Embed Size (px)
Citation preview

Journal of Cellular Biochemistry 103:920–930 (2008)
Wound Repair and Proliferation of BronchialEpithelial Cells Regulated by CTNNAL1
Yang Xiang,1 Yu-Rong Tan,1 Jian-Song Zhang,2 Xiao-Qun Qin,1* Bi-Bo Hu,3 Yue Wang,1
Fei Qu,1 and Hui-Jun Liu1
1Xiangya School of Medicine, Central South University, Changsha 410078, Hunan, PR China2Hunan Normal University, Changsha 410013, Hunan, PR China3Xiangtan central hospital, Xiangtan 411100, Hunan, PR China
Abstract Adhesion molecules play vital roles in airway hyperresponsiveness (AHR) or airway inflammation. Ourprevious study indicated that adhesion molecule catenin alpha-like 1 (CTNNAL1) is relevant closely to asthmasusceptibility, but its biological function or significance is still unclear. In the present study,we observed the temporal andspatial distribution of CTNNAL1 expression in mouse lung tissue with the OVA-sensitized asthma model and found thatthe level of CTNNAL1 mRNA showed a prominent negative correlation with pulmonary resistance (RL). To study thefunction of CTNNAL1 in airway, effects of CTNNAL1 on proliferation and wound repair activity of human bronchialepithelial cells (HBEC) was investigated with antisense oligonucleotide (ASO) technique. The results showed that:(1) CTNNAL1 ASO could decelerate the repairing velocity and proliferation of HBEC; (2) CTNNAL1 expression wasincreased on the edge cells of mechanic wounded area in culture; (3) extracellular matrix component fibronectin (Fn)obviously promoted wound repair activity and proliferation of HBEC, which could be blocked by CTNNAL1 ASO;(4) Western blot showed that Fn could promote FAK phosphorylation, which also be inhibited by CTNNAL1 ASO. Inconclusion, the level of CTNNAL1mRNA expression is highly correlated to airway resistance; CTNNAL1may contributeto thewound repair andproliferationofHBEC. Furthermore, itmay serve to Fnmediatedcell-extracellular adhesion and itssignal transduction. J. Cell. Biochem. 103: 920–930, 2008. � 2007 Wiley-Liss, Inc.
Key words: CTNNAL1; bronchial epithelial cells (BEC); asthma
In airway hyperresponsive diseases such asasthma, the bronchial epithelium is frequentlydesquamated, leaving areas of denuded base-ment membrane, under which submucosalterminal sensory nerve are directly exposed toinhaled air and the sensitivity of sensory nerveincrease aberrantly [Montefort et al., 1992]. Theepithelium both provides a physical barrier
to the external environment and regulatesseveral metabolic functions of airways, includ-ing fluid and ion transport to the airway lumen,mucociliary clearance, and airway diameter.Reconstitution of a functional epithelia barrierrequires an ordered sequence of events, includ-ing cell spreading and migration, adhesion,production of provisional matrix, proliferation,and differentiation into needed epithelial cellsubtypes. Each of these processes may dependon adequate expression and distribution ofadhesive molecules on epithelial cells. HBECexpress four kind of adhesive molecules, includ-ing cadherin, integrin, ICAM-1, and selectin.In normal state, HBEC express intergrinsand cadherins, which mediate and maintainintercellular homologous and heterogeneousadhesions. In injury state, HBEC can arrestleukocytes, evoke an inflammation reaction inairway by expressing selectins and intercellularadhesive molecule-1 (ICAM-1) [Campbell et al.,1994].
� 2007 Wiley-Liss, Inc.
Yang Xiang and Yu-Rong Tan contributed equally to thiswork.
Grant sponsor: National Natural Science Foundationof China; Grant number: #30270586; Grant sponsor:Natural Science Foundation of Hunan; Grant numbers:#05JJ30049, #03JJ3015.
*Correspondence to: Xiao-Qun Qin, Xiangya School ofMedicine, Central South University, Changsha 410078,Hunan, PR China. E-mail: [email protected]
Received 14 April 2007; Accepted 4 June 2007
DOI 10.1002/jcb.21461

Since the adhesionmolecule family has manytypes with diverse functions, the issue of whathomeostasis is maintained with regard to cells’constitutive adhesion and the inflammationadhesion mechanism is far from being clear.Therefore, we hypothesized that the defect instructures and functions of airway epitheliummay be the initiator of AHR. The imbalanceof the adhesion molecules expression maycontribute to the structural defect or functionaldisorder on airway epithelium and therefore beassociated with asthma pathogenicity. In orderto confirm the hypothesis, this present studyused a cDNA microarray to screen the differ-ential expression of adhesion molecules inhuman peripheral blood leukocytes, and foundthat an alpha-catenin-related protein, cateninalpha-like 1 (CTNNAL1) was downregulated inasthma patients, which was further confirmedby Real time PCR (These results will be publi-shed in an other article). However, the expres-sion of CTNNAL1 in airway and the functionassociated with airway epithelium are remai-ned unknown.CTNNAL1wasfirst characterizedas a2.45-kb
transcript that was down-regulated in humanpancreatic cancer cells [Zhang et al., 1998].CTNNAL1 shows extensive homology to a-catenin, a vinculin family member that plays acrucial role in cell differentiation and cancerinvasion. The human CTNNAL1 gene maps tochromosome 9q31–q32, a region where frequentalterations were observed in several types oftumors including bladder, ovarian, and esoph-ageal cancer. CTNNAL1 mRNA is expressedubiquitously, although the levels are lower inneural tissues [Janssens et al., 1999]. Eventhough a functional role has not been assignedto CTNNAL1, its extensive homology to a-catenin,and its chromosomal localization make it anintriguing candidate for further study relativeto cell differentiation, growth and motility.As of now, there have no papers published
about the expression of CTNNAL1 in adultlungs andwhether it is involved inwound repairor cell proliferation during airway inflam-mation or airway hyperresponsiveness (AHR).The aim of this study is to investigate thetemporal and spatial distribution of CTNNAL1mRNAinawell-characterizedOvalbumin (OVA)-sensitized Balb/c mice asthmamodel. CTNNAL1antisense oligonucleotide (ASO) was used toobserve the effect of CTNNAL1 on the repairand proliferation of HBEC.
