Embed Size (px)
Citation preview

NeuroImage 11, 708–734 (2000)doi:10.1006/nimg.2000.0562, available online at http://www.idealibrary.com on
Variability in fMRI: An Examination of Intersession DifferencesD. J. McGonigle, A. M. Howseman, B. S. Athwal, K. J. Friston, R. S. J. Frackowiak, and A. P. Holmes1
Wellcome Department of Cognitive Neurology, Institute of Neurology, London WC1N 3BG, United Kingdom
Received September 22, 1999
The results from a single functional magnetic reso-nance imaging session are typically reported as indic-ative of the subject’s functional neuroanatomy. Under-lying this interpretation is the implicit assumptionthat there are no responses specific to that particularsession, i.e., that the potential variability of responsebetween sessions is negligible. The present studysought to examine this assumption empirically. A totalof 99 sessions, comprising 33 repeats of simple motor,visual, and cognitive paradigms, were collected over aperiod of 2 months on a single male subject. For eachparadigm, the inclusion of session-by-condition inter-actions explained a significant amount of error vari-ance (P < 0.05 corrected for multiple comparisons)over a model assuming a common activation magni-tude across all sessions. However, many of those vox-els displaying significant session-by-condition interac-tions were not seen in a multisession fixed-effectsanalysis of the same data set; i.e., they were not acti-vated on average across all sessions. Most voxels thatwere both significantly variable and activated on av-erage across all sessions did not survive a random-effects analysis (modeling between-session variance).We interpret our results as demonstrating that correctinference about subject responses to activation taskscan be derived through the use of a statistical modelwhich accounts for both within- and between-sessionvariance, combined with an appropriately large ses-sion sample size. If researchers have access to only asingle session from a single subject, erroneous conclu-sions are a possibility, in that responses specific to thissingle session may be claimed to be typical responsesfor this subject. © 2000 Academic Press
INTRODUCTION
This study assesses the generality of results ob-tained from a single functional magnetic resonanceimaging (fMRI) session. fMRI is a noninvasive tech-nique that has revolutionized the study of human brainfunction (e.g., Belliveau et al., 1991; Ogawa et al., 1992;
1 Current address: Robertson Centre for Biostatistics, Boyd OrrBuilding, University Avenue, Glasgow G12 8QQ, Scotland, UK.
7081053-8119/00 $35.00Copyright © 2000 by Academic PressAll rights of reproduction in any form reserved.
Kwong et al., 1992). As with many brain imaging tech-niques, such as positron emission tomography, a num-ber of observations (scans) from each subject are col-lected. A single experimental examination of onesubject in this fashion constitutes a session. Althoughexceptions exist, it is unusual for a subject to bescanned on more than one occasion and, more oftenthan not, a single fMRI session is assumed to give anaccurate representation of a subject’s functional neu-roanatomy.
There may be problems, however, if one adopts this“one subject, one session” approach to neuroimagingexperiments. One session is only a single, discrete“snapshot” of the subject’s brain and may not epitomizeresponses to the sensorimotor or cognitive challengeemployed. Indeed, differences between sessions are in-evitable: for example, the BOLD response is an indirectand semiqualitative measure of neuronal activity, andthe relationship between BOLD contrast and cerebraloxygen metabolism is influenced by a number of phys-iological factors (e.g., for review see Ogawa et al.,1998). Furthermore, single-session results may be in-fluenced by slight variations in the hardware charac-teristics of the MR scanner, which are not systematicacross sessions (e.g., the shim performed to homoge-nize the B0 field of the scanner; Howseman et al., 1998).Any differences in subject position within the headcoilon separate scanning sessions may also result ingreater variability in voxel signal changes, due to par-tial volume effects, as may different patterns of subjectmovement between sessions. In addition to the above,nonspecific physiological effects such as the level ofarousal may further influence the neurovascular re-sponse to the activation task in question.
These effects are hard to control and may substan-tially influence a single session’s results, such that theexperiment may ultimately say as much about thecontext under which the data were acquired as theeffects of the experimental manipulation itself. Al-though few researchers would expect a precise replica-tion of the results if an experiment were repeated, it iscurrently unclear how generalizable single-session re-sults are with fMRI.

709INTERSESSION VARIANCE IN fMRI
FIG. 1. Design matrices used for analysis. The design matrix is a graphical representation of the experimental model. Each column ofthe design matrix represents a separate regressor within the statistical model, and each row represents a single fMRI volume. The gray-scalecolor value within each cell displays the value of the relevant regressor at that point in the fMRI time series before the model fit is estimated.For example, a simple boxcar regressor, before convolution, would occupy a single design matrix column, and each cell within the columnwould have a value of 0 or 1, depending on the experimental design used. (A) A single session design matrix with the regressor of interest(the CBC, column 1), the session mean effect (gi, column 2), and the set of discrete cosine basis functions used to effect high-pass filtering(columns 3–8). (B) A multisession design matrix, constructed from n single-session design matrices, where n is the number of sessionsanalyzed at the multisession level. The design matrix in A was used only for single-session analyses, whereas B was used for both the fixed-
and the random-effects multisession analyses.
710 MCGONIGLE ET AL.
This influence of session context on the activationeffects of a study constitutes a session-by-condition in-teraction. Although a number of studies have exam-ined the reproducibility of fMRI across a small numberof sessions (Cohen et al., 1999; Noll et al., 1997; Rom-bouts et al., 1998; Tegeler et al., 1999; Yetkin et al.,1996), our primary aim was to examine how well asingle session typifies a subject’s responses. Just as thesignificance of within-session experimental effects isassessed by sampling a number of scans for each con-dition, to assess between-session differences one mustsample multiple sessions. If a single session is to be agood exemplar of a subject’s functional neuroanatomy,session-by-condition interactions must be minimal.
The issue of single-session generality also influencesdata analysis. If activation effects do indeed vary sub-stantially between sessions, to generalize the results tothe subject an experiment will need to utilize multi-ple sessions and assess the data accounting for bothwithin- and between-session variability. Typically,these two levels of variability are not addressed, even if
FIG. 2. Single-session sagittal MIPs for the motor paradigm. Thecollected, only 30 are shown here (sessions 17, 23, and 24 were rejeccorrected for multiple comparisons unless otherwise stated.
multiple sessions are acquired; the experimental ef-
fects of interest are assessed using statistical modelsthat utilize within-session error variance (residualscan/scan variability) as the only component of vari-ance. Although session-by-condition interactions areoften modeled, the variability of these interaction ef-fects does not enter into the inference. Such a model,employing a single variance component, is a fixed-ef-fects model (Searle et al., 1992). These models havebeen the norm in neuroimaging analysis and assessonly the average experimental effect across the ob-served sessions. They do not take account of the vari-ability of responses between sessions and thereforecannot be used to draw conclusions about a subject’stypical response. For example, a spuriously large acti-vation in one voxel during only one session may belarge enough to dominate that voxel’s average re-sponses across sessions. In the case of a single sessioncollected from a single subject, the experiment is re-duced to a case study. Conclusions regarding the sub-ject’s typical response can be made only under theimplicit assumption that intersession variability of re-
mber of each session is displayed below it. Although 33 sessions weredue to movement artifacts). All results are thresholded at P , 0.05
nuted
sponse would be negligible were the experimental ses-

d
711INTERSESSION VARIANCE IN fMRI
sion repeated. As discussed above, this is highly un-likely.
If session-by-condition interactions are substantial,random-effects models are required. Random-effectsmodels allow for multiple variance components (Searleet al., 1992), so the effects of each session on the BOLDresponse are treated as a random variable. This re-flects the fact that a single session is considered asample from the population of all possible sessionsfrom the subject, and so significance can be computed,accounting for both between- and within-session vari-ance. Random-effects analyses have previously beenemployed to account for between-subject variability, orsubject-by-condition interactions, in fMRI studies(Holmes et al., 1998; Henson et al., 1999a,b).
As the random-effect analysis infers about the pop-ulation from which the samples were drawn, the N ofobservations is now the number of sessions. As thenumber of sessions is quite small, these analyses tendto have low power, that is, there is a high chance oftype II errors. An analysis of this type, however, isessential for the correct level of inference if session-by-
FIG. 3. Single-session sagittal MIPs for the cognitive paradigmisplayed. Sessions marked with “*” contain no significant voxels.
condition interactions are considerable. In the present
study we examined the reproducibility of the BOLDresponse in a single subject over multiple sessions forsimple motor, cognitive, and visual paradigms. We firstpresent results from each session analyzed in isola-tion, as if from a single-session experiment, using onlywithin-session variance to compute significance. Wethen show where significant session-by-condition inter-actions occur for each of our activation paradigms.Finally, we consider two analyses of the entire multi-session data set. The first is a fixed-effects analysis, thesecond a simple random-effects analysis.
METHODS
Subject and Session Details
The subject was a healthy 23-year-old right-handedmale. As our goal in the current study was to examinethe generality of a single session, each session wasconducted as if it were the first time the subject hadbeen examined: in effect, as if only one session was tobe obtained. Our motivation was therefore to control
imilar to Fig. 2, although 33 sessions were collected, only 30 are
. Sfor obvious and artifactual between-session differences

