Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 92, pp. 112-116, January 1995Cell Biology
The Saccharomyces cerevisiae phosphatidylinositol-transferprotein effects a ligand-dependent inhibition of choline-phosphatecytidylyltransferase activityHENRY B. SKINNER*, TODD P. MCGEE*, CHRISTOPHER R. MCMASTERt, MICHELLE R. FRY*, ROBERT M. BELLt,AND VYTAS A. BANKAITIS*t*Department of Cell Biology, University of Alabama, Birmingham, AL 35294-0005; and tDepartment of Molecular Cancer Biology, Duke University,Durham, NC 27710
Communicated by Eugene P. Kennedy, Harvard Medical School, Boston, MA, September 12, 1994
ABSTRACT The Saccharomyces cerevisiae protein SEC14pis required for Golgi function and cell viability in vivo. Thisrequirement is obviated by mutations that specifically inac-tivate the CDP-choline pathway for phosphatidylcholine bio-synthesis. The biochemical basis for the in vivo relationshipbetween SEC14p function and the CDP-choline pathway hasremained obscure. We now report that SEC14p effects an invivo depression of CDP-choline pathway activity by inhibitingcholine-phosphate cytidylyltransferase (CCTase; EC2.7.7.15), the rate-determining enzyme of the CDP-cholinepathway. Moreover, this SEC14p-mediated inhibition ofCCTase was recapitulated in vitro and was saturable. Finally,whereas the SEC14p-dependent inhibition of CCTase in vitrowas markedly reduced under assay conditions that wereexpected to increase levels of phosphatidylinositol-boundSEC14p, assay conditions expected to increase levels of phos-phatidylcholine-bound SEC14p resulted in significant poten-tiation of CCTase inhibition. The collective data suggest thatthe phosphatidylcholine-bound form of SEC14p effects anessential repression of CDP-choline pathway activity in Golgimembranes by inhibiting CCTase and that the phospholipid-binding/exchange activity of SEC14p represents a mecha-nism by which the regulatory activity of SEC14p is itselfcontrolled.
The Saccharomyces cerevisiae protein SEC14p is a member ofa ubiquitous class of proteins, the glycerophospholipid (PL)-transfer proteins, that were identified in assays measuringprotein-catalyzed transfer of lipids between membrane bilay-ers in vitro (1-3). While there has been much speculation withregard to the in vivo involvement of such proteins as diffusiblecarriers of lipid in intracellular lipid trafficking and lipidsorting events (1, 2), the availability of secl4 mutant strains ofyeast has provided the first system with which the in vivofunction of a PL-transfer protein can be studied in detail.SEC14p is a phosphatidylinositol/phosphatidylcholine (PI/
PC)-transfer protein that localizes as a peripheral Golgimembrane protein and is required for both transport ofsecretory glycoproteins from a late Golgi compartment andcell viability (4-6). Penetrating clues as to the function ofSEC14p in vivo were obtained from genetic studies revealingthat the SEC14p requirement for Golgi secretory function andcell viability was bypassed by specific inactivation of theCDP-choline pathway for PC biosynthesis (refs. 7 and 8; Fig.1A) and from biochemical studies indicating that the primaryconsequence of SEC14p dysfunction in vivo is a specificCDP-choline pathway-driven increase in Golgi PC (9). Thisincrease is accounted for by one of two mechanisms: (i) thatSEC14p maintains a reduced Golgi membrane PC content by
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.
PC removal from Golgi membranes via PC transfer or (ii) thatSEC14p represses the CDP-choline pathway in yeast Golgimembranes via a PL-sensing mechanism that effects down-regulation of CDP-choline pathway in response to the PLcontent of yeast Golgi membranes (9).The sensor model makes two distinguishing predictions.
First, it posits that SEC14p specifically represses the activity ofat least one of the three structural enzymes of the CDP-cholinepathway. Second, it predicts that the inhibitor activity ofSEC14p is controlled by whether PI or PC is bound to SEC14p.We now present in vivo and in vitro data to indicate thatSEC14p depresses CDP-choline pathway activity by inhibitingCCTase, the rate-determining enzyme of the CDP-cholinepathway. The data further suggest that SEC14p-PC is theinhibitory agent and that the phospholipid-binding/exchangeactivity of SEC14p represents a mechanism by which theCCTase-directed regulatory activity of SEC14p is itself con-trolled.
MATERIALS AND METHODS
Yeast Strains, Media, and Transformation. S. cerevisiaestrains included the wild-type strain CTY525 [MA Ta, ura3-52,Ahis3-200, lys2-801, Atrpl, leu2, YEp(TRPl)], the SEC14p-overproducing strain CTY521 [MARTq, ura3-52, Ahis3-200,lys2-801, Atrpl, leu2, YEp(SEC14, TRPI), and the SEC14psP-overproducing strain CTY457 (MATa, ura3-52, Ahis3-200,lys2-801, secl4A1l::HIS3, YEp(SEC14sP, URA3). StrainsCTY541, CTY553, CTY522, and CTY762 are cki-284::HIS3cct:URA3, cptl::LEU2, and YCP(PGAL::CCT) derivatives ofCTY525, respectively. Strains CTY540, CTY552, CTY526,and CTY761 are cki-284:HIS3, cct::URA3, cptl::LEU2, andYCp(PGAL::CCT) derivatives of CTY521, respectively. Yeasttransformation, YPD and YPGal media, yeast minimal medium,minimal defined inositol- and choline-free base media, and otherchemical reagents have been described (9-12). When appropri-ate, minimal defined medium was supplemented with 1 mMinositol and/or choline.
