Embed Size (px)
Citation preview

The Osteology of the Congrid Eel Gorgasia punctata and the
Relationships of the Heterocongrinae'
RICHARD H. ROSENBLATT2
ABSTRACT : Th e osteology of Gorgasia punctata is described, figured , and compared with that of other congrids . Gorgasia is clearly referable to the subfamilyHeterocongrinae. The heterocongrin es agree with the Congridae in several importantfeatu res, and do not differ in fund amental respects. Th erefore, the group is recognized as a subfamily of the Congridae. Gorgasia is the most primit ive heterocongrine, and agrees with the anagoine congrids in having a lateral ethmoid process.Because of this and other similarities it is suggested that the Anagoinae andHete rocongrin ae arose from a common stem. The genus X arifania was erected onthe erroneous basis of lack of caudal rays. It is synonymized with Tuenioconger,
THE CONGRID EEL Gorgasia ptl11ctata was placedby its describers in the little known apodalfamily Derichthyidae (Meek and H ildebrand,1923) . Bohlke (1951) was the first to pointout that the affinities of Gorgasia were withH eteroconger. Gosline (1952) provisionallyplaced H eteroconger and Gorgasia in the Congridae. Bohlke ( 1957) described the osteologyof the related N ystactichthys halis and placedthe eels allied to Heteroconger in the Congridae,but considered them to constitute the distinctsubfamily Heterocongrinae.
Bohlke considered Gorgasia to be the mostprimitive genus of the Heterocongrin ae on thebasis of its more complete complement of headpores, its uncoalesced upper labial flanges(called by him the "free edge of lip"), a welldeveloped pectoral fin, and its unspecializedmaxillary dentiti on. He considered Gorgasia tobe specialized, however, in that the caudal raysare much reduced and covered by thick skin.Internal characters were not considered, sincethe only complete specimen then available wasthe holotype.
Recent collections made by personnel of theScripps Institution of Oceanography haveamassed rich material of several species of
1 Contribution from the Scripps Institution ofOceanography, University of California, San Diego,and the Institute of Fisheries, University of BritishColumbia. Manuscript received December 22, 1965.
2 Scripps Institution of Oceanography, Universityof California , San Diego, La Jolla, California .
eastern Pacific heterocongrines , including Gorgasia punctata. Because of previous uncertaintiesregarding the exact position of Gorgasia in eelclassification, a study of the osteology of thisspecies seemed worthwhile.
ACKNOWLEDGMENTS
The figures were drawn under my supervisionby E. David Lane. Part of the cost of thisinvestigation was defrayed by a grant to theInstitute of Fisheries, University of BritishColumbia, from the H . R. McMill an Expeditionary Fund . Th e specimen of Tnenioconger hassiused in this study was collected as a part of theU. S. Biological Program, Ind ian Ocean Expedition.
MATERIALS AND METHODS
The two adults of Gorgasi« pnnctat« weretaken from a series of 69 (SI062-720-26A,Bahia Magdalena, Baja Californ ia, Mexico) .Th ese were bone-stained with alizarin andcleared in glycerine. The neurocranium was dissected out of one specimen, and the drawingswere made from the dried preparation . Inaddition I have utilized single-stained andcleared specimens of Taeniocongel' diglletiChabanaud (SI065-278, Gulf of Californ ia) ,T. berrei W ade (51061-261, Gulf of California), T . hassi3 (Klausewitz and Eibl-Eibesfeldt )
3 Taenioconger hassi was originally described inthe genus X arifania, of which it is the type species
91

92 PACIFIC SCIENCE, Vol. XXI, January 1967
K)p SR ""O-",5,,,,m,,--~~_
o.e crn
~" "~~, '
• ~ C --::r- EO
PAS PTS $PO PRO BO
. o.s cm
MX PAS PP
o
c
B
A
SUSPENSORIUM AND JAWS (F ig. 1): As
but in another they are fused for the anteriorone-quarter of their lengths . Th e frontals arecompletely fused, with no sign of a suture ormedian ridge. There are well-developed canalsalong the lateral margins of the pterotics andfrontals, with two large foramina anteriorly,but there is no transverse canal across thefrontals.