w
712 MCGONIGLE ET AL.
while ensuring that sources of typical between-sessionvariability (scanner hardware and subject physiology)would be sampled in an unbiased manner. The follow-ing precautions were taken: the same operators alwayscontrolled the scanner, ambient light and sound levelswere similar between sessions, and spoken instruc-tions to the subject were always exactly the same. Oneobvious factor that we could not control was that oursubject was always aware that he had performed thetask before in the scanner, only under slightly differentcircumstances. We called this the “Groundhog Day”effect.
Ninety-nine individual sessions were acquired fromthe subject over a period of 2 months. Each scanningsession consisted of one run of a motor, cognitive, orvisual paradigm. To minimize scanning time, sessionswere acquired in blocks of three. Each block of threesessions comprised a motor, visual, and cognitive ses-sion. The order of sessions across scanning blocks was
FIG. 4. Single-session sagittal MIPs for the visual paradigm. Simith “*” contain no significant voxels.
randomized to balance any possible order effects. Ses-
sion paradigms were designed to reduce the effects ofvariable task performance. For example, the subjectwas familiarized with both the random number gener-ation and the finger-tapping task before performingthem in the scanner, in an attempt to eliminate per-formance effects. In addition, the rates at which bothtasks were performed were chosen to ensure that sub-ject performance would be stable across sessions.These decisions were informed by studies which usedsimilar paradigms (motor paradigm—Blinkenberg etal., 1996; cognitive paradigm—Jahanshahi et al., sub-mitted for publication).
Motor Paradigm
The subject tapped his right index finger, paced byan auditory tone (1.5 Hz). The subject’s hand was re-strained within a custom-built thermoplastic splint,which ensured that the amplitude of the finger move-
r to Figs. 2 and 3, only 31 sessions are displayed. Sessions marked
ilament was consistent both across and within sessions.

713INTERSESSION VARIANCE IN fMRI
TABLE 1
Area Cluster size (k)a Z scoreb
Talairach coordinates
X Y Z
(a) Local maxima for the motor fixed-effects model
Left precentral gyrus (SM1) 5958 9.77 238 210 52Left precentral gyrus (SM1) 9.63 262 220 38Left postcentral gyrus (SM1) 9.51 260 218 46
Right cerebellum, anterior lobe 1709 9.50 20 254 218Right cerebellum, anterior lobe 9.19 2 254 224Right cerebellum, anterior lobe 9.14 8 258 220
Left supplementary motor area (SMA) 927 9.39 22 22 52Right SMA 8.75 8 2 58Right SMA 8.50 4 8 66
Left inferior frontal gyrus 227 9.23 62 6 16Left inferior frontal gyrus 6.98 62 4 4
Right postcentral gyrus (SM1) 197 9.07 58 212 50Right precentral gyrus (SM1) 8.29 52 26 38Right precentral gyrus (SM1) 7.71 62 22 38
Left inferior thalamus 466 8.84 212 218 2Left ventral midbrain 6.15 28 212 214Left midbrain 5.90 28 220 212
Right lateral premotor cortex 70 8.80 38 28 50Right inferior frontal gyrus 62 8.60 64 0 16Right inferior parietal lobule 604 8.54 50 228 24Right transverse temporal gyrus 8.36 52 218 12
Right superior temporal gyrus 7.99 68 226 16Left posterior cingulate gyrus 100 8.32 28 226 42
Left posterior cingulate gyrus 7.11 22 222 48Right inferior frontal operculum 228 8.23 0 6 4
Right insula 6.12 0 6 10Left cerebellum, anterior lobe 66 7.85 20 256 220Right cerebellum, anterior lobe 98 7.73 2 258 248
Right cerebellar tonsil 7.43 6 270 240Left posterior postcentral gyrus 67 7.63 18 246 64Left superior frontal gyrus 79 7.49 28 42 26Left superior thalamus 46 7.40 18 210 18
Left thalamus 5.73 212 26 12Left medial frontal gyrus 223 7.27 238 58 22
Left medial frontal gyrus 7.27 230 62 4Left inferior frontal gyrus 7.20 234 54 28
Third ventricle 22 7.27 6 242 26Right putamen 171 7.26 8 6 22
Right putamen 6.68 0 6 212Right anterior cingulate gyrus 27 6.74 14 8 42Right postcentral gyrus 7 6.74 40 230 58Right thalamus 40 6.70 14 214 2Right inferior frontal gyrus 18 6.46 30 60 22White matter, subcortical 33 6.46 16 0 12
White matter, subcortical 5.13 12 6 2Right posterior cingulate 19 6.43 12 224 46Right cerebellum 18 5.94 48 250 238Right thalamus 16 5.79 20 0 26Left cerebellum 12 5.76 236 264 226Left superior frontal gyrus 5 5.64 228 28 56Right thalamus 9 5.55 8 24 24Right cerebellum 12 5.51 40 262 250Left superior frontal gyrus 8 5.42 222 38 48Right caudate nucleus 9 5.30 20 26 20
(b) Local maxima for the motor random-effects analysis
Left precentral gyrus (SM1) 4253 8.76 236 210 52Left precentral gyrus (SM1) 8.45 260 218 38Left frontal operculum 8.37 248 4 0
Right cerebellum, anterior lobe 951 8.04 18 252 218Right cerebellum, anterior lobe 7.55 30 252 226Right cerebellum, anterior lobe 7.34 36 262 224

msswMaf
(dtwa
cte
714 MCGONIGLE ET AL.
Each activation epoch was alternated with a rest ep-och, in which the pacing tone was delivered to controlfor auditory activation. Thirteen blocks were collectedper session (7 rest and 6 active). The subject main-tained fixation on a cross that was backprojected ontoa transparent screen by an LCD video projector. Theprojector was similarly employed to deliver visual in-structions to the subject before each block (either“Move” or “Rest”).
Cognitive Paradigm
The subject generated random numbers from 1 to 9,paced by an auditory tone (0.66 Hz). In the rest condi-tion the subject counted from 1 to 9, similarly paced bythe auditory tone. The subject fixated in a fashionsimilar to that used before. Thirteen epochs were col-lected in total (7 rest and 6 active).
Visual Paradigm
A reversing black and white checkerboard flickeringat 8 Hz (Fox and Raichle, 1985) was presented to thesubject. The subject focused on a central fixation spotthat was constant across both activation (reversing
TABLE 1
Area Cluster size (k)a
Left inferior frontal gyrus 151Left inferior precentral gyrus
Left supplementary motor area (SMA) 545SMARight superior SMA
Left thalamus 173Left midbrain (red nucleus)
Right postcentral gyrus (SM1) 104Right inferior precentral gyrusRight inferior precentral gyrus
Left supramarginal gyrus 44Right lateral premotor cortex 46Right frontal operculum 104Right parietal operculum 256
Right transverse temporal gyrusRight transverse temporal gyrus
Left posterior cingulate gyrus 39Right inferior frontal gyrus 32Right inferior postcentral gyrus 25Cerebellar vermis 25Left superior frontal gyrus 21Right cerebellar tonsil 8Left superior thalamus 12Left posterior postcentral gyrus 14Left cerebellum, anterior lobe 10Right putamen 20
Note. SM1, primary somatomotor cortex; SMA, supplementary moa Only clusters with k $ 5 are listed. Where multiple foci exist forb All foci reported survive a statistical threshold of P , 0.05 corre
checkerboard stimulation) and rest (fixation spot only) S
blocks. Six epochs were acquired in total (three activa-tion and three rest).
Scanning Parameters
The data were acquired on a Siemens MagnetomVision (Siemens, Erlangen, Germany) at 2 T. EachBOLD–EPI volume scan consisted of 48 transverseslices (in-plane matrix 64 3 64; voxel size 3 3 3 3 3
m; TE 5 40 ms; TR 5 4.1 s). Seventy-eight volumecans were collected during each cognitive and motoression, and 36 scans per visual session (epoch lengthas always 6 scans). A T1-weighted high-resolutionRI of the subject (1 3 1 3 1.5 mm resolution) was
cquired to facilitate anatomical localization of theunctional data.
Image Preprocessing
Data preprocessing was carried out using SPM99Wellcome Department of Cognitive Neurology, Lon-on, UK; http:/www.fil.ion.ucl.ac.uk/spm). All func-ional volumes, independent of session or paradigm,ere realigned to the first volume acquired (Friston etl., 1995) and a mean realigned volume was created.
ontinued
Z scoreb
Talairach coordinates
X Y Z
8.00 262 8 166.96 260 22 167.62 28 22 487.34 0 2 567.15 4 10 667.44 212 220 25.47 212 214 2107.40 58 212 506.93 50 24 365.75 60 22 407.27 258 244 187.19 38 28 527.10 50 6 47.07 50 228 266.94 48 218 146.16 68 226 166.88 28 224 446.71 64 0 166.56 62 212 126.22 6 270 2406.04 228 42 265.88 12 256 2485.81 220 212 185.73 218 246 645.68 220 256 2205.57 28 8 22
cortex.cluster, the three most significant are reported.d for multiple comparisons.
—C
tora
essions containing obvious movement artifacts (de-

