Embed Size (px)
Citation preview

Tn
PI
a
ARRAA
KABBMM
1
tercattti
srnsmsac
0h
Biological Psychology 96 (2014) 8– 19
Contents lists available at ScienceDirect
Biological Psychology
journa l h om epa ge: www.elsev ier .com/ locate /b iopsycho
urning bias in virtual spatial navigation: Age-related differences andeuroanatomical correlates
eng Yuan, Ana M. Daugherty, Naftali Raz ∗
nstitute of Gerontology and Department of Psychology, Wayne State University, 87 East Ferry Street, 226 Knapp Building, Detroit, MI 48202, United States
r t i c l e i n f o
rticle history:eceived 26 June 2013eceived in revised form 24 October 2013ccepted 27 October 2013vailable online 2 November 2013
eywords:gingrain volumerain asymmetry
a b s t r a c t
Rodents frequently exhibit rotational bias associated with asymmetry in lesions and neurotransmittersin the striatum. However, in humans, turning preference is inconsistent across studies, and its neuralcorrelates are unclear. We examined turning bias in 140 right-handed healthy adults (18–77 years old),who navigated a virtual Morris Water Maze. On magnetic resonance images, we measured volumes ofbrain regions relevant to spatial navigation. We classified turns that occurred during virtual navigationas veering (less than 10◦), true turns (between 10◦ and 90◦) and course reversals (over 90◦). The resultsshowed that performance (time of platform search and distance traveled) was negatively related to age.The distance traveled was positively associated with volume of the orbito-frontal cortex but not with thevolumes of the cerebellum, the hippocampus or the primary visual cortex. Examination of turning behav-
RIorris Water Maze
ior showed that all participants veered to the right. In turns and reversals, although on average there wasno consistent direction preference, we observed significant individual biases. Virtual turning preferencecorrelated with volumetric asymmetry in the striatum, cerebellum, and hippocampus but not in the pre-frontal cortex. Participants preferred to turn toward the hemisphere with larger putamen, cerebellumand (in younger adults only) hippocampus. Advanced age was associated with greater rightward turningpreference. Men showed greater leftward preference whereas women exhibited stronger rightward bias.
. Introduction
Changing direction is an integral part of search and naviga-ion in complex and dynamic environments encountered in dailyxperience. Surprisingly, in the extant literature, this essential skilleceived substantially lesser attention than other component pro-esses and skills related to spatial navigation (see Moffat, 2009 for
review). It is unclear, for example, how changes of search direc-ion affect overall success of navigation, and whether the decisiono alter the search path is based solely on the incoming informa-ion or is influenced by individual biases of the navigators and theirndividual characteristics such as age and sex.
Investigation of rotational asymmetry has a long history intudies of animal behavior. These studies show that ambulatingats exhibit a significant degree of turning bias that depends oneurochemical characteristics of the neostriatum and other braintructures that are associated with execution and planning ofovement. Rats tend to turn in the direction of the brain hemi-
phere with lower striatal dopamine content (Zimmerberg, 1974),nimals trained in circling show increased dopamine content in theaudate nucleus contralateral to the circling direction (Yamamoto &
∗ Corresponding author. Tel.: +1 313 577 2297; fax: +1 508 256 5689.E-mail address: [email protected] (N. Raz).
301-0511/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.biopsycho.2013.10.009
© 2013 Elsevier B.V. All rights reserved.
Freed, 1982; Yamamoto, Lane, & Freed, 1982), but unilateral lesionsin the caudate nucleus cause transient contralateral rotation bias(Glick, 1978). Notably, in contrast to multiple findings pertainingto neurochemical basis of turning behavior, there is no informationabout the relationship between structural (e.g., volumetric) asym-metry of the neostriatum and other brain structures and turningbias in intact animals.
In healthy humans, behavioral studies that measured veeringand spontaneous turning in straight-line walking tasks producedinconsistent results. Some reported a leftward turning preference(Lenoir, Van Overschelde, De Rycke, & Musch, 2006; Mohr, Brugger,Bracha, Landis, & Viaud-Delmon, 2004; Toussaint & Fagard, 2008),whereas others found a rightward bias (Lyon & Satz, 1991; Mead &Hampson, 1997) or no preference at all (Bracha, 1987). Several fac-tors have been proposed to explain the predilection for turning ina specific direction, including handedness (Bradshaw & Bradshaw,1988; Mohr & Bracha, 2004; Mohr, Landis, Bracha, & Brugger, 2003),positional constraints on the starting point (Lenoir et al., 2006),hormonal fluctuations (Mead & Hampson, 1997), and differencesin task demands (Bradshaw & Bradshaw, 1988; Mohr et al., 2004).None of these studies evaluated the relationship between turning
preference and structural or functional properties of the brain.Information about turning bias in humans comes mostly fromnaturalistic studies, in which the investigators observed changesin whole-body position in real world environments during

cal Psy
aoLpfepitoiM
tMseatgitw
iiscp(M(stdrtpinihbIiniuAah2
rrsosAmwbett2
P. Yuan et al. / Biologi
ctivities such as running, stepping, forward-backward walking,r long-term spontaneous ambulation (for a summary see Mohr &ievesley, 2007). Thus, it is unclear whether the observed turningreference reflects asymmetry in gross motor activity or stemsrom differences in cognitive processing. To the best of our knowl-dge, none of the extant studies assessed rotation and turningreferences in a goal oriented navigation task, real or virtual. Thus,
t is important to examine turning preference in a navigation taskhat is independent of gross motor function. Such disengagementf navigation-related cognition and gross motor activity is possiblen a virtual environment exemplified by a virtual Morris Water
aze task (vMWM, Moffat & Resnick, 2002).In this computerized adaptation of one of the most popular
asks for studying rodent navigation and memory, Morris Wateraze (Morris, 1984; Morris, Garrud, Rawlins, & O’Keefe, 1982), a
earch for a hidden goal platform happens in a circular open fieldnvironment that does not enforce one direction over another,s do naturalistic environments that include barriers and struc-ured paths. Unlike the animal MWM that requires whole-bodyross-motor behavior, the vMWM yields the data that are largelyndependent of the whole-body responses and changes therein ashe participants “swim” to the platform by controlling a joystickithout altering their body position.
The neural basis of turning preference is poorly understood, butt is plausible that asymmetry in several brain regions that arenvolved in navigation may constitute its neuroanatomical sub-trate. These regions include hippocampus, neostriatum, frontalortex, and cerebellum. Most studies of rodents and humans sup-ort the critical role of the hippocampus in spatial navigationKaplan et al., 2012; Maguire et al., 1998; Maguire et al., 2000;
orris et al., 1982). However, this may not always be the caseMoffat, Kennedy, Rodrigue, & Raz, 2007). Notably, most in vivotudies have not assessed the contribution of subcortical struc-ures to individual differences in navigational skills and the one thatid revealed that age-related differences in the caudate nucleus,ather than hippocampal, volume correlated with success in findinghe goal in a virtual environment (Moffat et al., 2007). A com-lex skill, such as navigation, relies on multiple brain circuits and
nteractions among them. A recent study shows that during virtualavigation, both the hippocampus and the caudate are involved
n managing contextual information via the orbital frontal corticalub (Brown, Ross, Tobyne, & Stern, 2012). Thus, at least three majorrain structures contribute to one aspect of navigational behavior.
nterestingly, the extant literature on spatial navigation pays littlef any attention to the cerebellum, a functionally lateralized majoreural system involved in tasks that call for coordination of visual
nput and motor response, as well as in more complex cognitivendertakings that engage working memory (Botez, Botez, Elie, &ttig, 1989; Cabeza & Nyberg, 2000; Holmes, 1939). In one study,
non-significant trend for association between larger cerebellaremispheres and better virtual navigation was noted (Moffat et al.,007).