CTNNAL1 is a member of the vinculin super-family of proteins, which includes both a-catenin and vinculin. Vinculin is present bothin cadherin–catenin adhesion complexes andin focal adhesion complexes that mediate cell-extracellular matrix adhesion. As vinculinproteins are linked via talin to integrins, whichare transmembrane receptors for the extracel-lular matrix such as fibronectin (Fn) [Weisset al., 1998], and the linkage of Fn receptorproteins to the actin cytoskeleton are requiredfor proper tissue differentiation and homeo-stasis, we also detect the effect of CTNNAL1,a member of vinculin family, on Fn mediatedwound repair, cell proliferation, and focal adhe-sion kinase phosphorylation.
MATERIALS AND METHODS
Animal Model of Asthma
Balb/c mice were obtained from the experi-mental animal center, XiangYaMedical College,Central SouthUniversity. All mice were housedunder specific pathogen-free conditions and hadad libitum access to food and water.
Asthma models were induced by intraperito-neal injection with 50 mg of OVA (grade V;Sigma, St. Louis, MO) and 1 mg aluminumhydroxide (EM Industries, Hawthorne, NY)on day 0, and then boost with a second injection7–10 days later. On day 21, we challenge theanimals via the intratracheal route with 50 mgof antigen in 30 ml aliquots. We repeat thechallenge on days 22 and 23. This schedule anddose pattern allows optimal responses in bothinflammatory infiltrate and AHR [Isenberg-Feig et al., 2003; Nikolaidis et al., 2003]. Thirtymice were randomly divided into five experi-ment groups and one control group. Controlmice were sensitized or challenged with saline.Airway parameters measurement and lungtissues collection were performed on days 24,26, 30, 32 and 34, corresponding to the 1, 2, 4, 6,and 8 days after the last challenge, respectively.
Measurement of AirwayResponsiveness In Vivo
Airway responsiveness was measured usingbarometric whole-body plethysmography byrecording airflow and respiratory pressure cur-ves (Buxco; EMKATechnologies, Paris, France)in response to inhaled histamine (Sigma).
Function Research on CTNNAL1 921

Airway responsiveness was expressed in pul-monary resistance (RL), which was determinedby multiple linear regressions from trans-pulmonary pressure and airflow.
Briefly, mice were placed in a whole-bodychamber, and basal readingswere obtained andaveraged for 3 min. Aerosolized saline, followedby histamine (1.28 mg/ml), were nebulized for3 min, and readings were taken and averagedfor 3 min after each nebulization.
Cells Culture
The 16HBE14o�cells, a generous gift ofDr. Dieter Gruenert, University of CaliforniaSan Francisco, were SV40-transsformed humancentral airway epithelial cells. Cells werecultured in a mixture medium of DMEM: F12(1:1) (Sigma) containing 100 U/ml penicillin,100 U/ml streptomycin, and 10% fetal bovineserum (Hangzhou Sijiqing Biotech., Hangzhou,China) and incubated at 378C in 5% CO2.
Real Time PCR Measurement for Expressionof CTNNAL1 mRNA in Lung Tissues
and Cultured HBEC
RNA was extracted from mouse lung tissuesand cultured HBEC and reverse transcriptionwas performed by AMV reverse transcriptase(QIAGEN,Germany). PCRwas then carried outusing primers and taqman probes (TaKaRa,China). Taqman probes were labeled at their 50-end with the reporter dye 5-carboxyfluorescein(FAM) and at their 30-end with the quencher 6-carboxylelra-methyl rhodamine (TAMRA). Theprimers and taqmanprobeswere synthesizedasTable I. Briefly, 1 ml of the reverse-transcripts
was added to a 30 ml PCR mixture for 40 cycles.Each cycle included 938C for 30 s, 608C for 60 sby using Taq polymerase. Negative controlsconsisted of an equal volume of water substi-tuted for the volume of RNA in the RT reaction.Normalization of mRNA expression data forsample-to-sample variability in RNA input,RNA quality, and reverse transcription effi-ciency was achieved by comparing the copynumbers of targetmRNAswith that ofmouse orhuman GAPDH for each run.
In Situ Hybridization (ISH) Detectionfor CTNNAL1 mRNA in Cultured
HBEC and Lung Tissue
The following probes labeled 5-end withdigoxin (purchased from TaKaRa BiotechnologyCompany) were used for ISH to localize mouseCTNNAL1 mRNA: 50-ATCACCACACTTATA-AACCATAAAGATAATACCAA-30, 50-GCATA-CATGCAGAGGAGACATTTCAGGTGACTGGC-30, 50-ATGTCCAGGATGGCCTATTCTCTGTA-TTTATTTAC-30, or human CTNNAL1 mRNA: 50-ACCTTCCGGAGAAGCCCCAGGAAGACATTA-30, 50-ATGGAAGGGGTCTGAGAAGAGATAA-GGGCG-30, 50-GATCAAAACTCGCTCGGTGG-AGCAGACGCT-30. Briefly, the lung tissueswere fixed in 4% paraformaldehyde, embededin paraffin, cut into 6 mm paraffin sections andplaced on slides, the coverglasseswithHBEConthem were fixed in 4% paraformaldehyde. Theywere incubatedwith 30%H2O2 and 5%pepsin inturn and then hybridized at 428C overnightwith labeled probes (2.0 g/ml). After washing,the slides and coverglasses were sequentiallyincubated with blocking buffer, biotin labeledrabbit-anti-digoxin antibody, streptavidin bio-tin peroxidase complex (sABC) and biotinlabeled peroxidase, followed by rinsing withPBS after every step. The peroxidant activitywas visualized by the 3, 3-diaminobenzidinetetrahydrochloride (DAB).Thecoverglasseswereincubated with normal mouse serum instead ofthe mouse-anti-digoxin antibody in absence ofthe labeled probe for the negative control.
