Embed Size (px)
Citation preview

The Role of the Mandible in
Mouse Palatal Development Revisited
RICHARD JELINEK, MUDr., C.Sce.
MIROSLAV PETERKA, MUDr.,Prague, Czechoslovakia
Growth changes associated with formation of the secondary palate were studied inH-Velaz mouse embryos by morphometric measurement of medial sagittal sections ofthe heads between days 13 and 15 of embryonic development. Horizontalization of thepalatine processes depends primarily on the length of the mandible and is effected by a shiftof palatine process material into the supralingual space. The administration of 7.5mg cortisone acetate im to the female on day 12, of 0.5 mg 6-aminonicotinamide imon day 14, or amniocentesis on day 14 significantly retards growth of the embryo ingeneral and of the mandible in particular without significantly affecting deflexion ofthe skull base. The different spatial conditions created by disproportion betweendeflexion of the base and the length of the mandible cause irregularities in thecourse of horizontalization which lead, as a rule, to cleft palate.
Introduction
Although a promising experimental model
of cleft palate (CP) was discovered 25 years ago(Baxter, Fraser, 1950), the development of CPis still veiled in mystery and obscurity. The
application of a whole battery of morphologicaland biochemical techniques from cutting with arazor to the use of radioisotopes has led to theconclusion that normal palatal development isthe outcome of synchronized interaction be-
tween the processes, the tongue, the mandible,
and the cranium (Green and Kochhar, 1975).This explains why the instant of horizontaliza-
tion of the palatine processes is normally dis-tributed (Fraser, 1969) even in a highly stand-
ardized embryo population (Jelinek, Dostal,1972). However, it does not provide a causal
explanation of the action of teratogens whichproduce CP or a theoretical foundation for therational prevention of CP in man.
Since any systematic study of a morphoge-netic process should begin with an orientative
morphometric analysis, continue with an analy-
sis of elementary morphogenetic processes(Jelinek, 1973), and then go to the heart of thephenomena at molecular biology and biochemi-cal levels, it was decided some years ago toinvestigate the development of the palate alongthese lines. We succeeded in demonstrating
MUDr Richard Jelinek and MUDr Miroslav Pe-terka are affiliated with the Institute of ExperimentalMedicine CSAV, Teratology Department, Legerova61, 120 00 Praha 2, Czechoslovakia
211
that delayed horizontalization is not essential toCP formation (Dostal, Jelinek, 1974) eventhough it usually accompanies it. We also dem-onstrated that non-closure of the gap betweenthe palatine processes was associated with hypo-plasia as a result of inhibited proliferation.
Mitotic activity in the palatine process mesen-chyme of H-Velaz mouse embryos culminatesabout 24 hours prior to horizontalization (Jeli-nek, Dostal, 1974), and corticoids injected in-
traamniotically or administered via the mater-nal organism shear off this proliferation apex
(Jelinek, Dostal, 1975). This formation of CP onthe basis of hypoplasia of the palatine processesis only one case, and by no means a typical one,in a whole range of mechanisms. It is common
knowledge, and was demonstrated experimen-
tally long ago (Walker, Fraser, 1957), that CP
formation is usually connected with delayed on-set or a prolonged course of horizontalization.Data obtained by measuring proliferative activ-
ity show that the interval between the phase ofmaximum growth of the palatine processes andmaxilla and the instant of horizontalization istoo long for the two phenomena to be causallyassociated (Jelinek, Dostal, 1974, 1975). Fur-thermore, CP is readily induced by certaingrowth inhibitors (e.g. 6-azauridine - Dostal,
Jelinek, 1974) even after the phase of peak pro-liferation in the palatine processes. The maincause of CP formation after corticoid adminis-tration, which is evidently connected with im-pairment of the horizontalization mechanism,is still unknown (Greene, Kochhar, 1975). Us-

212
ing simple morphometry, we therefore at-
tempted a fresh revision of the marked growthchanges which take place about the time ofhorizontalization and verified the results of astudy of normal material in three etiologically
different experimental situations. Our aim wasto find the main factor responsible for the
change in the position of the palatine processes,although we fully realized that the horizontali-
zation of these processes is a classic example ofa multifactorially determined process. _
Materials and Method
All measurements and experiments wereperformed on random-bred H-Velaz albinomice. Females (mean weight 32.3 g) were matedover night, and the day a vaginal plug wasfound was counted as day one of pregnancy.
After killing the females by cervical dislocation,
embryos were collected at stages 14/12 (12thhour of day 14), 14/24, 15/12, and 15/24. After
freeing them from the uterus and the foetalmembranes, the embryos were weighed, di-vided into weight classes (25 mg intervals), andimmediately placed in Bouin-Hollande fixing
fluid, where they were kept for a week.They were then carefully decapitated, and
using a sharp safety razor blade set in a glassholder, the heads were cut vertically in the linerunning through the central canal of the spinalcord and the middle of the upper lip. Theplane and course of the cut were controlledunder a dissecting microscope. Fixation with a
pin inserted through the postero-lateral headregion, apropriate adjustment with a pair ofpincers, and advantageous properties of the
TABLE 1. Materials.
