Embed Size (px)
Citation preview

J. Cell Sci. 13, 69-81 (1973) 69
Printed in Great Britain
ULTRASTRUCTURE OF MEIOSIS AND
CENTRIOLE BEHAVIOUR IN ULVA
MUTABILIS F0YN
T. BRATENElectron Microscopical Unit for Biological Sciences,University of Oslo, Blindern, Oslo 3, Norway
AND 0. NORDBYDepartment of Zoology, University of Oslo, Blindern, Oslo 3, Norway
SUMMARY
The present work deals with the ultrastructure of the meiotic processes leading to theformation of zoospores. The formation of an exit pore on the outer surface of the sporangiumis the first visible sign that the cell will undergo meiosis and not just a somatic division. Pro-phase I nuclei differ from nuclei in mitotic prophase by having one or several invaginationsof the nuclear membrane. Electron-dense lines probably representing the lateral elements ofthe 8ynaptinemal complex are observed during prophase I. The central element of the complexhas, however, never been observed.
The fate of the centrioles during meiosis is described in detail. The number of duplicationsof the centrioles is found to be the same at meiotic and mitotic zooid formation. The numberof cell divisions is, however, different in the 2 cases.
INTRODUCTION
Among organisms suitable for the study of specific preparations for meioticor reductional division, the sea-lettuce Ulva and other members of the order Ulo-trichales possess the outstanding property that closely similar meiotic and mitoticzooid formation processes can be compared in division-synchronized diploid andhaploid plants. This fact makes it possible to separate processes concerned with thedifferentiation of reproductive cells, from processes involved with the meiotic processitself.
Ulva mutabilis Foyn has now for several years served as an object for genetical,physiological and morphological studies in our laboratories (Lovlie, 1968). As partof current morphological and biochemical studies of synchronized zooid formation(meiotic and mitotic) in this organism (Nordby & Hoxmark, 1972), the present workcontains a description of some ultrastructural features of the meiotic zoosporogenesis.
Particular attention is paid to the replication of the centrioles, the number ofwhich is 4 per cell in the haploid zoospore and 2 per cell in the haploid gamete.Several light-microscopical investigations exist on meiosis in green algae (e.g. Foyn,1934; Ramanathan, 1939). To the author's knowledge no investigations have, however,been made on the ultrastructure of meiosis, in these plants.

70 T. Brdten and 0. Nordby
MATERIAL AND METHODS
Diploid sporophytes of the fast-growing mutant 'Slender' (Sl/Sl) were used in thisinvestigation. The algae were grown at 18 °C in enriched seawater as described elsewhere(Lovlie, 1969). After approximately 5 weeks' growth in a I7h/7h light/dark programme withthe light period running from 04.00 to 21.00 hours, the algae were harvested at 14.00 hoursand cut into small fragments. The thoroughly washed fragments were transferred to freshgrowth medium in Petri dishes and placed in the sporulation chamber. This chamber has atemperature of 21 °C, and an illumination of 10-500 lux measured at dish centre. Here thelight period in the 17 h/7 h light/dark programme runs from 21.00 to 14.00 hours. Underthese conditions more than 90 % of the cells in the fragments will regularly carry out syn-chronous meiosis 40 h later (Nordby & Hoxmark, 1972).
Specimens for light microscopy were fixed in Bouin-Dubosque's solution (Lovlie, 1964)and stained in Gomori's haematoxylin according to Melander & Wingstrand (1953).
Preparation for electron microscopy was as described in a previous paper (Braten, 1971).
