Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE CIÊNCIAS BIOLÓGICAS DEPARTAMENTO DE BIOQUÍMICA
MESTRADO EM BIOQUÍMICA
ROBERTA LUCIANA DO NASCIMENTO GODONE
ISOLAMENTO DE UMA QUITINASE EXTRAÍDA DO
CEFALOTÓRAX DO CAMARÃO MARINHO Litopenaeus schmitti (BURKENROAD, 1936): AVALIAÇÃO DE SUAS ATIVIDADES
ANTIMICROBIANA E LARVICIDA
NATAL/RN 2011

ROBERTA LUCIANA DO NASCIMENTO GODONE
ISOLAMENTO DE UMA QUITINASE EXTRAÍDA DO CEFALOTÓRAX DO CAMARÃO MARINHO Litopenaeus schmitti
(BURKENROAD, 1936): AVALIAÇÃO DE SUAS ATIVIDADES ANTIMICROBIANA E LARVICIDA
Orientadora: Profa. Dra. Adriana Ferreira Uchôa Co-orientadora: Profa. Dra. Luciana Duarte Martins da Matta
Natal/RN 2011
Dissertação apresentada ao
Departamento de Bioquímica da
Universidade Federal do Rio Grande do
Norte como requisito parcial à obtenção
do título de Mestre em Bioquímica.

Catalogação da Publicação na Fonte
Universidade Federal do Rio Grande do Norte - UFRN
Biblioteca Central Zila Mamede
Godone, Roberta Luciana do Nascimento.
Isolamento de uma quitinase extraída do cefalotórax do camarão marinho
Litopenaeus Schmitti (BURKENROAD, 1936) : avaliação de suas atividades
antimicrobiana e larvicida / Roberta Luciana do Nascimento Godone. – Natal, 2011.
100 f. : il.
Orientadora: Adriana Ferreira Uchôa.
Co-orientadora: Luciana Duarte Martins da Matta.
Dissertação (Mestrado) – Universidade Federal do Rio Grande do Norte. Centro
de Biociências. Departamento de Bioquímica. Mestrado em Bioquímica.
1. Biotecnologia – Caracterização e aplicação – Dissertação. 2. Invertebrado –
Dissertação. 3. Enzimas – Dissertação. 4. Glicosidases – Dissertação. I. Uchôa,
Adriana Ferreira. II. Matta, Luciana Duarte Martins. III. Universidade Federal do Rio
Grande do Norte. IV. Título.
RN/UF/BCZM CDU 577.15(043.3)


Dedico esta obraDedico esta obraDedico esta obraDedico esta obra
A DeusDeusDeusDeus, fonte de aguá viva, meu pai fiel, estais comigo e eu contigo a todo o momento, seja ao meu lado ou me carregando nos braços, obrigada por todas as graças derramadas em minha
vida. Amém!
À minha mãe Aneide Aneide Aneide Aneide e o meu irmão MatheusMatheusMatheusMatheus, por toda paciência, carinho e compreensão, durante toda a minha vida, agradeço a Deus todos os dias desde o momento que acordo ate ir dormir, sei
que sou uma pessoa abençoada, pois tenho vocês.
À minha Orientadora, ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Luciana Duarte Martins da . Luciana Duarte Martins da . Luciana Duarte Martins da . Luciana Duarte Martins da MattaMattaMattaMatta, pela grande oportunidade de ingressar na ciência, pela amizade, atenção, carinho, compreensão e ensinamentos. Para
mim é uma honra inesplicavél, fazer parte do seu tão maravilhoso grupo de pesquisa, e sinto-me mais honrada ainda em poder dizer que fui preparada pela senhora. Quando vejo este trabalho concluído, me emociono, pois trabalhamos juntas e finalmente conseguimos o que tanto sonhavamos. Obrigada
sempre...
Ao Prof. Dr. Maurício Pereira de Sales (in memoriam),Prof. Dr. Maurício Pereira de Sales (in memoriam),Prof. Dr. Maurício Pereira de Sales (in memoriam),Prof. Dr. Maurício Pereira de Sales (in memoriam), pela oportunidade que me ofereceu, me aceitando como aluna de mestrado e me fazendo participar de seu grupo de pesquisa. Por depositar em mim, uma grande confiança, por acreditar no meu potencial, e ainda me fazer mais forte e determinada. Sou muito grata porisso e levarei comigo grandes lições de vida que aprendi
convivendo ao seu lado. Muito obrigada e sinto saudades.

AGRADECIMENTOS
A DeusDeusDeusDeus meu pai e amigo sinto-me tão feliz, pois sei que nunca me abandonaste e nunca me abandonará.Te amo Deus.
À minha mãezinhamãezinhamãezinhamãezinha e meu irmãozinhoirmãozinhoirmãozinhoirmãozinho por sempre acreditarem em mim, até quando eu não acreditava. Obrigada!
Á minha Orientadora ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Luciana da Matta (Profa). Luciana da Matta (Profa). Luciana da Matta (Profa). Luciana da Matta (Profa), pela confiança e apoio, em minha caminhada científica. A senhora é simplesmente Demais!!!!
Aos meus grandes amigos e cúmplices: Caroline Gabriela (Caroline Gabriela (Caroline Gabriela (Caroline Gabriela (GabiGabiGabiGabi)))), , , , Nerivaldo (Nerivaldo (Nerivaldo (Nerivaldo (NeríNeríNeríNerí)))), Stélio (Stélio (Stélio (Stélio (StelhinhoStelhinhoStelhinhoStelhinho)))) e Simony (Júlia)Simony (Júlia)Simony (Júlia)Simony (Júlia) que a todo tempo sempre estão de prontidão a me ouvir e ajudar, vocês são exemplo de caráter e otimismo. Simplesmente adoro vocês.
Ao Prof. Dr. LuizProf. Dr. LuizProf. Dr. LuizProf. Dr. Luiz RobertoRobertoRobertoRoberto pela ajuda, disponibilidade e contribuições valiosas em minha banca de qualificação, obrigada por ter participado desta etapa tão importante de minha vida.
Ao Prof. Dr. Maurício Pereira de SalesProf. Dr. Maurício Pereira de SalesProf. Dr. Maurício Pereira de SalesProf. Dr. Maurício Pereira de Sales (in memorian) por ter me aceitado como aluna de mestrado e ter acreditado no meu potencial. Obrigada!
À ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Adriana Uchôa. Adriana Uchôa. Adriana Uchôa. Adriana Uchôa por ter me aceitado como orientanda. Serei sempre grata porisso.
À ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Ana Heloneida. Ana Heloneida. Ana Heloneida. Ana Heloneida por ter aceitado participar de minha banca de qualificação, muitíssimo obrigada.
À ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Vânia Andrade , . Vânia Andrade , . Vânia Andrade , . Vânia Andrade , por ter aberto as portas de seu laboratório, pela orientação nos ensaios microbiológicos, por ter colocando uma de suas melhores alunas para me ajudar e por ter aceitado em participar de minha banca de qualificação, fazendo apontamentos valiosíssimos. Muito Obrigada.
Aos Professores Doutores José Luiz José Luiz José Luiz José Luiz Lima FilhoLima FilhoLima FilhoLima Filho e João Paulo MatosJoão Paulo MatosJoão Paulo MatosJoão Paulo Matos, por terem aceitado participar de minha banca de defesa. Muito Obrigada.

As meninas do LAMEA: Gabriela (Gabi), Raísa e LuizaGabriela (Gabi), Raísa e LuizaGabriela (Gabi), Raísa e LuizaGabriela (Gabi), Raísa e Luiza. Obrigada meninas!!!
O anjinho de candura, Gabriela (Gabi),Gabriela (Gabi),Gabriela (Gabi),Gabriela (Gabi), a qual sem ela eu não teria conseguido realizar nem entender os experimentos na Microbiologia. Obrigada pela paciência e percistência. Gabi você é mil...
À ProfProfProfProfaaaa. Dr. Dr. Dr. Draaaa. Fátima Ximenes. Fátima Ximenes. Fátima Ximenes. Fátima Ximenes, por ter disponibiizado o seu laboratório para a realização dos ensaios com os Aedes aegypti.
À Veterinária Patrícia BarraPatrícia BarraPatrícia BarraPatrícia Barra (Pati)(Pati)(Pati)(Pati), pela amizade incondicional, pelos momentos de descontração e passeios tão divertidos. Sem sua ajuda e ídeias tão pespícazes, eu não teria feito muita coisa. Sei que além de uma colega de trabalho, ganhei uma amiga para vida toda, você foi imprecindível na realização deste trabalho.
A Rafaella (Rafinha)Rafaella (Rafinha)Rafaella (Rafinha)Rafaella (Rafinha), pela amizade, palavras de força e dedicação, sei que sempre poderei contar contigo. E como eu ando dizendo pra você “Tô aqui viu”...
A JoyceJoyceJoyceJoyce, pela amizade tão sincera e carinho para comigo, adorei ter te conhecido e ser sua amiga, conta sempre comigo!
A HenriqueHenriqueHenriqueHenrique, pela amizade tão querida, pela companhia no Lab e pelas conversas tão divertidas e peculiares, que por muitas vezes me fizeram rir horrores... Te amo Lindão!!
Aos meus queridos amigos e companheiros da tão maravilhosa convivência no nosso “céuzinho” chamado LQFP: BrunaBrunaBrunaBruna (Brunete)(Brunete)(Brunete)(Brunete),,,, HenriqueHenriqueHenriqueHenrique (Amore)(Amore)(Amore)(Amore), Juliana, Juliana, Juliana, Juliana (Jú)(Jú)(Jú)(Jú), Paula, Leonardo, Paula, Leonardo, Paula, Leonardo, Paula, Leonardo (Léo)(Léo)(Léo)(Léo), Daniela, Daniela, Daniela, Daniela (Dani)(Dani)(Dani)(Dani) e SíSíSíSílvialvialvialvia, obrigada pelo apoio e ajuda nos experimentos, saibam que estarão sempre em meu coração.
À minha “IC/especial preferida” Bruna (Bruninha/Brunete)Bruna (Bruninha/Brunete)Bruna (Bruninha/Brunete)Bruna (Bruninha/Brunete), pela ajuda incondicional nos experimentos, força, amizade e companheirismo, sempre me colocando pra cima. Sem sua ajuda não sei o que seria de mim. Adoro você minha florzinha.
A Ticiana (Tici)Ticiana (Tici)Ticiana (Tici)Ticiana (Tici) e a CarolCarolCarolCarol (De boa na lagoa)(De boa na lagoa)(De boa na lagoa)(De boa na lagoa) pela convivência, paciência e carinho que sempre tiveram comigo, me lembrarei de vocês com carinho imenso, Obrigada.

A Sheyla VarelaSheyla VarelaSheyla VarelaSheyla Varela e a Dayse CarolineDayse CarolineDayse CarolineDayse Caroline, pela acolhida em minha mudança de laboratório, no início do mestrado e pela convivência tão especial, vocês duas foram muito importantes nesta etapa de minha vida. Meninas, muito obrigada!
Ao pessoal tão querido do BIOPOL PrPrPrProfofofof.... Dr.Dr.Dr.Dr. HugoHugoHugoHugo OliveiraOliveiraOliveiraOliveira, Sara, , Sara, , Sara, , Sara, Mariana, Leandro, PopóMariana, Leandro, PopóMariana, Leandro, PopóMariana, Leandro, Popó, Rafael, , Rafael, , Rafael, , Rafael, NedinaldoNedinaldoNedinaldoNedinaldo, , , , Arthur Arthur Arthur Arthur e JailmaJailmaJailmaJailma.
A JailmJailmJailmJailminhainhainhainha minha amiguinha do peito, muito obrigada pelo apoio, amizade e carinho, nunca esquecerei o que você fez e até hoje faz por mim, pode ter certeza você nunca sairá de meu coração. Convivendo com pessoas como você, só me faz ter mais certeza que ainda existem “os puros de coração”. Os meus mais sinceros agradecimentos.
Aos professores do Departamento de Bioquímica que direta ou indiretamente contribuirão na minha formação.
Aos amigos de departamento que colaborarão com a realização de experimentos: Patrícia,Patrícia,Patrícia,Patrícia, Norberto,Norberto,Norberto,Norberto, Nathália,Nathália,Nathália,Nathália, RaRaRaRaffffaelaelaelaelLLLLa, Joyce, a, Joyce, a, Joyce, a, Joyce, RafaelRafaelRafaelRafael e Ana PaulaAna PaulaAna PaulaAna Paula (minha 1º IC/querida).
Ao querido amigo Paulo Ricardo (Paulindo)Paulo Ricardo (Paulindo)Paulo Ricardo (Paulindo)Paulo Ricardo (Paulindo), pela amizade e ajuda na utilização dos programas estatísticos. Se não fosse você, nem sei. Brigadinha....
Aos companheiros da turma de Mestrado 2009: Clarissa, Alisson, Clarissa, Alisson, Clarissa, Alisson, Clarissa, Alisson, Renata, Ana Keyla, Larissa, Leandro Karlan, Dayse, Dayse Caroline, Renata, Ana Keyla, Larissa, Leandro Karlan, Dayse, Dayse Caroline, Renata, Ana Keyla, Larissa, Leandro Karlan, Dayse, Dayse Caroline, Renata, Ana Keyla, Larissa, Leandro Karlan, Dayse, Dayse Caroline, Dayse Santos, Nathália, Janisson, Angélica, Jana, Rafael, LeonardoDayse Santos, Nathália, Janisson, Angélica, Jana, Rafael, LeonardoDayse Santos, Nathália, Janisson, Angélica, Jana, Rafael, LeonardoDayse Santos, Nathália, Janisson, Angélica, Jana, Rafael, Leonardo NobreNobreNobreNobre, Marcos, Marcos, Marcos, Marcos FelipeFelipeFelipeFelipe, Ruth, Luciana, Leonardo Rêgo, Ruth, Luciana, Leonardo Rêgo, Ruth, Luciana, Leonardo Rêgo, Ruth, Luciana, Leonardo Rêgo e DieDieDieDiegogogogo.
Aos funcionários do Departamento de Bioquímica: Seu Marcos, Creuza, Seu Marcos, Creuza, Seu Marcos, Creuza, Seu Marcos, Creuza, Seu ItamaSeu ItamaSeu ItamaSeu Itamar, Jonas, Ângela, Eliene, Ricardo, Seu Rogérior, Jonas, Ângela, Eliene, Ricardo, Seu Rogérior, Jonas, Ângela, Eliene, Ricardo, Seu Rogérior, Jonas, Ângela, Eliene, Ricardo, Seu Rogério e Dona Dona Dona Dona MargaritaMargaritaMargaritaMargarita.
Aos amigos queridos do DBQ, Virgínia, Raquel, Érika, Ricardo, Marília, Virgínia, Raquel, Érika, Ricardo, Marília, Virgínia, Raquel, Érika, Ricardo, Marília, Virgínia, Raquel, Érika, Ricardo, Marília, Tuane, Luiza, CeTuane, Luiza, CeTuane, Luiza, CeTuane, Luiza, Celina, Carol, Gabriel, Raniere, Cínthia, Richelle, lina, Carol, Gabriel, Raniere, Cínthia, Richelle, lina, Carol, Gabriel, Raniere, Cínthia, Richelle, lina, Carol, Gabriel, Raniere, Cínthia, Richelle, Alexandre, JéssicaAlexandre, JéssicaAlexandre, JéssicaAlexandre, Jéssica,Thiago, Rafael Russi, Hugo,Thiago, Rafael Russi, Hugo,Thiago, Rafael Russi, Hugo,Thiago, Rafael Russi, Hugo e VanessaVanessaVanessaVanessa.
A todos que contribuíram de alguma forma para realização deste trabalho.
A UFRN e agências financiadoras CAPES e CNPq.

“... Não desanime de você, ainda que a colheita de hoje não seja muito feliz. Não coloque um ponto final nas suas esperanças. Ainda há muito o que fazer, ainda há muito o que plantar, e o que amar nessa vida. Ao invés de ficar parado no que você fez de errado, olhe para frente, e veja o que ainda pode ser feito...
A vida ainda não terminou. E já dizia o poeta "que os sonhos não envelhecem...por isso devemos sonhar sempre, sonhar grande,
sonhar lindo, sonhar rindo..."
Padre Fábio de MeloPadre Fábio de MeloPadre Fábio de MeloPadre Fábio de Melo

RESUMO
As quitinases são enzimas envolvidas na degradação da quitina e estão presentes em uma gama de organismos, inclusive os que não contêm quitina, tais como bactérias, vírus, plantas e animais desempenhando importantes papeis fisiológicos e ecológicos. A quitina é hidrolisada por um sistema quitinolítico classificado como: Endo-quitinases, Exo-quitinases e N-acetil-β-D-glucosaminidases. Neste trabalho, a Litochitinase1 foi extraída do cefalotórax do camarão marinho Litopenaeus schmitti e purificada 987,32 vezes utilizando-se cromatografia de troca iônica DEAE-Biogel e de exclusão molecular Sephacryl S-200. Esta apresentou massa molecular em torno de 28,5 kDa. Os resultados obtidos, após os testes cinéticos com a Litochitinase1 utilizando-se como substrato o p-nitrofenil-N-acetil-β-D-glucosaminideo, mostraram Km aparente de 0,51 mM, atividade ótima em pH variando de 5,0 a 6,0, temperatura ótima a 55°C e estabilidade de atividade quando pré-incubada nas temperaturas de 25, 37, 45, 50 e 55°C. A enzima apresentou uma faixa de estabilidade nos pHs de 4,0 a 5,5. O HgCl2, inibiu significativamente a Litochitinase1 enquanto que o MgCl2 potencializa levemente sua atividade. Os testes antimicrobianos realizados mostraram que a Litochitinase1 apresenta atividade contra a bactéria gram-negativa Escherichia coli numa concentração que varia 800 a 500 µg/mL. A atividade larvicida contra Aedes aegypti foi investigada com os extratos brutos, F-III (50-80%) e na Litochitinase1 com tempo de 24 e 48 horas. Os resultados obtidos mostraram atividade larvicida em todas estas amostras com valores de EC50 de 6,59 mg/mL para o extrato bruto, 5,36 mg/mL para F-III e 0,71 mg/mL para Litochitinase1 com 24 horas de ensaio e 3,22 e 0,49 mg/mL, para F-III e Litochitinase1 no tempo de 48 horas. Outros experimentos realizados confirmaram a presença de quitina no intestino médio das larvas de Aedes aegypti, as quais podem estar sofrendo a ação da Litochitinase1 provocando suas mortes, como ainda a ausência de proteínas bioativas como inibidores de proteases serínicas e lectinas no extrato bruto, F-III e Litochitinase1, indicando que a morte das larvas é mesmo por ação da Litochitinase1. Observou-se ainda que, as enzimas extraídas do homogenato intestinal das larvas não interferiram na atividade da Litochitinase1. Estes resultados indicam que esta enzima pode ser usada como alternativa para controlar infecções causadas por Escherichia coli, como também na redução da infestação do mosquito vetor da dengue.
Palavras Chaves: Glicosidases, Invertebrados, Purificação, Caracterização e Aplicação Biotecnológica.

ABSTRACT
Chitinases are enzymes involved in degradation of chitin and are present in a range of organisms, including those that do not contain chitin, such as bacteria, viruses, plants and animals, and play important physiological and ecological roles. Chitin is hydrolyzed by a chitinolytic system classified as: endo-chitinases, exo-chitinases and N-acetyl-β-D-glucosaminidases. In this study a Litochitinase1 extracted from the cephalotorax of the shrimp Litopenaeus Schmitt was purified 987.32 times using ion-exchange chromatography DEAE-Biogel and molecular exclusion Sephacryl S-200. These enzyme presented a molecular mass of about 28.5 kDa. The results, after kinetic assay with the Litochitinase1 using as substrate p-nitrophenyl-N-acetyl-β-D-glucosaminideo, showed apparent Km of 0.51 mM, optimal activity at pH ranging from 5.0 to 6.0, optimum temperature at 55°C and stability when pre-incubated at temperatures of 25, 37, 45, 50 and 55°C. The enzyme showed a range of stability at pH 4.0 to 5.5. HgCl2 inhibited Litochitinase1 while MgCl2 enhances its activity. Antimicrobial tests showed that Litochitinase1 present activity against gram-negative bacterium Escherichia coli in the 800 µg/mL concentration. The larvicidal activity against Aedes aegypti was investigated using crude extracts, F-III (50-80%) and Litochitinase1 at 24 and 48 hours. The results showed larvicidal activity in all these samples with EC50 values of 6.59 mg/mL for crude extract, 5.36 mg/mL for F-III and 0.71 mg/mL for Litochitinase1 at 24 hours and 3.22 and 0.49 mg/mL for the F-III and Litochitinase1 at 48 hours, respectively. Other experiments confirmed the presence of chitin in the midgut of Aedes aegypti larvae, which may be suffering the action of Litochitinase1 killing the larvae, but also the absence of contaminating proteins as serine proteinase inhibitors and lectins in the crude extract, F-III and Litochitinase1, indicating that the death of the larvae is by action of the Litochitinase1. We also observed that the enzymes extracted from intestinal homogenate of the larvae no have activity on Litochitinase1. These results indicate that the enzyme can be used as an alternative to control of infections caused by Escherichia coli and reducing the infestation of the mosquito vector of dengue. Keywords: Glucosidases, Invertebrate, Purification, Characterization and Biotechnology application.

LISTA DE FIGURAS
FIGURA 01 Estrutura Química da quitina. (Modificado de SEIDL, 2008).............................................................................................
21
FIGUTA 02 Representação esquemática das estruturas polimórficas de quitina, sendo que as setas representam as cadeias poliméricas no sentido do terminal não redutor para o redutor. (CAMPANA-FILHO, 2006)............................................................................................
22
FIGURA 03 Esquema dos padrões de clivagem de enzimas quitinolíticas. As subunidades da cadeia da quitina são mostradas em azul claro e o açúcar redutor em azul escuro. As tesouras representam as ações enzimáticas. (Modificado de SEIDL, 2008)............................................................................................
23
FIGURA 04 Mapa da distribuição geográfica do camarão branco Litopenaeus schmitti. (MARTINELI, 2005)....................................
29
FIGURA 05 Litopenaeus schmitti..................................................................... 32
FIGURA 06 Fluxograma do fracionamento com sulfato de amônio.................. 37
FIGURA 07 Atividades enzimáticas presentes no extrato bruto e nas frações obtidas da precipitação com sulfato de amônio............................
51
FIGURA 08 Perfil de eluição protéico da F-III em cromatografia de troca iônica e de atividade quitinásica ...................................................
52
FIGURA 09 Perfil de eluição da Litochitinase1 e atividade quitinásica em cromatografia de exclusão molecular (Sephacryl S-200).............
53
FIGURA 10 SDS-PAGE das amostras protéicas durante o processo de purificação.....................................................................................
55
FIGURA 11 Determinação do Km da Litochitinase1.......................................... 56
FIGURA 12 Efeito da temperatura sobre a atividade da
Litochitinase1.................................................................................
57
FIGURA 13 Efeito da pré-incubação a diferentes temperaturas na atividade da Litochitinase1............................................................................
58
FIGURA 14 Efeito do pH sobre a atividade da Litochitinase1.......................... 59
FIGURA 15 Efeito da pré-incubação a diferentes pHs na atividade da Litochitinase1...................................................................................
60

FIGURA 16 Curva de temperatura ótima para a detecção da atividade das enzimas proteolíticas presentes no homogenato intestinal de larvas de A.aegypti........................................................................
67
FIGURA 17 Efeito da pré-incubação com as enzimas digestivas de A.aegypti na atividade da Litochitinase1.......................................................
68

LISTAS DE QUADROS
QUADRO 1 Técnica de Difusão em Disco, cepas selecionadas e concentrações da quitinase utilizada..........................................
43
QUADRO 2 Técnica de Microdiluição em Caldo, cepas selecionadas e concentrações da quitinase ulitizada..........................................
44

LISTA DE TABELAS TABELA 1
Família das glicosil-hidrolases que possuem atividade N-
acetil-β-D-glucosaminidásica..............................................
20
TABELA 2 Algumas quitinases já isoladas e caracterizadas............... 28
TABELA 3 Etapas de purificação da Litochitinase1............................. 54
TABELA 4 Efeitos de íons e compostos sobre a
atividade da Litochitinase1..............................
61
TABELA 5 Avaliação da Concentração Inibitória Mínima (CIM) da
Litochitinase1 do cefalotórax do camarão marinho
Litopenaeus schimitti..........................................................
62
TABELA 6 Detecção de quitina em intestinos médios de Aedes
aegypti................................................................................
63
TABELA 7 Concentração efetiva do extrato bruto, F-III e
Litochitinase1 necessária para matar 50% (EC50) das
larvas L4 de Aedes aegypti em 24 e 48 h..........................
64
TABELA 8 Inibição de proteases serínicas.......................................... 65
TABELA 9 Atividade Hemaglutinante no extrato bruto, F-III e
Litochitinase1 com eritrócitos nativos e tratados
enzimaticamente................................................................
66

LISTA DE ABREVIATURAS
α Denota anomericidade alfa
ATCC American Type Culture Collection
β Denota anomericidade beta
BHI Brain Heart Infusion
°C Grau Celsius
cm Centímetros
CTT Cloreto de 2,3,5 – trifenil - tetrazólio
DO Densiometria óptica
EB Extrato Bruto
EDTA Acido etilenodiamino tetra-acético
F-I Extrato protéico precipitado com 0-30% de sulfato de amônio
F-II Extrato protéico precipitado com 30-50% de sulfato de amônio
F-III Extrato protéico precipitado com 50-80% de sulfato de amônio
x g Gravidade
g Grama
GlcNAc N-acetil-β-D-glucosamina
HCl Ácido Clorídrico
IUMBM International Union of Biochemistry and Molecular Biology
HI Homogenato intestinal
kDa Kilodaltons
Km Constante de Michaelis-Menten
EC50 Concentração Efetiva
M Molar
mM Milimolar
min. Minuto
mg Miligramas
µµµµg Microgramas
mL Mililitro
µµµµl Microlitro
NaCl Cloreto de Sódio
NaOH Hidroxido de Sódio

nm Nanômetro
pH Potencial hidrogeniônico
PNF p-NITROFENIL
SDS Dodecil sulfato de sódio
t Tonelada
TCA Ácido tricloro acético
Tris Tris hidroximetil aminometano
TEMED
[S]
UH
Tetra metileno diaminoetano
Concentração Molar de Substrato
Unidade de Hemaglutinação

