Embed Size (px)
Citation preview

INTRODUCTION
The vertebral column is derived entirely from cells in theventral halves of the somites. In morphological terms theevents leading to axial skeleton formation have been exten-sively studied in several species, and are well described(Verbout, 1985; Christ and Wilting, 1992). The vertebralcolumn is the most conspicuously segmented structure of thevertebrate body. The metameric pattern is initially laid downduring the formation of somites, yet there is not a clear rela-tionship between the vertebral units and the initial somites.Upon somite differentiation the ventral parts de-epithelializeand form the mesenchymal sclerotomes, which give rise to pre-cartilaginous structures as well as connective tissue. Thisprocess is dependent on signals from the notochord (Wattersonet al., 1954; Pourquie et al., 1993). Some sclerotome cellsmigrate medially and form the perichordal tube, which isinitially unsegmented and uniform in density. Thereafter asegmented pattern of condensations starts to appear, the lateralregions slightly preceding the axial ones. The perichordal tube
gives rise to vertebral bodies and intervertebral discs whereasthe caudal halves of the paired lateral sclerotome areas are theorigins of neural arches, pedicles and ribs (Verbout, 1985;Christ and Wilting, 1992). The axial regions of highest celldensity, the intervertebral disc anlagen, are positioned approx-imately midsegmentally with respect to the former somiteboundaries. By cell labelling experiments it has been shownthat one vertebral body is derived from cells originating fromtwo adjacent somites (Bagnall, 1992; Ewan and Everett, 1992).The mechanisms that control these morphological events inaxial and lateral regions have not been studied, however.
The existence of a large number of mouse mutants withspecific malformations in vertebral column development(Grüneberg, 1963; Theiler, 1988) is an important resource forinvestigating the molecular and morphogenetic events that leadto the development of this structure. One of these mutants,
undulated (un; Wright, 1947), carries a point mutation in thePax-1 gene (Balling et al., 1988), which is transcriptionallyactivated in sclerotome cells at the time of differentiation ofthe epithelial somites (Deutsch et al., 1988). A detailed mor-
1109Development 120, 1109-1121 (1994)Printed in Great Britain © The Company of Biologists Limited 1994
Previous studies have identified a single amino-acid substi-tution in the transcriptional regulator Pax-1 as the causeof the mouse skeletal mutant
undulated (un). To evaluatethe role of Pax-1 in the formation of the axial skeleton wehave studied Pax-1 protein expression in early sclerotomecells and during subsequent embryonic development, andwe have characterized the phenotype of three differentPax-1 mouse mutants, un, undulated-extensive (unex) andUndulated short-tail (Uns). In the Uns mutation the wholePax-1 locus is deleted, resulting in the complete absence ofPax-1 protein in these mice. The other two genotypes areinterpreted as hypomorphs. We conclude that Pax-1 isnecessary for normal vertebral column formation along theentire axis, although the severity of the phenotype isstrongest in the lumbar region and the tail. Pax-1-deficientmice lack vertebral bodies and intervertebral discs. Theproximal part of the ribs and the rib homologues are alsomissing or severely malformed, whereas neural arches arenearly normal. Pax-1 is thus required for the developmentof the ventral parts of vertebrae. Embryonic analysesreveal that although sclerotomes are formed in mutant
embryos, abnormalities can be detected from day 10.5 p.conwards. The phenotypic analyses also suggest that thenotochord still influences vertebral body formation somedays after the sclerotomes are formed. Furthermore, thenotochord diameter is larger in mutant embryos from day12 p.c., due to increased cell proliferation. In the stronglyaffected genotypes the notochord persists as a rod-likestructure and the nucleus pulposus is never properlyformed. Since the notochord is Pax-1-negative thesefindings suggest a bidirectional interaction betweennotochord and paraxial mesoderm. The availability ofthese Pax-1 mutant alleles permitted us to define an earlyrole for Pax-1 in sclerotome patterning as well as a late rolein intervertebral disc development. Our observationssuggest that Pax-1 function is required for essential stepsin ventral sclerotome differentiation, i.e. for the transitionfrom the mesenchymal stage to the onset of chondrogene-sis.
Key words: Pax-1, vertebral column, sclerotome, notochord, mousemutants
SUMMARY
The role of
Pax-1 in axial skeleton development
Johan Wallin1, Jörg Wilting2, Haruhiko Koseki1, Rüdiger Fritsch3, Bodo Christ2 and Rudi Balling1,4,*1Department of Developmental Biology, Max-Planck Institute of Immunobiology, Stübeweg 51, D-79108 Freiburg, FRG2Department of Anatomy, University of Freiburg3Max-Planck Institute for Biophysical Chemistry, Göttingen4Institut für Säugetiergenetik, GSF-Forschungszentrum Neuherberg, FRG
*Author for correspondence at address4
1110
phological analysis of the skeletal phenotype and theembryonic development of this mutant has been made byGrüneberg (1950, 1954). He described malformations ofvertebral bodies and intervertebral discs as well as vertebralprocesses, and came to the conclusion that these result fromdecreased sizes of the mesenchymal condensations thatprecede chondrification. Two alleles of un, undulated-extensive (unex; Wallace, 1985) and Undulated short-tail (Uns;Blandova and Egorov, 1975), both being phenotypically moreaffected than un, have subsequently been isolated. Both thesemutants have Pax-1 gene deletions, the latter one removing theentire Pax-1 locus (Balling et al., 1992; Dietrich S., Gruss P.,and R.B., unpublished observations).
The Pax genes form a family of developmental control genesthat have recently been the focus of great interest (Chalepakiset al., 1992; Gruss and Walther, 1992; Noll, 1993). They encodesequence-specific transcription factors that contain the DNA-binding paired-domain (Bopp et al., 1986; Chalepakis et al.,1991; Adams et al., 1992; Zannini et al., 1992), and in additionsome of the Pax proteins also contain a paired-type home-odomain (Walther et al., 1991). In the mouse, nine Pax geneshave been identified to date (Walther et al., 1991; Wallin et al.,1993), all being expressed in a spatially restricted mannerbeginning during early organogenesis. In addition to Pax-1, twoother members of the Pax gene family have been shown to playimportant roles in mouse embryogenesis. Mutations in the Pax-3 and Pax-6 genes cause the Splotch and Small-eye phenotypes,respectively (Epstein et al., 1991; Hill et al., 1991).
To gain further insight into early mechanisms of vertebralcolumn formation we have carried out a comparative pheno-typic analysis of the three Pax-1 mouse mutants, un, unex andUns, of which the latter two have not been investigated before.By analysis of mutant embryonic and new-born mice we show
that Pax-1 is required for normal development of ventralvertebral structures, but is dispensable for sclerotomeformation per se.
MATERIALS AND METHODS
Miceundulated (un) mice were purchased from the Jackson laboratory.Undulated-short tail (Uns) and undulated-extensive (unex) mutantmice were kindly provided by Dr A. M. Malashenko, Krosnogorsk,Russia and Dr J. L. Cruickshank, Leeds, England, respectively. Thesemutants were backcrossed onto the C57BL/6 strain. On this geneticbackground homozygous un and unex mice show reduced fertility, par-ticularly unex. Homozygous unex mice are often born small, in whichcase they also remain smaller and die earlier than their littermates. N1to N5 generations were used for analysis. Embryos were recoveredon days 9.5-18.5 p.c. where day 0.5 was 12 a.m. on the day ofdetection of the vaginal plug. On average, litters were born on day 19p.c. As controls, +/+ or un/+ littermates were normally used. In excep-tional cases C57BL/6 embryos of the corresponding stages were used.
