Embed Size (px)
Citation preview

Kidney International, Vol. 7 (1975), P. 145—153
Ultrastructure of a specialized neck region in therabbit nephron
HENRIK C. SCHØNHEYDER and ARVID B. MAUNSBACH
Department of Cell Biology, Institute of Anatomy, University of Aarhus, Aarhus, Denmark
Ulfrastructure of a specialized neck region in the rabbit nephron.The transition between the glomerulus and the proximal tubulewas studied by light and electron microscopy in perfusion-fixedrabbit kidneys. In most nephrons a transitional segment oftubule occurred between the Bowman's capsule and the proxi-mal tubule proper. This initial part of the tubule was lined byflattened cells and formed a cytologically distinct neck segment.The nephron population was heterogeneous with respect to thelength of this neck segment regardless of the level of the cortex.The neck cells contained an abundance of thin filaments oftenoriented at right angles to the long axis of the tubule and com-parable in thickness to actin filaments. The presence of actin-likefilaments suggests that neck cells may effect changes in the dia-meter of the neck segment and, thus, influence single nephronglomerular filtration rate.
Ultrastructure d'une region spécialisée du collet du néphron delapin. La transition entre le glomérule et Ic tube proximal a étéétudiée par microscopie photonique et electronique dans desreins de lapin fixes par perfusion. Dans la plupart des néphrons,un segment transitionnel du tubule existe entre la capsule deBowman et le tube proximal proprement dit. Cette partie initialedu tubule est bordée de cellules aplaties et forme un collet cyto-logiquement distinct. La population des néphrons est heterogCneen ce qui concerne la longueur de ce collet et ceci a tous lesniveaux du cortex. Les cellules du collet contiennent en abon-dance des filaments fins souvent orientés perpendiculairement al'axe du tubule et d'épaisseur comparable a celle des filamentsd'actine. La presence de ces filaments semblables a de l'actinesuggére que les cellules du collet peuvent determiner tine modi-fication du diamètre de ce collet et ainsi influencer la filtrationglomérulaire du même néphron.
It is well documented that in the kidney of amphibiaand most fishes a specialized, ciliated segment, fre-quently referred to as the neck segment, is interposedbetween the glomerulus and the proximal tubule [1—3].A similar cytologically distinct neck segment is notgenerally recognized in mammals [3—6] except in cer-tain pathologic conditions [7—9]. However, a constric-tion in the initial part of the proximal tubule has beenreported to occur occasionally in some species [10,11].
Received for publication May 20, 1974;and in revised form August 13, 1974.© 1975, by the International Society of Nephrology.
145
Other early light microscope studies quoted by vonMöllendorff [4] have indicated that in the cat and someother mammalian species the initial part of the proxi-mal tubule may be lined by flattened cells. Besides thisspecies variation, the glomerulotubular junction maydiffer somewhat from individual to individual [4, 10,11]. Since the glomerulotubular junction is criticallylocated for influencing the effective glomerular filtra-tion pressure and the passage of fluid into the proximaltubule, we have carried out an electron microscopestudy in the rabbit kidney to extend previous lightmicroscope observations. We have analyzed in somedetail the ultrastructure of the mammalian neck cellsand report here, among other data, an abundance ofactin-like filaments in these cells.
Methods
Five male adult white rabbits of the New Zealandstrain were used. A special procedure for anesthesiawas employed to avoid vascular reactions during sur-gery prior to fixation of the kidneys (de Mello, per-sonal communication). The rabbits were first given i.p.injections of 1 ml of sterile 0.9% sodium chloride dailyfor one week to condition the animals for the anesthe-sia. Lidocaine (0.5) was infiltrated around an earvein and the rabbit anesthetized by an i.v. injection of1 ml of 5% thiopental sodium (Pentothal, Abbott)followed by an i.p. injection of 3 ml of 5% pentobarbitalsodium (Nembutal, Abbott) and 0.5 ml of 0.1 % atro-pine [12]. A rubber band was placed around the thoraxto stimulate respiration. About eight minutes after theinitial dose, the rabbit was given another 0.3 ml of 5%thiopental sodium i.v. The kidneys were then fixed byperfusion with glutaraldehyde through the abdominalaorta as previously described for rats (method 1 in[13]). The glutaraldehyde was purified with activated
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by Elsevier - Publisher Connector

146 Schunheyder/Maunsbach
charcoal prior to use. The fixative solution contained2% glutaraldehyde in 0.1M cacodylate buffer and 4°/polyvinylpyrrolidone (PYP, mol wt about 40,000,Sigma Chemical Company). The fixation was precededby a short rinse with a mammalian Ringer's solutionwhich was contained in the tubing connecting the con-tainer of the fixative and the perfusion needle. Thefixative was perfused for three to five minutes andpieces of the kidneys then excised and kept in the samefixative at 4°C for 24 hr or more. The tissue was rinsedin 0.1M cacodylate buffer containing 4% PVP and post-fixed in 1% 0s04 for one hour, dehydrated in ethanoland embedded in epoxy resin (Epon 812). Sections werecut on an ultratome (LKB Ultratome III). To identifythe initial part of the proximal tubule, long series ofsemithin sections were cut from tissue blocks from allcortical levels. Within some of these series, ultrathinsections were cut for electron microscopy. Semithinsections were stained with toluidine blue. Ultrathinsections were stained with uranyl acetate and leadcitrate and examined with an electron microscope(Siemens Elmiskop 1A or JEOL JEM bOB).