715INTERSESSION VARIANCE IN fMRI
TABLE 2
Area Cluster size (k)a Z scoreb
Talairach coordinates
X Y Z
(a) Local maxima for the cognitive fixed-effects analysis
Right inferior parietal lobule 888 9.27 50 230 44Right inferior parietal lobule 8.63 34 244 44Right supramarginal gyrus 7.95 66 224 40
Left medial precentral gyrus (FEF) 1716 9.26 224 0 50Left lateral precentral gyrus 8.96 250 4 46Left middle frontal gyrus 8.51 254 10 26
Left precuneus 1812 9.17 210 266 44Left superior parietal lobule 8.86 242 236 50Left superior parietal lobule 8.77 218 262 64
Right medial precentral gyrus (FEF) 776 9.17 26 24 58Right superior frontal gyrus 8.70 20 6 60Right superior frontal gyrus 7.68 12 14 66
Right superior parietal lobule 876 8.99 20 266 58Right superior parietal lobule 8.74 26 270 50Right superior parietal lobule 8.42 22 258 62
Right anterior precentral gyrus 238 8.84 62 4 28Left middle frontal gyrus 454 8.63 234 36 22
Left middle frontal gyrus 8.22 248 40 24Left middle frontal gyrus 7.11 230 46 34
Right lateral premotor cortex 85 8.39 56 0 46Left supplementary motor area 421 8.30 22 18 50Right inferior frontal gyrus 406 8.30 56 16 22
Right inferior frontal gyrus 7.44 48 26 28Right superior temporal gyrus 7.44 234 16 6
Left inferior frontal gyrus 414 8.24 256 12 0Left inferior frontal gyrus 8.09 242 22 28Left insula 7.23 234 16 6
Right cerebellum, anterior lobe 190 8.00 30 262 230Right cerebellum, anterior lobe 7.32 34 254 236Right cerebellum, anterior lobe 5.86 40 260 236
Left tempero-occipital sulcus 41 7.96 252 252 218Right inferior parietal lobule 44 7.92 68 238 24
Right inferior parietal lobule 5.48 68 230 32Right inferior parietal lobule 5.33 64 244 34
Right middle frontal gyrus 311 7.91 44 40 32Right middle frontal gyrus 7.81 44 42 20Right middle frontal gyrus 7.80 34 36 18
Right anterior cingulate gyrus 37 7.75 12 12 38Left inferior parietal lobule 80 7.67 262 236 32Right middle temporal gyrus 72 7.65 58 220 26Right superior temporal gyrus 67 7.62 48 232 22Right middle occipital gyrus 27 7.21 50 274 12Left precentral gyrus 11 6.99 254 28 14Left middle frontal gyrus 28 6.99 230 56 26Left superior temporal gyrus 43 6.96 254 240 4Left inferior frontal gyrus 16 6.67 254 34 8Right cerebellum 16 6.58 36 238 242Right calcarine cortex (V1) 17 6.45 12 278 8Right insula 11 6.39 40 6 0Left cerebellum 30 6.36 222 256 234Left anterior cingulate 9 6.35 212 16 32Left inferior frontal gyrus 23 6.02 236 58 22
Left inferior frontal gyrus 5.53 232 62 4Right insula 11 5.90 42 16 4Left calcarine cortex (V1) 17 5.80 26 272 20Right middle frontal gyrus 7 5.77 40 30 24Left calcarine cortex (V1) 6 5.50 24 29 0Left hemisphere, white matter. 9 5.44 222 54 0Right superior frontal gyrus 5 5.24 16 38 54
L
L
L
R
R
R
L
716 MCGONIGLE ET AL.
cided by two of the authors with previous experience oftypical artifacts) were discarded at this stage: threemotor sessions, two visual sessions, and three cognitivesessions were excluded in this manner. The subject’sT1-weighted structural scan was coregistered to themean functional volume, and the mean volume used todetermine the parameters applied to all volumes dur-ing spatial normalization and resampling (Ashburner
TABLE 2
Area Cluster size (k)a
(b) Local maxima for the cog
eft superior parietal lobule 690Left posterior postcentral gyrusLeft superior parietal lobule
eft medial precentral gyrus (FEF) 1134Left middle frontal gyrusLeft ventral precentral gyrus
eft superior parietal lobule 156Left superior parietal lobuleLeft superior parietal gyrusight inferior parietal lobule 460Right inferior parietal lobuleRight supramarginal gyrusight superior parietal lobule 540Right superior parietal lobuleRight superior parietal lobuleight medial precentral gyrus (FEF) 569Right superior frontal gyrusRight superior frontal gyrus
eft precuneus 157Right anterior precentral gyrus 173Left middle frontal gyrus 276
Left middle frontal gyrusRight cerebellum, anterior lobe 78
Right cerebellum, anterior lobeRight inferior parietal lobule 22Right middle temporal gyrus 55Right lateral premotor cortex 41Left inferior frontal gyrus 139
Left inferior frontal gyrusRight inferior frontal gyrus 122Left temporal-occipital sulcus 18Left supplementary motor area (SMA) 201
Left supplementary motor area (SMA)Left supplementary motor area (SMA)
Right anterior cingulate gyrus 15Right middle frontal gyrus 98
Right middle frontal gyrusRight middle frontal gyrus
Left inferior parietal lobule 33Right middle frontal gyrus 19Right superior temporal gyrus 26Left middle frontal gyrus 7Right superior temporal gyrus 16Right inferior frontal gyrus 16Left superior frontal gyrus 7
Note. FEF, frontal eye fields; V1, primary visual cortex; SMA, supa,b See footnotes to Table 1.
et al., 1997; Ashburner and Friston, 1999) to a stan-
dard template (Evans et al., 1993). As the volume ofbrain sampled in each study was affected by the posi-tion of the subject within the scanner’s field of view, wefound that the extreme superior and inferior portionsof the subject’s brain were sparsely sampled. To ad-dress this, voxels not sampled in every session wereeliminated during normalization. All functional vol-umes were then smoothed with a FWHM Gaussian
ontinued
Z scoreb
Talairach coordinates
X Y Z
ive random-effects analysis
8.13 240 236 487.44 248 228 407.28 230 248 428.07 226 22 508.06 250 8 447.74 252 10 267.87 218 264 646.58 226 262 605.59 234 260 607.87 48 230 447.50 56 232 446.84 64 224 407.75 18 264 586.97 22 256 626.93 26 264 347.73 28 22 607.62 20 6 626.47 12 12 687.72 28 266 427.56 62 6 287.38 234 38 246.81 250 36 247.24 30 262 2306.56 32 256 2367.03 68 238 246.95 60 220 286.94 54 2 446.92 242 22 286.80 256 12 06.89 56 16 226.77 252 252 2186.71 22 18 486.66 26 10 546.64 0 16 586.66 14 12 386.53 38 38 345.93 44 40 205.66 34 44 386.26 264 238 266.26 34 36 186.01 46 230 245.98 230 54 285.90 54 18 2205.79 50 26 285.73 210 12 70
mentary motor area.
—C
nit
ple
kernel. Global changes in fMRI response from scan to
717INTERSESSION VARIANCE IN fMRI
TABLE 3
Area Cluster size (k) Z score
Talairach coordinates
X Y Z
(a) Local maxima for the visua fixed-effects analysis
Right calcarine cortex (V1) 11002 9.72 14 286 2Right calcarine cortex (V1) 9.63 6 274 24Left calcarine cortex (V1) 9.57 28 282 0
Right superior temporal gyrus 80 7.96 42 228 18Right precuneus 54 7.93 8 280 44Left lateral geniculate nucleus 91 7.92 218 226 24Right superior temporal gyrus 51 7.81 70 232 14Left inferior parietal lobule 152 7.79 232 238 54Left superior parietal lobule 130 7.75 228 254 54
Left superior parietal lobule 6.66 220 252 54Left superior parietal lobule 5.08 228 248 62
Left superior temporal gyrus 32 7.60 256 242 16Right lateral geniculate nucleus 83 7.59 24 224 4Right postcentral gyrus 95 7.52 36 232 54Left superior temporal gyrus 31 7.17 242 228 20Right lateral ventricle 112 6.82 20 226 24Left precuneus 25 6.76 218 262 48Right midbrain 27 6.71 8 226 26Right middle frontal gyrus 22 6.58 60 2 38Right inferior frontal gyrus 26 6.46 56 36 12
Right inferior frontal gyrus 5.50 52 42 16Left lateral ventricle 58 6.30 216 222 26Right precuneus 12 6.24 28 262 48Right cerebellum 18 6.18 12 270 246Right lateral ventricle 11 6.18 4 12 12Right parietal lobe, white matter 7 6.18 36 258 64Right supplementary motor area 8 5.50 2 2 64Right cerebellum 6 5.49 32 272 230Right fusiform gyrus 5 5.48 48 250 216Right temporal lobe, white matter 6 5.40 34 240 10Right superior parietal lobule 5 5.20 40 252 54
(b) Local maxima for the visual random-effects analysis
Right calcarine cortex (V1) 8766 8.66 10 280 28Right extrastriate cortex 8.59 24 296 16Right extrastriate cortex 8.54 22 276 218
Right precuneus 46 7.48 8 278 44Right parieto-occipital fissure 7.23 12 286 38
Right superior temporal gyrus 80 6.68 42 228 18Left superior parietal lobule 37 6.49 226 256 54Left lateral geniculate nucleus 27 6.21 216 228 24Left inferior parietal lobule 47 6.06 232 238 56Right postcentral gyrus 17 6.04 34 232 52Right lateral geniculate nucleus 21 5.98 22 226 0Right superior temporal gyrus 17 5.96 70 234 12Right inferior precentral gyrus 6 5.87 64 10 6Left intraparietal sulcus 13 5.86 220 270 34Left superior temporal gyrus 9 5.72 240 226 20Right intraparietal sulcus 26 5.71 32 270 26Left superior temporal gyrus 7 5.70 256 242 18Right middle frontal gyrus 6 5.57 58 4 38Right intraparietal sulcus 8 5.34 24 276 38Left inferior occipital gyrus 5 5.16 240 264 28
Note. V1, primary visual cortex.
l(w
ris
718 MCGONIGLE ET AL.
scan were removed by proportionally scaling each scanto have a common global mean voxel value.
Theory and Implementation
Statistical analysis was carried out using the generalinear framework described by Worsley and Friston1995). The sessions for each paradigm were modeledith a simple linear model for the data at each voxel:
Yij 5 gi 1 aif~ j! 1 OK bikgk~ j! 1 eij. (1)
FIG. 5. Multisession analyses of the motor paradigm, analyzedandom-effects model (c). Voxels surviving the statistical threshold adentification of activated areas. Each transverse slice is 2 mm thickcores having a brighter color.
k51
Here Yij denotes the value of the voxel of scan j ( j 51, . . . , J ) of session i (i 5 1, . . . , I ). gi is the mean(block) effect for session i. f( j) is a reference waveform,a function of the scan index within session which hasthe same form for all sessions. Here we shall use asimple “convolved boxcar” reference waveform (CBC),consisting of a boxcar function of zeros and ones rep-resenting the experimental time course, convolvedwith the expected hemodynamic response function.The parameter ai is the amplitude of the CBC responsefor session i. Differences in the session response am-
ng a fixed-effects model (a), extra-sum of squares F test (b), and adisplayed on a coregistered structural scan of the subject to aid thehe color bar represents statistical significance, with higher Z and F
usire. T
plitudes ai constitute session-by-condition interac-