SUMÁRIO
1. INTRODUÇÃO................................................................................................ 18 1.1. GLICOSIDASES.......................................................................................... 18
1.1.1. 1.1.1.Glicosil-Hidrolases................................................................................... 19
1.1.2. 1.1.2.Quitinases................................................................................................. 21
1.1.2.1. Ocorrência e Função das Quitinases................................................. 23
1.1.2.2. Aplicações Biotecnologicas................................................................ 26
1.2. Litopenaeus schmitti (BURKENROAD, 1936) COMO FONTE DE QUITINASES..............................................................................................
28
2. OBJETIVOS................................................................................................... 31 2.1. GERAIS....................................................................................................... 31 2.2. ESPECÍFICOS.......................................................................................... 31 3. MATERIAIS................................................................................................... 32
3.1. MATERIAL BIOLÓGICO............................................................................... 32
3.1.1. Camarão Litopenaeus schmitti............................................................. 32 3.1.1. 3.1.2. Insetos..................................................................................................... 32 3.1.2. 3.1.3. Eritrócitos Humanos............................................................................... 33 3.1.3. 3.1.4. Microorganismos..................................................................................... 33
3.2. SUBSTRATOS SINTÉTICOS DERIVADOS DE AÇUCAR....................... 33 3.3. REAGENTES, SOLUÇÕES, MEMBRANAS E MATRIZES
CROMATOGRÁFICAS............................................................................. 33
3.4. APARELHOS............................................................................................ 34 4. 4. MÉTODOS...................................................................................................... 36
4.1. OBTENÇÃO DO EXTRATO BRUTO........................................................... 36 4.2. FRACIONAMENTO COM SULFATO DE AMÔNIO...................................... 36
4.3. DETERMINAÇÃO DAS ATIVIDADES ENZIMÁTICAS COM p-NITROFENIL DERIVADOS DE AÇÚCAR....................................................
37
4.4. DOSAGEM DE PROTEÍNAS....................................................................... 38 4.5. PURIFICAÇÃO DA ENZIMA........................................................................ 38
4.5.1. Cromatografia de Troca Iônica DEAE-Biogel...................................... 38 4.5.2. Cromatografia de Exclusão Molecular – Sephacryl S – 200.............. 38
4.6. ELETROFORESE EM GEL DE POLIACRILAMIDA EM CONDIÇÕES DESNATURANTES-SDS/PAGE................................................................
39
4.7. ESTUDOS CINÉTICOS.............................................................................. 40 4.7.1. Determinação da Constante de Michaelis- Menten (Km).................... 40 4.7.2. Efeito da temperatura sobre a atividade enzimática........................... 40

4.7.3. Influência do pH sobre a atividade enzimática.................................... 41
4.7.4. Influência de diversos sais e EDTA na hidrólise do p-nitofenil-N-acetil-ββββ-D-glucosaminídeo.....................................................................
41
4.8. PREPARAÇÃO DA QUITINASE PARA OS ENSAIOS BIOLÓGICOS......... 42
4.9. AVALIAÇÃO DA ATIVIDADE ANTIMICROBIANA...................................... 42 4.9.1. Análise Qualitativa- Técnica de Difusão em Disco.............................
42
4.9.2. Análise Quantificativa- Determinação da Concentração Inibitória Mínima (CIM)............................................................................................
43
4.10. ENSAIOS COM Aedes aegypti................................................................ 44 4.10.1. Obtenção das larvas de Aedes aegypti.............................................. 44 4.10.2. Dissecação das larvas de Aedes aegypti.......................................... 44 4.10.3. Preparo do homogenato intestinal das larvas.................................. 45 4.10.4. Detecção de quitina no intestino das larvas de Aedes aegypti...... 45
4.10.5. Ensaios com larvas de Aedes aegypti............................................... 45 4.10.6. Determinação da EC50......................................................................... 46 4.10.7. Análises Estatísticas........................................................................... 46 4.10.8. Investigação de proteínas bioativas interferentes no processo
digestório nas larvas de Aedes aegypti.............................................
46 4.10.8.1. Ensaio de inibição de proteases serínicas...................................... 46 a) Preparo do substrato................................................................................... 46 b) Atividade anti-quimotriptica....................................................................... 46 c) Atividade anti-triptica.................................................................................. 47 4.10.8.2. Ensaios de hemaglutinação............................................................. 47 a) Preparo do sangue..................................................................................... 47 b) Tratamento dos eritrócitos com papaína................................................. 47 c) Tratamento dos eritrócitos com tripsina.................................................. 48 d) Ensaio........................................................................................................... 48 4.10.8.3. Efeito do homogenato intestinal das larvas de Aedes aegypti
sobre a atividade quitinolítica...........................................................
48 a) Determinação de atividade azocaseínolítica............................................. 48 b) Efeito da temperatura sobre a atividade azocaseínolitica a pH 7,5........ 49
c) Influencia das enzimas digestivas do Aedes aegypti na hidrólise do p-
nitrofenil-N-acetil-ββββ-D-glucosaminídeo.......................................................
49
5. RESULTADOS ............................................................................................. 50
5.1.IDENTIFICAÇÃO DE GLICOSIDASES E SULFATASES NOS CEFALOTÓRAX DOS CAMARÕES MARINHOS litopenaeus
schmitti...........................................................................................................
50
5.2. PURIFICAÇÃO DA QUTINASE.................................................................... 51 5.2.1. Em cromatografia de troca iônica......................................................... 51

5.2.2. Em cromatografia de exclusão molecular S-200................................. 52 5.2.3. Resumo das etapas de purificação....................................................... 53 5.3. PERFIL ELETROFORÉTICO DAS AMOSTRAS EM SDS-PAGE............... 54
5.4. EXPERIMENTOS CINÉTICOS.................................................................... 55 5.4.1. Determinação do Km............................................................................. 55 5.4.2. Efeito da temperatura sobre a atividade enzimática........................... 56 5.4.3. Efeito do pH sobre a atividade da Litochitinase1............................... 58 5.4.4. Efeito de sais e EDTA sobre a atividade enzimática........................... 60 5.5. AVALIAÇÃO DA ATIVIDADE ANTIMICROBIANA...................................... 61 5.5.1. Utilizando a técnica de difusão em disco............................................ 61 5.5.2. Utilizando a técnica de microdiluição em caldo para a
determinação da concentração inibitória mínima (CIM)....................
62 5.6. ENSAIO COM Aedes aegypti...................................................................... 62 5.6.1. Detecção de quitina no intestino médio de larvas de Aedes aegypti 62 5.6.2. Ensaio larvicida em Aedes aegypti...................................................... 63 5.6.3. Detecção de inibidores de proteases serínicas no EB, FIII e
Litochitinase1.........................................................................................
64 5.6.4. Atividade hemaglutinante...................................................................... 65 5.6.5. Determinação da temperatura ótima das atividades proteolíticas
presentes no homogenato intestinal das larvas de Aedes
aegypti.....................................................................................................
66 5.6.6. Influência das enzimas digestivas de Aedes aegypti sobre a
Litochitinase1.........................................................................................
67 6. DISCUSSÃO............................................................................................... 70 7. CONCLUSÕES........................................................................................... 79
REFERÊNCIAS .......................................................................................... 81

18
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
1. INTRODUÇÃO
1.1 GLICOSIDASES
As glicosidases são um grupo de enzimas que clivam O-glicosídeos, N-
glicosídeos e tio-glicosídeos (DUMONT, 2008; WITHERS, 2010). Estando desta
forma diretamente envolvidas no metabolismo de monossacarídeos, dissacarídeos,
oligossacarídeos e até polissacarídeos (FERREIRA, 2001). Dentre elas destacam-se
as endoglicosidases e exoglicosidases. As endoglicosidases são enzimas que
clivam cadeias liberando dissacarídeos ou oligossacarídeos. Já as exoglicosidases
são um grupo de enzimas que clivam a cadeia oligossacarídica através de sua
porção não redutora liberando monossacarídeos (MALLEY et al.,1989).
As glicosidases formam um grupo altamente heterogêneo de enzimas
hidrolíticas (BHATIA et al., 2002). Tais enzimas são encontradas em diversos
organismos, como bactérias (KATAYEVA et al., 1992; PAAVILAINEN et al., 1993;
SESTELO et al., 2004; ARO et al., 2005) fungos (RICCIO et al., 1999; JAGER et
al., 2001; YUN et al., 2001; BELANCIC et al., 2003) plantas (SUE et al., 2000;
CAMERON et al., 2001; GERARDI, 2001; SARRY; GÜNATA, 2004) e animais
(MARANA et al., 1995; HAYS et al., 1998; MARANA, 1999), (PONTOH; LOW, 2002;
DRUMONT, 2008),como por exemplo em crustáceos (KOSTANJNEK et al., 2010;
ALLARDYCE et al., 2010) e o homem (GRACE et al., 1994; DAY et al., 1998;
NEMETH et al., 2003; GERMAIN, 2004).
A principal reação catalisada por estas enzimas é a hidrólise de ligações β-
glicosídicas. Estas enzimas possuem uma carboxila e um carboxilato responsáveis
pela catálise, um funcionando como nucleófilo e o outro como doador de prótons
(JONES, 2002). A especificidade das β-glicosidases pode variar, pois algumas
destas enzimas podem ser capazes de atuar sobre uma gama enorme de substratos
e outras podem ser mais restritas (FERREIRA, 1998, 2001, 2003; AZEVEDO, 2003).
Desta forma o nome β-glicosidase é dado a diferentes tipos de enzimas capazes de
hidrolisar ligações β-glicosídicas em dissacarídeos, oligossacarídeos e glicosídeos
conjugados (COULON et al., 1998; BHATIA et al., 2002).

19
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
1.1.1 GLICOSIL-HIDROLASES
As glicosil-hidrolases são glicosidases que hidrolisam ligações O-glicosídicas
e são classificadas, pela Enzyme Commission Numbers (EC) a International Union of
Biochemistry and Molecular Biology (IUBMB), com base na sua especificidade pelo
substrato, por exemplo: β-glucosídeo ou β-galactosídeo, e no tipo de ligação, por
exemplo: β-1,4 ou β-1,3. Esta classificação é útil, pois unifica a nomenclatura e evita
as ambigüidades dos nomes triviais. Entretanto, tal classificação não reflete as
características estruturais e correlações evolutivas entre as enzimas (MARANA,
1999).
A fim de estabelecer uma classificação que contemplasse o aspecto estrutural
e evolutivo das glicosil-hidrolases, HENRISSAT (1991), partindo do pressuposto de
que existe uma correlação entre similaridade da seqüência de aminoácidos, e a
estrutura terciária das proteínas, foi realizado um alinhamento de seqüências e
conjuntos de aminoácidos hidrofóbicos das glicosil-hidrolases disponíveis e montou-
se um sistema de classificação para estas enzimas(CHOTIA; LESK, 1986).
Esta classificação baseou-se em um total de 291 seqüências correspondendo
a 39 diferentes EC que puderam ser classificadas em 35 famílias em seguida
HENRISSAT; BAIROCH, (1993), ampliaram ainda mais o sistema de classificação
atingindo um total de 45 famílias. Agrupando as enzimas de origem evolutiva
comum, independentemente das reações que elas catalisam aproximadamente 90
famílias de glicosil-hidrolases são conhecidas (WITHERS, 2002).
Além do critério de similaridade na seqüência de aminoácidos, elas também
podem ser classificadas segundo mecanismos de reação, que as categoriza
segundo dois mecanismos: o de retenção e o de inversão da configuração do
anômero C1 do anel glicosídico. A hidrólise se dá via catálise ácida, onde há um
doador de prótons e um nucleófilo. Em ambos, o doador de prótons está próximo ao
oxigênio glicosídico e o nucleófilo fica na posição vicinal do carbono 2 do anel. A
diferença é que no mecanismo de inversão a distancia do nucleófilo é maior, a fim
de comportar uma molécula de água. Essa distância é cerca de 5,5 Å no mecanismo
de retenção e ~10 Å no de inversão na formação do complexo enzima substrato.
(DAVIS, 1995; KOSHALND, 1953). O ataque nucleofílico é feito pelos dois resíduos

20
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
aproximando-se pelo lado oposto da ligação glicosídica e há a retenção ou inversão
da configuração anomérica do anel glicosídico (HENRISSAT, 1995).
A clivagem da ligação glicosídica entre um resíduo de N-acetil-β-D-
glucosamina e outro resíduo adjacente, pode ser do tipo exo ou endo-
glucosidasicas, e essas se encontram distribuídas em seis famílias de glicosil-
hidrolases: GH3, GH18, GH19, GH20, GH73 e GH84. (LIMA, 2006). A tabela 1
abaixo sumariza as características das seis famílias citadas.
O sistema de classificação glicosil-hidrolase (GH), introduzido e desenvolvido
por HENRISSAT, (1991) e HENRISSAT; BAIROCH, (1993) está disponível em um
banco de dados geral (http://afmb.cnrs-mrs.fr/~CAZY/index.html) (COUTINHO;
HENRISSAT, 1999; LIMA, 2006).
Tabela 1: Família das glicosil-hidrolases que possuem atividade N-acetil-β-D-glucosaminidásica.
Família Atividades conhecidas
GH3 β-glicosidase (EC 3.2.1. 21); xilano 1,4-β-xilosidase (EC 3.2.1. 37);
β-N-acetilhexosaminidase (EC 3.2.1. 52); glicano 1,3-β-glicosidase
(EC 3.2.1. 58); glicano 1,4-β-glicosidase (EC 3.2.1. 74); exo-1,3-
1,4-glicanase (EC 3.2.1. -); α-L-arabinofuranosidase (EC 3.2.1. 55)
GH18 Quitinase (EC 3.2.1. 14); endo-β-N-acetilglicosaminidase (EC
3.2.1. 96)
GH19 Quitinase (EC 3.2.1. 14)
GH20 β-hexosaminidase (EC 3.2.1. 52); lacto-N-biosidase (EC 3.2.1.
140)
GH73 Endo-β-N-acetilglicosaminidase (EC 3.2.1. 96); β-1,4-N-
acetilmuramoilhidrolase
GH84 β-N-acetilglicosaminidase (EC 3.2.1. 52); hialuronidase (EC 3.2.1.
35)
Fonte: (modificado de HENRRISAT ; COUTINHO, 1999).

21
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
1.1.2 QUITINASES
As quitinases são enzimas que clivam ligações O-glicosídicas entre os
carbonos C1 e C4 dos resíduos de β-1,4-N-acetilglucosamina (GlcNAc), constituintes
do polímero de quitina (Figura 01). Estas enzimas são classificadas como glicosil
hidrolases e compreendem cerca de 90 famílias de enzimas descritas. As quitinases
foram colocadas na família 18, 19 e 20, na qual, as da família 18 são encontradas
em vírus, bactérias, fungos filamentosos ou leveduriformes, plantas e animais, e,
portanto, a família é diversa em termos evolucionários. Membros da família 19 são
quase que exclusivas de plantas. A família 20 consiste em N-acetil-β-D-
hexosaminidase ou N-acetil-β-D-glucosaminidases de bactérias, fungos e seres
humanos (HORSCH et al., 1997; WITHERS, 2002; SEIDEL, 2008).
A quitina é a segunda substância orgânica até então conhecida na biosfera
sendo superada apenas pela celulose, mas supera esta última em termos de taxa de
reposição, que chega a ser duas vezes maior que a da celulose (THARANATHAN;
KITTUR 2003; YEN et al., 2009).
Quitina e celulose possuem características estruturais semelhantes e atuam
como invólucros protetores e materiais de suporte e defesa nos organismos em que
ocorrem. A quitina e/ou polímeros de N-acetil-glucosamina encontra-se na matriz da
estrutura esquelética de invertebrados, como artrópodes, anelídeos, moluscos e
cnidários, em algas diatomáceas, no peptideoglicano da parede celular das bactérias
e também está presente nas paredes celulares de alguns fungos, como
ascomicetos, zigomicetos, basidiomicetos e deuteromicetos (KARAMANOS, 1997;
RIVAS et al., 2002; DUO-CHAN, 2006; ZHANG et al., 2010).
Figura 01: Estrutura Química da quitina. Fonte: (Modificado de SEIDL, 2008)

22
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Este biopolímero ocorre naturalmente em três diferentes formas α-, β- e γ-
quitina (Figura 02). A α-quitina é encontrada em estruturas rígidas e resistentes,
como a cutícula de artrópodes, e nesses casos ocorre fortemente associada a
proteínas, materiais inorgânicos ou ambos. A α-quitina é a forma mais abundante e
é também considerada a mais estável, visto que a conversão das duas últimas
formas na primeira é irreversível (ROBERTS, 1992). As formas β- e γ-quitina ocorre
em estruturas flexíveis embora também resistentes. Nas lulas do gênero Loligo a α-
quitina constitui uma fina capa que reveste as paredes do esôfago e do estômago, a
β-quitina ocorre como o principal componente das conchas, ou plumas, e a γ-quitina
integra uma espessa cutícula que recobre outras zonas do estômago (ABRAM;
HIGUERA, 2004).
Figura 02: Representação esquemática das estruturas polimórficas de quitina, sendo que as setas representam as cadeias poliméricas no sentido do terminal não-redutor para o redutor.
Fonte: (CAMPANA-FILHO, 2006).
As quitinases podem ser classificadas ainda, quanto ao seu modo de ação
sobre o substrato. A nomenclatura enzimática oficial (EC) classifica as enzimas
quitinolíticas em apenas dois grupos de enzimas: as quitinases, também conhecidas
como endoquitinases, que catalizam a hidrólise randômica de ligações β-1,4 de N-
acetilglucosamina (GlcNac) liberando produto solúveis como quitotetraoses,
quitotriose e diacetilquitobiose; e as N-acetilglucosaminidases, também conhecidas
como exoquitinases, que clivam a quitina em monômeros N-acetilglucosamina.
Entretanto algumas enzimas quitinolíticas não se encaixam nessa
classificação. Assim diferentes classificações foram sugeridas. Dependendo dos
seus padrões de clivagem as quitinases foram divididas em três tipos:
endoquitinases, exoquitinases e N-acetil-β-D-glucosaminidases. As endoquitinases

23
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
clivam as ligações β-1,4 da quitina a partir de qualquer ponto ao longo da cadeia
polimérica liberando produtos (oligossacarídeos) de tamanho aleatório, enquanto as
exoquitinases atuam a partir da porção não-redutora da cadeia e seus produtos
finais geralmente são dímeros de N-acetil-β-D-glucosamina (GlcNAc)2 (SAHAI;
MANOCHA, 1993; SEIDEL, 2008). As N-acetil-β-D-glucosaminidases são um grupo
de enzimas que hidrolisam componentes O-glicosídicos, removendo resíduos
terminais não-redutores de N-acetil-β-D-glucosamina, sendo assim, considerada
uma exoglicosidase (HORSCH et al., 1997). Estas enzimas também são
denominadas de N-acetil-β-D-hexosaminidases (EC 3.2.1. 52), devido ao fato de
possuírem atividade N-acetil-β-glucosaminidásica bem como atividade N-acetil-β-
galactosaminidásica (KRESSE; GLOSSL, 1987; HORSCH et al., 1997; NIIMI et al.,
2001).( Figura, 03).
Figura 03: Esquema dos padrões de clivagem de enzimas quitinolíticas. As subunidades da cadeia da quitina são mostradas em azul claro e o açúcar redutor em azul escuro. As tesouras representam as ações enzimáticas
Fonte: (Modificado de SEIDL, 2008).
1.1.2.1 OCORRÊNCIA E FUNÇÕES DAS QUITINASES
Diversos estudos apontam o papel fisiológico e ecológico das quitinases. Na
maioria destes estudos conclui-se que elas estão envolvidas em processos de
defesa contra patógenos, mas que também desempenham outros papéis
importantes que serão abordados adiante.
Endoquitinases
Exoquitinases
N-acetilglucosaminidases

24
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
A quitina, além de estar presente em fungos, também se encontra em insetos
e crustáceos (LENARDON et al., 2010; CHEN et al., 2010; QIN et al., 2010). As
quitinases em crustáceos são associadas à degradação parcial do exoesqueleto
durante seu desenvolvimento, por um mecanismo que parece estar sob o controle
hormonal. Assim como nos crustáceos, o desenvolvimento de insetos está
diretamente relacionado com a regulação da síntese e degradação de quitina, que é
o principal componente do seu exoesqueleto (SPINDLER-BARTH, 1993). Além do
papel no desenvolvimento, as quitinases também exercem o papel de defesa contra
patógenos em outros invertebrados
Segundo MULLEN et al. (2004), estudos preliminares mostraram a presença
de altos níveis de exo e endoquitinases em corais infectados por fungos.
As quitinases em fungos parecem ser um fator decisivo na colonização e
predação para obtenção de alimentos (BISHOP et al., 2000). A atividade quitinolítica
é de fundamental importância para patogênese de infestação por fungos, como por
exemplo, Metarhizium anisopliae sf. acridum (SCREEN et al., 2001). Através da
indução com quitina, este microorganismo foi capaz de secretar isoformas básicas e
acidas de endoquitinases (KANG et al.,1999). Estas enzimas são largamente
utilizadas por M. anisopliae na colonização de lagartas de Manduca sexta, digerindo
a cutícula e a membrana peritrófica (SCREEN et al., 2001).
No culicídeo Aedes aegypti, a quitina é um dos componentes principais da
membrana peritrófica (MP) formada no meio intestinal da fêmea adulta após o
repasto sanguineo, o que ocasiona a proteção do sangue ingerido contra micróbios
patogênicos, promovendo assim uma barreira a distribuição e crescimento
subseqüente desses patógenos. (FILHO et al., 2002), desta forma a degradação da
membrana peritrófica nesses insetos pode ocasionar crescimento de
microorganismos promovendo sua infestação como também pode ocasionar a
ruptura no tecido intestinal, o vazamento do fluido celular e o desbalanceamento
osmótico, matando o inseto por desidratação ou inanição (BISHOP et al., 2000).
Estudos mostraram que quando o culicídeo Aedes aegypti se alimenta de
alosamidina X, ocorre a formação atípica da membrana peritrófica (MP), sugerindo a
presença de um sistema quitinolítico no intestino do inseto, responsável pela
modificação e controle da formação da MP. O conhecimento de como este sistema
quitinolítico funciona e sua função na formação da MP pode facilitar o

25
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
desenvolvimento de estratégias que podem ser usadas para manipular o processo
natural, incluindo a interrupção da invasão de patógenos em mosquitos vetores
(FILHO et al., 2002).
As quitinases são também podem ser encontradas em alguns organismos que
não apresentam a quitina em sua constituição, como em bactérias, nematódeos e
plantas (COHEN-KUPIEC; CHET, 1998). Em bactérias como Serratia e
Streptomyces como também em nematódeos (Onchocerca gibsoni) (McNAB;
GLOVER, 1991), sua função está relacionada à nutrição. Dessa forma, a hidrólise de
uma variedade de quitinas encontradas na natureza permite ao organismo utilizar
esses polímeros como fonte de energia (VAN AALTEN et al., 2001; COHEN-
KUPIEC; CHET, 1998). Em nematódeos, as quitinases estão presentes na casca do
ovo e contribuem para a manutenção de sua integridade (ARNOLD et al., 1993).
Quitinases em plantas estão associadas à defesa contra patógenos e fazem
parte do grupo de proteínas denominadas PR (Pathogenesis Related Proteins), que
são proteínas capazes de induzir resistência local ou sistêmica ao ataque de fungos
e outros fitopatógenos. Evidências indicam que as quitinases têm um papel direto
nessa defesa por atacar diretamente os polímeros de quitina, que é o maior
componente da parede celular da maioria dos fungos (COLLINGE et al., 1993;
KUPIEC; CHET, 1998; HODGE et al., 1996).
Nesses casos, essas enzimas possuem um papel de defesa contra
organismos que possuem quitina em sua composição, inibindo in vitro o crescimento
de fungos filamentosos, especialmente em combinação com β-1,3-glucanase
(COHEN-KUPIEC; CHET, 1998; ISELI et al.,1996).
Embora animais vertebrados não sejam produtores de quitina, a presença de
quitinases tem sido verificada no organismo de várias espécies desses animais.
Essas enzimas são encontradas em vários órgãos e tecidos, como no trato
digestivo, participando do metabolismo de carboidratos (JEUNIAUX, 1993). Peixes
que se alimentam de crustáceos ou insetos contêm uma grande concentração de
quitinases em seu suco pancreático (ROTTA, 2003).
A expressão de quitinases em mamíferos não havia sido reportada até 1994
quando HOLLACK et al. identificaram a primeira quitinase humana em
pacientes com a Doença de Gaucher. Em outro estudo BOOT et al. (2001)
identificaram no trato gastrintestinal e pulmões, outra quitinase denominada
AMCase (acidic mammalian chitinase).