All of the studies of neural foundations of navigation in space,eal or virtual, have focused on the time and distance that wasequired to reach the goal as the primary outcome and none con-idered the components of the search such as turns and reversalsf course. Moreover, none of these studies examined the relation-hip between navigation and structural volumetric asymmetry.lthough the majority of studies of structural hemispheric asym-etry revealed left-sided specialization in the areas associatedith language, reports of volumetric asymmetries of brain regions
eyond language areas have produced inconsistent results (Good
t al., 2001; Raz et al., 2004; Zilles et al., 1996). It is possiblehat regional volumetric asymmetry is unreliable at the popula-ion level, although it may be stable in individual brains (Raz et al.,004).chology 96 (2014) 8– 19 9
To summarize, the extant literature indicates that rodents andhumans exhibit turning bias in free movement conditions. Animalturning preference may be related to neurochemical asymmetry inthe neostriatum. Although neural substrates of turning preferencehave not been studied in humans, human brains exhibit signifi-cant gross asymmetry and regional brain volumes are associatedwith proficiency in spatial navigation. Thus, in this study, our goalwas threefold. First, we sought to establish if indeed healthy adultsexhibited turning preference while navigating a virtual environ-ment. Second, we inquired whether turning asymmetry, if found,would be related, like other aspects of navigational performance,to age and sex. Third, we examined the possible neuroanatomicalcorrelates of turning behavior and turning asymmetry in humanvirtual navigation; based on rodent studies, we hypothesized thatvolume asymmetry of the neostriatum but not the hippocampusor prefrontal, orbito-frontal or primary visual cortices would berelated to contralateral rotational preference. We also hypothe-sized that because of its importance in perceptual motor tasks thatrequire continuous tracking of visual targets, hemispheric asym-metry in cerebellar volume would be also associated with turningpreference.
2. Materials and methods
2.1. Participants
The sample was part of a larger ongoing study of neural correlates of cognitiveaging. The participants were recruited from the metropolitan Detroit area throughadvertisement in the local media and screened by telephone interview and a healthquestionnaire. The participants included in the current study attained a minimum ofhigh school education, were native English speakers and were strongly right-handed(75% and above on the Edinburgh Handedness Questionnaire; Oldfield, 1971). Per-sons who reported a history of cardiovascular disease, neurological or psychiatricconditions, diabetes, head trauma with a loss of consciousness for more than 5 min,hypo- or hyperthyroidism, or drug and alcohol problems were excluded from partic-ipation in the study. Individuals who were taking anti-seizure medication, anxiolyt-ics, or antidepressants were also excluded. Mini Mental State Examination (MMSE;Folstein, Folstein, & McHugh, 1975) and Geriatric Depression Questionnaire (CES-D;Radloff, 1977) were used to screen for dementia and depression with only individu-als who scored at least 26 on the MMSE and below 16 on the CES-D admitted in thestudy. Participants, who had a diagnosis of hypertension, were taking antihyper-tensive medications, or had a measured blood pressure (averaged over three testingdays) in excess of 140 mm Hg (systolic) or 90 mm Hg (diastolic) were classified ashypertensive and excluded from the analyses. All participants provided informedconsent for participation in this study, which was approved by university humaninvestigations committee. Of 241 persons who completed the navigation study, 18were excluded because they were not strongly right-handed, 45 because of diag-nosed hypertension and additional 15 because of observed hypertension. Further-more, 23 subjects of did not have MRI scans. Thus, the sample consisted of 140 per-sons (93 women), who were 18–77 years old (M = 48.5 years, SD = 15.8 years). Therewere no significant sex difference in age (M = 48.8, SD = 14.9 in females; M = 48.0,SD = 17.5 in males, t (138) = 0.277, p = .782), education, MMSE or CESD (all p > .1).
2.2. MRI processing
Imaging was performed on a Bruker-Varian 4-Tesla MRI system with an8-channel radio frequency coil. Magnetization-prepared rapid gradient echo(MPRAGE) T1-weighted images were acquired in the coronal plane with thefollowing parameters: TE = 4.38 ms, TR = 1600 ms, TI = 800 ms, FOV = 256 mm, res-olution = .67 mm × .67 mm × 1.34 mm, matrix size = 384 × 384, and flip angle = 8◦ .
Images were rotated to adjust for variation in head pitch (forward or backward),tilt (to the left or right) and rotation (to the right or to the left), so that the potentialasymmetrical positioning in the scanner was corrected. The images were reformat-ted in native space to correct for undesired head tilt, pitch, and rotation, with Analyze10.0 software (Biomedical Imaging Resource, Mayo Clinic College of Medicine). Allimages were processed by the same group of expert operators who attained reli-ability of at least .90 as measured by the intraclass correlation formula ICC(2) thatassumes random raters (Shrout & Fleiss, 1979).
2.3. Volumetry
The volumes of the caudate nucleus (Cd), putamen (Pt), hippocampus (Hc),cerebellum (Cb), orbito-frontal (OFC) and dorsal lateral prefrontal cortex (DLPFC)in the left and right hemispheres were measured following the rules that weredescribed in detail in previous publications (e.g., Raz, Ghisletta, Rodrigue, Kennedy,& Lindenberger, 2010; Raz et al., 2005). The volumes of all six brain regions were

10 P. Yuan et al. / Biological Psychology 96 (2014) 8– 19
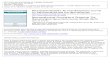
F ram op
aaTmrilca
wlbsh
Pbcta
ttttdH
ogigo
ldfws
sbmv
tla
2
d
ig. 1. A typical view from within the virtual environment (left) and overhead diagerspective.
djusted for individual differences in intracranial volume (ICV), using the ANOCVApproach described in previous publications (Jack et al., 1989; Raz et al., 2004).he volume asymmetries of these regions of interest (ROIs) were defined as asym-etry = (volumeright − volumeleft)/(volumeright + volumeleft). The asymmetry indices
anged between −1 and 1. Positive asymmetry scores indicated larger ROI volumen the right hemisphere than in the left hemisphere and negative scores indicatedarger ROI volume in the left than in the right hemisphere. The rules of region demar-ation, previously described in detail (see Raz et al., 2004; Raz et al., 2005, for rulesnd illustrations) are summarized below.
Caudate nucleus (Cd). The volume of the Cd was measured from the slice onhich the Cd first appeared, usually lateral to the lateral ventricles, until it was no
onger identifiable. The Cd was bordered medially by the lateral ventricle, laterallyy the internal capsule, dorsally by the subcortical white matter, and ventrally bytria terminalis (rostral part) and septal nucleus (caudal part). The ROI included theead and the body of Cd but not the tail.
Putamen (Pt). The volume of the Pt was measured from the slice on which thet first appeared until it was no longer identifiable. The Pt was bordered laterallyy the external capsule, medially by the internal capsule (anterior to the anteriorommissure) or globus pallidus (posterior to the anterior commissure), dorsally byhe subcortical white matter, and ventrally by the temporal stem, optic radiations,mygdala, temporal horn, and anterior commissure.
Hippocampus (Hc). Hc volume was measured on slices aligned perpendicular tohe long axis of the right Hc between the mammillary bodies and the slice showinghe fornices rising from the fimbria. The Hc included CA1–CA4, dentate gyrus andhe subiculum. The medial border of the Hc was defined by tracing the subiculumo its most medial position and drawing a horizontal line at the medial curve. Theorsal medial boundary was the ambient cistern above the Hc. The lateral border ofc was the lateral ventricle or the temporal lobe white matter.
Dorsal lateral prefrontal cortex (DLPFC). The volume of the DLPFC was measuredn the coronal slices located within the posterior 40% of the distance between theenu of the corpus callosum and the tip of the frontal pole. The described ROIncluded superior, middle, and inferior frontal gyri. The DLPFC was defined as theray matter located between the most dorsomedial point of the cortex and therbital sulcus.
Orbito-frontal cortex (OFC). The range of the OFC was identical to DLPFC. The mostateral branch of the orbital sulcus that breaches the external aspect of the brainefines the lateral boundary of the OFC, which also served as the lower boundaryor the DLPFC. The medial boundary was defined by the olfactory sulcus. The OFCas defined as the gray matter located between the orbital sulcus and olfactory
ulcus.Cerebellum (Cb). The cerebellar hemispheres were measured on the coronal
lices from the first slice on which it appeared to the last slice that it was visi-le. The hemispheric gray matter, the cerebellar tonsils, the vellum, and the corpusedullare were included, whereas the vermis, cerebellar peduncles, and the fourth
entricle were excluded in the tracing of each cerebellar hemisphere.Primary visual cortex (VC). This region was defined as the cortex located along
he banks of the calcarine sulcus. It was measured on the anterior 50% coronal slicesocated between the mid-vermis slice and the occipital pole. This ROI was a part ofrea 17, although it does not constitute the entirety of the primary visual cortex.