[14C]Choline Labeling and Phospholipid Quantitation.Procedures for choline labeling, quantitation and normaliza-tion of 14C incorporation, phospholipid extraction, and thin-layer chromatography have been described (9, 13). [14C]Cho-line-labeled phospholipids were visualized by autoradiogra-phy, excised, and quantitated by scintillation counting. Formeasurement of bulk cellular phospholipids, the appropriateyeast strains were grown in the appropriate defined ethanol-
Abbreviations: PI, phosphatidylinositol; PC, phosphatidylcholine; PE,phosphatidylethanolamine; PS, phosphatidylserine; PL, glycerophos-pholipid; SEC14psP, Schizosaccharomyces pombe SEC14p; CCTase,choline-phosphate cytidylyltransferase; CKIase, choline kinase;CPTase, cholinephosphotransferase.tTowhom reprint requests should be addressed at: Department of CellBiology, University of Alabama, Birmingham, AL 35294-0005.
112
Proc. NatL Acad Sci USA 92 (1995) 113
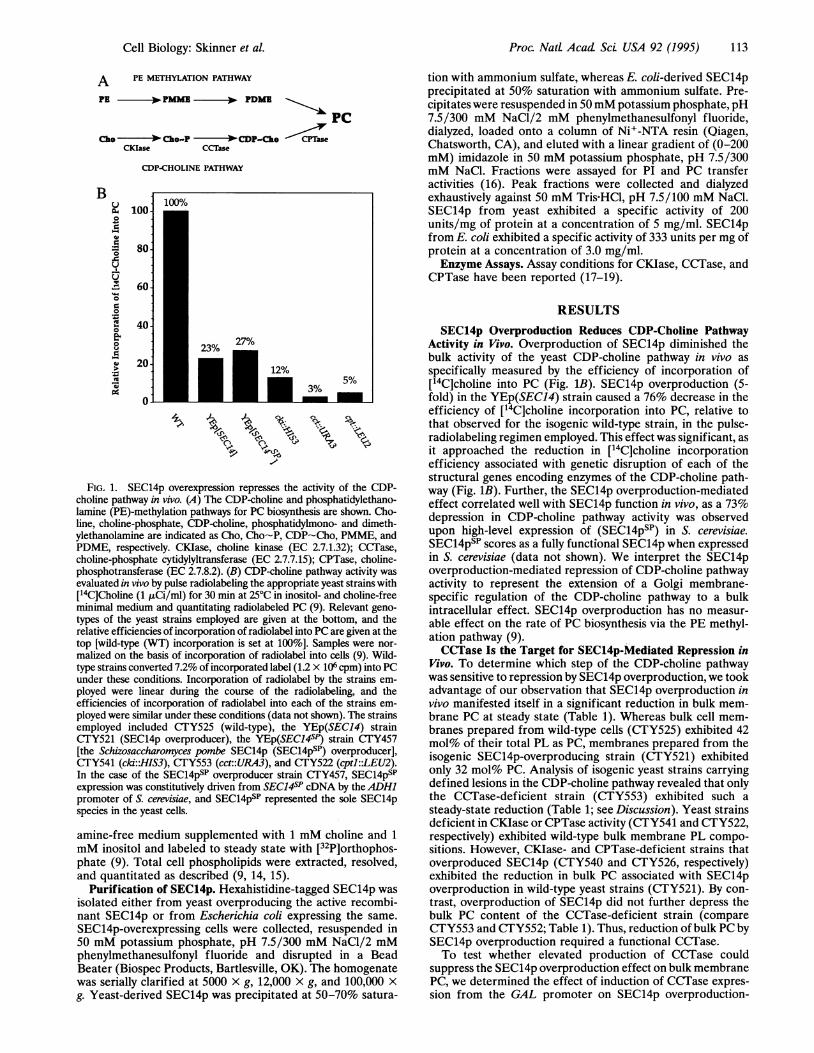
A PE METHYLATION PATHWAY
PE 00 PamIE PDMI
P
Cho ch.o-P CDP-cho CPTaseCKlase CCTase
CDP-CHOLINE PATHWAY
Bu
ODa._
-
0u1-
0
0
*.;a
uC:
100
5..
6-
FIG. 1. SEC14p overexpression represses the activity of the CDP-choline pathway in vivo. (A) The CDP-choline and phosphatidylethano-lamine (PE)-methylation pathways for PC biosynthesis are shown. Cho-line, choline-phosphate, CDP-choline, phosphatidylmono- and dimeth-ylethanolamine are indicated as Cho, Cho-P, CDP-Cho, PMME, andPDME, respectively. CKIase, choline kinase (EC 2.7.1.32); CCTase,choline-phosphate cytidylyltransferase (EC 2.7.7.15); CPTase, choline-phosphotransferase (EC 2.7.8.2). (B) CDP-choline pathway activity wasevaluated in vivo by pulse radiolabeling the appropriate yeast strains with[14C]Choline (1 ,uCi/ml) for 30 min at 25°C in inositol- and choline-freeminimal medium and quantitating radiolabeled PC (9). Relevant geno-
types of the yeast strains employed are given at the bottom, and therelative efficiencies of incorporation of radiolabel into PC are given at thetop [wild-type (WT) incorporation is set at 100%]. Samples were nor-
malized on the basis of incorporation of radiolabel into cells (9). Wild-type strains converted 7.2% of incorporated label (1.2 x 106 cpm) into PCunder these conditions. Incorporation of radiolabel by the strains em-
ployed were linear during the course of the radiolabeling, and theefficiencies of incorporation of radiolabel into each of the strains em-ployed were similar under these conditions (data not shown). The strainsemployed included CTY525 (wild-type), the YEp(SEC14) strainCTFY521 (SEC14p overproducer), the YEp(SEC14sP) strain CTY457[the Schizosaccharomyces pombe SEC14p (SEC14psP) overproducer],CTY541 (cki:.HIS3), CTY553 (cct::URA3), and CTY522 (cptl::LEU2).In the case of the SEC14psP overproducer strain CTY457, SEC14psPexpression was constitutively driven from SEC14SP cDNA by theADH1promoter of S. cerevisiae, and SEC14psP represented the sole SEC14pspecies in the yeast cells.