719INTERSESSION VARIANCE IN fMRI
tions. The additional reference functions gk( j) are a setof discrete cosine basis functions, effecting a simple“high-pass” filter, as described by Holmes et al. (1997),with cutoff (specified by K) set at twice the experimen-tal period. We shall assume that this model fits, suchthat the residual errors (eij) have zero mean and exhibitonly short-term autocorrelation within session. In thefollowing we shall refer to the CBC amplitudes ai sim-ply as the response for session i.
Individual-Session Analyses
Each session was analyzed alone as a single fMRIsession, as if it were the only session acquired, using a
FIG. 5—
“standard” SPM analysis. The Groundhog Day effects
aside, this enables a comparison of how the results of asingle-session experiment can vary and illustrates whydrawing conclusions about a subject from a single ses-sion can be dangerous. The model used is that of Eq.(1), but considering only a single session (i) at a time.The residual errors are assumed to be Normally dis-tributed with variance si(e)
2 , estimated individually foreach session. Temporal autocorrelation was dealt withusing the method of Worsley and Friston (1995) bytemporally smoothing the session time series with aGaussian kernel of 6-s FWHM. The design matrix foreach session is illustrated in Fig. 1A. A t statisticassessing the null hypothesis of zero response (ai 5 0)
ntinued
Cowas constructed for each voxel, giving an SPM{t} for

HtNs
720 MCGONIGLE ET AL.
each session indicating the significance of the response.For display, each session-specific SPM{t} was trans-formed to an equivalent SPM{Z} by probability integraltransform. This was effected by replacing each t valuewith the standard Normal ordinate with the same up-per tail probability.
Multiple-Session Analyses—Session-by-ConditionInteractions
To assess whether there were significant session-by-condition interactions, we compared the model ofEq. (1) for all I sessions (design matrix shown in Fig.
FIG. 5—
1B) with a reduced model in which the response was
identical for all sessions (a i 5 a9, i 5 1, . . . , I ).ere we assume that the residual variance is iden-
ical across sessions, such that the residuals areormally distributed with zero mean and variancee2. The additional variance modeled by the full
model (including session-by-condition interactions)was compared with the residual variance using anextra sum-of-squares F test (Draper and Smith,1981), modified to account for temporally autocorre-lated residuals using the method of Worsley andFriston (1995). The resulting SPM{F} identifies vox-els that display significant session-by-condition in-
ntinued
Coteractions.

a
ArFt
ca
tn
M
amw
a
sis
ctmsissaWhim
pa
bTb
pas
721INTERSESSION VARIANCE IN fMRI
Multiple-Session Analyses
If there are substantial differences in response fromsession to session a single-session experiment is inad-equate if one wishes to examine a subject’s response toexperimental stimuli in general, and so a multiple-session experiment is necessitated.
Multiple-Session Analyses—Fixed-Effects Model
Given a multiple-session data set, modeled with Eq.(1) (design matrix shown in Fig. 1B), a fixed-effectsanalysis proceeds by assuming that the session-specificresponses ai themselves are of interest. The residualerrors eij are assumed Normally distributed with zeromean and constant variance se
2. Evidence of a responseacross sessions can be tested by examining a# •, theverage of the I session-specific responses
a# • 5 Oi51
I
ai .
gain, short-term temporal autocorrelation in the er-ors were handled using the method of Worsley andriston (1995), temporally smoothing each sessionime series with a Gaussian kernel of 6-s FWHM.
However, since the session-specific responses areonsidered fixed, only one component of variance isccounted for (the residual error variance se
2), and in-ference from the resulting SPM{t} is limited to theaverage response for the observed sessions. As such,his analysis is sensitive to large effects in a smallumber of sessions.
ultiple-Session Analyses—Random-Effects Model
To extend inference beyond the particular sessionscquired, we must recognize that these sessions areerely a sample of possible sessions, each of whichould have its own response ai. Thus, we regard the ai
of Eq. (1) as random effects, accepting that the responsemplitudes ai for the sessions under consideration are
merely one sample from the (hypothetical) distributionof response amplitudes for a session chosen at random.A simple second-level (between-session) model wouldbe
ai 5 a 1 ei, (2)
where the ai are from Eq. (1) (the within-sessionmodel), and the between-session errors ei have zeromean and variance sa
2 and can be considered indepen-dent. Thus, the random-effects model has two compo-nents of variance, between session, sa
2, and within ses-ion (residual), se
2. Using this model we can considernference regarding a, the underlying average re-
ponse across all possible sessions. cIn general, analysis of such random-effects modelsan be difficult (Searle and Casella, 1992). However,he simple models considered here are balanced (theodels for each session are exactly the same) and
eparable (the only common parameter across sessionss the intrasession (residual) variance se
2, assumed con-tant for all sessions). This permits a simple “summarytatistic” approach (Frison and Pocock, 1992). Such anpproach was first described for neuroimaging data byorsley et al. (1992), and its importance subsequently
ighlighted by Holmes et al. (1998), who describe themplementation (in SPM) used here. In essence, the
odel of Eq. (1) is fitted to yield estimates ai of theresponse amplitude ai at each voxel for each session.The variance of the estimated response amplitudes ai
across sessions incorporates both within- (se2) and be-
tween-session variability (sa2) in the appropriate pro-
ortions to assess the significance of the overall subjectctivation effect a (Frison and Pocock, 1992). Thus,
each session data set is surmised by a single contrastimage whose voxel values are the fitted response am-plitudes. These contrast images can then be assessedat the intersession level for a significant average effect,with inference extending to the subject in general (un-der similar experimental conditions) rather than justthe particular sessions acquired.
To conduct a parametric analysis, it remains tochoose a specific model for the between-session errorsei. In the absence of any evidence (yet) to suggestotherwise, consider a simple Normal model
ai 5 a 1 ei, ei , N~0, s a2!. (3)
Our approach here is pragmatic: we know nothingabout ei’s distribution. Our assumption of Normalityallows us to introduce random effects analyses simplyand logically as an extension of the parametric statis-tical tests used by SPM. We will discuss the validity ofthis assumption in the discussion.
With the models of Eqs. (1) and (3), the random-effects analysis can be effected as a simple one-samplet test on the contrast images, yielding an SPM{t}.
RESULTS
All tabular data referenced in the results section cane accessed at http://www.fil.ion.ucl.ac.uk/;davem.he coordinate system used is that of the space definedy Talairach and Tournoux (1988).
Individual Session Results
Figures 2, 3, and 4 show sagittal maximum intensityrojections (MIPs) per session for the motor, cognitive,nd visual tasks, respectively. Each SPM{Z} MIPhows voxels that survive a threshold of P , 0.05,
orrected for multiple comparisons.
T
s
722 MCGONIGLE ET AL.
It is immediately obvious that the pattern of acti-vated voxels varies widely between repeated singlesessions in our subject. While a grossly homogeneouspattern is evident across single-session MIPs of thesame paradigm, the spatial distribution of voxels ineach MIP is highly variable. Even though striking sim-ilarity is evident between certain data sets (e.g., visualsessions 10 and 12, Fig. 4), a large number of sessionsfrom all three paradigms display no significantly acti-vated voxels (e.g., visual sessions 4 and 30). The differ-ences are best exemplified by comparing the SPM{Z} ofmotor session 1 (Fig. 2), which contains 1076 voxels
FIG. 6. Multisession analyses of the cognitive paradigm, analyzerandom-effects model (c). Voxels surviving the statistical threshold aidentification of activated areas. Each transverse slice is 2 mm thickcores having a brighter color.
above threshold, and motor session 33, which contains
only 5. Results from the cognitive paradigm (Fig. 3) arebroadly similar: while the spatial distribution of voxelsbetween MIPs is more comparable than in the motorand visual paradigms, a large number of sessions con-tain no significantly activated voxels at the chosenthreshold.
MIPs are binary statistical images, in which voxelsare classified as “active” or “inactive” according to ac-cepted but arbitrary statistical thresholds (for discus-sions of this issue, see Poline et al., 1996; Genovese etal., 1997; Noll et al., 1997; Cohen and DuBois, 1999;
egeler et al., 1999). In any of the MIPs of Figs. 2, 3,
sing a fixed-effects model (a), extra-sum of squares F test (b), and adisplayed on a coregistered structural scan of the subject to aid thehe color bar represents statistical significance, with higher Z and F
d ure. T
and 4, a voxel i could have very different ai’s between