Antisense Oligonucleotide (ASO) of CTNNAL1
CTNNAL1 ASO was designed according tothe humanCTNNAL1mRNAsequence (nucleo-tides 1–20 fromATG, AUGGCCGCCUCUCCC-GGACC). The ASO was synthesized at TaKaRaBiotechnology Company as a 20-base phosphor-othioate chemic oligonucleotides, where bases
TABLE I. Oligonucleotide Primers Used forReal-Time PCR Analysis
Sequence of primers
CTNNAL1-humanForward 50-TCTCGCAACTATGGAAAG-30
Reverse 50-GTAGGCAGAATCAGTAAAG-30
FAMþAACTGCGAATCAGCCCATþTAMRAGAPDH-human
Forward 50-CCACTCCTC CACCTTTGAC-30
Reverse 50-ACCCTGTTGCTGTAGCCA-30
FAMþTTGCCCTCAACGACCACTTTGTCþTAMRACTNNAL1-mouse
Forward 50-TCTTCGGGAGAATGTTTGCTT-30
Reverse 50-TGTGCTCGTGGCTGGTGTAG-30
FAMþCCGCATTGGAGGCCGTCTTGþTAMRAGAPDH-mouse
Forward 50-TGTGTCCGTCGTGGATCTGA-30
Reverse 50-CCTGCTTCACCACCTTCTTGA-30
FAMþ CCGCCTGGAGAAACCTGCCAAGTATGþ TAMRA
922 Xiang et al.

1–5 and 16–20 were modified with 20-O-(2-methoxy)-ethyl.CTNNAL1ASOwas transfectedinto cellswith lipofectin (Sigma) for 4h in serum-free DMEM. Then the ASO reaction mixturewas replaced with normal growth media (with10% FBS) and the cells were incubated undernormal conditions for an additional 16–20 h.The efficacy of theASO in inhibiting the activityof CTNNAL1 was verified by real time PCR. Anonsense oligonucleotide (50-TCTGGACTGTC-CCCCATTTC-30) was used as control.
Monolayer Wound Repair Assay
This assaywas used to demonstrate the effectof CTNNAL1 ASO on epithelial cell migrationand wound repair. We have previously pub-lished details of this method [Guan et al., 2006;Tan et al., 2006]. Briefly, 16HBE14o� cells weregrown until confluent in 12-well plates withDMEM: F12 (1:1), and a small woundwasmadein the confluentmonolayer with a rubber stylet.The edge of wound was recognized and theremaining wound area was measured seriallyper 4 h in 24 h by video microscopy (OlympusCompany, Japan). A linear regression equationof the remaining wound area to time was obta-ined. Repair index (RI), equal to the absolutevalue of slope, was used to judge the repairspeed of HBEC. HBEC were pretreated withCTNNAL1 ASO (0.4, 0.8, 1.0 mM) or Fn (10 mg/ml, Sigma) or both. A positive control, 10 mg/mlEGF, was done at the same time to ensure validexperiments.
Proliferation Assay
The cells were trypsinized in a 0.25% trypsinsolution and seeded in a 96-well plate at adensity of 104 cells (0.1 ml/well). After the cellsgrown for 24 h to approximately 80% sub-confluent state, 0.1 ml serum-free medium wasadded to each well to synchronize the growth ofcells for 24 h. Then EGF (10 mg/ml), Fn (10 mg/ml), CTNNAL1 ASO (0.4, 0.8, 1.0 mM) or both ofFn and CTNNAL1ASOwere added to each welland incubated for another 24 h. Each treatmentwas tested in at least six wells. Next, 15 ml of0.5% MTT solution was added to each well andincubated for 4 h. Then, the medium and MTTwere removed and 150 ml of DMSO was addedto eachwell. Themixturewas shaken for 10minto dissolve the crystal. The OD of each wellsolution was determined at 570 nm by using anELISA reader.
Western Blot Analysis for FAK Phosphorylation
HBE cells(5� 106) were lysed in ice-cold celllysis buffer (1% Triton X-100, 150 mM NaCl,10 mM Tris-HCl (pH 7.4), 2 mM sodium ortho-vanadate, 10 mg/ml leupeptin, 50 mM NaF,5mMEDTA, 1mMEGTA, and 1mMPMSF) bystirring for 1 h at 48C. The lysate was obtainedafter a centrifugation at 13,000g for 15 min. Anequal volume of Laemmli sample buffer wasadded to each cell lysate. Samples were boiledfor 10 min, and then equal amounts of proteinwere separated by 7.5% SDS–PAGE beforebeing transferred to nitrocellulose membrane.The membrane were blocked with 3% BSAin PBS for 2 h and then incubated with theanti-FAK mAb or anti-phospho-FAK mAb.Following incubation with the appropriatehorseradish peroxidase-conjugated secondaryantibody, detection was made using the enhan-ced chemiluminescene system.
Statistical Analyses
The data were analyzed using unpairedStudent’s t-test. Values were expressed bymean�SE. P< 0.05 was considered as statisti-cally significant.
RESULTS
The Expression of CTNNAL1 mRNAin Mouse Lung Tissue
To fully understand the dynamic changes ofCTNNAL1 expression during the developmentof asthma,weobserved the temporal and spatialdistribution of CTNNAL1 expression in mouselung tissue with the OVA-sensitized asthmamodel.