Results
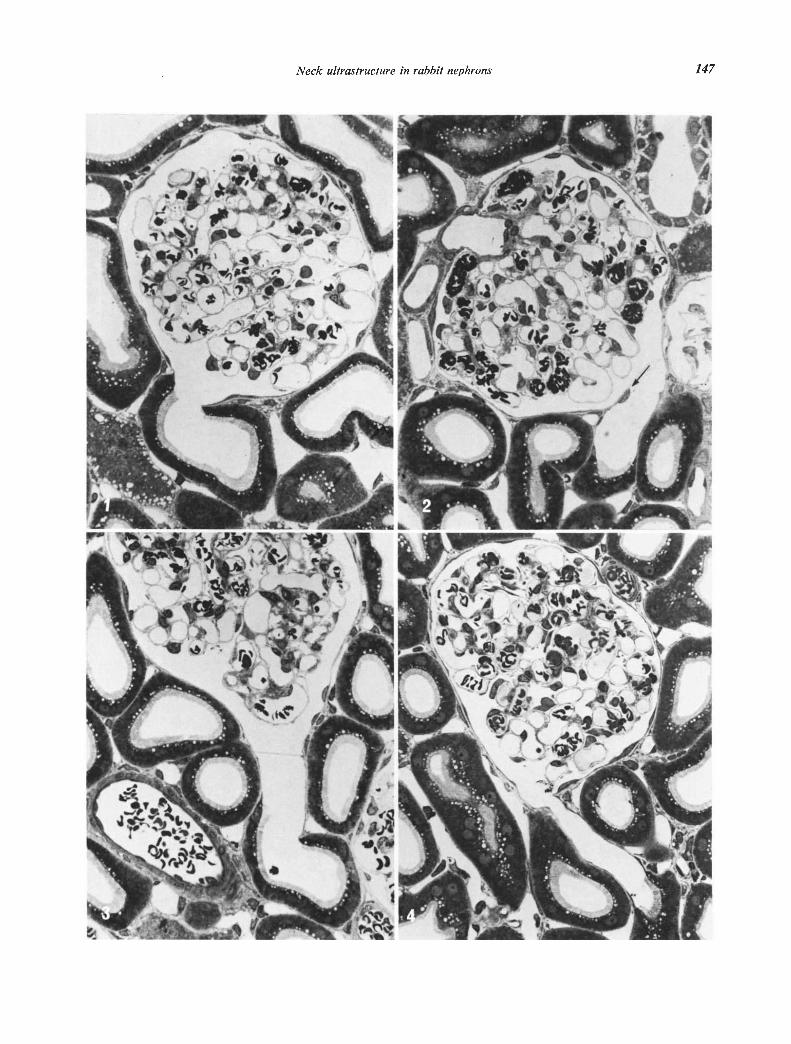
The location of the transition from the parietal layerof the capsule of Bowman to the proximal tubule isvariable in the rabbit kidney (Figs. 1—4). Occasionallythe typical epithelium of the proximal tubule beginsprecisely at the urinary pole of the glomerulus (Fig. 1).In other nephrons this transition occurs within thetubule at some distance from its beginning, thus form-ing a cytologically specialized initial segment of thetubule (Figs. 2—5). We find it convenient to call thisinitial part of the tubule the neck segment, although itis not homologous to the ciliated neck segment of sub-mammalian classes and does not necessarily representa constriction of the tubule.