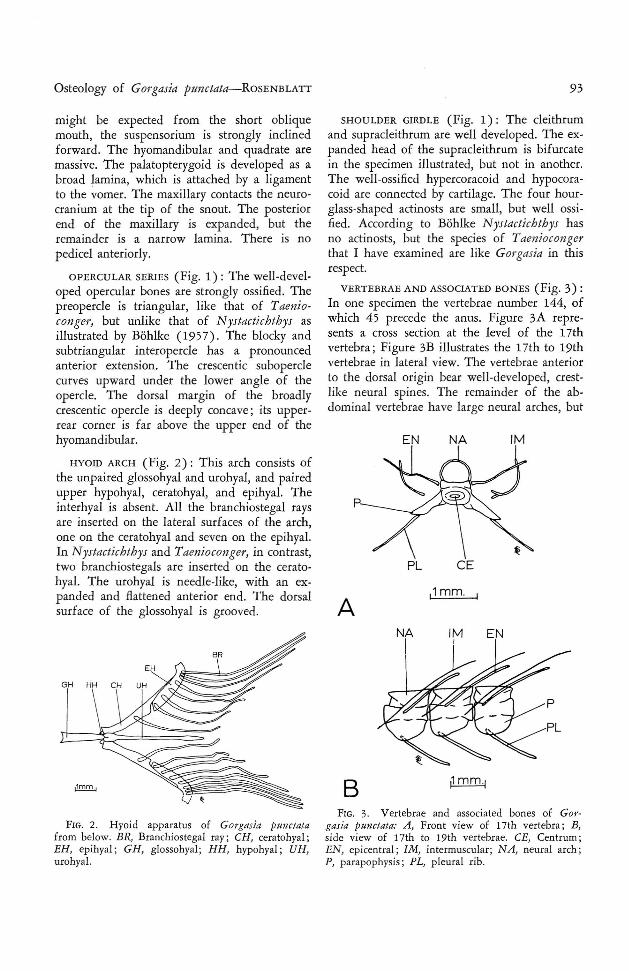
FIG. 1. H ead skeleton of Gorgasia punctata. A ,Lateral view, includin g pectoral girdle ; B, neu rocranium, lateral view ; C, neurocranium, top view ; D,neurocranium, bottom view. A A , Articular angular;BO, basioccipi tal; BR, branchiostegal ray; CL, cleith rum ; CO, circumorbital; D, dentary; EO, exoccipital ;EP, epiotic ; FR, frontal; H MD; hyomandibular ;H OC, hypocoracoid ; H RC, hypercoracoid ; l OP, interopercle ; LEP, lateral ethmoid process; MX, maxillary; N, nasal ; OBS, orbitosphenoid; OP, opercle ;PAR, par ietal; PAS, parasphen oid; PEV, premaxillary ethm ovomerine block ; POP, preopercle; PP,palatopt erygoid ; PR, pectoral ray; PRO, proo tic ;PT, pterygiophore; PT O, p terot ic ; PTS, pterosp henoid ; QU , quadrate; SCL, supracleithrum; S0 , supraoccipital ; SOP, subopercle ; SPO, sphen otic.
N E URO CRANIUM (Fi g. 1) : The skull istruncated posteriorly, except where the exoccipital flanges break the outl ine. The pre maxillaries, ethmoid, and vomer are fused, withno suggestion of articulations, such as werereported by Bohlke (1 957) for N ystactichthyshalis. The anterior, triangular tooth -patch mayrepresent the premaxillary dent ition. The dorsalor ethmoid portion of the complex is very thin,and is reduced medially to a septum, so thatit is shaped much like an I-beam in cross section.Laterally the ethmoid portion is expanded, andgives rise to two heavy, forward-curving processes. These are very similar to the "lateralethmoid processes" reported by Asano (1962)for A nago and Alloconger. Below this are twosmall proj ections from the lateral face of thevomer. These vomerine processes are difficultto distinguish from the base of the lateralethmoid process. They are more evident inTnenioconger and, judging from Bohlke'sfigures 3B and 3C, are developed in Nystactichthys as well. Posteriorly on the under side of thecranium, parts of the prootic and basioccipitalare expanded to form a prominent auditorybulla, which contains a large otolith (p resumably the sagitta). The foramen magnum issurrounded by exoccipital flanges, which graspthe first vertebra. Th e supraoccipital is welldeveloped, but does not completely separate theepiotics, which are in contact posteriorly. Theparietals are sutured in the specimen figured,
(unnumbered, D 'Arros Island , Amir antes Is- CL
lands) , T. n.sp. (SI062-42, Bahia Banderas,Mexico) , and A riosoma gilberti (Ogilby)(SI062-77, Sinaloa, Mexico).