RESULTS
General
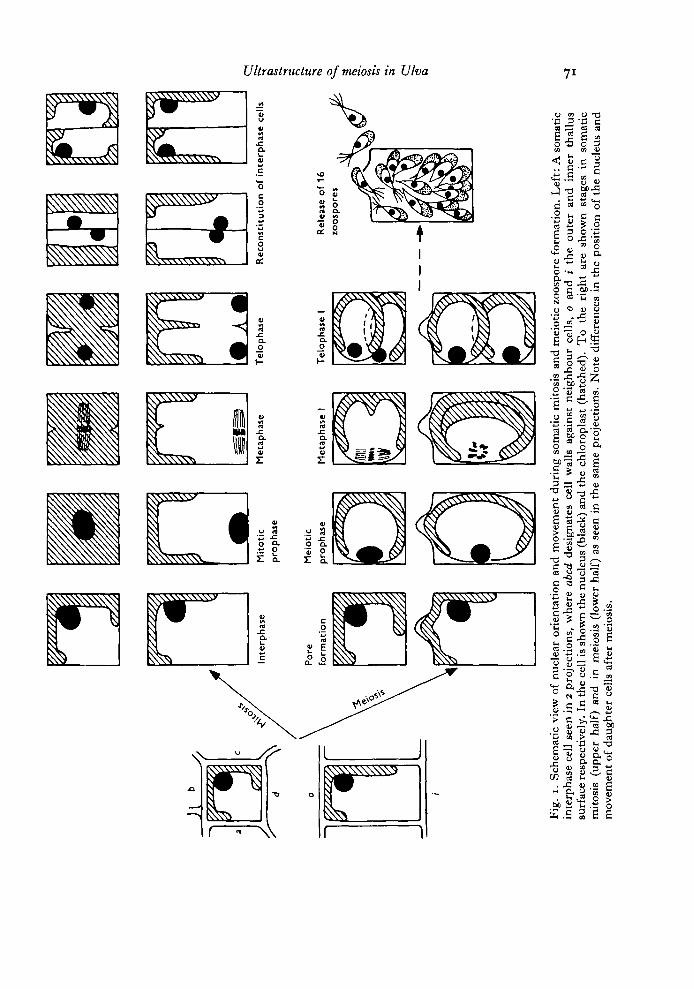
Principally JJlva grows as a hollow tube, the wall of the tube being made up of asingle coherent layer of cells. As described by Lovlie & Braten (1970) and indicatedin Fig. 1, the somatic mitosis is initiated by a wandering of the cell nucleus towardsthe inner surface of the cell. The nucleus divides while lying close to this wall, andalways with its spindle axis parallel to the inner surface. After completed division theformed daughter nuclei approach each other and glide outwards along the newlyformed division furrow to take up their interphase position in the vicinity of thecup-shaped chloroplast. A new cell wall is then laid down vertical to the surfaceplane. A detailed light-microscopical description of the meiotic division is in prepara-tion. The result is summarized as follows:
Meiotic formation of motile zoospores deviates in several respects from somaticmitosis. The first sign of zooid formation is the appearance of a bulge on the outersurface of the cell. This will later become the exit pore for the fully formed spores.Shortly thereafter the cells round up within the cell wall envelopes, apparentlylosing their turgor. The meiotic division is also initiated by a wandering of thenucleus from its interphase position close to the chloroplast, to a position close tothe cell wall opposite the chloroplast. This cell wall is, however, not the inner surfacewall as in mitosis. In meiosis the nucleus takes up a position at the centre of thewall shared with the neighbouring cell. The spindle axis lies parallel with the outerand inner surface plane, but already during the anaphase the spindle and the formingdaughter cells rotate so (hat they end up lying on top of each other (Fig. 1). Asduring mitosis the division furrow appears as a ring around the whole cell.
After the completion of the first division, essentially the same division and move-ment patterns are repeated 3 more times giving rise to 16 daughter cells within thecell wall of each spore mother-cell.
In the usual light/dark regime used in these experiments meiotic prophase stagesappear some 40 h after transfer to the sporulation chamber. The 16 zoospores persporangium are formed within the next 12 h, and they will be released through theexit pore at some time during the next light period (Fig. 12).
I IIM
ito
tic
pro
ph
ase
Mei
otic
prop
haie
Met
aph
ase
Met
aph
ase
I
Tel
opha
se
Tel
op
has
e I
3
Rec
on
stlt
uti
on
of
inte
rph
ase
cells
Rel
ease
of
16zo
osp
ore
s
3 I I"
Fig.
i.
Sche
mat
ic v
iew
of
nucl
ear
orie
ntat
ion
and
mov
emen
t du
ring
som
atic
mito
sis
and
mei
otic
zoo
spor
e fo
rmat
ion.