Fifty-five transitions between the parietal epitheliumof Bowman's capsule and the proximal tubule wereidentified by light microscopy in serial sections thatpassed closely parallel to the long axis of the initialpart of the tubule. All transitions which were not posi-tively identified by serial sectioning were excluded from
analysis. The sections were obtained from blocks ran-domly selected from all parts of the cortex in five ani-mals. The length of the neck segment was variable(Figs. 2—4) and ranged up to a maximum of 90 i. Eachrabbit exhibited a wide range of different types oftransitions. A neck segment was completely absent inonly five nephrons (Fig. 1), and in a few nephrons therewere only a few scattered flat cells at the transition.
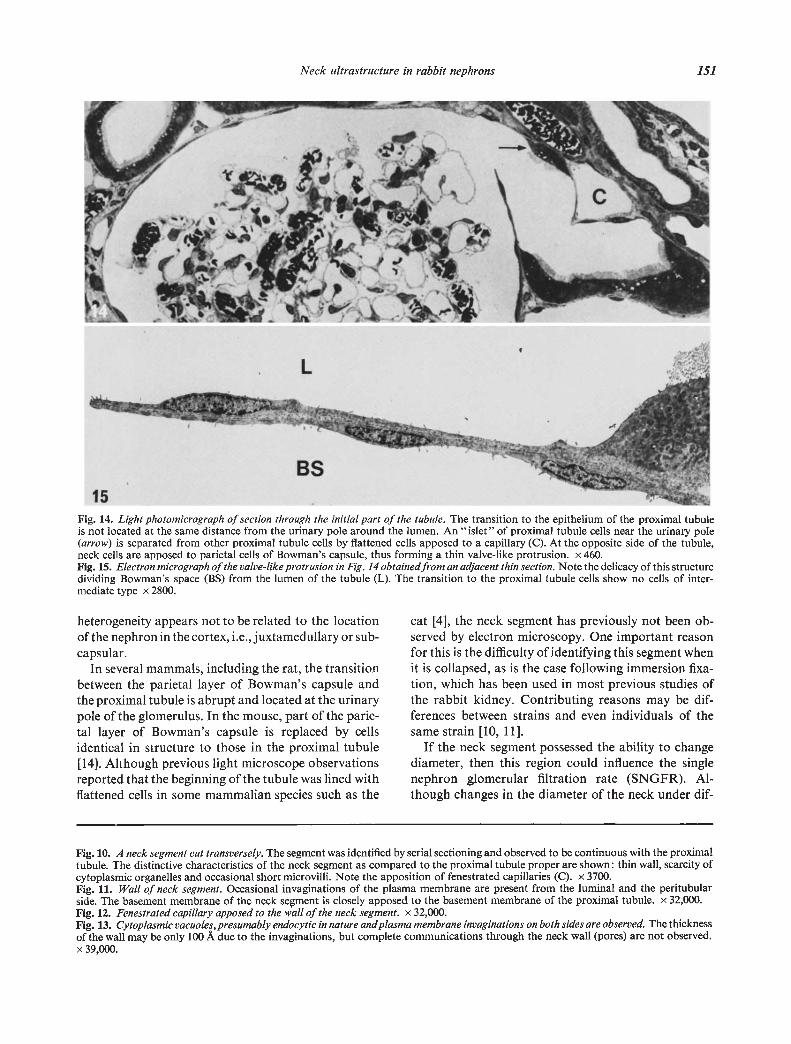
It was repeatedly noted that the axis of the neck seg-ment was oriented tangential to the glomerulus (Figs.2 and 4). Due to tangential orientation, part of thewall of the neck facing the center of the glomerulussometimes formed a valve-like protrusion which separ-ated the space of Bowman and the lumen of the neck(Figs. 2, 14 and 15).
