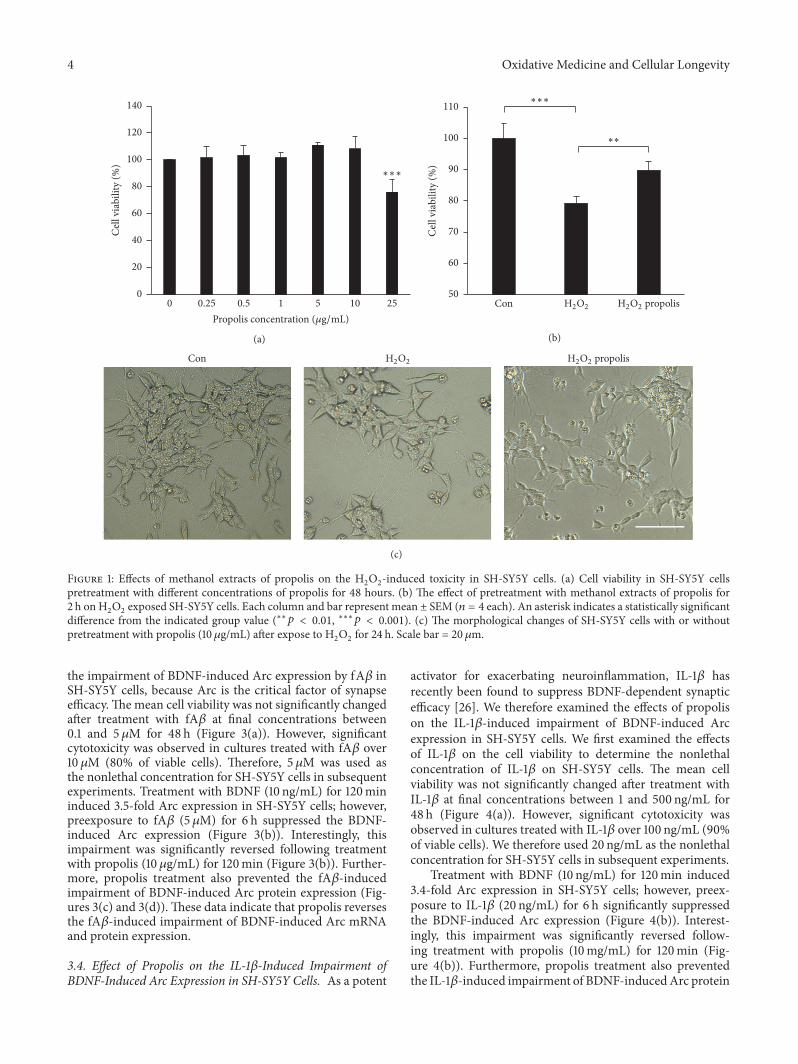
4 Oxidative Medicine and Cellular Longevity
0 025 05 1 5 10 25
lowastlowastlowast
0
20
40
60
80
100
120
140C
ell v
iabi
lity
()
Propolis concentration (gmL)
(a)
Con
lowastlowastlowast
lowastlowast
H2O2 H2O2 propolis50
60
70
80
90
100
110
Cel
l via
bilit
y (
)
(b)
Con H2O2 H2O2 propolis
(c)
Figure 1 Effects of methanol extracts of propolis on the H2O2-induced toxicity in SH-SY5Y cells (a) Cell viability in SH-SY5Y cells
pretreatment with different concentrations of propolis for 48 hours (b) The effect of pretreatment with methanol extracts of propolis for2 h on H
2O2exposed SH-SY5Y cells Each column and bar represent mean plusmn SEM (119899 = 4 each) An asterisk indicates a statistically significant
difference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001) (c) The morphological changes of SH-SY5Y cells with or withoutpretreatment with propolis (10120583gmL) after expose to H
2O2for 24 h Scale bar = 20 120583m
the impairment of BDNF-induced Arc expression by fA120573 inSH-SY5Y cells because Arc is the critical factor of synapseefficacyThemean cell viability was not significantly changedafter treatment with fA120573 at final concentrations between01 and 5 120583M for 48 h (Figure 3(a)) However significantcytotoxicity was observed in cultures treated with fA120573 over10 120583M (80 of viable cells) Therefore 5 120583M was used asthe nonlethal concentration for SH-SY5Y cells in subsequentexperiments Treatment with BDNF (10 ngmL) for 120mininduced 35-fold Arc expression in SH-SY5Y cells howeverpreexposure to fA120573 (5 120583M) for 6 h suppressed the BDNF-induced Arc expression (Figure 3(b)) Interestingly thisimpairment was significantly reversed following treatmentwith propolis (10 120583gmL) for 120min (Figure 3(b)) Further-more propolis treatment also prevented the fA120573-inducedimpairment of BDNF-induced Arc protein expression (Fig-ures 3(c) and 3(d)) These data indicate that propolis reversesthe fA120573-induced impairment of BDNF-induced Arc mRNAand protein expression
34 Effect of Propolis on the IL-1120573-Induced Impairment ofBDNF-Induced Arc Expression in SH-SY5Y Cells As a potent
activator for exacerbating neuroinflammation IL-1120573 hasrecently been found to suppress BDNF-dependent synapticefficacy [26] We therefore examined the effects of propolison the IL-1120573-induced impairment of BDNF-induced Arcexpression in SH-SY5Y cells We first examined the effectsof IL-1120573 on the cell viability to determine the nonlethalconcentration of IL-1120573 on SH-SY5Y cells The mean cellviability was not significantly changed after treatment withIL-1120573 at final concentrations between 1 and 500 ngmL for48 h (Figure 4(a)) However significant cytotoxicity wasobserved in cultures treated with IL-1120573 over 100 ngmL (90of viable cells) We therefore used 20 ngmL as the nonlethalconcentration for SH-SY5Y cells in subsequent experiments
Treatment with BDNF (10 ngmL) for 120min induced34-fold Arc expression in SH-SY5Y cells however preex-posure to IL-1120573 (20 ngmL) for 6 h significantly suppressedthe BDNF-induced Arc expression (Figure 4(b)) Interest-ingly this impairment was significantly reversed follow-ing treatment with propolis (10mgmL) for 120min (Fig-ure 4(b)) Furthermore propolis treatment also preventedthe IL-1120573-induced impairment of BDNF-inducedArc protein
Oxidative Medicine and Cellular Longevity 5
Hoechst MitoSOX Merge
Con
H2O2
H2O2propolis
(a)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
1
2
3
4
5
6
Relat
ive M
itoSO
X flu
ores
cenc
e den
sity
(b)
Hoechst Merge
Con
H2O2
H2O2propolis
8-oxo-dG
(c)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
05
1
15
2
25
3
35
Relat
ive 8
-oxo
-dG
fluo
resc
ence
den
sity
(d)
Figure 2 Effects of methanol extracts of propolis on the H2O2-induced oxidative stress in SH-SY5Y cells (a) Fluorescent images ofMitoSOX
Red signals in SH-SY5Y cells exposed to H2O2for 1 h with or without propolis (10120583gmL) for 2 h Scale bar = 15 120583m (b) The quantitative
analyses of MitoSOX Red signal intensity in (a) (c) Immunofluorescent CLMS images of 8-oxo-dG (red) with Hoechst-stained nuclei (blue)in SH-SY5Y cells exposed to 100 120583M of H
2O2for 4 h with or without pretreatment with propolis (10120583gmL) for 2 h Scale bar = 10 120583m (d)
The quantitative analyses of 8-oxo-dG immunofluorescence signal intensity in (c) The results are expressed as the mean plusmn SEM (119899 = 4 each)and the asterisks indicate a statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
expression (Figures 4(c) and 4(d)) These data indicate thatpropolis reverses the IL-1120573-induced impairment of BDNF-induced Arc mRNA and protein expression