amine-free medium supplemented with 1 mM choline and 1mM inositol and labeled to steady state with [32P]orthophos-phate (9). Total cell phospholipids were extracted, resolved,and quantitated as described (9, 14, 15).
Purification of SEC14p. Hexahistidine-tagged SEC14p wasisolated either from yeast overproducing the active recombi-nant SEC14p or from Escherichia coli expressing the same.
SEC14p-overexpressing cells were collected, resuspended in50 mM potassium phosphate, pH 7.5/300 mM NaCl/2 mMphenylmethanesulfonyl fluoride and disrupted in a BeadBeater (Biospec Products, Bartlesville, OK). The homogenatewas serially clarified at 5000 x g, 12,000 x g, and 100,000 xg. Yeast-derived SEC14p was precipitated at 50-70% satura-
tion with ammonium sulfate, whereas E. coli-derived SEC14pprecipitated at 50% saturation with ammonium sulfate. Pre-cipitates were resuspended in 50mM potassium phosphate, pH7.5/300 mM NaCl/2 mM phenylmethanesulfonyl fluoride,dialyzed, loaded onto a column of Ni+-NTA resin (Qiagen,Chatsworth, CA), and eluted with a linear gradient of (0-200mM) imidazole in 50 mM potassium phosphate, pH 7.5/300mM NaCl. Fractions were assayed for PI and PC transferactivities (16). Peak fractions were collected and dialyzedexhaustively against 50 mM Tris-HCl, pH 7.5/100 mM NaCl.SEC14p from yeast exhibited a specific activity of 200units/mg of protein at a concentration of 5 mg/ml. SEC14pfrom E. coli exhibited a specific activity of 333 units per mg ofprotein at a concentration of 3.0 mg/ml.Enzyme Assays. Assay conditions for CKlase, CCTase, and
CPTase have been reported (17-19).
RESULTS
SEC14p Overproduction Reduces CDP-Choline PathwayActivity in Vivo. Overproduction of SEC14p diminished thebulk activity of the yeast CDP-choline pathway in vivo asspecifically measured by the efficiency of incorporation of[14C]choline into PC (Fig. 1B). SEC14p overproduction (5-fold) in the YEp(SEC14) strain caused a 76% decrease in theefficiency of [14C]choline incorporation into PC, relative tothat observed for the isogenic wild-type strain, in the pulse-radiolabeling regimen employed. This effect was significant, asit approached the reduction in [14C]choline incorporationefficiency associated with genetic disruption of each of thestructural genes encoding enzymes of the CDP-choline path-way (Fig. 1B). Further, the SEC14p overproduction-mediatedeffect correlated well with SEC14p function in vivo, as a 73%depression in CDP-choline pathway activity was observedupon high-level expression of (SEC14psP) in S. cerevisiae.SEC14psp scores as a fully functional SEC14p when expressedin S. cerevisiae (data not shown). We interpret the SEC14poverproduction-mediated repression of CDP-choline pathwayactivity to represent the extension of a Golgi membrane-specific regulation of the CDP-choline pathway to a bulkintracellular effect. SEC14p overproduction has no measur-able effect on the rate of PC biosynthesis via the PE methyl-ation pathway (9).CCTase Is the Target for SEC14p-Mediated Repression in
Vivo. To determine which step of the CDP-choline pathwaywas sensitive to repression by SEC14p overproduction, we tookadvantage of our observation that SEC14p overproduction invivo manifested itself in a significant reduction in bulk mem-brane PC at steady state (Table 1). Whereas bulk cell mem-branes prepared from wild-type cells (CTY525) exhibited 42mol% of their total PL as PC, membranes prepared from theisogenic SEC14p-overproducing strain (CTY521) exhibitedonly 32 mol% PC. Analysis of isogenic yeast strains carryingdefined lesions in the CDP-choline pathway revealed that onlythe CCTase-deficient strain (CTY553) exhibited such asteady-state reduction (Table 1; see Discussion). Yeast strainsdeficient in CKlase or CPTase activity (CTY541 and CTY522,respectively) exhibited wild-type bulk membrane PL compo-sitions. However, CKIase- and CPTase-deficient strains thatoverproduced SEC14p (CTY540 and CTY526, respectively)exhibited the reduction in bulk PC associated with SEC14poverproduction in wild-type yeast strains (CTY521). By con-trast, overproduction of SEC14p did not further depress thebulk PC content of the CCTase-deficient strain (compareCTY553 and CTY552; Table 1). Thus, reduction of bulk PC bySEC14p overproduction required a functional CCTase.To test whether elevated production of CCTase could
suppress the SEC14p overproduction effect on bulk membranePC, we determined the effect of induction of CCTase expres-sion from the GAL promoter on SEC14p overproduction-
Cell Biology: Skinner et al.