OSTEOLOGY
(Klausewitz and Eibl-Eibesfeldt, 1959 ) . The soledis tinctio n of the genus X arijan ia was the supposedlack of cauda l rays. Th e tail -tip of X. hassi is fleshybut flexible and clearly contains well-developed cauda lrays. These are visible und er direct light and are obvious when transmitt ed light is used. In addi tion tothe Arnirantes specimen, I have examined a para typeof X . hassi (ANSP 94706) through the courtesy of]. Boh lke. I can lind no other important differences,either in external morphology or osteology, betweenX . hassi and the species of T'aeniocanger examin ed.Th e nominal genus X arijania is consider ed, therefore,to be a synonym of Taenioconger .
Osteology of Gorgasia p1t11ctata- RoSENBLATT 93
FIG. 3. Vertebrae and associated bones of G Of
gasia punctata: A, Front view of 17th vertebra; B,side view of 17th to 19th vertebrae. CE, Centr um;EN, epicentral ; 1M , intermuscular; NA, neural arch;P, parapoph ysis; PL, pleura l r ib.
SHOULDER GIRDLE (Fig. 1): The cleithrumand supracleithrum are well developed. The expanded head of the supracleithrum is bifurcatein the specimen illustrated, but not in another.The well-ossified hypercoracoid and hypocoracoid are connected by cartilage. Th e four hourglass-shaped actinosts are small, but well ossified. According to Bohlke N ystactichthys hasno actinosts, but the species of Taeniocongerthat I have examined are like Gorgasin in thisrespect.
VERTEBRAE AND ASSOCIATED BONES (Fig. 3) :In one specimen the vertebrae number 144, ofwhich 45 precede the anus. Figure 3A represents a cross section at the level of the 17thvertebra ; Figure 3B illustrates the 17th to 19thvertebrae in lateral view. The vertebrae anteriorto the dorsal origin bear well-developed, crestlike neural spines. The remainder of the abdominal vertebrae have large neural arches, but
P
PL
PL CE
,1mm. I
ANA 1M ENI I
B
GH
might be expected from the short obliquemouth, the suspensorium is strongly inclinedforward. Th e hyomandibular and quadrate aremassive. The palatopterygoid is developed as abroad lamina, which is attached by a ligamentto the vomer. The maxillary contacts the neurocranium at the tip of the snout. The posteriorend of the maxillary is expanded, but theremainder is a narrow lamina. There is nopedicel anteriorly.
OPERCU LAR SERIES ( Fig. 1) : Th e well-developed opercular bones are strongly ossified. Thepreopercle is triangular, like that of Taenioconger, but unlike that of N ystactichthys asillustrated by Bohlke (1957). The blocky andsubtriangular interopercle has a pronouncedanterior extension. The crescentic suboperclecurves upward under the lower angle of theopercle. The dorsal margin of the broadlycrescentic opercle is deeply concave; its upperrear corner is far above the upper end of thehyomandibular.
HY OID ARCH (Fig. 2): Thi s arch consists ofthe unpaired glossohyal and urohyal, and pairedupper hypohyal, ceratohyal, and epihyal. Th einterhyal is absent. All the branchiostegal raysare inserted on the lateral surfaces of the arch,one on the ceratohyal and seven on the epihyal.In N ystactichthys and Tnenioconger, in contrast,two branchiostegals are inserted on the ceratehyal. The urohyal is needle-like, with an expanded and flattened anterior end. Th e dorsalsurface of the glossohyal is grooved.
FIG. 2. H yoid app aratus of Gorgasia punctatafrom below. BR, Branchios tegal ray ; CH, ceratohyal;EH, epihyal ; GH , glossohyal; HH, hypohyal ; UH ,urohyal.