Cleft Palate Journal, July 1977, Vol. 14 No. 3
Bouin-Hollande-treated fetal tissues greatly re-duced distortion of the section plane. More-over, the slices were carefully checked for ir-regularities, and those in which the hypophy-seal stalk, symphysis of Meckel's cartilage, andthe vertebral canal were not clearly visible, wereeliminated. In this way, the possible artificialmovement between the upper and lower facialregions was kept within the limits of systematicerror. The sagittal surface of the cut wasstained with Mayer's haematoxylin, and weagain checked its position and angle to exclude
any irregularities.The findings were evaluated in drawings
made under a Wild (Heerbrugg) projecting de-
vice at 13-fold total enlargement. This tech-nique was used both for the control group andfor the three experimental groups of embryos.
In the first experimental group, the femaleswere injected im with 0.5 mg 6-aminonicotin-amide on day 14 and in the second group with
7.5 mg cortisone acetate (also im) on day 12.These doses induced CP in about 50 per cent ofembryos. In the third experimental group,
about half of the embryos of each pregnantfemale were deprived of amniotic fluid by am-
niocentesis on day 14. Data on the amount ofmaterial in the various groups are given in Ta-ble 1. For statistical evaluation, we used routineparametric and nonparametric methods, whichwill be mentioned separately in the relevant
paragraphs.
Results
A) RELATIONSHIPS IN NoRMAL EmBR¥yos. In
sagittal sections of the heads, we chose four
meanfetal weight number of
q ___, number 77mm of resorptionstreatment ay of sacrifice offemales iving fe- _ and auto-
tuses X S |yses
6-AN day 14 14/12 3 27 203.7 16.5 114/24 4 47 239.2 26.8 215/12 4 33 277.1 24.7 015/24 4 47 333.1 31.3 2
Cortisone acetate day 12 14/24 2 22 217.2 27.5 115/12 2 26 298.3 18.4 215/24 2 22 365.0 53.2 3
Amniocentesis day 14 14/12 1 13 166.5 13.2 014/24 6 16 174.6 21.6 1315/12 3 9 270.0 12.8 1015/24 5 7 317.6 21.7 19
Littermates to amniocentesis 14/24 28 185.9 20.4 1215/12 16 265.5 21.4 515/24 12 323.6 30.4 16
Controls 14/12 5 51 211.1 20.6 114/24 5 52 252.0 27.8 115/12 5 59 332 4 37.1 115/24 3 34 382.5 42.0 0

Jelinek and Peterka, PALATAL DEVELOPMENT REVISITED
structures, clearly distinguishable after stainingwith haematoxylin, which furnished good basicinformation on growth changes. They were thepituitary or, to be more exact, the division be-tween the neurohypophysis and the infundibu-lar (Figure 1 - 1), the arcus anterior atlantis (2),
the hyoid (3), and the apex of Meckel's cartilage
(4). The lines joining the nearest facing pointsof these structures form two triangles with acommon side, I - 3 (pituitary-hyoid).
Measurement of all these distances in the
various embryo weight categories showed thatthe period in which horizontalization of the
palatine processes takes place is characterizedby a marked change in the more ventral of thetwo triangles, while the only change in the dor-sal triangle is slight widening of the base (2 - 3).
The pronounced change in the ventral triangleis caused by a rapid shift of the apex of Meckel'scartilage away from the pituitary-hyoid line,resulting chiefly in an increase in distances I -#(pituitary - Meckel) (Figure 1, 3 - 4, hyoid -Meckel) and (Figure 2, A, horizontal component
of change). This is the first of three strikingphenomena which precede horizontalization ofthe palatine processes.
The second event is growth of the base of theskull, usually described as deflection. In fact, itsgrowth is oriented primarily in a ventrocranial
direction, and one of the results is marked ele-vation of the snout in relation to the basal hori-zontal (Figure 1,2 - 3, atlas - hyoid) and (Figure1 and Figure 2 v, vertical component of change.
The third striking phenomenon is the for-mation of a relatively large space between thedorsum of the tongue and the roof of the prim-
itive oral cavity (Figure 3). The space makes its
.
FIGURE 1. Morphometric points on medial sagittal sec-tion of head of mouse fetus and mean difference betweendistances measured at beginning and end of observationperiod (i.e. between weight categories 151-175 mg and 376-
. 400 mg) in projection millimetres (mm x 13). Point 1: pitui-tary, point 2: arcus anterior atlantis, point 3: hyoid, point 4:Meckel's cartilage, point 5: tip of tongue. Distance v is de-fined as the position of the inferior margin of the philtrumrelation to the basal horizontal 2-3. At the beginning of thegiven observation period, it has negative value.
213
FIGURE 2. Change in configuration of head of mousefetus at end of observation period (stippled area) comparedwith initial state (white area). Main growth changes distrib-uted over vertical (v - deflexion of base) and horizontal (h -growth of mandible) components. Both projections of headoriented to a common point (arcus anterior atlantis) andbasal horizontal (2 - 3, i.e. atlas - hyoid).