35 Effect of Propolis on BDNF Expression in SH-SY5YCells We next examined the effects of propolis on BDNFexpression in SH-SY5Y cells because BDNF is the criticalneurotrophic factor that is important for synapse efficacyPretreatment with propolis significantly increased the BDNFmRNA expression in SH-SY5Y cells from 60min up to240min even at a low concentration (1120583gmL) (Figure 5(a))Furthermore treatment with propolis for 60min increasedthe BDNF mRNA expression in SH-SY5Y cells in a dose-dependent manner (Figure 5(b)) The propolis-increased
BNDF expression was completely abolished by preincubationwith Wortmannin (200 nM PI-3K inhibitor) but not bytreatment with Y27632 (1 120583M ROCK inhibitor) or GFX(200 nM PKC inhibitor) (Figure 5(c)) BDNFTrkB signalinghas been reported to play an important role in long termpotential to further confirm the BDNF signaling to Arc weuse a Carbachol to stimulate SH-SY5Y and the responses ofCarbachol were examined in the presence of U0126 (ERKinhibitors 10 120583M) or ANA-12 (TrkB selective antagonist100 nM)
1 h of treatment with Carbachol can significantly increasethe protein expression level of Arc However pretreatmentwith U0126 and ANA significantly reduced the Carbachol-increased Arc (Figures 5(d) and 5(e))) Therefore the
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
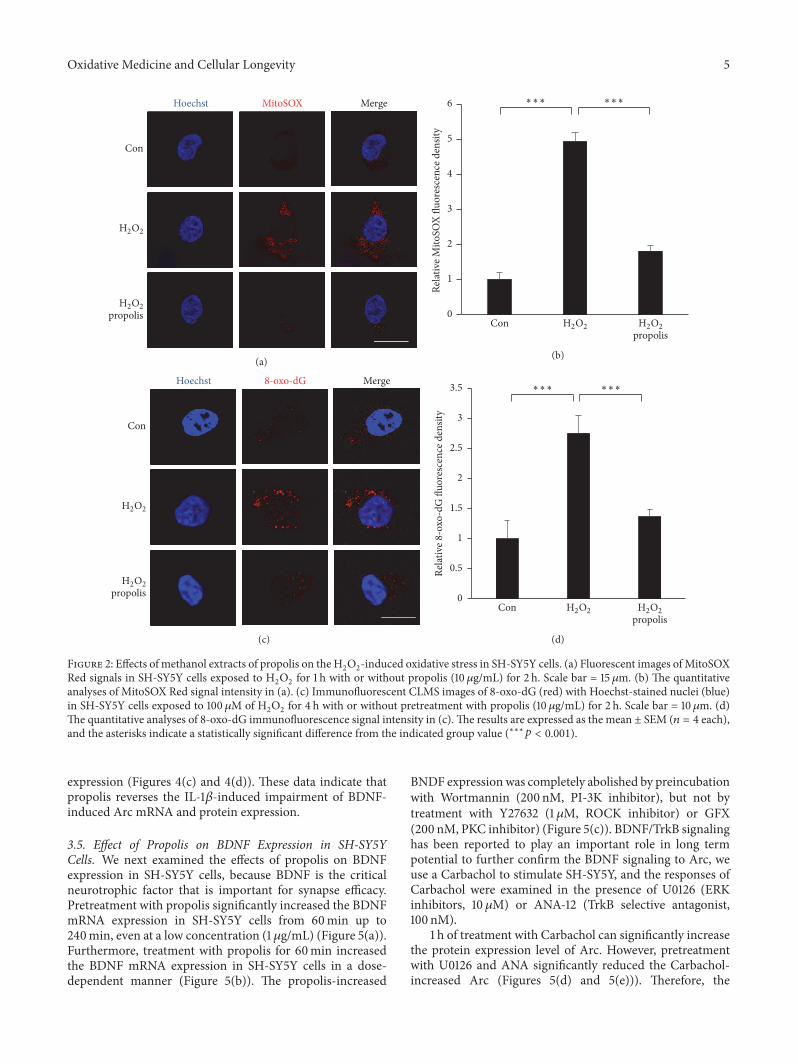
Oxidative Medicine and Cellular Longevity 5
Hoechst MitoSOX Merge
Con
H2O2
H2O2propolis
(a)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
1
2
3
4
5
6
Relat
ive M
itoSO
X flu
ores
cenc
e den
sity
(b)
Hoechst Merge
Con
H2O2
H2O2propolis
8-oxo-dG
(c)
Con H2O2 H2O2propolis
lowastlowastlowast lowastlowastlowast
0
05
1
15
2
25
3
35
Relat
ive 8
-oxo
-dG
fluo
resc
ence
den
sity
(d)
Figure 2 Effects of methanol extracts of propolis on the H2O2-induced oxidative stress in SH-SY5Y cells (a) Fluorescent images ofMitoSOX
Red signals in SH-SY5Y cells exposed to H2O2for 1 h with or without propolis (10120583gmL) for 2 h Scale bar = 15 120583m (b) The quantitative
analyses of MitoSOX Red signal intensity in (a) (c) Immunofluorescent CLMS images of 8-oxo-dG (red) with Hoechst-stained nuclei (blue)in SH-SY5Y cells exposed to 100 120583M of H
2O2for 4 h with or without pretreatment with propolis (10120583gmL) for 2 h Scale bar = 10 120583m (d)
The quantitative analyses of 8-oxo-dG immunofluorescence signal intensity in (c) The results are expressed as the mean plusmn SEM (119899 = 4 each)and the asterisks indicate a statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
expression (Figures 4(c) and 4(d)) These data indicate thatpropolis reverses the IL-1120573-induced impairment of BDNF-induced Arc mRNA and protein expression
35 Effect of Propolis on BDNF Expression in SH-SY5YCells We next examined the effects of propolis on BDNFexpression in SH-SY5Y cells because BDNF is the criticalneurotrophic factor that is important for synapse efficacyPretreatment with propolis significantly increased the BDNFmRNA expression in SH-SY5Y cells from 60min up to240min even at a low concentration (1120583gmL) (Figure 5(a))Furthermore treatment with propolis for 60min increasedthe BDNF mRNA expression in SH-SY5Y cells in a dose-dependent manner (Figure 5(b)) The propolis-increased
BNDF expression was completely abolished by preincubationwith Wortmannin (200 nM PI-3K inhibitor) but not bytreatment with Y27632 (1 120583M ROCK inhibitor) or GFX(200 nM PKC inhibitor) (Figure 5(c)) BDNFTrkB signalinghas been reported to play an important role in long termpotential to further confirm the BDNF signaling to Arc weuse a Carbachol to stimulate SH-SY5Y and the responses ofCarbachol were examined in the presence of U0126 (ERKinhibitors 10 120583M) or ANA-12 (TrkB selective antagonist100 nM)
1 h of treatment with Carbachol can significantly increasethe protein expression level of Arc However pretreatmentwith U0126 and ANA significantly reduced the Carbachol-increased Arc (Figures 5(d) and 5(e))) Therefore the
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
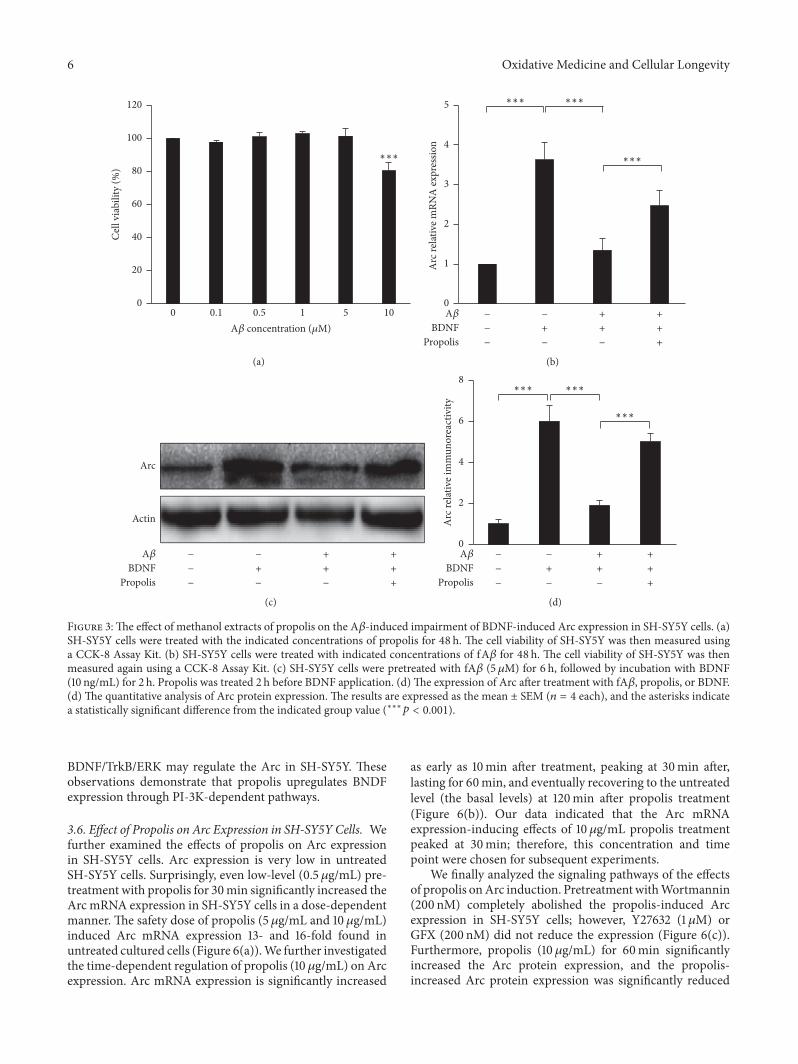
6 Oxidative Medicine and Cellular Longevity
0 01 05 1 5 10
lowastlowastlowast
0
20
40
60
80
100
120C
ell v
iabi
lity
()
A concentration (M)
(a)
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
0
1
2
3
4
5
Arc
relat
ive m
RNA
expr
essio
n
BDNFPropolis
A +
+
+
+
+ +
minus
minus
minusminus
minus
minus
(b)
Actin
Arc
minusBDNFPropolis
A+
+ +
+
+
+
minus
minus minus
minus
minus
(c)
lowastlowastlowast lowastlowastlowast
lowastlowastlowast
BDNFPropolis
A+
+ +
+
+
+
minus
minus
minus minus minus
minus0
2
4
6
8