Lef
t: A
som
atic
inte
rpha
se c
ell
seen
in
2 pr
ojec
tions
, w
here
abe
d de
sign
ates
cel
l w
alls
aga
inst
nei
ghbo
ur c
ells
, 0
and
i th
e ou
ter
and
inne
r th
allu
ssu
rfac
e re
spec
tivel
y. I
n th
e ce
ll is
show
n th
e nu
cleu
s (b
lack
) and
the
chlo
ropl
ast
(hat
ched
). T
o th
e ri
ght
are
show
n st
ages
in
som
atic
mito
sis
(upp
er h
alf)
and
in
mei
osis
(lo
wer
hal
f) a
s se
en i
n th
e sa
me
proj
ectio
ns.
Not
e di
ffer
ence
s in
the
pos
ition
of
the
nucl
eus
and
mov
emen
t of
dau
ghte
r ce
lls a
fter
mei
osis
.

72 T. Brdten and 0. Nordby
Formation of the exit pore
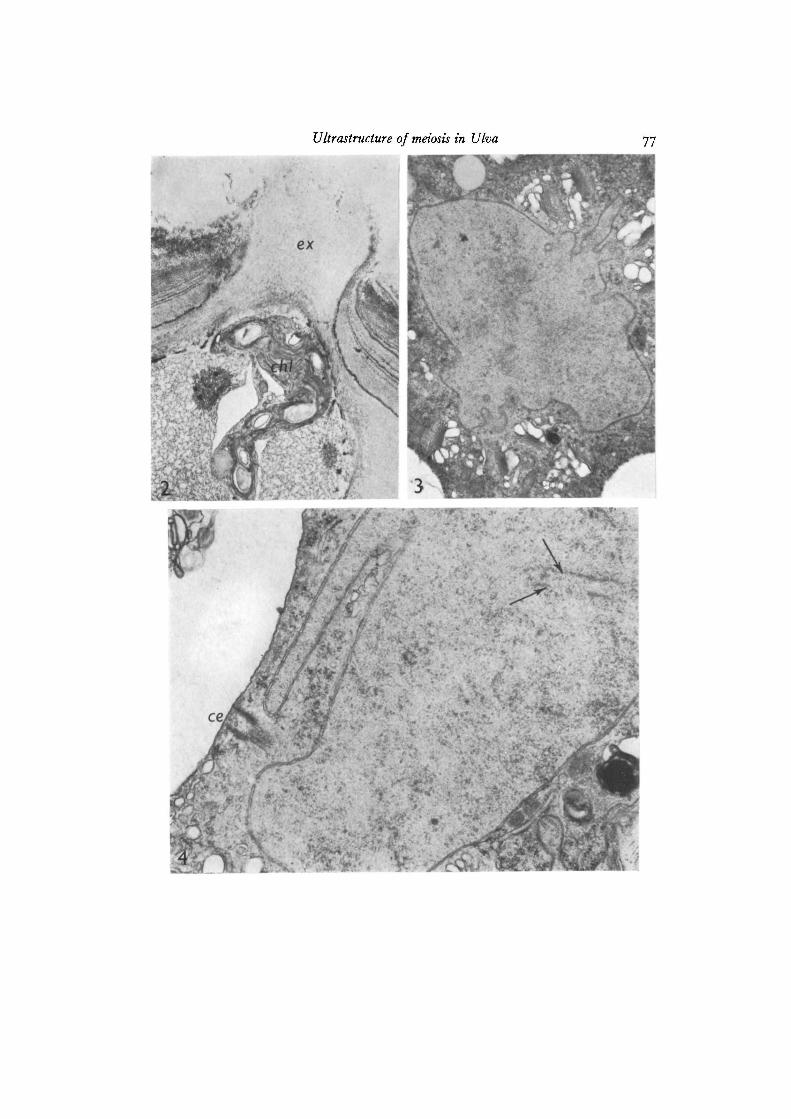
The exit pore is always formed on the outer surface of the sporangia. Onset ofspore formation is the first visible sign that the cell will undergo meiosis and notjust a somatic division. Spore formation starts by an outward bulging of the outercell wall. At the same time a weakening of the wall at this place can be seen (Fig. 2).On electron micrographs this weakening is seen as less densely packed and morerandomly oriented fibrils than in the rest of the cell wall. Membrane-bound vesiclesare frequently observed at the site of cell wall weakening.