26
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
A ocorrência de quitinases em mamíferos e humanos sugere que sua função
esteja relacionada com a metabolização de carboidratos e de participação em
mecanismos de defesa contra agentes patogênicos. No entanto, seu papel
fisiológico em mamíferos e especialmente em humanos ainda não é completamente
conhecido e parece ser um paradoxo, sendo presença da quitotriosidase fortemente
associada também à arteriosclerose e a presença da AMCase associada à asma.
Esses dados sugerem que essas enzimas possam participar de desordens
inflamatórias (DONNELY; BARNES, 2004).
1.1.2.2 APLICAÇÕES BIOTECNOLÓGICAS
Extensos estudos sobre quitinases estão sendo desenvolvidos devido ao fato
de sua ampla capacidade de aplicação na biotecnologia, nas mais diversas áreas
como na agricultura, medicina e o setor industrial.
A aplicação de quitinases na agricultura objetiva principalmente o controle
biológico seja pelo desenvolvimento de bioinseticidas para controle de insetos,
fungos fitopatogênicos e bactérias ou pelo desenvolvimento de plantas transgênicas
resistentes ao ataque de microorganismos ou ainda na produção de
quitooligossacarídeos biologicamente ativos (PATIL et al., 2000; FLEURI, 2008).
Muitas quitinases de várias fontes têm apresentado atividade contra patógenos de
plantas e contra insetos que causam pestes na agricultura (DAHYIA et al., 2006).
Quitinases de T. harzianum (DEMARCO et al., 2000), Enterobacter sp. NRG4
(DAHYIA et al., 2005), Cellulosimicrobium cellulans 191 (FLEURI; SATO, 2008)
Clonostachys rósea (LU BECK et al., 2009) e de sementes de Acacia confusa (NG et
al., 2010) , já mostraram atividade inibitória contra alguns desses patógenos,
podendo assim ser utilizados como suplementos na produção de fungicidas e
inseticidas. A partir da bactéria Bacillus circulans foi produzido uma quitinase para
ser usada como suplemento bioinseticida da bactéria entomopatogênica Bacillus
thuringiensis para controle de larvas de lepidópteras. Um sistema lítico composto
das seguintes enzimas: quitinase, protease, β-glucanase e lipase foram utilizadas
para o controle de manchas nas folhas de centeio, causadas por Biopolaris
sorokiniana (FLEURI; SATO, 2005).

27
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tem sido extensamente pesquisada a utilização das quitinases na produção
de quitooligossacarídeos com atividades biológicas. Estes são potencialmente úteis
na medicina. Por exemplo, a quitohexaose e quitoheptaose mostraram atividade
antitumoral. A combinação específica de enzimas quitinolíticas com altos níveis de
endo-quitinases e baixa atividade de N-acetilglucosaminidases e exo-quitinases
podem ser necessárias para obter o oligossacarídeo desejado. Em contrapartida a
produção de N-acetilglucosamina pode ser obtida utilizando altas proporções de
exo-quitinases e N-acetilglucosaminidases. Este açúcar quando aplicado por via
intramuscular ou oral é um potente antiinflamatório podendo ser utilizado no
tratamento de colites ulcerativas ou doenças gastrointestinais (PATIL et. al., 2000).
Quitinases purificadas podem liberar oligômeros de quitina, a partir da parede
celular fúngica (SCHLUMBAUM et al., 1986) como também podem agir
semelhantemente à lisozima, sobre paredes celulares bacterianas (HERGET et al.,
1990; STINTZI et al., 1993).
A quitina também está presente na membrana peritrófica (MP) de insetos
como o Aedes Aegypti. A MP protege o epitélio intestinal e promove a
compartimentalização das enzimas digestivas, sendo fundamental para o processo
digestório. As quitinases podem ter ação inseticida por interferirem na integridade da
MP. Etapas adicionais de purificação dessa (s) quitinase(s) poderão indicar essas
proteínas como candidatas no controle do inseto transmissor da dengue.
Outro campo em que pode haver utilização de quitina é a bioconversão. O
consumo de camarões no mundo é muito grande, principalmente em países
costeiros. No entanto, somente a carne é consumida e o cefalotórax e a casca, são
descartados. Há dados em que alguns países são produzidos mais de 2,5 milhões
toneladas de lixo dessa natureza (HAKI et al., 2003; DAHYIA, 2006). Assim, o
desenvolvimento de métodos de conversão desse rejeito em produtos derivados de
quitina é de grande importância. E esses derivados de quitina podem ser obtidos
pela degradação desses pelas quitinases. Nos últimos anos várias quitinases
derivadas de diversos organismos foram isoladas, clonadas ou expressas na forma
recombinante (Tabela 2).

28
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 2: Algumas quitinases já isoladas e caracterizadas.
Origem Massa Molecular
(kDa)
Autor/Ano
Streptomyces sp. TH-11 29 HOANG et al., 2011
Penaeus monodon 72,4 e 51,9 PROESPRAIWONG et al., 2010
Ipomoea carnea 30,6 PATEL et al., 2010
Pennahia argentatus 42 IKEDA et al., 2009
Pieris rapae 59,5 e 57,2 SHI et al., 2006
Bacillus thuringiensis 74 BARBOZA-CORONA et
al., 2002
Pseudomonas aeruginosa 385
58 THOMPSON et al., 2001
Pseudomonas sp YHS-A2 67 LEE et al., 2000
1.2 Litopenaeus schmitti (BURKENROAD, 1936) COMO FONTE DE
QUITINASES
O camarão branco Litopenaeus schmitti, é o único pertencente ao gênero
Litopenaeus que ocorre em águas brasileiras. Assim como várias outras espécies da
família Penaeidae, possuem alta fecundidade, procriam em águas costeiras e
apresentam estágio larval planctônico por uma semana ou mais, sendo facilmente
disperso pelas correntes (LUVESUTO, 2006).
As áreas estuarinas são zonas adequadas para a nutrição e desenvolvimento
dos camarões (GAMBA; RODRIGUEZ, 1987), onde crescem aproximadamente de
10 mm a 75 mm, durante aproximadamente de 6 a 9 meses (EWALD, 1965; NEIVA
et al., 1971). Os machos da espécie crescem em torno de 170 mm e as fêmeas
comumente atingem um tamanho de 200 mm. Os maiores indivíduos devem ter em
torno de 2 anos, uma vez que os camarões peneídeos possuem baixa longevidade
(1,5 a 2 anos) (ROTHLISBERG et al.,1985).

29
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
A espécie Litopenaeus schmitti popularmente conhecida como “Vila Franca”,
ocorre no Atlântico ocidental desde as Antilhas até o Rio Grande do Sul (Figura 04),
sendo os adultos encontrados em pequenas profundidades de até 47 metros
(PÉREZ-FARFANTE, 1970; SILVA, 1977).
Figura 04: Mapa da distribuição geográfica do camarão branco Litopenaeus schmitti .
Fonte: (MARTINELI, 2005).
O camarão branco apresenta características relacionadas à sua
distribuição e ao seu ciclo de vida que podem influenciar a estruturação em escala
fina de suas populações, como a ocorrência do acasalamento e postura próximos
a costa (entre 20 e 30 metros) e a permanência dos pré-adultos nas proximidades
de regiões estuarinas (SILVA, 1977). Nesta espécie, a postura aparentemente é
realizada em águas marinhas de pequena profundidade e de salinidade elevada
(EWALD, 1965; PÉREZ-FARFANTE, 1970; COELHO; SANTOS, 1994). A maior
parte das fêmeas em postura tem 7 meses de idade, sendo a idade média da

30
INTRODUÇÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
primeira maturação inferior a 6 meses; poucas se reproduzem novamente aos 10-
12 meses de idade (COELHO; SANTOS, 1994).
Sua distribuição está associada a substratos maciços e lamacentos com um
conteúdo orgânico relativamente alto e alta turbidez da água, característico de águas
estuarinas (DALL et al., 1990).
Há poucos estudos sobre essa espécie de camarão, principalmente quando
se diz respeito à enzimologia. Os trabalhos existentes abrangem outras espécies de
camarão ou aspectos ligados a identificação de genes relacionados ao sistema
imune de Litopenaeus vannamei (LIMA NETO, 2006), bioconversão de resíduos
para produção de quitosana (ASSIS et al., 2008), efeitos de íons mercúrio na
atividade da qutinase (LIN et al., 2005), caracterização dos glicosaminoglicanos
isolados do L. schmitti (SANTOS, 2006) e análise filogenética de quitinases do
camarão Penaeus monodon (PROESPRAIWONG et al., 2010).
Os estudos realizados em nosso laboratório sugerem uma ampla perspectiva
de trabalhos a serem realizados com endo-, exoglicosidases e sulfatases
encontradas em aninais marinhos como, por exemplo, a identificação de
glicodidases e sulfatases em diferentes tecidos do molusco Aplysia cervina (MATTA;
ABREU, 2005), indentificação de uma nova β-N-acetylhexosaminidase do cnidário
Palythoa caribaeorum (SOUZA et al., 2008) e a purificação de uma β-N-
acetylhexosaminidase do mamífero marinho Sotalia fluviatilis (GOMES JÚNIOR et
al., 2010).
O estudo e a purificação destas enzimas e suas utilizações biotecnológicas
como antifúngicos, antibacterianos e bioinseticidas é uma das propostas desta
pesquisa e servirão como ferramentas moleculares na elucidação de estrutura
química de glicoconjugados, dissacarídeos e/ou oligossacarídeos e de
polissacarídeos marinhos de algas marinhas, como ainda a produção de
oligossacarídeos com potenciais atividades farmacológicas.

31
OBJETIVOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
2. OBJETIVOS:
2.1. Gerais
Isolar, caracterizar cineticamente e realizar ensaios microbiológicos e larvicida
com a quitinase, obtida do camarão Litopenaeus schmitti.
2.2. Específicos
• Extrair glicosidases e sulfatases do cefalotórax do camarão marinho
Litopenaeus schmitti;
• Fracionar estas enzimas através de precipitação com sulfato de amônio
e identificar glicosidases e sulfatases nestas frações utilizando
substratos sintéticos específicos;
• Purificar a quitinase utilizando cromatografias;
• Propor a massa molecular da quitinase isolada;
• Realizar testes cinéticos com a Litochitinase1;
• Avaliar a atividade antibacteriana e antifúngica da Litochitinase1 sobre
cepas de Staphylococcus aureus, Candida albicans, Candida tropicalis,
Escherichia coli e Pseudomonas aeruginosa;
• Investigar a presença de quitina nos intestinos médios das larvas de
Aedes aegypti;
• Avaliar a atividade larvicida das frações ricas em Litochitinase1 contra
larvas de Aedes aegypti;
• Identificar a presença de proteínas bioativas como: proteinases serínicas
e lectinas, nas frações ricas em Litochitinase1;
• Avaliar a influência das enzimas digestivas de A. aegypti sobre a
Litochitinase1.

32
MATERIAIS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
3. MATERIAIS 3.1. Material Biológico
3.1.1 Camarão Litopenaeus schmitti
Figura 05: Litopenaeus schmitti. Fonte: Arquivo Próprio
Os camarões da espécie Litopenaeus schmitti foram obtidos no Mercado de
peixes situado na cidade do Natal, Estado do Rio Grande do Norte. Estes foram
mantidos a 4°C durante o transporte e depois armazenados a -20°C até seu
processamento.
3.1.2. Insetos
As larvas do quarto instar de Aedes aegypti foram obtidas da criação mantida
no Laboratório de Entomologia (LABENT) do Departamento de Microbiologia e
Parasitologia da UFRN. Estes experimentos foram realizados com o auxílio da aluna
de pós-graduação, a Veterinária (CRMV- RN 373) Patrícia Barra e da Profa. Dra.
Maria de Fátima F. de M. Ximenes.
Surperfilo: Artropoda
Filo: Crustacea
Classe: Malacostraca
Ordem: Decapoda
Família: Penaeidae
Gênero: Litopenaeus
Espécie: Litopenaeus schmitti

33
MATERIAIS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
3.1.3. Eritrócitos Humanos
Os eritrócitos humanos foram obtidos através de doações de bolsas de
sangue pelo HEMOCENTRO-RN. As bolsas fornecidas encontravam-se fora do
prazo de validade para transfusões.
3.1.4. Microorganismos
As cepas de Staphylococcus aureus (25923), Candida tropicalis (13803),
Escherichia coli (25922) e Pseudomonas aeruginosa (27853), foram obtidas do Banco
de Cepas ATCC (Americam Type Culture Collection) e Candida albicans (12) obtida
do banco ICB e mantidas no Laboratório de Micologia Médica e Ambiental (LAMEA)
no Departamento de Microbiologia e Parasitologia da UFRN. Estes experimentos
foram realizados sob a supervisão e orientação da Profa. Dra. Vânia Sousa Andrade e
com o auxílio da aluna de Iniciação Cientifica (Bolsista PROPESQ) Gabriela Medeiros
Araújo.
3.2. Substratos Sintéticos derivados de açúcares
p-nitofenil-sulfato; p-nitrofenil-N-acetil-β-D-Glucosaminídeo; p-nitrofenil-β-D-
glucopiranosídeo; p-nitrofenil-α-D-glucopiranosídeo; p-nitrofenil-N-acetil-α-D-
galactosaminídeo; 4-Nitrofenil N, N’-diacetil-β-D-quitobiosideo; 4-Nitrofenil β-D-N, N’,
N’’-triacetilquitotriose foram adquiridos da Sigma Chemical Company (St Louis, MO,
EUA).
3.3. Reagentes, Soluções, membranas e matrizes cromatográficas
• Acrilamida e bisacrilamida da Bio-Rad Laboratories (Hercules, CA, EUA),
dodecil sulfato de sódio (SDS) e ácido acético (C2H4O2) da Reagen, Quimibras
Indústrias Químicas S.A. Indústria Brasileira (Rio de Janeiro, RJ, Brasil);
• Hidróxido de sódio (NaOH), Álcool etílico (C2H6O) foi obtido da CIRQ
Cromato Produtos Químicos LTDA (São Paulo, SP, Brasil);

34
MATERIAIS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
• Albumina sérica bovina, ácido etilenodiamino tretra-acético (EDTA),
persulfato de amônio, glicina, N, N,N’ tetra-metileno diamino (TEMED), nitrato de
prata, “Coomassie brilhante blue” G 250, Azocaseína, Proteases Serínicas: Tripsina
e Quimotripsina, Protease Cisteínica: Papaína, da Sigma Chemical Company (St
Louis, MO, EUA);
• Padrões de massa molecular para eletroforese da Fermentas Life Sciences
(Ontário, Canadá);
• Tiossulfato de sódio (Na2S2O3), acetato de sódio (NaC2H3O2), cloreto de
sódio (NaCl), ácido clorídrico (HCL), cloreto de potássio (KCL), sulfato de amônio
((NH2)2SO4), e fosfato de sódio monobásico (Na2HPO4.H2O), acetona, sulfato de
sódio (Na2SO4), fosfato de sódio dibásico (NaH2PO4) e citrato de sódio (NaC6H5O7)
foram adquiridos da VETEC Química Fina LTDA (Rio de Janeiro, RJ, Brasil); ácido
tricloroacético (CCl3COOH) da MERCK (Darmstadt, Germany);
• Membranas de diálise (limite de exclusão: 6.000-8.000 e 12.000-14.000
dáltons) da Spectrum Medical Industries, Inc.(Houston TX, EUA);
• Matriz cromatográfica DEAE-Biogel A 1,5m da Bio Rad Laboratories
(Richmond, CA, EUA);
• Matriz cromatográfica Sephacryl S-200 da Sigma Chemical Company (St
Louis, MO, EUA).
3.4. Aparelhos
• Agitador orbital modelo 2525, Banhos e estufas de temperaturas constantes
da FANEM LTDA (São Paulo, SP, Brasil);
• Balança analítica – SCIENTECH, obtidos da QUIMIS Aparelhos Científicos
LTDA (Diadema – SP, Brasil);
• Bomba peristáltica modelo 18-1110-91 e coletor de frações modelo 18-1003-
64, Pharmacia Biotech, (Uppsala, Sweden);
• Centrifuga refrigerada modelo CR 21 da Hitachi Koki Co. LTDA (Tóquio,
Japão);
• Concentrador 5301 Eppendorf (Hamburgo, Germany);
• Espectrofotômetro Hitachi U 2000 (Tóquio, Japão) ;
• Lupa, Olympus (California, USA)

35
MATERIAIS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
• Medidor de pH Digimed (São Paulo, SP, Brasil);
• Microcentrífuga Eppendorf 5410 (Hamburgo, Germany);
• Sistema de eletroforese em gel vertical, modelo Mini-VE da Amersham
Biosciences (Uppsala, Suécia);
• Sistema de ultrapurificação de água Millipore modelo Milli-Q Plus (NYE, EUA)
• Fluxo Lâminar-Purifier Class II Biosafety Cabinet (Kansas City, EUA).

36
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
4. MÉTODOS
4.1. Obtenção do Extrato Bruto
Os camarões da espécie Litopenaeus schmitti foram descongelados, os
cefalotórax foram cuidadosamente separados da musculatura e homogeneizados
com dois volumes de tampão acetato de sódio 0,1 M pH 5,0, em banho de gelo. Em
seguida centrifugados a 12.000 x g (4°C) e as fases solúveis (extratos brutos)
utilizados para posterior fracionamento utilizando-se sulfato de amônio.
4.2. Fracionamento com Sulfato de Amônio
O fracionamento com sulfato de amônio foi efetuado em três etapas de
saturação: 0-30%, 30-50% e 50-80%. A concentração de sal, correspondente a cada
saturação, foi adicionado lentamente sobre a amostra sofrendo leve agitação e,
posteriormente deixado por 18 horas a 4°C (geladeira). Em seguida foram
centrifugados a 12.000 x g, os precipitados ressuspensos em tampão acetato de
sódio 0,1 M pH 5,0 e dialisados por 18 horas contra o mesmo tampão (quatro
vezes). Todos estes processos foram executados a 4°C. Estas frações foram
denominadas F-I, F-II e F-III, referindo-se aos percentuais de saturação 0-30%, 30-
50% e 50-80%, respectivamente (Figura 06).

37
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Figura 06: Fluxograma do fracionamento com sulfato de amônio.
4.3. Determinação das Atividades Enzimáticas Com p-Nitrofenil Derivados de
Açúcar
As incubações com p-nitrofenil derivados de açúcar (p-nitrofenil-α-D-
galactopiranosideo, p-nitrofenil-sulfato, p-nitrofenil-β-D-glucopiranosídeo, p-nitrofenil-
β-D-N-acetilglucosaminídeo e p-nitrofenil-α-D-glucopiranosídeo, 4-Nitrofenil N, N’-
diacetil-β-D-quitobiosideo, 4-Nitrofenil β-D-N, N’, N’’-triacetilquitotriose) foram
realizadas utilizando-se 10 mM de cada substrato sintético com alíquotas do extrato
bruto, das frações precipitadas com sulfato de amônio e cromatográficas (volume
final de 100 µL). Os ensaios foram realizados no tempo de 30 minutos a 37°C para o
EB e F-III e 2 horas a 55°C para as cromatografias. As reações foram interrompidas
com 1 mL de hidróxido de sódio 0,25 N e o p-nitrofenol liberado lido em
Homogenizado com tampão acetato de sódio 0,1M pH 5,0 12.000 x g – 30’, 4 ºC
0-30% Sulfato de amônio 12.000 xg –30’, 4 ºC
30-50% Sulfato de amônio 12.000 xg –30’, 4 ºC
50-80% Sulfato de amônio 12.000 xg –30’, 4 ºC
Sobrenadante (Extrato Bruto)
Precipitado (F-I) Sobrenadante
Precipitado (F-II)
Precipitado (F-III)
Sobrenadante

38
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
espectrofotômetro a 405 nm. Para cada ensaio foram feitos controles negativos dos
substratos (substrato e tampão no mesmo volume e concentração do ensaio). Uma
unidade de atividade foi definida como a quantidade de enzima capaz de aumentar
em 0,01 unidade de absorbância a 405 nm.
4.4. Dosagem de Proteínas
Para a quantificação das proteínas utilizamos o método de SEDMAK;
GROSSBERG (1977) com albumina sérica bovina como padrão.
4.5. Purificação da Enzima
4.5.1. Cromatografia de Troca Iônica DEAE-Biogel
A fração F-III (9,32 mg), proveniente do fracionamento com sulfato de amônio,
foi submetida à cromatografia de troca-iônica DEAE-Biogel. A coluna com cerca de
10 cm3 de gel foi equilibrada em tampão fosfato de sódio 0,02 M pH 8,0. Para a
remoção das proteínas, que não se complexaram com o gel, foi feita eluição (fluxo
de 1 mL/3min) com o mesmo tampão até que nenhuma proteína fosse detectada no
eluato. Este pico protéico é, geralmente, denominado de não-retido. Em seguida, as
proteínas retidas, foram eluídas da coluna utilizando-se o mesmo tampão, acrescido
de cloreto de sódio nas concentrações de 0,2; 0,4; 0,6; 0,8 e 1,0M. Todas as etapas
foram realizadas a 4°C. O perfil de eluição protéica das frações da coluna foi
determinado por absorção a 280 nm. Para determinação das atividades enzimáticas,
alíquotas de 20 µL (0,062 µg) destas frações foram ensaiadas a 37°C durante uma
hora com p-nitrofenil-N-acetil-β-D-glucosaminídeo. O p-nitrofenol liberado foi lido a
405 nm em espectrofotômetro.
4.5.2. Cromatografia de Exclusão Molecular – Sephacryl S – 200
Cerca de 2 mL (1 mg), da fração eluída a 0,2 M de NaCl proveniente da
cromatografia de troca iônica DEAE-Biogel, foi aplicada em cromatografia de
exclusão molecular Sephacryl S-200 (com diâmetro de 104 cm x 0,9 cm), equilibrada

39
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
com tampão acetato de sódio 0,1 M pH 5.0. As proteínas foram eluídas utilizando-se
o mesmo tampão com o fluxo de 0,4 mL por minuto, sendo coletadas frações de 2
mL/5 min e armazenadas a 4°C até o uso. O perfil de eluição protéica foi monitorado
em espectrofotômetro a 280 nm e para a determinação das atividades enzimáticas,
alíquotas de 50 µL destas frações foram avaliadas a 45°C durante 2 horas com p-
nitrofenil-N-acetil-β-D-glucosaminídeo. O p-nitrofenol liberado foi lido a 405 nm em
espectrofotômetro.
4.6. Eletroforese em gel de Poliacrilamida em condições desnaturantes-
SDS/PAGE
Para avaliar o grau de pureza da quitinase e determinar a massa molecular
aparente, alíquotas (20 µg) das frações: extrato bruto, F-III, pico protéico de maior
atividade quitinásica eluído da cromatografia de troca iônica DEAE-Biogel e exclusão
molecular Sephacryl S-200 foram submetidos a uma eletroforese em gel de
poliacrilamida 10% em presença de SDS, de acordo com a metodologia descrita por
LAEMMLI, (1970). As frações foram precipitadas com acido tricloroacético 90% na
razão de 9:1. Após 30 minutos de precipitação em banho de gelo, as amostras foram
centrifugadas durante 15 minutos a 15.000 x g, o sobrenadante foi descartado e ao
precipitado adicionado acetona e uma nova centrifugação foi realizada nas mesmas
condições da anterior. Uma vez descartado o sobrenadante, o precipitado foi
ressuspenso em 20 µL de tampão de amostra (azul de bromofenol 5%, SDS 20% e
sacarose 10%). A corrida eletroforética foi realizada a uma corrente de 30mA por
aproximadamente 2 horas. Para a revelação das bandas protéicas, o gel foi corado
com coomassie blue R 250 0,1% em solução de metanol 40% e ácido acético 10%,
e descorado com a solução de metanol 40% e ácido acético 10%.
Para a detecção de proteínas na ordem de nanogramas, o gel foi corado com
nitrato de prata de acordo com a seguinte metodologia: O gel corado, como já
descrito, foi desidratado seqüencialmente com três lavagens em solução de etanol
50%, por 20 minutos cada. Em seguida, o gel foi colocado sob agitação durante 1
minuto em solução de tiossulfato de sódio 0,02%. Este foi lavado rapidamente três
vezes com água destilada e, em seguida, foi adicionado solução de nitrato de prata
0,2% acrescida de 74 µl de formaldeído 37% mantendo-se por 20 minutos sob

40
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
agitação. O gel foi lavado novamente três vezes com água destilada e adicionou-se
solução reveladora de carbonato de sódio 6%, acrescida de 50 µl de formaldeído
37% e 2 mL de tiossulfato de sódio 0,02%. A reação foi interrompida com acido
acético 13% após obtida a coloração desejada.
4.7. Estudos Cinéticos
Os experimentos cinéticos foram realizados incubando-se a quitinase, obtida
purificada da cromatografia de exclusão molecular Sephacryl S-200, com o p-
nitrofenil-N-acetil-β-D-glucosaminídeo como substrato. Os ensaios foram realizados
em duplicatas, com seus respectivos controles negativos (tampão e substrato no
mesmo volume, concentrações, temperaturas e pH dos ensaios).
4.7.1. Determinação da constante de Michaelis-Menten (Km)
Com o objetivo de se determinar o Km aparente da quitinase para a hidrólise
do p-nitrofenil-N-acetil-β-D-glucosaminídeo, alíquotas de 50 µl (4,05 µg) da enzima
foram incubadas a 55°C durante 2 h com diferentes concentrações finais do p-
nitrofenil-N-acetil-β-D-glucosaminídeo (0,25; 0,50; 0,75; 1,0; 1,25; 1,5; 2,0; 2,5; 3,0 e
3,5 mM). Os ensaios foram feitos em duplicatas, sendo interrompidos com 1 mL de
NaOH 0,25 N e o p-nitrofenol liberado lido em espectrofotômetro a 405 nm.
Controles negativos dos substratos foram feitos para cada concentração utilizada.
Os valores de Km foram calculados por meio do programa de computador ORIGIN
5.0.
4.7.2. Efeito da temperatura sobre a atividade enzimática
A influência da temperatura sobre a atividade da quitinase foi realizada
incubando-se 50 µl (4,05 µg) desta e 1,5 mM do p-nitrofenil-N-acetil-β-D-
glucosaminídeo durante 2 horas nas temperaturas de 4; 25; 37; 45; 50; 55; 60; 70 e
80°C. Transcorrido cada ensaio as reações foram interrompidas como descrito em
3.3.