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 3The effect of methanol extracts of propolis on the A120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells (a)SH-SY5Y cells were treated with the indicated concentrations of propolis for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were treated with indicated concentrations of fA120573 for 48 h The cell viability of SH-SY5Y was thenmeasured again using a CCK-8 Assay Kit (c) SH-SY5Y cells were pretreated with fA120573 (5120583M) for 6 h followed by incubation with BDNF(10 ngmL) for 2 h Propolis was treated 2 h before BDNF application (d)The expression of Arc after treatment with fA120573 propolis or BDNF(d) The quantitative analysis of Arc protein expression The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicatea statistically significant difference from the indicated group value (lowastlowastlowast119901 lt 0001)
BDNFTrkBERK may regulate the Arc in SH-SY5Y Theseobservations demonstrate that propolis upregulates BNDFexpression through PI-3K-dependent pathways
36 Effect of Propolis on Arc Expression in SH-SY5Y Cells Wefurther examined the effects of propolis on Arc expressionin SH-SY5Y cells Arc expression is very low in untreatedSH-SY5Y cells Surprisingly even low-level (05 120583gmL) pre-treatment with propolis for 30min significantly increased theArc mRNA expression in SH-SY5Y cells in a dose-dependentmanner The safety dose of propolis (5 120583gmL and 10 120583gmL)induced Arc mRNA expression 13- and 16-fold found inuntreated cultured cells (Figure 6(a))We further investigatedthe time-dependent regulation of propolis (10120583gmL) on Arcexpression Arc mRNA expression is significantly increased
as early as 10min after treatment peaking at 30min afterlasting for 60min and eventually recovering to the untreatedlevel (the basal levels) at 120min after propolis treatment(Figure 6(b)) Our data indicated that the Arc mRNAexpression-inducing effects of 10120583gmL propolis treatmentpeaked at 30min therefore this concentration and timepoint were chosen for subsequent experiments
We finally analyzed the signaling pathways of the effectsof propolis onArc induction PretreatmentwithWortmannin(200 nM) completely abolished the propolis-induced Arcexpression in SH-SY5Y cells however Y27632 (1120583M) orGFX (200 nM) did not reduce the expression (Figure 6(c))Furthermore propolis (10 120583gmL) for 60min significantlyincreased the Arc protein expression and the propolis-increased Arc protein expression was significantly reduced
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
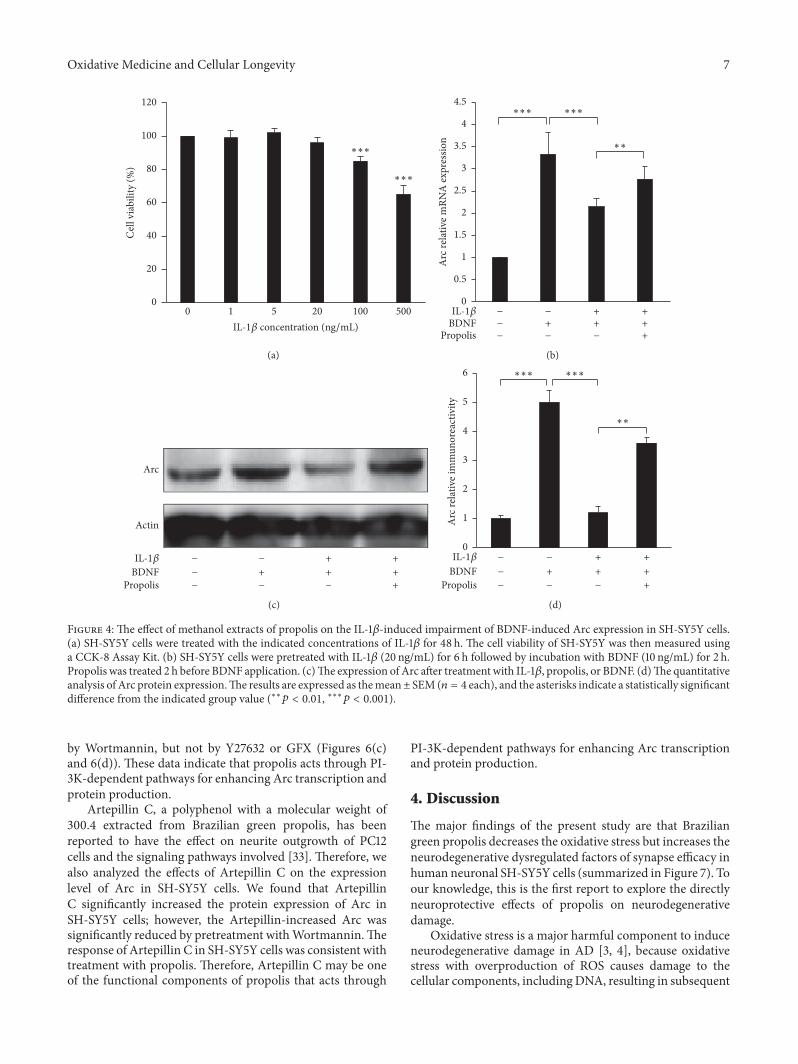
Oxidative Medicine and Cellular Longevity 7
0 1 5 20 100 5000
20
40
60
80
100
120
Cel
l via
bilit
y (
)
lowastlowastlowast
lowastlowastlowast
IL-1 concentration (ngmL)
(a)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
05
1
15
2
25
3
35
4
45
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
minusminusminus
minus+minus
+++
++minus
BDNFPropolis
IL-1
(c)
lowastlowastlowast lowastlowastlowast
lowastlowast
BDNFPropolis
minus minus ++minus + ++minus minus +minus
IL-10
1
2
3
4
5
6
Arc
relat
ive i
mm
unor
eact
ivity
(d)
Figure 4 The effect of methanol extracts of propolis on the IL-1120573-induced impairment of BDNF-induced Arc expression in SH-SY5Y cells(a) SH-SY5Y cells were treated with the indicated concentrations of IL-1120573 for 48 h The cell viability of SH-SY5Y was then measured usinga CCK-8 Assay Kit (b) SH-SY5Y cells were pretreated with IL-1120573 (20 ngmL) for 6 h followed by incubation with BDNF (10 ngmL) for 2 hPropolis was treated 2 h before BDNF application (c)The expression of Arc after treatment with IL-1120573 propolis or BDNF (d)The quantitativeanalysis of Arc protein expressionThe results are expressed as themeanplusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the indicated group value (lowastlowast119901 lt 001 lowastlowastlowast119901 lt 0001)
by Wortmannin but not by Y27632 or GFX (Figures 6(c)and 6(d)) These data indicate that propolis acts through PI-3K-dependent pathways for enhancing Arc transcription andprotein production
Artepillin C a polyphenol with a molecular weight of3004 extracted from Brazilian green propolis has beenreported to have the effect on neurite outgrowth of PC12cells and the signaling pathways involved [33] Therefore wealso analyzed the effects of Artepillin C on the expressionlevel of Arc in SH-SY5Y cells We found that ArtepillinC significantly increased the protein expression of Arc inSH-SY5Y cells however the Artepillin-increased Arc wassignificantly reduced by pretreatment withWortmanninTheresponse of Artepillin C in SH-SY5Y cells was consistent withtreatment with propolis Therefore Artepillin C may be oneof the functional components of propolis that acts through
PI-3K-dependent pathways for enhancing Arc transcriptionand protein production
4 Discussion