assoc
723INTERSESSION VARIANCE IN fMRI
sessions, yet still pass the threshold and appear to beconsistently activated.
Multiple-Session Analyses
Figures 5, 6, and 7 show the results of the motor,cognitive, and visual multiple-session analyses, respec-tively. As noted above, merely examining thresholdedstatistical maps is perhaps not the best way to examinesimilarities between sessions. Our use of the ESS-F testllowed us to examine which voxels showed statisticallyignificant variability across all sessions for our singleubject (Figs. 5b, 6b, and 7b). If a single session typifiesur subject’s response, there should be few session-by-ondition interactions, and thus the SPM{F} maps from
FIG. 6—
each analysis should display relatively few voxels.
By specifically examining the variability of session-by-condition interactions, we implicitly limit our analysis tovoxels that are activated on at least one session by thetask. Noise that has a truly random expression over timeis unlikely to be modeled sufficiently well by each ses-sion’s regressor of interest; however, task-correlatednoise, such as movement, will still present a problem.
ESS{F} Analyses
Figures 5b, 6b, and 7b show the results of eachmultisession ESS-F test. These SPMs were thresh-olded at P , 0.05 corrected as for the fixed-effectsSPM{Z}’s, reflecting that we did not have any a priorihypotheses concerning where we expected to see
ntinued
Cogreater variability. An important point to note at this

t
724 MCGONIGLE ET AL.
stage is that the ESS-F test is free of any constraintsabout the direction of activation effects observed. Assuch, although our main concern was to examine thevariability of activation effects, each SPM{F} also con-ains voxels that had highly variable deactivations. In
the interest of parsimony, these results will not bediscussed here.
Somewhat surprisingly, each fixed-effects SPM{Z}did not display a high degree of overlap with its corre-sponding SPM{F}. This is because each SPM{F} iden-tifies voxels that show high variability, even if they arenot classified as activated on average. Reflecting thisfact, the area displaying the highest degree of variabil-ity in signal intensity between sessions in the motor
FIG. 6—
paradigm (Fig. 5b) is located within the white matter of
the temporal lobe (228, 242, 228, F 5 7.88)—an areawhich does not appear on the fixed-effect SPM{Z} map(Fig. 5a). A similar area is observed in the cognitiveparadigm’s SPM{F} (238, 240, 6, F 5 7.40; Fig. 6b);again, this area is not present on the fixed-effectsSPM{Z} (Fig. 6a). There was some overlap betweenvoxels which displayed significant variability in eachSPM{F} and the corresponding fixed-effects SPM{Z}:for example, posterior SMA (22, 28, 52; F 5 4.67),ipsilateral cerebellum (26, 238, 222; F 5 5.68), andcontralateral precentral gyrus (226, 218, 70; F 54.66). These voxels were typically located at the edge ofa larger cluster of activated voxels. The variability seenmay reflect subtle differences in the areal extent of
ntinued
Coactivations at the periphery of large clusters.

5c22(avp(s
ase79sKpcw5
tade
aeva
R
obstvdwweae
725INTERSESSION VARIANCE IN fMRI
Fixed-Effects Analyses
The fixed-effects analyses of all three tasks (Figs.5a, 6a, and 7a) displayed areas of activation concor-dant with previous studies employing a similar task.A number of fMRI studies have used finger-tappingas a stereotypical motor task (e.g., Rao et al., 1993),and we found similar results (Table 1a), includingthe contralateral SM1 (Talairach coord. 238, 210,
2; Z score 5 9.77), the anterior lobe of the ipsilateralerebellum (20, 254, 218; Z 5 9.50), the SMA (22,2, 52; Z 5 9.39), the contralateral thalamus (212,18, 2; Z 5 8.84), and the ipsilateral premotor cortex
38, 28, 50; Z 5 8.80). It is notable that the SPM{Z}lso contains areas not previously reported as acti-ated by a simple externally paced finger-tappingaradigm, such as the right inferior parietal lobule50, 228, 24; Z 5 8.54). This is not surprising, as aingle a i of sufficient magnitude may be adequate for
any voxel to pass the average significance thresholdover sessions and so appear on the multisessionfixed-effects SPM{Z}. If the fixed-effects SPM{Z} isviewed in isolation, it is impossible to know if theseare “true” activated voxels which have not been re-ported in previous studies due to a lack of sensitivityor areas which display an activation effect signifi-cantly large to appear in the multisession fixed-ef-fects maps, yet are not consistently activated acrosssessions.
Similar patterns of results were observed uponinspection of the multisession fixed-effects SPM{Z}’sfrom the cognitive and visual paradigms (Figs. 6aand 7a). Although less is known about the functionalneuroanatomy of paced random-number generation,we found areas similar to a previous study (Table 2a;Jahanshahi et al., submitted for publication). In com-mon with Jahanshahi and colleagues, we found acti-vation in the anterior cingulate cortex, but againnoticed discrepancies between our results and theirs,e.g., our finding of bilateral calcarine cortex (12, 278,8, Z 5 6.45 and 26, 272, 20, Z 5 5.80) and left SMA
ctivation (22, 18, 50, Z 5 8.30). Similarly, our vi-ual paradigm activated, as expected, striate andxtrastriate areas around the calacarine sulcus (Fig.a, Table 3a), including bilateral V1 (14, 286, 2, Z 5.72 and 28, 282, 0, Z 5 9.57), in common withtudies employing a comparable stimulus (e.g.,wong et al., 1992). However, as with the otheraradigms, a number of areas not previously impli-ated in the functional neuroanatomy of this taskere activated (e.g., the right SMA: 2, 2, 64, Z 5.50).Clearly, these effects beg closer scrutiny. If we wish
o examine repeated trials of the same activation par-digm within a particular subject, it is necessary toefine variability within the same subject. The fixed-
ffects SPM{Z}’s tell us where voxels are active onverage across the observed sessions. If we wish toxamine the generality of a single fMRI session, theariability of each voxel across all sessions must beddressed.
andom-Effects Analyses
Figures 5c, 6c, and 7c show random-effects analysesf each multisession data set. These SPM{Z}’s haveeen weighted by both between-session and withinses-ion variances of each data set. Upon visual inspection,he random-effects SPM{Z}’s resemble a “cleaned-up”ersion of the fixed-effects SPM{Z}’s, and each para-igm’s pattern of results is now more in concordanceith previous studies. There are still, however, areasithin the random-effects SPM{Z} that one would notxpect, a priori, to be involved in the functional neuro-natomy of each task (Tables 1b, 2b, and 3b). Forxample, the motor random-effects SPM{Z} (Fig. 5c)
displays prominent bilateral auditory cortex activation(242, 228, 18, Z 5 8.14 and 48, 218, 14, Z 5 6.16). Wedid not expect this, as pacing tones were played duringboth rest and activation epochs during each motor ses-sion. This result may reflect attentional modulation ofauditory areas (Woodruff et al., 1996; Grady et al.,1997), as the tones’ salience was different between therest and the activation conditions. The neurobiologicalexplanation for this result need not concern us here: itis sufficient to recognize that we did not predict thispattern of activation. If we had access to only a singlesession from our subject, we would have been suspi-cious about their true nature. Even a multisessionfixed effects would not have helped: we would not beable to identify if the activation was driven by a smallnumber of sessions only or was indeed a true positive.This reasoning demonstrates that multiple scanningsessions analyzed with an appropriate statisticalmodel can reduce ambiguous interpretations.
The majority of voxels present in both the fixed-effects SPM{Z} and SPM{F} do not appear in the ran-dom-effects SPM{Z}’s. Properly accounting for be-tween-session variance means that these voxels nolonger survive a threshold of P , 0.05, corrected formultiple comparisons. This demonstrates that combin-ing multiple sampling of sessions with a statisticalmodel with more than one component of variance cor-rectly accounts for even small session-by-condition in-teractions.
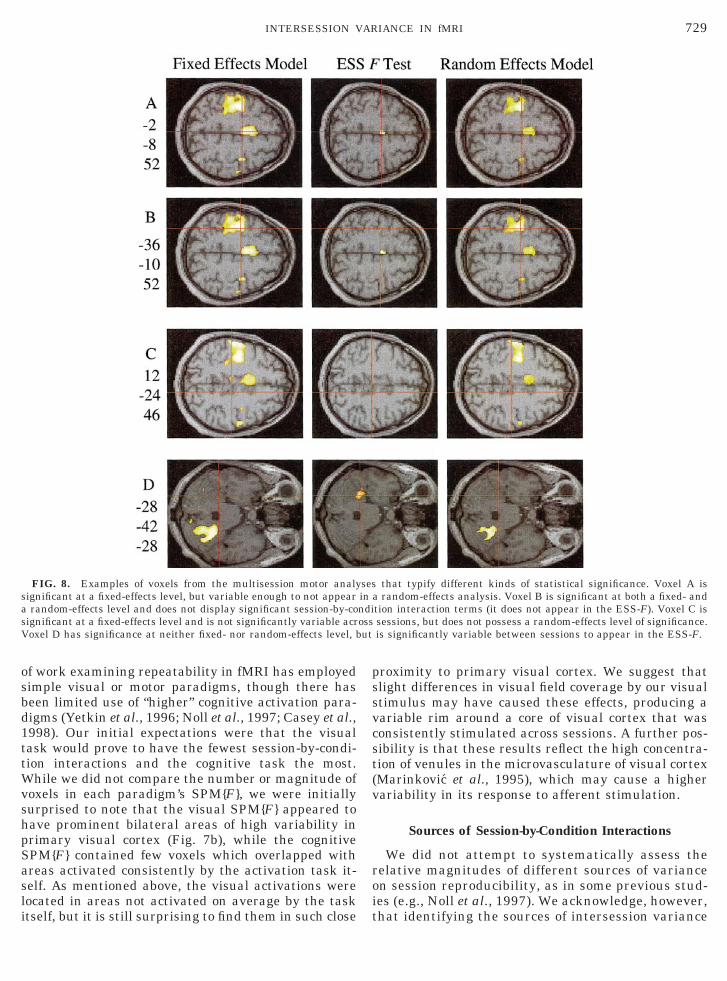
Figures 8 and 9 show voxels that typify differentpatterns of behavior across sessions, using the motorparadigm as an example. Figures 8A and 9A show avoxel in posterior SMA (22, 28, 252) which survives athreshold of P , 0.05, corrected for multiple compari-sons, in our multisession fixed-effects analysis. How-ever, this voxel displays significant session-by-condi-tion interactions (as seen by its appearance on the
ESS-F map) and thus fails to survive correction when a
ris
726 MCGONIGLE ET AL.
random-effects model is used. This voxel is an excellentexample of variability in “active” voxels. When oneexamines its parameter estimates by session (Fig. 9A),it is striking how stable it appears over some sessions(for example, sessions 15 to 18) and yet how variable itsbehavior is over all sessions. The histogram of param-eter estimates in Fig. 9A shows that although only onesession has a parameter estimate of greater than 1.5,this can still weigh the average activation effect overall sessions. When the variability of responses oversessions is addressed in the random-effects analysis,
FIG. 7. Multisession analyses of the visual paradigm, analyzedandom-effects model (c). Voxels surviving the statistical threshold adentification of activated areas. Each transverse slice is 2 mm thickcores having a brighter color.
the voxel loses significance.
The voxel in left primary motor cortex (236, 210, 52)displayed in Figs. 8B and 9B typifies voxels that sur-vive statistical thresholds in both fixed- and random-effects analyses. This voxel shows remarkably similarparameter estimates over all sessions (Fig. 9B). Thevoxel in Figs. 8C and 9C is one that, although notsignificantly variable (not shown on the ESS{F} map inFig. 8C), does not survive correction when a random-effects model is used.
Voxels within each SPM{F} can be thought of asbelonging to various classes: those which are not
ng a fixed-effects model (a), extra-sum of squares F test (b), and adisplayed on a coregistered structural scan of the subject to aid thehe color bar represents statistical significance, with higher Z and F
usire. T
activated by each paradigm, but display high vari-