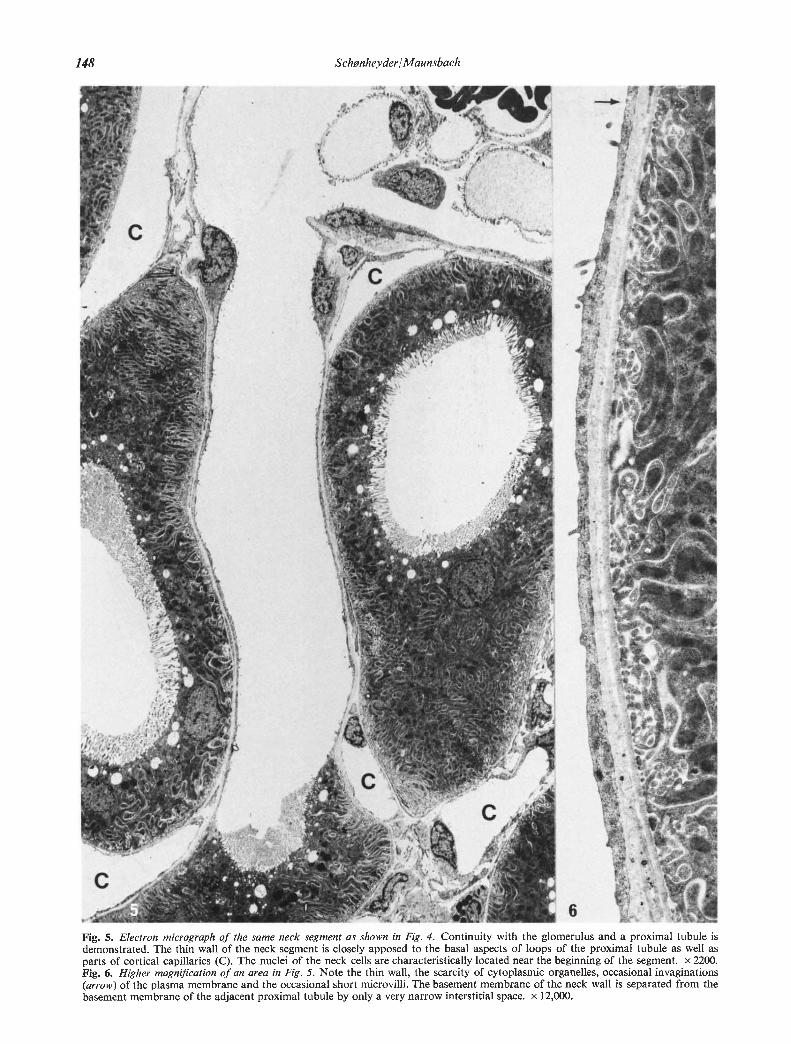
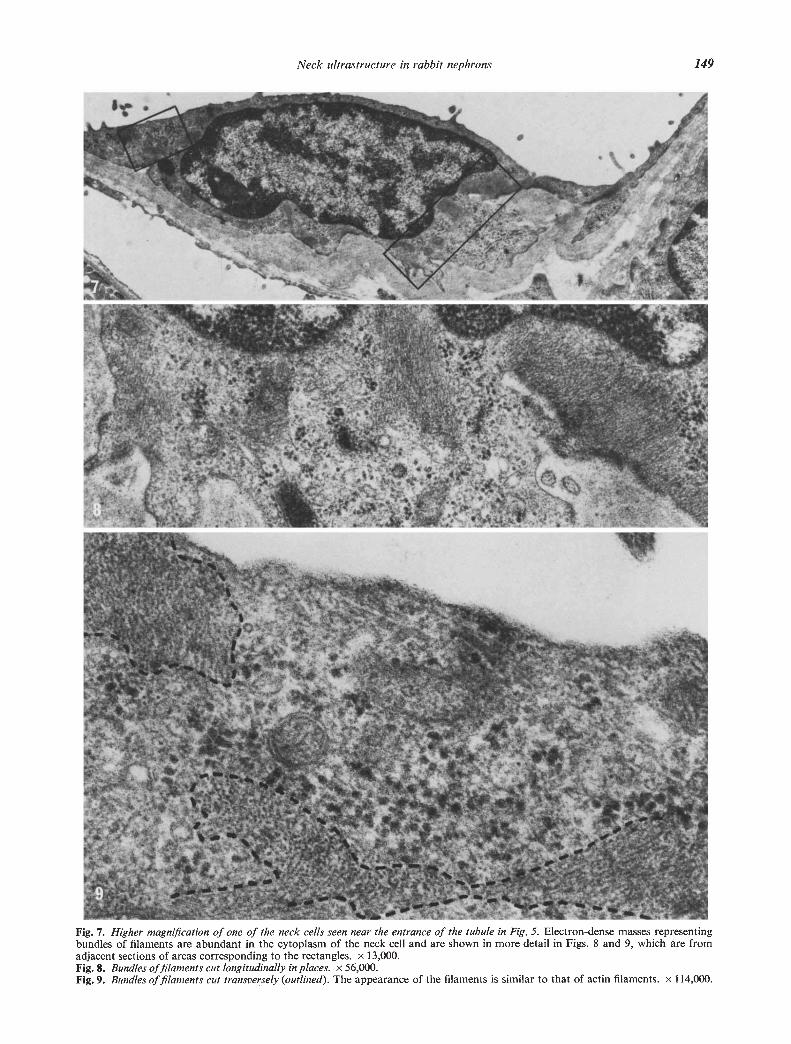
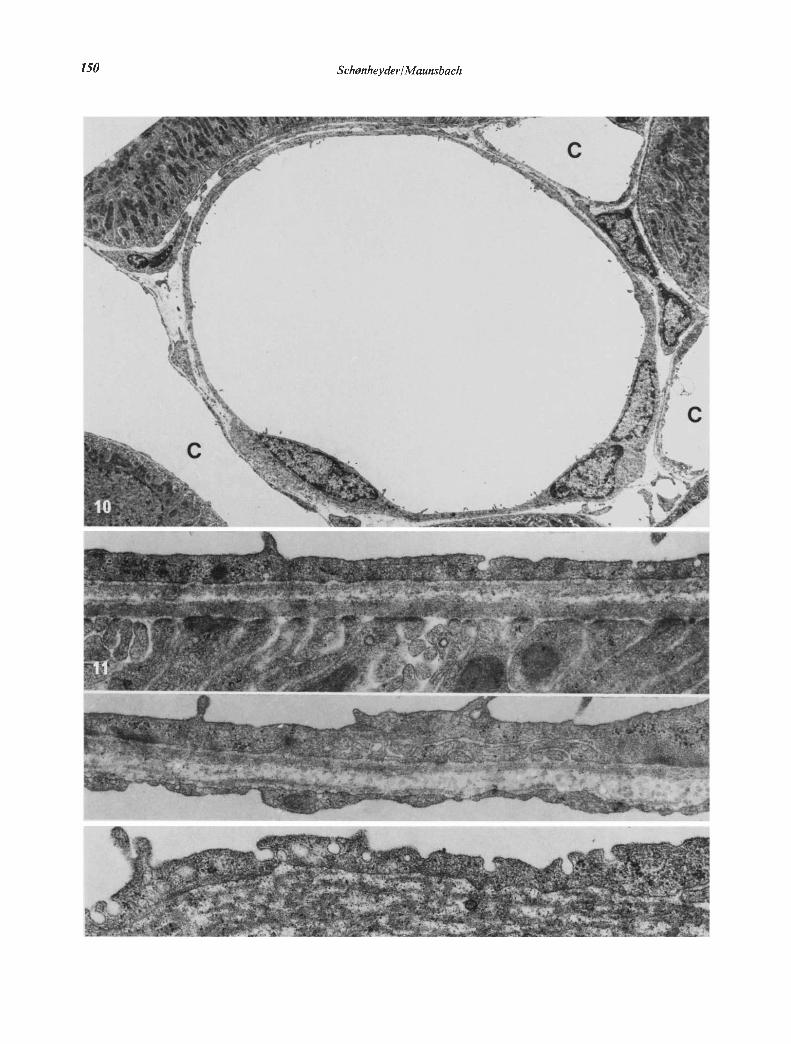
The neck cells were similar in fine structure to thecells of the parietal, epithelium of Bowman's capsule.The nuclei were frequently located at the very begin-ning of the neck (Fig. 5). Beyond the region of thenucleus, the thickness of the cells was 1000 to 3000 A(Figs. 6 and 10). The cytoplasm contained small num-bers of the usual cell organelles including microtubulesand, in addition, exhibited filaments with a thicknessof 50 to 80 A (Figs. 7—9). In many parts of the cellsthese filaments formed thick bundles which were oftenoriented at right angles to the axis of the tubule. Thebundles of filaments were particularly prominent in thecytoplasm adjacent to the nuclei.
The cytoplasm in many regions contained smallcytoplasniic vesicles. In the same regions the cell mem-brane formed small invaginations, mainly from theluminal side but also from the peritubular side (Figs. 11and 13). As a result the cytoplasmic layer which separ-ated apposing cell membrane was sometimes onlyabout 100 A in thickness. The basement membranewas similar to that of the proximal tubule. Capillariesapposed to the neck segment usually showed a fene-strated endothelium (Fig. 12).