The major findings of the present study are that Braziliangreen propolis decreases the oxidative stress but increases theneurodegenerative dysregulated factors of synapse efficacy inhuman neuronal SH-SY5Y cells (summarized in Figure 7) Toour knowledge this is the first report to explore the directlyneuroprotective effects of propolis on neurodegenerativedamage
Oxidative stress is a major harmful component to induceneurodegenerative damage in AD [3 4] because oxidativestress with overproduction of ROS causes damage to thecellular components includingDNA resulting in subsequent
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
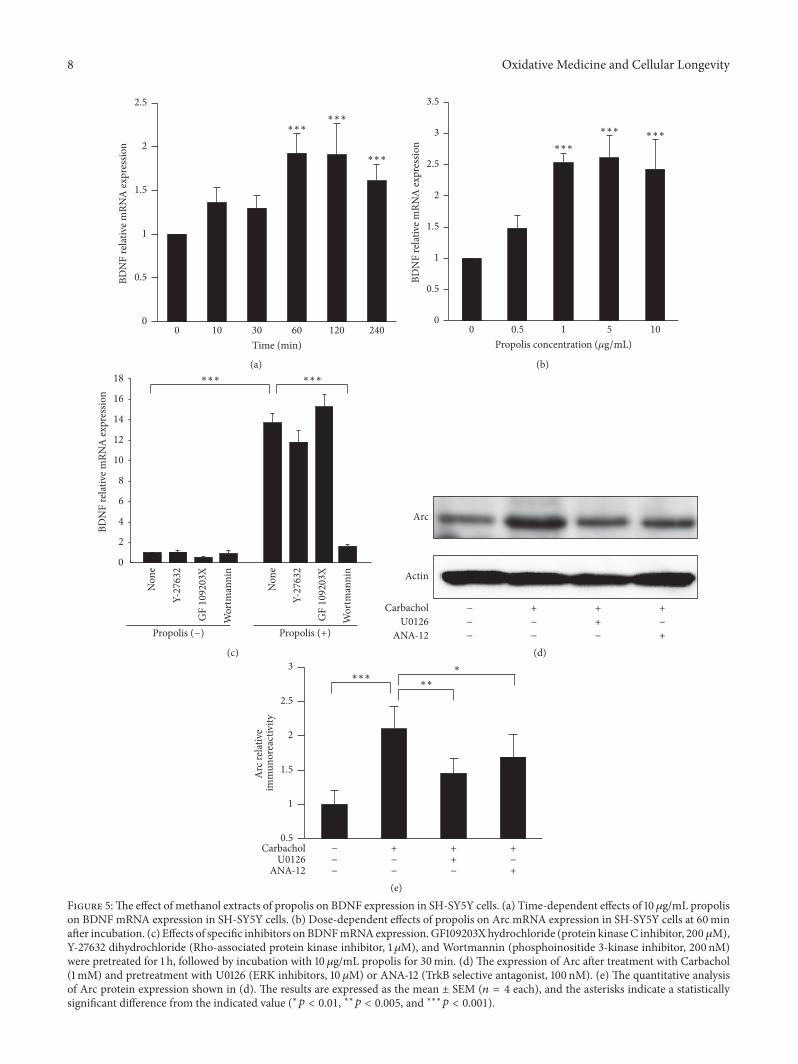
8 Oxidative Medicine and Cellular Longevity
0 10 30 60 120 240Time (min)
0
05
1
15
2
25
BDN
F re
lativ
e mRN
A ex
pres
sion
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
(a)
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
05
1
15
2
25
3
35
BDN
F re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(b)
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
lowastlowastlowast lowastlowastlowast
Propolis (+)Propolis (minus)
BDN
F re
lativ
e mRN
A ex
pres
sion
18
14
16
10
12
2
6
4
0
8
(c)
Actin
Arc
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
(d)
U0126Carbachol
ANA-12
minusminusminus
+minusminus
+minus+
++minus
Arc
relat
ive
lowastlowastlowastlowastlowast
lowast
05
1
15
2
25
3
imm
unor
eact
ivity
(e)
Figure 5The effect of methanol extracts of propolis on BDNF expression in SH-SY5Y cells (a) Time-dependent effects of 10120583gmL propolison BDNF mRNA expression in SH-SY5Y cells (b) Dose-dependent effects of propolis on Arc mRNA expression in SH-SY5Y cells at 60minafter incubation (c) Effects of specific inhibitors onBDNFmRNAexpression GF109203Xhydrochloride (protein kinaseC inhibitor 200 120583M)Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) and Wortmannin (phosphoinositide 3-kinase inhibitor 200 nM)were pretreated for 1 h followed by incubation with 10 120583gmL propolis for 30min (d) The expression of Arc after treatment with Carbachol(1mM) and pretreatment with U0126 (ERK inhibitors 10 120583M) or ANA-12 (TrkB selective antagonist 100 nM) (e) The quantitative analysisof Arc protein expression shown in (d) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statisticallysignificant difference from the indicated value (lowast119901 lt 001 lowastlowast119901 lt 0005 and lowastlowastlowast119901 lt 0001)
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003
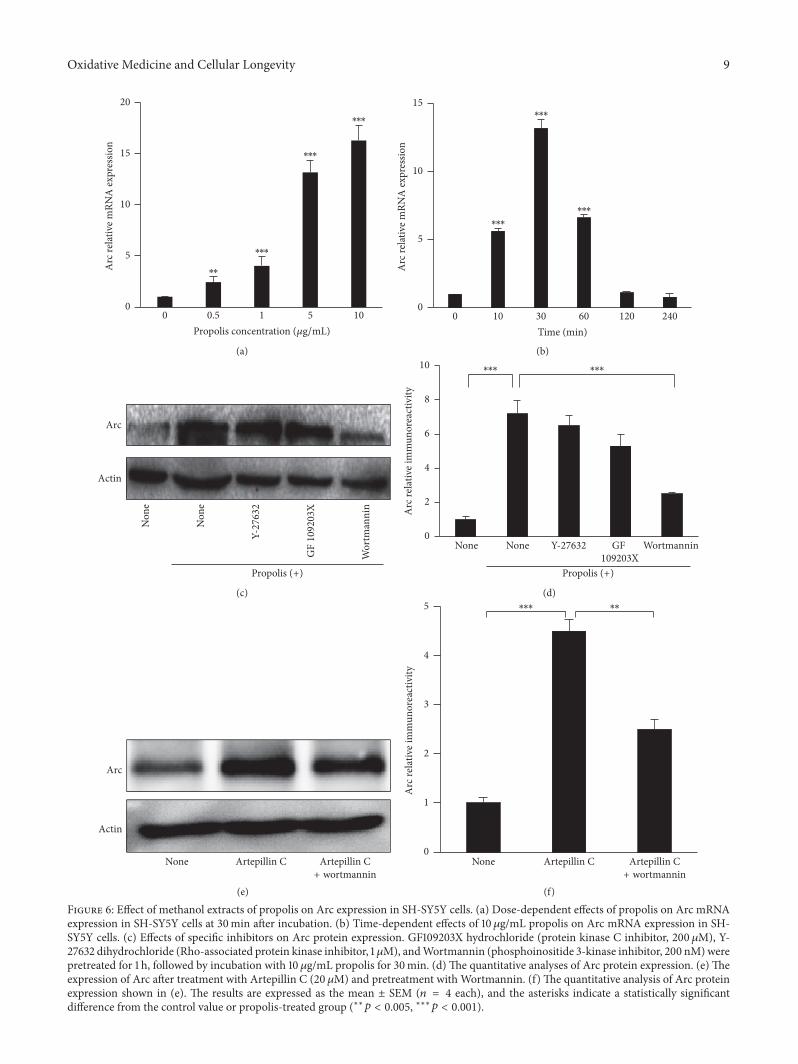
Oxidative Medicine and Cellular Longevity 9
0 05 1 5 10
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
0
5
10
15
20A
rc re
lativ
e mRN
A ex
pres
sion
Propolis concentration (gmL)
(a)
0 10 30 60 120 240Time (min)
lowastlowastlowast
lowastlowastlowast
lowastlowastlowast
0
5
10
15
Arc
relat
ive m
RNA
expr
essio
n
(b)
Actin
Arc
Non
e
Non
e
Y-27
632
GF
1092
03X
Wor
tman
nin
Propolis (+)
(c)
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowastlowast
None None Y-27632 GF 109203X
Wortmannin0
2
4
6
8
10
Propolis (+)
(d)
Actin
Arc
None Artepillin C Artepillin C+ wortmannin
(e)
0
1
2
3
4
5
None Artepillin C Artepillin C+ wortmannin
Arc
rela
tive i
mm
unor
eact
ivity
lowastlowastlowast lowastlowast
(f)
Figure 6 Effect of methanol extracts of propolis on Arc expression in SH-SY5Y cells (a) Dose-dependent effects of propolis on Arc mRNAexpression in SH-SY5Y cells at 30min after incubation (b) Time-dependent effects of 10120583gmL propolis on Arc mRNA expression in SH-SY5Y cells (c) Effects of specific inhibitors on Arc protein expression GF109203X hydrochloride (protein kinase C inhibitor 200120583M) Y-27632 dihydrochloride (Rho-associated protein kinase inhibitor 1120583M) andWortmannin (phosphoinositide 3-kinase inhibitor 200 nM)werepretreated for 1 h followed by incubation with 10120583gmL propolis for 30min (d) The quantitative analyses of Arc protein expression (e) Theexpression of Arc after treatment with Artepillin C (20120583M) and pretreatment with Wortmannin (f) The quantitative analysis of Arc proteinexpression shown in (e) The results are expressed as the mean plusmn SEM (119899 = 4 each) and the asterisks indicate a statistically significantdifference from the control value or propolis-treated group (lowastlowast119901 lt 0005 lowastlowastlowast119901 lt 0001)
10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

10 Oxidative Medicine and Cellular Longevity
AIL-1
Synaptic efficacy
PI3K
Alzheimerrsquos disease
Normalaging
BDNF
Propolis
Arc
Oxidativestress
Figure 7 A schematic representation of the effects and the principlemolecular mechanisms of Brazilian green propolis on neurode-generative damage-induced oxidative stress and downregulation ofsynaptic efficacy