GenotypingGenotypings were made either by PCR analysis on embryonic DNAprepared from yolk-sacs or by Southern blotting of placental DNA.Genomic Southern blots were performed according to standard pro-cedures (Sambrook et al., 1989). For PCRs two different primer pairswere used: (1) with primers flanking the mutated HaeIII restrictionsite in un and (2) specific for the fifth exon deleted in unex. These Pax-1-specific oligonucleotides were: (1) 5
′-CAGAGCAGACGTACGGCGAAG-3′ and 5′-AGGCAAA-GATGCCAGGATCCC-3′ and for (2) 5′-AGAGCCATCAGCATGGTTTCG-3′ and 5′-TGGAGGG-AGTCCAGATTAAGC-3′. PCR reaction conditions were: 30 cyclesof denaturation for 1 minute at 94°C, reannealing for 1 minute at 60°C
J. Wallin and others
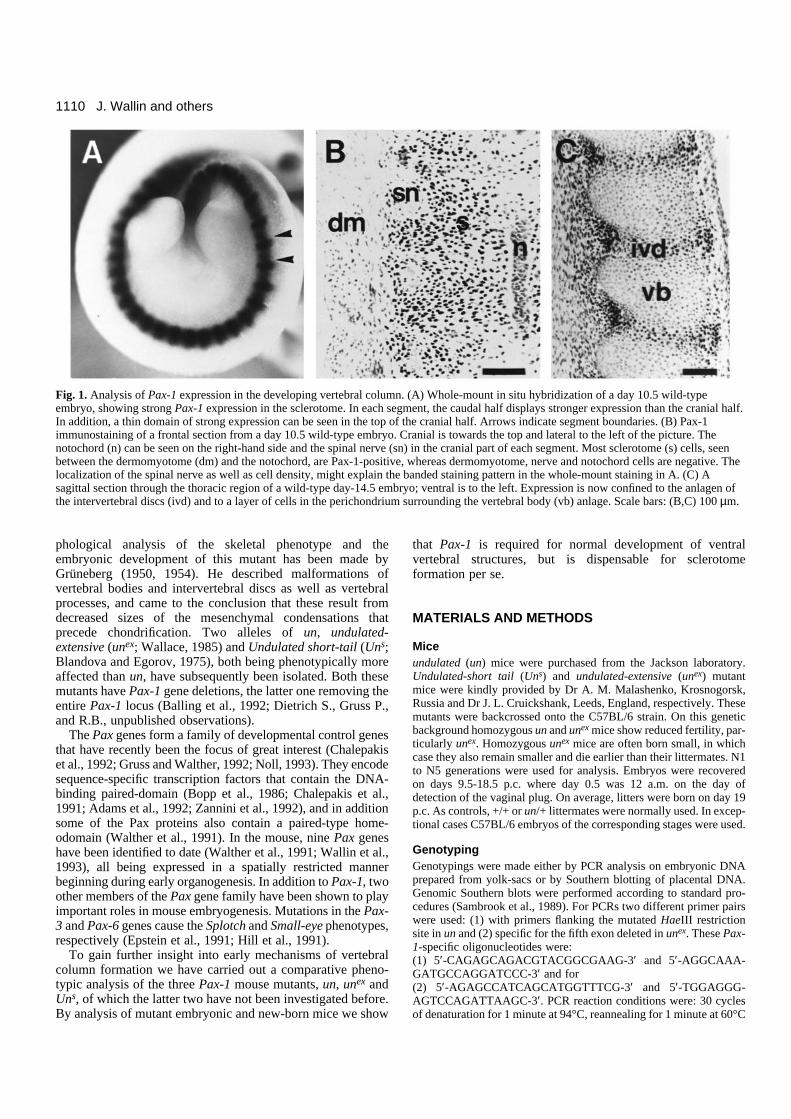
Fig. 1. Analysis of Pax-1 expression in the developing vertebral column. (A) Whole-mount in situ hybridization of a day 10.5 wild-typeembryo, showing strong Pax-1 expression in the sclerotome. In each segment, the caudal half displays stronger expression than the cranial half.In addition, a thin domain of strong expression can be seen in the top of the cranial half. Arrows indicate segment boundaries. (B) Pax-1immunostaining of a frontal section from a day 10.5 wild-type embryo. Cranial is towards the top and lateral to the left of the picture. Thenotochord (n) can be seen on the right-hand side and the spinal nerve (sn) in the cranial part of each segment. Most sclerotome (s) cells, seenbetween the dermomyotome (dm) and the notochord, are Pax-1-positive, whereas dermomyotome, nerve and notochord cells are negative. Thelocalization of the spinal nerve as well as cell density, might explain the banded staining pattern in the whole-mount staining in A. (C) Asagittal section through the thoracic region of a wild-type day-14.5 embryo; ventral is to the left. Expression is now confined to the anlagen ofthe intervertebral discs (ivd) and to a layer of cells in the perichondrium surrounding the vertebral body (vb) anlage. Scale bars: (B,C) 100 µm.

1111
Pax-1 and axial skeleton development
and extension for 1 minute at 72°C for 1 minute; 2.5 units Taq poly-merase (Amersham) using assay conditions specified by the manu-facturer in a 50 µl reaction volume. PCR products were separated inagarose gels and visualized by ethidium bromide staining. For typingof un, PCR products were digested with HaeIII before gel separation,producing allele-specific band sizes that could easily be scored. Fordiagnosis of Uns and unex homozygotes, a primer pair specific for anunlinked gene (Pax-9) was included as an internal control. A largenumber of un and unex embryos were also obtained from crosses ofhomozygous mice.
Histology For histological analysis, day 9.5-18.5 embryos were fixed in Bouin’ssolution and embedded in paraffin. 8 µm sections were stained eitherin haematoxylin-eosin or Azan. In addition, for day 10.5-12.5 p.c.embryos of the Uns/Uns, Uns/un and +/+ genotypes, semi-thinsections were made, focusing on the strongly affected lumbar region.Embryos were fixed in Karnovsky’s solution, embedded in plastic and0.75 µm sections were stained with toluidine blue.
Skeletal preparationsSkeletal preparations were made by a slight modification of the Alcianblue/alizarin red staining procedure described by Kessel et al. (1990).Specimens were fixed in 99% ethanol for 24 hours (fetuses older thanday 15 p.c. were first deskinned and eviscerated), and then kept inacetone for another 24 hours. Incubation in staining solution (1 vol.of 0.3% Alcian blue in 70% ethanol, 1 vol. of 0.1% alizarin red S in96% ethanol, 1 volume of absolute acetic acid, and 17 volumes of70% ethanol) was performed for 4-6 hours at 37°C and then over-night at room temperature. Samples were rinsed in water and kept in1% potassium hydroxide/20% glycerol at 37°C over-night, with addi-tional incubation at room temperature until complete clearing. Forlong term storage, specimens were transferred into 50%, 80% andfinally 100% glycerol.
In situ hybridizationPax-1 riboprobes were generated from a HincII-SacI paired boxfragment using either 35S-UTP or digoxigenin-11-UTP labelling for
in situ hybridization on sections or in whole embryos, respectively.Fixation, hybridization and subsequent detection procedures wereessentially as described by Kessel and Gruss (1991) and Rosen andBeddington (1993).
BrdU labellingCell proliferation was monitored by incorporation of BrdU intoembryos in utero. Pregnant mice were injected intraperitoneally witha 5 mg/ml BrdU (Sigma) solution in PBS. The amount injected wasapproximately 50 µg/g body weight. After 2 hours the mice werekilled and embryos isolated, fixed and embedded in paraffin. 8 µmsections were stained with an anti-BrdU antibody as described belowand counterstained with basic fuchsin or hematoxylin. The number oflabelled cells as well as total cell numbers were scored.
ImmunohistochemistryFor immunostainings, embryos were fixed in 3% acetic acid inabsolute ethanol at 4°C over-night, incubated first in absolute ethanoland then in xylene, both twice for 30 minutes, followed by a 1:1mixture of xylene:paraffin for 30 minutes at 55°C, infiltrated withparaffin by three incubations for 1 hour at 55°C and embedded.
The generation of the Pax-1-specific antiserum has been previouslydescribed (Chalepakis et al., 1991). The mouse monoclonal anti-BrdUantibody was purchased from Dako.
For BrdU immunocytochemistry, sections were deparaffinized andair-dried over-night. They were then incubated in 2 N HCl for 30minutes, washed in PBS, pH 6.0, for 1 minute and in PBS, pH 7.4(hereafter called PBS) for 5 minutes. Primary antibody was diluted1:50 in PBS, 1% BSA, 0.25% Tween-20 and incubated for 1 hour.Following washes, detection was made with the biotin-streptavidin-peroxidase amplification procedure, UniTect ABC-kit (Dianova).
Pax-1 immunostaining was made on deparaffinized sections thatwere first bleached in 0.3% H2O2 in methanol for 30 minutes, washedin PBS, blocked with 10% normal goat serum, in PBS for 60 minutesand then incubated with the antiserum diluted 1:200 in the blockingsolution. Detection was made with peroxidase-conjugated goat-anti-rabbit IgG (Sigma), and diaminobenzidine was used as chromogen.
Fig. 2. A schematic representation of the molecular basis of the Pax-1 deficiency of the un and Uns alleles (A), and the external appearance ofadult homozygous un (B), homozygous unex (C) and heterozygous Uns mice (D). In the upper part of A, the exon-intron organization of thePax-1 gene is depicted (Deutsch, 1990) The coding region is indicated as filled boxes and the paired box is hatched. The position of the pointmutation in un is marked by a cross. The exact locations of the deletion breakpoints in Uns have not been determined.
1112
RESULTS
Pax-1 expression during sclerotome developmentTo determine the role of Pax-1 in sclerotome differentiation,we have analysed Pax-1 RNA and protein expression in earlysomites and in subsequent axial skeleton development. Pax-1transcription can first be detected around day 8.5 p.c. in theventromedial part of newly formed somites (data not shown).This corresponds to the time of de-epithelialization of the ventralsomite half, i.e. the emergence ofthe sclerotome. At first, expressionis uniformly distributed in sclero-tome cells. In whole-mountexpression analysis of day-10.5embryos with a Pax-1 RNA probe,it was possible to distinguishbetween cranial and caudal parts ofthe sclerotomes, the caudal part ofeach segment being more stronglypositive and extending moredorsally than the cranial one (Fig.1A). At day 12.5, the Pax-1 tran-scripts are mainly confined to theanlagen of the intervertebral discand weaker domains can be seen inthe perichondria lining the cartilageblastemas of the vertebral bodies,the pedicles and the proximal ribs(data not shown). These findingsare in good accordance with thoseof Deutsch et al. (1988).
Pax-1 protein expression wasdetected with a rabbit antiserumraised against a peptide in thecarboxy-terminal end of the protein(Chalepakis et al., 1991). Theprotein is expressed with a delay ofabout 1 day compared to the onsetof transcription. First, at day 9.5,Pax-1 protein can be detected insclerotome cells, albeit weakly(data not shown). Immunostainingof day-10.5 embryos revealedexpression in almost all sclerotomecells (Fig. 1B). These cellscontinue to express Pax-1 up to thetime when mesenchymal cells startto chondrify in order to form thevertebral bodies, which takes placearound day 12.0 p.c. At day 12.5p.c., the cells of the anlagen of thevertebral bodies are not stained,while the anlagen of the interverte-bral discs remain strongly positive(Fig. 7E). Cells surrounding thevertebral body anlagen, those in thedisc anlagen as well as those in thecranial half of each lateral sclero-tome (forming connective tissuearound the spinal nerves and dorsal
root ganglia) remain strongly Pax-1-positive at day 14.5 p.c.(Figs 1C, 7G). In the vertebral column of new-born mice onlya small number of Pax-1-positive cells can be observed in theventral part of the annulus fibrosus (data not shown).