My ~- /y bap
FIGURE 3. Formation of space between dorsum oftongue and roof of primitive oral cavity (space denoted byarrows). Projection of palatine process indicated by stippling;projection of position of Meckel's cartilage illustrated in stage14/24.
first appearance dorsally in the area where thepalatine processes are situated above the
tongue from the very beginning. It then growssteadily higher and spreads forward. As thepalatine processes are horizontalized, they pro-trude into the space and, as they flatten the
dorsum of the tongue, suddenly enlarge it. Inthe graph illustrating its height, this is mani-
fested in distinct discontinuity (Figure 5 and 6).Other findings also emerged: 1) Microdissec-
tion demonstrated that line 1 - 4 (pituitary -Meckel), which crosses the developing spaceobliquely in a ventro-caudal direction, corre-sponds to the course of Meckel's cartilage (Fig-ure 3 - 14/24) and can thus be regarded as a
reliable index of growth of the mandible. 2)The initial relationship between the inferiormargin of the palatine processes and the dor-sum of the tongue precludes horizontalization
of the processes without their conformation(Figure 3). Evidence of conformation of theprocesses is provided by the progressive de-crease in the height of their lateral projection,
which, just before horizontalization, is mani-fested in elevation of the projection of thelower margin of the processes to the level of
the dorsum of the tongue. 3) Extension of the

214 Cleft Palate Journal, July 1977, Vol. 14 No.
prom Ix \//so //\// _
r= 0%942 “vi”
o/{o
“J f}...-
o
oo /+..oo,‘/oo
30 o,ea/ .o
/o/oo
20
T4 _____A_ A_ ,
0 20 30 40 S0pum
FIGURE 4. Correlation between increase in distance 1 - 4
(pituitary - Meckel's cartilage; on abscissa) and distance 1 - 5
(pituitary - tip of tongue ; on ordinate). The high correlation
coefficient and the distribution of the points show that exten-
sion of the tongue is a function of growth of Meckel's carti-
lage. Pmm - projection millimetres (mm X 13); beginning
and maximum of horizontalization of palatine processes de-
noted by white triangles.
tongue closely follows the growth of Meckel's
cartilage (Figure 4). Line I - 4 (pituitary - Mec-
kel) runs, throughout the observation period,
approximately down the centre of the fan of
highly differentiated fibres of the genioglossus
muscle, which radiates into the dorsum of the
tongue precisely in the area where the space
appears (Figure 3).
B) HyroTruEsis or THE HoRIZONTALIZATION
MrcHanIsm. On the basis of the above find-
ings, we formulated the hypothesis that, during
horizontalization, the palatine processes shift
into a pre-prepared space produced by exten-
sion of the tongue in association with growth of
the mandible. The time and manner of hori-
zontalization are the outcomes of interaction
between growth of the base of the skull, the
volume of maxillary mesenchymal material,
and the activity of the glossomandibular com-
plex.
C) CORRELATION OF THE HricHtT or THE SU-
PRALINGUAL SPACE To THE LENGTH OF THE
MaANDIBLE AND GROWTH oF THE BAsE (Figures 5
and 6). The height of the space increases lin-
early with the increase in distance 1 - 4 to a
mean value of 2 pmm*, which corresponds to a
mandibular length index value of 33-34 pmm.
At this point the course of the curve is broken
owing to penetration of the palatine processes
into the space, which is suddenly raised by 4
pmm. The curve continues from this point with
about the same trend. The height of the space
in embryos with unilaterally horizontalized or
* pmm = projections millimetres (mm X 13)
still separated processes forms a short, discon-
tinuous transition. Figure 5 demonstrates the
positive and very definitive relationship be-
tween the length of the mandible, the size of
the space, and horizontalization of the palatine
processes. It also shows that the space exists
before it is filled by the processes.
The correlation of horizontalization to
growth of the base, the index of which is rela-
tionship of the inferior margin of the philtrum
to the basal horizontal, 2 - 3, (Figure 3), is far
less unequivocal (Figure 6). The space is
formed while the lower margin of the snout still
lies well below the basal horizontal, and hori-
zontalization of the palatine processes begins at
values of between -2 and =-1 pmm. Disconti-
nuity is not of the point type, and the curves for
the height of the space in embryos with hori-
zontalized and non horizontalized processes
overlap over the relatively wide range of -2 to
+3 pmm. The relationship between growth of
the mandible and the instant of horizontaliza-
pmm l
8
ro [ _®H +
6 1o
4 -
2 U m-
IL-f- r r v B r -
0 19 23 27 31 35 39 43 pmm
FIGURE 5. Increase in height of supralingual space (on
ordinate) in correlation to value of mandibular length index
(1 - 4; on abscissa). H-: height of space before commence-
ment of horizontalization of palatine processes, H+: height
of space after completion of horizontalization. Intermediary
line: height of space in embryos during horizontalization. s -
standard deviation, pmm - projection millimetres (mm x 13).
Height of space defined as distance between lines parallel
with basal horizontal, one of which touches convexity of
dorsum of tongue and other roof of primitive oral cavity.
pmm
8 {
e 100 OEE S ,
- 8 » o0000Q 0000-o4 00008
pmm
FIGURE 6. Increase in height of supralingual space (on
ordinate) in correlation to deflexion of base of skull, evalu-
ated from distance of inferior margin of philtrum from basal
horizontal (2 - 3, on abscissa). Details as Figure 5.