.4. vMWM apparatus and procedures
Apparatus. The vMWM task (Moffat & Resnick, 2002) was administered on a Dellesktop computer, with a 17-inch TFT LCD SONY monitor. Participants were seated
f the vMWM map (right). Participants viewed the environment from a first person
in a chair and their heads were approximately 50 cm from the screen. Participantsviewed the virtual environment from a first person perspective (Fig. 1) and guidedtheir movements with the use of a Thrustmaster Top Gun Fox 2 pro joystick.
Pretest training. Pretest training familiarized participants with the virtual envi-ronment and movement in the virtual environment with a joystick. After theexperimenter’s instruction, there was a period of free exploration of virtual envi-ronment using the joystick. Participants practiced until they were comfortable withthe joystick and could control movement in the virtual environment while navi-gating to targets designated by experimenter. To demonstrate their abilities to usethe joystick, participants completed two tasks in pretest training: moving onto topof small squares in the center of platforms; moving as quickly as possible to reacha goal along a hallway with long straight parts as well as turns in it. In addition,the participants practiced vMWM with a virtual circular pool filled with water andsurrounded by prominent objects. Participants were instructed to move to a plat-form that alternated between being visible and invisible in five successive trials. Theposition of the platform remained constant across trials. Each participant completedthe pretest training tasks, though the training time could be different among par-ticipants. Participants were instructed to move only forward and were informedthat backward movement was against the rules. Although the software did notpreclude participants from moving backwards in the virtual environment, if theyattempted such backwards movement, it was disallowed by experimenter, whowas present throughout the experiment. Movement speed varied and was individ-ually controlled by changing the force applied to the joystick. Participants were ableto change the direction of virtual movement either during forward motion or afterstopping in the vMWM.
vMWM learning trials. The vMWM was represented on a computer screen as acircular pool filled with water (Fig. 1). The diameter of the virtual pool was 4415virtual units. The pool was presented with an image of an irregularly shaped roomthat contained several objects that could serve as visual cues: a lamp, a group of trees,a flag, a picture, and a tree. A rectangular platform, with side length of 883 virtualunits, was hidden beneath the surface of water and participants were instructedto find the hidden platform as quickly as possible. When participants passed overthe platform, it would emerge from the water, thus terminating the trial. Therewas a 12-s interval between two successive trials. During the inter-trial interval,participants were able to move on the platform, although they were not explicitlyinstructed to do so. Participants were informed that the platform remained in thesame location on each trial and that the environment did not change. On each of thesix trials, participants were placed into one of the three quadrants of the pool notcontaining the platform, facing a different direction at the beginning of each trialwith the platform behind their field of view. Thus, participants were compelled tomake turns before they were able to orient toward the platform. Although the startlocations and directions differed between trials, the settings were identical for eachparticipant. There was no time limit on the learning trials.
Probe trial and visible platform trial. Immediately after 6 vMWM learning tri-als, the participants performed a 1-min probe trial, in which the environment wasidentical to the learning trials, except that the platform was removed. After theprobe trial, there was a visible platform trial, in which the water maze was the sameas the learning trials, but the platform was not submerged, and participants wereinstructed to move onto the visible platform as fast as possible.
2.5. Turning angle
Cursor position and the azimuth of movement to the platform were continu-ously sampled at a rate of 85 Hz. Moment-by-moment change in azimuth was an

P. Yuan et al. / Biological Psychology 96 (2014) 8– 19 11
iot(
2
1nasrlr
2
gpwswTrddi
etavfdvtbpda
cltraic
3
3
3
31
Table 1Correlations between age and vMWM performance in each trial.
Trial 1 Trial 2 Trial 3 Trial 4 Trial 5 Trial 6
Age–Distance traveled .145 .362** .270** .338** .293** .320**
Age–Time of search .340** .412** .351** .421** .352** .394**
Note: Travel distance and time were log-transformed to reduce skewness. Higher
Fig. 2. Examples of turn typology.
ndex of moving segment. Three types of moving segments were defined: continu-us clockwise turning (right rotation, Fig. 2, from A to B), continuous anti-clockwiseurning (left rotation, Fig. 2, from C to D and from O to A), and no change of azimuthstraight segment, Fig. 2, from B to C).
.6. Laterality indices
A regular turn was defined as a continuous change of movement direction by0–90◦; small deviations (veers) were defined as continuous changes of direction ofo more than 10◦; and a reversal was defined as continuous change of direction byt least 90◦ . To investigate rotation preference, we computed a laterality index byubtracting the number of left turns (or deviations, reversals) from the number ofight turns (or deviations, reversals), and dividing this difference by their sum. Thisaterality index ranged between −1 and 1. A positive laterality index score indicatedight rotation bias and negative scores indicated left rotation bias.
.7. Data analyses
First, we examined age and sex differences in traditional indices of virtual navi-ation performance: search distance and time of search. After establishing a typicalattern of age and sex differences and the course of navigation skill acquisition,e examined the pattern of direction changes exhibited in the process of virtual
earch. Age was centered at the sample mean. The other continuous variables,hen served as independent variables, were also centered at the sample mean.
he performance in visible platform session served as control for possible age-elated difference in visual and motor skill. All first-order interactions would beropped from models if found non-significant (p > .1). Benjamini–Hochberg proce-ure (Benjamini & Hochberg, 1995) was applied to control false discovery rate (FDR)
n multiple hypothesis testing.To examine age and sex differences in vMWM performance, we fitted a gen-
ral linear model (GLM) to the data. In this GLM, the travel distance or time washe dependent variable, age, sex and performance in visible platform trial serveds between-subject independent variables, and trial was a six-level within-subjectariable. To test the associations between regional brain volumes and vMWM per-ormance, we evaluated separate GLMs for each of the seven regions, with the travelistance as the dependent variable, age, sex and regional volume as independentariables and trial as a repeated measure. The distance traveled in visible platformrial was also included in models. In order to verify the relationship between regionalrain volumes and vMWM skill in the previous report (Moffat et al., 2007), in whicharticipants were drawn from two extreme age groups and age was treated as aichotomous variable, we also analyzed our data using a median split of age, inddition to treating age as a continuous variable.
To examine age and sex differences in virtual turning preference and the asso-iation between turning bias and regional hemispheric asymmetry, we analyzedaterality scores of all three types of turns in a GLM framework. In this model, theurn laterality index was the dependent variable, the turn type (veering, turns, andeversals) was a three-level repeated measure, whereas age, sex and volumetricsymmetry of the Hc, Cb, Cd, Pt, DLPFC, OFC and the VC served as between-subjectndependent variables. The performance in visible platform trial was included as aovariate as well.
. Results
.1. Behavioral findings
.1.1. Virtual navigation performance: search distanceThe mean distance traveled in the learning 6 trials was
1,769 (SD = 21,684) virtual units. The main effects of age (F (1,36) = 33.859, p < .001) and sex (F (1, 136) = 4.819, p = .030) were
scores reflect poorer performance.** p < .01.
significant, with women and older participants traveling longer dis-tance in search of the platform. There was also a main effect of trial(Fig. 3, F (5, 680) = 21.257, p < .001), indicating improvement in per-formance across trials. The interaction effects of trial × age (Fig. 4,F (5, 680) = 3.252, p = .007), trial × sex (F (5, 680) = 2.342, p = .040)and trial × performance in visible platform trial (F (5, 680) = 2.704,p = .020) were also significant. The linear component of the trialeffect was significant (F (1, 136) = 81.766, p < .001), as well as thequadratic component for trial × age interaction (F (1, 136) = 5.528,p = .020). Although performance improved with practice, age dif-ferences increased after the first trial (Table 1). The correlationbetween age and travel distance was smaller on trial 1 than thaton trial 2: Steiger’s Z* = −1.970, p < .05.