a1
727INTERSESSION VARIANCE IN fMRI
ability of their parameter estimates (Figs. 8D and9D); “true” active or deactivated voxels, survivingboth fixed- and random-effects definitions of variabil-ity (Figs. 8B and 9B); voxels which are significant ata fixed-effects level but are significantly variable anddo not survive correction for between-session vari-ance (Figs. 8A and 9A); and voxels which, while notsurviving a random-effects analysis, are not signifi-cantly variable as defined by the ESS{F} map (Figs.8C and 9C).
DISCUSSION
The generality of any experimental result is an issue
FIG. 7—
which confronts all researchers, independent of exper-
imental discipline (Abelson, 1995). The results of anyisolated experiment are always open to contamination,and fMRI is no exception. As fMRI is an ideal experi-mental technique to examine questions that requireserial scanning sessions, there have been a number ofprevious studies that sought to examine the reproduc-ibility of fMRI data. Researchers have examined simi-lar activation paradigms across laboratories (Casey et
l., 1998), imaging modalities (Ojemann et al.,998),and sessions (Le et al., 1997; Noll et al., 1997;
Rombouts et al., 1998; Cohen et al., 1999). These stud-ies sought to characterize the reproducibility of fMRIdata and so tried to ensure that each session was
ntinued
Cocarried out similarly to those preceding it.

728 MCGONIGLE ET AL.
Examining the reproducibility of fMRI data is animportant question, but our own question was subtlydifferent: we chose to examine how well a single-session data set from a single subject typified thesubject’s response across multiple sessions, using avariety of activation paradigms. By examining thevariability in the magnitude of activation effectsacross a large number of sessions we accepted thateach session would be different. Indeed, it was ex-actly this between-session variability that we wishedto quantify.
We found that significant session-by-condition inter-actions occurred in each of the multisession data setsthat we examined, as illustrated by the respective ESS
FIG. 7—
SPM{F}’s. Our results are evidence of the influence of
session context on the results of any individual sessionand show the potential danger of drawing general con-clusions from an individual session analyzed in isola-tion with nothing known about reproducibility. If onesamples more sessions, each successive session ac-quired facilitates a better estimation of between-ses-sion variance, thereby increasing power to detect theunderlying response.
Differences in the Generality of DifferentActivation Paradigms
We chose to examine different activation paradigmsto ensure that the results of our study would not be
ntinued
Colimited to a single class of activation task. The majority
1ttWvshpSasli
v
roi
729INTERSESSION VARIANCE IN fMRI
of work examining repeatability in fMRI has employedsimple visual or motor paradigms, though there hasbeen limited use of “higher” cognitive activation para-digms (Yetkin et al., 1996; Noll et al., 1997; Casey et al.,998). Our initial expectations were that the visualask would prove to have the fewest session-by-condi-ion interactions and the cognitive task the most.
hile we did not compare the number or magnitude ofoxels in each paradigm’s SPM{F}, we were initiallyurprised to note that the visual SPM{F} appeared toave prominent bilateral areas of high variability inrimary visual cortex (Fig. 7b), while the cognitivePM{F} contained few voxels which overlapped withreas activated consistently by the activation task it-elf. As mentioned above, the visual activations wereocated in areas not activated on average by the task
FIG. 8. Examples of voxels from the multisession motor analysignificant at a fixed-effects level, but variable enough to not appeara random-effects level and does not display significant session-by-cosignificant at a fixed-effects level and is not significantly variable acrVoxel D has significance at neither fixed- nor random-effects level, b
tself, but it is still surprising to find them in such close t
proximity to primary visual cortex. We suggest thatslight differences in visual field coverage by our visualstimulus may have caused these effects, producing avariable rim around a core of visual cortex that wasconsistently stimulated across sessions. A further pos-sibility is that these results reflect the high concentra-tion of venules in the microvasculature of visual cortex(Marinkovic et al., 1995), which may cause a higherariability in its response to afferent stimulation.
Sources of Session-by-Condition Interactions
We did not attempt to systematically assess theelative magnitudes of different sources of variancen session reproducibility, as in some previous stud-es (e.g., Noll et al., 1997). We acknowledge, however,
that typify different kinds of statistical significance. Voxel A isa random-effects analysis. Voxel B is significant at both a fixed- andtion interaction terms (it does not appear in the ESS-F). Voxel C issessions, but does not possess a random-effects level of significance.is significantly variable between sessions to appear in the ESS-F.
sesinndiossut
hat identifying the sources of intersession variance

T
730 MCGONIGLE ET AL.
FIG. 9. Session-by-session plots of the parameter estimates (ai) and their standard deviations (vertical bars) of the voxels from Fig. 8.he histograms below each plot show the spread of values of a across all sessions.