Discussion
The present observations provide clear evidencefor structural heterogeneity among nephrons withrespect to the beginning of the tubule. However, this
Figs. 1 to 4. Light photomicrographs of sections of the initial part of proximal tubules oriented close to the long axis of the tubule.Erythrocytes are present in renal vessels because some blood was allowed to flow into the kidneys during the late phase of perfusionfixation to act as markers of the arterial tree. Fig. 1. The epithelium of this proximal tubule begins directly at the urinary pole of theglomerulus. The transition from the cells of the parietal layer of Bowman's capsule is abrupt with no intermediate cell types. x 460.Fig. 2. The epithelium of this proximal tubule begins about 35 from the entrance of the tubule. The initial part of the tubule is linedwith flattened cells and forms a distinct neck segment. Due to the tangential orientation of the neck segment in relation to the glomerulusthe neck epithelium is directly apposed to the outside of Bowman's capsule, thus forming a thin, valve-like protrusion (arrow). x 460Fig. 3. A neck segment is present between the glomerulus and the proximal tubule but no valve-like protrusion or constriction is observedx 460. Fig. 4. Long neck segment measuring about 75g. The transition from flattened neck cells to proximal tubule cells is abrupt. x 460.
Neck ultrastructure in rabbit nephrons 147
I-
—
—
_ A
I
N
'1
'.4 I
148 Schanheyder/Maunsbac/z
Fig. 5. Electron micrograph of the same neck segment as shown in Fig. 4. Continuity with the glomerulus and a proximal tubule isdemonstrated. The thin wall of the neck segment is closely apposed to the basal aspects of loops of the proximal tubule as well asparts of cortical capillaries (C). The nuclei of the neck cells are characteristically located near the beginning of the segment. x 2200.Fig. 6. Higher magnification of an area in Fig. 5. Note the thin wall, the scarcity of cytoplasmic organelles, occasional invaginations(arrow) of the plasma membrane and the occasional short microvilli. The basement membrane of the neck wall is separated from thebasement membrane of the adjacent proximal tubule by only a very narrow interstitial space. x 12,000.
-_;,.
i4'M
--
— -
I
C)
I -
—
4
——
C-
'I
a
—ç
e -1
Neck ultrastructure in rabbit nephrons 149
Fig. 7. Higher magnification of one of the neck cells seen near the entrance of the tubule in Fig. 5. Electron-dense masses representingbundles of filaments are abundant in the cytoplasm of the neck cell and are shown in more detail in Figs. 8 and 9, which are fromadjacent sections of areas corresponding to the rectangles. x 13,000.Fig. 8. Bundles of filaments cut longitudinally in places. x 56,000.Fig. 9. Bundles of filaments cut transversely (outlined). The appearance of the filaments is similar to that of actin filaments. x I 14,000.
'I T
4'!I
3--.
•
F.
1
S
150 Schønlieyder/Maunsbach
-
C
a
Neck ultrastructure in rabbit nephrons 151
Fig. 14. Light photomicrograph of section through the initial part of the tubule. The transition to the epithelium of the proximal tubuleis not located at the same distance from the urinary pole around the lumen. An "islet" of proximal tubule cells near the urinary pole(arrow) is separated from other proximal tubule cells by flattened cells apposed to a capillary (C). At the opposite side of the tubule,neck cells are apposed to parietal cells of Bowman's capsule, thus forming a thin valve-like protrusion. x 460.Fig. 15. Electron micrograph of the valve-like protrusion in Fig. 14 obtained from an adjacent thin section. Note the delicacy of this structuredividing Bowman's space (BS) from the lumen of the tubule (L). The transition to the proximal tubule cells show no cells of inter-mediate type x 2800.
heterogeneity appears not to be related to the locationof the nephron in the cortex, i.e., juxtamedullary or sub-capsular.
In several mammals, including the rat, the transitionbetween the parietal layer of Bowman's capsule andthe proximal tubule is abrupt and located at the urinarypole of the glomerulus. In the mouse, part of the pane-tal layer of Bowman's capsule is replaced by cellsidentical in structure to those in the proximal tubule[14]. Although previous light microscope observationsreported that the beginning of the tubule was lined withflattened cells in some mammalian species such as the
cat [4], the neck segment has previously not been ob-served by electron microscopy. One important reasonfor this is the difficulty of identifying this segment whenit is collapsed, as is the case following immersion fixa-tion, which has been used in most previous studies ofthe rabbit kidney. Contributing reasons may be dif-ferences between strains and even individuals of thesame strain [10, 11].