Jelinek and Peterka, PALATAL DEVELOPMENT REVISITED
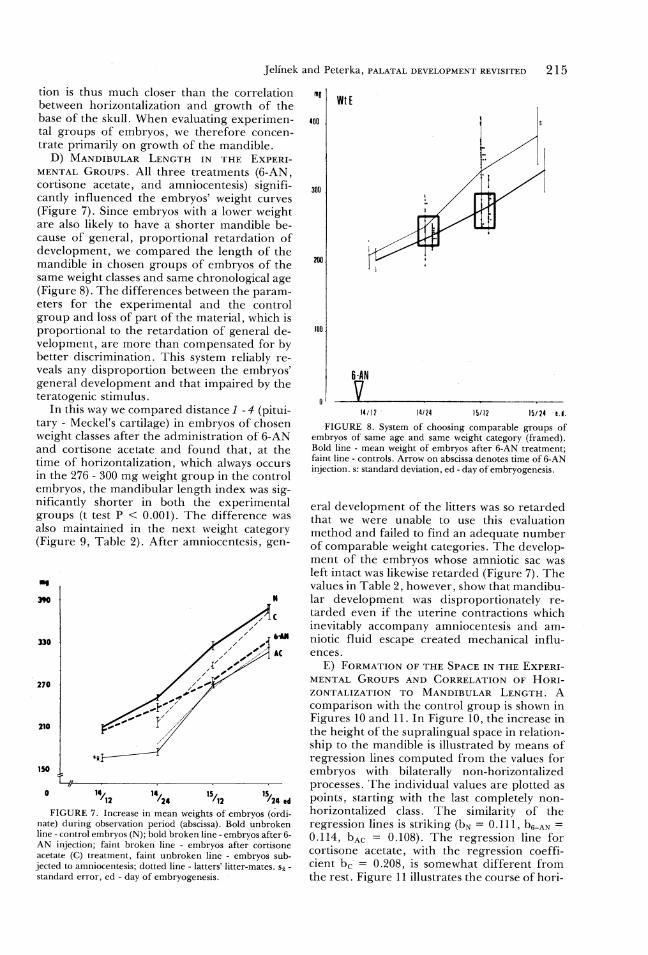
tion is thus much closer than the correlationbetween horizontalization and growth of thebase of the skull. When evaluating experimen-tal groups of embryos, we therefore concen-
trate primarily on growth of the mandible.D) MANDIBULAR LENGTH IN THE EXPERI-
MENTAL Groups. All three treatments (6-AN,cortisone acetate, and amniocentesis) signifi-cantly influenced the embryos' weight curves(Figure 7). Since embryos with a lower weightare also likely to have a shorter mandible be-cause of general, proportional retardation ofdevelopment, we compared the length of themandible in chosen groups of embryos of the
same weight classes and same chronological age(Figure 8). The differences between the param-
eters for the experimental and the controlgroup and loss of part of the material, which isproportional to the retardation of general de-velopment, are more than compensated for by
better discrimination. This system reliably re-veals any disproportion between the embryos'
general development and that impaired by theteratogenic stimulus.
In this way we compared distance I - 4 (pitui-tary - Meckel's cartilage) in embryos of chosenweight classes after the administration of 6-ANand cortisone acetate and found that, at thetime of horizontalization, which always occursin the 276 - 300 mg weight group in the control
embryos, the mandibular length index was sig-nificantly shorter in both the experimentalgroups (t test P < 0.001). The difference was
also maintained in the next weight category(Figure 9, Table 2). After amniocentesis, gen-
330
270
210
150 |
I T T v a
° "/p "/ze "/> "/2n edFIGURE 7. Increase in mean weights of embryos (ordi-
nate) during observation period (abscissa). Bold unbrokenline - control embryos (N); bold broken line - embryos after 6-AN injection; faint broken line - embryos after cortisoneacetate (C) treatment, faint unbroken line - embryos sub-jected to amniocentesis; dotted line - latters' litter-mates. S5 -standard error, ed - day of embryogenesis.
215
WtE
400
300
100 .
6 -AN
14/12 ° 14724 1§/12 15724 e.
FIGURE 8. System of choosing comparable groups ofembryos of same age and same weight category (framed).Bold line - mean weight of embryos after 6-AN treatment;faint line - controls. Arrow on abscissa denotes time of 6-ANinjection. s: standard deviation, ed - day of embryogenesis.
eral development of the litters was so retardedthat we were unable to use this evaluationmethod and failed to find an adequate numberof comparable weight categories. The develop-
ment of the embryos whose amniotic sac wasleft intact was likewise retarded (Figure 7). Thevalues in Table 2, however, show that mandibu-lar development was disproportionately re-tarded even if the uterine contractions which
inevitably accompany amniocentesis and am-niotic fluid escape created mechanical influ-ences. ‘
E) FORMATION OF THE SPACE IN THE EXPERI-MENTAL GROUPS AND CORRELATION OF Hor-ZONTALIZATION To MANDIBULAR LENGTH. Acomparison with the control group is shown inFigures 10 and 11. In Figure 10, the increase inthe height of the supralingual space in relation-ship to the mandible is illustrated by means ofregression lines computed from the values forembryos with bilaterally. non-horizontalizedprocesses. The individual values are plottedaspoints, starting with the last completely non-horizontalized class. The similarity of theregression lines is striking (by = 0.111, bean =0.114, bac = 0.108). The regression line forcortisone acetate, with the regression coeffi-cient be = 0.208, is somewhat different fromthe rest. Figure 11 illustrates the course of hori-

216
AN
20 -
100%--_2==--
-20 |
\\m /225\" /21s\ , /ns\ /37$mq
/12 /a "A2 "ae
FIGURE 9. Relative values of mandibular length (1 - 4)index for individual embryos after injecting 6-AN (above)and cortisone acetate (below), expresses as percentage ofmean control values in comparable groups (for manner ofchoice see Figure 8). The zone round 100% level denotes theconfidence interval +s for the control group. Black circles:embryos before horizontalization of palatine processes, whitecircles: embryos with incompletely horizontalized processes,crossed black circles: embryos with completed horizontaliza-tion. A 6-pointed asterisk denotes a statistically significantdifference at the 1% level of confidence; a 5-pointed asteriska significant difference at the 5% level of confidence.