The nuclear division
The marked difference in structure between the 'synizesis'-like and 'spiremal'stage of meiotic prophase as seen in light-microscope preparations is not encounteredin the corresponding electron micrographs. It is however, a widespread notion thatthe tight 'synizesis knot' is actually a fixation artifact of light microscopy (Rhoades,1961).
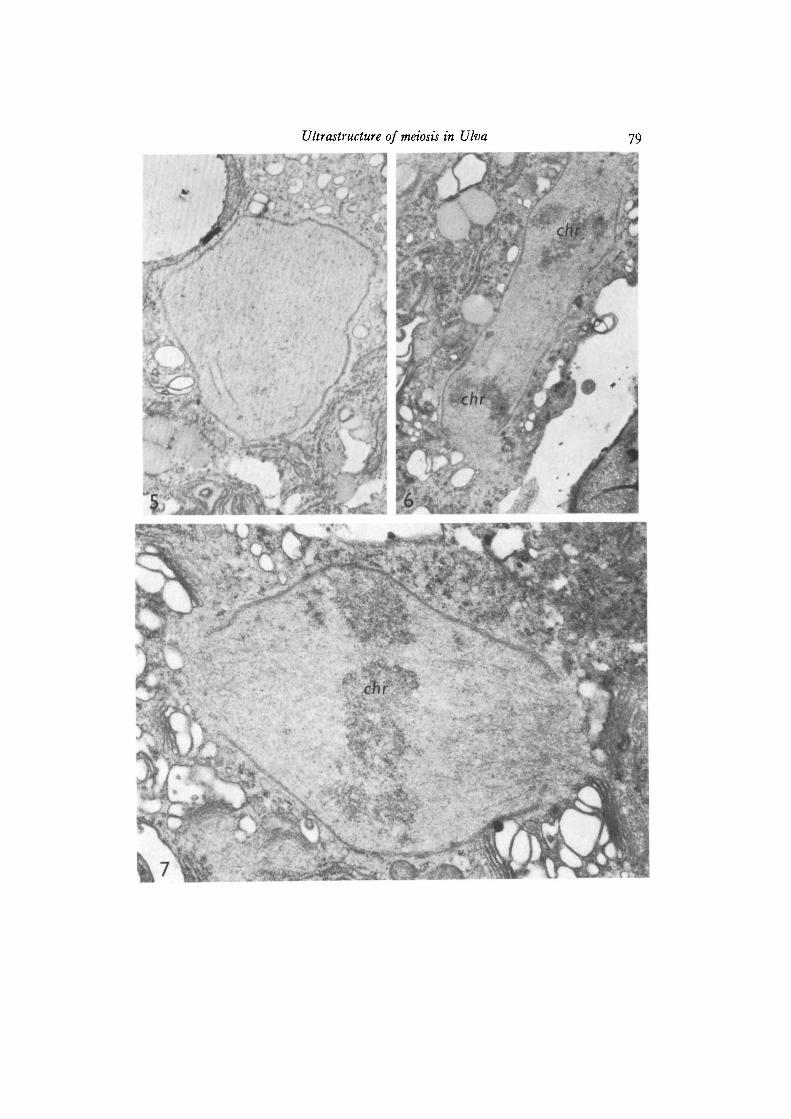
The morphology of the nucleus at prophase I is strikingly different from what isseen at mitosis. The nucleus has an irregular surface due to usually one extensiveinvagination of the nuclear membrane (Fig. 4). Nuclei with large numbers of mem-brane imaginations have also been found (Fig. 3). Characteristically the centriolesoccupy a position at the opening of one of the invaginations of the nuclear membrane.Microtubules are found in the cytoplasm of this invagination. One other characteristicof the prophase I nucleus is the appearance of electron-dense lines often found tobe paired (Figs. 4, 5). Each line measures approximately 30 run in width and the 2paired lines are separated by a space of 0-15-0-2 /tm. We think that these lines rep-resent the lateral element of the synaptinemal complex (see Moses, 1968, for termi-nology). Unpaired lines may represent the so-called axial elements thought to beearly formation stages of the synaptinemal complex. We have, in spite of extensivesearch, never observed complete synaptinemal complexes containing the centralelement.