If the neck segment possessed the ability to changediameter, then this region could influence the singlenephron glomerular filtration rate (SNGFR). Al-though changes in the diameter of the neck under dif-
Fig. 10. A neck segment cut transversely. The segment was idçntified by serial sectioning and observed to be continuous with the proximaltubule. The distinctive characteristics of the neck segment as compared to the proximal tubule proper are shown: thin wall, scarcity ofcytoplasmic organelles and occasional short microvilli. Note the apposition of fenestrated capillaries (C). x 3700.Fig. 11. Wall of neck segment. Occasional invaginations of the plasma membrane are present from the luminal and the peritubularside. The basement membrane of the neck sement is closely apposed to the basement membrane of the proximal tubule. x 32,000.Fig. 12. Fenestrated capillary apposed to the wall of the neck segment. x 32,000.Fig. 13. Cytoplasmic vacuoles,presumably endocytic in nature andplasma membrane invaginations on both sides are observed. The thicknessof the wall may be only 100 A due to the invaginations, but complete communications through the neck wall (pores) are not observed.x 39,000.
L
BS

152 Schonheyder/Maunsbach
ferent functional conditions have yet to be demon-strated, the presence of both the filaments in the neckcells and the valve-like protrusions provide some basisfor speculation. The filaments observed in the neckcells are similar in structure to filaments observed inother types of kidney cells, including cells of the pane-tal layer of Bowman's capsule [15—19] and proximaltubule cells [6, 16—18, 20]. In the neck cells it is parti-cularly intriguing that many filament bundles are pre-sent in cells located at the very beginning of the necksegment and that they are oriented more or less at rightangles to the axis of the tubule. If the filaments arecontractile, they may function in much the same way asa sphincter. The possibility that the filaments are con-tractile in nature is suggested by their similarity in boththickness and arrangement to actin filaments that havebeen identified in other kidney cells [20]. However,direct evidence to support this postulated functionalrole is lacking at this time. Alternatively, it is also quitepossible that the filaments are actually noncontractilein nature and serve to prevent closure of the valve-likeregion at the neck by acting as a "cytoskeleton" [19].The valve-like protrusions observed in some nephronsas the common wall of the neck segment and the panic-tal layer of Bowman's capsule may narrow the entranceto the tubule at high pressures in the space of Bowman.
The presence of a neck segment, cytologically quitedifferent from the proximal tubule, in the beginning ofthe tubule may also be of significance for the transportof water and solutes across the tubule wall. Since necksegment cells contain only few mitochondria, it appearsunlikely that the cells have much active transport.However, endocytic activity is suggested by the cyto-plasmic vacuoles and cell membrane invaginations atthe luminal and the peritubular sides of the cells. Al-though unproved, the possibility exists that the vacu-oles and invaginations form part of a transport systemacross the tubule wall corresponding to the large poresystem in nonfenestrated capillaries [12]. It is alsopossible that the neck cells offer a different resistanceto passive transport across the tubule wall than doother tubule cells. In this context it is of interest thatthe neck segment is often closely apposed by fenes-trated capillaries.
The well known lability of the rabbit glomerularfiltration rate to various stimuli, such as apnea, isgenerally considered a vasomotor response [12]. Theexistence of neck segments containing filament-richcells may suggest a greater complexity of the renal re-sponse in the rabbit.
Acknowledgments
This work was supported by grants from the DanishMedical Research Council and by a scholarship from
the University of Aarhus to H. C. Schonheyder. Dr.G. B. de Mello, provided information about his pro-cedure to anesthetize rabbits. Mrs. Marianne Ellegaardprovided technical assistance.
Reprint requests to Dr. Henrik C. Schønheyder, Department ofCell Biology, Institute of Anatomy, University of Aarhus, Aarhus,Denmark.