cell death Antioxidant therapy therefore is considered asan approach in prevention and clinical management ofAD [6] We used an in vitro model of human neuronalSH-SY5Y cells to investigate the direct effect of Braziliangreen propolis (propolis) and its signaling transductions onArc expression to avoid other effectors Pretreatment withpropolis could protect the H
2O2-induced cell death which
agreed with those of the previous reports showing baccharinp-Coumaric acid and Artepillin C the active componentsof propolis are effective in reducing neurotoxicity [29ndash32]We found that propolis significantly inhibited the H
2O2-
induced mitochondria ROS generation as well as nuclearDNA damage in SH-SY5Y cells (Figures 1 and 2) Thuswe provide the first evidence that propolis attenuates theoxidative stress-induced DNA damage directly in neuronalcells
The dysfunction of synaptic efficacy is an early neurode-generative damage in AD [1 21] because the impairmentof synaptic activation precedes substantial A120573 accumulationand neuron loss in the brain [10 41 42] BDNF-inducedArc expression is widely used as a marker of defectivesynaptic efficacy [21 26 43] In the present study we foundthat preexposure to fA120573 (a neurodegenerative hallmark) ata nonlethal concentration (5120583M) impaired BDNF-inducedArc expression (Figure 3(b)) however fA120573 at a nonlethalconcentration did not affect Arc expression in SH-SY5Ycells (data not shown) These data were consistent with thefindings of previous studies showing that BDNF-induced Arcexpression was inhibited by pretreatment with oligomeric A120573or fA120573 in cultured cortical neurons [21 26 44] Surprisinglytreatment with (10 120583gmL) for 120min significantly reversedthe fA120573-induced impairment of BDNF-induced Arc expres-sion in SH-SY5Y cells Since a decline in Arc expression is
correlated with cognitive impairment in AD [45 46] BDNF-induced Arc expression fundamentally regulates the synapticefficacy [17 18]These observations indicate that propolis canprevent the fA120573-induced dysfunction of synaptic efficacy inneuronal cells
Increasing evidence indicates that microglia-related neu-roinflammation contributes to the decline in cognitive func-tion during aging as well as in AD [47 48] As a potentactivator for exacerbating neuroinflammation IL-1120573has beenreported to suppress BDNF-dependent synaptic efficacyresulting in cognitive impairment [26 49] Furthermorethe overexpression of IL-1120573 in a transgenic mouse modelresulted in increased microglia activation with a significantreduction in behaviorally induced Arc levels and impairedcontextual and spatial memory [50] In the present studywe demonstrated that nonlethal concentrations of IL-1120573(20 ngmL) impaired BDNF-induced Arc expression in SH-SY5Y cells (Figures 4(b)ndash4(d)) a finding that agreed withthose of a previous report showing that IL-1120573 suppressed theBDNF-induced Arc expression in cultured brain slices [26]Importantly treatment with propolis significantly reversedthe IL-1120573-induced impairment of BDNF-inducedArc expres-sionThese observations indicate that propolis can reverse theneuroinflammation-induced dysfunction of synaptic efficacyin neuronal cells
BDNF as a neurotropic factor fundamentally controlsthe synaptic efficacy [17 51] and closely correlates withcognitive functions [14 19] Indeed BDNF levels are reducedeven in the preclinical stages of AD [52] In the presentstudy treatment with propolis significantly increased theBDNF expression in SH-SY5Y cells in dose- and time-dependent manners Furthermore the propolis-upregulatedBNDF expression was completely abolished by preincubationwith PI3K inhibitor indicating that the effects of propolison BDNF expression are mediated by the PI3K signalingpathway
Arc expression was low in the untreated SH-SY5Y cellswhich agreedwith the findings of other reports using culturedprimary cortical neurons [53 54] Exogenous treatment withpropolis induced Arc expression in SH-SY5Y cells in bothdose- and time-dependent manners Of note exposure topropolis induced rapid robust Arc mRNA expression asquickly as 10min after exposure and peaking at 30min whichparalleled the Arc expression peaks around 30min afterbehavior induction [55]Therefore Arc may serve as an earlybeginning effector molecule for propolis-induced neuronalactivity
The PI3K signaling pathway plays a pivotal role in synap-tic efficacy and memory consolidation [56ndash58] The obser-vations of which the effects of propolis on Arc expressionare mediated by the PI-3K signaling pathway thus providethe underlying mechanism of propolis in directly regulat-ing the molecule for synaptic efficacy in human neuronalcells Artepillin C a major component of Brazilian greenpropolis has also been found to act as a neurotrophic-likefactor for promoting NGF-induced neurite outgrowth [33]Our ongoing human research at high altitude shows thatelderly individuals take propolis score significantly higher oncognitive tests than nontreated individuals (Wu amp Zhu et al
Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

Oxidative Medicine and Cellular Longevity 11
unpublished data) The neuroprotective effects of Braziliangreen propolis on neurodegenerative damage might providea valuable therapeutic strategy for prevention of cognitiveimpairment in AD as well as aging
5 Conclusion
Brazilian green propolis could reduce oxidative stress andprevent the neurodegenerative damaged synapse efficacy(schematic represented in Figure 7) Therefore we pro-vide the principle molecular mechanisms of the benefits ofpropolis as the therapeutic agent for maintenance cognitivefunction of brain
Competing Interests
Theauthors declare no conflict of interests in associationwiththis study
Authorsrsquo Contributions
Junjun Ni and Zhou Wu contributed equally to this work
Acknowledgments
This work was supported by a Yamada Research Grant toZhou Wu (N00183)
References
[1] D J Selkoe ldquoAlzheimerrsquos disease is a synaptic failurerdquo Sciencevol 298 no 5594 pp 789ndash791 2002
[2] R Brookmeyer D A Evans LHebert et al ldquoNational estimatesof the prevalence of Alzheimerrsquos disease in the United StatesrdquoAlzheimerrsquos amp Dementia vol 7 no 1 pp 61ndash73 2011
[3] L Y Al-Ayadhi ldquoOxidative stress and neurodegenerative dis-easerdquo Neurosciences vol 9 no 1 pp 19ndash23 2004
[4] N H Zawia D K Lahiri and F Cardozo-Pelaez ldquoEpigeneticsoxidative stress and Alzheimer diseaserdquo Free Radical Biologyand Medicine vol 46 no 9 pp 1241ndash1249 2009
[5] H-U Simon A Haj-Yehia and F Levi-Schaffer ldquoRole of reac-tive oxygen species (ROS) in apoptosis inductionrdquo Apoptosisvol 5 no 5 pp 415ndash418 2000
[6] M Manczak P Mao M J Calkins et al ldquoMitochondria-targeted antioxidants protect against amyloid-120573 toxicity inAlzheimerrsquos disease neuronsrdquo Journal of Alzheimerrsquos Diseasevol 20 S2 pp S609ndashS631 2010
[7] H Akiyama T Arai H Kondo E Tanno C Haga andK Ikeda ldquoCell mediators of inflammation in the Alzheimerdisease brainrdquo Alzheimer Disease and Associated Disorders vol14 supplement 1 pp S47ndashS53 2000
[8] E G McGeer and P L McGeer ldquoThe importance of inflamma-torymechanisms inAlzheimer diseaserdquo Experimental Gerontol-ogy vol 33 no 5 pp 371ndash378 1998
[9] E M Blalock K-C Chen K Sharrow et al ldquoGene microar-rays in hippocampal aging statistical profiling identifies novelprocesses correlated with cognitive impairmentrdquo Journal ofNeuroscience vol 23 no 9 pp 3807ndash3819 2003
[10] C A Dickey J F Loring J Montgomery M N Gordon PS Eastman and D Morgan ldquoSelectively reduced expression of
synaptic plasticity-related genes in amyloid precursor proteinplus presenilin-1 transgenic micerdquo Journal of Neuroscience vol23 no 12 pp 5219ndash5226 2003
[11] S Rosi ldquoNeuroinflammation and the plasticity-related imme-diate-early gene Arcrdquo Brain Behavior and Immunity vol 25no 1 pp S39ndashS49 2011
[12] D F Swaab andA-M Bao ldquo(Re-)activation of neurons in agingand dementia lessons from the hypothalamusrdquo ExperimentalGerontology vol 46 no 2-3 pp 178ndash184 2011
[13] G K Gouras D Tampellini R H Takahashi and E Capetillo-Zarate ldquoIntraneuronal 120573-amyloid accumulation and synapsepathology in Alzheimerrsquos diseaserdquo Acta Neuropathologica vol119 no 5 pp 523ndash541 2010
[14] J F Guzowski G L Lyford G D Stevenson et al ldquoInhibition ofactivity-dependent arc protein expression in the rat hippocam-pus impairs the maintenance of long-term potentiation and theconsolidation of long-term memoryrdquo Journal of Neurosciencevol 20 no 11 pp 3993ndash4001 2000
[15] J J RodriguezH ADavies A T Silva et al ldquoLong-termpoten-tiation in the rat dentate gyrus is associated with enhancedArcArg31 protein expression in spines dendrites and gliardquoEuropean Journal of Neuroscience vol 21 no 9 pp 2384ndash23962005
[16] O Steward and PWorley ldquoLocal synthesis of proteins at synap-tic sites on dendrites role in synaptic plasticity and memoryconsolidationrdquo Neurobiology of Learning and Memory vol 78no 3 pp 508ndash527 2002
[17] C R Bramham and E Messaoudi ldquoBDNF function in adultsynaptic plasticity the synaptic consolidation hypothesisrdquoProgress in Neurobiology vol 76 no 2 pp 99ndash125 2005
[18] S-W Ying M Futter K Rosenblum et al ldquoBrain-derivedneurotrophic factor induces long-term potentiation in intactadult hippocampus requirement for ERK activation coupledto CREB and upregulation of Arc synthesisrdquo Journal of Neuro-science vol 22 no 5 pp 1532ndash1540 2002