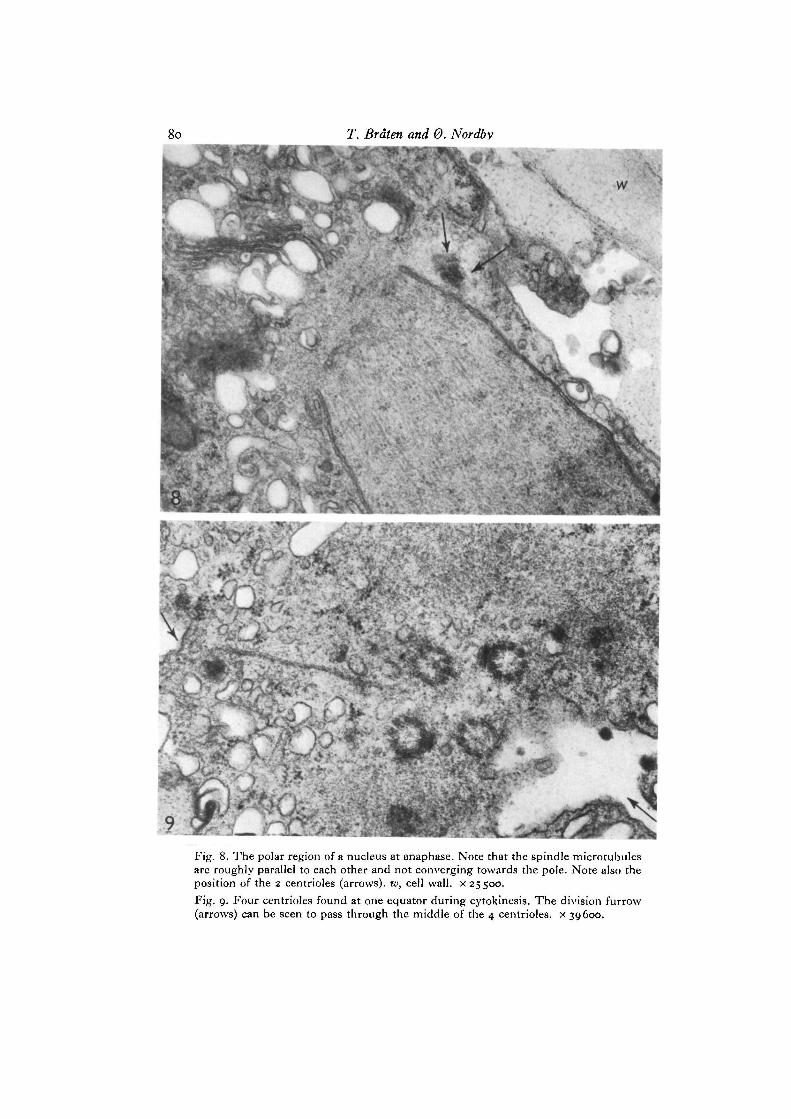
The metaphase picture is the same as seen during mitosis. The nuclear surfacehas regained a smooth appearance except at the 2 poles where large fenestrae can beseen (Fig. 7). The polar region is a homogeneous mass of cytoplasm devoid ofribosomes and other cell organelles. Spindle microtubules are numerous and seemto stop rather abruptly at the level of the openings in the nuclear envelope (Fig. 8).
Anaphase proceeds without any sign of cytokinesis (Fig. 6). The nucleus takes theshape of a long cylinder open at both ends, but with the nuclear envelope intact overthe rest of the surface.
The behaviour of the centrioles during division and cytokinesis
When the centrioles are first observed during meiosis they lie in the narrow spacebetween the prophase nucleus and the cell wall (Fig. 4). As mentioned in the previoussection, when an invagination develops into the interior of the nucleus the centriolesare found just outside this invagination. In this position the centrioles duplicate.