References
I. EDWARDS JO, SCHNInER C: The renal unit in the kidney ofvertebrates. AmJAnat 53:55—87, 1933
2. BARGMANN W, KNOOP A, ScmaELaR TH: Histologische,cytochemische und elektronenmikroskopische Untersuch-ungen am Nephron (mit Berticksichtigung der Mitochond-rien). Z Zellforsch Mikrosk Anat 42:386-422, 1955
3. ROUTLLER C: General anatomy and histology of the kidney,in The Kidney, Morphology, Biochemistry, Physiology,edited by ROUILLER C, MULLaR AF, New York, AcademicPressmc, 1969, p. 61
4. vori MdLLENDORFF W: Der Exkretionsapparat, in Handbuchder Mikroskopischen Anatomie des Menschen. Berlin,Springer-Verlag, 1930, vol. VII, p. 1
5. TRUMP BF, BULOER RE: Morphology of the kidney, inStructural Basis of Renal Disease, edited by BECKER EL,New York, Hoeber Medical Division of Harper and Row,1968, p. 1
6. MAUNSBACH AB: Ultrastructure of the proximal tubule, inHandbook of Physiology, Section 8: Renal Physiology,edited by ORL0FF J, BERLINER RW, Washington D.C.,American Physiological Society, 1973, p. 31
7. CLAY RD, DARMADY EM, HAWKINS M: The nature of therenal lesion in the Fanconi syndrome. .JPathol Bacteriol 65:551—558, 1953
8. JACKSON JD, SMITH FG, LITMAN NN, YUILE CL, LAnA H:The Fanconi syndrome with cystinosi: Electron microscopy
of renal biopsy specimens from five patients. Am J Med 33:893—910, 1962
9. DARMADY EM: The renal changes in some metabolic
diseases, in The Kidney, edited by MosloFI FK, SMITH DE,Baltimore, The Williams and Wilkins Company, 1966,
p. 253
10. PETER K: Die Nierenkanaichen des Menschen und einigerShugetiere, in Untersuchungen ilber Bau und Entwickelungder Niere, lena, Fischer, 1909
11. SPERBER I: Studies on the mammalian kidney. Zoo BidragUppsala 22:249—431, 1944
12. FOR5TER RP: Comparative vertebrate physiology and renalconcepts, in Handbook of Physiology, Section 8: Renal
Physiology, edited by ORLOFF J, BERLINER RW, WashingtonD.C., American Physiological Society, 1973, p. 161
13. MAUNsBACH AB: The influence of different fixatives and
fixation methods on the ultrastructure of rat kidney proxi-
mal tubule cells: I. Comparison of different perfusion fixa-
tion methods and of glutaraldehyde, formaldehyde andosmium tetroxide fixatives. J Ultrastruct Res 15:242—282,1966
14. DIETERT SE: The columnar cells occurring in the parietal
layer of Bowman's capsule: Cellular fine structure and pro-
tein transport. J Cell Biol 35:435—444, 196715. ANDERsON WA: The fine structure of compensatory growth
in the rat kidney after unilateral nephrectomy. Am .1 Anat121:217—248,1967

Neck ultrastructure in rabbit nephrons 153
16. PEASE DC: Myoid features of renal corpuscles and tubules.J Ultrastruct Res 23:304—320, 1968
17. Ross MH, REITH EJ: Myoid elements in the mammaliannephron and their relationship to other specializations in thebasal part of kidney tubule cells. Am J Anat 129:399—416,1970
18. NEWSTEAD JD: Filaments in renal parenchymal and inter-stitial cells. J Ultrastruct Res 34:316—328, 1971
19. WEBBER WA, WONO WT: The function of the basal fila-
ments in the parietal layer of Bowman's capsule. Can JPhysiol Pharmacol 51:53—60, 1973
20. ROSTGAARD J, KRISTENSEN BI, NIELSEN LE: Electron micros-copy of filaments in the basal part of rat kidney tubule cells,and their in situ interaction with heavy meromyosin. ZZellforsch Mikrosk Anat 132:497—521, 1972
21. BRUNS RR, PALADE GE: Studies on blood capillaries: II.Transport of ferritin molecules across the wall of musclecapillaries. J Cell Biol 37:277—299, 1968

![Practice For May: Cell Ultrastructure [114 marks]blogs.4j.lane.edu/.../2018/02/Cell-Ultrastructure-Test-1.pdfPractice For May: Cell Ultrastructure [114 marks]1. Which structure found](https://img.pdfslide.net/doc/110x75/5eda4db5b3745412b5711d9c/practice-for-may-cell-ultrastructure-114-marksblogs4jlaneedu201802cell-ultrastructure-test-1pdf.jpg)






![Identification and Characterization of the Rhoptry Neck ... · rective 86/609/EC]) as fully described elsewhere (27). Rabbit sera were producedbyGenmedSynthesis,Inc.,SanAntonio,TX(OLAWassurance](https://img.pdfslide.net/doc/110x75/5f3ceb80403206049d243b5f/identification-and-characterization-of-the-rhoptry-neck-rective-86609ec.jpg)




