94
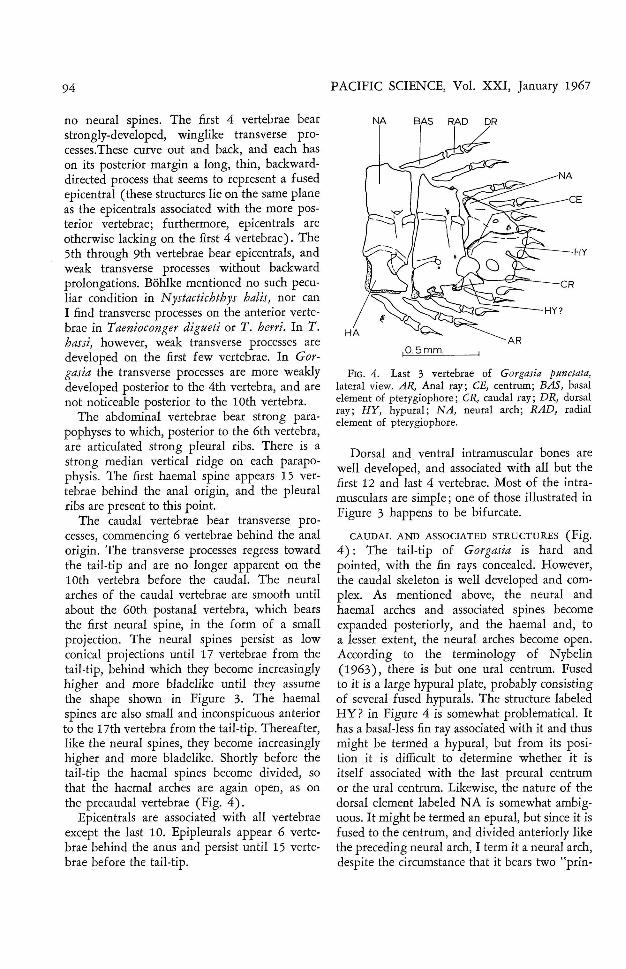
no neural spines. The first 4 vertebrae bearstrongly-developed, winglike transverse processes.These curve out and back, and each hason its posterior margin a long, thin, backwarddirected process that seems to represent a fusedepicentral (these structures lie on the same planeas the epicentrals associated with the more posterior vertebrae; furthermore, epicentrals areotherwise lacking on the first 4 vertebrae). The5th through 9th vertebrae bear epicentrals, andweak transverse processes without backwardprolongations. Bohlke mentioned no such peculiar condition in N ystactichthys balis, nor canI find transverse processes on the anterior vertebrae in Tuenioconger digueti or T. berri . In T.hassi, however, weak transverse processes aredeveloped on the first few vertebrae. In Gorgasia the transverse processes are more weaklydeveloped posterior to the 4th vertebra, and arenot noticeable posterior to the 10th vertebra.
The abdominal vertebrae bear strong parapophyses to which, posterior to the 6th vertebra,are articulated strong pleural ribs. There is astrong median vertical ridge on each parapophysis. The first haemal spine appears 15 vertebrae behind the anal origin, and the pleuralribs are present to this point.
The caudal vertebrae bear transverse processes, commencing 6 vertebrae behind the analorigin. The transverse processes regress towardthe tail-tip and are no longer apparent on the10th vertebra before the caudal. The neuralarches of the caudal vertebrae are smooth untilabout the 60th postanal vertebra, which bearsthe first neural spine, in the form of a smallprojection. The neural spines persist as lowconical projections until 17 vertebrae from thetail-tip, behind which they become increasinglyhigher and more bladelike until they assumethe shape shown in Figure 3. The haemalspines are also small and inconspicuous anteriorto the 17th vertebra from the tail-tip. Thereafter,like the neural spines, they become increasinglyhigher and more bladelike. Shortly before thetail-tip the haemal spines become divided, sothat the haemal arches are again open, as onthe precaudal vertebrae (Fig. 4).
Epicentrals are associated with all vertebraeexcept the last 10. Epipleurals appear 6 vertebrae behind the anus and persist until 15 vertebrae before the tail-tip .