3.1.2. Virtual navigation performance: time of searchThe mean time to complete six vMWM trials was 186 s
(SD = 153 s). We examined age and sex differences in the timeof search by using an identical GLM approach. Similar as in thesearch distance analyses, there were significant effects of age (F(1, 136) = 44.421, p < .001) and sex (F (1, 136) = 6.490, p = .012),with older participants and women evidencing longer searchtimes. There was a significant main effect of trial (Fig. 3, F (5,680) = 9.218, p < .001) and a trend for trial × age interaction (Fig. 4,F (5, 680) = 2.140, p = .059). The linear component of the trial effect(F (1, 136) = 40.867, p < .001) and the quadratic component fortrial × age interaction (F (1, 136) = 5.250, p = .024) were also signif-icant. The positive correlations between age and search time weresignificant across all six trials (Table 1).
3.1.3. Side preference in turning behaviorWe observed a clear rightward bias in small deviations (veering)
from the search course. The values of the laterality index rangedfrom .400 to 1.000 (M = .794, SD = .106). However, there was nobias in course reversals; the laterality score did not differ fromzero: M = .063, SD = .458, median = .060. Although the mean lateral-ity index for regular turns differed from zero (M = −.080, SD = .413, t(139) = −2.298, p < .05), the median of 0.000 indicated the apparentmean bias attributable to the influence of a few extreme values.
Older age was associated with a greater number of rever-sals: r = .333 and turns: r = .361; both log transformed to reduceskewness, for both p < .001. Number of reversals and turns posi-tively correlated with time and distance of search across trials: allr > .6, p < .001. There were significantly more turns than reversals: t(139) = 3.952, p < .001. Although the asymmetry in turns and rever-sals correlated modestly (r = .202, p = .017), neither correlated withthe asymmetry of veering (r = .156, p = .065 for turns and r = .022,p = 799 for reversals).
3.1.4. Performance in probe trial and visible platform trialIn probe trial, the proportion of distance and time spent in tar-
get quadrant were both significantly greater than 25% (distance:
M = 0.347, SD = 0.199, t (139) = 5.797, p < .0001; time: M = 0.337,SD = 0.195, t (139) = 5.262, p < .0001). The association between ageand proportion of time or distance in target quadrant was not sig-nificant (age–time percent: r = .129, p = .13; age–distance percent:
12 P. Yuan et al. / Biological Psychology 96 (2014) 8– 19
Fig. 3. Sex differences in vMWM performance: Distance and time of search by trial.
F as a
i
rt
t(spi
3
3
essra(ropppF
right-sided asymmetry was greater in the VC than in caudate andhippocampus (Table 2).
A GLM with the asymmetry index as the dependent variable, ageand sex as independent variables and the ROI as repeated measures
Table 2Descriptive and inferential statistics of volume asymmetry indices.
Region Mean SD 95% confidence interval t value
Putamen −.032 .028 −.037 to −.028 −13.741OFC −.018 .048 −.026 to −.010 −4.384Cerebellum −.005 .011 −.007 to −.003 −5.101Caudate nucleus .014 .023 .010 to .018 7.201Hippocampus .015 .041 .008 to .021 4.171
ig. 4. Distance traveled and time spent in search of a hidden platform in the vMWMndex, distance and time.
= .148, p = .08). Thus, older participants were not performing worsehan the younger participants did in the probe trial.
In visible platform trial, the travel distance and time spento complete visible platform session varied with age (distance: F1, 137) = 10.558, p = .002; time: F (1, 137) = 34.994, p < .001) andex (distance: F (1, 137) = 9.165, p = .003; time: F (1, 137) = 5.819,
= .017). However, they did not significantly affect the performancen learning trials when age and gender were controlled (both p > .3).
.2. Neuroanatomical findings
.2.1. Regional brain volumesWe fitted a GLM with the ICV-adjusted volume as a depend-
nt measure, ROI as a seven-level repeated measure, and age andex as independent variables to the data. The analysis revealedignificant main effects of age (F (1, 136) = 79.978, p < .001) andegion (F (6, 816) = 9912.976, p < .001), as well as interactions ofge × region (F (6, 816) = 35.991, p < .001) and age × sex × region (F6, 816) = 5.013, p < .001). The follow-up GLM analyses for each ROIevealed associations between advanced age and smaller volumesf all regions: Cd: F (1, 137) = 48.697, p < .001; Pt: F (1, 137) = 57.553,
< .001; Hc: F (1, 137) = 11.909, p < .001; OFC: F (1, 137) = 74.271, < .001; DLPFC: F (1, 137) = 65.786, p < .001; Cb: F (1, 136) = 43.822, < .001; VC: F (1, 136) = 6.043, p = .015; p values were adjusted forDR; see the scatterplots of age differences in volume in Fig. 5.
function of age and trial number. The color codes the magnitude of the performance
3.2.2. Regional volumetric asymmetryWe observed anatomic asymmetry in the seven examined
regions. The left hemisphere was larger than the right one in theputamen, the OFC and cerebellum. Examination of the confidencelimits overlap among the regional indices of asymmetry (Table 2)revealing that the asymmetry was greater in the Pt than in the OFC,which in turn showed greater asymmetry than Cb did. In contrast,the right hemispheres were larger than the left ones in the cau-date, the hippocampus, the DLPFC and the VC. The magnitude of
DLPFC .023 .036 .017 to .029 7.591Primary visual cortex .037 .067 .026 to .048 6.539
Note: Asymmetry direction: R > L positive; t values present the result of one-samplet-test for volume asymmetry index of each ROI against zero, all p < .001.

P. Yuan et al. / Biological Psychology 96 (2014) 8– 19 13
in re
rpos
3
3
bvc
Fig. 5. Age differences
evealed a significant main effect of region (F (6, 822) = 46.168, < .001), that reflected the differences in direction and magnitudef asymmetry reported above. The effects of age and sex were notignificant.
.3. Neuroanatomical correlates of navigational performance
.3.1. Performance in vMWM and regional brain volumes
Except the age and sex effects, reported above, the associationsetween the indices of vMWM performance and regional brainolumes were not significant in any region (all p > .2), which is inontrast to the findings in the previous report (Moffat et al., 2007).
gional brain volumes.
However, once age was dichotomized, we replicated some of theresults of the previous study. Consistent with Moffat et al. (2007)findings, there was a main effect of OFC volume on the travel dis-tance, F (1, 135) = 7.572, p = .049. Larger Pt and DLPFC volumes werealso not associated with better vMWM performance, although non-significant trends were observed: Pt: F (1, 135) = 4.853, p = .068;DLPFC: F (1, 135) = 5.752, p = .063. All p values were adjusted forFDR. The effects of VC, Cd, Hc and Cb volumes were not significant.
3.3.2. Turning preference and neuroanatomical asymmetryAge (F (1, 128) = 12.515, p < .001), sex (F (1, 128) = 7.970, p = .006),
Pt asymmetry (F (1, 128) = 8.309, p = .005), Cb asymmetry (F (1,

1 cal Psychology 96 (2014) 8– 19
1Htme(pttttaoatiFtw
nsrTaf
t1rt1FtctapeFDo
4
idtoop
4
pdadr
cai
betw
een
age,
vMW
M
per
form
ance
, reg
ion
al
volu
mes
, vol
um
e
asym
met
ries
and
asym
met
ries
in
rota
tion
s.