114 Cell Biology: Skinner et al.
mediated reduction in bulk membrane PC. The SEC14poverproduction effect was apparent in yeast strains grown ingalactose-containing medium (Table 1). While the wild-typestrain (CTY525) exhibited 44 mol% of total PL as PC, theisogenic SEC14p-overproducing strain exhibited only 33 mol%PC. Expression of CCTase from the GAL promoter (2.5-foldoverproduction), while not affecting bulk membrane PC levelsin the wild-type strain (CTY762), completely suppressed theSEC14p overproduction-mediated reduction in bulk mem-brane PC (strain CTY761). These collective data identifiedCCTase as the enzyme through which SEC14p effected therepression of CDP-choline pathway activity in vivo.SEC14p-Mediated Inhibition of CCTase Is Recapitulated in
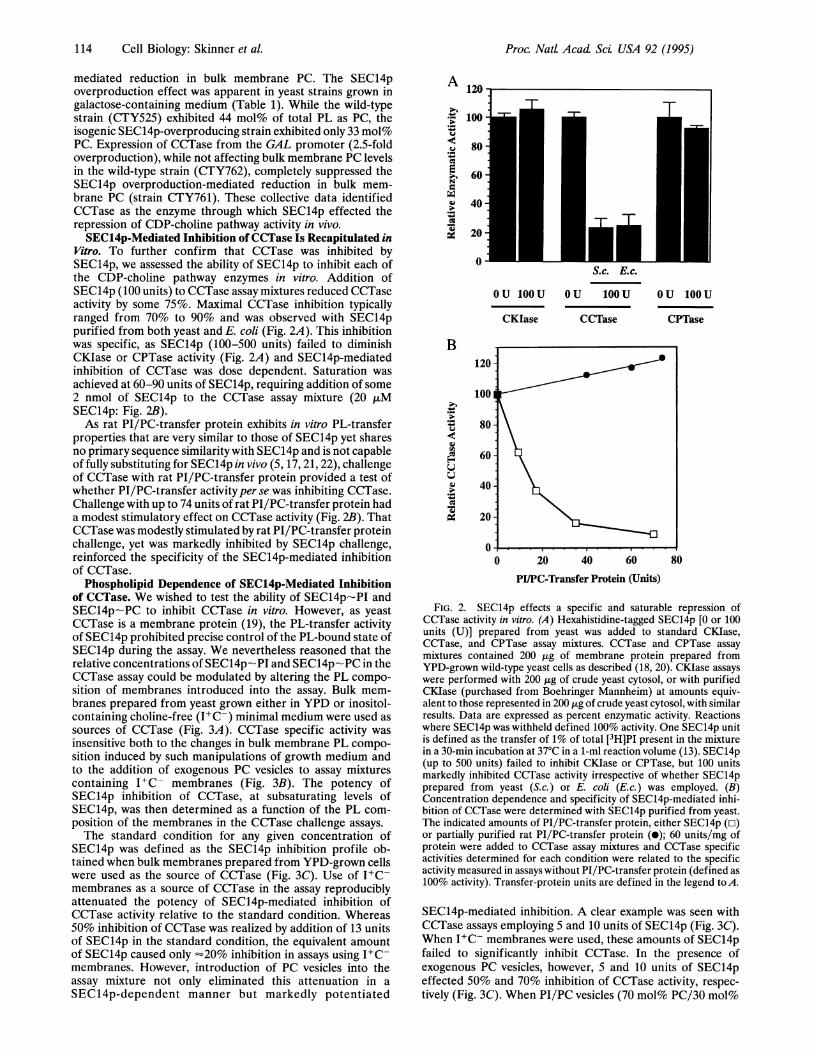
Vitro. To further confirm that CCTase was inhibited bySEC14p, we assessed the ability of SEC14p to inhibit each ofthe CDP-choline pathway enzymes in vitro. Addition ofSEC14p (100 units) to CCTase assay mixtures reduced CCTaseactivity by some 75%. Maximal CCTase inhibition typicallyranged from 70% to 90% and was observed with SEC14ppurified from both yeast and E. coli (Fig. 2A). This inhibitionwas specific, as SEC14p (100-500 units) failed to diminishCKIase or CPTase activity (Fig. 2A) and SEC14p-mediatedinhibition of CCTase was dose dependent. Saturation wasachieved at 60-90 units of SEC14p, requiring addition of some2 nmol of SEC14p to the CCTase assay mixture (20 ,uMSEC14p: Fig. 2B).As rat PI/PC-transfer protein exhibits in vitro PL-transfer
properties that are very similar to those of SEC14p yet sharesno primary sequence similarity with SEC14p and is not capableof fully substituting for SEC14p in vivo (5, 17,21,22), challengeof CCTase with rat PI/PC-transfer protein provided a test ofwhether PI/PC-transfer activityper se was inhibiting CCTase.Challenge with up to 74 units of rat PI/PC-transfer protein hada modest stimulatory effect on CCTase activity (Fig. 2B). ThatCCTase was modestly stimulated by rat PI/PC-transfer proteinchallenge, yet was markedly inhibited by SEC14p challenge,reinforced the specificity of the SEC14p-mediated inhibitionof CCTase.