In the normal group, the structure of bronchiwalls and pulmonary alveoli were intact andregular, which showed no sign of inflamma-tionary infiltration, mucosal exudation, andcavitary stricture. However, inflammationaryinfiltration, mucosal exudation, cavitary stric-ture, and bronchial epithelial denudation wereobserved in asthma groups.
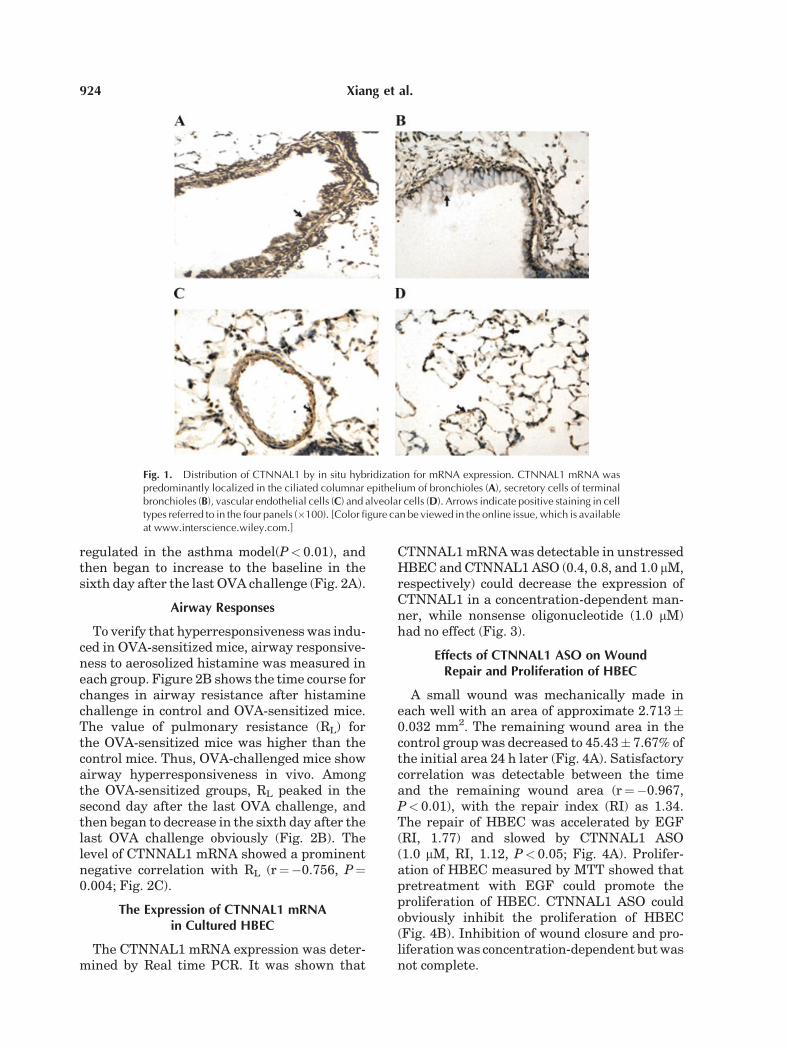
In situ hybridization showed that CTNNAL1mRNA was predominantly localized in theciliated columnar epithelium of bronchioles(Fig. 1A), secretory cells of terminal bronchioles(Fig. 1B), vascular endothelial cells (Fig. 1C)and alveolar cells (Fig. 1D).
Real time PCR showed that the expressionof CTNNAL1 mRNA was significantly down
Function Research on CTNNAL1 923
regulated in the asthma model(P< 0.01), andthen began to increase to the baseline in thesixth day after the last OVA challenge (Fig. 2A).
Airway Responses
To verify that hyperresponsivenesswas indu-ced in OVA-sensitized mice, airway responsive-ness to aerosolized histamine was measured ineach group. Figure 2B shows the time course forchanges in airway resistance after histaminechallenge in control and OVA-sensitized mice.The value of pulmonary resistance (RL) forthe OVA-sensitized mice was higher than thecontrol mice. Thus, OVA-challenged mice showairway hyperresponsiveness in vivo. Amongthe OVA-sensitized groups, RL peaked in thesecond day after the last OVA challenge, andthen began to decrease in the sixth day after thelast OVA challenge obviously (Fig. 2B). Thelevel of CTNNAL1 mRNA showed a prominentnegative correlation with RL (r¼�0.756, P¼0.004; Fig. 2C).
The Expression of CTNNAL1 mRNAin Cultured HBEC
The CTNNAL1 mRNA expression was deter-mined by Real time PCR. It was shown that
CTNNAL1mRNAwas detectable in unstressedHBEC and CTNNAL1ASO (0.4, 0.8, and 1.0 mM,respectively) could decrease the expression ofCTNNAL1 in a concentration-dependent man-ner, while nonsense oligonucleotide (1.0 mM)had no effect (Fig. 3).
Effects of CTNNAL1 ASO on WoundRepair and Proliferation of HBEC
A small wound was mechanically made ineach well with an area of approximate 2.713�0.032 mm2. The remaining wound area in thecontrol group was decreased to 45.43� 7.67% ofthe initial area 24 h later (Fig. 4A). Satisfactorycorrelation was detectable between the timeand the remaining wound area (r¼�0.967,P< 0.01), with the repair index (RI) as 1.34.The repair of HBEC was accelerated by EGF(RI, 1.77) and slowed by CTNNAL1 ASO(1.0 mM, RI, 1.12, P< 0.05; Fig. 4A). Prolifer-ation of HBEC measured by MTT showed thatpretreatment with EGF could promote theproliferation of HBEC. CTNNAL1 ASO couldobviously inhibit the proliferation of HBEC(Fig. 4B). Inhibition of wound closure and pro-liferationwas concentration-dependent butwasnot complete.