AG
E
vMW
M
per
form
ance
Reg
ion
al
volu
me
(ad
just
ed
for
ICV
)
Asy
mm
etry
of
regi
onal
volu
me
Turn
ing
asym
met
ry
Dis
tan
ce
Tim
e
CD
V
PTV
HC
V
DLP
FCV
OFC
CB
V
VC
V
CD
PT
HC
DLP
FC
OFC
CB
VC
Vee
r
Turn
.47
.53
.92
−.51
−.26
−.29
−.54
−.34
−.39
.67
−.28
−.15
−.19
.36
.42
−.57
−.32
−.35
.48
.46
.35
−.59
−.34
−.38
.54
.49
.40
.77
−.47
−.26
−.30
.46
.41
.39
.37
.39
−.18
−.04
−.05
.28
.27
.22
.22
.28
.28
−.07
−.02
.00
−.01
.01
−.02
.09
.12
−.05
−.18
−.15
−.19
−.15
−.06
−.01
−.02
.13
.17
.05
.14
−.05
−.13
−.12
−.15
.14
.11
.08
.17
.19
.07
−.08
−.14
.13
m
.11
−.02
−.03
−.04
−.01
−.02
−.14
−.13
−.06
.05
−.01
.02
−.39
−.01
.01
−.02
−.01
−.01
−.15
−.10
−.05
.01
.06
.14
−.01
−.21
−.09
.03
−.06
−.09
−.03
.10
−.07
−.04
−.05
.00
−.08
.01
−.12
.08
.08
−.01
.14
−.02
−.00
−.07
−.15
.01
.04
.10
−.13
−.05
.04
−.11
.09
−.05
−.09
.10
−.03
−.02
−.14
−.06
−.06
−.01
−.09
−.01
−.02
.06
.08
.05
.00
.15
.02
.07
.00
.19
.26
.25
−.13
−.06
.02
−.05
.00
−.05
.01
.00
.10
−.01
−.02
−.02
.21
.17
.16
rsal
.19
−.01
.07
−.09
−.08
−.14
−.22
−.25
−.08
.07
−.20
.15
.07
.04
−.16
.19
.03
.02
.20
ym, P
T
Asy
m, H
C
Asy
m, D
LPFC
Asy
m, O
FC
Asy
m, C
B
Asy
m, V
C
Asy
m: v
olu
met
ric
asym
met
ry
ind
ices
of
Cd
, Pt,
Hc,
DLP
FC, O
FC, C
b,
VC
, res
pec
tive
ly. A
sym
veer
, Asy
m
turn
, Asy
m
reve
rsal
: asy
mm
etry
ind
ices
of
smal
lla
r
turn
s
and
reve
rsal
s,
resp
ecti
vely
. Asy
mm
etry
dir
ecti
on:
R
>
L
pos
itiv
e.
4 P. Yuan et al. / Biologi
28) = 11.133, p = .001), as well as the interaction between age andc asymmetry (F (1, 128) = 4.420, p = .038) were associated with
urning preference. Within-subjects analysis revealed a significantain effect of rotation type (F (2, 256) = 280.812, p < .001) and sev-
ral significant interactions: rotation type × age × Hc asymmetryF (2, 256) = 3.107, p = .046), rotation type × age (F (2, 256) = 5.259,
= .006), rotation type × sex (F (2, 256) = 3.706, p = .026) and rota-ion type × Cb asymmetry (F (2, 256) = 3.142, p = .045). In addition,here were two nonsignificant trends for interactions: rotationype × Cd asymmetry (F (2, 256) = 2.500, p = .084) and rotationype × Pt asymmetry (F (2, 256) = 2.212, p = .112). The correlationsmong relevant variables are presented in Table 3. The asymmetryf veering (small deviations) was unrelated to age, sex or volumesymmetry: all p > .05, as depicted in Fig. 6a. The interaction of turnypes with age reflected greater counter-clockwise (left) preferencen turns among younger participants (F (1, 128) = 9.277, p = .008,ig. 6b) and greater clockwise (right) preference in reversals amonghe older adults (F (1, 128) = 7.353, p = .011, Fig. 6c). Both p valuesere adjusted for FDR.
Men evidenced bias toward leftward (counter-clockwise) onormal turns (M = −.184, t (46) = −2.912, p = .006) but not on rever-als, whereas women showed rightward (clockwise) preference oneversals (M = .111, t (92) = 2.274, p = .025) but not on regular turns.he sex differences were significant: F (1, 128) = 5.538 for turns,nd F (1, 128) = 5.345 for reversals, both p = .034 after controllingor FDR.
The asymmetry of Cb volume was associated with ipsilateralurning bias: F (1, 128) = 6.553, p = .021 for turns, Fig. 7a; F (1,28) = 6.198, p = .021 for reversals, Fig. 8a; i.e., the participantsotated toward the larger hemisphere. Ipsilateral Pt asymme-ry was also marginally related to lateral bias in reversals (F (1,28) = 5.665, p = .056, Fig. 8b) and turns (F (1, 128) = 3.366, p = .10,ig. 7b); i.e., course reversals were more likely in the direction ofhe larger hemisphere. In contrast, Cd asymmetry was not asso-iated with turning bias (F (1, 128) = 3.881, p = .153). Althoughhe main effect of Hc asymmetry was not significant, there wasn interaction of age with Hc asymmetry (F (1, 128) = 7.950,
= .006) for regular turns. Hc asymmetry was related to ipsilat-ral turn preference in young (aged < 41, N = 41, r = .310, p = .049,ig. 9) but not in older participants. The asymmetry of the VC,LPFC and OFC were unrelated to the turning bias for any typef turn.
. Discussion
We found that in a manually executed desk-top virtual nav-gation task, healthy adults exhibited a wide range of individualifferences in turning preference. The direction and magnitude ofurning bias differed according to participants’ age and sex. More-ver, the turning asymmetry correlated with volume asymmetryf the neostriatum, cerebellum, and (among younger people) hip-ocampus but not of the prefrontal cortex.
.1. Age differences in turning asymmetry
In their deviation from the search path, the participants dis-layed a wide range of deviation angles, and the magnitude ofeviation mattered. Right-side preference was clear for small devi-tions from the course – veering. We can only speculate that sucheviations are likely non-volitional and reflect participants’ strongight-handedness.
In contrast to veering, when participants decided to change theourse or reverse the direction of the virtual search, they showed, as
group, no directional preference, while exhibiting a wide range ofndividual differences in turning bias. The latter were related to age Ta
ble
3C
orre
lati
ons
lgD
ista
nce
lgTi
me
CD
V
PTV
HC
V
DLP
FCV
OFC
V
CB
V
VC
V
CD
Asy
m
PT
Asy
m
HC
Asy
m
DLP
FC
Asy
OFC
Asy
m
CB
Asy
m
VC
Asy
m
Asy
m
veer
Asy
m
turn
Asy
m
reve
Not
es: C
D
As
veer
ing,
regu

P. Yuan et al. / Biological Psychology 96 (2014) 8– 19 15
FR
aTpptpsc
ig. 6. Age differences in lateral bias for three types of turns. Asymmetry direction: > L positive.
nd the type of turns that were executed during vMWM navigation.he younger participants evidenced counter-clockwise (leftward)references in true turns (10–90◦), but with increasing age of thearticipants, progressively smaller preference was observed, with
he older adults showed no consistent turning bias at all. A contraryattern was noticed for course reversals: the younger participantshowed no directional bias, whereas the older adults evidencedlockwise (right-side) preference.Fig. 7. Lateral bias for regular turns vs. volume asymmetry. Asymmetry direction:R > L positive.
In as much as the virtual manual navigation corresponds toa full body movement in a naturalistic setting, the leftward tur-ning bias of the younger adults is in accord with the reportedcounter-clockwise bias in running within an arena configuredsimilarly to a vMWM (Toussaint & Fagard, 2008). It is unclearwhy such bias is absent in the older adults and why youngerparticipants exhibited no bias in course reversals. Increases inturns and reversals were associated with advanced age, regard-less of the direction, and the number of turns and reversalscorrelated with longer time of search (i.e., worse performance).Thus, both types of deviations from course indicated maladap-tive search behavior. However, reversals were less frequent thanturns and it is possible that the young participants just did notproduce a sufficient number of reversals to yield valid indices ofasymmetry.
Because the indices of turning and reversals asymmetry cor-related weakly among themselves and did not show significantcorrelations with veering asymmetry, it is unclear whether turningrepresents a coherent construct. Veering asymmetry may reflect abias in unilateral joystick use by strongly right-handed participants.Indeed, veering preference for the right side was almost universaland unrelated to age, sex or brain asymmetry. In contrast, indi-vidual asymmetry in true turns and course reversals may reflect
a combination of automatized movement programs and deliber-ate data-driven decisions. Course reversals, in addition may reflectsignificant disorientation in the search space. Which of those playa greater role in spatial navigation is unclear, but the observed
16 P. Yuan et al. / Biological Psy
Fp
rs
4
t
Fp
ig. 8. Lateral bias in reversals vs. volume asymmetry. Asymmetry direction: R > Lositive.
elationships between turning bias and brain asymmetries provideome clues.