PACIFIC SCIENCE, Vol. XXI, January 1967
NA
NA
CE
rr---HY
HY?
HA
FIG . 4. Last 3 vertebrae of Gorgasia punctata,lateral view. AR, Anal ray ; CE, centrum ; BAS, basalelement of pterygiophore ; CR, caudal ray; DR, dorsalray; HY, hypural; NA, neural arch ; RAD, radialelement of pterygiophore.
Dorsal and ventral intramuscular bones arewell developed, and associated with all but thefirst 12 and last 4 vertebrae. Most of the intramusculars are simple; one of those illustrated inFigure 3 happens to be bifurcate.
CAUDAL AND ASSOCIATED STRUCTURES (Fig.4): The tail-tip of Gorgasia is hard andpointed, with the fin rays concealed. However,the caudal skeleton is well developed and complex. As mentioned above, the neural andhaemal arches and associated spines becomeexpanded posteriorly, and the haemal and, toa lesser extent, the neural arches become open.According to the terminology of Nybelin(1963), there is but one ural centrum. Fusedto it is a large hypural plate, probably consistingof several fused hypurals. The structure labeledHY? in Figure 4 is somewhat problematical. Ithas a basal-less fin ray associated with it and thusmight be termed a hypural, but from its position it is difficult to determine whether it isitself associated with the last preural centrumor the ural centrum. Likewise, the nature of thedorsal element labeled NA is somewhat ambiguous. It might be termed an epural, but since it isfused to the centrum, and divided anteriorly likethe preceding neural arch, I term it a neural arch,despite the circumstance that it bears two "prin-

Osteology of G orgasia pm1etata-RoSE NBLATT
cipal" caudal rays. A strong process curvesforwar d and downward from the neural archelement. It probably serves as a muscle attachment and may be associated with tail-first digging in sand. An essentially similar caudalskeleton has been figured by Bohlke for N ystaetiehth ys balis, and I have found th e caudalskeleton of Taenio conger digtteti, T. berrei, T.hassi, and T. n.sp. to be basically the same.
CIRCUMORBITALS AN D LATERAL LINE CA
N ALS: The circumorbital ring is complete, consisting of at least five weakly-ossified and roofless bones. Apparently the small "supraorbital"illustrated by Asano (1 962) for several Japanese congr ids is absent. The temporal canal isencased in bone in G. pm1etata only, amongthe species examined. The lateral-line canalalong the body is contained in a series of ossicles (l ateral-line scales?) . Ant eriorly these aredeveloped as unconnected but closely opposedtubes, which posteriorly gradually become lessstrongly ossified, so that along the midbodythere is an open trough consisting of a seriesof ossified half-rings. In the species of Taenio conger examined, the lateral-line ossicles aredeveloped as short, widely-spaced, troughlikeossifications.
RELATIO NSHIPS OF THE H ETE ROCONGRINAE
The heterocongrines resemble the Ophichthidae in several respects. In both groups the ribsare laminar, and the neural spines reduced(vestigial in the ophichth ids) , as are the circumorbita ls. In Gorgasia and in some speciesof Taenioconger the caudal fin is shor t and thetail-tip fleshy. In all, the body is elongate andcircular in cross section. The pectoral fin, as inmany ophichthids, is reduced (varying fromsmall in Gorgasia to minu te in Nystaetiehthysand Tuenioconger to absent in H eteroeonger) .In addition to these structural characters, bothgroups share the sand-dwelling habit. However,the characters given by Gosline (1 951) tosepara te the Congridae and the Ophi chth idae(except that it is now known that many congrids have an auditory bulla) serve to distinguish Gorgasia and its allies from the ophichthids as well. In addition, it may be noted thatthe Congri dae have the parapophyses dividedby a vertical ridge and have a vomerine process,
95
to which the palatopterygoid is ligamentouslyattached-features that appear to be lacking inthe Ophichthidae.
The superficial similarities between Gorgasiaand the ophichthids are certainly para llel adaptations to a similar mode of life, and the minorosteological similarities may be adaptations aswell.