41
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
A fim de testar a estabilidade da quitinase frente a variadas temperaturas,
foram feitas pré-incubações usando 50 µl (4,05 µg) da enzima durante 30 minutos a
25, 37, 45, 50, 55, 60, 70, 80 e 100°C na ausência do substrato. Transcorrido o
tempo, o p-nitrofenil-N-acetil-β-D-glucosaminídeo foi adicionado para uma
concentração final de 1,5 mM e incubado a 55°C durante 2 h. Em seguida as
reações foram interrompidas como descrito em 3.3.
4.7.3. Influência do pH sobre a atividade enzimática
Para a realização destes experimentos alíquotas de 200 µl (16,2 µg) da
quitinase foram dialisadas durante 18 horas contra os tampões: glicina-HCl pH 3,0;
3,5 e 4,0; acetato de sódio pH 4,0; 5,0 e 5,5; fosfato de sódio pH 5,5; 6,0 e 7,0 e
Tris-HCl pH 7,0; 8,0 e 9,0. Todos os tampões estavam na concentração de 0,1 M.
Após a diálise alíquotas de 50 µl (4,05 µg) da enzima dialisada, nos tampões já
citados, foram incubadas com 1,5 mM de p-nitrofenil-N-acetil-β-D-glucosaminídeo
durante 2 horas a 55°C. Transcorrido o tempo de ensaio as reações foram
interrompidas e quantificadas a 405 nm em espectrofotômetro.
A fim de testar a estabilidade da quitinase frente a variações de pHs,
alíquotas de 200 µl (16,2 µg) da quitinase foram dialisadas durante 18 horas contra
os tampões: glicina-HCl pH 3,0; 3,5 e 4,0; acetato de sódio pH 4,0; 5,0 e 5,5; fosfato
de sódio pH 5,5; 6,0 e 7,0 e Tris-HCl pH 7,0 ;8,0 e 9,0. Foram feitas pré-incubações
usando 50 µl (4,05 µg) da enzima durante 30 minutos a 50°C na ausência do
substrato. Transcorrido o tempo, o p-nitrofenil-N-acetil-β-D-glucosaminídeo foi
adicionado para uma concentração final de 1,5 mM e incubado a 55°C durante 2 h.
Em seguida as reações foram interrompidas como descrito em 3.3.
4.7.4. Influência de diversos sais e EDTA na hidrólise do p-nitrofenil-N-acetil-ββββ-
D-Glucosaminídeo.
Alíquotas de 50 µL (4,05 µg) da enzima foram pré-incubadas a 50°C, durante
15 minutos, na presença de diversos sais e EDTA a uma concentração final de 10
mM. Transcorrido este tempo, foi adicionado o p-nitrofenol-N-Acetil-β-D-
Glucosaminídeo numa concentração final de 1,5 mM e procedemos com o ensaio a

42
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
55°C, e após 2 h, os ensaios foram interrompidos com 1mL de NaOH 0,25 N e o p-
nitrofenil liberado, lido em espectrofotômetro a 405 nm. Os sais utilizados nesse
experimento foram: Cloreto de Magnésio (MgCl2), Cloreto de Potássio (KCl), Cloreto
de Cálcio (CaCl2), Tiossulfato de sódio pentahidratado (Na2S2O25H2O), Sulfato de
sódio (Na2SO4), Fosfato de sódio dibasico (NaH2PO4), Fosfato de sódio monobásico
(Na2HPO4.H2O), Citrato de sódio(NaC6H5O7), cloreto de sódio (NaCl), além do ácido
etilenodiamino tetra-acético (EDTA).
4.8. Preparação da quitinase para os ensaios biológicos
A Litochitinase1, foi submetida a precipitação em 2 volumes de acetona,na
qual esta foi adicionada lentamente sobre a amostra sofrendo leve agitação e,
posteriormente deixada por 18h a 4°C (geladeira). Em seguida foi centrifugada a
12.000 x g, o sobrenadante descartado e o precipitado ressuspenso em tampão
acetato de sódio 0,1 M pH 5,0 para a realização dos ensaios biológicos. Todos estes
processos foram executados a 4°C.
4.9. Avaliação da atividade antimicrobiana
4.9.1. Análise Qualitativa- Técnica de Difusão em Disco
Para avaliação in vitro da atividade antimicrobiana da quitinase foi
inicialmente aplicada à técnica de difusão em disco (BAUER et al., 1966). A partir
de cultivo em Ágar Mueller-Hinton e Ágar Saboraund foram preparadas suspensões
ajustadas pela escala 0,5 Mc Farland, para as bactérias (Staphylococcus aureus) e
leveduras (Candida albicans e Candida tropicalis). Em seguida as suspensões foram
plaqueadas em Ágar Mueller-Hinton e Saboraund para as bactérias e leveduras,
respectivamente. Posteriormente Discos de papel filtro (Whatman 6 mm), foram
impregnados, com 20 µL da quitinase nas concentrações de 676 µg/mL e 1600
µg/mL, e fixados sobre a superfície do meio inoculado (Quadro 1). As placas foram
incubadas em estufa bacteriológica por 24 horas, a 35 ºC. Os halos de inibição do
crescimento bacteriano e fúngico foram medidos em milímetros com auxílio de uma
régua milimetrada. Foi realizado o controle negativo utilizando, água destilada e/ ou
tampão acetato de sódio 100 mM, pH 5,0.

43
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Quadro 1: Técnica de Difusão em Disco, cepas selecionadas e concentrações da quitinase utilizada
TÉCNICA MICROORGANISMO AMOSTRA TESTADA
(QUITINASE)
Difusão em disco
S. aureus 676 µg/mL Acetato de sódio 100 mM, pH 5,0
C. albicans 1600 µg/mL Acetato de sódio 100 mM, pH 5,0
1600 µg/mL água
C. tropicalis 1600 µg/mL Acetato de sódio 100 mM, pH 5,0
4.9.2. Ánalise Quantificativa- Determinação da Concentração Inibitória
Mínima (CIM)
A CIM foi obtida por meio do método de Microdiluição em caldo, de acordo
com a metodologia descrita no Clinical and Laboratory Standards Institute (CLSI
2003). Os microorganismos utilizados nesta técnica foram: Staphylococcus aureus,
Candida albicans, Escherichia coli e Pseudomonas aeruginosa.
A quitinase apartir de diluições seriadas foi testadas nas concentrações finais
de 800, 400, 200 e 100 µg/mL. O teste foi realizado pela distribuição em placas de
microdiluição estéreis contendo 96 poços, em cada orifício teste e de controle de
crescimento foram adicionados 20 µL de inoculo microbiano e 100 µL dos meios BHI
e caldo Saboraund para bactérias e leveduras, respectivamente. Os orifícios testes
receberam também 50 µL da quitinase na concentração inicial de 1.600 mg/mL, já os
orifício contendos os controles negativos continham 50 µL de tampão acetato de
sódio 100 mM, pH 5,0 e os controles positivos continham 20 µL de gentamicina na
concentração de 40 µg (Quadro 2). Em seguida, as placas foram incubadas em
estufa bacteriológica por 24 horas, a 37 ºC para as bactérias e por 24 horas, a 28 ºC
para as leveduras. Para leitura, que foi realizada visualmente, após o tempo de 3 h,
foram adicionados 20 µL de cloreto de, 2,3,5 – trifenil – tetrazólio (CTT) em cada
orifício, indicador de multiplicação bacteriana, que apresenta coloração
avermelhada na presença de células viáveis.

44
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Quadro 2: Técnica de Microdiluição em Caldo, cepas selecionadas e concentrações da quitinase utilizada
TÉCNICA MICROORGANISMO AMOSTRA TESTADA
(QUITINASE)
Microdiluição em Caldo
C. albicans 800µg/mL Acetato de sódio 100 Mm, pH 5,0
800 µg/mL água
S. aureus 800 µg/mL Acetato de sódio 100 Mm, pH 5,0
E. coli
800µg/mL Acetato de sódio 100 Mm, pH 5,0
800 µg/mL água
P.aeruginosa 800µg/mL Acetato de sódio 100 Mm, pH 5,0
*Para os ensaios de microdiluição em caldo foram realizadas diluições seriadas
4.10. Ensaios com Aedes aegypti
4.10.1. Obtenção das larvas de Aedes aegypti
Armadilhas (Ovitrampas) foram colocadas em diversos pontos no Campus
Universitário da UFRN e no Bairro Nordeste (elevado índice de infestação predial),
Natal-RN. Semanalmente as palhetas foram trazidas para o LABENT, Laboratório
de Entomologia Médica da UFRN, e após a contagem dos ovos, foram submersas
em uma bandeja plástica (30 cm x 20 cm x 15 cm) com água limpa contendo ração
para peixes autoclavada. Após a eclosão as palhetas foram removidas e as larvas
identificadas em microscópio estereoscópico como sendo de Aedes aegypti, onde
posteriormente foram utilizadas nos ensaios.
4.10.2. Dissecação das larvas de A. aegypti
Cem larvas de A. aegypti no quarto instar foram dissecadas em banho de
gelo, com o auxílio de uma lupa, e trato intestinal das larvas foi exposto utilizando-se
pinças e transferido para microtubos de ensaio contendo solução tampão Tris-HCl

45
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
50 mM pH 7,5. Todos os materiais coletados foram mantidos a -20°C, até a extração
do homogenato intestinal e utilização nos ensaios de inibição.
4.10.3. Preparo do homogenato intestinal das larvas
O homogenato intestinal foi preparado seguindo a metodologia estabelecida
por TERRA et al. (1997), com algumas modificações. Os intestinos das larvas dos
insetos foram homogeneizados com o auxílio de um pistilo em tubos eppendorff, em
banho de gelo, por aproximadamente 10 minutos, usando-se como extrator 800 µL
de Tris-HCl 50 mM pH 7,5. Em seguida os homogenatos foram centrifugados a
10.000 x g durante 15 minutos a 4°C. Os sobrenadantes obtidos foram utilizados
posteriormente para os ensaios de inibição.
4.10.4. Detecção de quitina no intestino das larvas de A. aegypti
Os intestinos das larvas foram retirados com auxílio de pinça e bisturi, e em
seguida reservados. Os intestinos foram então lavados para a remoção dos
conteúdos luminais remanescentes. A presença da quitina foi verificada por meio do
teste de Von Wisselingh (ROGER; PERKINS, 1968). Este teste qualitativo detecta
quitosana produzida após tratamento térmico do material contendo quitina, com
NaOH saturado por 15 minutos a 160 °C. Resíduos foram lavados com diferentes
concentrações de álcool etílico: 10, 20, 30, 40, 80 e 95%. Após a reação, a presença
de quitina foi observada com solução de lugol e, em seguida, ácido sulfúrico a 1%.
Controle positivo foi feito utilizado quitina de lagosta e para controle negativo foi
utilizado celulose.
4.10.5. Ensaios com larvas de A. aegypti
A atividade larvicida da Litochitinase1 foi avaliada utilizando uma adaptação
da Organização Mundial de Saúde (1981). Cinco larvas do quarto estádio (L4) foram
colocadas em recipientes plásticos descartáveis contendo os extratos brutos, F-III e
S-200. As concentrações utilizadas para o extrato bruto foram 1,55; 3,8; 5,7 e 7,22
mg/mL. Já as concentrações para a F-III foram 1,23; 1,84; 3,08 e 4,31 mg/mL e para
a Litochitinase1 foram utilizadas as seguintes concentrações: 0,472; 0,675; 0,9 e
1,12 mg/mL. O volume final do ensaio larvicida foi de 20 mL de solução de teste ou
controle negativo. Os ensaios foram realizados em quadruplicatas e a taxa

46
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
de mortalidade das larvas foi determinada após 24 e 48 h de incubação a 28 ± 2 ° C
com fotoperíodo de 12/12 h (claro-escuro). As larvas foram consideradas mortas
quando não respondiam a estímulos mecânicos como, por exemplo, a agitação aos
recipientes plásticos.
4.10.6. Determinação da EC50
A concentração da quitinase que mata 50% das larvas L4 da Aedes aegypti
foi determinada pela construção de uma curva regressão linear simples a qual
relaciona o percentual de mortalidade com a concentração de enzima utilizada no
ensaio.
4.10.7. Análises Estatísticas
A análise estatística dos dados experimentais foi realizada utilizando o
software de computador GraphPad Prisma 5® para Windows® (GraphPad Software,
Inc, USA), para determinar a significância das diferenças (p <0,05) entre os
tratamentos, por meio dos testes ANOVA e Kruskal-Wallis.
4.10.8. Investigação de proteínas bioativas interferentes no processo
digestório nas larvas de Aedes aegypti
4.10.8.1. Ensaio de inibição de proteases serínicas
a) Preparo do Substrato
Azocaseína a 1% e 1,5% (substrato protéico) foi solubilizada em 50 mL de
tampão Tris-HCL 50 mM pH 7,5. A solução foi fervida por cerca de 10 minutos. Após
resfriamento o volume foi completado com água destilada. A solução foi conservada
a -20 ºC até sua utilização.
b) Atividade anti-quimotríptica
A atividade inibitória sobre a quimotripsina foi determinada utilizando 20 µL de
solução de quimotripsina bovina (0,2 mg/mL em tampão Tris-HCL 50 mM, pH 7,5,
contendo CaCl2 20 mM) pré-incubada em tampão Tris-HCL 50 mM, pH 7,5, contendo
CaCl2 20 mM e 100 µL das amostras enzimáticas (EB (1557,7 µg) , F-III (1121,4 µg)

47
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
e a Litochitinase1 (8,1µg)) por 15 minutos a 37 °C. Após esse período, a reação foi
iniciada adicionando-se 200 µL de azocaseína 1%. Descorridos 30 minutos, a
reação foi interrompida adicionando-se 300 µL de solução de TCA 20%. A mistura
da reação foi centrifugada a 12.000 x g por 10 minutos e o sobrenadante
alcalinizado com NaOH 2 N na proporção de 1:1. A absorbância foi medida em
espectrofotômetro a 440 nm. Provas em branco foram realizadas e os ensaios foram
realizados em triplicatas.
c) Atividade anti-tríptica
A atividade anti-tríptica foi determinada utilizando azocaseína como substrato
alíquotas de 20 µL de solução de tripsina bovina (0,3 mg/mL em tampão Tris-HCl 50
mM pH 7,5), foram pré-incubadas por 15 minutos a 37 °C, com 120 µL de HCl 2,5
mM, 350 µL de tampão Tris-HCl 50 mM pH 7,5 e 200 µL da amostra (EB (3115,4
µg), F-III (2242,8 µg) e a Litochitinase1(16,1 µg)). Após isso a reação foi iniciada
adicionando-se 200 µL de azocaseína 1%. Descorridos 30 minutos, a reação foi
interrompida adicionando-se 300 µL de solução de TCA 20%. A mistura da reação
foi centrifugada a 12.000 x g por 10 minutos e o sobrenadante alcalinizado com
NaOH 2 N na proporção de 1:1. A absorbância foi medida em espectrofotômetro a
440 nm. Provas em branco foram realizadas e os ensaios foram realizados em
triplicatas.
4.10.8.2. Ensaios de Hemaglutinação
a) Preparo do sangue
Alíquotas de 2 mL de sangue foram lavadas com 8 mL de solução salina
fisiológica e centrifugadas a 924 x g, até a obtenção de uma massa de eritrócitos
íntegros livre de soro e material hemolisado.
b) Tratamento dos eritrócitos com Papaína
Uma solução estoque de papaína de 1 mg/mL em solução salina foi
adicionada aos eritrócitos, previamente lavados na proporção de 1:1 (v/v). A mistura

48
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
foi incubada por 30 minutos a 37 °C, com leve agitação ocasional, Em seguida foi
centrifugada a 924 x g, por 5 minutos e seu precipitado foi lavado 6 vezes com
solução salina. Realizou-se o hematócrito e em seguida uma solução de eritrócitos
na concentração de 4% foi preparada.
c) Tratamento dos eritrócitos com Tripsina
Uma solução estoque de tripsina na concentração de 1mg/mL foi preparada e
adicionada a 1 mL de eritrócitos, previamente lavados. A mistura foi deixada em
repouso por 1 hora, a temperatura ambiente, com leve agitação ocasional. Em
seguida, foi centrifugada a 924 x g, por 5 minutos e seu precipitado lavado por 6
vezes com solução salina. Realizou-se o hematócrito e uma solução de eritrócitos
diluída a 4% em solução salina foi preparada.
d) Ensaio
Os testes de hemaglutinação foram realizados por meio de diluição seriada
das amostras testes em microplacas com fundo em “V”. No primeiro poço foram
adicionados 25 µL da amostra (EB (389,4 µg) e/ou F-III (280,3 µg) e/ou
Litochitinase1 (2 µg)) e 25 µL de uma suspensão de eritrócitos a 4% tratados
enzimaticamente enquanto que, a partir do segundo em diante foram adicionados 25
µL de solução salina, 25 µL da amostra diluída seriadamente e 25 µL da suspensão
de eritrócitos a 4%, submetido a tratamentos enzimáticos. A reação foi incubada por
1 hora, a temperatura ambiente. O grau de aglutinação foi observado visualmente e
o título expresso em unidade de hemaglutinação (UH), que é definido como o
inverso da maior diluição da amostra que tenha apresentado nítida aglutinação
(MOREIRA et al., 1977).
4.10.8.3. Efeito do homogenato intestinal das larvas de A. aegypti sobre a
atividade quitinolítica
a) Determinação de atividade Azocaseinolítica
A atividade azocaseinolítica foi pesquisada por incubação de 30, 50 e 70 µL
de homogenato intestinal com 450 µL de tampão Tris-HCl 50mM pH 7,5 e 500 µL de

49
MÉTODOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
solução de azocaseína 1,5% por 30 minutos a 37 °C. A reação foi paralisada com
150 µL de TCA 20%. Transcorridos 30 minutos, a suspensão foi centrifugada por 15
minutos, a 12000 x g a temperatura ambiente. Alíquotas de 800 µL do sobrenadante
foram alcalinizadas com 800 µL de NaOH 2 N. Todos os ensaios foram feitos em
triplicatas e provas em branco foram realizadas. A atividade azocaseinolítica foi
medida pela absorbância a 440 nm.
b) Efeito da temperatura sobre a atividade azocaseinolítica a pH 7,5
O efeito da temperatura foi avaliado utilizando as seguintes temperaturas: 5,
10, 20, 30, 40, 50, 60, 70, 80 e 90 °C. Os ensaios foram realizados com descrito no
item 4.11.4.
c) Influência das enzimas digestivas do A. aegypti na hidrólise do p-
nitrofenil-N-acetil-ββββ-D-Glucosaminídeo
Alíquotas de 50 µL (4,05 µg) da Litochitinase1 foram pré-incubadas a 50°C,
durante 15 minutos, na presença do homogenato intestinal das larvas do mosquito
A. aegypti. Transcorrido este tempo, foi adicionado o p-nitrofenol-N-Acetil-β-D-
Glucosaminídeo numa concentração final de 1,5 mM e procedemos com o ensaio a
55°C, e após 2 h, o ensaio foi interrompido com 1 mL de NaOH 0,25 N e o p-
nitrofenil liberado, lido em espectrofotômetro a 405 nm.

50
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
5. RESULTADOS
5.1. Identificação de glicosidases e sulfatases nos cefalotórax dos camarões
marinhos Litopenaeus schmitti
A identificação das atividades enzimáticas foi pesquisada no EB e nas frações
precipitadas com sulfato de amônio (F-I, F-II e F-III) obtidas dos cefalotórax dos
camarões. Para isso foram utilizados alguns substratos sintéticos derivados de
açúcar. Os resultados mostraram a presença de quase todas as atividades
enzimáticas pesquisadas no extrato bruto, F-I, F-II e F-III (Figura 07).
É possível observar ainda, no gráfico da figura 07, que os ensaios utilizando o
EB e as amostras F-I, F-II e F-III, mostraram que, de todas as atividades
pesquisadas, a N-acetil-β-D-glucosaminidase (quitinase) foi a que mais se
sobressaiu em todos estes testes apresentado maior atividade específica na F-III.
Devido a isto a F-III, foi submetida ao fracionamento em cromatografia de troca
iônica DEAE-Biogel.

51
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
5.2. Purificação da Quitinase
5.2.1. Em cromatografia de troca-iônica DEAE-Biogel
A fração F-III, por ser mais rica na atividade quitinásica, foi submetida (9,32
mg) a cromatografia de troca iônica DEAE-Biogel. Nesta etapa de purificação
observou-se inicialmente a eluição de um pico não retido, eluído com o tampão de
equilíbrio da coluna e, em seguida, a eluição de picos protéicos utilizando-se as
concentrações de NaCl de: 0,2; 0,4; 0,6; 0,8 e 1,0M (Figura 08). Todas as frações
eluídas da cromatografia foram incubadas com o p-nitrofenil-N-acetil-β-D-
glucosaminídeo para o acompanhamento da atividade enzimática e eluição da
quitinase. Estes testes mostraram a eluição da quitinase apenas no pico protéico
eluído com 0,2 M de NaCl. Nas outras molaridades de NaCl, não observou-se a
Figura 07: Atividades enzimáticas presentes no extrato bruto e nas frações obtidas da precipitação com sulfato de amônio do cefalotórax do camarão marinho Litopenaeus schmitti. Alíquotas das frações do extrato bruto, F-I, F-II e F-III foram incubadas com cada p-nitrofenil para uma concentração final de 10 mM a 37 ºC, durante 30 minutos. Estas foram interrompidas pela adição de 1 mL de NaOH 0,25 N e o p-nitrofenol liberado é lido em espectrofotômetro a 405 nm. Onde: Bruto, é o extrato bruto antes da precipitação com sulfato de amônio, F-I, F-II e F-III, frações precipitadas com 0-30%, 30-50% e 50-80% de sulfato de amônio, respectivamente. A atividade específica foi encontrada através da relação entre a quantidade de p-nitrofenol liberado e a massa em µg de proteína contida no volume de cada fração ensaiada, por um período de 30 minutos.
Frações
Bruto F-I F-II F-III
β-D-glucosaminidase
α-D-glucosaminidase
Sulfatase
α-D-galactosaminidase
N-acetil-β-D-glucosaminidase
0
0,005
0,01
0,015
0,02
0,025
0,03

52
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
presença de atividade enzimática (Figura 08). O pico protéico e de atividade
quitinásica eluído com 0,2 M de NaCl foram reunidos, dialisados contra tampão
acetato de sódio 0,1 M pH 5,0, concentrados com dois volumes de acetona e
aplicado em cromatografia de exclusão molecular Sephacryl S-200.
5.2.2. Em cromatografia de Exclusão molecular
O pico protéico eluído com 0,2 M de NaCl oriundo da cromatografia de troca-
iônica que apresentou atividade enzimática, foi aplicado em cromatografia de
exclusão molecular Sephacryl S-200. Esta cromatografia revelou um pico protéico de
elevada absorbância a 280 nm, mas sem atividade enzimática (Figura 09). A
quitinase foi eluída em um pequeno pico protéico, anterior ao principal, e com
elevada atividade específica. Este pico protéico foi visualizado em SDS-PAGE e
Figura 08: Perfil de eluição protéico da F-III em cromatografia de troca iônica e de atividade quitinásica. Cerca de 2 mL da F-III (~9,32 mg) foi submetida a cromatografia de troca iônica. A coluna com cerca de 10 cm3 de gel foi equilibrada em tampão fosfato de sódio 0,02 M pH 8,0. A eluição (fluxo de 1 mL/3 min) foi realizada com o mesmo tampão até que a leitura de proteínas se aproximasse de zero, em seguida, utilizamos como tampão de eluição (fosfato de sódio 0,02 M pH 8,0 acrescido de NaCl nas concentrações de 0,2; 0,4; 0,6; 0,8 e 1 M). As proteínas das frações eluidas (1 mL) foram lidas a 280 nm e o perfil das atividades realizado com o p-nitrofenil-N-acetil-β-D-glucosaminideo com leituras a 405 nm.
00,050,10,150,20,250,30,350,40,450,5
00,020,040,060,08
0,10,120,140,160,18
0,2
1 9 17 25 33 41 49 57 65 73 81 89 97 105
D.O
. 40
5 n
m
D.O
. 28
0 n
m
Frações
proteina N-acetil-beta-D-glucosaminidase
NR 0,2 M 0,4 M 0,6 M 0,8 M 1 M
Atividade

53
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
posteriormente utilizado nos testes cinéticos. Esta fração por ser rica em atividade
quitinásica, a enzima foi denominada Litochitinase1.
5.2.3. Resumo das etapas de purificação protéica
As etapas utilizadas na purificação da Litochitinase1 estão sumarizadas na
tabela de purificação (Tabela 3). É possível identificar um índice de purificação final
de 987,32 vezes e uma recuperação de 2,62%, com a remoção de uma grande
quantidade de proteínas durante o processo de isolamento entre as etapas F-III e as
cromatografias.
Figura 09: Perfil de eluição da Litochitinase1 e atividade quitinásica em cromatografia de exclusão molecular (Sephacryl S-200). A coluna foi equilibrada e eluída com tampão acetato de sódio 0,1 M pH 5,0. Foram coletadas frações de 2 mL/5 min. A eluição das proteínas foi monitorada em espectrofotômetro a 280 nm e o perfil das atividades com o p-nitrofenil-N-acetil-β-D-glucosaminídeo determinadas por leitura a 405

54
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Etapas Proteína
total (mg)
Ativ. total
(U)
Ativ.
Específica
(U)
Purificação
(X)
Recuperação
(%)
EB 4501,06 1551,200 0,34 1 100
F-III 396,414 265,625 0,67 2 17,12
DEAE-Biogel 51,026 37,050 0,72 2,12 2,38
Litochitinase1 0,12129 40,716 335,69 987,32 2,62
5.3. Perfil eletroforético das amostras em SDS-PAGE
Para avaliar o grau de pureza e determinar a massa molecular aproximada da
Litochitinase1, alíquotas do EB, F-III, pico 0,2 M da DEAE-Biogel e pico de atividade
da S-200 foram submetidas à eletroforese em gel de poliacrilamida na presença de
SDS (Figura 10). A análise do gel mostrou um grau de pureza relevante do pico com
atividade da S-200 onde apenas uma banda protéica é observada na região de 28,5
kDa. Esse valor foi determinado pela correlação entre o logarítimo do peso
molecular das proteínas padrões versus o logarítimo de migração das proteínas.
Tabela 3: Etapas de purificação da Litochitinase1
Tabela construída utilizando-se os resultados obtidos nos ensaios enzimáticos com o p-nitrofenil-N-acetil-β-D-glucosaminideo. Onde: Atividade Total (U) = 1U corresponde a 0,01 unidade de absorbância a 405 nm; Purificação = Razão entre a atividade específica em cada passo de purificação e a atividade específica do extrato bruto; Recuperação = Percentual de atividade total em cada passo de purificação em relação a atividade total no extrato bruto (100%).