Phospholipid Dependence of SEC14p-Mediated Inhibitionof CCTase. We wished to test the ability of SEC14p-PI andSEC14p-PC to inhibit CCTase in vitro. However, as yeastCCTase is a membrane protein (19), the PL-transfer activityof SEC14p prohibited precise control of the PL-bound state ofSEC14p during the assay. We nevertheless reasoned that therelative concentrations ofSEC14p-PI and SEC14p-PC in theCCTase assay could be modulated by altering the PL compo-sition of membranes introduced into the assay. Bulk mem-branes prepared from yeast grown either in YPD or inositol-containing choline-free (I+C-) minimal medium were used assources of CCTase (Fig. 3A). CCTase specific activity wasinsensitive both to the changes in bulk membrane PL compo-sition induced by such manipulations of growth medium andto the addition of exogenous PC vesicles to assay mixturescontaining I+C- membranes (Fig. 3B). The potency ofSEC14p inhibition of CCTase, at subsaturating levels ofSEC14p, was then determined as a function of the PL com-position of the membranes in the CCTase challenge assays.The standard condition for any given concentration of
SEC14p was defined as the SEC14p inhibition profile ob-tained when bulk membranes prepared from YPD-grown cellswere used as the source of CCTase (Fig. 3C). Use of I+C-membranes as a source of CCTase in the assay reproduciblyattenuated the potency of SEC14p-mediated inhibition ofCCTase activity relative to the standard condition. Whereas50% inhibition of CCTase was realized by addition of 13 unitsof SEC14p in the standard condition, the equivalent amountof SEC14p caused only '20% inhibition in assays using I+C-membranes. However, introduction of PC vesicles into theassay mixture not only eliminated this attenuation in aSEC14p-dependent manner but markedly potentiated
A 120
*: 100*
80
; 60N
v 40
; 20
0
0U lOOU
CKIase
B
;O-
,*Cu
E
v
Cu
120 -
100 1
80-
60-
40-
20-
S.c. E.c.
OU lOOU
CCTase
r 20 4
0 20 40
OU lOOU
CPTase
60 80
PIMPC-Transfer Protein (Units)
FIG. 2. SEC14p effects a specific and saturable repression ofCCTase activity in vitro. (A) Hexahistidine-tagged SEC14p [0 or 100units (U)] prepared from yeast was added to standard CKIase,CCTase, and CPTase assay mixtures. CCTase and CPTase assaymixtures contained 200 ,tg of membrane protein prepared fromYPD-grown wild-type yeast cells as described (18, 20). CKIase assayswere performed with 200 ,g of crude yeast cytosol, or with purifiedCKIase (purchased from Boehringer Mannheim) at amounts equiv-alent to those represented in 200 tig of crude yeast cytosol, with similarresults. Data are expressed as percent enzymatic activity. Reactionswhere SEC14p was withheld defined 100% activity. One SEC14p unitis defined as the transfer of 1% of total [3H]PI present in the mixturein a 30-min incubation at 37°C in a 1-ml reaction volume (13). SEC14p(up to 500 units) failed to inhibit CKIase or CPTase, but 100 unitsmarkedly inhibited CCTase activity irrespective of whether SEC14pprepared from yeast (S.c.) or E. coli (Ec.) was employed. (B)Concentration dependence and specificity of SEC14p-mediated inhi-bition of CCTase were determined with SEC14p purified from yeast.The indicated amounts of PI/PC-transfer protein, either SEC14p (z1)or partially purified rat PI/PC-transfer protein (0); 60 units/mg ofprotein were added to CCTase assay mixtures and CCTase specificactivities determined for each condition were related to the specificactivity measured in assays without PI/PC-transfer protein (defined as100% activity). Transfer-protein units are defined in the legend to A.