175
zontalization in the experimental groups as
compared to the normal in relationship to man-
dibular length.
The distribution curves for embryos with
processes undergoing horizontalization (in-
cluding unilateral) were computed by retro-
grade subtraction of the cumulative frequencies
estimated from the mean number of embryos
with horizontalized and non-horizontalized
processes for each pair of pmm mandibular
length (Figure 10). The course of horizontaliza-
tion after cortisone acetate again differs sharply
from the rest by its two-peak maximum,
whereas discontinuities in the control and am-
niocentesis groups are the result of the small
amount of material. The distribution of em-
bryos in which horizontalization is taking place
is in every case within satisfactory mandibular
length index limits (about 30 pmm) except for
the second cortisone acetate maximum. Back-
conversion to general development parameters
shows no significant differences in the mean
weight of embryos in which horizontalization is
taking place, whereas the mean chronological
age of the experimental embryos is distinctly
higher.
Cleft Palate Journal, July 1977, Vol. 14 No. 3
Discussion
The immediate factor in horizontalization of
the palatine processes is extension of the
tongue, caused by growth of Meckel's cartilage.
Delayed growth of the mandible in the experi-
mental groups is part of the retardation of
growth responsible for the anomalous course of
weight curves of these embryos, although it
does not involve growth of the base of the skull.
The course of elevation of the snout in relation
to the basal horizontal shows no statistically sig-
nificant difference between any of the groups
when related to the chronological age of the
embryos, whereas growth of the mandible in
the experimental groups is significantly re-
tarded precisely in the crucial phase of normal
horizontalization (Figure 12). The chief differ-
ence in the experimental embryos compared
with the controls is that the mandible does not
attain the critical length necessary for extension
of the tongue and origination of the supralin-
a | Nbp 4
6o ** %
4 4q e
_§2 ~~~o -__Sin -
20 30 40 pmpmm ~
8 "AN
6& 1 ‘ @
P3 4 <e. ;* Spe *-e zoo. © e
t ..o-o—-..—“:’
J ==--_ *e - <9 r -r20 30 40 pmm
MM
a C+@+
oa e o * a *
F1 .- .~"f.
© (IR /”,,o1
///// baso T Ls ¥ °C ¥"
130 30 40 pmm
pmm
& AC
zfl’ @6 4 co
e 0. 0
a 20s
R P3 o eo _L
_---*yt ~"0 - --- a v ~~
20 30 40 pmm
FIGURE 10. Increase in height of supralingual space
(ordinate) in correlation to mandibular length (1 - 4) index
(abscissa). Regression lines computed from values before
horizontalization of palatine processes started. (Only the last
group with 100% non-horizontalization is illustrated - black
circles). White circles - values for embryos with incomplete
horizontalization, crossed black circles - embryos with com-
pleted horizontalization. N - control group; 6-AN - values
after injection of 6-AN; C -values after injection of cortisone
acetate; AC - values for all embryos of females subjected to
amniocentesis; pmm - projection millimetres (mm

TABLE
2.Mean
values
fordistance
pituitary-Meckel's
cartilage
asin
dica
torof
mandibulargrowth.
weig
ht150
175
clas
smg
stag
e
6-AN
1021
.312
14/1
2C AC
819.1
0.6
519
.61.5
o" b~ 6"
«O-
1C) CO& m-N O
O if)
&raC
r
6-AN
117
.014/24
C.
..
AC
619
.31.6
420
.00.8
15/1
2C
6-AN
15/24
C AC
N=
norm
alcontrolgr
oup.
6-AN=
group
trea
tedwi
th6-
amin
onic
otin
amid
e.C=
grou
ptreatedwi
thco
rtis
one
acet
ate.
AC=
grou
psubmitted
toamniocentesis.
200
| ig
22A
22.3
23.7
o «om/ rd
2235
lm
25.0
25.0
28.6
25.8
25.1
25.0
2.0 Co=< r= 2.7
250
bad
29.8
27.5
27.0
32.0
28.3
32.0
27.3
oo nnre- or- & I~m- /m- 1.6
275
likes
32.1
28.0
34.0
31.0
30.1
28.0
3.1
0.8
1.7
1.6
1.6
J <
300
as
36.6
33.5
32.9
36.0
33.9
36.0
29.0
1.6
13 in CJ 66 <G
325
las36.3
37.6
34.1
38.0
36.8
1.4
0.9
2.0
1.5
350
'*
38.7
38.0
38.3
38.6
0.8
r- aC)
375
400
ip
likes
38.0
38.0
36.8
3.8
38.7
0.8
218
Cleft Palate Journal, July 1977, Vol. 14 No. 3
o,A
20(Cal A __
; i40 pmm
o 20 war "° oas;s
%% 245 me 283 mg
100 ‘.AN
20
---
0 30 ao‘ 40 pmm18/2e
oA 315 mg
100 C
20
P r r
0 20
%
100 AC
30
re - por-} _
© 20 30 40 pmm18/1o 16] 3
298 mq 334 mg
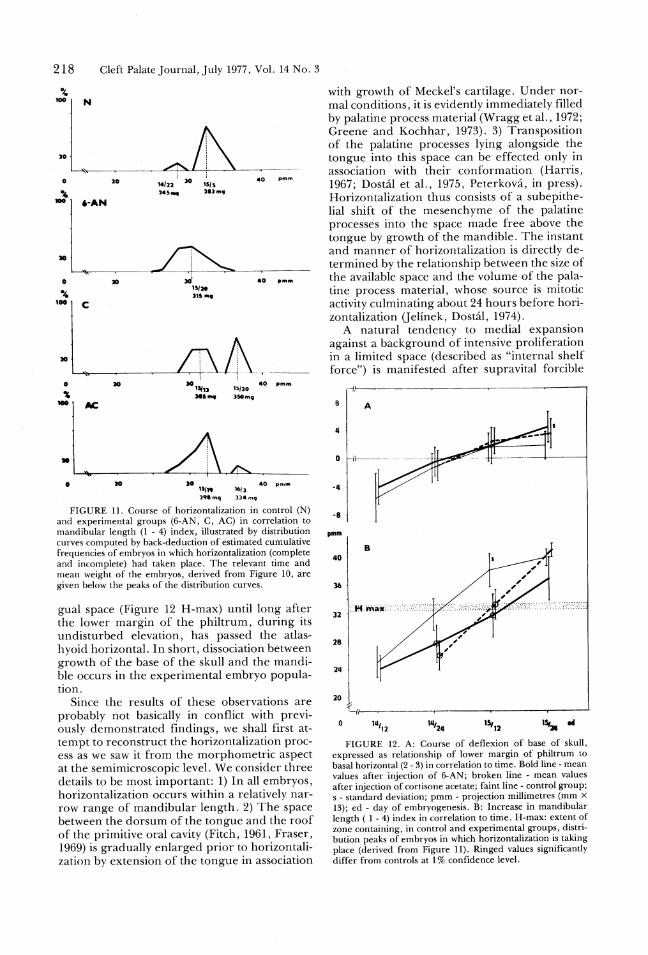
FIGURE 11. Course of horizontalization in control (N)
and experimental groups (6-AN, C, AC) in correlation to
mandibular length (1 - 4) index, illustrated by distribution
curves computed by back-deduction of estimated cumulative
frequencies of embryos in which horizontalization (complete
and incomplete) had taken place. The relevant time and
mean weight of the embryos, derived from Figure 10, are