55
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
5.4. Experimentos cinéticos
A Litochitinase1 foi utilizada para realizar os estudos cinéticos. Nestes
experimentos foi utilizado p-nitrofenil-N-acetil-β-D-glucosaminideo como substrato,
variando-se sua concentração, como também as condições de temperatura e pH dos
ensaios.
5.4.1. Determinação do Km
Estes experimentos foram realizados incubando-se a Litochitinase1 com
diferentes concentrações do substrato (Figura 11). Os resultados mostraram um Km
aparente de 0,51 mM obitido pelo calculo utilizando o programa ORIGIN 5.0.
Observa-se ainda que a partir da concentração de 1,5 mM de substrato, a atividade
enzimática tende a estabilizar até atingir a velocidade máxima da reação.
Figura 10: SDS-PAGE das amostras protéicas durante o processo de purificação
Onde: M- mistura de proteínas de diferentes pesos moleculares utilizadas como padrão, EB- extrato bruto, F-III, fração precipitada com 50-80% de sulfato de amônio, 0,2M de NaCl na cromatografia de troca-iônica e Litochitinase1 eluída na cromatografia de exclusão molecular.
kDa
M EB F-III 0,2 M Litochitinase1
28,5 kDa
230
130
95
72
56
36
28

56
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
0,5 1,0 1,5 2,0 2,5 3,0 3,5 4,0
0,20
0,25
0,30
0,35
0,40
0,45
0,50
0,55
Data: Data1_meanModel: HyperblChi^2 = 0.00046P1 0.61317 ±0.02175P2 0.51232 ±0.0667
Abs
orbâ
ncia
(40
5 nm
)
[S]
5.4.2. Efeito da temperatura sobre a atividade enzimática
Neste estudo incubou-se a Litochitinase1 com o substrato sob diferentes
temperaturas. Após o ensaio foi observado que a 55°C ocorre maior atividade
quitinásica (Figura 12), embora na temperatura de 50°C ainda se observe quase
73% da atividade enzimática, mas esta decai consideravelmente em temperaturas
acima de 55°C.
Figura 11: Determinação do Km da Litochitinase1 50 µl da quitinase (4,05 µg) foram incubadas a 55 ºC por 2 horas com diferentes concentrações (0,25; 0,50; 0,75; 1; 1,25; 1,5; 2; 2,5; 3 e 3,5 mM) de p-nitrofenil-N-acetil-β-D-glucosaminideo. Transcorrido este tempo, as reações foram interrompidas e o p-nitrofenil lido a 405 nm. O experimento foi realizado em duplicatas e o resultado apresentado, no gráfico representa as médias com seus respectivos desvios padrões.
Km 0, 51232
V
eloc
idad
e (V
)

57
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
4 °C 25°C 37°C 45°C 50°C 55°C 60°C 70°C 80°C
0
20
40
60
80
100
120
Ativ
idad
e %
Temperatura 0C
A estabilidade térmica foi pesquisada pré-incubando a Litochitinase1, em
diferentes temperaturas. A Figura 13 mostra que nas temperaturas de 25, 37 e 45°C
a atividade enzimática permanece estável. No entanto observa-se que nas
temperaturas de 50 e 55°C ocorre uma potencialização da atividade enzimática e
que coincide com a temperatura ótima de atividade da enzima. Ainda observa-se
uma atividade considerável a uma temperatura de 60°C e que cai até a completa
perda de atividade enzimática a 70, 80 e 100°C.
Figura 12: Efeito da temperatura sobre a atividade da Litochitinase1
50 µl da quitinase (4,05 µg) foram incubados com 1,5 mM de p-nitrofenil-N-acetil-β-D-glucosaminideo em diferentes temperaturas durante 2 horas. Transcorrido o tempo, as reações foram interrompidas como descrito em métodos. O experimento foi realizado em duplicatas e o resultado apresentado no gráfico representa as médias com seus respectivos desvios padrões.

58
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
30 40 50 60 70 80 90 100 110
0
20
40
60
80
100
120
140
Ativ
idad
e re
sidu
al (
%)
Temperatura de pré-incubação (0C)
5.4.3. Efeito do pH sobre a atividade da Litochitinase1
O efeito do pH sobre a catálise do substrato foi avaliado. Observou-se que a
maior atividade enzimática sobre o substrato ocorreu na faixa entre os pH 5,0 e 6,0
(Figura 14), embora o gráfico mostre atividade ótima em pH 5,5, os erros
experimentais (não plotados) demonstram que estas diferenças não são
significativas. Em pH 4,0 e 8,0 a atividade da enzima cai consideravelmente.
Figura 13: Efeito da pré-incubação a diferentes temperaturas na atividade da Litochitinase1 50 µl da quitinase (4,05 µg) foi pré-incubada durante 30 minutos a 25, 37, 45, 50, 55, 60, 70, 80 e 100°C na ausência do substrato. Transcorrido o tempo, o p-nitrofenil-N-acetil-β-D-glucosaminídeo foi adicionado para uma concentração final de 1,5 mM e incubado a 55°C durante 2 h. Em seguida as reações foram interrompidas como descrito em métodos.

59
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
3 4 5 6 7 8 9
0
20
40
60
80
100
120
Tampão Glicina Tampão Acetato de sódio Tampão Fosfato de sódio Tampão Tris
Ativ
idad
e (%
)
pH
A estabilidade frente ao pH foi estimada pela determinação da atividade da
Litochitinase1, após pré-incubação por 30 minutos a diferentes pHs. De acordo com
o gráfico da figura 15, a quitinase não é estável em pHs muito ácidos e muito
básicos, perdendo completamente a sua atividade. A estabilidade da enzima é
atingida numa faixa de pH entre 4,0 – 5,5, embora a Litochitinase1 apresente um
pico de atividade no pH 7,0.
Figura 14: Efeito do pH sobre a atividade da Litochitinase1
Alíquotas de 200 µl (16,2 µg) da quitinase foram dialisadas durante 18 horas contra os tampões: glicina-HCl pH 3,0; 3,5 e 4,0; acetato de sódio pH 4,0; 5,0 e 5,5; fosfato de sódio pH 5,5; 6,0 e 7,0 e Tris-HCl pH 7,0;8,0 e 9,0. Todos os tampões estavam na concentração de 0,1 M. Após a diálise alíquotas de 50 µl da enzima foram incubadas com 1,5 mM de p-nitrofenil-N-acetil-β-D-glucosaminídeo durante 2 horas a 55°C. Transcorrido o tempo de ensaio as reações foram interrompidas e quantificadas a 405 nm em espectrofotômetro.

60
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
3 4 5 6 7 8 9-50-40-30-20-10
0102030405060708090
100110
Glicina-HCl Acetato de Sódio Fosfato de Sódio Tris-HCl
Ativ
idad
e R
esid
ual (
%)
pH
Alíquotas de 200 µl da quitinase foram dialisadas durante 18 horas contra os tampões: glicina-HCl pH 3,0 e 3,5; acetato de sódio pH 4,0 e 5,0; fosfato de sódio pH 5,5 e 6,0; e Tris-HCl pH 7,0, 8,0 e 9,0. Todos os tampões estavam na concentração de 0,1 M. Após a diálise alíquotas de 50 µl das enzimas foram pré-incubadas durante 30 minutos em suas temperaturas ótimas, na ausência do substrato. Transcorrido o tempo, o p-nitrofenil-N-acetil-β-D-glucosaminídeo foi adicionado na concentração de 10 mM e incubado na temperatura ótima da enzima durante 2 h. Em seguida as reações foram interrompidas como descrito na metodologia.
5.4.4. Efeito de sais e EDTA sobre a atividade enzimática
O efeito de sais e EDTA sobre a atividade da Litochitinase1 foi estimada pela
determinação da atividade, após a pré-incubação com vários compostos. De acordo
com a tabela 4, a Litochitinase1 apresentou sua atividade enzimática levemente
reduzida na presença do Na2HPO4 (5%). Por outro lado, os sais NaCl, KCl, NaH2PO4
e HgCl2, afetaram de forma expressiva a atividade enzimática, reduzindo-a em
aproximadamente: 28, 30, 40 e 98%, respectivamente (valores aproximados). O sal
MgCl2 foi o único que estimulou (~8%) levemente a atividade enzimática da
quitinase. CaCl2, Na2SO4, EDTA e o NaC6H5O7 não interferiram na atividade da
enzima.
Figura 15: Efeito da pré-incubação a diferentes pHs na atividade da Litochitinase1

61
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 4: Efeitos de íons e compostos sobre a atividade da Litochitinase1
Sais e EDTA Ativ. Residual (%)
MgCl2 107,6 ± 4,2
KCl 71,4 ± 3,4
CaCl2 100,09 ± 5,3
Na2S2O25H2O 99,09± 4,07
Na2SO4 99,59 ± 2,26
HgCl2 1,09 ± 0,137
NaH2PO4 57,635 ± 7,04
Na2HPO4 91,405 ± 3,68
NaC6H5O7 98,025 ± 1,58
NaCl 80,39 ± 9,07
EDTA 96,075 ± 0,40
A enzima foi pré-incubada durante 15 minutos com cada sal e/ou EDTA antes de ser iniciado o ensaio. Os resultados foram apresentados com as médias seguidas de seus respectivos desvios padrões.
5.5. Avaliação da atividade antimicrobiana
5.5.1. Utilizando a técnica de difusão em disco
Para se avaliar as atividades antimicrobianas foi, inicialmente, utilizada a
técnica de difusão em disco. Os resultados obtidos com a Litochitinase1 em estudo,
não produziram halos de inibição para os microrganismos testados, sugerindo
ausência de atividade antimicrobiana sobre estirpes de: S. aureus, C. tropicalis, e C.
albicans. Os discos utilizados como controle negativo (tampão acetato de sódio 100
mM, pH 5,0/água), também não demonstraram qualquer atividade inibitória sobre os
microorganismos (Dados não mostrados).

62
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
5.5.2. Utilizando a técnica de microdiluição em caldo para determinação da
Concentração Inibitória Mínima (CIM)
Na técnica de microdiluição em caldo os valores de CIM foram obtidos por
diluições sucessivas e determinados por leitura visual. Os resultados obtidos com a
Litochitinase1, não apresentaram atividade contra os microorganismos: S. aureus, C.
albicans e P. aeruginosa. Apresentando atividade antimicrobiana apenas para o
microorganismo E. coli variando de uma concentração de 800 a 500 µg/mL (Tabela
5).
Tabela 5: Avaliação da Concentração Inibitória Mínima (CIM) da Litochitinase1 do cefalotórax do camarão marinho Litopenaeus schmitti.
Cepas utilizadas Concentração
utilizada
Atividade
antimicrobiana
S. aureus - -
C. albicans - -
E. coli 800 - 500 mg/mL +
P. aeruginosa - -
(-) Não apresentou atividade antimicrobiana nas concentrações testadas; (+) Apresentou atividade antimicrobiana nas concentrações testadas.
5.6. Ensaios com os Aedes aegypti
5.6.1. Detecção de quitina no intestino médio de larvas de Aedes aegypti
A fim de detectar a presença de quitina no intestino médio das larvas de Aedes
aegypti foi utilizado o método químico qualitativo de Von Wisseling. Os resultados
confirmaram a presença de quitina no intestino médio de larvas destes insetos
(Tabela 6).

63
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 6: Detecção de quitina em intestinos médios de Aedes aegypti
Teste de Von Wisseling Coloração marrom (lugol)
Coloraçao violeta (H2SO4)
Quitina de lagosta + +
Intestinos de Aedes aegypti
+ +
Algodão ND ND
(ND) não detectado; (+) detecção. Controle positivo: quitina de lagosta; Controle negativo: algodão
5.6.2. Ensaio larvicida em Aedes aegypti
Estes experimentos foram feitos com o intuito de analisar o efeito larvicida da
Litochitinase1 sobre larvas do quarto instar de A. aegypti. Cinco larvas (L4) foram
colocadas em recipientes plásticos descartáveis contendo alíquotas do EB, F-III e a
Litochitinase1, em concentrações crescentes, e seus respectivos controles
negativos, com volume final do ensaio de 20 mL. Os ensaios foram realizados em
quadruplicatas e a taxa de mortalidade das larvas foi determinada após 24 e 48 h de
incubação a 28 ± 2 ° C com fotoperíodo de 12/12 h (claro-escuro). Os resultados
obtidos apontam que todas as amostras testadas apresentaram atividade larvicida
contra A. aegypti de forma dose dependente, onde a Litochitinase1 revelou-se mais
eficiente do que as outras amostras testadas, tendo como valores de EC50 para 24
horas de ensaio 0,71 mg/mL e para 48 horas de 0,49 mg/mL. Para as outras frações
como EB o valor de EC50 foi de 6,59 mg/mL e para F-III a EC50 para 24 e 48 horas
foi de 5,36 e 3,22 respectivamente. No ensaio com o EB, no tempo de 48 horas,
todas as larvas morreram. As análises estatísticas realizadas por meio dos testes
ANOVA e Kruskal-wallis, mostraram significância entre os resultados destes quando
se utilizou o EB, FIII e a Litochitinase1 em diferentes concentrações, nos tempos de
24 e 48 horas, com p< 0,05. As larvas do grupo controle apresentaram grande
mobilidade, cuja locomoção foi percebida por meio das contrações do corpo,
reagindo rapidamente a qualquer toque. Os valores da EC50 estão sumarizados na
tabela 7.

64
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 7: Concentração Efetiva do Extrato bruto, F-III e Litochitinase1 necessária para matar 50% (EC50) das larvas L4 de Aedes aegypti em 24 e 48 h.
Amostras EC/ Tempo (h)
EC50
/24h EC50
/48h
EB 6,59 mg/mL *
F-III 5,36 mg/mL 3,22 mg/mL
Litochitinase1 0,71 mg/mL 0,49 mg/mL
*Todas as larvas mortas. Onde EC: concentração letal de proteínas necessárias para matar 50% (EC) de larvas de A. aegypti L4 em 24 e 48 h.
5.6.3. Detecção de inibidores de proteases serínicas no EB, FIII e
Litochitinase1
Considerando que as enzimas digestivas de Aedes aegypti são proteases
serinicas e que inibidores destas enzimas poderiam comprometer o processo
digestório e o conseqüente desenvolvimento dessas larvas, foi verificada a presença
de inibidores de proteases. O EB, a F-III e a Litochitinase1, oriundas do cefalotórax
do camarão marinho Litopenaeus schmitti, foram submetidas à avaliação de
atividade inibitória sobre tripsina e quimotripsina. Os resultados confirmaram a
ausência de inibidores de proteinases serinicas em todas as amostras analisadas
(Tabela 8).

65
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 8: Inibição de Proteases Serínicas
Enzimas Amostras Inibição
Tripsina
EB ND
FIII ND
Litochitinase1 ND
Quimotripsina
EB ND
FIII ND
Litochitinase1 ND
(ND) não detectado
5.6.4. Atividade hemaglutinante
Com o objetivo de avaliar a presença de lectinas nas amostras do cefalotórax
do camarão aliquotas do EB, F-III e Litochitinase1, foram submetidas a ensaios de
hemaglutinações com eritrócitos do sistema ABO tratados ou não com enzimas,
visando elucidar qual tipo sanguíneo apresentaria maior título de hemaglutinação e
qual tratamento enzimático se apresentaria mais eficiente na exposição de
carboidratos na superfície das células. Os resultados obtidos não apontaram a
presença de atividade hemaglutinante em nenhuma das frações testadas tanto com
eritrócitos sem tratamento nem com os tratados. No entanto a F-III apresentou um
leve potencial hemolítico em todos os tipos sanguíneos não tratados ou tratados
(Tabela 9).

66
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Tabela 9: Atividade hemaglutinante no extrato bruto, F-III e Litochitinase1
com eritrócitos nativos e tratados enzimaticamente
SISTEMA ABO FRAÇÕES PROTEÍCAS (UH)*
Sem tratamento EB F-III Litochitinase1
A ND hemólise ND
B ND hemólise ND
O ND hemólise ND
Tratamento com tripsina
A ND hemólise ND
B ND hemólise ND
O ND hemólise ND
Tratamento com papaína
A ND hemólise ND
B ND hemólise ND
O ND hemólise ND
*UH: Unidade de hemaglutinação; (ND) não- detectado
5.6.5. Determinação da temperatura ótima das atividades proteolíticas
presentes no homogenato intestinal das larvas de A. aegypti
Para analisar a temperatura ótima de atividade das enzimas proteolíticas no
homogenato intestinal das larvas de A.aegypti, as mesmas foram incubadas com o
substrato azocaseína 1,5% em diferentes temperaturas. As enzimas presentes nos
homogenatos intestinais foram ativas na faixa de temperatura de 10 a 60°C (Figura
16). Sendo assim a temperatura do ensaio realizado para analisar se as enzimas
digestivas dos insetos inibiriam a atividade da Litochitinase1 poderia ser feita na
temperatura ótima desta que é de 55°C.
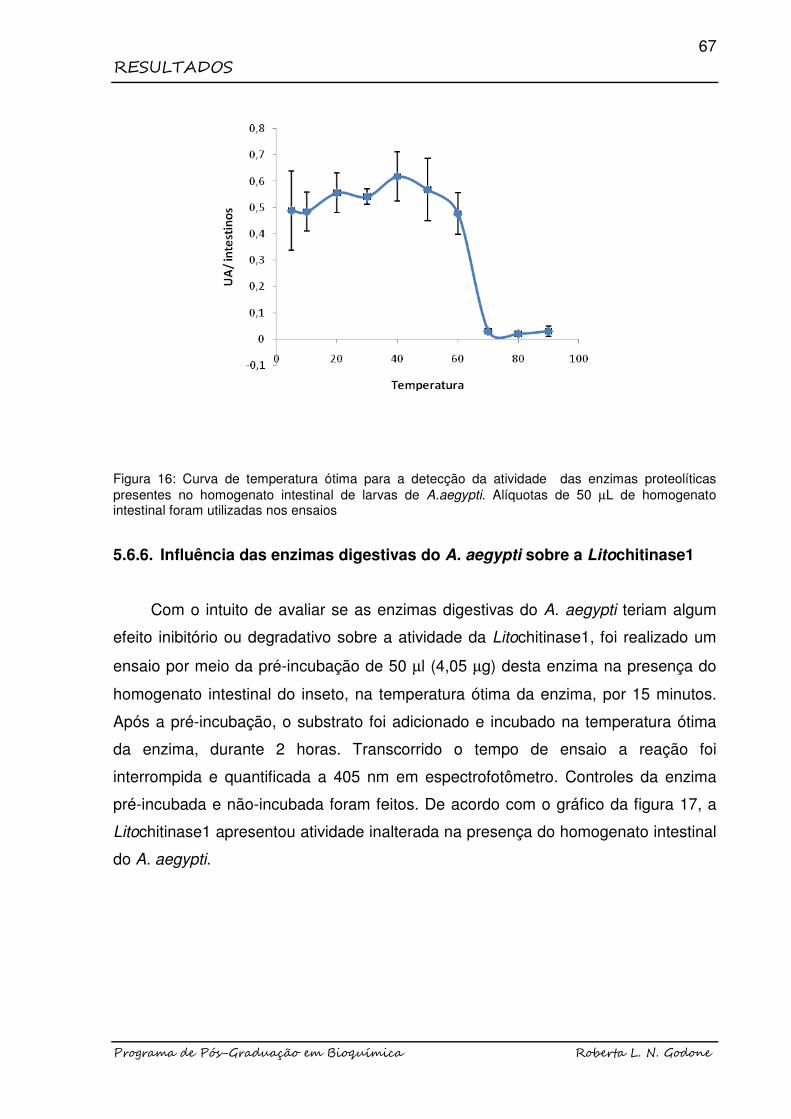
67
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Figura 16: Curva de temperatura ótima para a detecção da atividade das enzimas proteolíticas presentes no homogenato intestinal de larvas de A.aegypti. Alíquotas de 50 µL de homogenato intestinal foram utilizadas nos ensaios
5.6.6. Influência das enzimas digestivas do A. aegypti sobre a Litochitinase1
Com o intuito de avaliar se as enzimas digestivas do A. aegypti teriam algum
efeito inibitório ou degradativo sobre a atividade da Litochitinase1, foi realizado um
ensaio por meio da pré-incubação de 50 µl (4,05 µg) desta enzima na presença do
homogenato intestinal do inseto, na temperatura ótima da enzima, por 15 minutos.
Após a pré-incubação, o substrato foi adicionado e incubado na temperatura ótima
da enzima, durante 2 horas. Transcorrido o tempo de ensaio a reação foi
interrompida e quantificada a 405 nm em espectrofotômetro. Controles da enzima
pré-incubada e não-incubada foram feitos. De acordo com o gráfico da figura 17, a
Litochitinase1 apresentou atividade inalterada na presença do homogenato intestinal
do A. aegypti.

68
RESULTADOS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
Figura 17: Efeito da pré-incubação com as enzimas digestivas de A.aegypti na atividade da Litochitinase1. 50 µl da Litochitinase1 foram pré-incubados durante 15 minutos com o homogenato intestinal, na ausência do substrato. Transcorrido o tempo, o p-nitrofenil-N-acetil-β-D-glucosaminídeo foi adicionado na concentração de 10 mM e incubado na temperatura ótima da enzima durante 2 h. Em seguida as reações foram interrompidas como descrito na metodologia. Onde: CNI- Controle da Litochitinase1 não pré-incubado; CI- Controle da Litochitinase1 pré-incubado sem enzimas do homogenato e EHI- Litochitinase1 pré-incubada com enzimas do homogenato intestinal.

70
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
6. DISCUSSÃO
Em 2003, a produção mundial de camarão foi de 4.630.000 toneladas sendo
que 1.630.000 t (35,21% do total) foram produzidas por cultivo (REVISTA
PANORAMA DA AQÜICULTURA, 2004). No Brasil, a pesca extrativa marinha
produziu 35.451,5 toneladas de camarão em 2007 (ESTATÍSTICA DA PESCA,
2007). A carcinicultura, após um pico de produção de 90.190 t em 2003, manteve
uma produção de 65.000 t anuais de camarão no período de 2005 a 2007 (ROCHA,
2008). O processamento de cerca de 100.000 t anuais de camarão no Brasil gera
uma enorme quantidade de rejeitos, principalmente durante a etapa de retirada da
casca e do cefalotórax (GOMES et al., 2010).
Algumas das propostas para eliminar este resíduo da indústria pesqueira
incluem a elaboração de farinha de casca de camarão (para a utilização em
alimentos) e o aproveitamento da quitina para diversos usos, como na produção de
catalisadores, em complementos alimentares para redução do colesterol e controle
do peso (SHAHIDI et al., 1999; KIM; RAJAPAKSE, 2005) ou como agente
imobilizador de enzimas e microorganismos (PALLA et al., 2011). A transformação
dos rejeitos da indústria de processamento do camarão em quitosanas ou
quitinooligossacarídeos com massas molares médias e níveis de desacetilação
controlados seria uma alternativa viável, gerando produtos com alto valor agregado
para usos biotecnológicos (GOMES et al., 2010).
Dentre as principais tecnologias que surgiram desde a década de 70, a
biotecnologia tem atraído maior atenção por ter se revelado capaz de gerar enorme
riqueza e influência em cada setor da economia com destaque para a saúde,
produção e processamento de alimentos, agricultura, proteção ambiental e produção
de materiais (GAVRILESCU; CHISTI, 2005).
A indústria de enzimas é resultado de um desenvolvimento rápido, alcançado
nas últimas quatro décadas, graças à evolução da biotecnologia moderna. O
desenvolvimento dos processos de purificação e fermentação, ocorrido durante a
última metade do século passado, permitiu a produção de enzimas mais puras, bem
caracterizadas e em larga escala (KIRK, 2002). O mercado mundial de enzimas
movimentou em 2007 cerca de 2,3 bilhões de dólares e espera-se para 2012 um
movimento superior a 2,7 bilhões, com um aumento anual de 4% (THAKORE, 2008).