Comparative phenotypic analysis of three Pax-1mutant alleles We have chosen to study the phenotype of three spontaneously
J. Wallin and others
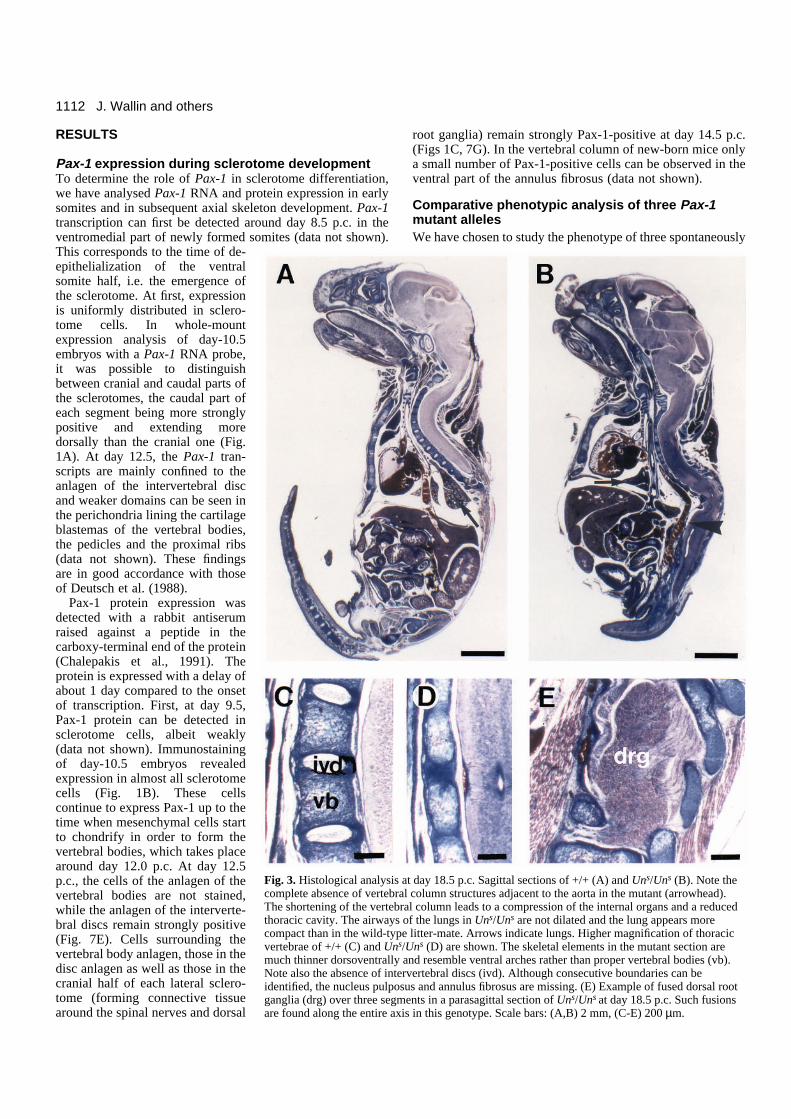
Fig. 3. Histological analysis at day 18.5 p.c. Sagittal sections of +/+ (A) and Uns/Uns (B). Note thecomplete absence of vertebral column structures adjacent to the aorta in the mutant (arrowhead).The shortening of the vertebral column leads to a compression of the internal organs and a reducedthoracic cavity. The airways of the lungs in Uns/Uns are not dilated and the lung appears morecompact than in the wild-type litter-mate. Arrows indicate lungs. Higher magnification of thoracicvertebrae of +/+ (C) and Uns/Uns (D) are shown. The skeletal elements in the mutant section aremuch thinner dorsoventrally and resemble ventral arches rather than proper vertebral bodies (vb).Note also the absence of intervertebral discs (ivd). Although consecutive boundaries can beidentified, the nucleus pulposus and annulus fibrosus are missing. (E) Example of fused dorsal rootganglia (drg) over three segments in a parasagittal section of Uns/Uns at day 18.5 p.c. Such fusionsare found along the entire axis in this genotype. Scale bars: (A,B) 2 mm, (C-E) 200 µm.
1113Pax-1 and axial skeleton development
arisen mouse mutants, un, unex andUns, since the molecular character-ization of these mice has shown thatthere is a point mutation in thepaired domain of un (Balling et al.,1988), whereas unex and Uns
display Pax-1 gene deletions. InUns the entire locus is deleted(schematically depicted in Fig. 2A).The absence of the Pax-1 gene inthis mutant has previously beendemonstrated for the paired boxregion (Wallin et al., 1993), and hasbeen extended to the 5′-flankingsequences and 3′-untranslatedregion (data not shown). The size ofthe Uns deletion has not beenexactly determined, but a majorchromosomal deletion has beenexcluded by karyotyping and bydemonstration of the presence, inUns homozygous DNA, of severalclosely linked DNA markers(GBASE, March, 1993, A. Y.Hillyard, D. P. Doolittle, M. T.Davisson, and T. H. Roderick, TheJackson Laboratory, Bar Harbor,ME) (data not shown). In unex thefifth exon, which includes partof the carboxy-terminal codingsequence, is deleted (Dietrich S.,Gruss P., and R. B., unpublishedobservations). The allelic nature ofall three mutations has also beenconfirmed by inter-crosses. In nocombination of the differentmutants was a complementationeffect observed.
The phenotypic appearance ofadult mutant mice is characterizedby their shortened and kinky tails(Fig. 2B-D). un is known as arecessive mutation (Wright, 1947),but in our colony we have observeda small percentage of heterozygousanimals that have slight but signifi-cant distal tail kinks, which is alsotrue for unex (data not shown). Thusalthough un/+ embryos occasionallydo display very mild abnormalities,they serve as good controls for thestronger genotypes for all practicalpurposes. Uns is invariably semi-dominant and produces a clearphenotype in the heterozygoussituation. Uns/Uns mice die shortlyafter birth. Their tails are only rudi-mentary and the trunks considerablyshorter than those of wild-type lit-termates (Fig. 3A,B), neverthelesspups show normal movements at
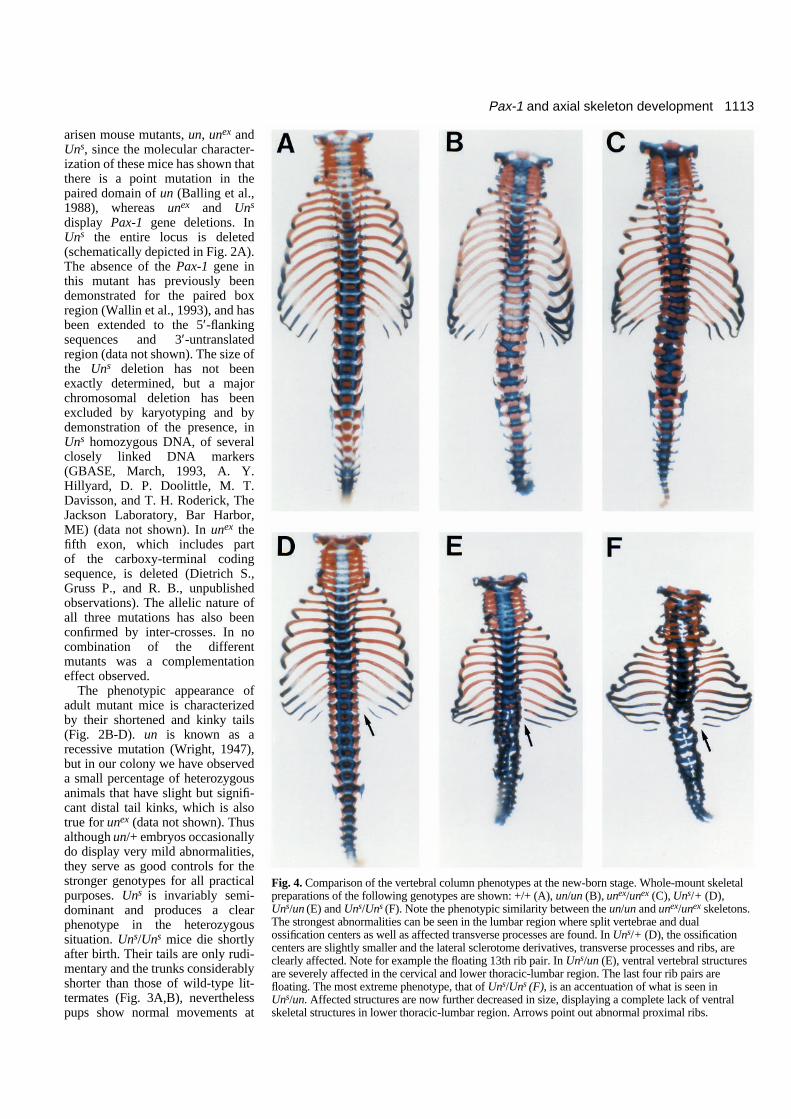
Fig. 4. Comparison of the vertebral column phenotypes at the new-born stage. Whole-mount skeletalpreparations of the following genotypes are shown: +/+ (A), un/un (B), unex/unex (C), Uns/+ (D),Uns/un (E) and Uns/Uns (F). Note the phenotypic similarity between the un/un and unex/unex skeletons.The strongest abnormalities can be seen in the lumbar region where split vertebrae and dualossification centers as well as affected transverse processes are found. In Uns/+ (D), the ossificationcenters are slightly smaller and the lateral sclerotome derivatives, transverse processes and ribs, areclearly affected. Note for example the floating 13th rib pair. In Uns/un (E), ventral vertebral structuresare severely affected in the cervical and lower thoracic-lumbar region. The last four rib pairs arefloating. The most extreme phenotype, that of Uns/Uns (F), is an accentuation of what is seen inUns/un. Affected structures are now further decreased in size, displaying a complete lack of ventralskeletal structures in lower thoracic-lumbar region. Arrows point out abnormal proximal ribs.
1114 J. Wallin and others
1115Pax-1 and axial skeleton development
birth. The reason for the postnatal lethality, which is alsoobserved for the Uns/un and Uns/unex compound heterozygotes,could not be exactly determined. The most likely explanationis respiratory failure as a result of lung space constraints. His-tological sections of Uns/Uns at day 18.5 p.c. show lungs withdrastically reduced airway lumina (Fig. 3A,B).