Fig. 1. Distribution of CTNNAL1 by in situ hybridization for mRNA expression. CTNNAL1 mRNA waspredominantly localized in the ciliated columnar epithelium of bronchioles (A), secretory cells of terminalbronchioles (B), vascular endothelial cells (C) and alveolar cells (D). Arrows indicate positive staining in celltypes referred to in the four panels (�100). [Color figure can be viewed in the online issue, which is availableat www.interscience.wiley.com.]
924 Xiang et al.

CTNNAL1 Expression on the Edge ofCells in Wounded Cell Culture
To clarify the biological role of CTNNAL1 inrepair and proliferation, examination of woundedges creation in monolayer for CTNNAL1expression by in situ hybridization was pre-
formed. The results showed that few expressionof CTNNAL1 mRNA was detectable in theinitial wound area, but its expression explicitlyincreased on the cells in wound edges 18 h aftercreation (Fig. 5).
CTNNAL1 ASO Inhibited Fn Mediated WoundRepair and Proliferation of HBEC
The results showed that Fn obviously pro-moted wound repair and proliferation of HBEC.HBEC incubatedwith 1.0mMofCTNNAL1ASOfor 30 min blocked the acceleration of bothwound repair and proliferation induced by Fn(Fig. 6A,B).
Effects of CTNNAL1 ASO on FAK Phosphorylation
Western blot showed that Fn could promoteFAK phosphorylation, which reached the peakin 30 min, then weakened and disappeared90 min later (Fig. 7A). CTNNAL1 ASO alonehad no effects on FAK phosphorylation. How-ever, it could inhibit the effects induced by Fn(Fig. 7B).
DISCUSSION
Destruction in airway epithelium structureand integrity, which is often induced by flawsof constitutive adhesion, is the essential link
Fig. 2. The relationship of CTNNAL1 expression and airwayresponse. A: Real time PCR showed the time course ofCTNNAL1expression in mice lung tissue. Group 1 day, 2 days,4 days, 6 days, and 8 days corresponding to the 1, 2, 4, 6, and8 days after the last challenge respectively. The result showedthat the expression of CTNNAL1 mRNA was significantly downregulated in the asthma model(P<0.01), and then began toincrease to the baseline in the sixth day after the last OVAchallenge (n¼5, **P<0.01). B: The time course for changes inpulmonary resistance after histamine challenge in control andOVA-sensitized mice. The value of RL for the OVA-sensitizedmice was higher than the control mice. Thus, OVA-challengedmice show airway hyperresponsiveness in vivo. Among theOVA-sensitized groups, RL peaked in the secondday after the lastOVA challenge, and then began to decrease in the sixth dayafter the last OVA challenge obviously. (n¼ 5,*P< 0.05). C: Thecorrelation of CTNNAL1 mRNA level and pulmonary resistance(RL).There is a negative correlation between them (r¼�0.756,P¼ 0.004).
Fig. 3. The expression of CTNNAL1 on HBEC assayed byreal time PCR. Real time PCR showed the expression ofCTNNAL1 on a human epithelial cell line, 16HBE14o-.CTNNAL1 ASO (0.4, 0.8, 1.0 mM, respectively) could inhibitCTNNAL1 expression in a concentration-dependent manner,while nonsense oligonucleotide(1.0 mM) had no effect (n¼3,*P<0.05 vs. control; **P< 0.01 vs. Control).
Function Research on CTNNAL1 925

of asthma pathogenicity. Therefore it can bereasonably speculated that abnormal expres-sion of adhesive molecules in airway epithelialcells, which might be possibly due to eithercorrespond coding gene alteration or imbalanceexpression among various kind of adhesionmolecules, contribute to aberrant susceptibilityto asthma. Based on the result of asthma-associated adhesion molecules expression spec-trum, CTNNAL1, whose expression was downregulated in asthma patients, was selected as acandidate in this present study for a furtherfunctional investigation. Using a well-charac-terized OVA-sensitized Balb/c mice asthmamodel, we carried out a time course study ofthe expression of CTNNAL1 in lung followingthe AHR progression. Our data provide adynamic picture of CTNNAL1 expression andairway responses. For the first time we demon-
strated that, CTNNAL1 may participate in thewound repair and proliferation of HBEC invitro. Our data indicates that the down regu-lation of CTNNAL1 expression might contrib-ute to the asthma development.
CTNNAL1 is firstly identified and chromoso-mal localized by Zhang et al. [1998]. It is a novelprotein homologous to a-catenin and its func-tion remains unclear.
a-catenin is essential for cadherin adhesiveactivity which forms functional cell–cell adhe-sion and links cadherin complexes to the actincytoskeleton, so that is able to provide flexibledynamic adhesion during wound repair of adulttissues and embryonic development [Tepass,2002; Perez-Moreno et al., 2003]. In bronchialepithelium, expression ofa-cateninwasnotedatcell–cell borders in both bronchial pseudostra-tified columnar and basal cells at relatively highlevels. Reduced expression of a-catenin wasclosely correlated with an atypical grade ofdysplasia in the basal layer andwas observed insquamous cell carcinoma [Pirinen et al., 2001;Kato et al., 2005].
With734aminoacids, thepredictedCTNNAL1polypeptide is smaller than a-catenin. It is notyet knownwhether CTNNAL1 localizes to cell–cell borders or is a component of adherensjunctions.However, the amino-terminal amphi-pathic helices of a-catenin that are thought toensure an interaction with b-catenin are alsopresent in CTNNAL1. Furthermore, amphi-pathic helices in the carboxy-terminal partof vinculin and a-catenin, which allow theseproteins to bind to the actin cytoskeleton, arealso present in CTNNAL1 [Park, 2002]. There-fore, sequence homology between CTNNAL1anda-cateninorvinculin implies thatCTNNAL1,whose function was not described previously,might have the potential to mediate cell–celladhesion.