Ultrastructure of mewsis in Ulva 73
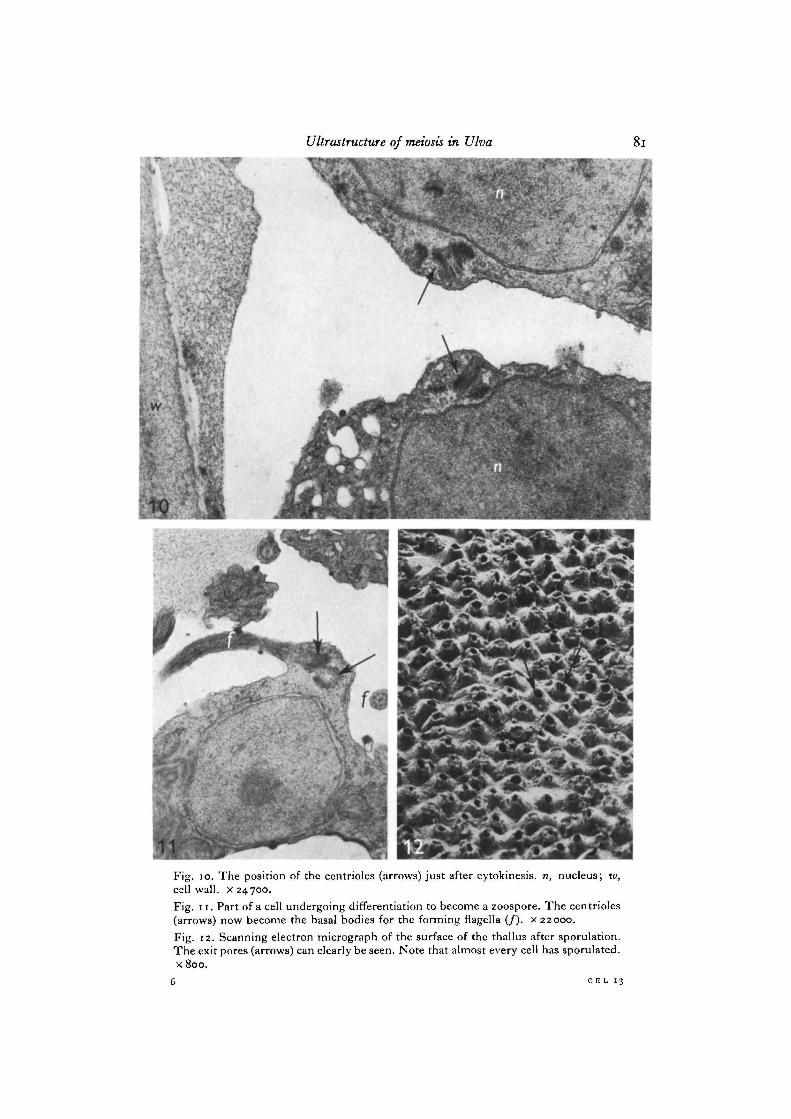
The 2 daughter pairs now move towards the spindle polar areas and are found tolie close to this area during metaphase. The position of the centrioles at early anaphaseis to one side of the polar region between the nucleus and the cell wall (Fig. 8).We have no pictures showing the movement of the centrioles during ana- and telo-phase. When reconstituted nuclei are found after telophase, however, these lie closetogether. Between the nuclei all 4 centrioles may be found packed closely together.Figs. 9 and 10 clearly show that the subsequent division furrow will cut right throughthe centre of the centriole group, distributing 2 centrioles to each daughter cell.This equatorial position with the 4 centrioles close together is interpreted as beingdue to the fact that the 2 nuclei start to rotate and move together immediately afterreconstitution (0. Nordby, in preparation). The same pattern of centriole duplicationand movement is seen during the following 3 mitotic divisions. It is to be notedhere that a distinct interphase stage appears between each of these divisions, andthat the centrioles duplicate before any sign of nuclear division is apparent.