Whereas the kinks in un/un are soft and can easily becorrected upon pressure, the tail abnormalities in unex/unex andUns/+ are more pronounced and rigid, partially due to vertebralfusions. The increase in severity of the phenotypes, can beinterpreted as a gradual loss of Pax-1 function in the differentmutants. In general, the axial skeleton abnormalities in thesemutants are qualitatively similar but quantitatively different.This is more apparent when a detailed analysis of the vertebralcomponents is made. As the strongest phenotypes result inpostnatal death, we have concentrated our analyses and com-parisons on embryos up to the new-born stage.
Abnormalities in the vertebral column of new-bornmiceAll undulated mutants display phenotypic abnormalities of thevertebral column along the entire axis. A list of typicalvertebral column aberrations is given in Table 1. The abnor-malities affecting vertebral bodies or other axial skeletalelements are similar in all the mutants, but can be found todifferent degrees; the Uns/Uns genotype being the moststrongly affected. The severity of defects is not evenly distrib-uted along the axis and is most pronounced in the lumbar
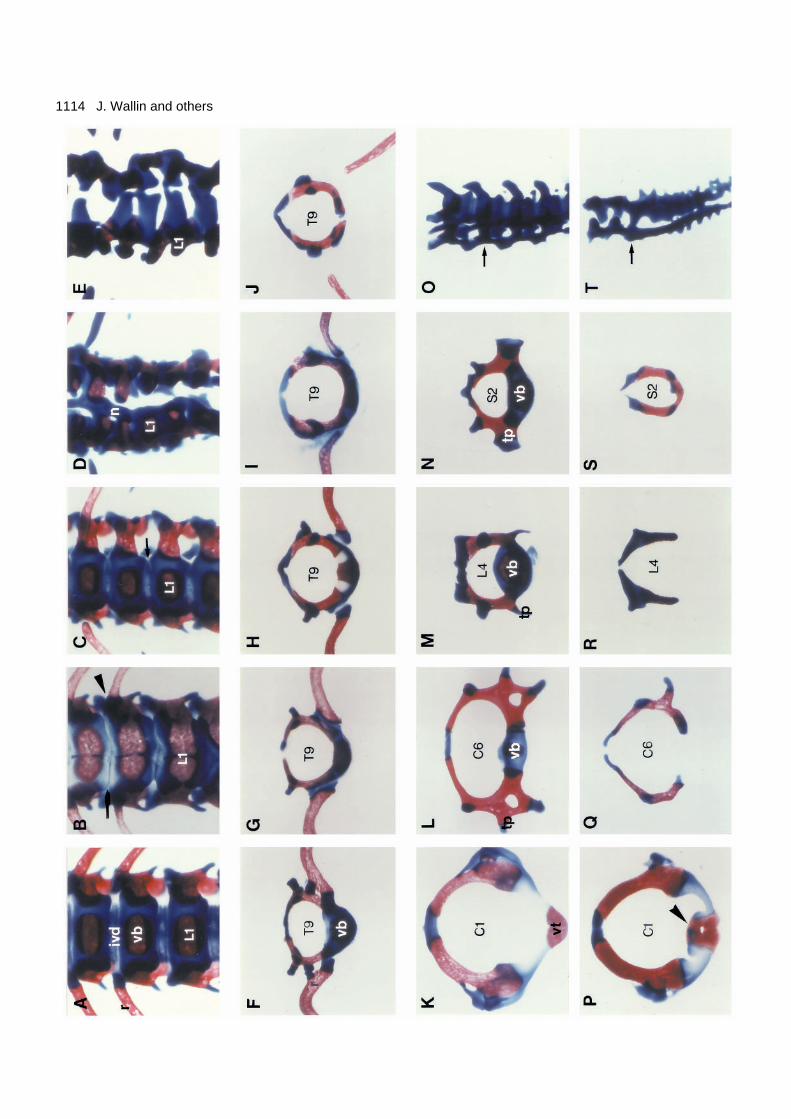
Fig. 5. Analysis of separate skeletal elements at the new-born stage.Frontal views of the lower thoracic-upper lumbar region of the +/+,un/un, Uns/+, Uns/un and Uns/Uns genotypes (A-E). In the new-bornwild-type skeleton (A) the ribs form joints at the lateral aspects ofthe intervertebral discs, which at this stage are well-definedstructures. In un/un, however, the 13th rib pair fuses to the vertebralbody (arrowhead) and the discs are virtually absent (arrow). In Uns/+(C), the annulus fibrosus is thinner than in the wild-type (arrow),whereas Uns/un (D) and Uns/Uns (E) have no bodies and discs. Notealso the notochord remnant in Uns/un (D) which is covered by acartilage layer. The ninth thoracic vertebrae of the same genotypes asA-E are compared in F-J, respectively. The principal similarity of thephenotypes can clearly be seen; vertebral bodies are reduced orabsent and the proximal part of the rib is abnormal or missing. Acomparison of isolated vertebrae from different craniocaudal levelsof +/+ (K-O) and Uns/Uns (P-T) new-born mice are also shown.Atlas (K,P), sixth cervical vertebrae (L,Q), fourth lumbar vertebrae(M,R), second sacral (N,S) and tails (O,T). The +/+ tail specimen inO shows the last sacral and the first three caudal vertebrae, while theUns/Uns (T) tail is composed of the last two sacral vertebrae as wellas remnants of several caudal vertebrae. Arrows indicate the positionof the first caudal vertebrae. With the exception of the atlas and thetail, the vertebral abnormalities at different levels are principallysimilar, i.e. lack of ventral and ventrolateral elements. A slight effecton of the neural arches can also be noted as a reduction of thespineous processes. The dens axis is fused to the atlas in Uns/Uns
(arrowhead; P). The tails are viewed at a slight angle and theabnormal tail of Uns/Uns has a trifurcated appearance (T). Oneventral and two lateral structures are observed. These vertebraeremnants are fused along the craniocaudal axis. ivd (intervertebraldisc), vb (vertebral body), n (notochord), L1 (first lumbar vertebra),T (thoracic), C (cervical), S (sacral), vt (ventral tubercle), tp(transverse process).
Table 1. Types of skeletal abnormalities that are observedin the vertebral column of un, unex and Uns mice
Perichordal tube derivatives Lateral sclerotome derivatives
Absence of vertebral body Absent or reduced transverse Split vertebral body processes“Ventral arches” Absent or reduced ribsFusions between atlas and dens axis Fusions between ribs and vertebral Reduced and malformed vertebral bodies
body (dual ossification centers) Fusions between adjacent neural Absent or reduced intervertebral arches
discs Reduced spinous processes“Haemal arches” (in proximal part Absent pedicles (only in caudal
of tail) vertebrae)Fusions between vertebral bodies
(mainly seen in tail)
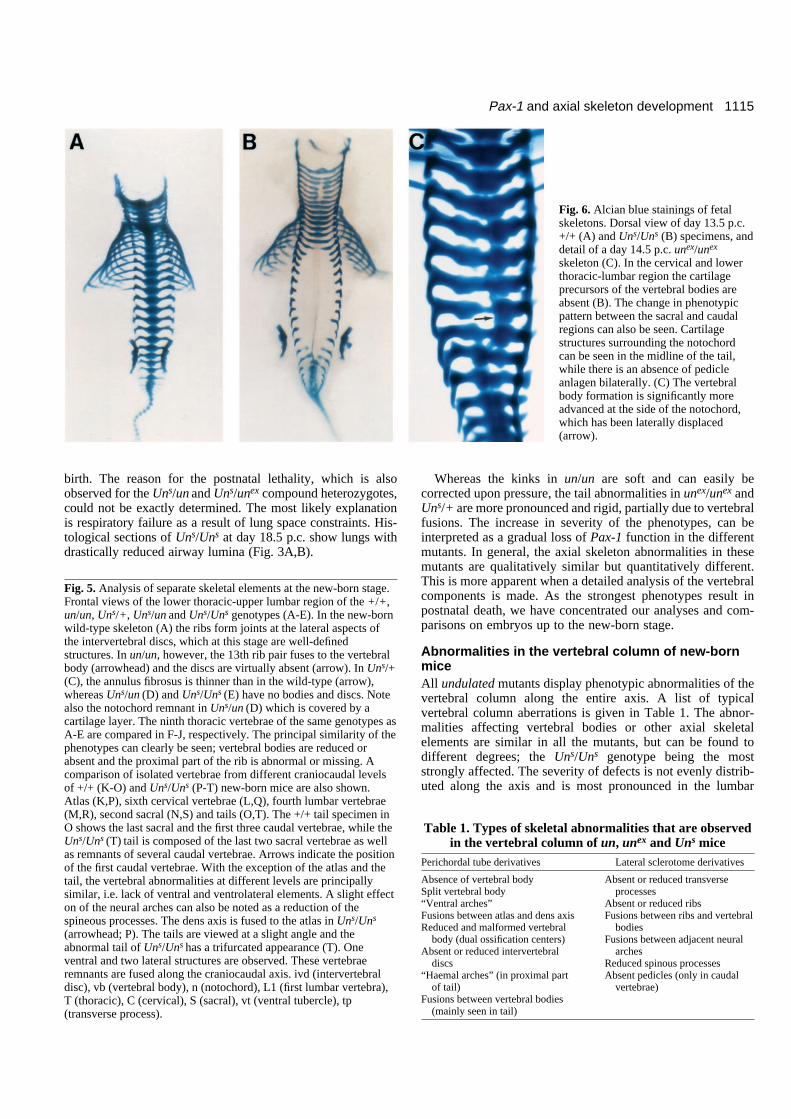
Fig. 6. Alcian blue stainings of fetalskeletons. Dorsal view of day 13.5 p.c.+/+ (A) and Uns/Uns (B) specimens, anddetail of a day 14.5 p.c. unex/unex
skeleton (C). In the cervical and lowerthoracic-lumbar region the cartilageprecursors of the vertebral bodies areabsent (B). The change in phenotypicpattern between the sacral and caudalregions can also be seen. Cartilagestructures surrounding the notochordcan be seen in the midline of the tail,while there is an absence of pedicleanlagen bilaterally. (C) The vertebralbody formation is significantly moreadvanced at the side of the notochord,which has been laterally displaced(arrow).