Alth ough recognizing the close relationshipbetween the two groups, Klausewitz and EiblEibesfeldt (19 59) maintain ed the family Heterocongridae as distinct from the Congridae.Their action was based on behavioral differences and on bone reduction and "Fensterbildung" (fenestration ?) in the skeleton. However, their illustration of the head skeleton ofXarifania h. hassi shows a well-developed skulland well-integrated jaws, suspensorium, andopercular series. On the basis of the presentinvestiga tion there are no grounds (except thatthe circumorbital series is less well developed )for the contention that the skeleton has undergone important reduction in comparison withthat of the Congridae.
There is, in fact, nothing in the osteology ofthe heterocongrines I have examined that wouldpreclude the inclusion of the group in the Congr idae. The heterocongrines possess the basiccongri d characters of ankylosed frontals, forward-inclined suspensorium, few and non overlapping branchiostegals, maxillary-ethmoid articulation near tip of snout, caudal vertebraewith transverse processes, skull truncate posteriorly, parapophyses divided by a vertical ridge,and a lateral process on the vomer.
The chief osteological differences are: neuralspines absent on most abdominal vertebrae (andon most caudal vertebrae as well in Gorgasia ) ;neural and haemal arches becoming high andbladelike near the tail-tip; urostylar vertebrabetter developed, and supporting structure ofcaudal more complex; epineurals and epip leurals lost 10- 15 vertebrae before tail-tip ; circumorbital series reduced and less ossified.Stronger divergences from the basic congridtype are found in nonosteological characters.In most of the Congr idae the muzzle is elongate, and the olfactory organ is well developed,with numerous lamellae. In the heterocongrinesthe muzzle is short, the eye is relatively large,and the olfactory rosette is much smaller, with

96
few lamellae (ca. 20) . Th is distinction is nodoubt correlated with a change in food habits(Klausewitz and Eibl-Eibesfeldt, 1959). Alsothe habit of living colonially in sand tubes isunknown in other congrids.
These differences and similarities seem tobear out Bohlke's (1957) contention that theHeterocongrinae should be regarded as a welldefined subfamily within the Congridae. Thatthere are profo und differences in behavior andecology is undoubted , but these have not involved any fundamental changes in the basiccongrid body plan.
Until recently, little information has beenavailable on the osteology of the family Congridae. However, Asano (1962) has presenteddetailed information on the anatomy of 10genera and 14 species of Japanese congrids.On the basis of his study, Asano recognizedtwo subfamilies, the Anagoinae and the Congrinae (the Heterocongrinae were not considered) . The Anagoinae were said to differ fromthe Congrinae in that there is a forward andlaterally directed process on the ethmoid, thesupraoccipital is absent, there are only foursuborbitals, the abdominal and caudal vertebraeare about equal in number, the gas bladder isattached to the parapophyses, the tail-tip is hard,the caudal rays are short, the fin rays are unsegmented, and the lateral-line scales are welldeveloped.
Asano assigned two genera, A nago and AUoconger, to the Anagoinae. I can confirm thatA riosoma belongs here, as does the recentlydescribed Paraconger Kanazawa 1961.
The heterocongrines share characters withboth the Anagoinae and the Congrinae. Theyagree with the congrines in that the supraoccipital is present, there are many more caudalthan abdominal vertebrae, and the gas bladderis free from the parapophyses. They agree withthe anagoines in that the fin rays are unsegmented, the caudal is reduced, and the lateralline scales are well ossified (corr esponding toAsano's "Allago type") . I have been unable todetermine with certainty the number of suborbitals in the heterocongrines.
Gorgasia alone agrees with the Anagoinaein having a lateral ethmoid process. In thisconnection it is important to establish the evolutionary position of Gorgasia. Bohlke (1957)
PACIFIC SCIENCE, Vol. XXI, January 1967
gave reasons for considering Gorgasia to be inmost respects the most prim itive of the Heterocongrinae. H is conclusions are borne out in thisstudy, except for the discovery in Gorgasi« ofpeculiar, expanded transverse processes on theanterior vertebrae, and the loss of an anteriormaxillary pedicel. These specializations probably preclude Gorgasia as an ancestor, but theydo not militate against the hypoth esis thatGorgasia is more generalized over-all, and prob ably was an earlier offshoot of the heterocongrine line.