SEC14p-mediated inhibition. A clear example was seen withCCTase assays employing 5 and 10 units of SEC14p (Fig. 3C).When I+C- membranes were used, these amounts of SEC14pfailed to significantly inhibit CCTase. In the presence ofexogenous PC vesicles, however, 5 and 10 units of SEC14peffected 50% and 70% inhibition of CCTase activity, respec-tively (Fig. 3C). When PI/PC vesicles (70 mol% PC/30 mol%
in
11 Iv
Proc Natl Acad Sci USA 92 (1995)
I
Proc. NatL Acad Sci USA 92 (1995) 115
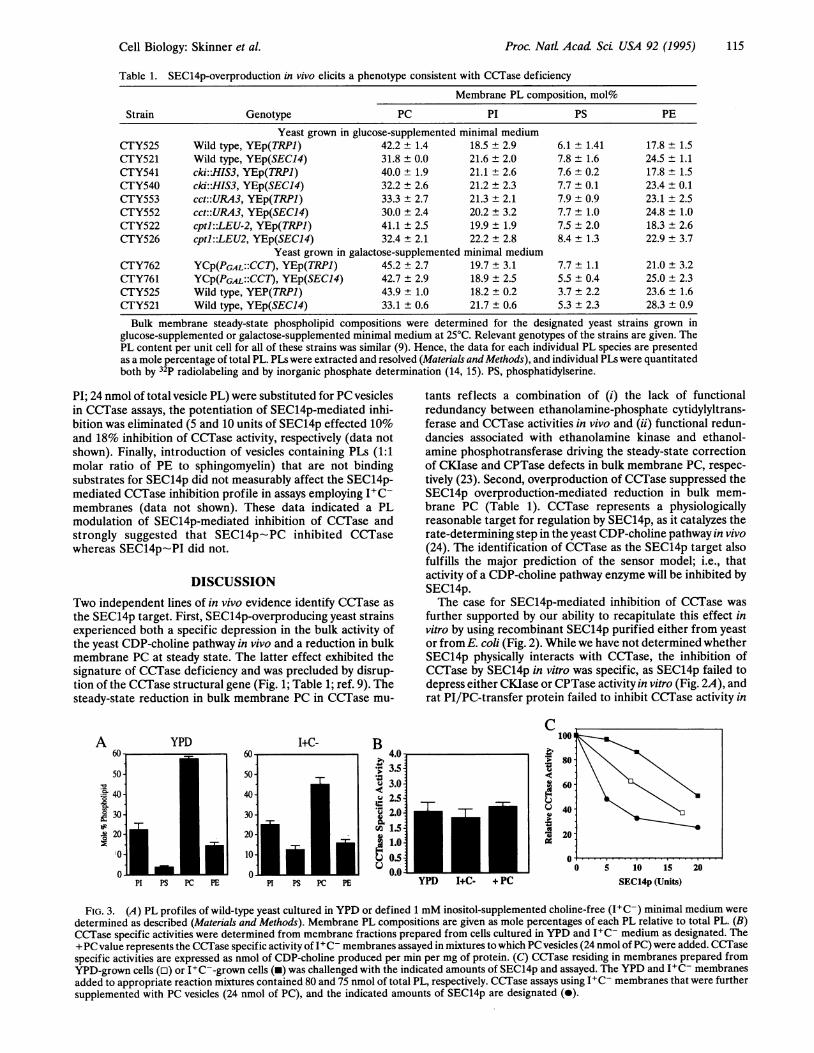
Table 1. SEC14p-overproduction in vivo elicits a phenotype consistent with CCTase deficiency
Membrane PL composition, mol%
Strain Genotype PC PI PS PE
Yeast grown in glucose-supplemented minimal mediumCTY525 Wild type, YEp(TRPI) 42.2 ± 1.4 18.5 ± 2.9 6.1 ± 1.41 17.8 ± 1.5CTY521 Wild type, YEp(SEC14) 31.8 ± 0.0 21.6 ± 2.0 7.8 ± 1.6 24.5 ± 1.1CTY541 cki:1IS3, YEp(TRPI) 40.0 ± 1.9 21.1 ± 2.6 7.6 ± 0.2 17.8 ± 1.5CTY540 cki:HIS3, YEp(SEC14) 32.2 ± 2.6 21.2 ± 2.3 7.7 ± 0.1 23.4 ± 0.1CTY553 cct::URA3, YEp(TRPI) 33.3 ± 2.7 21.3 ± 2.1 7.9 ± 0.9 23.1 ± 2.5CTY552 cct::URA3, YEp(SEC14) 30.0 ± 2.4 20.2 ± 3.2 7.7 ± 1.0 24.8 ± 1.0CTY522 cptl::LEU-2, YEp(TRPI) 41.1 ± 2.5 19.9 ± 1.9 7.5 ± 2.0 18.3 ± 2.6CTY526 cptl::LEU2, YEp(SEC14) 32.4 ± 2.1 22.2 ± 2.8 8.4 ± 1.3 22.9 ± 3.7
Yeast grown in galactose-supplemented minimal mediumCTY762 YCP(PGAL::CCT), YEp(TRPJ) 45.2 ± 2.7 19.7 ± 3.1 7.7 ± 1.1 21.0 ± 3.2CTY761 YCP(PGAL::CCT), YEp(SEC14) 42.7 ± 2.9 18.9 ± 2.5 5.5 ± 0.4 25.0 ± 2.3CTY525 Wild type, YEP(TRPI) 43.9 ± 1.0 18.2 ± 0.2 3.7 ± 2.2 23.6 ± 1.6CfTY521 Wild type, YEp(SEC14) 33.1 ± 0.6 21.7 ± 0.6 5.3 ± 2.3 28.3 ± 0.9
Bulk membrane steady-state phospholipid compositions were determined for the designated yeast strains grown inglucose-supplemented or galactose-supplemented minimal medium at 25°C. Relevant genotypes of the strains are given. ThePL content per unit cell for all of these strains was similar (9). Hence, the data for each individual PL species are presentedas a mole percentage of total PL. PLs were extracted and resolved (Materials and Methods), and individual PLs were quantitatedboth by 32P radiolabeling and by inorganic phosphate determination (14, 15). PS, phosphatidylserine.
PI; 24 nmol of total vesicle PL) were substituted for PC vesiclesin CCTase assays, the potentiation of SEC14p-mediated inhi-bition was eliminated (5 and 10 units of SEC14p effected 10%and 18% inhibition of CCTase activity, respectively (data notshown). Finally, introduction of vesicles containing PLs (1:1molar ratio of PE to sphingomyelin) that are not bindingsubstrates for SEC14p did not measurably affect the SEC14p-mediated CCTase inhibition profile in assays employing I+C-membranes (data not shown). These data indicated a PLmodulation of SEC14p-mediated inhibition of CCTase andstrongly suggested that SEC14p-PC inhibited CCTasewhereas SEC14p-PI did not.