[19] J F Guzowski B LMcNaughton C A Barnes and P FWorleyldquoEnvironment-specific expression of the immediate-early geneArc in hippocampal neuronal ensemblesrdquoNature Neurosciencevol 2 no 12 pp 1120ndash1124 1999
[20] E Messaoudi S-W Ying T Kanhema S D Croll andC R Bramham ldquoBrain-derived neurotrophic factor triggerstranscription-dependent late phase long-term potentiation invivordquo Journal of Neuroscience vol 22 no 17 pp 7453ndash74612002
[21] D-CWang S-S Chen Y-C Lee andT-J Chen ldquoAmyloid-120573 atsublethal level impairs BDNF-induced arc expression in corticalneuronsrdquoNeuroscience Letters vol 398 no 1-2 pp 78ndash82 2006
[22] M V Chao R Rajagopal and F S Lee ldquoNeurotrophin sig-nalling in health and diseaserdquo Clinical Science vol 110 no 2pp 167ndash173 2006
[23] H Akiyama ldquoInflammation and Alzheimerrsquos diseaserdquo Neurobi-ology of Aging vol 21 no 3 pp 383ndash421 2000
[24] M T Heneka M K OrsquoBanion D Terwel and M P KummerldquoNeuroinflammatory processes in Alzheimerrsquos diseaserdquo Journalof Neural Transmission vol 117 no 8 pp 919ndash947 2010
[25] P L McGeer J Rogers and E G McGeer ldquoInflammation anti-inflammatory agents and Alzheimer disease the last 12 yearsrdquoJournal of Alzheimerrsquos Disease vol 9 no 3 pp 271ndash276 2006
[26] L Tong G A Prieto E A Kramar et al ldquoBrain-derivedneurotrophic factor-dependent synaptic plasticity is suppressedby interleukin-1120573 via p38 mitogen-activated protein kinaserdquoJournal of Neuroscience vol 32 no 49 pp 17714ndash17724 2012
12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

12 Oxidative Medicine and Cellular Longevity
[27] V S Bankova S L de Castro and M C Marcucci ldquoPropolisrecent advances in chemistry and plant originrdquo Apidologie vol31 no 1 pp 3ndash15 2000
[28] MMarcucci ldquoPropolis chemical composition biological prop-erties and therapeutic activityrdquoApidologie vol 26 no 2 pp 83ndash99 1995
[29] J-H Yoon K Youn C-T Ho M V Karwe W-S Jeongand M Jun ldquop-coumaric acid and ursolic acid from Cornifructus attenuated 120573-amyloid
25minusminus35-induced toxicity through
regulation of the NF-120581B signaling pathway in PC12 cellsrdquoJournal of Agricultural and Food Chemistry vol 62 no 21 pp4911ndash4916 2014
[30] M O Villareal K Sasaki D Margout et al ldquoNeuroprotectiveeffect of Picholine virgin olive oil and its hydroxycinnamicacids component against 120573-amyloid-induced toxicity in SH-SY5Y neurotypic cellsrdquo Cytotechnology vol 68 no 6 pp 2567ndash2578 2016
[31] Y Nakajima M Shimazawa S Mishima and H Hara ldquoNeu-roprotective effects of Brazilian green propolis and its mainconstituents against oxygen-glucose deprivation stress with agene-expression analysisrdquo Phytotherapy Research vol 23 no 10pp 1431ndash1438 2009
[32] Y Nakajima M Shimazawa S Mishima and H Hara ldquoWaterextract of propolis and its main constituents caffeoylquinicacid derivatives exert neuroprotective effects via antioxidantactionsrdquo Life Sciences vol 80 no 4 pp 370ndash377 2007
[33] Y Kano N Horie S Doi et al ldquoArtepillin C derived frompropolis induces neurite outgrowth in PC12m3 cells via ERKand p38 MAPK pathwaysrdquo Neurochemical Research vol 33 no9 pp 1795ndash1803 2008
[34] Y Liu Z Wu X Zhang et al ldquoLeptomeningeal cells transduceperipheral macrophages inflammatory signal to microglia inreponse to Porphyromonas gingivalis LPSrdquoMediators of Inflam-mation vol 2013 Article ID 407562 11 pages 2013
[35] Z Wu L Sun S Hashioka et al ldquoDifferential pathways forinterleukin-1120573 production activated by chromogranin A andamyloid 120573 in microgliardquo Neurobiology of Aging vol 34 no 12pp 2715ndash2725 2013
[36] D Uberti L Piccioni A Colzi D Bravi P L Canonico andMMemo ldquoPergolide protects SH-SY5Y cells against neurodegen-eration induced by H
2O2rdquo European Journal of Pharmacology
vol 434 no 1-2 pp 17ndash20 2002[37] ZWu A Zhu F Takayama et al ldquoBrazilian green propolis sup-
presses the hypoxia-induced neuroinflammatory responses byinhibiting NF-120581B activation in microgliardquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 906726 10 pages2013
[38] J Ni Z Wu C Peterts K Yamamoto H Qing and H Nakan-ishi ldquoThe critical role of proteolytic relay through cathepsins Band E in the phenotypic change of microgliamacrophagerdquoTheJournal of Neuroscience vol 35 no 36 pp 12488ndash12501 2015
[39] J Choi M C Sullards J A Olzmann et al ldquoOxidative damageof DJ-1 is linked to sporadic Parkinson andAlzheimer diseasesrdquoJournal of Biological Chemistry vol 281 no 16 pp 10816ndash108242006