HereweshowedthatmRNAlevel ofCTNNAL1was decreased in our asthma animals. Therewas a negative correlation between the pulmo-nary resistance (RL) in asthma mice and thelevels of CTNNAL1 mRNA in the 8 days’ timecourse after the last OVA challenge. It isconceivable that CTNNAL1 contribute to BECconstitutive adhesion. The down regulation of itmight hinder the maintenance of epitheliumintegrity in stressed condition. It was convincedthat the integrity of airway epithelium inasthmatic patients or normal people is signifi-cantly relevant with airway responsiveness
Fig. 4. The effects of CTNNAL1 ASO treatments on HBECwound repair (A) and proliferation (B). EGF could promoteHBECwound repair and proliferation. CTNNAL1 ASO (0.4, 0.8,1.0 mM, respectively) could partly inhibit the HBEC woundrepair and proliferation (n¼18, **P< 0.01 vs. control, *P< 0.05vs. Control). [Color figure can be viewed in the online issue,which is available at www.interscience.wiley.com.]
926 Xiang et al.

[Takizawa, 2005]. The reduction of CTNNAL1could account for the observed augmentationof the RL.In order to verify the role ofCTNNAL1 inBEC
constitutive adhesion, we performed in vitroexperiments to get further understanding ofassociated functions of CTNNAL1 in airway.Here we report the expression of CTNNAL1
on a human epithelial cell line, 16HBE14o-, byin situ hybridization and real timePCR, and theexpression was obviously decreased after thetreatment of CTNNAL1 ASO.Repair of a damaged epithelium may be a
necessary process in restoring airway functionto its normal state. Repair generally involvesseveral steps, including migration and spread-ing of epithelial cells at the margin of the injuryinto the damaged region and proliferation ofnew epithelial cells [Keenan et al., 1982a,b,c].The migration and dissemination to the woundoccur within 16–20 h after damage and theproliferation occurs around 24 h after thedamage [Kim et al., 1998]. It is reported thata-catenin links to growth signaling pathways[Barth et al., 1997] and regulates skin cellproliferation through a mitogen-activated pro-tein kinase dependent pathway [Vasioukhinet al., 2001]. Interestingly, CTNNAL1 mRNAappears to be differentially regulated followinggrowth arrest of cells, suggesting that it may
play a role in growth regulation [Zhang et al.,1998], although not any function of it was des-cribed previously. In this study, we found thatthe repair and proliferation of HBEC wereslowed down after treated with CTNNAL1ASO, and CTNNAL1 expression was explicitlyincreased on the cells in wound edges, whichindicating that CTNNAL1 might involved ingrowth regulation and may be beneficial for therecovery of epithelium damage. Our findingsshowed some functional similarities betweenCTNNAL1 and a-catenin.
Yeast two-hybrid analyses have uncoveredthe lymphoid blast crisis oncogene (LBC) Rhoguanine nucleotide-exchange factor as a part-ner for CTNNAL1 but not for a-catenin, whichindicates that CTNNAL1might have a new rolein modulating signaling by the Rho pathway[Park, 2002]. a-catenin and CTNNAL1 havedistinct activities that down regulate, respec-tively, b-catenin and Ras signals converging onthe cyclin D1 promoter [Merdek et al., 2004]. Itwill be interesting to see how the functions andassociations of CTNNAL1 differ from those ofthe ’classic’ a-catenins.
The binding of fibronectin (Fn), a extracellu-larmatrix protein, to integrins has been showedto alter expression of several genes, and toregulate adhesion, migration, proliferation andapoptosis of epithelial cells [Harkonen et al.,
Fig. 5. CTNNAL1 expression on the edge of cells in wounded cell culture. Few expression of CTNNAL1mRNA was detectable in the initial wound area (A1, 50�; A2, 200�), but its expression was explicitlyincreasedon the cells inwoundedges 18 h after creation (B1, 50�;B2, 200�). [Color figure canbe viewed inthe online issue, which is available at www.interscience.wiley.com.]
Function Research on CTNNAL1 927

1995; Matter and Ruoslahti, 2001; Ohke et al.,2001; Giuffrida et al., 2004; Jarvis and Bryers,2005; Yamazaki et al., 2005]. Signal trans-duction from integrins after binding of matrixprotein proceeds via several intermediate path-ways, such as alteration in calcium influx [Cantiet al., 2005], changes in inositol phosphatemetabolism [Jones et al., 2005; Ruzzi et al.,2005], and activation of multiple protein tyro-sine kinases, including focal adhesion kinase(FAK) and certain MAP kinases [Wierzbicka-Patynowski and Schwarzbauer, 2002].
Vinculin is present both in cadherin-cateninadhesion complexes that mediate cell-cell adhe-sion and in focal adhesion complexes that medi-ate cell-extracellular matrix adhesion, whilea-catenin is present only in cadherin-catenin
adhesion complexes [Weiss et al., 1998]. Asa member of the vinculin superfamily, thegenomic organization of CTNNAL1 is closelyrelated to that of the aE-catenin gene CTNNA1,but not at all to that of the vinculin gene[Janssens et al., 1999]. Here we showed Fncould obviously promoted wound repair andproliferation of HBEC, these effects can be sup-pressed by pretreatment of CTNNAL1 ASO.CTNNAL1 might be concerned with the migra-tion and proliferation mediated by Fn. Westernblot showed that Fn could promote FAK phos-phorylation in a transient time-dependentmanner. CTNNAL1 ASO alone did not causeFAK phosphorylation. However, it could inhibitFAKphosphorylation induced by Fn, indicatingthatCTNNAL1might have a role inmodulatingmigration and proliferation via FAK phospho-rylation signal transduction from Fn. Ourresults showed a novel link between CTNNAL1with Fn mediated cell-extracellular matrixadhesion. However, the role of CTNNAL1 infibronectin induced FAK-phosphorylation and
Fig. 6. The effects of CTNNAL1binding with Fn on woundclosure (A) and proliferation (B) in HBEC monolayers. Fn couldaccelerate the repair and proliferation of HBEC. CTNNAL1 ASO(0.4, 0.8, 1.0 mM, respectively) could partly inhibit the HBECwound repair and proliferation induced by Fn. (n¼ 18,**P<0.01 vs. control, *P<0.05 vs. control, #P<0.05 vs. Fn).[Color figure canbeviewed in theonline issue,which is availableat www.interscience.wiley.com.]