.2. Sex differences in turning asymmetry
In true turns, men showed stronger leftward bias relativeo women, whereas in reversals, men shifted toward no clear
ig. 9. Lateral bias in regular turns vs. hippocampal volume asymmetry in youngerarticipants (less than 41 years of age). Asymmetry direction: R > L positive.
chology 96 (2014) 8– 19
directional preference, whereas women exhibited propensity toturn rightward. This sex difference is consistent with the previousexperimental (Mead & Hampson, 1996) and questionnaire (Stochl& Croudace, 2012) studies that reported greater rightward bias inwomen compared to men. However, our findings are in contrastwith an observational study that reported rightward turning biasin right-handed males and leftward bias in right-handed females(Bracha, Seitz, Otemaa, & Glick, 1987). This discrepancy may arisefrom the difference in measures of rotational preference. The firststudy (Mead & Hampson, 1996) tested turning preference in a lim-ited and structured environment: the participants were instructedto turn 180◦ and walk directly to a tape recorder located behindthem. The second study (Bracha, Seitz et al., 1987) investigatedhuman turning preference during long-term spontaneous activity.Different tasks may evoke different rotational preference even inthe same participants (Mohr et al., 2004). Besides the differencein the context and structure of navigation tasks, the definition of aturn also varies among the studies. For example, in the assessmentof bias in long-term spontaneous turning, investigators (Bracha,Seitz et al., 1987) counted right and left full turns of at least 360◦,whereas Mead (Mead & Hampson, 1996) counted 180◦ turns. Inthe current study, these types of turns would be viewed as coursereversals. These discrepancies hamper the comparison of results.
4.3. Neuroanatomical correlates of navigation and turning bias
In accord with the results of a previous study on the same task(Moffat et al., 2007), better performance in vMWM was associatedwith larger OFC and (marginally) larger DLPFC volumes. There wasalso a trend that better navigational performance correlated withlarger volumes of brain regions in a cortico-striatal system, whichplays an important role in supporting computational and strategicaspects of the search. However, neither the volume of the hip-pocampus, a putative neural substrate of spatial mapping, nor thevolume of the cerebellum, which supports various cognitive andperceptual-motor operations were related to speed and efficiencyof navigation. These results are in accord with the previous report(Moffat et al., 2007), in which the effect of hippocampus volumewas nonsignificant, whereas the effect of cerebellum volume wasonly marginally significant on vMWM performance. Future stud-ies measuring the volume of hippocampal subfields may clarifyfurther the association between hippocampal volume and naviga-tional performance.
To the best of our knowledge, this is the first study to exam-ine the structural brain correlates of turning preference in healthyhumans. We observed that the lateral, age and sex differencesin turning preference and its brain correlates varied across threetypes of turning behavior. Asymmetry in turning behavior, espe-cially in course reversals, was associated with ipsilateral volumeasymmetry in the cerebellum and putamen. Hippocampal volumeasymmetry was related to ipsilateral turn preference, but only inyounger (under the age of 41) participants. The asymmetry of thecortical regions, VC, DLPFC and OFC was unrelated to the turningbias. Notably, veering, the activity with consistent lateral bias wasunrelated to anatomical asymmetry in any of the examined brainregions.
The absence of associations between prefrontal asymmetry andany sort of turning bias suggests that the latter was unlikely toreflect bias in higher cognitive processes that rely on the DLPFCand OFC. In contrast, the observed correlations between hemi-spheric asymmetry in motor structures (Cb and Pt) and rotationalbias indicate that asymmetric structure of the motor, primarily
extrapyramidal, system may be responsible for at least some of theobserved bias.What aspect or property of these motor structures supports theobserved behavioral asymmetry is unknown and in the absence of

cal Psy
nughcTmSto
dotNauCRptmof
etct2naIotmleloamrmoic
bsmdD(r
4
vttaoir
P. Yuan et al. / Biologi
eurochemical measures we can only speculate about the possiblenderlying mechanisms. Given the association of dopaminer-ic activity with rotational bias in rodents (Zimmerberg, 1974),emispheric asymmetry in dopaminergic activity may be a goodandidate for a neurochemical substrate of the observed bias.he literature suggests that asymmetry of spontaneous rotationay depend on asymmetric striatal dopaminergic activity (Bracha,
hultz, Glick, & Kleinman, 1987; Lyon & Satz, 1991). However, inhe absence of measurement of DA activity, the link between thebserved turning asymmetry and striatal DA remains a speculation.
To the best of our knowledge, in contrast to the relative abun-ance of studies on neostriatal dopamine, hemispheric asymmetryf this neurotransmitter in the Cb remains unexplored. Althoughhe Cb controls ipsilateral finger movements (Solodkin, Hlustik,oll, & Small, 2001; Wiestler, McGonigle, & Diedrichsen, 2011), thessociation of volumetric Cb asymmetry with turning preference isnlikely to reflect exclusively asymmetry of hand movement. Theb is involved in spatial navigation (Rondi-Reig & Burguiere, 2005;ondi-Reig, Le Marec, Caston, & Mariani, 2002) as it shapes hip-ocampal spatial representation and therefore affects navigationhat is based on self-motion cues (Rochefort et al., 2011). The Cb
ay affect the turning asymmetry by influencing the processingf optic flow cues in the hippocampus, and this hypothesis meritsurther investigation.
In the current study, the association between turning prefer-nce and volumetric asymmetry of the Hc was significant amonghe younger participants only. A similar observation of the asso-iation between the Hc and navigational performance as specifico younger participants has been reported by others (Moffat et al.,007). Moreover, the extant findings linking hippocampal size andavigational process (e.g., Maguire et al., 1998; Maguire et al., 2000)re limited to samples of younger and middle-aged participants.t is possible that younger adults adopted an allocentric (place-riented) strategy that relies on the Hc, whereas older individualsend to adhere to an egocentric (self-referential) strategy that
akes lesser use of the Hc circuitry. Indeed, older adults are lessikely than their younger counterparts to adopt an allocentric strat-gy in virtual navigation (Rodgers, Sindone, & Moffat, 2012) and areess likely to abandon an egocentric strategy even in the presencef corrective feedback (Harris, Wiener, & Wolbers, 2012). Because
desktop vMWM task allows for subject’s head movements, itay invoke activity in the hippocampal place cells that is greatly
educed by the absence of vestibular cues in head-restrained ani-als (Chen, King, Burgess, & O’Keefe, 2013). In this context, lack
f association between turning bias and hippocampal asymmetryn older participants may be related to lesser dependence on placeell activity in older adults.
Advanced age was associated with weaker leftward turningias in true turns (10–90◦) and greater rightward bias in rever-als (90◦ and above). The observed age-related turning preferenceight be related to age-related differences in asymmetry of striatal
opamine availability. Rightward asymmetry of dopamine D2 and3 receptors availability in the caudate nucleus decreases with age
Vernaleken, 2007), and thus, can attenuate leftward or exaggerateightward turning bias.
.4. Limitations of the study
Although it may seem obvious, it is important to emphasize thatirtual and real navigations are not identical, and interpretation ofhe extant virtual navigation experiments conducted at the desk-op or in an MRI scanner must proceed with caution. Navigation in
virtual reality environment executed by a person sitting in frontf a computer screen shares many characteristics with navigationn real space: reliance on visual cues to solve a spatial problem,oot planning, and use of working memory for spatial locations and
chology 96 (2014) 8– 19 17
management of search history. However, the view field affordedby the 17-in. monitor used in this study was limited, and usinglarger screen or goggles would make the virtual environment moresimilar to the real world. In addition, unlike its real counterpart,virtual navigation at the desktop gives little opportunity for use ofmotor and proprioceptive information, i.e., idiothetic cues (Taube,Valerio, & Yoder, 2013). Nonetheless, unlike executing virtual spa-tial search from supine position with constrained head movementsin fMRI experiments, conducting a search in virtual environmentwhile being deprived of whole-body motor cues may be not of aconsiderable detriment to navigation if at least some head positioncues are available (see Taube et al., 2013 for a review), as is the casein the desktop vMWM task used in our experiment.