given below the peaks of the distribution curves.
gual space (Figure 12 H-max) until long after
the lower margin of the philtrum, during its
undisturbed elevation, has passed the atlas-
hyoid horizontal. In short, dissociation between
growth of the base of the skull and the mandi-
ble occurs in the experimental embryo popula-
tion.
Since the results of these observations are
probably not basically in conflict with previ-
ously demonstrated findings, we shall first at-
tempt to reconstruct the horizontalization proc-
ess as we saw it from the morphometric aspect
at the semimicroscopic level. We consider three
details to be most important: 1) In all embryos,
horizontalization occurs within a relatively nar-
row range of mandibular length. 2) The space
between the dorsum of the tongue and the roof
of the primitive oral cavity (Fitch, 1961, Fraser,
1969) is gradually enlarged prior to horizontali-
zation by extension of the tongue in association
with growth of Meckel's cartilage. Under nor-
mal conditions, it is evidently immediately filled
by palatine process material (Wragg et al., 1972;
Greene and Kochhar, 1973). 3) Transposition
of the palatine processes lying alongside the
tongue into this space can be effected only in
association with their conformation (Harris,
1967; Dostal et al., 1975, Peterkova, in press).
Horizontalization thus consists of a subepithe-
lial shift of the mesenchyme of the palatine
processes into the space made free above the
tongue by growth of the mandible. The instant
and manner of horizontalization is directly de-
termined by the relationship between the size of
the available space and the volume of the pala-
tine process material, whose source is mitotic
activity culminating about 24 hours before hori-
zontalization (Jelinek, Dostal, 1974).
A natural tendency to medial expansion
against a background of intensive proliferation
in a limited space (described as "internal shelf
force") is manifested after supravital forcible
JL11
pmm
40
32
28
24
20h
AN
|N _L =e
0 Ill/12 15,12 IS,” ed
FIGURE 12. A: Course of deflexion of base of skull,
expressed as relationship of lower margin of philtrum to
basal horizontal (2 - 3) in correlation to time. Bold line - mean
values after injection of 6-AN; broken line - mean values
after injection of cortisone acetate; faint line - control group;
s - standard deviation; pmm - projection millimetres (mm %
13); ed - day of embryogenesis. B: Increase in mandibular
length ( 1 - 4) index in correlation to time. H-max: extent of
zone containing, in control and experimental groups, distri-
bution peaks of embryos in which horizontalization is taking
place (derived from Figure 11). Ringed values significantly
differ from controls at 1% confidence level.
14,24

Jelinek and Peterka, PALATAL DEVELOPMENT REVISITED
extension of the tongue, in "lifting" of the pala-tine processes into a neutral position about halfway to the horizontal (Fitch, 1961; Ross andWalker, 1967). Under normal conditions, i.e. ifthe space is sufficiently large (if the mandible issufficiently long) and there is enough material,transposition of both palatine processes is ef-fected in a single instant (Figure 5). If the spaceis small in relation to the size of the palatine
processes (a short mandible), horizontalizationis asymmetrical. Only the palatine processespenetrate into the small space in the anterior
part of the oral cavity, thereby causing devia-tion of the tongue, which presses against the
contralateral, non-horizontalized process andprevents its mediocranial transposition. We en-
counter this picture in a normal population ofindividuals with prematurely horizontalizedprocesses, in the group of embryos treated with
6-AN (the shift to the left in distribution of the
course of horizontalization in Figure 11), inwhich administration of the teratogen on day 14did not influence the main proliferation phasein the palatal mesenchyme, and in the group ofembryos treated with cortisone acetate. In thislast group, the situation is complicated by thefact that, when administered on day 12, corti-sone acetate "erases" the peak of mitotic activityin the palatine processes at stage 13/24 (Jelinek,Dostal, 1975); and, as demonstrated in part ofthe population, it has a negative effect on the
growth of the mandible. Hypoplasia of themaxillary mesenchyme sharply reduces medialexpansion, and in some of the embryos this ismanifested by failure of the processes to behorizontalized even when the supralingual
space is more than adequate (the regression line
with the larger angle in Figure 10, the secondhorizontalization maximum in Figure 11).