The retention in Gorgasia of a lateral ethmoidprocess indicates relationship with the anagoineline. It seems unlikely that the agreement represents convergence. Eels have evolved a number of structures bracing the maxillary, correlated with elongation of the gape and with theuse of the jaws in biting and crushing (Gos line, 1951; Asano, 1962). However, the trendin heterocongrine evolution has been in theother direction, toward shortening of the gapeand development of a jaw structure and dentition suitable for snapping at planktonic prey.It may be that the retention of the lateral ethmoid process in Gorgasia has allowed the lossof the maxillary pedicel.
It seems plausible to hypothesize that theHeterocongrinae and Anagoinae arose from acommon ancestor which had a lateral ethmoidprocess, a supraoccipital, unsegmented fin rays,and well-developed lateral-line scales. It seemslikely that the sand-burrowing habit (knownfor Anago) had already been developed. Thetwo groups have diverged sharply, however.The development of the plankton-feeding habitin the heterocongrines has been accompaniedby important changes in the head. The mouthhas become short and oblique, and the dentition specialized. The development of a shortoblique mouth as an adaptation to snapping atplankton or small prey has taken place in anumber of fishes. Compare, for example, theserranid genus Epinepbelus, which feeds onrelatively large prey, with the plankton-feedingParantbias. A similar phenomenon can be seenif the bottom-feeding embiotocid genus Micrometres is compared with the closely relatedgenus Bracbyistius, which feeds in midwater(Hubbs and Hubbs, 1954). Walter A. Starck IIhas pointed out to me that the shorten ing of the

Osteology of Gorgasia punctata- RoSENBLATT
muzzle in these fishes results in the placementof the eye close to the tip of the snout, and thusallows for close-up binocular vision. Thus ,vision has become more important in prey finding in heterocongrines, and the eye is muchenlarged and the olfactory organ much reduced. The lateral-line system on the head haslikewise become reduced, again probably correlated with the increased dependence on vision.The great elongation of the slender body wouldseem to be an adaptation to getting the headwell off the bottom, and yet maintaining contact with the sand tube which is used for cover.(The normal posture of a heterocongrine isvertical, with the anterior one-half to two-thirdsof the body out of the sand tube.)
On the other hand, the anagoines, except inthe loss of the supraoccipital, have divergedmuch less from the basic congrid type, either instructure or in behavior.
REFERENCES
ASANo, H. 1962. Studies on the congrid eelsof Japan. Bull. Misaki Mar. BioI. Inst. 1 :1143.
BOHLKE, J. 1951. A new eel of the genus Taenioconger from the Philippines. Copeia1951 (1) :32-35 .
97
- - - 1957. On the occurrence of garden eelsin the western Atlantic, with a synopsis ofthe Heterocongrinae. Proe. Acad. Nat. Sci.Philadelphia 109 :59-79.
GOSLINE, W . A. 1951. The osteology and classification of the ophichthid eels of the Hawaiian Islands. Pacific Sci. 5(4) :298- 320.
--- 1952. Notes on the systematic statusof four eel families. J. W ashington Acad.Sci. 42 (4) :130-135.
HUBBS, C. L., and L. C. HUBBS. 1954. Data onthe life history, variation, ecology and relationships of the Kelpperch, Brachyistius frenatus, an embiotocid fish of the Californias.Calif. Fish and Game 40(2) :183- 198.
KANAZAWA, R. 1961. Paraconger, a new genuswith three new species of eels (family Congridae) . Proe. U. S. N atl. Mus. 113(3450):1-14.
KLAUSEWITZ, W. , and I. EIBL-EIBESFELDT.1959. Neue Rohrenaale von den Maldivenund Nikob aren (Pisces, Apodes, Heterocongridae) . Senck. BioI. 40(3/4) :135- 153.
MEEK, S. E., and S. F. HILDEBRAND. 1923.The marine fishes of Panama, Pt. I. FieldMus. Nat. Hist. Zool. Ser, 15. 330 pp.
NYBELIN, O. 1963. Zur Morphologie und Terminologie des Schwanzskelettes der Actinopterygier. Ark. Zool. (2) 15 (35) :485-516.