1116
region and the tail. A comparison of skeletal preparations atthe new-born stage is shown in Fig. 4. In the wild type, singleossification centres are formed in all vertebral bodies (Fig. 4A).The un and unex homozygous new-borns show split vertebrae,vertebrae with dual ossification centers and missing transverseprocesses primarily in the lumbar region (Fig. 4B,C). Thesephenotypes are very similar, although unex has slightly moresevere malformations and a few more segments are affected.In the case of Uns a mild phenotype can already be seen in het-erozygous mice (Fig. 4D). Vertebral body ossification centersare smaller than normal and transverse processes as well as theproximal parts of ribs are missing or reduced. Uns/Uns mice,with a total Pax-1 deficiency, completely lack vertebral bodiesin the lower thoracic and lumbar regions (Fig. 4F). Thecompound heterozygote Uns/un is intermediate in phenotypebetween Uns/Uns and un/un mice (Fig. 4E).
On closer inspection of the lower thoracic and upper lumbarregions, it is evident, that with the exception of Uns/+, thedifferent mutants lack intervertebral discs (Figs 3C,D and 5A-E). In Uns/+ the discs are thinner in the periphery, indicatinga disturbance in the formation of the annulus fibrosus. Theintervertebral disc is, at the new-born stage, composed partlyof hyaline cartilage, and can therefore be stained with Alcianblue. In adult mice the hyaline cartilage has been replaced byfibrous cartilage (Theiler, 1988) and appears clear in whole-mount skeletal preparations. In adult skeletal preparations thelack of discs in undulated could be confirmed (data not shown).In extreme cases the lack of discs leads to vertebral bodyfusions. On rare occasions mice of the Uns/unex genotype havesurvived after birth and at most lived up to five months of age.The vertebral column of a few of these mice displayed multiplefusions (data not shown).
Thus, in all mutants we detect disturbed formation of bothvertebral bodies and intervertebral discs. Furthermore, in allcases the proximal parts of the ribs as well as the rib homo-logues, the transverse processes, are affected. In un and unex
the last pairs of ribs are shorter and fuse abnormally to thevertebral bodies. Uns/+, Uns/un and Uns/Uns display moresevere abnormalities, with several floating rib pairs and the13th pair is completely missing in Uns/Uns (Fig. 5A-E).
A systematic comparison of the +/+, un/un, Uns/+, Uns/unand Uns/Uns genotypes at the level of the ninth thoracicvertebra clearly shows that the basic phenotype is essentiallythe same, and varies only in degree of severity between thealleles (Fig. 5F-J). Although we observe small variations inphenotypic severity within one allele (none of the strains arecompletely inbred), these are insignificant compared to thevariation between alleles.
Comparison of single vertebrae was made by dissection ofwhole-mount skeletal preparations. In Fig. 5K-T representativevertebrae from cervical, lumbar, sacral and caudal regions fromwild-type and Uns/Uns mice are shown. From this analysis itis evident that vertebral body and rib/transverse processformation are abnormal at all levels. Where a ventral ossifiedstructure can be seen, it resembles a pair of ventrally fusedneural arches rather than a vertebral body (Fig. 5J,S).Therefore, Uns/Uns mice are deficient in structures derivedboth from the perichordal tube and the ventralmost parts oflateral sclerotomes. The neural arches are fairly normallyshaped. Reductions of the spinous and articular processes aswell as rare fusions between adjacent neural arches can be
detected. These deformities are discrete in comparison to theventral ones and may to a large extent be due to the compres-sion of the vertebral column. The tail vertebrae display aphenotype which is different from the rest of axis. The tail-remnant in Uns/Uns gives a trifurcated appearance, represent-ing vestigial vertebral bodies and paired neural arches (Fig.5T). The pedicles, the structures connecting bodies and neuralarches, are lost. These malformed structures display only scantsigns of segmentation and are fused to the tip of the tail.
First phenotypic sclerotome abnormality at day 10.5p.c. of developmentTo study when the Pax-1 protein starts to exert its effect wehave conducted a detailed morphological analysis of mutantembryos. We have concentrated our study primarily on thelumbar region in the different mutants with particular emphasison the strongest genotypes Uns/un and Uns/Uns. The compoundheterozygote Uns/un has the advantage that the Pax-1 RNA andprotein expressed from the un allele can be detected in themutant embryos, although functionally this mutant is almost asaffected as the null-allele Uns/Uns.
Analyses of cartilage formation in skeletal clearance prepa-rations, revealed that axial structures in the vicinity of thenotochord never chondrify in mutant embryos (Fig. 6B). Themutant phenotype is visible from day 11.5 p.c., the first time-point when Alcian blue can be used to stain the developingskeleton. Therefore, in agreement with the earlier results ofGrüneberg (Grüneberg, 1954), it can be concluded that themalformations are formed at an early embryonic stage and thatthe primary function of Pax-1 should be looked for beforechondrogenesis commences. This lack of axial chondrogene-sis most often leads to a lateral displacement of the notochord.Vertebral body formation is always more normal on the sideof the notochord. This phenomenon, best observed in unex/unex
skeletal preparations (Fig. 6C), suggests that vertebral bodyformation is also dependent on a late notochord function.
Histological analysis of early vertebral column formation,reveals that sclerotome formation occurs largely normally inthe Pax-1-deficient mice. Morphological abnormalities canfirst be detected at day 10.5 p.c., when the lumbar sclerotomeregion already is abnormal compared with controls. In the wildtype, densely and loosely arranged areas are present and theaxial cells are oriented towards the notochord, whereas in themost affected mutants the sclerotome cells are evenly distrib-uted (Fig. 7A,B).
Abnormalities in the perichordal tube and lateralsclerotome regions At day 12.5 p.c. more profound changes can be detected. Inmid-sagittal sections of Uns/Uns and Uns/un embryos, in theregion of the perichordal tube, cell numbers are stronglyreduced, corresponding to the loss of the anlagen for thevertebral bodies and intervertebral discs (Fig. 7E,F). The fewcells present in the perichordal tube of Uns/un embryos at thisstage do not display the normal metameric arrangement ofcondensed intervertebral disc anlagen and cell-sparse vertebralbody anlagen. We performed immunostainings on mutant andwild-type embryos to compare the distribution of Pax-1-expressing cells. As can be seen in Fig. 7, the Pax-1-positivedomains are much wider in the craniocaudal dimension in theUns/un mutant. The observation that most cells are Pax-1-
J. Wallin and others
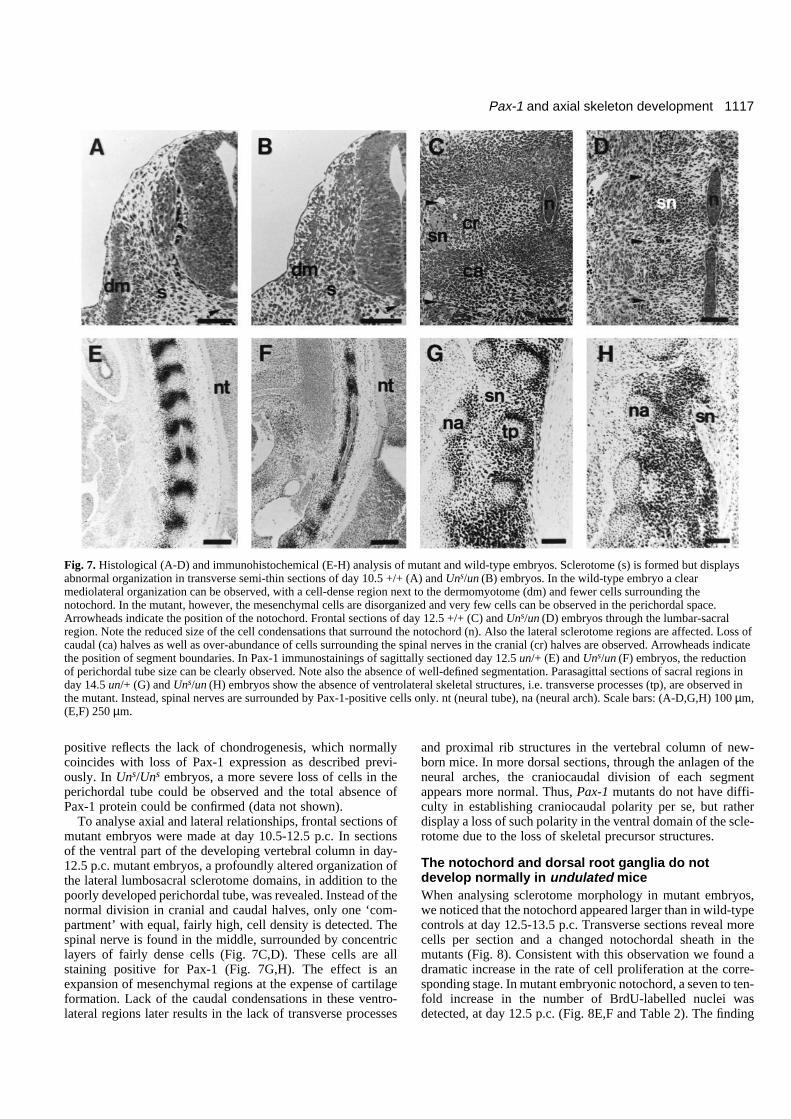
1117Pax-1 and axial skeleton development
positive reflects the lack of chondrogenesis, which normallycoincides with loss of Pax-1 expression as described previ-ously. In Uns/Uns embryos, a more severe loss of cells in theperichordal tube could be observed and the total absence ofPax-1 protein could be confirmed (data not shown).