Fig. 7. Western blot showed that the effects of varioustreatments on FAK phosphorylation. A: The time course of FAKphosphorylation caused by Fn. The result showed that FAKphosphorylation became strong from 10 to 30 min, and thendeclined gradually. B: Western blot showed that Fn couldpromote the phosphorylation of FAK.CTNNAL1 ASO alone didnot affect FAKphosphorylation.However, CTNNAL1ASOcouldinhibit FAK phosphorylation induced by Fn (n¼ 3, **P<0.01 vs.control, #P< 0.05 vs. Fn).
928 Xiang et al.

its position in signaling cascade need to bestudied in our next step.Our result showed that CTNNAL1ASO could
decelerate the repairing velocity and prolifer-ation of HBECmediated by Fn or alone in vitro.One potential explanation is that CTNNAL1activate processes that lead to migration andproliferation in addition to Fn mediated tyro-sine kinase cascade signals. Inhibition of HBECmigration and proliferation in the present studywith the use of CTNNAL1 ASO, then, may bedue to blockade of not only cell-matrix but alsocell–cell interactions, as either or both proc-esses may be available during repair.In asthma, the bronchial epithelium shows
evidence of damage with loss of columnar cellsfrom their basal cell attachments [Takizawa,2005] and the enhanced expression of the epid-ermal growth factor receptor (EGFR, HER1,c-erbB1) [Puddicombe et al., 2000]. Epithelialdamage in asthma might due to increasedsusceptibility to a damaging agent and/or aninadequate repair response.Ourfindings revealthat the level of CTNNAL1 mRNA is highlycorrelated to the degree of pulmonary resist-ance; CTNNAL1 may contribute to the woundrepair andproliferation ofHBEC.Furthermore,it may serve to Fn mediated cell–extracellularadhesion and its signal transduction via FAKphosphorylation. These results raise the possi-bility that the down regulation of CTNNAL1might contribute to the asthma developmentbecause of the attenuated cell–cell and cell–matrix adhesion, which lead to the bronchialepithelium desquamated and AHR. Detailedfunctional analysis of CTNNAL1 may provideimportant clues to the understanding of for-mation and regulation of cell adhesion.
GLOSSARY
AHR airway hyperresponsivenessCTNNAL1 catenin alpha-like 1RL pulmonary resistanceHBEC human bronchial epithelium cellsASO antisense oligonucleotideISH in situ hybridizationRI repair indexFn fibronectin
ACKNOWLEDGMENTS
We thank Professor Gruenert from SanFrancisco Branch Campus, the University of
California for the gift of an immortalizedhumanBEC line 16HBE14o�.
REFERENCES
Barth AI, Nathke IS, NelsonWJ. 1997. Cadherins, cateninsand APC protein: Interplay between cytoskeletal com-plexes and signaling pathways. Curr Opin Cell Biol 9:683–690.
Campbell AM, Vignola AM, Godard P. 1994. Epithelial cellsand adhesion molecules. Respir Med 88:329–331.
Canti C, Nieto-Rostro M, Foucault I, Heblich F, Wratten J,Richards MW, Hendrich J, Douglas L, Page KM, DaviesA, Dolphin AC. 2005. The metal-ion-dependent adhesionsite in the Von Willebrand factor-A domain of alpha2-delta subunits is key to trafficking voltage-gated Ca2þchannels. Proc Natl Acad Sci USA 102:11230–11235.
Giuffrida A, Scarpa S, Birarelli P, Modesti A. 2004. Theinteraction of tenascin-C with fibronectin modulates themigration and specific metalloprotease activity in humanmesothelioma cell lines of different histotype. Int J Oncol25:745–750.
Guan CX, ZhangM, Qin XQ, Cui YR, Luo ZQ, Bai HB, FangX. 2006. Vasoactive intestinal peptide enhances woundhealing and proliferation of human bronchial epithelialcells. Peptides 27:3107–3114.
Harkonen E, Virtanen I, Linnala A, Laitinen LL, KinnulaVL. 1995. Modulation of fibronectin and tenascin pro-duction in human bronchial epithelial cells by inflam-matory cytokines in vitro. Am J Respir Cell Mol Biol 13:109–115.
Isenberg-Feig H, Justice JP, Keane-Myers A. 2003. Animalmodels of allergic asthma. Curr Allergy Asthma Rep 3:70–78.
Janssens B, Staes K, van Roy F. 1999. Human alpha-catulin, a novel alpha-catenin-like molecule with con-served genomic structure, but deviating alternativesplicing. Biochim Biophys Acta 1447:341–347.
Jarvis RA, Bryers JD. 2005. Effects of controlled fibronectinsurface orientation on subsequent Staphylococcus epi-dermidis adhesion. J Biomed Mater Res A 75:41–55.
Jones NP, Peak J, Brader S, Eccles SA, Katan M. 2005.PLCgamma1 is essential for early events in integrinsignalling required for cell motility. J Cell Sci 118:2695–2706.
Kato Y, Hirano T, Yoshida K, Yashima K, Akimoto S, TsujiK, Ohira T, Tsuboi M, Ikeda N, Ebihara Y, Kato H. 2005.Frequent loss of E-cadherin and/or catenins in intra-bronchial lesions during carcinogenesis of the bronchialepithelium. Lung Cancer 48:323–330.