Asymmetrical horizontalization is an unde-sirable phenomenon, which at best leads to the
prolongation of bilateral movement of the pala-tine processes and to delayed closure of thepalate. In a normal embryo population, thisstate is rectified with intensive prolongation ofthe mandible. In embryos treated with 6-AN or
cortisone acetate, however, the state is perma-nent (Peterka and Dostal, 1977), and differen-tiation of the preossification blastema in the
palatine processes fixes it. In the group of em-bryos treated with 6-AN, we find yet anothermodification of the horizontalization process.In association with the relatively short mandi-ble, the dorsum of the the tongue is retained inthe posterior part of the space between thepalatine processes. With normal growth of the
base of the skull (manifested in continuous de-flexion), the space becomes free anteriorly and
is penetrated by the anterior part of the pala-
219
tine processes so that the normal posteroante-rior sequence of the horizontalization process isreversed.
Various authors have drawn attention to thelength of the mandible as a factor influencing
closure of the secondary palate. In accordancewith the Sicher's hypothesis (1915), Hart et al.(1969) pointed out that growth of the mandiblein mice spurted between 13 and 16 days with themaximum on day 14. Schwartz and Chaudhry
(1968) found micrognathia after administering
cortisone acetate, and Harris and Morgan(1969) regarded the length of Meckel's cartilageas an important factor in the development of
cleft palate. Poswillo (1966), Deuschle and War-kany (1956), and Asling et al. (1960) described ashort mandible in association with cleft palate inrat embryos. In homozygous embryos with ge-
netically determined cleft palate ("short headmutation"), Fitch (1961) demonstrated a shortmandible. Thesereports presented no evidenceconfirming the influence of mandibular lengthon the commencement of horizontalization,however. A study by Shih et al. (1974) wasintended to furnish objective data through aquantitative evaluation of the length of themandible. After the administration of 6-AN,cortisone acetate, and vitamin A, no differenceswere found in the length of the mandible at thetime of horizontalization. Shih's results cannotbe compared with ours, however, as the initialhyoid - Meckel distance which she used doesnot correspond to the course of Meckel's carti-lage and is probably not a reliable index since itsvalues increase in association with deflexion of
the base of the skull, which is closely followedby the mandible (Figure 13). An interesting
cephalometric study of developing orofacial
I
FIGURE 13. Apparent prolongation of mandible on eval-uating its length from distance 3 - 4 (hyoid - Meckel's carti-lage), during deflexion of base of skull manifested in eleva-tion of snout, which is passively followed by mandible. In thistheoretical case, the mandibular length (1 - 4, pituitary -Meckel's cartilage) index remains the same after rotation toposition 1 - 4.

220
structures in the rat was published by Diewert(1974) stressing the possible importance ofMeckel's cartilage and the median palatal proc-
ess in triggering the palatal-shelves elevation. Itseems that palatal development in mice is simi-lar to that in rats. Finally, it may be stated that
our conclusions, based on the action of exoge-nous teratogens, correspond to findings in micehomozygous for the chondrodysplasia gene
(Seegmiller, 1976) a further confirmation of thewell-known fact that both genetically condi-tioned and environmentally induced defectsfollow common developmental pathways.
Summary
Growth changes associated with formation of
the secondary palate were studied in H-Velazmouse embryos by morphometric measure-ment of medial sagittal sections of heads be-
tween days 13 and 15 of embryonic develop-ment. The marked change in the configurationof the head in this phase is due primarily togrowth of the base of the skull (manifested indeflection) and to intensivegrowth of the man-
dible, which causes extension of the tonguefrom the posterior part of the roof of the primi-
tive oral cavity. Horizontalization of the pala-tine processes depends primarily on the lengthof the mandible and is effected by a shift ofpalatine process material into the supralingualspace. The administration of 7.5 mg of corti-sone acetate im to the female on day 12, of 0.5mg 6-aminonicotinamide im onday 14, or ofamniocentesis on day 14 significantly retardsgrowth of the embryo in general and of the
mandible in particular without significantly af-
fecting deflexion of the base of the skull. Hori-zontalization again occurs after the critical
length of the mandible has beenattained, i.e.
eight to 15 hours later than in the controlgroup. The different spatial conditions createdby disproportion between deflexion of the base
and the length of the mandible cause irregular-ities in the course of the horizontalization whichlead, as a rule, to cleft palate. After the admin-istration of cortisone acetate, the horizontaliza-tion process is further complicated by depletion
of the palatal mesenchyme, based, as we dem-onstrated in an earlier study, on inhibition ofproliferative activity at the end of day 13.
References
AsLING, C. W., NELson, M. M., DouvenE®RrtTy, H. L.,WrrcHtT, H. V., Anp Evans, H. M., The develop-ment of cleft palate resulting from maternal peter-oylglutamic (folic) acid deficiency during the latterhalf of gestation in rats, Surg. Gyn. Obst., 111, 19-21, 1960.