To analyse axial and lateral relationships, frontal sections ofmutant embryos were made at day 10.5-12.5 p.c. In sectionsof the ventral part of the developing vertebral column in day-12.5 p.c. mutant embryos, a profoundly altered organization ofthe lateral lumbosacral sclerotome domains, in addition to thepoorly developed perichordal tube, was revealed. Instead of thenormal division in cranial and caudal halves, only one ‘com-partment’ with equal, fairly high, cell density is detected. Thespinal nerve is found in the middle, surrounded by concentriclayers of fairly dense cells (Fig. 7C,D). These cells are allstaining positive for Pax-1 (Fig. 7G,H). The effect is anexpansion of mesenchymal regions at the expense of cartilageformation. Lack of the caudal condensations in these ventro-lateral regions later results in the lack of transverse processes
and proximal rib structures in the vertebral column of new-born mice. In more dorsal sections, through the anlagen of theneural arches, the craniocaudal division of each segmentappears more normal. Thus, Pax-1 mutants do not have diffi-culty in establishing craniocaudal polarity per se, but ratherdisplay a loss of such polarity in the ventral domain of the scle-rotome due to the loss of skeletal precursor structures.
The notochord and dorsal root ganglia do notdevelop normally in undulated miceWhen analysing sclerotome morphology in mutant embryos,we noticed that the notochord appeared larger than in wild-typecontrols at day 12.5-13.5 p.c. Transverse sections reveal morecells per section and a changed notochordal sheath in themutants (Fig. 8). Consistent with this observation we found adramatic increase in the rate of cell proliferation at the corre-sponding stage. In mutant embryonic notochord, a seven to ten-fold increase in the number of BrdU-labelled nuclei wasdetected, at day 12.5 p.c. (Fig. 8E,F and Table 2). The finding
Fig. 7. Histological (A-D) and immunohistochemical (E-H) analysis of mutant and wild-type embryos. Sclerotome (s) is formed but displaysabnormal organization in transverse semi-thin sections of day 10.5 +/+ (A) and Uns/un (B) embryos. In the wild-type embryo a clearmediolateral organization can be observed, with a cell-dense region next to the dermomyotome (dm) and fewer cells surrounding thenotochord. In the mutant, however, the mesenchymal cells are disorganized and very few cells can be observed in the perichordal space.Arrowheads indicate the position of the notochord. Frontal sections of day 12.5 +/+ (C) and Uns/un (D) embryos through the lumbar-sacralregion. Note the reduced size of the cell condensations that surround the notochord (n). Also the lateral sclerotome regions are affected. Loss ofcaudal (ca) halves as well as over-abundance of cells surrounding the spinal nerves in the cranial (cr) halves are observed. Arrowheads indicatethe position of segment boundaries. In Pax-1 immunostainings of sagittally sectioned day 12.5 un/+ (E) and Uns/un (F) embryos, the reductionof perichordal tube size can be clearly observed. Note also the absence of well-defined segmentation. Parasagittal sections of sacral regions inday 14.5 un/+ (G) and Uns/un (H) embryos show the absence of ventrolateral skeletal structures, i.e. transverse processes (tp), are observed inthe mutant. Instead, spinal nerves are surrounded by Pax-1-positive cells only. nt (neural tube), na (neural arch). Scale bars: (A-D,G,H) 100 µm,(E,F) 250 µm.
1118
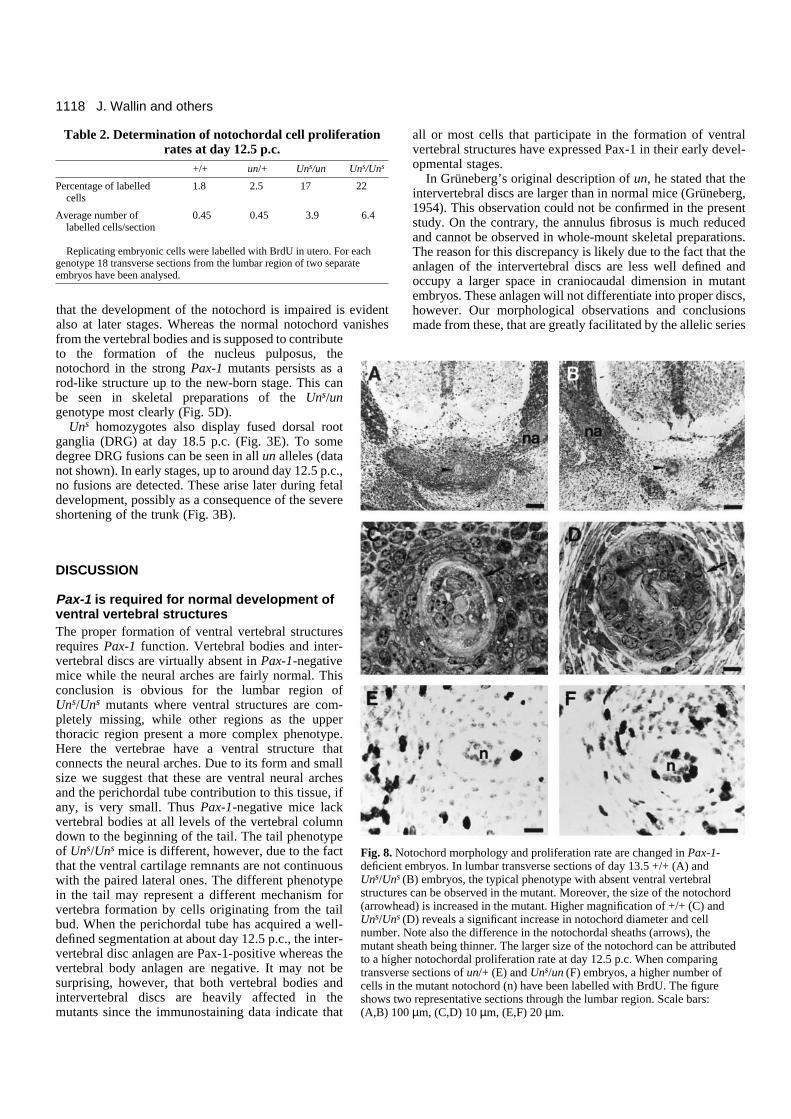
that the development of the notochord is impaired is evidentalso at later stages. Whereas the normal notochord vanishesfrom the vertebral bodies and is supposed to contributeto the formation of the nucleus pulposus, thenotochord in the strong Pax-1 mutants persists as arod-like structure up to the new-born stage. This canbe seen in skeletal preparations of the Uns/ungenotype most clearly (Fig. 5D).
Uns homozygotes also display fused dorsal rootganglia (DRG) at day 18.5 p.c. (Fig. 3E). To somedegree DRG fusions can be seen in all un alleles (datanot shown). In early stages, up to around day 12.5 p.c.,no fusions are detected. These arise later during fetaldevelopment, possibly as a consequence of the severeshortening of the trunk (Fig. 3B).
DISCUSSION
Pax-1 is required for normal development ofventral vertebral structuresThe proper formation of ventral vertebral structuresrequires Pax-1 function. Vertebral bodies and inter-vertebral discs are virtually absent in Pax-1-negativemice while the neural arches are fairly normal. Thisconclusion is obvious for the lumbar region ofUns/Uns mutants where ventral structures are com-pletely missing, while other regions as the upperthoracic region present a more complex phenotype.Here the vertebrae have a ventral structure thatconnects the neural arches. Due to its form and smallsize we suggest that these are ventral neural archesand the perichordal tube contribution to this tissue, ifany, is very small. Thus Pax-1-negative mice lackvertebral bodies at all levels of the vertebral columndown to the beginning of the tail. The tail phenotypeof Uns/Uns mice is different, however, due to the factthat the ventral cartilage remnants are not continuouswith the paired lateral ones. The different phenotypein the tail may represent a different mechanism forvertebra formation by cells originating from the tailbud. When the perichordal tube has acquired a well-defined segmentation at about day 12.5 p.c., the inter-vertebral disc anlagen are Pax-1-positive whereas thevertebral body anlagen are negative. It may not besurprising, however, that both vertebral bodies andintervertebral discs are heavily affected in themutants since the immunostaining data indicate that
all or most cells that participate in the formation of ventralvertebral structures have expressed Pax-1 in their early devel-opmental stages.
In Grüneberg’s original description of un, he stated that theintervertebral discs are larger than in normal mice (Grüneberg,1954). This observation could not be confirmed in the presentstudy. On the contrary, the annulus fibrosus is much reducedand cannot be observed in whole-mount skeletal preparations.The reason for this discrepancy is likely due to the fact that theanlagen of the intervertebral discs are less well defined andoccupy a larger space in craniocaudal dimension in mutantembryos. These anlagen will not differentiate into proper discs,however. Our morphological observations and conclusionsmade from these, that are greatly facilitated by the allelic series
J. Wallin and others
Fig. 8. Notochord morphology and proliferation rate are changed in Pax-1-deficient embryos. In lumbar transverse sections of day 13.5 +/+ (A) andUns/Uns (B) embryos, the typical phenotype with absent ventral vertebralstructures can be observed in the mutant. Moreover, the size of the notochord(arrowhead) is increased in the mutant. Higher magnification of +/+ (C) andUns/Uns (D) reveals a significant increase in notochord diameter and cellnumber. Note also the difference in the notochordal sheaths (arrows), themutant sheath being thinner. The larger size of the notochord can be attributedto a higher notochordal proliferation rate at day 12.5 p.c. When comparingtransverse sections of un/+ (E) and Uns/un (F) embryos, a higher number ofcells in the mutant notochord (n) have been labelled with BrdU. The figureshows two representative sections through the lumbar region. Scale bars: (A,B) 100 µm, (C,D) 10 µm, (E,F) 20 µm.
Table 2. Determination of notochordal cell proliferationrates at day 12.5 p.c.
+/+ un/+ Uns/un Uns/Uns
Percentage of labelled 1.8 2.5 17 22cells
Average number of 0.45 0.45 3.9 6.4labelled cells/section
Replicating embryonic cells were labelled with BrdU in utero. For eachgenotype 18 transverse sections from the lumbar region of two separateembryos have been analysed.