Because virtual environments lack proprioceptive cues andvisual flow that are inherent to real world navigation, spatial infor-mation acquired from the virtual environment is less accurate thanthat from a real environment (Wilson, 1997). However, humans cantransfer spatial knowledge acquired from virtual environments tothe real world (Wilson, 1997), and the spatial knowledge in a vir-tual environment is highly predictive of performance in a similarreal-world maze (Waller, 2000). Thus, although it remains unclearhow the results of two-dimensional joystick-powered virtual nav-igation are related to true body movement in space, the observedassociations between turning laterality and regional volume asym-metry contribute to the understanding of the neural basis of spatialcognition. Systematic manipulation of availability of vestibular andmotor cues in virtual navigation is necessary for clarification ofconditions, under which a task such as vMWM can be treated asa window into real navigational skills.
Several limitations of the current study reflect selection of sub-jects and task parameters. Relative rarity of left-handers in thepopulation (Tan, 1988) prompted us to limit our sample to strongright-handed participants, who, with very rare exceptions, oper-ated the joystick exclusively with their right hand. None of theparticipants consistently used left hand, although in 3–4 cases theybriefly switched to non-dominant hand or used both hands forunknown reasons. Thus, our findings are limited to persons whoexhibit strong right-hand preference. Moreover, with age, the pro-portion of right-handers in the population steadily increases (Coren& Halpern, 1991; Hugdahl, Satz, Mitrushina, & Miller, 1993; Porac &Friesen, 2000). As a result of that age-related variation at the popu-lation level, the representativeness of participants with a particularhand preference might vary with age.
Because of its cross-sectional design, this study cannot informabout age-related change in turning preference, cerebral asymme-try and the relationship between them. In addition, generalizabilityof our findings is limited by selection of participants with noreported health problems.
Finally, currently available structural MRI methods do not allowfor a precise measurement of volumes in the regions that aredimmed critical of utilization of head direction cues in rodents– the lateral mammillary nuclei, anterior dorsal thalamus andpost-subiculum (Taube et al., 2013). Future improvements in MRIinstrumentation and refinement of currently available sequencesare needed for closing this gap between animal studies and humanvirtual navigation experiments. In future research it is also impor-tant to explore the roles of hippocampus sub-regions (CA1, CA3,the dentate gyrus) in navigation. Precise measure of sub-region vol-umes would help to reveal their relation to turning behavior duringnavigation. Unfortunately, the sequence that would allow such pre-cision of Hc parcellation (see Bender, Daugherty, & Raz, 2013) wasunavailable to us at the time of this study. Use of semi-automatic
measurement tools, such as Freesurfer, has its own share of problemwhen applied to aging, For example, Hc volume is overestimated byabout 20% in Freesurfer and the correlations between Freesurfer andmanually traced volumes are around .75 at best (Cherbuin, Anstey,
1 cal Psy
RPtnm
nahciniWabofc
A
ID
R
B
B
B
B
B
B
B
B
C
C
C
C
F
G
G
H
8 P. Yuan et al. / Biologi
églade-Meslin, & Sachdev, 2009; Hasan & Pedraza, 2009; Pardoe,ell, Abbott, & Jackson, 2009; Shen et al., 2010). Thus, clarifica-ion of the role played by specific Hc regions in various aspects ofavigation would require further development of neuroanatomicaleasurement techniques.In summary, turning preference observed during goal-directed
avigation in a virtual environment correlates with age, sexnd hemispheric asymmetry of the putamen, cerebellum, andippocampus. The observed differences in associations betweenortical and subcortical structural asymmetries and turning behav-or point to a previously undeclared source of variance in virtualavigation. Although landmark orientation based on spatial cues
s the crucial part of navigation in mammals (Taube et al., 2013;ang & Spelke, 2002), the observed bias in turning could introduce
n unconscious component that might impair course correctionased on noisy visual-spatial information. When under conditionsf uncertainty that are especially likely in a search for a hidden plat-orm, the cortex commands to make a left, cerebellum and striatuman give an unwanted nudge to the right.
cknowledgments
The study was supported in part by a grant from the Nationalnstitute on Aging (R37 AG011230) to NR. We are grateful to Cherylahle, Andrew Bender, and Yiqin Yang for help in data collection.
eferences
ender, A. R., Daugherty, A. M., & Raz, N. (2013). Vascular risk moderates associa-tions between hippocampal subfield volumes and memory. Journal of CognitiveNeuroscience, 25, 1851–1862.
enjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practicaland powerful approach to multiple testing. Journal of the Royal Statistical Society,Series B, 57, 289–300.
otez, M. I., Botez, T., Elie, R., & Attig, E. (1989). Role of the cerebellum in complexhuman-behavior. Italian Journal of Neurological Sciences, 10, 291–300.
racha, H. S. (1987). Asymmetric rotational (circling) behavior, a dopamine-related asymmetry: Preliminary findings in unmedicated and never-medicatedschizophrenic patients. Biological Psychiatry (1969), 22, 995–1003.
racha, H. S., Seitz, D. J., Otemaa, J., & Glick, S. D. (1987). Rotational movement (cir-cling) in normal humans: Sex difference and relationship to hand, foot and eyepreference. Brain Research, 411, 231–235.
racha, H. S., Shults, C., Glick, S. D., & Kleinman, J. E. (1987). Spontaneous asymmetriccircling behavior in hemi-parkinsonism; a human equivalent of the lesioned-circling rodent behavior. Life Sciences, 40, 1127–1130.
radshaw, J. L., & Bradshaw, J. A. (1988). Rotational and turning tendencies inhumans: An analog of lateral biases in rats? International Journal of Neuroscience,39, 229–232.
rown, T. I., Ross, R. S., Tobyne, S. M., & Stern, C. E. (2012). Cooperative interac-tions between hippocampal and striatal systems support flexible navigation.Neuroimage, 60, 1316–1330.
abeza, R., & Nyberg, L. (2000). Imaging cognition II: An empirical review of 275 PETand fMRI studies. Journal of Cognitive Neuroscience, 12, 1–47.
hen, G., King, J. A., Burgess, N., & O’Keefe, J. (2013). How vision and movementcombine in the hippocampal place code. Proceedings of the National Academy ofSciences of the United States of America, 110, 378–383.
herbuin, N., Anstey, K. J., Réglade-Meslin, C., & Sachdev, P. S. (2009). In vivo hip-pocampal measurement and memory: A comparison of manual tracing andautomated segmentation in a large community-based sample. PLoS One, 4,e5265.
oren, S., & Halpern, D. F. (1991). Left-handedness: A marker for decreased survivalfitness. Psychological Bulletin, 109, 90–106.
olstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). “Mini-mental state”. A practicalmethod for grading the cognitive state of patients for the clinician. Journal ofPsychiatric Research, 12, 189–198.
lick, S. D. (1978). Nocturnal rotation in normal rats: Correlation withamphetamine-induced rotation and effects of nigro-striatal lesions. BrainResearch, 150, 149–161.
ood, C. D., Johnsrude, I., Ashburner, J., Henson, R. N., Friston, K. J., & Frackowiak, R.S. (2001). Cerebral asymmetry and the effects of sex and handedness on brain
structure: A voxel-based morphometric analysis of 465 normal adult humanbrains. Neuroimage, 14, 685–700.arris, M. A., Wiener, J. M., & Wolbers, T. (2012). Aging specifically impairs switch-ing to an allocentric navigational strategy. Frontiers in Aging Neuroscience,4, 29.
chology 96 (2014) 8– 19
Hasan, K. M., & Pedraza, O. (2009). Improving the reliability of manual and automatedmethods for hippocampal and amygdala volume measurements. Neuroimage,48, 497–498.
Holmes, G. (1939). The cerebellum of man. Brain, 62, 1–30.Hugdahl, K., Satz, P., Mitrushina, M., & Miller, E. N. (1993). Left-handedness and old
age: Do left-handers die earlier? Neuropsychologia, 31, 325–333.Jack, C. R., Jr., Twomey, C. K., Zinsmeister, A. R., Sharbrough, F. W., Petersen, R. C., &
Cascino, G. D. (1989). Anterior temporal lobes and hippocampal formations: Nor-mative volumetric measurements from MR images in young adults. Radiology,172, 549–554.