731INTERSESSION VARIANCE IN fMRI
is important. For example, it is possible that spatialpreprocessing may affect intersession variance quiteindependent of underlying physical or physiologicalvariability. The realignment procedure used (Ash-burner et al., 1997) seeks to minimize the sum-of-squares differences between successive volumes anda reference (here, the first volume in the time series).It is a facile point that each paradigm induces inten-sity changes in voxels (i.e., it “activates” them), andso volumes acquired during the “on” period of eachparadigm will contain focal intensity differencesfrom volumes acquired during “rest.” As a successfulrealignment between two volumes relies on the vol-umes used being similar rigid bodies, differing onlyin their alignment in space, the paradigm-inducedintensity changes will affect the efficacy of alignmentand may ultimately raise intersession variance. Inaddition, similar effects in voxels lying at a tissueboundary (voxels in the walls of the ventricles, forexample) may result from simple repositioning of thesubject between sessions, causing session-specificpartial volume effects.
Examples like the above make it difficult to con-clude if the variability which we observe is attribut-able to differences in: (i) the scanning environment(e.g., position of subject within headcoil) or (ii) pre-processing (misalignment). We are more confidentthat we were able to minimize session-by-conditioninteractions that could be attributable to perfor-mance differences. We treated each successive scan-ning session as though it was the first time that oursubject had been scanned, to examine the potentialinfluence of session context on a single-session ex-periment (acknowledging that we cannot overcomethe Groundhog Day effect). It could be argued thatsystematic differences in our subject’s performanceacross sessions may have resulted in the session-by-condition interactions which we observed, as the re-peated execution of any active task or protocol ofsensory stimulation may result in habituation orlearning effects (e.g., Karni et al., 1995). We choseour activation tasks to minimize this possibility. Oursubject was pretrained on the motor task, and ourtask frequency was chosen to lie within a rangepreviously demonstrated by Blinkenberg and col-leagues (1996) to have a low error rate (between 1and 2 Hz). Similarly, we chose a stable rate of num-ber generation for our cognitive task (informed bythe results of Jahanshahi et al., submitted for pub-lication). Although we did not record subject perfor-mance on this task (primarily because of the motionit would produce), performance at random numbergeneration remains stable over a number of repeti-tions (Evans et al., 1980). Furthermore, a prelimi-nary MANCOVA analysis of our motor data set ex-amining the effects of session revealed no systematic
expression of the experimental variance over subse-quent sessions (data not shown). Although we acceptthat learning effects may exhibit complex temporaldynamics, the structure of our parameter estimatesover sessions suggests random variation (Fig. 8)around a “true” mean parameter estimate. However,without independent measures of task performance,we cannot entirely rule out between-session habitu-ation or learning-related changes in activation.
Stability of fMRI Results acrossSessions—Consequences for Longitudinal Studies
As noted previously, fMRI is ideally suited to theexamination of learning or recovery-of-function stud-ies. These studies are typically predicated on the as-sumption that the experimental effects will be largeenough to ensure their detection compared to nonspe-cific between-session effects. Because of the consider-able time that must typically be devoted to such stud-ies, it would be useful to have some idea of the relativemagnitudes of each effect before beginning. Althoughin the present study we have examined the similarityof the results between sessions while accepting a cer-tain degree of difference in the practical implementa-tion of each session, our results do not really addressthe issue of signal to noise in longitudinal fMRI stud-ies. As any difference between sessions is a session-by-condition interaction, any study which purports to fo-cus on session-by-condition interactions produced bythe experimental manipulation must ensure that non-specific session-by-condition interactions can be effi-ciently controlled (for a discussion of these issues, seePetersson et al., 1999). We examined three tasks thatwere designed to show limited session-by-condition in-teractions in subject performance. As such, our resultscannot be used to address the validity of longitudinalfMRI studies. We note, however, that the stability ofour results suggests that longitudinal studies that pro-duce unambiguous results should be feasible.
Use of Thresholded Statistical Mapsto Analyze Session Generality
Typically, the results of neuroimaging experimentsare displayed using binarized statistical maps. In thisfashion, voxels that pass a predetermined statisticalthreshold are classified as active and other voxels asinactive. Although the utility and clarity of the resultsmotivate this approach, much of the richness of func-tional neuroimaging data sets is removed. Attempts toexamine the test–retest reliability of fMRI using mea-sures such as “voxel counting” on thresholded mapstherefore suffer from two problems: an essentially ar-bitrarily defined statistical threshold and the loss ofcomplexity which accompanies any method that has toclassify voxels as either active or inactive. We sought,instead, to characterize our data sets in terms of the
between-session variance of the activation effects and
trF
st
732 MCGONIGLE ET AL.
not merely examine when voxels passed an arbitrarilyset threshold on successive sessions. The differencesbetween the two approaches are apparent when onecompares the results of our single-session analyses(Figs. 2, 3, and 4) with our later multisession analyses(Figs. 5, 6, and 7). Generally a failure to detect activa-tion may say more about the sensitivity of the experi-ment than the presence of the effect itself (Poline et al.,1996). Certain areas may therefore appear more vari-able than they truly are.
Effects of Sample Size on the Analysis of Generality
Our results demonstrate the need for a large samplesize when examining how well a single fMRI sessionexemplifies a subject’s responses. The plots of param-eter estimates by session in Fig. 9 show that voxels inwhich we found significant session-by-condition inter-actions over all of our sessions appear surprisinglystable when examined over a small number of sessions(for example, sessions 27–29 in Fig. 9C are almostidentical). This effect has been termed “the law of smallnumbers” (Tversky et al., 1971)—the tendency to as-cribe a lack of variability to small sample groups. Ouruse of a large number of sessions allowed us to char-acterize variability that may have been missed by pre-vious studies employing five repeated sessions at moston the same subject. However, the opposite argumentmay be leveled at our results: if the sample size is largeenough, then a statistically significant difference willalways be found—this is merely an example of thefallacy of classical hypothesis testing. We accept thiscriticism, but believe that an analysis of 30 sessions isan appropriate sample size for the purposes of thisstudy. The existence of significantly variable voxelsnecessitates the use of a random-effects model to allowthe experiments to truly generalize their results to thesubject.
Levels of Inference Arising from Fixed- andRandom-Effects Models
Worlsey and colleagues (1992) first suggested the useof a “summary statistic” approach to the analysis offunctional neuroimaging data. However, the imple-mentation we used in the current study is that ofHolmes and Friston (1998), who suggested random-effects analyses for balanced designs in neuroimagingemploying a general linear framework to allow for thebetween-subject variance component in multisubjectdesigns. As discussed previously, the random-effectsanalysis confers generality, but with a concomitantloss of sensitivity due to the inevitable low degrees offreedom. We assumed that the ei were Normally dis-ributed and incorporated this assumption into ourandom-effects level model. However, by examiningig. 9 it is clear that the ei do not necessarily conform
to this distribution. If we examine the case of the voxel
in Fig. 9A, it is clear that this voxel has a skewed-rightdistribution. Indeed, if one asks a simpler question ofthe voxel in Fig. 9A (how often is a . 0) and employ aimple sign test, the probability of getting 31/33 posi-ive a’s is ,7 3 1028. Yet this voxel does not pass the
random-effects analysis used here. Although we acceptthat this is only one voxel, it casts doubts on assump-tions of Normality for the eI, and it is clear that furtherinvestigation is needed into the distribution of be-tween-session variance. The development of random-effects models that do not require prior assumptions ofthe distribution of residuals may be needed to addressthis issue.
The use of random-effects models in the analysis offMRI data is a recent addition to the canon of neuro-imaging analysis methods, and it is wise to note aprevious adoption of this measure. In the analysis ofbehavioral data from human subjects, Clark’s (1973)initial proposal that the model should be used morefrequently highlighted an obvious problem: treating asample from a population as random and selecting asample randomly from a population are clearly not thesame. Although we could argue that by using a ran-dom-effects analysis in our study, we can generalizeour results to our subject as a putative population, wehave performed a very limited sampling of our subject’sresponses. Each scanning session was performed overa 2-month period only, and session times were selectedin a biased manner: near midday and near 6 PM in theevening. However, it is not elegant to have to state that“our results generalize to the population of possiblesessions sampled from our subject over a period of 2months, using the resources available in our labora-tory.” In practice, these caveats are usually accepted.Indeed, the use of random-effects models to ensure thecorrect level of inference in multisubject fMRI analysesrarely addresses the other sources of systematic vari-ation in the population that the investigators are gen-eralizing to (usually male, Caucasian right-handerswho respond to advertisements and financial reward).However, adopting a random-effects model does affordsome protection against inappropriate generalizationof results, as noted by Abelson (1995).