DISCUSSIONTwo independent lines of in vivo evidence identify CCTase asthe SEC14p target. First, SEC14p-overproducing yeast strainsexperienced both a specific depression in the bulk activity ofthe yeast CDP-choline pathway in vivo and a reduction in bulkmembrane PC at steady state. The latter effect exhibited thesignature of CCTase deficiency and was precluded by disrup-tion of the CCTase structural gene (Fig. 1; Table 1; ref. 9). Thesteady-state reduction in bulk membrane PC in CCTase mu-
A YPD60-
50-cl 40 -.sa 230
.v 20-
PI PS PC PE
I+C- B
s*;l
Qglav
Pi PS PC PE
tants reflects a combination of (i) the lack of functionalredundancy between ethanolamine-phosphate cytidylyltrans-ferase and CCTase activities in vivo and (ii) functional redun-dancies associated with ethanolamine kinase and ethanol-amine phosphotransferase driving the steady-state correctionof CKIase and CPTase defects in bulk membrane PC, respec-tively (23). Second, overproduction of CCTase suppressed theSEC14p overproduction-mediated reduction in bulk mem-brane PC (Table 1). CCTase represents a physiologicallyreasonable target for regulation by SEC14p, as it catalyzes therate-determining step in the yeast CDP-choline pathway in vivo(24). The identification of CCTase as the SEC14p target alsofulfills the major prediction of the sensor model; i.e., thatactivity of a CDP-choline pathway enzyme will be inhibited bySEC14p.The case for SEC14p-mediated inhibition of CCTase was
further supported by our ability to recapitulate this effect invitro by using recombinant SEC14p purified either from yeastor from E. coli (Fig. 2). While we have not determined whetherSEC14p physically interacts with CCTase, the inhibition ofCCTase by SEC14p in vitro was specific, as SEC14p failed todepress either CKIase or CPTase activity in vitro (Fig. 2A), andrat PI/PC-transfer protein failed to inhibit CCTase activity in
C1001
~6040
20
0 5 10 15 20SEC14p (Units)
FIG. 3. (A) PL profiles of wild-type yeast cultured in YPD or defined 1 mM inositol-supplemented choline-free (I+C-) minimal medium weredetermined as described (Materials and Methods). Membrane PL compositions are given as mole percentages of each PL relative to total PL. (B)CCTase specific activities were determined from membrane fractions prepared from cells cultured in YPD and I+C- medium as designated. The+PC value represents the CCTase specific activity of I+C- membranes assayed in mixtures to which PC vesicles (24 nmol of PC) were added. CCTasespecific activities are expressed as nmol of CDP-choline produced per min per mg of protein. (C) CCTase residing in membranes prepared fromYPD-grown cells (o) or I+C--grown cells (m) was challenged with the indicated amounts of SEC14p and assayed. The YPD and I+C- membranesadded to appropriate reaction mixtures contained 80 and 75 nmol of total PL, respectively. CCTase assays using I+C- membranes that were furthersupplemented with PC vesicles (24 nmol of PC), and the indicated amounts of SEC14p are designated (0).
Cell Biology: Skinner et al.

116 Cell Biology: Skinner et al.
SEC14p-PI
I,l
SEC14p-PCSEC14p-PC
Cho + ATP
Cytosol
SEC14p-PL
>®^~~-...
Cho-P
ADP
,Lumen
Pi DAG
FIG. 4. SEC14p as a ligand-modulated inhibitor of yeast CCTaseactivity in Golgi membranes. SEC14p is concentrated on the cytosolicsurface of yeast Golgi membranes (7), where it is proposed to executea constitutive PL-exchange reaction that generates a pool of PL-boundSEC14p (SEC14p-PL) composed of both SEC14p-PC andSEC14p-PI, as indicated. SEC14p-PC serves as a negative effectorof CCTase activity in yeast Golgi membranes, and the extent ofCCTase inhibition is proportional to SEC14p-PC. Diacylglycerol isabbreviated to DAG.
vitro (Fig. 2B). Lastly, the finding that the potency of SEC14p-mediated inhibition of CCTase was a function of membranePC content suggested that SEC14p-PC was the CCTaseinhibitor (Fig. 3C). The indication that the regulatory functionof SEC14p was controlled by the ligand-bound state ofSEC14p provided strong support for a second major tenet ofthe sensor model.We propose that SEC14p functions to downregulate CDP-
choline pathway activity in yeast Golgi membranes in vivo byinhibiting CCTase and that SEC14p-PC is responsible forinhibition of CCTase (Fig. 4). Thus, a sensitive feedback loopis created that couples the action of CCTase to Golgi PC levels.We also propose that the CCTase-directed activity of SEC14pis itself regulated through a constitutive PL-exchange reactionexecuted by SEC14p on Golgi membranes, an activity thatmanifests itself as a PL-exchange activity in vitro. This modelpredicts that a mutant SEC14p able to bind/transfer only PCwill inhibit CCTase and score as a functional SEC14p in vivo,whereas a SEC14p capable of executing only PI-binding/transfer will neither inhibit CCTase nor score as a functionalSEC14p in vivo. Finally, we suggest that the PI-binding activityof SEC14p constitutes a competitive PL-binding reaction thatconstantly regulates the amount of SEC14-PC present inresponse to the PI/PC ratio present in Golgi membranes.
This proposed mechanism of SEC14p function is analogousto that established for Ras-like GTP-binding proteins, whereligand occupancy regulates signal transduction from a sensor
component to a downstream effector (25). For SEC14p, theinherent PL-exchange activity associated with this proteinprovides a mechanism for controlling its CCTase-directedregulatory activity. These data emphasize the potential for an
indirect relevance of the in vitro PL-transfer activity of aprotein to in vivo function and indicate that the mechanism offunction determined for one PL-transfer protein cannot beconfidently extrapolated to that of another: even to one thatexhibits similar PL-substrate specificities in vitro. Finally, thesensor model leaves open the possibility that, as in the case ofRas-like GTP-binding proteins, there may exist multiple mem-bers of a SEC14p family in cells, each dedicated to regulatinga specific cellular process. Consistent with this idea, we haveidentified a SEC14p homolog in S. cerevisiae that shares highprimary sequence homology, but not functional redundancy,with SEC14p (unpublished data).