Kaplan, R., Doeller, C. F., Barnes, G. R., Litvak, V., Duzel, E., Bandettini, P. A., et al.(2012). Movement-related theta rhythm in humans: Coordinating self-directedhippocampal learning. Plos Biology, 10, e1001267.
Lenoir, M., Van Overschelde, S., De Rycke, M., & Musch, E. (2006). Intrinsic andextrinsic factors of turning preferences in humans. Neuroscience Letters, 393,179–183.
Lyon, N., & Satz, P. (1991). Left turning (swivel) in medicated chronic schizophrenicpatients. Schizophrenia Research, 4, 53–58.
Maguire, E. A., Burgess, N., Donnett, J. G., Frackowiak, R. S., Frith, C. D., & O’Keefe, J.(1998). Knowing where and getting there: A human navigation network. Science,280, 921–924.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak,R. S., et al. (2000). Navigation-related structural change in the hippocampi oftaxi drivers. Proceedings of the National Academy of Sciences of the United Statesof America, 97, 4398–4403.
Mead, L. A., & Hampson, E. (1996). A sex difference in turning bias in humans.Behavioural Brain Research, 78, 73–79.
Mead, L. A., & Hampson, E. (1997). Turning bias in humans is influenced by phase ofthe menstrual cycle. Hormones and Behavior, 31, 65–74.
Moffat, S. D. (2009). Aging and spatial navigation: What do we know and where dowe go? Neuropsychology Review, 19, 478–489.
Moffat, S. D., Kennedy, K. M., Rodrigue, K. M., & Raz, N. (2007). Extra hippocampalcontributions to age differences in human spatial navigation. Cerebral Cortex, 17,1274–1282.
Moffat, S. D., & Resnick, S. M. (2002). Effects of age on virtual environment placenavigation and allocentric cognitive mapping. Behavioral Neuroscience, 116,851–859.
Mohr, C., & Bracha, H. S. (2004). Compound measure of hand-foot-eye preferencemasked opposite turning behavior in healthy right-handers and non-right-handers: Technical comment on Mohr et al. (2003). Behavioral Neuroscience,118, 1145–1146.
Mohr, C., Brugger, R., Bracha, H. S., Landis, T., & Viaud-Delmon, I. (2004). Human sidepreferences in three different whole-body movement tasks. Behavioural BrainResearch, 151, 321–326.
Mohr, C., Landis, T., Bracha, H. S., & Brugger, P. (2003). Opposite turningbehavior in right-handers and non-right-handers suggests a link betweenhandedness and cerebral dopamine asymmetries. Behavioral Neuroscience, 117,1448–1452.
Mohr, C., & Lievesley, A. (2007). Test–retest stability of an experimental measure ofhuman turning behaviour in right-handers, mixed-handers, and left-handers.Laterality, 12, 172–190.
Morris, R. (1984). Developments of a water-maze procedure for studying spatiallearning in the rat. Journal of Neuroscience Methods, 11, 47–60.
Morris, R. G., Garrud, P., Rawlins, J. N., & O’Keefe, J. (1982). Place navigation impairedin rats with hippocampal lesions. Nature, 297, 681–683.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The edinburghinventory. Neuropsychologia, 9, 97–113.
Pardoe, H. R., Pell, G. S., Abbott, D. F., & Jackson, G. D. (2009). Hippocampal volumeassessment in temporal lobe epilepsy: How good is automated segmentation?Epilepsia, 50, 2586–2592.
Porac, C., & Friesen, I. C. (2000). Hand preference side and its relation to handpreference switch history among old and oldest-old adults. Developmental Neu-ropsychology, 17, 225–239.
Radloff, L. S. (1977). The CES-D scale: A self-report depression scale forresearch in the general population. Applied Psychological Measurement, 1,385–401.
Raz, N., Ghisletta, P., Rodrigue, K. M., Kennedy, K. M., & Lindenberger, U. (2010). Tra-jectories of brain aging in middle-aged and older adults: Regional and individualdifferences. Neuroimage, 51, 501–511.
Raz, N., Gunning-Dixon, F., Head, D., Rodrigue, K. M., Williamson, A., & Acker, J. D.(2004). Aging, sexual dimorphism, and hemispheric asymmetry of the cerebralcortex: Replicability of regional differences in volume. Neurobiology of Aging, 25,377–396.
Raz, N., Lindenberger, U., Rodrigue, K. M., Kennedy, K. M., Head, D., Williamson,A., et al. (2005). Regional brain changes in aging healthy adults: Gen-eral trends, individual differences and modifiers. Cerebral Cortex, 15,1676–1689.
Rochefort, C., Arabo, A., Andre, M., Poucet, B., Save, E., & Rondi-Reig, L. (2011). Cere-bellum shapes hippocampal spatial code. Science, 334, 385–389.
Rodgers, M. K., Sindone, J. A., 3rd, & Moffat, S. D. (2012). Effects of age on navigationstrategy. Neurobiology of Aging, 33, 202 e15–22.
Rondi-Reig, L., & Burguiere, E. (2005). Is the cerebellum ready for navigation?Progress in Brain Research, 148, 199–212.
Rondi-Reig, L., Le Marec, N., Caston, J., & Mariani, J. (2002). The role of climbingand parallel fibers inputs to cerebellar cortex in navigation. Behavioural BrainResearch, 132, 11–18.

cal Psy
S
S
S
ST
T
T
V
P. Yuan et al. / Biologi
hen, L., Saykin, A. J., Kim, S., Firpi, H. A., West, J. D., Risacher, S. L., et al. (2010).Comparison of manual and automated determination of hippocampal volumesin MCI and early AD. Brain Imaging and Behavior, 4, 86–95.
hrout, P. E., & Fleiss, J. L. (1979). Intraclass correlations: Uses in assessing raterreliability. Psychological Bulletin, 86, 420–428.
olodkin, A., Hlustik, P., Noll, D. C., & Small, S. L. (2001). Lateralization of motor cir-cuits and handedness during finger movements. European Journal of Neurology,8, 425–434.
tochl, J., & Croudace, T. (2012). Predictors of human rotation. Laterality.an, U. (1988). The distribution of hand preference in normal men and women.
International Journal of Neuroscience, 41, 35–55.aube, J. S., Valerio, S., & Yoder, R. M. (2013). Is navigation in virtual real-
ity with fMRI really navigation? Journal of Cognitive Neuroscience, 25,
1008–1019.oussaint, Y., & Fagard, J. (2008). A counterclockwise bias in running. NeuroscienceLetters, 442, 59–62.
ernaleken, I. (2007). Asymmetry in dopamine D2/3 receptors of caudate nucleus islost with age. NeuroImage, 34, 870.
chology 96 (2014) 8– 19 19
Waller, D. (2000). Individual differences in spatial learning from computer-simulated environments. Journal of Experimental Psychology: Applied, 6, 307–321.
Wang, R., & Spelke, E. (2002). Human spatial representation: Insights from animals.Trends in Cognitive Sciences, 6, 376.
Wiestler, T., McGonigle, D. J., & Diedrichsen, J. (2011). Integration of sensory andmotor representations of single fingers in the human cerebellum. Journal ofNeurophysiology, 105, 3042–3053.
Wilson, P. N. (1997). Transfer of spatial information from a virtual to a real environ-ment. Human Factors, 39, 526–531.
Yamamoto, B. K., & Freed, C. R. (1982). The trained circling rat: A model for inducingunilateral caudate dopamine metabolism. Nature, 298, 467–468.
Yamamoto, B. K., Lane, R. F., & Freed, C. R. (1982). Normal rats trained to circle showasymmetric caudate dopamine release. Life Sciences, 30, 2155–2162.
Zilles, K., Dabringhaus, A., Geyer, S., Amunts, K., Qu, M., Schleicher, A., et al. (1996).Structural asymmetries in the human forebrain and the forebrain of non-humanprimates and rats. Neuroscience and Biobehavioral Reviews, 20, 593–605.
Zimmerberg, B. (1974). Neurochemical correlate of a spatial preference in rats. Sci-ence, 185, 623–625.