1119Pax-1 and axial skeleton development
now available, are otherwise in good agreement withGrüneberg’s work on un (1950, 1954).
The role of Pax-1 in intervertebral disc development mightrepresent a late function compared to the patterning of earlysclerotome cells. In later stages, Pax-1 expression is found indisc anlagen and lining the skeletal structures. Pax-1 may herehave a role in the differentiation of fibrous tissue, andtherefore, in the maintenance of the boundaries betweenskeletal elements. Indeed, fusions between axial skeletalelements are frequently seen in the mutants.
The strong phenotypic similarities in the three Pax-1mutants leave little doubt that undulated is due to a reductionof Pax-1 function. While in un the phenotype can be linked toa single point mutation in the Pax-1 gene, the deletion sizes ofthe other two alleles are not known. It thus remains an openquestion whether other deleted genes also significantly con-tribute to the strong phenotype seen in Uns/Uns mice. Webelieve this not to be the case, because of the apparent lack ofany phenotypic changes that cannot be observed in a milderform in un mice. This will not be strictly proven, however, untila clean Pax-1 null mutation has been accomplished via homol-ogous recombination in ES cells and introduced into thegermline or until Uns/Uns mice have been rescued via trans-genesis. We suggest that Uns/Uns mice display a Pax-1 nullphenotype and that the weaker mutations are hypomorphs. Anobservation in support of that idea is that Pax-1un protein stillretains DNA-binding activity, albeit much weaker than thewild-type protein (Chalepakis et al., 1991). Further support forthis notion is provided by the demonstration that the causativegenetic change in the Splotch-delayed mutant, is a pointmutation in the paired domain, similar to the situation in un.;the phenotype of this mutant was interpreted to be due to partialloss of Pax-3 function (Vogan et al., 1993).
Recently, a detailed study of the interaction of paireddomains with DNA target sequences revealed that this DNAbinding domain is composed of two subdomains (Czerny et al.,1993). These authors discuss that a mutation in one of the sub-domains may affect the binding to only a subset of target genes.This speculation is of great interest for the present study, as unis one of the potential models. Indeed, a differential effect onseparate targets is consistent with our phenotypic analyses.Although all abnormalities to some degree can be seen in allmutants, the relative strength of phenotypic alterations indifferent regions are not always proportional. Thus, when thephenotype of un is compared to that of Uns, it can be seen thatthe point-mutation in un affects development in axial sclero-tome derivatives relatively more than lateral ones, i.e. therib/transverse process phenotype is fairly weak in un.
We have recently identified a Pax gene that is highly homol-ogous to Pax-1 (Wallin et al., 1993). This gene, Pax-9, isexpressed in a similar but not identical pattern during sclero-tome development. To analyse whether the expression of thisgene is dependent on Pax-1 we have compared Pax-9expression domains in wild-type and Uns/un embryos (A.Neubüser, J. W. and R. B., unpublished observations). Wehave not been able to detect any changes in expression patternsthat may not primarily be due to morphological changes.
Sclerotome and notochord are interdependentstructuresA common denominator of the affected vertebral structures is
their proximity to the notochord. The development of thevertebral column is dependent on the presence of the notochord(Watterson et al., 1954), and a supernumerary notochordinduces the formation of additional vertebral body-like struc-tures, but represses myotome formation, in surgically manipu-lated chicken embryos (Brand-Saberi et al., 1993; Pourquie etal., 1993). Interestingly, it was also shown that the notochordis able to induce Pax-1 expression in dorsal somite derivatives,that are normally Pax-1 negative (Brand-Saberi et al., 1993).The induced Pax-1 expression is accompanied by a loss of der-momyotome cells and the appearance of sclerotome-like mes-enchymal cells. These observations make Pax-1 a very goodcandidate for a mediator of notochordal signals to the sclero-tome, and as such might function as an embryonic competencefactor. Based on the analysis of Pax-1 expression in thenotochord mouse mutant Danforth’s short-tail (Sd) and com-parison of the Sd and un phenotypes, we have suggested thatPax-1 might be one of the major mediators of inductive signalsfrom the notochord to the sclerotome (Koseki et al., 1993). Ourobservation that vertebral body formation is often differentiallyaffected on either side of the midline is correlated with a dis-placement of the notochord to the more normal side. Wesuggest that the development of the bodies is dependent onnotochord signals that come significantly later than thefactor(s) that initially induce Pax-1 expression. The simplestinterpretation would be that sustained notochord signalling isrequired for normal maintenance of Pax-1 expression, whichin turn controls the downstream cascade involved in theformation of vertebral bodies and intervertebral discs. Alter-natively, Pax-1 acts as a competence factor in sclerotome cellsenabling them to respond to further notochord signals.
A strong rib phenotype was observed in Uns/Uns. Theproximal part is absent or reduced while lateral rib structuresare unaffected. This division corresponds fairly well to thoseparts of the ribs that are apparently derived from medial andlateral somite-halves, respectively; the medial half gives riseto proximal ribs, whereas the lateral half gives rise to the moredistal part of the rib (Charles Ordahl, personal communica-tion). It may be that only the medial somite-half is dependenton notochord signals; in Sd, the rib phenotype is similar to theone in Uns/Uns (Koseki et al., 1993). Mutant mice with aphenotype that is virtually the opposite of that for Uns withrespect to the ribs have been recently described (Braun et al.,1992). These mice carry targeted mutations in the myf-5 gene.
To our surprise the morphological changes in the mutantalso involve the notochord. Enlarged notochords were shownto have increased proliferation rates, and abnormal develop-ment results in the persistence of the notochord in vertebrae.These observations indicate that proper notochord develop-ment is dependent on surrounding sclerotome cells, whichmeans that signalling may be bidirectional. Normally thenotochord expands significantly at the level of the interverte-bral discs, whereas it vanishes from the centres of the vertebralbodies. We suggest that notochordal cell proliferation isnormally suppressed by surrounding perichordal tube cells andthat subsequent differentiation of intervertebral discs leads toan expansion of the notochord only at these levels. A schematicmodel for the induction, expression and functional role of Pax-1 is presented in Fig. 9.
Another unexpected finding was the fusion of dorsal rootganglia. It is conceivable that this abnormality in a neural tissue

1120
is secondary to sclerotome abnormalities. It is well establishedthat only the cranial half of each lateral sclerotome segment ispermissive for neural crest cell migration and dorsal rootganglion growth (Keynes and Stern, 1984). In transplantationexperiments in the avian system, it has been shown that neuralcrest-derived cell development in the context of cranial halfsomites only resulted in nonsegmented coalesced ganglia(Kalcheim and Teillet, 1989). In the Pax-1 mutants, theabnormal sclerotome cells in the cranial compartment mayaffect ganglion growth in a way akin to the influence on thenotochord. Alternatively, the shortening of the vertebralcolumn may cause a mechanical compression that forces theganglia together, resulting in fusions.
Pax genes and organ developmentFrom our observations in un mutant mice, we speculate thatPax-1 may act as an embryonic differentiation factor, makingventral mesenchymal cells competent to differentiate intohyaline and fibrous cartilage. Several mutant phenotypesresulting from genetic alterations in Pax genes have beendescribed in diverse species (Chalepakis et al., 1992; Gruss andWalther, 1992; Noll, 1993), making the Pax gene family oneof the currently better understood groups of developmentalgenes. Studies on the different Pax mouse mutants have doc-umented a central role for Pax genes in morphogenesis.Splotch, a Pax-3 mutant, displays overgrowth of the neuraltube with exencephaly and spina bifida, affecting also themigration of pigment cells, derivatives of the neural crest(Auerbach, 1954). Small-eye embryos, which carry a mutationin the Pax-6 gene, also display neural crest cell migrationproblems (Matsuo et al., 1993; Schmahl et al., 1993). Our dataon the undulated phenotype might be interpreted as a problemof sclerotome cell proliferation, differentiation and/ormigration. The significant reduction of ventral sclerotome cellshas not been correlated to a readily detectable decrease in scle-rotome cell proliferation rate (J. W. and R. B., unpublishedobservations). These preliminary data are, however, compati-ble with a slight but significant change in proliferation rate.Compared to the notochord, which displayed a large increasein proliferation rate, the developing sclerotome is a complexstructure with regional differences in all three dimensions. Anextended analysis is therefore required to document changes inthe different sclerotome regions. One study that argues infavour of at least some proliferative role of Pax genes is the
demonstration of an oncogenic activity of these genes(Maulbecker et al., 1993). Recently, a role for Pax-2 in the con-version of mesenchyme to epithelium during kidney develop-ment was demonstrated in an organ culture system (Rothen-pieler and Dressler, 1993). This situation, where earlymesenchymal cells fail to aggregate and differentiate, bears astrong resemblance to the phenotype in undulated mice wheremesenchymal sclerotome cells fail to condense and initiatechondrogenesis. We propose that Pax-1 plays a crucial role inthe early differentiation of ventral sclerotome cells, leading toprofound changes in morphology and extracellular matrixcomposition. Very little is known about downstream targetgenes under control of Pax transcription factors. So far onlythree candidate target genes have been suggested; CD19 forPax-5 and thyroperoxidase and thyroglobulin for Pax-8(Kozmik et al., 1992; Zannini et al., 1992). The identificationof Pax-1 as an essential gene for early sclerotome developmentprovides a starting point, however, for a characterization of thecascade of events that underlie the morphogenesis of the axialskeleton.
We thank Peter Gruss for the gift of Pax-1 cDNA, Mary Dickinsonand Andy McMahon for the BrdU-labelling protocol, Bärbel Strack,Monika Schüttoff, Günther Frank and Sybille Antoni for technicalassistance, Uli Birsner for oligonucleotide synthesis and Lore Lay forphotographic work. J. W. was supported by an EMBO fellowship andthe Swedish Medical Research Council. H. K. was supported by theHuman Frontier Science Program. The work was supported by theMax-Planck-Society and the Deutsche Forschungsgemeinschaft (Ba869/3-1).