Although we have shown that with an appropriatestatistical model and a large sample of sessions we canobtain robust results, a number of issues remain un-answered. In particular, we would hesitate before gen-eralizing our own results to other centers, subjects, oractivation paradigms, as between-session variancemay vary greatly depending on the context underwhich it is studied.
CONCLUSIONS
In this paper we described the results of an experi-ment designed to examine intersession variance in
fMRI during the performance of simple visual, motor,
733INTERSESSION VARIANCE IN fMRI
and cognitive tasks by a single subject. First, analyzingour data session by session, we suggested that bina-rized statistical maps, though convenient, are not auseful tool for the evaluation of intersession variabil-ity. We then described an analytical framework thatallowed us to identify significantly variable voxels bysession across our multisession data sets. Each multi-session data set, by paradigm, showed evidence of sig-nificant session-by-condition interactions. This resultdemonstrates that session context effects have a sig-nificant effect on fMRI data and illustrates that a sin-gle session should be considered merely as a singlesample of a subject’s responses to the experimentalintervention employed. As we sampled a large numberof sessions across all paradigms, we then compared thedifferences between analyzing these data using eitherfixed- or random-effects linear models, the latter beinga recent addition to neuroimaging analysis. Althoughwe comment on the usefulness of random effects anal-yses, which allow inference about experimental effectsto be extended to the population which the sessionswere sampled from, we draw attention to the interses-sion distribution of voxel response amplitudes. Ourassumption of Normally distributed intersession resid-uals was not supported by close examination of some ofour data, and so we accept that future work is requiredbefore random-effects models can be used to their fullpotential. Finally, we acknowledge that identifying thesource and magnitude of the different sources of inter-session variance in fMRI is crucial. The ability to dif-ferentiate between variability caused by the neurovas-cular signals that fMRI measures, and variabilityintroduced by the means of measurement and analysisof these signals, is essential for the future of fMRI as anoninvasive imaging modality.
ACKNOWLEDGMENTS
We thank Jesper Andersson and John Ashburner for invaluablehelp and advice and our reviewers for their constructive and gener-ous comments. The Wellcome Trust supported this work.
REFERENCES
Abelson, R. P. 1995. Statistics as Principled Argument. Elbaum,Hillsdale, NJ.
Ashburner, J., and Friston, K. J. 1999. Nonlinear spatial normalisa-tion using basis functions. Hum. Brain Mapp. 7:254–266.
Ashburner, J., Neelin, P., Collins, D. L., Evans, A. C., and Friston,K. J. 1997. Incorporating prior knowledge into image registration.NeuroImage 6:344–352.
Belliveau, J. W., Kennedy, D. N., McKinstry, R. C., Buchbinder,B. R., Weisskoff, R. M., Cohen, M. S., Vevea, J. M., Brady, T. J.,and Rosen, B. R. 1991. Functional mapping of the human visual-cortex by magnetic-resonance-imaging. Science 254:716–719.
Blinkenberg, M., Bonde, C., Holm, S., Svarer, C., Andersen, J.,Paulson, O. B., and Law, I. 1996. Rate dependence of regionalcerebral activation during performance of a repetitive motor task:
A PET study. J. Cereb. Blood Flow Metab. 16:794–803.Casey, B. J., Cohen, J. D., O’Craven, K., Davidson, R. J., Irwin, W.,Nelson, C. A., Noll, D. C., Hu, X., Lowe, M. J., Rosen, B. R.,Truwitt, C. L., and Turski, P. A. 1998. Reproducibility of fMRIresults across four institutions using a spatial working memorytask. NeuroImage 8:249–261.
Clark, H. H. 1973. The language-as-fixed-effect fallacy. J. Verb.Learn. Verb. Behav. 12:335–359.
Cohen, M. S., and DuBois, R. M. 1999. Stability, repeatability, andthe expression of signal magnitude in functional magnetic reso-nance imaging. J. Magn. Reson. Imag. 10:33–40.
Draper, N. R., and Smith, H. 1981. Applied Regression Analysis, pp.97–98. Wiley, New York.
Evans, A. C., Collins, D. L., Mills, S. R., Brown, E. D., Kelly, R. L.,and Peters, T. M. 1993. 3D statistical neuroanatomical modelsfrom 305 MRI volumes. In Proceedings, IEEE Nuclear ScienceSymposium and Medical Imaging Conference, pp. 1813–1817.IEEE Inc., Piscataway, NJ.
Evans, F. J., and Graham, C. 1980. Subjective random numbergeneration and attention deployment during acquisition and over-learning of a motor skill. Bull. Psychonom. Soc. 15:391–394.
Fox, P. T., and Raichle, M. E. 1985. Stimulus rate determines re-gional blood flow in striate cortex demonstrated by positron emis-sion tomography. Ann. Neurol. 17:303–305.
Frison, L., and Pocock, S. J. 1992. Repeated measures in clinicaltrials: An analysis using mean summary statistics and its impli-cations for design. Stat. Med. 11:1685–1704.
Friston, K. J., Ashburner, J., Frith, C. D., Poline, J.-B., Heather,J. D., and Frackowiak, R. S. J. 1995. Spatial registration andnormalisation of images. Hum. Brain Mapp. 2:165–189.
Genovese, C. R., Noll, D. C., and Eddy, W. F. 1997. Estimatingtest–retest reliability in functional MR imaging. I. Statisticalmethodology. Magn. Res. Med. 38:497–507.
Grady, C. L., Van Meter, J. W., Maisog, J. M., Pietrini, P., Krasuski,J., and Rauschecker, J. P. 1997. Attention-related modulation ofactivity in primary and secondary auditory cortex. NeuroReport28:2511–2516.
Henson, R. N. A., Shallice, T., and Dolan, R. J. 1999a. Right prefron-tal cortex and episodic memory retrieval: A functional MRI test ofthe monitoring hypothesis. Brain 122:1367–1381.
Henson, R. N. A., Rugg, M. D., Shallice, T., Josephs, O., and Dolan,R. J. 1999b. Recollection and familiarity in recognition memory:An event-related functional magnetic resonance imaging study.J. Neurosci. 19:3962–3972.
Holmes, A. P., Josephs, O., Buchel, C., and Friston, K. J. 1997.Statistical modeling of low frequency confounds in fMRI. Neuro-Image 5:S480.
Holmes, A. P., and Friston, K. J. 1998. Generalisability, randomeffects and population inference. NeuroImage 7:S754.
Howseman, A. M., McGonigle, D. J., Grootoonk, S., Ramdeen, J.,Athwal, B. S., and Turner, R. 1998. Assessment of the variabilityin fMRI data sets due to subject positioning and calibration of theMRI scanner. NeuroImage 7:S599.
Karni, A., Meyer, G., Jezzard, P., Adams, M. M., Turner, R., andUngerleider, L. G. 1995. Functional MRI evidence for adult motorcortex plasticity during motor skill learning. Nature 377:155–158.
Kwong, K. K., Belliveau, J. W., Chesler, D. A., Goldberg, I. E.,Weisskoff, R. M., Poncelet, B. P., Kennedy, D. N., Hoppel, B. E.,Cohen, M. S., Turner, R., Cheng, H. M., Brady, T. J., and Rosen,B. R. 1992. Dynamic magnetic resonance imaging of human brainactivity during primary sensory stimulation. Proc. Natl. Acad. Sci.USA 89:5675–5679.
Le, T. H., and Hu, X. 1997. Methods for assessing accuracy and
reliability in functional MRI. NMR Biomed. 10:160–164.
734 MCGONIGLE ET AL.
Marinkovic, R., Cvejin, B., Markovic, L., and Budimlija, Z. 1995.[Morphologic characteristics of the vascular network in the striatearea in humans]. Med. Pregl. 48:7–9.
Noll, D. C., Genovese, C. R., Nystrom, L. E., Vazquez, A. L., Forman,S. D., Eddy, W. F., and Cohen, J. D. 1997. Estimating test–retestreliability in functional MR imaging. II. Application to motor andcognitive activation studies. Magn. Res. Med. 38:508–517.
Ogawa, S., Menon, R. S., Tank, D. W., Kim, S. G., Merkle, H.,Ellerman, J. M., and Urgubil, K. 1992. Intrinsic signal changesaccompanying sensory stimulation: Functional brain mappingwith magnetic resonance imaging. Proc. Natl. Acad. Sci. USA89:5951–5955.
Ogawa, S., Menon, R. S., Kim, S.-G., and Ugurbil, K. 1998. On thecharacteristics of functional magnetic resonance imaging of thebrain. Annu. Rev. Biophys. Biomol. Struct. 27:447–474.
Ojemmaan, J. G., Buckner, R. L., Akbudak, E., Snyder, A. Z.,Ollinger, J. M., McKinstry, R. C., Rosen, B. R., Petersen, S. E.,Raichle, M. E., and Conturo, T. E. 1998. Functional MRI studies ofword-stem completion: Reliability across laboratories and compar-ison to blood flow imaging with PET. Hum. Brain Mapp. 6:203–215.
Petersson, K. M., Elfgren, C., and Ingvar, M. 1999. Learning-relatedeffects and functional neuroimaging. Hum. Brain Mapp. 7:234–243.
Poline, J. B., Vandenberghe, R., Holmes, A. P., Friston, K. J., andFrackowiak, R. S. J. 1996. Reproducibility of PET activation stud-ies: Lessons from a multi-center European experiment. NeuroIm-age 4:34–54.
Rao, S. M., Binder, J. R., Bandettini, P. A., Hammeke, T. A., Yetkin,
F. Z., Jesmanowicz, A., Lisk, L. M., Morris, G. L., Mueller, W. M.,Estkowski, L. D., Wong, E. C., Haughton, V. M., and Hyde, J. S.1993. Functional magnetic resonance imaging of complex humanmovements. Neurology 43:2311–2318.
Rombouts, S. A. R. B., Barkhof, F., Hoogenraad, F. G. C., Sprenger,M., and Scheltens, P. 1998. Within-subject reproducibility of visualactivation patterns with functional magnetic resonance imagingusing multislice echo planar imaging. Magn. Reson. Imag. 16:105–113.
Searle, S. R., Casella, G., and McCulloch, C. E. 1992. VarianceComponents. Wiley, New York.
Talairach, P., and Tournoux, J. 1988. A Stereotactic Coplanar Atlasof the Human Brain. Thieme, Stuttgart.
Tegeler, C., Strother, S. C., Anderson, J. R., and Kim, S. G. 1999.Reproducibility of BOLD-based functional MRI obtained at 4T.Hum. Brain Mapp. 7:267–283.
Tversky, A., and Kahneman, D. 1971. Belief in the ‘law of smallnumbers.’ Psychol. Bull. 75:105–110.
Woodruff, P. W. R., Benson, R. R., Bandettini, P. A., Kwong, K. K.,Howard, R. J., Talavage, T., Belliveau, J., and Rosen, B. R. 1996.Modulation of auditory and visual cortex by selective attention ismodality-dependent. NeuroReport 7:1909–1913.
Worsley, K. J., Evans, A. C., Marrett, S., and Neelin, P. 1992. Athree-dimensional statistical analysis for CBF activation studiesin human brain. J. Cereb. Blood Flow Metab. 12:900–918.
Worsley, K. J., and Friston, K. J. 1995. Analysis of fMRI timeseriesrevisited—Again. NeuroImage 2:173–181.
Yetkin, F. Z., McAuliffe, T. L., Cox, R., and Haughton, V. M. 1996.Test–retest precision of functional MR in sensory and motor task
activation. AJNR 17:195–198.