We thank Jesse Hay and Tom Martin for their generous gift of ratPI/PC-transfer protein. This work was supported by grants from theU.S. Public Health Service (GM44530) and the American HeartAssociation (92-763) (to V.A.B.) and by a Basic Mechanisms of LungDisease postdoctoral training grant from the National Institutes ofHealth (5T32 HL07553) (to M.R.F.). C.R.M. and R.M.B. weresupported by National Institutes of Health Grant GM20015 (toR.M.B.).
1. Wirtz, K. W. A. (1991) Annu. Rev. Biochem. 60, 73-99.2. Rueckert, D. G. & Schmidt, K. (1990) Chem. Phys. Lipids 56,
1-20.3. Cleves, A., McGee, T. & Bankaitis, V. (1991) Trends Cell Biol. 1,
30-34.4. Novick, P., Field, C. & Schekman, R. (1980) Cell 21, 205-215.5. Bankaitis, V. A., Malehorn, D. E., Emr, S. D. & Greene, R.
(1989) J. Cell Biol. 108, 1271-1281.6. Bankaitis, V. A., Aitken, J. R., Cleves, A. E. & Dowhan, W.
(1990) Nature (London) 347, 561-562.7. Cleves, A. E., McGee, T. P., Whitters, E. A., Champion, K. M.,
Aitken, J. R., Dowhan, W., Goebl, M. & Bankaitis, V. A. (1991)Cell 64, 789-800.
8. Kennedy, E. P. & Weiss, S. B. (1956)J. Biol. Chem. 222,185-191.9. McGee, T. P., Skinner, H. B., Whitters, E. A., Henry, S. A. &
Bankaitis, V. A. (1994) J. Cell Biol. 124, 273-287.10. Ito, H., Fukuda, Y., Murata, K. & Kimura, A. (1983) J. Bacteriol.
153, 163-168.11. Sherman, F., Fink, G. R. & Hicks, J. B. (1983) Methods in Yeast
Genetics (Cold Spring Harbor Lab. Press, Plainview, NY).12. Klig, L. S., Homann, M. J., Carman, G. M. & Henry, S. A. (1985)
J. Bacteriol. 162, 1135-1141.13. Aitken, J. F., van Heusden, G. P. H., Temkin, M. & Dowhan, W.
(1990) J. Biol. Chem. 265, 4711-4717.14. Ames, B. N. (1966) Methods Enzymol. 8, 115-116.15. Atkinson, K. D. (1984) Genetics 108, 533-543.16. Bush, G. L., Tassin, A.-M., Friden, H. & Meyer, D. I. (1991) J.
Biol. Chem. 266, 13811-13814.17. Skinner, H. B., Alb, J. G., Whitters, E. A., Helmkamp, G. M., Jr.,
& Bankaitis, V. A. (1993) EMBO J. 12, 4775-4784.18. Hosaka, K., Kodake, T. & Yamashita, S. (1989) J. Biol. Chem.
264, 2053-2059.19. Nikawa, J., Yonemura, K. & Yamashita, S. (1983) Eur. J.
Biochem. 131, 223-229.20. Hjelmstad, R. H. & Bell, R. M. (1987) J. Biol. Chem. 262,
3909-3917.21. Daum, G. & Paltauf, F. (1984) Biochim. Biophys. Acta 794,
385-391.22. Dickeson, S. K., Lim, C. N., Schuyler, G. T., Dalton, T. P.,
Helmkamp, G. M., Jr., & Yarbrough, L. R. (1989) J. Biol. Chem.264, 16557-16564.
23. McGee; T. P., Skinner, H. B. & Bankaitis, V. A. (1994) J. Bac-teriol. 176, 6861-6868.
24. Tsukagoshi, Y., Nikawa, J. & Yamashita, S. (1987) Eur. J.Biochem. 169, 477-486.
25. Bourne, H. R., Sanders, D. A. & McCormick, F. (1991) Nature(London) 349, 117-127.
Proc- Natl Acad Sci USA 92 (1995)

















![FeatureawareDigitalSurfaceModelanalysisand ...1.1Motivation 3 (a)Earlyillustrationofthe2DMAT(consistingofedges andpoints)oftwo‘1anthropomorphs’[Blum,1967]. (b)The3DMAT(right)ofasurfacemodelofahand(left](https://img.pdfslide.net/doc/110x75/612f210b1ecc515869433f32/featureawaredigitalsurfacemodelanalysisand-11motivation-3-aearlyillustrationofthe2dmatconsistingofedges.jpg)






![A Mean Field View of the Landscape of Two-Layers Neural … · 2018. 6. 6. · AMeanFieldViewoftheLandscape ofTwo-LayersNeuralNetworks AndreaMontanari [withSongMei,Phan-MinhNguyen]](https://img.pdfslide.net/doc/110x75/611227ff8db59724b615a22f/a-mean-field-view-of-the-landscape-of-two-layers-neural-2018-6-6-ameanfieldviewofthelandscape.jpg)