REFERENCES
Adams, B., Dörfler, P., Aguzzi, A., Kozmik, Z., Urbanék, P., Maurer-Fogy,I. and Busslinger, M. (1992). Pax-5 encodes the transcription factor BSAPand is expressed in B lymphocytes, the developing CNS and adult testis.Genes Dev. 6, 1589-1607.
Auerbach, R. (1954). Analysis of the developmental effects of a lethalmutation in the house mouse. J. Exp. Zool. 127, 305-329.
Bagnall, K. M. (1992). The migration and distribution of somite cells afterlabelling with the carbocyanine dye, DiI: the relationship of this distributionto segmentation in the vertebrate body. Anat. Embryol. 185, 317-324.
Balling, R., Deutsch, U. and Gruss, P. (1988). Undulated, a mutationaffecting the development of the mouse skeleton, has a point mutation in thepaired box of Pax 1. Cell 55, 531-535.
Balling, R., Lau, C. F., Dietrich, S., Wallin, J. and Gruss, P. (1992).Development of the skeletal system. Ciba Foundation Symposium 165, 132-145. Chichester: Wiley.
J. Wallin and others
Fig. 9. A schematic model for the induction of Pax-1 expression and the role of Pax-1 in vertebral column development. (A) Notochord signalsinduce Pax-1 expression in the ventromedial part of the somite (Brand-Saberi et al., 1993; Koseki et al., 1993). (B) Pax-1-expressingsclerotome cells give rise to ventral parts of the vertebral column. In this developmental process, there is a bidirectional dependence of cells ofthe notochord and the anlagen of the vertebral bodies and intervertebral discs for their proper differentiation (arrows). (C) The majorconsequences of a Pax-1 deficiency are absence of vertebral bodies and intervertebral discs (black) and the loss of proximal rib structures(grey) or rib homologues, i.e. the transverse processes. The neural arches (white) are relatively unaffected.

1121Pax-1 and axial skeleton development
Blandova, Y. R. and Egorov, I. U. (1975). Sut allelic with un. Mouse NewsLett. 52, 43.
Bopp, D., Burri, M., Baumgartner, S., Frigerio, G. and Noll, M. (1986).Conservation of a large protein domain in the segmentation gene paired andin functionally related genes of Drosophila. Cell 47, 1033-1040.
Brand-Saberi, B., Ebensperger, C., Wilting, J., Balling, R. and Christ, B.(1993). The ventralizing effect of the notochord on somite differentiation inchick embryos. Anat. Embryol. 188, 239-245.
Braun, T., Rudnicki, M. A., Arnold, H. H. and Jaenisch, R. (1992). Targetedinactivation of the muscle regulatory gene myf-5 results in abnormal ribdevelopment and perinatal death. Cell 71, 369-382.
Chalepakis, G., Fritsch, R., Fickenscher, H., Deutsch, U., Goulding, M. andGruss, P. (1991). The molecular basis of the undulated/Pax-1 mutation. Cell66, 873-884.
Chalepakis, G., Tremblay, P. and Gruss, P. (1992). Pax genes, mutants andmolecular function. J. Cell Sci. 16 (suppl.), 61-67.
Christ, B. and Wilting, J. (1992). From somites to vertebral column. Ann.Anat. 174, 23-32.
Czerny, T., Schaffner, G. and Busslinger, M. (1993). DNA sequencerecognition by Pax proteins: bipartite structure of the paired domain and itsbinding site. Genes Dev. 7, 2048-2061.
Deutsch, U. (1990). Identifizierung der Paired-Box-Genfamile der Maus undCharakterisierung von Pax-1 als Entwicklungs-Kontrollgen. Inaugural-Dissertation. Heidelberg.
Deutsch, U., Dressler, G. R. and Gruss, P. (1988). Pax1, a member of a pairedbox homologous murine gene family, is expressed in segmented structuresduring development. Cell 53, 617-625.
Epstein, D. J., Vekemans, M. and Gros, P. (1991). Splotch (Sp2h), a mutationaffecting development of the mouse neural tube, shows a deletion within thepaired homeodomain of Pax-3. Cell 67, 767-774.
Ewan, K. B. R. and Everett, A. W. (1992). Evidence for resegmentation in theformation of the vertebral column using the novel approach of retroviral-mediated gene transfer. Exp. Cell Res. 198, 315-320.
Grüneberg, H. (1950). Genetical studies on the skeleton of the mouse. II.Undulated and its modifiers. J. Genet. 50, 142-173.
Grüneberg, H. (1954). Genetical studies on the skeleton of the mouse. XII. Thedevelopment of undulated. J. Genet. 52, 441-455.
Grüneberg, H. (1963). The Pathology of Development. Oxford: BlackwellScientific.
Gruss, P. and Walther, C. (1992). Pax in development. Cell 69, 719-722.Hill, R. E., Favor, J., Hogan, B. L. M., Ton, C. C. T., Saunders, G. F.,
Hanson, I. M., Prosser, J., Jordan, T., Hastie, N. D. and van Heyningen,V. (1991). Mouse Small eye results from mutations in a paired-likehomeobox-containing gene. Nature 354, 522-525.
Kalcheim, C. and Teillet, M.A. (1989) Consequences of somite manipulationon the pattern of dorsal root ganglion development. Development 106, 85-93.
Kessel, M., Balling, R. and Gruss, P. (1990). Variations of cervical vertebraeafter expression of a Hox-1.1 transgene in mice. Cell 61, 301-308.
Kessel, M. and Gruss, P. (1991). Homeotic transformations of murinevertebrae and concomitant alteration of Hox codes induced by retinoic acid.Cell 67, 1-20.
Keynes, R. J. and Stern, C. D. (1984). Segmentation in the vertebrate nervoussystem. Nature 310, 786-789.
Koseki, H., Wallin, J., Wilting, J., Mizutani, Y., Kispert, A., Ebensperger,C., Herrmann, B. G., Christ, B. and Balling, R. (1993). A role for Pax-1 as
a mediator of notochordal signals during the dorsoventral specification ofvertebrae. Development 119, 649-660.
Kozmik, Z., Wang, S., Dörfler, P., Adams, B. and Busslinger, M. (1992).The promoter of the CD19 gene is a target for the B-cell-specifictranscription factor BSAP. Mol. Cell. Biol. 12, 2662-2672.
Maulbecker, C. C. and Gruss, P. (1993). The oncogenic potential of Paxgenes. EMBO J. 12, 2361-2367.
Matsuo, T., Osumi-Yamashita, N., Noji, S., Ohuchi, H., Koyama, E.,Myokai, F., Matsuo, N., Taniguchi, S., Doi, H., Iseki, S., Ninomiya, Y.,Fujiwara, M., Watanabe, T. and Eto, K. (1993). A mutation in the Pax-6gene in rat small eye is associated with impaired migration of midbrain cells.Nature Genet. 3, 299-304.
Noll, M. (1993). Evolution and role of Pax genes. Curr. Opin. Genet. Dev. 3,595-605.
Pourquie, O., Coltey, M., Teillet, M.-A., Ordahl, C. and Le Douarin, N.(1993). Control of dorsoventral patterning of somitic derivatives bynotochord and floor plate. Proc. Natl. Acad. Sci. USA 90, 5242-5246.
Rosen, B. and Beddington, R. S. P. (1993). Whole-mount in situ hybridizationin the mouse embryo: gene expression in three dimensions. Trends Genet. 9,162-167.
Rothenpieler, U. W. and Dressler, G. R. (1993). Pax-2 is required formesenchyme-to-epithelium conversion during kidney development.Development 119, 711-720.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning. ALaboratory Manual. Cold Spring Harbour NY: Cold Spring HarbourLaboratory Press
Schmahl, W., Knoediseder, M., Favor, J. and Davidson, D. (1993). Defectsof neuronal migration and the pathogenesis of cortical malformations areassociated with Small eye (Sey) in the mouse, a point mutation at the Pax-6locus. Acta Neuropathol. 86, 126-135.
Theiler, K. (1988). Vertebral malformations Adv. Anat. Embryol. Cell Biol.112, 1-99.
Verbout, A. J. (1985). The development of the vertebral column. Adv. Anat.Embryol. Cell Biol. 90, 1-122.
Vogan, K. J., Epstein, D. J., Trasler, D. G. and Gros, P. (1993). The Splotch-delayed (Spd) mouse mutant carries a point mutation within the paired box ofthe Pax-3 gene. Genomics 17, 364-369.
Wallace, M. E. (1985). An inherited agent of mutation with chromosomedamage in wild mice. J. Heredity 76, 271-278.
Wallin, J., Mizutani, Y., Imai, K., Miyashita, N., Moriwaki, K., Taniguchi,M., Koseki, H. and Balling, R. (1993). A new Pax gene, Pax-9, maps tomouse chromosome 12. Mammalian Genome 4, 354-358.
Walther, C., Guenet, J.-L., Simon, D., Deutsch, U., Jostes, B., Goulding,M., Plachov, D., Balling, R. and Gruss, P. (1991). Pax: A murine multigenefamily of paired box containing genes. Genomics 11, 424-434.
Watterson, R. L., Fowler, I. and Fowler, B. J. (1954). The role of the neuraltube and notochord in development of the axial skeleton of the chick. Am. J.Anat. 95, 337-397.
Wright, M. E. (1947). Undulated: A new genetic factor in Mus musculusaffecting the spine and tail. Heredity 1, 137-141.
Zannini, M., Francis-Lang, H., Plachov, D. and Di Lauro, R. (1992). Pax-8,a paired domain-containing protein, binds to a sequence overlapping therecognition site of a homeodomain and activates transcription from twothyroid-specific promoters. Mol. Cell. Biol. 12, 4230-4241.
(Accepted 8 February 1994)