[40] E W Englander Z Hu A Sharma H-M Lee Z-H Wu andG H Greeley ldquoRat MYH a glycosylase for repair of oxidativelydamaged DNA has brain-specific isoforms that localize toneuronal mitochondriardquo Journal of Neurochemistry vol 83 no6 pp 1471ndash1480 2002
[41] C A Dickey M N Gordon J E Mason et al ldquoAmyloidsuppresses induction of genes critical formemory consolidation
in APP + PS1 transgenic micerdquo Journal of Neurochemistry vol88 no 2 pp 434ndash442 2004
[42] J J Palop J Chin N Bien-Ly et al ldquoVulnerability of dentategranule cells to disruption of Arc expression in human amyloidprecursor protein transgenicmicerdquoThe Journal of Neurosciencevol 25 no 42 pp 9686ndash9693 2005
[43] T-J Chen D-C Wang and S-S Chen ldquoAmyloid-120573 interruptsthe PI3K-Akt-mTOR signaling pathway that could be involvedin brain-derived neurotrophic factor-inducedArc expression inrat cortical neuronsrdquo Journal of Neuroscience Research vol 87no 10 pp 2297ndash2307 2009
[44] V Echeverria D E Berman and O Arancio ldquoOligomers of120573-amyloid peptide inhibit BDNF-induced arc expression incultured cortical neuronsrdquo Current Alzheimer Research vol 4no 5 pp 518ndash521 2007
[45] B Monti C Berteotti and A Constestabile ldquoDysregulation ofmemory-related proteins in the hippocampus of aged rats andtheir relation with cognitive impairmentrdquoHippocampus vol 15no 8 pp 1041ndash1049 2005
[46] D J Selkoe and D Schenk ldquoAlzheimerrsquos disease molecularunderstanding predicts amyloid-based therapeuticsrdquo AnnualReview of Pharmacology and Toxicology vol 43 pp 545ndash5842003
[47] H Nakanishi and Z Wu ldquoMicroglia-aging roles of microgliallysosome- andmitochondria-derived reactive oxygen species inbrain agingrdquo Behavioural Brain Research vol 201 no 1 pp 1ndash72009
[48] Z Wu and H Nakanishi ldquoConnection between periodontitisand Alzheimerrsquos disease possible roles of microglia and lep-tomeningeal cellsrdquo Journal of Pharmacological Sciences vol 126no 1 pp 8ndash13 2014
[49] C R Pugh M Fleshner L R Watkins S F Maier and J WRudy ldquoThe immune system and memory consolidation a rolefor the cytokine IL-1120573rdquoNeuroscience and Biobehavioral Reviewsvol 25 no 1 pp 29ndash41 2001
[50] A M Hein M R Stasko S B Matousek et al ldquoSustained hip-pocampal IL-1120573 overexpression impairs contextual and spatialmemory in transgenic micerdquo Brain Behavior and Immunityvol 24 no 2 pp 243ndash253 2010
[51] Y Yin G M Edelman and P W Vanderklish ldquoThe brain-derived neurotrophic factor enhances synthesis of Arc insynaptoneurosomesrdquo Proceedings of the National Academy ofSciences of the United States of America vol 99 no 4 pp 2368ndash2373 2002
[52] S Peng J Wuu E J Mufson and M Fahnestock ldquoPrecursorform of brain-derived neurotrophic factor and mature brain-derived neurotrophic factor are decreased in the pre-clinicalstages of Alzheimerrsquos diseaserdquo Journal of Neurochemistry vol93 no 6 pp 1412ndash1421 2005
[53] S Chamniansawat and S Chongthammakun ldquoEstrogen stim-ulates activity-regulated cytoskeleton associated protein (Arc)expression via the MAPK- and PI-3K-dependent pathways inSH-SY5Y cellsrdquoNeuroscience Letters vol 452 no 2 pp 130ndash1352009
[54] J Kremerskothen D Wendholt I Teber and A BarnekowldquoInsulin-induced expression of the activity-regulated cytos-keletonassociated gene (ARC) in human neuroblastoma cellsrequires p2l(ras) mitogen-activated protein kinaseextracellu-lar regulated kinase and src tyrosine kinases but is protein kin-aseC-independentrdquoNeuroscience Letters vol 321 no 3 pp 153ndash156 2002
[55] V Ramırez-Amaya A Vazdarjanova D Mikhael S Rosi PF Worley and C A Barnes ldquoSpatial exploration-induced Arc
Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003

Oxidative Medicine and Cellular Longevity 13
mRNA and protein expression evidence for selective network-specific reactivationrdquo Journal of Neuroscience vol 25 no 7 pp1761ndash1768 2005
[56] M Cammalleri R Lutjens F Berton et al ldquoTime-restrictedrole for dendritic activation of the mTOR-p70S6K pathway inthe induction of late-phase long-term potentiation in the CA1rdquoProceedings of the National Academy of Sciences of the UnitedStates of America vol 100 no 2 pp 14368ndash14373 2003
[57] J M Horwood F Dufour S Laroche and S Davis ldquoSignallingmechanisms mediated by the phosphoinositide 3-kinaseAktcascade in synaptic plasticity and memory in the ratrdquo EuropeanJournal of Neuroscience vol 23 no 12 pp 3375ndash3384 2006
[58] P Opazo A M Watabe S G N Grant and T J OrsquoDellldquoPhosphatidylinositol 3-kinase regulates the induction of long-term potentiation through extracellular signal-related kinase-independent mechanismsrdquoThe Journal of Neuroscience vol 23no 9 pp 3679ndash3688 2003





