Keenan KP, Combs JW, McDowell EM. 1982a. Regener-ation of hamster tracheal epithelium after injury. II.Focal lesions: Stathmokinetic and autoradiographicstudies of cell proliferation. Virchows Arch B Cell PatholIncl Mol Pathol 41:215–229.
Keenan KP, Combs JW, McDowell EM. 1982b. Regener-ation of hamster tracheal epithelium after mechanicalinjury. I. Focal lesions: Quantitative morphologic studyof cell proliferation. Virchows Arch B Cell Pathol Incl MolPathol 41:193–214.
KeenanKP, Combs JW,McDowell EM. 1982c. Regenerationof hamster tracheal epithelium after mechanical injury.III. Large and small lesions: Comparative stathmokineticand single pulse and continuous thymidine labeling
Function Research on CTNNAL1 929

autoradiographic studies. Virchows Arch B Cell PatholIncl Mol Pathol 41:231–252.
Kim JS, McKinnis VS, Nawrocki A, White SR. 1998.Stimulation of migration and wound repair of guinea-pig airway epithelial cells in response to epidermalgrowth factor. Am J Respir Cell Mol Biol 18:66–74.
Matter ML, Ruoslahti RA. 2001. A signaling pathway fronthe alpha5beta1 and alphavbeta3 integrins that elevatesbcl-2 trscription. J Biol Chem 276:27757–27763.
MerdekKD, NguyenNT, Toksoz D. 2004. Distinct activitiesof the alpha-catenin family, alpha-catulin and alpha-catenin, on beta-catenin-mediated signaling. Mol CellBiol 24:2410–2422.
Montefort S, Roberts JA, Beasley R, Holgate ST, RocheWR.1992. The site of disruption of the bronchial epithelium inasthmatic and non-asthmatic subjects. Thorax 47:499–503.
Nikolaidis NM, Zimmermann N, King NE, Mishra A, PopeSM, Finkelman FD, Rothenberg ME. 2003. Trefoil factor-2 is an allergen-induced gene regulated by Th2 cytokinesand STAT6 in the lung. Am J Respir Cell Mol Biol 29:458–464.
OhkeM, Tada S, NabeM,MatsuoK, KataokaM,HaradaM.2001. The role of fibronectin in bronchoalveolar lavagefluid of asthmatic patients. ActaMed Okayama 55:83–89.
Park Bea. 2002. Association of Lbc Rho guanine nucleotideexchange factor with a-catenin-related protein, a-catulin/CTNNAL1, supports serum response factor activation. JBiol Chem 277:45361–45370.
Perez-Moreno M, Jamora C, Fuchs E. 2003. Sticky busi-ness: Orchestrating cellular signals at adherens junc-tions. Cell 112:535–548.
Pirinen RT, Hirvikoski P, Johansson RT, Hollmen S,Kosma VM. 2001. Reduced expression of alpha-catenin,beta-catenin, and gamma-catenin is associated with highcell proliferative activity and poor differentiation in non-small cell lung cancer. J Clin Pathol 54:391–395.
Puddicombe SM, Polosa R, Richter A, Krishna MT,Howarth PH, Holgate ST, Davies DE. 2000. Involvementof the epidermal growth factor receptor in epithelialrepair in asthma. FASEB J 14:1362–1374.
Ruzzi L, Ciarafoni I, Silvestri L, Semeraro ML, Abeni D.2005. Association of PLA2 polymorphism of the ITGB3gene with early fetal loss. Fertil Steril 83:511–512.
Takizawa H. 2005. Bronchial epithelial cells in allergicreactions. Curr Drug Targets Inflamm Allergy 4:305–311.
Tan YR, Qi MM, Qin XQ, Xiang Y, Li X, Wang Y, Qu F, LiuHJ, Zhang JS. 2006. Wound repair and proliferation ofbronchial epithelial cells enhanced by bombesin receptorsubtype 3 activation. Peptides 27:1852–1858.
Tepass U. 2002. Adherens junctions: New insight intoassembly, modulation and function. Bioessays 24:690–695.
Vasioukhin V, Bauer C, Degenstein L, Wise B, Fuchs E.2001. Hyperproliferation and defects in epithelial polar-ity upon conditional ablation of alpha-catenin in skin.Cell 104:605–617.
Weiss EE, Kroemker M, Rudiger AH, Jockusch BM,Rudiger M. 1998. Vinculin is part of the cadherin-cateninjunctional complex: Complex formation between alpha-catenin and vinculin. J Cell Biol 141:755–764.
Wierzbicka-Patynowski I, Schwarzbauer JE. 2002. Regu-latory role for SRC and phosphatidylinositol 3-kinase ininitiation of fibronectin matrix assembly. J Biol Chem277:19703–19708.
Yamazaki D, Fujiwara T, Suetsugu S, Takenawa T. 2005. Anovel function of WAVE in lamellipodia: WAVE1 isrequired for stabilization of lamellipodial protrusionsduring cell spreading. Genes Cells 10:381–392.
Zhang JS, Nelson M, Wang L, Liu W, Qian CP, Shridhar V,Urrutia R, Smith DI. 1998. Identification and chromoso-mal localization of CTNNAL1, a novel protein homolo-gous to alpha-catenin. Genomics 54:149–154.
930 Xiang et al.














![Original Article PM2.5 exacerbate allergic asthma involved ... · kinase signaling pathway in human bronchial epithelial cells [17]. In this current study, we further investigated](https://img.pdfslide.net/doc/110x75/5e199bdefa15dc142f16e394/original-article-pm25-exacerbate-allergic-asthma-involved-kinase-signaling.jpg)