Subsequent to the last nuclear division, an additional centriole duplication takesplace, thus furnishing each future zoospore with 4 centrioles. These remain in right-angled pairs, and are seen to lie in a distinct bulge on the somewhat elongated cell.These 4 centrioles then form the basal bodies for the 4 flagella which emerge duringthe subsequent differentiation of the zoospores (Fig. 11). Usually the 16 zoosporesform a hollow sphere inside the sporangium with their flagella pointing towards eachother.
DISCUSSION
The irregularly shaped nuclei with one or several extensive invaginations of thenuclear membrane are characteristic of meiotic prophase. Nuclei undergoing ordinarymitosis show ovoid nuclei with a smooth surface (Lovlie & Braten, 1970). We havenot observed attachment of chromosomes to the invagination(s) of the nuclear mem-brane at prophase, as described in some organisms (e.g. Underbrink, Ting & Sparrow,1967; Moens & Perkins, 1969).
We are puzzled by the fact that no complete synaptinemal complex has beenfound, this, in spite of the extensive search through large numbers of prophase Inuclei. One sees electron-dense parallel lines, but the central element of the complexhas never been observed. Moses (1968) states that the appearance of the centralelement varies widely among species and even within an individual. Our failure toobserve it can hardly be due to extremely short duration of this particular stage.It is possible that our method of preparation fails to stain the central element. Thelow affinity of Ulva tissue for osmium is a general problem in preparation for electronmicroscopy. This for instance leads to difficulties in identifying hetero- and euchro-matic regions of chromosomes in ordinary vegetative tissue. Our measurements ofthe size of the synaptinemal complex lie within those reported from other organisms(Moses, 1968).
Two pairs of centrioles exist throughout meiosis except for early interphase.The position of the centrioles at anaphase, somewhat to one side of the polar region,

74 T. Brdten and 0. Nordby
confirms observations made from vegetative cells that centrioles do not take directpart in the formation of the spindle (Lovlie & Braten, 1970). Recent investigations(Szollosi, Calarco & Donahue, 1972) have shown that the presence of centrioles isnot necessary for the formation of meiotic spindles in mouse oocytes.
The position of the 4 centrioles at the equator during cytokinesis strongly con-firms the suggestion made by Johnson & Porter (1968) that the centrioles determinethe position of the cleavage plane. This is particularly evident in Fig. 9 where thecleavage furrow can be seen to pass right between the 4 centrioles.
The interpretation that the flagella are formed at the 16-cell stage is somewhatcircumstantial, since we have no serial sections of this stage. However, since flagellaare never seen in dividing cells, this interpretation appears to be the most probable-
The number of flagella is, apart from the difference in size, the only way of dis-tinguishing between zoospores and gametes on a morphological basis. Interestingin this connexion is the fact that each gametangium apparently produces 32 bi-flageUated gametes while each sporangium produces 16 quadrinagellated zoospores.This means that after the last centriole duplication, the future gametes undergoanother division, leaving each cell with 2 centrioles to become the basal bodies of the2 flagella. In the case of the zoospores no division takes place after the last centrioleduplication; thus each cell is given 4 centrioles to become the basal bodies of the 4flagella. In other words: the number of duplications of the centrioles is the sameat meiotic and mitotic zooid formation. The number of cell divisions is, however,different in the 2 cases.
We wish to thank Mrs Eva Jenssen for skilled technical assistance. Financial support fromRoyal Norwegian Council for Scientific and Industrial Research is gratefully acknowledged.
REFERENCES
BRATEN, T. (1971). The ultrastructure of fertilization and zygote formation in the green algaUlva mutabilis Foyn. J. Cell Set. 9, 621-635.
FOYN, B. (1934). Lebenszyldus, Cytologie und Sexualitat der Chlorophycee Ulva lactuca L.Arch. Protistenk. 83, 154-177.
JOHNSON, U. G. & PORTER, K. R. (1968). Fine structure of cell division in Chlamydoinonasreinliardi. Basal bodies and microtubules. J. Cell Biol. 38, 403-425.
LOVLIE, A. (1964). Genetic control of division rate and morphogenesis in Ulva mutabilis Foyn.C.r. Trav. Lab. Carhberg 34, 77-168.