BaxtTER, H., AND FRASER, F. C., Production of con-
Cleft Palate Journal, July 1977, Vol. 14 No. 3
genital defects in the offspring of female micetreated with cortisone, McGill Med. J., 19, 245-249, -1950.
DruscHLE, F. M., anp Warkany, J., Congenitaldentofacial malformation in rats induced by mater-nal nutritional deficiency, J. Dent. Res., 35, 674-684, 1956.
DIEwERT, V. M., A cephalometric study of orofacialstructures during secondary palate closure in therat, Archs. Oral Biol., 19, 3083-315, 1974.
DosT&L, M., aND JELINEK, R., Morphogenesis ofcleft palate induced by exogenous factors. VI. Thequestion of delayed palatal process horizontaliza-tion, Teratology, 10, 47-53, 1974.
DostTALr, M., PETERKOYA, R., AND JELiNEK, R., Hori-zontalization of the palatine processes in the mouseembryo, Folia Morph. (Prague), 23, 104-108, 1975.
FircH, N., Development of cleft palate in mice homo-zygous for the shorthead mutation, J. Morph., 109,151-157, 1961.
FRASER, F. C., Gene-environment interactions in theproduction of cleft palate. In "Methods For Tera-tological Studies In Experimental Animals AndMan" ed. Nishimura, H., and Miller, J. R., Tokyo,Shoin, 34-49, 1969.
GREENE, R. M., anp KocHmHar, D. M., Palatal closurein the mouse as demonstrated in frozen sections,Am. J. Anat., 137, 477-482, 1973.
GrrEnNr, R. M., anp Kocmmar, D. M., Some aspectof cortico-steroid-induced cleft palate: A review,Teratology, 11, 47-56, 1975.
HarRIs, J. W. S., Experimental studies on closureformation in the secondary palate,The
Scientific Basis of Medicine Ann. Rev., 356-370, 1967.Harris; J.W.:S., anp Morcan, P. R., Aetiology of
cleft palate induced by meclozine, buclizine andhydroxyzine, Excerpta Med. Int. Congr. Ser., 191, 47,1969 (Abstr.).
Hart, J. C., SmirEy, G. R., anp Drxon, A. D.,Sagittal growth of the craniofacial complex in nor-mal embryonic mice, Arch. Oral Biol., 14, 995-997,1969.
JriinExk, R., anp DostT&L, M., On the method ofevaluating the development of an embryonic com-ponent in experimental teratology, Folia Morph.(Prague), 20, 130-133, 1972.
JELiNEK, R., Contribution of embryological princi-ples to teratology, Acta Univ. Carolinae Med., Mono-graph., 56-57, 17-24, 1973.
R., anp DosTAL, M., Morphogenesis of cleftplate induced by exogenous factors. VII. Mitoticactivity during formation of the mouse secondarypalate, Folia Morph. (Prague), 22, 94-101, 1974.
JELiNEK, R., Anp DosTAL, M., Inhibitory effect ofcorticoids on the proliferative pattern in mousepalatal processes, Teratology, 11, 193-198, 1975.
PrTERKA, M., aAnp DostTAL, M., Influence of cleftpalate on growth of the maxilla in mouse embryos,Cleft Palate J., 14, 1977.
PrETERKOYVA, R., Positional changes of the palatineartery as an indicator of formation of the mousesecondary palate, Acta Univ. Carolinae Med., (inpress).
PoswILLO, D., Observations of fetal posture andcausal mechanisms of congenital deformity of pal- _

Jelinek and Peterka, PALATAL DEVELOPMENT REVISITED
ate, mandible and limbs, J. Dent. Res., Suppl. 3,45, 584-596, 1966.
Ross, L. M., AND WALKER, B. E., Movement of pala-tine shelves in untreated and teratogen treatedmouse embryos, Am. J. Anat., 121, 509-521, 1967.
ScHwaARTZz, D. M., anp CHnaupury, A. P., Planimet-ric studies of mandible in A/Jax mice born withcortisone-induced cleft palates, J. Dent. Res., 47,725-732, 1968.
SEEGMILLER, R. E.., Relationship between mandibulargrowth retardation and cleft palate in mice homo-zygous for the chondrodysplasia gene, Teratology,13, 5A (Abstract), 1976.
221
Sun, L. Y., TrasiEr, D. G., aAnD> FRASER, F. C.,Relation of mandible growth to palate closure inmice, Teratology, 9, 191-202, 1974.
SICHER, H., Entwickelung des sekundaeren Gaumensbeim Menschen, Anat. Anz., 47, 513-523, 545-562,1915.
WarkrErR®, B. E., AND FRASER, F. C., The embryologyof cortisone-induced cleft palate, J. Embryol. Exp.Morph., 5, 201-209, 1957. ‘
Wracc, L. E., DirwERT, V. M., anp KLEIN,; M.,Spatial relations in the oral cavity and the mecha-nism of secondary palate closure in the rat, Arch.Oral Biol., 17, 683-690, 1972.
![Traumatic bone cyst of mandible: a case series€¦ · the mandible; only a few cases in the condylar and anter-ior regions of the mandible have been reported [8, 9], whereas maxillary](https://img.pdfslide.net/doc/110x75/612a028722625b5ff82bcaf3/traumatic-bone-cyst-of-mandible-a-case-series-the-mandible-only-a-few-cases-in.jpg)