71
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
As enzimas quitinolíticas apresentam inúmeras aplicações biotecnológicas no
ramo da indústria e agricultura. As quitinases podem ser utilizadas no controle de
fungos patógenos de plantas e insetos, como ainda na produção de
quitinooligossacarídeos biologicamente ativos (PATIL, et al,2000).
O objetivo principal deste trabalho foi estudar uma quitinase purificada e
extraída do cefalotórax do camarão marinho Litopenaeus schmitti, porção esta
normalmente descartada sem qualquer tipo de aproveitamento tecnológico
(FOGAÇA; LEGAT, 2009) e avaliar sua aplicabilidade biotecnológica como agente
microbicida e larvicida.
Os resultados iniciais utilizando os p-nitrofrofenís derivados de açúcar como
substrato mostraram a presença de quase todas as atividades enzimáticas
pesquisadas no EB, F-I, F-II e F-III. Estes não foram tão surpreendentes, pois
extratos enzimáticos nos primeiros passos de purificação geralmente são ricos em
diversas atividades glicosidásicas e sulfatásicas. Fato este já observado nos extratos
dos celenterados Palythae coribaerum (SOUZA, 2003); Palythoa voriabitis
(FERREIRA, 2003); no molusco Aplysia cervina (MATTA; ABREU, 2005);
Echinometra lucunter (LIMA, 2006) e no microcrustáceo Artemia franciscana
(NASCIMENTO, 2008). As possíveis funções dessas enzimas seriam a degradação
de carboidratos complexos, como polissacarídeos e glicosaminoglicanos (KRESSE;
GROSSI, 1987; STAM et al., 2005). De todas as frações testadas com os substratos
sintéticos derivados de açúcar a F-III apresentou maior atividade específica,
ressaltando que nesta fração a atividade quitinásica se sobressai das demais.
Resultados semelhantes foram encontrados no molusco Chiton sp. (VIEIRA, 2002),
Strombus goliath (ARAÚJO, 2002), Paliythoa caribaeorum (SOUZA, 2003) e no
Echinoderma marinho Echinometra lucunter (LIMA, 2006) onde a enzima que
apresentou atividade quitinolítica também mostrou maior atividade específica nas
frações protéicas extraídas destes animais.
Para a obtenção de quitinases purificadas, diferentes métodos são utilizados
tais como, precipitação com sulfato de amônio (IKE et al., 2006), cromatografia de
interação hidrofóbica (DUO-CHAN, 2005), cromatografia de troca iônica (IKE et al.,
2006), cromatografia de afinidade a quitina (KIM et al., 2007) dentre outras. Para
uma purificação total geralmente se faz necessário a utilização de mais de um dos
métodos acima. Para a purificação da quitinase extraída do cefalotórax do camarão
marinho Litopenaeus Schmitti combinou-se cromatografia de troca iônica e exclusão

72
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
molecular S-200, onde foi obtida a enzima purificada 987,32 vezes com um
rendimento de 2,62 %. Este baixo rendimento pode ter sido causado pela perda de
atividade enzimática ao longo do processo de purificação. Acredita-se que, por
serem glicoproteínas, estas enzimas possam perder sua porção carboidrática ao
longo do processo de purificação, o que compromete sua atividade (PASTORE,
2003). A enzima assim purificada recebeu o nome de Litochitinase1.
Resultados de purificações elevadas foram encontrados na literatura com N-
acetilhexosaminidases como é o caso de SOUZA (2003) e GOMES-JÚNIOR (2010)
que obtiveram alto grau de purificação, 4826 vezes e 2232 vezes, respectivamente.
Para avaliar o grau de pureza e determinar a massa molecular aproximada da
Litochitinase1 uma eletroforese em gel de poliacrilamida em condições
desnaturantes revelou que esta possui uma banda protéica com o peso de 28,5 kDa.
Em geral a literatura traz exemplos de quitinases com massas moleculares variando
em uma ampla faixa que vai de 40 a 240 kDa (PETERS et al., 1998; CIFALI; DIAS
FILHO, 1999; AMUTHA, 1999), porém recentemente foi descrito por GOMES et al .
2010 duas quitinases com peso molecular de 24 e 30 kDa extraída de Vitis vinífera e
ainda L. KOPPARAPU et al. (2011) descreveram uma quitinase extraída de Punica
granatum cujo peso molecular é de 29 kDa, peso este bem próximo ao encontrado
no gel de SDS–PAGE apresentado neste trabalho.
O valor de Km aparente de 0,51 mM encontrado demonstra que essa enzima
possui grande afinidade por seu substrato e que sua hidrólise obedeceu à cinética
de Michaelis-Menten. O valor de Km obtido não diferiu significativamente de alguns
valores encontrados em outros trabalhos, como no caso da quitinase de Pennahia
argentatus onde foi observado um Km de 0,52 mM (IKEDA et al., 2009) e da
quitinase de Hexagrammos otakii (MATSUMIYA et al., 2006) que apresentou um Km
de 0,494 mM. Entretanto, muitos estudos encontraram valores de Km mais elevados
como no caso da quitinase de ovos de Halocynthia roretzi (MATSUURA et al., 1993)
que apresentou Km de 1,2 mM, e quitinase do vírus Autographa californica (DI MARO
et al., 2010) onde se observou um Km de 3,5 mM.
A temperatura ótima observada para a Litochitinase1, após duas horas de
ensaio a diferentes temperaturas, foi de 55°C. Essa atividade decai em temperaturas
abaixo e acima deste ponto, podendo indicar desnaturação proteíca. Estudos
cinéticos realizados com exoquitinases de diversas outras fontes demonstraram para
esse tipo de glicosidase temperaturas estáveis entre 37 e 50°C (TSUJIBO et al.,

73
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
1995; MATSUO et al., 1999; KAMEI et al., 1999; ARAÚJO, 2002; VIEIRA, 2002).
Recentemente KOPPARAPU et al. (2011) relataram uma temperatura ótima de 70°C
para a quitinase extraída da romã (Punica granatum). Outras quitinases purificadas
de uva, tamarindo, abacaxi, látex de Ficus microcarpa e do camarão Penaeus
monodon apresentaram temperaturas ótimas de 40, 45, 60 e 55°C, respectivamente
(ANO et al., 2003; TAIRA et al., 2005a, 2005b; RAO; GOWDA, 2008;
PROESPRAIWONG et al., 2010). Temperaturas estas, muito próximas a
temperatura ótima da quitinase estudada neste trabalho.
A pré-incubação da enzima por 30 minutos a várias temperaturas revelou que
a Litochitinase1 é estável de 25 a 60° C. Entre 50 e 55° C sofre uma leve ativação
aumentando em aproximadamente 20% sua atividade de catálise do substrato. Isto
indica que a Litochitinase1, ao ser submetida a estas temperaturas, de alguma
maneira elas afetam positivamente sua atuação sobre o substrato. Temperaturas
acima de 60ºC as inativaram rapidamente. Recentemente, PATEL et al., (2010),
isolaram uma quitinase do látex de Ipomoea carnea que apresentou estabilidade até
a temperatura de 80ºC. Uma estabilidade relativa até a temperatura de 40ºC foi
observada em uma quitinase recombinante, purificada do camarão tigre negro
(Penaeus monodon) (PROESPRAIWONG et al., 2010).
O pH ótimo encontrado para a hidrólise do p-nitrofenil-N-acetil-β-D-
glucosaminídeo foi entre os pH 5,0 e 6,0. Em pH 4,0 e 8,0 a atividade da enzima cai
consideravelmente. Uma quitinase purificada do vírus Autographa californica (DI
MARO et al., 2010) mostrou estabilidade em pH que variava de 5,0 a 7,0, resultado
este semelhante ao encontrado para a enzima aqui estudada. Da mesma forma
CHEN et al., (2011), purificou uma enzima presente no látex do mamão que
apresentava atividade ótima no pH 5,0 e PATEL et al., (2010), purificaram uma
quitinase do látex da Ipomoea carnea que apresentou atividade máxima no pH 5,5.
A Litochitinase1 não é estável em pHs muito ácidos e muito básicos, perdendo
completamente as suas atividades. Isso possivelmente é devido ao fato de que
nesses pHs a enzima perde sua conformação nativa, devido a mudanças no estado
de ionização das cadeias laterais dos aminoácidos localizados no sitio ativo desta,
alterando assim a distribuição de cargas, interferindo de sobremaneira na estrutura
da proteína e na atividade enzimática. A estabilidade da enzima é atingida numa
faixa de pH entre 4,0 – 5,5, embora apresente um pico de atividade no pH 7,0.

74
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
O efeito de vários sais e EDTA sobre a hidrólise do p-nitrofenil-N-acetil-β-D-
glucosaminídeo pela Litochitinase1 foi estudado. A enzima apresentou sua atividade
levemente reduzida na presença do Na2HPO4 (5%). Por outro lado, os sais KCl,
HgCl2, NaH2PO4, e NaCl afetaram de forma expressiva a atividade enzimática,
reduzindo sua atividade em 28, 98, 40 e 30%, respectivamente. O MgCl2 estimulou
só levemente (~8%) a atividade da Litochitinase1, porém concentrações mais
elevadas do sal, possam estimular ainda mais a enzima. CaCl2, Na2SO4, EDTA e o
NaC6H5O7 não interferiram na atividade da enzima. O efeito de sais (íons) e
compostos é amplamente variável, dependendo da enzima em estudo. O efeito
inibitório observado com a adição de HgCl2 (íon Hg2+ ) é geralmente observado
em outras glicosidases (CHEN et al., 1994; WEI et al., 1996; NAKANO et al., 1998;
RIOU et al., 1998; HAYASHI et al., 1999; LUCAS et al., 2000; YAZDI et al., 2003;
ZANOELO et al., 2004; LI et al., 2005). O íon Hg2+ é reconhecido agente oxidante de
grupos sulfidril (-SH), e a inibição provocada por este íon pode indicar o
envolvimento de importantes grupos sulfidril no sítio catalítico da enzima (PAINBENI
et al., 1992; GUEGUEN et al., 1995a, 1997b; WEI et al., 1996; NAKANO et al., 1998;
OH et al., 1999). No entanto, a inibição observada na presença de Hg2+ pode não
estar apenas relacionada à ligação deste íon com grupos sulfidril, mas também à
interação deste íon com resíduos de triptofano e/ou grupamentos carboxil em
aminoácidos da enzima (LUSTERIO et al., 1992).
O EDTA, reagente com a capacidade de quelar íons metálicos, é empregado
em diversos testes bioquímicos visando, entre outros, diminuir a ação de
metaloproteinases em extratos brutos. Entretanto, o EDTA pode tornar-se
problemático caso as enzimas dependam de íons como o Ca+2 (MATEO; DI
STEFANO, 1997). Nas concentrações utilizadas, este reagente não apresentou
efeito sobre a atividade da Litochitinase1, desta forma estes estudos sugerem que a
não interferência do EDTA indica que a catálise não é dependente de íons Ca+2,
similarmente a outros estudos, sugerindo que a enzima não requer co-fatores
metálicos (DRIDER et al., 1993; GUEGUEN et al., 1995a; NAKANO et al., 1998;
RIOU et al., 1998; YAN et al., 1998; KIMURA et al., 1999; OH et al., 1999;
ZANOELO et al., 2004; LI et al., 2005; FERREIRA, 2010).
A literatura a respeito da ação antifúngica e antibacteriana de produtos
naturais é escassa, sendo que a maioria dos estudos verifica seu potencial como
antiinflamatórios e produtos dermatológicos. Produtos naturais como substâncias

75
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
antimicóticas e bactericidas derivados de origem protéica não são muito estudados.
A maioria das substâncias empregadas como agentes microbicidas são de origem
vegetal, e já foi demonstrado, por exemplo, o efeito antifúngico de plantas como
estragão e tomilho (DUKE, 1985; BRANTNER et al., 1996), de substancias como o
própolis (NOSTRO et al., 2006; PONTIN et al., 2008), ação antibacteriana do
Anacardiun occidentale (cajueiro) que foi estudado mostrando-se efetivo para
bactérias gram-positivas e gram-negativas (KUDI, 1999) e a atividade
antimicrobiana do extrato bruto da folha de Baccharis dracunculifolia (DA
SILVA FILHO et al., 2008).
No presente estudo, testes microbiológicos utilizando a quitinase purificada
foram realizados e os resultados obtidos pela técnica de difusão em disco, para as
cepas testadas de S. aureus, as leveduras C. tropicalis e C. albicans não
apresentaram halos de inibição, logo, a Litochitinase1 não foi capaz de inibir o
crescimento desses microorganismos, isto pode ser atribuído a dificuldades na
padronização e comparação de técnicas de difusão em disco, devido à interferência
das características de algumas amostras na capacidade de difusão (SILVEIRA et
al., 2009), pois em se tratando de avaliação de produtos naturais RIBEIRO;
SOARES (2000) afirmam que as características de solubilidade do material em teste
interferem na difusão do mesmo no meio de cultura.
Diferente dos resultados obtidos neste estudo, já é relatado na literatura à
ação de quitinases extraídas do amedoim e do microorganismo Trichoderma
harzianum contra bactérias gram-positivas como S. aureus (WANG et al., 2007; LIN
et al., 2009). Várias enzimas têm sido estudadas como agentes antifúngicos em
potencial. A ação fungicida deve-se principalmente à hidrólise dos componentes da
parede celular (FLEURI; SATO, 2007). Porém a maioria dos estudos relacionados à
ação de quitinases como agentes antifúngicos é aplicada em fungos fitopatogênicos
como revela estudos desenvolvidos por YE; NG (2005), WANG (2007), FLEURI;
SATO (2008) e LAM; NG (2010).
Para os resultados de microdiluição em caldo dentre as quatro cepas testadas,
a Litochitinase1 não apresentou atividade antimicrobiana para S. aureus, C. albicans
e P. aeruginosa. Estudos realizados com quitinases de outras fontes como a da
planta Brassica Junceae se mostraram ativas contra estes microorganismos (GUAN
et al., 2008). No entanto a Litochitinase1 apresentou-se efetiva contra o
microorganismo E. coli, inibindo-o partindo de uma concentração inicial de 800

76
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
µg/mL, concentração bem inferior a de outros trabalhos que utilizaram a mesma
cepa e o mesmo teste de microdiluição, como mostrado no estudo de
CHOWDHURY (2008) onde uma proteína extraída da Solanum villosum consegue
efeito bactericida contra E. coli partindo de uma concentração de 1800 µg/mL.
Escherichia coli é a espécie predominante entre os diversos microrganismos
anaeróbios facultativos e comumente encontrada na microbiota normal no trato
gastrointestinal humano (GILL et al., 2006) e outros vertebrados, com o qual
geralmente estabelece associações comensais. Esse microrganismo pertence à
família Enterobacteriaceae e entre suas principais características destacam-se:
bacilos gram-negativos, não esporulados capazes de fermentar glicose com
produção de ácido e gás (FRANCO; LANDGRAF, 2003).
Seres humanos saudáveis tipicamente, têm mais de um bilhão de células de
E. coli no intestino. Estima-se que metade das células vivas de E. coli estão fora do
seu hospedeiro, em seu habitat secundário (SAVAGEAU, 1983). Além destes
habitats, algumas estirpes têm o potencial de causar um amplo espectro de
doenças intestinais e extra-intestinais, como infecção do trato urinário, septicemia,
meningite e pneumonia em humanos e animais (DONNENBERG, 2002) e como
também aumento da inflamação da mucosa gástrica em pacientes com doença de
Crohn (BARNICH et al., 2010). Com a sua vasta gama de patologias, a E. coli é uma
das principais causas de morbidade e mortalidade em todo o mundo, causando a
cada ano mais de dois milhões de mortes por diarréia infantil (BISSON et al., 2002;
RODRÍGUEZ-BAÑO et al., 2006) e infecções extra-intestinais (principalmente
septicemia derivados de infecção urinária) e também é responsável por
aproximadamente 150 milhões de casos de cistite simples (DU et al., 2002).
Durante a última década, tem havido um marcado aumento nas infecções por
E. coli resistentes a antibióticos, tais como: Fluoroquinolona. Esta crescente
resistência limita as opções de tratamento e podem afetar o prognóstico da doença
(LAUTENBACH et al., 2001), como também ocorrer o uso inadequado de
antimicrobianos ocasionando atrasos na terapia correta. O aumento da resistencia
bacteriana, justifica a necessidade urgente por novos agentes antibióticos que atuem
por mecanismos de ação diferentes dos fármacos em uso (PAYNE et al., 2007;
COATES; HU, 2007).

77
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
As novas estratégias da pesquisa em produtos naturais, envolvendo a busca
de substâncias pouco exploradas para o acesso da descoberta de novos antibióticos
extremamente importantes num cenário de rápido desenvolvimento de resistência
pelas bactérias aos agentes terapêuticos disponíveis, indica que a Litochitinase1
pode vir a ser utilizada como uma possível alternativa no combate das doenças
causadas pela E.coli.
A dengue é a mais importante e global arbovirose da atualidade, causando de
50-100 milhões de casos/ano em mais de 100 países (REGIS et al., 2008) e
submetendo 2,5 bilhões de pessoas ao risco de se infectarem, principalmente em
países tropicais e subtropicais onde as condições climáticas (temperatura e
umidade) são favoráveis a proliferação do mosquito transmissor, o Aedes aegypti
(POLANCZYK et al., 2003, SIMAS et al., 2004). Como não há vacina disponível para
prevenção, o controle da dengue tem sido limitada ao combate do vetor e centrado
na aplicação de inseticidas químicos (CARVALHO et al., 2004; LIMA et al., 2005a;
MACORIS et al., 2007; SILVA; MENDES, 2007), especialmente os organofosforados
e piretróides para os adultos e organofosforados para as larvas (LIMA et al., 2006).
Todavia, relatos de populações de insetos resistentes (RODRIGUES et al, 2001) a
esses inseticidas apontam para a necessidade de pesquisas que busquem novas
alternativas de controle, isentas de risco para a saúde humana e ao meio ambiente.
Não obstante os trabalhos realizados por COSTA et al. (2005); SILVA et al.
(2007); LUZ et al. (2008); MEDEIROS, (2007) e FERREIRA et al. (2009) utilizando
como fontes produtos naturais contra as larvas de Aedes nenhum utilizavam
quitinases. Por isso resolveu-se verificar o potencial larvicida da Litochitinase1.
Os ensaios in vivo apontam que todas as amostras testadas apresentaram
atividade contra larvas do quarto estádio de A. aegypti de forma dose dependente.
Essa atividade aumenta à medida que se utiliza material com maior grau de
purificação (EB: EC50 6,59 mg/mL, F-III: EC50 5,36, Litochitinase1: EC500,71 mg/mL)
sugerindo que a molécula responsável pela morte das larvas tenha sido a
Litochitinase1, e que, supostamente esta poderia estar atuando na quitina presente
no exoesqueleto e/ou na membrana peritrófica do inseto. A membrana peritrófica é
um revestido do intestino médio constituída por proteínas e quitina, cuja presença foi
confirmada pelo teste de Von Wisselingh, e vem sendo bastante estudada como alvo
para a ação de bioinseticidas (HARPER, HOPKINS; CZAPLA, 2001). A favor dessa
segunda hipótese verificamos a não degradação da Litochitinase1 pelas enzimas

78
DISCUSSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
digestivas de A. aegypti e, adicionalmente, a ausência de outras moléculas bioativas
que poderiam também interferir no processo digestório das larvas, como inibidores
de proteases serinicas e lectinas nas frações testadas, indicando mais uma vez que
a morte das larvas está ocorrendo por ação da Litochitinase1.
As enzimas digestivas de A. aegypti são proteases serinicas tipo tripsina e
quimotripsina (KUNZ, 1978). Ensaios com outros insetos mostram que a inibição
destas enzimas compromete o processo digestório e o consequente
desenvolvimento e/ou morte das larvas, por privação de nutrientes. A maioria dos
inibidores de proteases tem sido obtidos de fontes como órgãos de reservas de
plantas (XAVIER-FILHO, 1992; ARAÚJO et al., 2004), todavia trabalhos recentes na
literatura descrevem inibidores de proteinases serínicas em invertebrados marinhos
como no camarão Fenneropenaeus chinensis (KONG et al., 2009) e Penaeus
momodon (VISETNAN et al., 2009; DONPUDSA et al., 2010). .
Lectinas tem sido encontradas em quase todos os organismos, sendo
amplamente distribuídas entre vegetais, vírus, bactérias, mamíferos e vários grupos
de invetebrados (GERLACH et al., 2005) como por exemplo: nos camarões Penaeus
paulensis (MARQUES; BARRACO, 2000), Penaeus monodon (LUO et al., 2005) e
Penaeus japonicus (YANG et al., 2007). As lectinas ligantes de N-
acetilglucosamina, oriundas das plantas Moringa Oleifera (COELHO et al., 2009) e
Myracroduom urundeuva (SÁ et al., 2009) mostraram atividade larvicida contra
Aedes aegypti.
Mesmo diante da necessidade de estudos complementares que esclareçam o
mecanismo pelo qual a quitinase provoca a morte de larvas de A. aegypti, e
comprovem a ausência de efeitos deletérios ambientais e sanitários, percebe-se o
potencial dessas moléculas para serem utilizadas como ferramentas auxiliares no
controle do vetor transmissor da dengue.

79
CONCLUSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
7. CONCLUSÃO
• Pôde-se observar a presença de atividades glicosidásicas e sulfatásicas nos
extratos enzimáticos obtidos do cefalotórax do camarão marinho Litopenaeus
schmitti;
• A fração F-III (50-80%) apresentou maior atividade quitinolítica;
• A Litochitinase1 foi purificada na ordem de 987,32 vezes e com recuperação de
2,62% apresentando massa molecular de aproximadamente 28,5 kDa;
• A temperatura ótima para a catálise do substrato sintético pela Litochitinase1 é
de 55°C e elevada estabilidade térmica permanecendo com 100% de sua
atividade quando pré-incubada nas temperaturas de 25, 37 e 45°C e
apresentando aumento desta nas temperaturas de 50 e 55°C;
• A atividade ótima para atuação da Litochitinase1 ocorre em uma faixa de pH,
entre 5,0 e 6,0, com faixa de estabilidade nos pHs 4,0 a 5,5;
• O HgCl2 na concentração de 10mM inibe significativamente a Litochitinase1
enquanto que o MgCl2 na mesma molaridade potencializa sua atividade;
• A Litochitinase1 apresentou constante de Michaelis-Menten (Km) de 0,51 mM,
indicando que a enzima possui alta afinidade pelo substrato quando comparadas
com dados da literatura;
• A Litochitinase1 demonstrou potencial microbicida contra a bactéria gram-
negativa E. coli demonstrado pela inibição de seu crescimento, avaliado pelo
método de microdiluição em caldo, podendo ser utilizada no combate de
infecções causadas por este patógeno ;
• A Litochitinase1 apresentou atividade larvicida contra larvas de Aedes aegypti de
maneira dose dependente e, de acordo com os outros resultados obtidos como:
ausência de proteínas contaminantes como inibidores de proteinases serínicas e
lectinas, esta atividade se confirma indicando-a para ser usada como alternativa
de controle eficaz na redução da infestação pelo mosquito vetor.

80
CONCLUSÃO
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
• No Brasil, a carcinicultura marinha é uma das atividades agroindustriais mais
atrativas economicamente, concentrando 93% de sua produção no Nordeste. No
entanto, durante o processamento do camarão, cerca de 50% do peso do animal
resulta em subproduto (cefalotórax, segmentos abdominais e carapaças),
descartado sem qualquer tipo de aproveitamento tecnológico (FOGAÇA; LEGAT,
2009). Diante de tal fato, a utilização dos cefalotórax para extração de enzimas
com enormes aplicabilidades, constitui-se numa via alternativa de
aproveitamento destes resíduos.

81
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
REFERENCIAS
ABRAM, A. P.; HIGUERA, I. Em Quitina y quitosano: obtencion, caracterizacion y aplicaciones;. ed.; Programa Cyted 2004, - Pontificia Universidad Catolica Del Peru/Fondo Editorial: Lima, 2004, cap 1.
ALLARDYCE, B. J; LINTON, S. M; SABOROWSKI, REINHARD. The last piece in the cellulase puzzle: the characterisation of β-glucosidase from the herbivorous gecarcinid land crab Gecarcoidea natalis. The Journal of Experimental Biology. v, 213. p, 2950-2957, 2010.
AMUTHA, B.; KHIRE, J. M.; KHAN, M. I. Active site characterization of the exo-N acetyl-β-D-glucosaminidase from thermotolerant Bacillus sp. NCIM 5120: involvement of tryptophan, histidine and carboxylate residues in catalytic activity. Biochimica et Biophysica Acta BBA-General Subjects, v. 1427, p.121-132, 1999.
ARAÚJO, C. L..; BEZERRA, I. W.L.; DANTAS, I. W. L; LIMA, T. V. S; OLIVEIRA, A. S.; MIRANDA, M. R. A; LEITE, E. L.; SALES, M. P. Biological activity of proteins from pulps of tropical fruits. Food Chemistry. V. 85, p. 107-110, 2004.
ARAÚJO, C. M. D. B. Purificação e caracterização parcial de uma ββββ-N acetilglucosaminidase extraída do molusco Strombus goliath. Dissertação de Mestrado em bioquímica – Universidade Federal do Rio Grande do Norte, Rio Grande do Norte- RN, 2002.
ARO, N.; PAKULA, T.; PENTTILA, M. Transcriptional regulation of plant cell wall degradation by filamentous fungi. FEMS Microbiology Reviews, Oxford, v. 29, p. 719-739, 2005.
ASSIS, A. S.; STAMFORD, T. C. M.; STAMFORD, T. L. M. Bioconversão de Resíduos de camarão Litopenaeus vannamei (Bonner, 1931) para produção de Biofilme de Quitosana. Revista Iberoamericana de Polímeros, v. 9, p. 480-499, 2008.
BARBOZA-CORONA. J. E; MAZZOCCO, E. N; VELAZQUEZ-ROBLEDO, R; HERNANDEZ, RUBE´N SALCEDO-; BAUTISTA, M; JIMENEZ, B; IBARRA, J. E. Cloning, Sequencing, and Expression of the Chitinase Gene chiA74from Bacillus thuringiensis. APPLIED ANDENVIRONMENTALMICROBIOLOGY. p. 1023–1029, 2003. BARNICH, N; DENIZOT, J; DARFEUILLE-MICHAUD, A. E. coli-mediated gut inflammation in genetically predisposed Crohn’s disease Patients. Pathologie Biologie. 2010. BAUER, A.W., KIRB, W.M.M., SHERRIS, J.C. Antibiotics susceptibility testing by a standardized single disk method. American Journal of Pathology, v.19, p.493 496, 1966.
BELANCIC, A.; GUNATA, Z.; VALLIER, M. J.; AGOSIN E. β-glucosidase from the native yeast Debaryomyces vanrijiae: purification, characterization, and its effect on