LOVLIE, A. (1968). On the use of a multicellular alga {Ulva mutabilis Foyn) in the study ofgeneral aspects of growth and differentiation. Nytt Mag. Zool. 16, 39—49.
LeVLiE, A. (1969). Cell size, nucleic acids and synthetic efficiency in the wild type and agrowth mutant of the multicellular alga Ulva mutabilis Foyn. Devi Biol. 20, 349-367.
LevuE, A. & BRATEN, T. (1970). On mitosis in the multicellular alga Ulva mutabilis Foyn.J. Cell Set. 6, 109-129.
MELANDER, Y. & WINGSTRAND, K. G. (1953). Gomori's hematoxylin as a chromosome stain.Stain Technol. 28, 217-223.
MOENS, P. B. & PERKINS, F. O. (1969). Chromosome number of a small protist: Accurate deter-mination. Science, N.Y. 166, 1289-1291.
MOSES, M. J. (1968). Synaptinemal complex. A. Rev. Genet. 2, 363-412.NORDBY, 0. & HOXMARK, R. C. (1972). Changes in cellular parameters during synchronous
meiosis in Ulva mutabilis Foyn. Expl Cell Res. 75, 321-328.RAMANATHAN, K. R. (1939). The morphology, cytology and alternation of generations in Entero-
morpha compressa (L) Grev. var. lingulata (J. AG.) Hauck. Ann. Bot. 3, 375-398.

Ultrastructure of meiosis in Ulva 75
RHOADES, M. M. (1961). Meiosis. In The Cell, vol. 3 (ed. J. Brachet & A. E. Mirsky), pp. 1-76.New York and London: Academic Press.
SZOLLOSI, D., CALARCO, P. & DONAHUE, R. P. (1972). Absence of centrioles in the first andsecond meiotic spindles of mouse oocytes. J. Cell Set. 11, 521-541.
UNDERBRINK, A. G., TING, Y. C. & SPARROW, A. H. (1967). Note on the occurrence of a synap-tinemal complex at meiotic prophase in Zea mays. Can. J. Genet. Cytol. 9, 606-609.
(Received 6 December 1972)

76 T. Brdten and 0. Nordby
Fig. 2. Part of a cell in an eaily stage of meiosis, but showing an advanced state ofexit pore formation. Part of the chloroplast (cJit) can be seen close to the future exitpore (ex), x IOIOO.
Fig. 3. Prophase I nucleus showing a highly irregular surface. X 14300.Fig. 4. Part of a prophase I nucleus showing an extensive invagination of the nuclearmembrane. One centriole (ce) can be seen just outside this invagination. Arrowsindicate lines probably representing the synaptinenial complex, x 25 200.
Ultrastructure of meiosis in Ulva
• 1 * \

78 T. Brdlen and 0. Nordby
Fig. 5. Prophase I nucleus showing typical dense lines thought to be the synapti-nemal complex, x 9600Fig. 6. The nucleus at anaphase. Note an intact nuclear envelope except at the poleswhere large fenestrae can be seen. Electron-dense areas (chr) represent chromosomes,x 10200.
Fig. 7. The nucleus at metaphase I. Chromosomes (chr) and spindle microtubulescan be seen, x 28900.
Ultrastructure of meiosis in Ulva
T. Brdten and 0. Nordby
? : ^ _Fig. 8. The polar region of a nucleus at anaphase. Note that the spindle microtubulesare roughly parallel to each other and not converging towards the pole. Note also theposition of the 2 centrioles (arrows), w, cell wall, x 25500.Fig. 9. Four centrioles found at one equator during cytokinesis. The division furrow(arrows) can be seen to pass through the middle of the 4 centrioles. x 39600.
Ultraslructure of meiosis in Uha 81
Fig. 10. The position of the centrioles (arrows) just after cytokinesis, n, nucleus; w,cell wall, x 24700.Fig. 11. Part of a cell undergoing differentiation to become a zoospore. The centrioles(arrows) now become the basal bodies for the forming flagella (/). x 22000.Fig. 12. Scanning electron micrograph of the surface of the thallus after sporulation.The exit pores (arrows) can clearly be seen. Note that almost every cell has sporulated.x 800.
.& CI5L 13