82
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
monoterpene content of a Muscat grape juice. Journal of Agricultural and Food Chemistry, Washington, v. 51, p. 1453-1459, 2003.
BHATIA, Y., MISHRA, S., BISARIA, V. S. Microbial β-glucosidases: Cloning, properties and applications. Critical Reviews in Biotechnology, Philadelphia; v. 22, p. 375-407, 2002.
BISHOP, J. G.; DEAN; A. M.; MITCHELL-OLDS, T. Rapid evolution in plant chitinases: Molecular targets of selection in plant-pathogen coevolution. PNAS. V. 97, p. 5322–5327, 2000. BISSON, G; FISHMAN, NO; PATEL, JB. Extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella species: risk factors for colonization and impact of antimicrobial formulary interventions on colonization prevalence. Infect Control Hosp Epidemiol. v. 23, p. 254-60, 2002. BOOT, R.G; BLOMMAART, E. F. C.; SWART, E.;GHAUHARALI-VAN DER VLUGT, K; BIJL, N.; MOE, C.; PLACE, A.; AERTS, J. M. F. G. Identification of a Novel Acidic Mammalian Chitinase Distinct from Chitotriosidase. The Journal of Biological Chemisty. V. 276, p. 6770-6778, 2001. BRANTNER, A; MALES, Z; PEPELJNJAK, S; ANTOLIC, A. Antimicrobial activity of Paliurus spina-christi Mill. Journal of Ethonpharm. V. 52, p. 119-122, 1996.
CAMERON, R. G.; MANTHEY, J. A.; BAKER, R. A.; GROHMANN K. Purification and characterization of a beta-glucosidase from Citrus sinensis var. Valencia fruit tissue. Journal of Agricultural and Food Chemistry, Washington, v. 49, p. 4457-4462, 2001.
CAMPANA FILHO, S. P.; BRITTO, D.; CURTI, E.; ABREU, F. R.; CARDOSO, M. B.; BATTISTI, M. V.; SIM, P. C.; GOY, R. C.; SIGNINI, R.; LAVALL, R. L. Extração, estruturas e propriedades de αααα- e ββββ- quitina. Disponível em: <http://www.scielo.br/scielo.php?pid=S010040422007000300026&script=sci_a ttext&tlng=pt> Acesso em: 28.Out.2008.
CARVALHO, M. S. L.; CALDAS, E. D.; DEGALLIER, N.; VILARINHOS, P. T. R.; SOUZA, L. C. K. R.; YOSHIZAWA, M. A. C.; KNOX, M. B.; OLIVEIRA, C. Suscetibilidade de larvas de Aedes aegypti ao inseticida temefós no Distrito Federal. Revista de Saúde Pública, v.38(5). p, 623-629, 2004.
CHEN, H.; LI, X.; LJUNGDAHL, L. G. Isolation and properties of an extracellula β-glucosidase from the polycentric rumen fungus Orpinomyces sp. strain PC-2 Applied and Environmental Microbiology, Washington, v. 60, p. 64-70, 1994. CHEN, J; TANG, B; CHEN, H; YAO, Q; HUANG, X; CHEN, J; ZHANG, D; ZHANG, W. Different Functions of the Insect Soluble and Membrane- Bound Trehalase Genes in Chitin Biosynthesis Revealed by RNA Interference. PLoS ONE. v. 5, p. 10133, 2010,

83
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
CHEN, L; CHUNG, Y. C; CHANG, YA-M; CHANG, CHEN-T. Characterisation of a b N-acetylhexosaminidase from a commercial papaya latex preparation. Food Chemistry. v. 124, p. 1404–1410, 2011. CHOTIA,C. AND LESK,A.M. The relation between the divergence of sequence and structure in proteins. EMBO J. v. 5, p. 823–826.1986. CHOWDHURY, NANDITA; LASKAR, SUBRATA; CHANDRA, GOUTAM. Mosquito larvicidal and antimicrobial activity of protein of Solanum villosum leaves. BMC Complementary and Alternative Medicine, v. 62, p.1-6, 2008.
CIFALI, A. P.; DIAS FILHO, B. P. Purification and partial chatacterization of N-acetyl-beta-D-glucosaminidase from Tritrichomonas foetus. Parasitol. Res, v. 85, p. 256-262, 1999.
CLSI (Clinical and Laboratory Standards Institute). Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically. v. 23, n. 2. 6 ed. Approved standard M7-A6. 2003. COATES, A. R. M; HU, Y. BR. J. Pharmacol, v. 152, p. 1147, 2007. COELHO, J. S; SANTOS, N. D.L; NAPOLEÃO, T. H; GOMES, F. S; FERREIRA, R. S; ZINGALI, R. B; COELHO, L. C.B.B; LEITE, S.P; NAVARRO, D. M.A.F; PAIVA, P. M.G. Effect of Moringa oleifera lectin on development and mortality of Aedes aegypti larvae. Chemosphere. v. 77, p. 934–938, 2009.
COELHO, P. A.; SANTOS, M. C.F. Ciclo biologico de Penaeus schmitti em Pernambuco (Crustacea, Decápoda, Penaeidae). Bol. Tecn. Cient.. CEPENE, Rio Formoso, v. 2, p. 35-50, 1994.
COHEN-KUPIEC, R; CHET, I. The molecular biology of chitin digestion. Currrent Opinion in Biotechnology. v. 9, p, 270-277, 1998. COLLINGE, D. B; KRAGH, K. M; MIKKELSEN, J. D; NIELSEN, K. K.; RASMUSSEN, U. & VAD, K. Plant chitinases. The Plant Jounal v. 3, p. 31–40, 1993. COSTA, J. G. M.; RODRIGUES, F. F. G. ANGÉLICO, E. C., SILVA, M. R. MOTA, M. L. SANTOS, N. K. A.; CARDOSO, A. L. H.; LEMOS, T. L. G. Estudo químico-biológico dos óleos essenciais de Hyptis martiusii, Lippia sidoides e Syzigium aromaticum frente às larvas do Aedes aegypti. Revista Brasileira de Farmacognosia, v.15(4), p.304-309, 2005.
COULON, S.; CHEMARDIN, P.; GUEGUEN, Y.; ARNAUD, A.; GALZY, P. Purification and characterization of an intracellular β-glucosidase from Lactobacillus casei ATCC 393. Applied Biochemistry and Biotechnology, Totowa, v. 74, p. 105-114, 1998.

84
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
COUTINHO, P. M; HENRISSAT, B. Carbohydrate-active enzymes: na integrated database approach. In Gilbert, H.J., Davies, g., Henrissat, B. and Svensson, B. (eds), Recent Advances in Carbohydrate Bioengineering. The Royal Society of Chemistry, Cambridge, p, 3-12, 1999.
DA SILVA FILHO, A.A.; SOUSA, J.P.B.; SOARES, S. Antimicrobial activity of the extract and isolated compounds from Baccharis dracunculifolia D. C. (Asteraceae). Journal of Biosciences, v.63 p.40-46, 2008.
DALL, W.; HILL, B.; ROTHLISBERG, P .C.; SHARPLES, D . J. The biology of Penaeidae. Advances in Marine Biology, Academic Press, London. v. 27, p. 489, 1990.
DAVIES, G.; HENRISSAT, B. Structures and mechanisms of glycosyl Hydrolases. Structure. v. 3, p.853-859, 1995. DAY, A. J.; DUPONT, M. S.; RIDLEY, S.; RHODES, M.; RHODES, M. J. C.; MORGAN, M. R. A.; WILLIAMSON, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Letters, Amsterdam, v. 436, p. 71-75. 1998. DE MARCO,J.L; LIMA, L. H. C; SOUSA, M. V; FELIX, C. R.A. Trichoderma harzianum chitinase destroys the cell wall of the phytopathogen Crinipellis the causal agent of witches beoom disease of cocoa. World Journal of Microbiology & Biotechnology, v. 16, p. 383-386, 2000. DI MARO, A; TERRACCIANO, I; STICCO, L; FIANDRA, L; RUOCCO, M; CORRADO, G; PARENTE, A; RAO, R. Purification and characterization of a viral chitinase active against plant pathogens and herbivores from transgenic tobacco. Journal of Biotechnology. v. 147, p. 1–6, 2010. DONNENBERG, MS. Escherichia coli Virulence mechanisms of a versatile pathogen. Baltimore: Academic press, Elsevier Science, 2002. DONNELLY, L. E.; BARNES, P. J. Acidic mammalian chitinase – a potencial target ashma therapy . Trends in pharmacological Sciences. v. 25, p. 509 511, 2004. DONPUDSA, S; PONPRATEEP, S; PRAPAVORARAT, A; VISETNAN, S; TASSANAKAJON, A; RIMPHANITCHAYAKIT, V. A Kazal-type serine proteinase inhibitor SPIPm2 from the black tiger shrimp Penaeus monodon is involved in antiviral responses. Developmental and Comparative Immunology. v. 34, p.1101 1108, 2010. DRIDER, D. et al. Isolation and characterization of the exocellular β-glucosidase of Candida cacaoi: possible use in carbohydrates degradation. LWT – Food Science and Technology, Amsterdam, v. 26, p. 198-204, 1993.

85
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
DUMONT, A. F. β β β β-glicosidases intestinais da larva de Diatrea saccharalis: clonagem e seqüenciamento dos cDNA, expressões e algumas propriedades.Dissertação de Mestrado em Ciências Bioquímicas – Universidade de São Paulo – SP, 2008.
DUO-CHUAN, L. Review of fungal chitinases. Mycopathologia, v. 60, p.161:345, 2006. DU, B; LONG, Y; LIU, H. Extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae bloodstream infection: risk factors and clinical outcome. Intensive Care Med. v, 28. p, 1718-23, 2002.
ESTATÍSTICA DA PESCA, 2007, Brasil Grandes Regiões e Unidades da Federação, Brasília- DF, Dezembro 2007.
EWALD, J .J. Estúdios sobre la biologia del camarón blanco, Penaeus schmitti. Acta Cientifica Venezuelana, Venezuela. v. 3, p. 190-200, 1967.
FERREIRA, A. H. P; MARANA, S.; TERRA, W. R; FERREIRA, C. Purification, sequencing, and properties of a beta-glycosidase purified from midgut lúmen of Tenebrio molitor (Coleoptera) larvae. Insect Biochem. Molec. Biol. v. 31, p. 1065 1076, 2001.
FERREIRA, A. H. P; TERRA, W. R; FERREIRA, C. Characterization of a beta glycosidase higghly active on disaccharides and of a beta-galactosidase from Tenebrio molitor midgut lúmen, Insect Biochem. Molec. Biol. v. 33, p. 253-265, 2003.
FERREIRA, A. H. P; TORRES, B. B.; TERRA, W. R. Substrate specificities of midgut beta-glycosidases from insects insects of different orders. Comp. Biochem. Physiol. v. 119b, p. 219-225, 1998.
FERREIRA, H. J. PURIFICAÇÃO PARCIAL E ESTUDOS CINÉTICOS DE N-ACETIL-ββββ-D-GLUCOSAMINIDASES EXTRAÍDAS DO HEPATOPÂNCREAS DA LAGOSTA PANULIRUS ARGUS (LATREILLE, 1804). Monografia- Universidade Federal do Ri Grande do Norte- RN, 2010.
FERREIRA, P. A. Purificação parcial e caracterização de uma ββββ-N acetilglucosaminidase de Palythoa variabilis. Dissertação de Mestrado em bioquímica – Universidade Federal do Rio Grande do Norte, Rio Grande do Norte- RN, 2003.
FERREIRA, P.M.P., CARVALHO, A.F.U., FARIAS, D.F., CARIOLANO, N.G., MELO, V.M.M., QUEIROZ, M.G.R., MARTINS, A.M.C., MACHADO-NETO, J.G.,. Larvicidal activity of the water extract of Moringa oleifera seeds against Aedes aegypti and its toxicity upon laboratory animals. Anais Acad. Brasil. Ciên. v.81, p. 207–216, 2009.
FLEURI, L. F.; SATO, H. H. β-1,3 glucanase. Biotecnologia: Ciência e Desenvolvimento, v. 37, p. 40-43, 2007.

86
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
FLEURI, L. F; SATO, H. H. PRODUÇÃO, PURIFICAÇÃO, CLONAGEM E APLICAÇÃO DE ENZIMAS LÍTICAS. Quim. Nova, v. 28, p. 871-879, 2005.
FOGAÇA, F. H. S.; LEGAT, J. F. A. Ensilagem de resíduos do beneficiamento do camarão marinho. Sigera. Florianópolis, SC. 2009.
FRANCO, B.D.G.M.; LANDGRAF, M. Microrganismos Patogênicos de Importância em Alimentos In: FRANCO, B.D.G.M.; LANDGRAF, M. Microbiologia dos alimentos. São Paulo: Atheneu, p. 33-81 , 2003.
GAMBA, A. L. & RODRIGUEZ, G. Migratory behavior of postlarval white, Penaeus schmitti, and river shrimps, Macrobrachium olfersi and Macrobrachium acanthurus in their zone of overlap in a tropical lagoon. Bulletin of Marine Science, v. 40, p. 454 463,1987.
GAVRILESCU, M. CHISTI, Y. Biotechnology a sustainable alternative for chemical industry. Review. Biotechnol. Adv., v. 23, p.471-499. 2005.
GERARDI, C.; BLANDO, F.; SANTINO, A.; ZACHEO, G. Purification and characterization of a β-glucosidase abundantly expressed in ripe sweet cherry (Prunus avium L.) fruit. Plant Science, Clare, v. 160, p. 795-805, 2001.
GERLACH, D; SCHLOTT, B. ZAHRINGER, V; SCHIMIDIT, K. N-acetil-D galactosamine/N-acetil-D-glucosamina-recognized lectin from the snail Cepaea hortensis: purification, chemical characterization, cloning and expression in E. coli. Immunology and Medicinal Microbiology, v. 43. p. 223-232, 2005.
GERMAIN, D. P. Gaucher’s disease: a paradigm for interventional genetics. Clinical Genetics, Oxford, v. 65, p. 77-86, 2004.
GILL, S. R., M. POP, R. T. DEBOY, P. B. ECKBURG, P. J. TURNBAUGH, B. S. SAMUEL, J. I. GORDON, D. A. RELMAN, C. M. FRASER-LIGGETT, AND K. E. NELSON. Metagenomic analysis of the human distal gut microbiome. Science. v. 312, p.1355–1359, 2006. GOMES - JÚNIOR, J. E; SOUZA, D. S. L; NASCIMENTO, R. M ; LIMA, Á. L. M ; MELO, J. A. T.; MILLER, R. N. G.; FRANCO, O. L.; GROSSI-DE-SA, M. F.; ABREU, L. R. D. . Purification and Characterization of a Liver-derived b-N Acetylhexosaminidase from Marine Mammal Sotalia fluviatilis. The Protein Journal. v. 29, p. 188-194, 2010. GOMES, L. P; OLIVEIRA, C. I. R; SILVA, M. C; ANDRADE, C.T; AGUILA, E. M. D; SILVA, J. T; PASCHOALIN, V. M. F. PURIFICAÇÃO E CARACTERIZAÇÃO DA QUITINASE DE UVA (Vitis vinífera L. CV RED GLOBE) PARA A PRODUÇÃO DE QUITOSANA A PARTIR DE QUITINA DE CAMARÃO. Quim. Nova, v. 33, p. 1882-1886, 2010.

87
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
GRACE, M. E.; NEWMAN, K. M.; SCHEINKER, V.; BERG-FUSSMAN, A.; GRABOWSKI, G. A. Analysis of human acid beta-glucosidase by site-ditected mutagenesis and heterologous expression. Journal of Biological Chemistry, Bethesda, v. 269, p. 2283-2291, 1994.
GUAN, Y; RAMALINGAM, S; NAGEGOWDA, D; TAYLOR, P. W. J.; CHYE, MEE L. Brassica juncea chitinase BjCHI1 inhibits growth of fungal phytopathogens and agglutinates Gram-negative bacteria. Journal of Experimental Botany, v. 59, p. 3475–3484, 2008. GUEGUEN, Y. et al. Enzymatic synthesis of dodecil.β-D-glucopyranoside catalyzed by Candida molischiana 35M5N β-glucosidase. Bioresource Technology, Oxford, v. 53, p. 263-267, 1995b. GUEGUEN, Y. et al. Purification and characterization of an intracellular β glucosidase from Botrytis cinerea. Enzyme and Microbial Technology, New York, v. 17, p. 900-906, 1995a. GUEGUEN, Y. et al. Purification and characterization of an intracellular β glucosidase from a new strain of Leuconostoc mesenteroides isolated from cassava. Jounal of Applied Microbiology, Oxford, v. 82, p. 469-476, 1997b. HAKI, G. D.; RAKSHIT, S.KDevelopments in industrially important thermostable enzymes: a review. Bioresource Technology.v. 89, p.17-34,2003. HAYASHI, S. Purification and characterization of the intracellular β-glucosidase from Aureobasidium sp ATCC 20524. Journal of Industrial Microbiology & Biotechnology, Heidelberg, v. 22, p. 160 – 163, 1999.
HAYS, W. S.; VANDERJAGT, D. J.; BOSE, B.; SERIANNI, A. S.; GLEW R. H. Catalytic mechanism and specificity for hydrolysis and transglycosylation reactions of cytosolic β-glucosidase from guinea pig liver. Journal of Biological Chemistry, Bethesda, v. 273, p. 34941-34948, 1998.
HENRISSAT, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochemistry Journal. v. 280, p. 309-316, 1991. HENRISSAT, B.; BAIROCH, A. New families in the classification of glycosyl hydrolases based on amino acid sequence similarities. Biochemical Journal. v. 293, p. 781–788, 1993. HENRISSAT, B.; CALLBAUT, I.; FABREGA, S.; LEHN, P.; MORNON, J.P.; DAVIES, G. Conserved catalytic machinery and prediction of a common fold for several families of glycosyl hidrolases. PNAS. v. 92, p. 7090-7094, 1995.
HOANG, KIM-CHI; LAI, TZU-HSUAN; LIN, CHUNG-SHENG; CHEN, YING-TSONG; LIAU, CHUN-YI. The Chitinolytic Activities of Streptomyces sp. TH-11. International Journal of Molecular Sciences. v, 12. p, 56-65, 2011.

88
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
HOLLAK, C.E.; VAN WEELY, S.; VAN OERS, M.H.; AERTS, J.M. Marked elevation of plasma chitotriosidase activity: A novel hallmark of Gaucher disease. J. Clin. Invest. v. 93,p. 1288–1292,1994.
HORSCH, M.; MAYER, C.; SENNHAUSER, U.; RAST, D. M. β-N acetylhexosaminidase: A target for the Design of Antifungical Agents. Pharmacol. Ther, v. 76, p. 187-218, 1997.
HOPKINS, T. L.; HARPER, M. S. Lepidopteran Peritrophic Membranes and Effects of Dietary Wheat Germ Agglutinin on Their Formation and Structure. Archives of Insect Biochemistry and Physiology. v. 47, p. 100-109, 2001.
IKEDA M, MIYAUCHI K, MOCHIZUKI A, MATSUMIYA M. Purification and characterization of chitinase from the stomach of silver croaker Pennahia argentatus. Protein Expression and Purification. v.65(2), p.214-222, 2009.
ISELI, B.; ARMAND, S.; BOILER, T.; NEUHAUS, J.M.; HENRISSAT, B. Plant chitinases use two different hydrolytic mechanisms. FEBS Letters, v. 382:186- p.188, 1996.
JAGER, S.; BRUMBAUER, A.; FEHÉR, E.; RÉCZEY, K.; KISS, L. Production and characterization of β-glucosidase from different Aspergillus strains. World Journal of Microbiology & Biotechnology, New York, v. 17, p. 455-461, 2001.
JEUNIAUX, C. Chitinolytic systems in the digestive tract of vertebrates: a review. In Chitin Enzymology. 233-244 R.A.A. Muzzarelli, ed European Chitin Society, Italy. 1993.
KAMEI, A.; HAYASHI, S. Properties of partially purified beta-N acetylglucosaminidase from bovine crystalline lens. Biol. Pharm. Bull. v. 22(8), p. 866 869, 1999.
KARAMANOS, Y. Endo- N-acetyl-β-D-glucosaminidase and their potential substrates: structure/function relations ships. Res Microbiol, v. 148, p. 661- 671, 1997.
KATAYEVA, I. A.; GOLOVCHENKO, N. P.; CHUVILSKAYA N, A.; AKIMENKO, V. K. Clostridium thermocellum β-glucosidases A and B: purification, properties, localization, ans regulation of biosynthesis. Enzyme and Microbial Technology, New York, v. 14, p. 407, 1992.
KIM, S. K. AND RAJAPAKSE, N. Enzymatic production and biological activities of chitosan oligosaccharides (COS): A review. Carbohydrate Polymers. v. 62, p.357–368, 2005.
KIMURA, I.; YOSHIOKA, N.; TAJIMA, S. Purification and characterization of a β glucosidase with β-xylosidase activity from Aspergillus sojae. Journal of Bioscience and Bioengineering, Osaka, v. 87, p. 538-541, 1999.

89
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
KIRK, O., BORCHERT, T.B., FUGLSANG, C.C. Industrial enzyme applications. Curr. Op. Biotechnol. v. 13, p.345-351. 2002. KITTL, R; WITHERS, S. G. New approaches to enzymatic glycoside synthesis through directed evolution. Carbohydrate Research. v. 345, p.1272–1279, 2010. KONG, H. J. ; CHO, H. K ; PARK, E. M ; HONG, G. E; KIM; Y. O; NAM; B. H; KIM; W. J; LEE, S.J; HAN, H. S; JANG, I. K; LEE, C. H; CHEONG, J; CHOI, T. J. Molecular cloning of Kazal-type proteinase inhibitor of the shrimp Fenneropenaeus chinensis. Fish & Shellfish Immunology. v. 26, p. 109-114, 2009. KOPPARAPU, N. K; LIU, Z; YAN, Q; JIANG, Z; ZHANG, S. A novel thermostable (PJC) from pomegranate (Punica granatum) juice. Food Chemistry, v. 127, p. 1569 1575, 2011. KOSHLAND, D.E. Stereochemistry and the mechanism of enzymatic reactions. Biol. Reviews. v. 28, p.416-436, 1953. KOSTANJNEK, R; MILATOVIB, M; KTRUS, J. Endogenous origin of endo-β-1,4-glucanase in common woodlouse Porcellio scaber (Crustacea, Isopoda). J Comp Physiol B. v. 180, p. 1143–1153, 2010.
KRESSE, H.; GLOSSL, J. Glycosaminoglycans degradation. Adv. Enzymol., v. 60,p. 217-311, 1987.
KUNZ, A. S. Resolution and properties of the proteinase in the larva of the mosquito, Aedes aegypti. Insect Biochem. v. 8, p. 43-51, 1978.
KUSAYKIN, M.I.; BURTSEVA, Y.V.; SVETASHEVA, T.G., SOVA, V.V.; ZVYAGINTSEVA, T.N. Distribution of O-glycosilhydrolases in marine invertebrates. Enzymes of the marine mollusk Littorina kurila that catalyse fucoidan transformation. Biochemistry (Moscow), v.68, n.3, p.384- 392, 2003.
LAEMMLI, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, August 1970, v. 227, no. 5259, p. 680-685.
LAM, S. K; NG, T. B. Acaconin, a chitinase-like antifungal protein with cytotoxic and anti-HIV-1 reverse transcriptase activities from Acacia confuse seeds. ACTAABP. v. 57, 2010. LAUTENBACH, E; STROM, BL; BILKER, WB. Epidemiological investigation of fluoroquinolone resistance in infections due to extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Clin Infect Dis. v. 33, p. 1288-1294, 2001. LEE, H.S; HAM, D.S; CHOI, S.J; KIM, D,S; BAI, D.H; YU, J.H. Purification, characterization, and primary structure of a chitinase from Pseudomonas sp. YHS-A2. App. Microbiol. Biotechnol.v, 54. p, 397-405, 2000.

90
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
LENARDON, M. D; MUNRO, C. A; GOW, N. AR. Chitin synthesis and fungal pathogenesis. Current Opinion in Microbiology. v, 13. p, 416–423, 2010. LE TRAON-MASSON, M. P.; PELLERIN, P. Purification and characterization of two β-D-glucosidases from an Aspergillus niger enzyme preparation: affinity and specificity toward glucosylated compounds characteristic of the processing of fruits. Enzyme and Microbial Technology, New York, v. 22, p. 374-382, 1998.
LI, X. et al. Expression, purification and characterization of a recombinant β- glucosidase from Volvariella volvacea. Biotechnology Letters, Dordrecht, v. 27, p. 1369-1373, 2005.
LIMA NETO, E. C. Identificação de genes Relacionados ao Sistema Imune do Camarão marinho Litopenaeus vannamei. Dissertação de Mestrado em Recursos Pesqueiros e Aqüicultura – Universidade Federal Rural de Pernanbuco, Pernambuco-PE, 2006.
LIMA, A. L. M. Purificação e caracterização parcial de duas N-acetil-ββββ hexosaminidases do equinoderma marinho Echinometra lucunter. Dissertação de Mestrado em Bioquímica - Universidade Federal do Rio Grande do Norte - RN, 2006.
LIMA, E. P.; OLIVEIRA FILHO, A. M.; LIMA, J. W. O.; RAMOS JUNIOR, A. N.; CAVALCANTI, L. P. G.; PONTES, R. J. S. Resistência do Aedes aegypti ao Temefós em Municípios do Estado do Ceará. Revista da Sociedade Brasileira de Medicina Tropical. v.39(3), p.259-263, 2006.
LIMA, J. B. P.; MELO, N. V.; VALLE, D. Residual effect of two Bacillus thuringiensis var. israelensis products assayed against Aedes aegypti (diptera: culicidae) in laboratory and outdoors at Rio de Janeiro, Brazil. Revista do Instituto de Medicina Tropical de São Paulo. v. 47(3), p.125-130. 2005b.
LIN, J. C.; XIE, X. L.; GANG, M.; WANG, Q.; CHEN, Q. X. Effects of mercuric íon on the conformation and activity of Penaeus vannamei β-N-acetyl-D-glucosaminidase. International Journal of Biological Macromolecules. v. 36, p. 327-330, 2005. LIN, S. B., LIN, Y. C., & CHEN, H. H. Low molecular weight chitosan prepared with the aid of cellulase, lysozyme and chitinase: Characterisation and antibacterial activity. Food Chemistry. v, 116. p. 47–53, 2009. LINEWEAVER, H.; BURK, D. The determination of enzyme dissociation constants. Journal of the American Chemical Society, Washington, v. 56, p. 658-666, 1934. LOURENÇO-DE-OLIVEIRA R, VAZEILLE M, DE FILIPPIS AMB, FAILLOUX AB. Aedes aegypti in Brazil: genetically differentiated populations with high susceptibility to dengue and yellow fever viruses. American Journal of Tropical Medicine & Hygiene.;v. 98, P. 43-54. 2004.

91
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
LUCAS, R. et al. β-glucosidase from Chalara paradoxa CH32: purification and properties. Journal of Agricultural and Food Chemistry, Washington, v. 48, p. 3698-3703, 2000.
LUNA, J. D. S. M. F; ANJOS, A. F; KUWABARA, E. F. NAVARRO-SILVA, M. A. Susceptibility of Aedes aegypti to temephos and cypermethrin insecticides. Revista de Saúde Publica. Brazil, v. 38, p. 1-2, 2004.
LUO, T; YANG, H; LI, F; ZHANG, X; XU, X. Purification, characterization and cDNA cloning of a novel lipopolysaccharide-binding lectin from the shrimp Penaeus monodon. Developmental & Comparative Immunology, v. 20, p. 1-11, 2005. LUSTERIO, D. D. et al. Alkali resistant, alkaline endo-1,4-β-glucanase produced by Bacillus sp. PKM-5430. Bioscience, Biotechnology and Biochemistry, Tokyo, v. 56, p. 1671-1672, 1992.
LUVESUTO, E. Analise genética e morfométrica da estrutura populacional do camarão branco Litopenaeus schmitti (Decápoda, Crustácea) na costa do Rio Grande do Norte, Brasil: uma abordagem em fina escala. Dissertação de Mestrado em Ecologia em Recursos Naturais- Universidade Federal de São Carlos, São Paulo-SP, 2006.
LUZ, C.; TAI, M. H. H.; SANTOS, A. H.; SILVA, H. H. G. Impact of moisture on survival of Aedes aegypti eggs and ovicidal activity of Metarhizium anisopliae under laboratory conditions. Memórias do Instituto Oswaldo Cruz, v. 103(2), p.214-215, 2008.
MACORIS, M. L. G.; ANDRIGHETTI, M. M. T.; OTRERA, V. C. G,; CARVALHO, L. R.; CALDAS JUNIOR, A. L., BROGDON, W. G. Association of insecticide use and alteration on Aedes aegypti susceptibility status. Memórias do Instituto. Oswaldo Cruz. v. 102(8),p. 895-900, 2007.
MALEY, F.; TRIMBLE, R.B.; TARENTINO, A. L.; PLUMMER, T. H. JR. Characterization of glycoproteins and their associated oligosaccharides through the use of endoglicosidases. Analytical Biochemisty, v. 180, p. 195-204, 1989.
MARANA, S. R.; TERRA, W, R; FERREIRA, C. Midgut β-glucosidases from Abracris flavolineta (Orthoptera: Acrididae). Physycal properties, substrate soecificities and function. Insect Biochemistry ans Molecular Biology, Oxford, v. 25, p. 835-843, 1995.
MARANA, S.R. Purificação e caracterizaçã das ββββ-glicosidases digestivas de Spodoptera frugiperda (Lepidóptera). Tese de Doutorado em Bioquímca-Universidade de São Paulo - São Paulo, SP, 1999.
MARQUES, M. R.F; BARRACCO, M. A. Lectins, as non-self-recognitin factors, in crustaceans. Aquaculture, v. 191, p. 23-44, 2000.

92
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
MARTINELLI, J. M. Estrutura populacional dos camarões Penaeidae no estuário do rio Caeté, litoral Norte do Brasil. Tese de Doutorado. Universidade Federal do Pará, p. 174, 2005.
MATEO, J. J.; DI STEFANO, R. Description of the β-glucosidase activity of wine yeasts. Food Microbiology, London, v. 14, p. 583-591, 1997.
MATSUMIYA, M; ARAKANE, Y; HAGA, A; MUTHUKRISHNAM, S; KRAMER, K. J. Substrate specificity of chitinases from two species of fish, greenling, Hexagrammos otakii, and common mackerel, Scomber japonicas, and the insect, tobacco hornworm, Manduca sexta. Biosci Biotechnol Boichem. v. 70, p. 971-979, 2006.
MATSUO, Y.; KURITA, M.; PARK, J. K.; TANAKA, K.; NAKAGAWA, T.; KAWAMUKAI, M.; MATSUDA, H. Purification, characterization and gene analysia of N-acetylglucosaminidase from Enterobacter sp. G-1. Biosci. Biotechnol. Biochem., v. 63, p. 1261-1268, 1999.
MATSUURA, K.; SAWADA, H.; YOKOSAWA, H. Purification and properties of N acetylglucosaminidase from eggs os the ascidian, Halocynthia roretzi. Eur. J. Biochem., v. 218, n. 2, p. 535-541, 1993.
MATTA, L. D. M.; ABREU, L. R. D. Glicosidases e sulfatases no molusco marinho Aplysia cervina. Arq. Inst. Biol., v. 72, p. 205-210, 2005.
MEDEIROS, V. F. Potencial larvicida de extratos de plantas regionais no controle de larvas de Aedes aegypti (Diptera: Culicidae). Dissertação de Mestrado em Ciências Biológicas – Universidade Federal do Rio Grande do Norte – RN, 2007.
MORAIS, H. Liquid chromatographic and electrophoretic characterization of extracellular β-glucosidase of Pleurotus ostreatus grown in organic waste. Journal of Chromatography B, Amsterdam, v. 770, p. 111-119, 2002.
MOREIRA, R. A; MONTEIRO, A. C. O; HORTA, A. C. G; OLIVEIRA, J. T. A; CAVADA, B. S. Isolation and characterization of Dioclea altíssima var. megacarpa seed lectin. Phytochemistry, v. 46, p. 139-144, 1977.
MULLEN, K. M.; PETERS, E. C.; HARVELL, C. D. Coral Resistance to Disease. In E.Rosenberg & Y.Loya (eds) Coral Health & Disease, Springer-Verlag. 2004. NAKANO, H. et al. Purification and characterization of a novel β-glucosidase from Clavibacter michiganense that hydrolyzes glucosyl ester linkage in steviol glycosides. Journal of Fermentation and Bioengineering, Osaka, v. 85, p. 162 168, 1998.
NASCIMENTO, R. M. Purificação e caracterização parcial de uma ββββ-D glicosidase de Artemia franciscana com ação sobre celobiose e lactose. Dissertação de Mestrado em bioquímica – Universidade Federal do Rio Grande do Norte, Rio Grande do Norte- RN, 2008.

93
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
NEIVA, G. S.; MOURA, S. J. C. Sumário sobre a exploração de recursos marinhos do litoral brasileiro: situações atuais e pespectivas. PDP, Série de documentos ocasionais, v. 27, p. 44, 1971.
NEMETH, K.; PLUMB, G. W.; BERRIN, JG.; JUGE N,; JACOB, R.; NAIM, H. Y.; WILLIAMSON, G.; SWALLOW, D. M.; KROON, P. A. Deglycosylation by small intestinal epithelial cell β-glucosidases is a critical step in the absorption and metabolism of dietary flavonoid glycosides in humans. European Journal of Nutrition, Darmastadt, v. 42, p. 29-42, 2003.
NIIMI, K.;SHEPHERED, M. G; CANNON, R. D. Distinguishing Candida Species by β-N-acetylhexosaminidase activity. Journal of Clinical Microbiology, v. 39, p. 2089-2097, 2001.
NOSTRO, A; CELLINI, L; DI BARTOLOMEO, S; DI CAMPLI, E; GRANDE, R; CANNATELLI, MA. Antibacterial effect of plant extracts against Helicobacter pylori. Phytother Res. v. 19, p. 198–202, 2005.
OH, K. B. et al. Isolation and properties of an extracellular β-glucosidase from a filamentous fungus, Cladosporium resinae, isolated from kerosene. Bioscience, Biotechnology and Biochemistry, 1999.
PAAVILAINEN, S.; HELLMAN, J.; KORPELA, T. Purification, characterization, gene cloning, and sequencing of a new β-glucosidase from Bacillus circulans subsp. Alkalophilus. Applied and Enviromenmental Microbiology, Washington, v. 59, p. 927-932, 1993.
PAINBENI, E. et al. Purification and characterization of a Bacillus polymyxa β- glucosidase expressed in Escherichia coli. Journal of Bacteriology, Washington, v. 174, p. 3087-3091, 1992. PAYNE, D. J; GWYNN, M. N; HOLMES, D. J; PAMPLIANO, D. L. Drug Discovery Nat. Rev. v. 6, p. 29, 2007. PALLA, C. A; PACHECO, C; CARRÍN, M. E. Preparation and modification of chitosan particles for Rhizomucor miehei lipase immobilization. Biochemical Engineering Journal. 2011.
PASTORE GM, R S, KOBLITZ MGB. Purificação parcial e caracterização bioquímica de lipase extracelular produzida por nova linhagem de Rhizopus sp . v.23(4), p.135-140, 2003.
PATEL, A. K; SHING, V. K; YADAV, R. P; et al.Purification and characterization of a new chitinas e from latex of Ipomoea carnea. Process Biochemistry.v.45(5), p.675-681, 2010.
PATIL, R. S.; GHORMADE, V.; DESHPANDE, M. V.; Enzyme Microb. Technol. v. 26, p. 473, 2000.

94
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
PÉREZ-FARFANTE, I. Sinopsis de datos biológicos sobre el camarón blanco Penaeus schmitti. FAO Fishery Reports, Bunkenroad, v. 37, p. 1417-1438, 1970.
PETERS, G.; SABOROWSKI, R.; MENTLEIN, R.; BUCHHOLZ, F. Isoforms of an N-acetyl-β-D-glucosaminidase and antibody production. Comparative Biochemistry and Physiology part B 120, 1998.
POLANCZYK, R. A.; GARCIA, M. O.; ALVES, S. B. Potencial de Bacillus thuringiensis israelensisBerliner no controle de Aedes aegypti. Revista de Saúde Pública. v. 37(6), p.813-816, 2003.
PONTIN, K; DA SILVA FILHO, AA; SANTOS, FF; ANDRADE E SILVA, ML. CUNHA, WR; NANAYAKKARA, NPD. In vitro and in vivo antileishmanial activities of a Brazilian green propolis extract. Parasitol Res. v, 103. p, 487-92, 2008.
PONTOH, J.; LOW, N. H. Purification and characterization of β-glucosidase from honey bees (Apis mellifera). Insect Biochemistry and Molecular Biology, Oxoford, v. 32, p. 67690, 2002.
PROESPRAIWONG, P; TASSANAKAJON, A.; RIMPHANITCHAYAKIT, V. Chitinases from the black tiger shrimp Penaeus monodon: Phylogenetics, expression and activities. Comparative Biochemistry and Physiology, Part B. v. 156, p. 86 96, 2010. QIN, Y; LU, X; SUN, N; ROGERS, R. D. Dissolution or extraction of crustacean shells using ionic liquids to obtain high molecular weight purified chitin and direct production of chitin films and fibers. Green Chem. v, 12. p, 968–971, 2010. RAO, D., & GOWDA, L. Abundant class III acidic chitinase homologue in Tamarind (Tamarindus indica) seed serves as the major storage protein. Journal of Agricultural Food Chemistry. v. 56,p. 2175–2182, 2008. REGIS, L.; MONTEIRO, A. M.; MELO-SANTOS, M. A. V.; SILVEIRA JR, J. C.; FURTADO, A. F.; ACIOLI, R. V.; SANTOS, G. M.; NAKAZAWA, M.; CARVALHO, M. S.; RIBEIRO JR, P. J.; SOUZA, W. V. Developing new approaches for detecting and preventing Aedes aegypti population outbreaks: basis for surveillance, alert and control system. Memórias do Instituto Oswaldo Cruz. v. 103(1), p. 50-59, 2008.
REVISTA PANORAMA DA AQUICULTURA, 2004, 14, 21, http://www.panoramadaaquicultura.com.br, acessada em Janeiro 2010.
RIBEIRO, M.C. & SOARES, M.M.S.R. Microbiologia prática: roteiro e manual. São Paulo: Atheneu, 2000.
RICCIO, P. et al. Extraction and immobilization in one step of two β-glucosidases released from a yeast strain of Debaryomyces hansenii. Enzyme and Microbial Technology, New York, v. 24, p. 123-129, 1999.

95
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
RIOU, C. et al. Purification, characterization, and substrate specificity of a novel highly glucose-tolerant β-glucosidase from Aspergillus oryzae. Applied and Environmental Microbiology, Washington, v. 64, p. 3607-3614, 1998.
RIVAS, B.; GARCIA, J. L.; LOPES, R.; GARCIA, P. Purification and polar localization of pneumococcal LytB, a putative endo-beta-N acetylglucosaminidase: the chain-dispersing murein hydrolases. J. Bacteriol, v. 184, p. 4988-5000, 2002.
ROBERTS, G. A. F.; Chitin Chemistry, Mc Millan Press Ltd: London, 1992, cap 1.
ROCHA, I. P.; Revista da ABCC. V. 20. 2008.
RODRÍGUEZ-BAÑO, J; NAVARRO, MD; ROMERO, L. Clinical and molecular epidemiology of extended-spectrum β-lactamase-producing Escherichia coli as a cause of nosocomial infection or colonization: implications for control. Clin Infect Dis. v. 42, p. 37-45, 2006. RODRIGUES, M. M.; BISSET, J.; FERNANDEZ, D. M.; LAUZÁN, L.; SOCA, A. Detection of insecticide resistance in Aedes aegypti (Diptera, Culicidae) from Cuba and Venezuela. Journal of Medical Entomology. v. 38, p.623-628, 2001.
ROGER, H. J; PERKINS, H.R. Cell walls of filamentous fungi. E. and F. N. Spon: London, 23, cap 9, 153-160. 1968.
ROTA, M. A. Aspectos Gerais da Fisiologia e Estrutura do Sistema Digestivo dos Peixes Relacionados a Piscicultura. Embrapa Pantanal, p. 48. 2003.
ROTHLISBERG, P.; JACKSON, C. J.; PENDREY, R. C. Distribution and abundance of early penaeid larvae in the Gulf of Carpentaria, Second Australian National Prawn Seminal NPS2. Australia, p. 23-30, 1985.
SÁ, R. A; SANTOS, N. D. LI; SILVA, C. S. B; NAPOLEÃO, T. H; GOMES, F. S; CAVADA, B. S; COELHO, L. C. B. B; NAVARRO, D. M. A. F; BIEBER, L. W; PAIVA, P. M. G. Larvicidal activity of lectins from Myracrodruon urundeuva on Aedes aegypti. Comparative Biochemistry and Physiology, Part C. v. 149, p. 300–306, 2009. SAHAI, A.S; MANOCHA, M.S. Chitinases of fungi and plants: their involvement in morphogenesis and host-parasite interaction. FEMS Microbiol. Rev. v. 11, p. 317-338, 1993. SAMOSHINA, N. M; SAMOSHIN, V. V. The Michaelis constants ratio for two substrates with a series of fungal (mould and yeast) β-galactosidases. Enzyme and Microbial Technology, New York, v. 36, p. 239-251, 2005. SANTANA, T. T. S. Imobilização de uma n-acetil-β-d-glucosaminidase extraída do camarão marinho Litopenaeus schmitti em Dacron ferromagnético. Monografia – Universidade Federal do Rio Grande do Norte, Rio Grande do Norte- RN, 2009.

96
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
SANTOS, V. O. Polissacarídeos Sulfatados de Interesse Farmacológico no Camarão Marinho Litopenaeus schmitti. Dissertação de Mestrado em Bioquímica – Universidade Federal do Rio Grande do Norte - RN, 2006.
SARRY, J. E.; GÜNATA, Z. Plant and microbial glycoside hydrolases: volatile release from glycosidic aroma precursors. Food Chemistry, Oxford, v. 87, p. 509 521, 2004. SAVAGEAU, MA. Escherichia coli habitats, cell types, and molecular mechanisms of gene control. Am Nat. v, 122. p, 732–744, 1983. SHAHIDI, F; ARACHCHI, J. K. V; JEON, Y. J. Food applications of chitin and chitosans. Trends in Food Science & Technology. v, 10. p, 37 – 51, 1999.
SCHLUMBAUM, A.; MAUCH, F.; VÖGELI, U.; BOLLER, T. Plant chitinases are potent inhibitors of fungal growth. Nature, v. 324, p.365–367, 1986.
SEDMAK J & GROSSBERG SE. A rapid, sensitive, and versatile assay for protein using coomassie brilhant blue G250. Analytical Biochemistry. 1977, 79, 544-552.
SEIDL, V. Chitinases of filamentous fungi: a large group of diverse proteins with multiple physiological functions. Fungal Biology Reviews, Austria, v. 22, p. 36-42, 2008.
SESTELO, A. B. F.; POZA, M.; VILLA, T. G. β-glucosidase activity in a Lactobacillus plantarum wine strain. World Journal of Microbiology & Biotechnology, New York, v. 20, p. 633-637, 2004.
SHI, Y; JIANG, Z; HAN, P; ZHENG G-X; SONG, K-K; CHEN, Q-X. Purification and some properties of b-N-acetyl- D-glucosaminidase from the cabbage butterfly (Pieris rapae). Biochimie.v. 89, p. 347-354, 2007. SILVA, H. H. G.; GERIS, R.; RODRIGUES FILHO, E.; ROCHA, C.; SILVA, I. G. Larvicidal activity of oil-resin fractions from the Brazilian medicinal plant Copaifera reticulata Ducke (Leguminosae-Caesalpinoideae) against Aedes aegypti (Diptera, Culicidae) Revista da Sociedade Brasileira de Medicina Tropical. v. 40(3), p.264-267, 2007.
SILVA, J. J.; MENDES, J. Susceptibility of Aedes aegypti (L) to the insect growth regulators diflubenzuron and methoprene in Uberlândia, State of Minas Gerais. Revista da Sociedade Brasileira de Medicina Tropical, v.40(6), p. 612-616, 2007.
SILVA, O. Aspectos Bioecológicos e pesqueiros de três especies de camarões do gênero Penaeus nas costa do estado do Rio de Janeiro e Experimentos de cultivo. Dissertação de Mestrado, Pós-Graduação em Zoologia, Universidade Federal do Rio de Janeiro, RJ, 1977.

97
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
SILVA, W.J., DÓRIA, G.A.A., MAIA, R.T., NUNES, R.S., CARVALHO, G.A., BLANK, A.F., ALVES, P.B., MARÇAL, R.M., CAVALCANTI, S.C.H.,. Effects of essential oils on Aedes aegypti larvae: alternatives to environmentally safe insecticides. Bioresour. Technol.v. 99, p.3251–3255, 2008. SILVA, W.J., DÓRIA, G.A.A., MAIA, R.T., NUNES, R.S., CARVALHO, G.A., BLANK, A.F., ALVES, P.B., MARÇAL, R.M., CAVALCANTI, S.C.H. Effects of essential oils on Aedes aegypti larvae: alternatives to environmentally safe insecticides. Bioresour. Technol. v. 99,p. 3251–3255, 2008. SILVEIRA, L.M.S.; OLEA, R.S.G.; MESQUITA, J.S. et al. Metodologias de atividade antimicrobiana aplicadas a extratos de plantas: comparação entre duas técnicas de Agar difusão. Revista Brasileira de Farmacognosia, v.90, n.2, p.124-128, 2009. SIMAS, N. K.; LIMA, E. C.; CONCEIÇÃO, S. R.; KUSTER, R. M.; OLIVEIRA FILHO, A. M. Produtos naturais para o controle da transmissão da dengue - Atividade larvicida de Myroxylon balsamum (óleo vermelho) e de terpenóides e fenilpropanóides. Química Nova, v.27(1), p.46-49, 2004.
SOUZA, DJAIR S L ; GROSSI SÁ, M. F. ; FRANCO, O. L. ; G JÚNIOR, JOSÉ EDILSON ; ABREU, L. R. D. Identification of a novel b-N-acetylhexosaminidase (Pcb-NAHA1) from marine Zoanthid Palythoa caribaeorum (Cnidaria, Anthozoa, Zoanthidea). Protein Expression and Purification.v, 58. p, 61-69. 2008.
SPINDLER-BARTH, M. Hormonal regulation of chitin metabolism in insect cell lines. In Chitin Enzymology. p. 75-82. R.A.A. Muzzarelli, ed. European Chitin Society, Italy. 1993.
STINTZI, A.; HEITZ, T.; WIEDEMANN, M. S.; KAUFFMANN, S.; GEOFFROY, P. Plant “pathogenesis-related” proteins and their role in defense against pathogens. Biochemie. v. 75, p. 687-706, 1993.
SUE, M.; ISHIHARA, A.; IWAMURA, H. Purification and characterization of a β glucosidase from rue (Secale cereal L.) seedlings. Plant Sciences, Clare, v. 155, p. 67-74, 2000.
TAIRA, T., OHDOMARI, A., NAKAMA, N., SHIMOJI, M., & ISHIHARA, M. Characterization and antifungal activity of Gazyumaru (Ficus microcarpa) latex chitinases: Both the chitin binding and the antifungal activities of class I chitinase are reinforced with increasing ionic strength. Bioscience, Biotechnology, and Biochemistry.v. 69,p. 811–818, 2005a. TAIRA, T., TOMA, N., & ISHIHARA, M. Purification, characterization, and antifungal activity of chitinases from pine apple (Ananas comosus) leaf. Bioscience, Biotechnology, and Biochemistry. v.69, p. 189–196, 2005b. THAKORE, Y.B. Enzymes for Industrial Applications. BBC RESEARCH. BIO030 Published: January 2008.

98
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
TERRA, W. R; FERREIRA, C; DE BIANCHI, A.G. Action pattern, kinetical properties and electophoretical studies of an alpha-amylase present in midgut homogenates from Rhynchonsciara Americana (Diptera) larvae. Comparative Biochemistry and Physiology, v.56B, p. 201-209. 1997.
THARANATHAN, R.N.; KITTUR, F.S. Chitin—the undisputed biomolecule of great potential, Critical Reviews in Food Science and Nutrition. v. 43, p. 61–87, 2003.
THOMPSON SE, SMITH M, WILKINSONS MC, PEEK K. Identification and characterization of a chitinase antigen from Pseudomonas aeruginosa strain 385. Appl Environ Microbiol. v. 67, p.4001-4008, 2001.
TSUJIBO, H., FUJIMOTO, K., TANNO, H., MIYAMOTO, K., KIMURA, Y; IMADA, C., OKAMI, Y. AND INAMORI, Y. Molecular cloning of the gene which encodesb-N-acetylglucosaminidase from a marine bacterium,Alteromonassp. Strain O-7.Appl Environ Microbiol. v. 61, p. 804–806,1995.
TU , S; QIU, X; CAO, L; HAN, R; ZHANG, Y; LIU, X. Expression and characterization of the chitinases from Serratia marcescensGEI strain for the control of Varroa destructor, a honey bee parasite. Journal of Invertebrate Pathology.v. 104, p.75–82, 2010.
VIEIRA, V. K. B. Detecção e caracterização parcial da ββββ-N acetilglucosaminidase em extratos de gônadas do molusco Chiton sp. Dissertação de Mestrado em bioquímica – Universidade Federal do Rio Grande do Norte, Rio Grande do Norte-RN, 2002.
VISETNAN, S; DONPUDSA, S; SUPUNGUL, P; TASSANAKAJON, A; RIMPHANITCHAYAKIT, V. Kazal-type serine proteinase inhibitors from the black tiger shrimp Penaeus monodon and the inhibitory activities of SPIPm4 and 5. Fish & Shellfish Immunology. v. 27, p. 266–274, 2009. YAN, T. R.; LIAU, J. C. Synthesis of cello-oligosaccharides from cellobiose with β-glucosidase II from Aspergillus niger. Biotechnology Letters, Dordrecht, v. 20, p. 591-594, 1998. YANG, H; LUO, T; LI, F; LI,S ; XU, X. Purification and characterization of a calcim-independent lectin (PjLec) from the haemolymph of the shrimp Penaeus japonicus. Fish & Shellfish Immunology, v. 22, p. 88-97, 2007.
YE X; NG T. B. A chitinase with antifungal activity from the mung bean. Protein Expression and Purification. v. 40, p. 230–236, 2005.
YEN, M. T.; YANG, J. H.; MAU, J. H. Physicochemical characterization of chitin and chitosan from crabs shells. Carbohydrate polymers. v. 75, p. 15-21, 2009. YUN, S. I.; JEONG, C. S.; CHUNG, D. K.; CHO, H. S. Purification and some properties of a β-glucosidase from Trichoderma harzianum type C-4. Bioscience, Biotechnology and Biochemistry, Tokio, v. 65, p. 2028-2032, 2001.

99
REFERÊNCIAS
Programa de Pós-Graduação em Bioquímica Roberta L. N. Godone
YAZDI, M. T. et al. Purification and characterization of two intracellular β- glucosidases from the Neurospora crassa mutant cell-1. World Journal of Microbiology & Biotechnology, New York, v. 19, p.79-84, 2003.
XAVIER-FILHO, J. The biological roles of serine and cysteine proteinase inhibitors in plants. Revista Brasileira de Fisiologia Vegetal. v. 4, 1992.
ZANOELO, F. F. et al. β-Glucosidase activity from the thermophilic fungus Scytalidium thermophilum is stimulated by glucose and xylose. FEMS Microbiology Letters, Oxford, v. 240, p. 137-143, 2004.
ZHANG, J; SUN, Y; LI, F; HUANG, B; XIANG, J. Molecular characterization and expression analysis of chitinase (Fcchi-3) from Chinese shrimp, Fenneropenaeus chinensis. Mol Biol Rep.v, 37. p. 1913–1921, 2010. WANG, S; SHAO, B; RAO, P; LEE; Y; YE; X . Hypotin, a Novel Antipathogenic and Antiproliferative Protein from Peanuts with a Sequence Similar to Those of Chitinase Precursors. J. Agric. Food Chem., v. 55, p. 9792–9799, 2007. WEI, D. L. et al. Purification and characterization of an extracellular β- glucosidase from the wood-grown fungus Xylaria regalis. Current Microbiology, New York, v. 33, p. 297-301, 1996. World Health OrganizationInstructions for Determining the Susceptibility or Resistance of Mosquito Larvae to Insecticides. WHO/VBC/81.807, pp. 1-6. , 1981.





























