Embed Size (px)
Citation preview

Ans dem Institut f u r Tierphysiologie und Tierernahrung der Universitat Gottingen Direktor: Prof . D r . D r . W . Lenkeit
Zur Abhangigkeit des Mineralumsatzes unter Beachtung der negativen Ca-, P- Anfangsbilanz wahrend der Laktation
von der Mineralstoffversorgung wahrend der Graviditat
7. Mitteilung der ,Langfristigen Untersuchungen zum aui3eren und inneren Stofiwechsel des graviden u n d laktierenden Schweines"
Von W. LENKEIT, J. 0. GUTTE, W. KIRCHHOFF und F. K. SOEHNGEN
Bekannt ist, da8 nach den Beobachtungen von FORBES (7), ELLENBERGER (5) u. a. !xi laktierenden Kuhen kurz nach den1 Kalben trotz ausreichender Versorgung die Cal- cium- wie Phosphorbilanzen negativ werden und gegen Ende der Laktationsperiode w i d e r positive Werte erreichen. Diese negative Bilanzphnse zu Beginn der Laktation wird jedoch nicht regelmaflig gefunden (14,28) .
Bei unseren Untersuchungen uber die Beziehungen des N-Umsatzes wahrend der Laktation zum N-Umsatz wahrend der Tragezeit beim Schwein wurde eine negative N-Bilanz wahrend der Laktation nur bei reichlicher Nahrstoff versorgung wahrend der Tragezeit gefundeii (21,23). Diese negativeBilanz in den ersten Laktationswochcn konnte namlich weder mit der Zufuhr wahrend dieser Zeit noch mit der Ausscheidung mit der Milch in Beziehung gebracht werden. Das in ublich kurzfristigen Perioden an- genommene Gleichgewicht zwischen Ausscheidung und Aufnahme ist hier nicht ge- geben.
In der vorliegenden Arbeit war zu prufen, in welchem Umfange die Mineralbilanz wahrend tder Laktation von der Griifle der Mineralzufuhr (Ca, P, Mg, K, Na) wah- rend der Tragezeit abhangt. In beiden Phasen wurden daneben Blutuntersuchungen iiber den Gehalt an Ca, P und an Phosphatase durchgefuhrt. Hier seien zunachst vor- wiegend die Untersuchungen uber den Ca-P-Umsatz mitgeteilt.
Zur Versuchsmethodik
Die Vercuche wurden mit Sauen als langfristige Stoffwehselversuche in der im Got- tinger Institut angewandten und fruher bereits mitgeteilten Weise durchgefuhrt (19, 23, 21, 23). Die Gewiniiung der Exkremente, die Bestimmung der Milchmenge, die Probenahmen der Milch wie der Placenta und des Fruchtwassers zur Analyse ist in

4 W'. Lenkert, J . 0. Gutte, W . Kirchhofi und F . K. Soehngen
den Mitteilungen 1 bis 4 beschrieben worden. Insgesamt werden hier zu der obigen Fragestellung die Untersuchungen uber den Mineralstoffwechsel an 5 Sauen herange- zogen. Da an den gleichen Versuchstieren auch der N-Umsatz gepriifi wurde, ist die allgemeine Versuchsiikrsicht (Korperentwicklung u. a.) hier nicht wiederholt wonden, es sei in dieser Baiehung auf die Mitteilungen 2, 3 und 4 vcrwiesen (20, 21, 23). Die Analysen wutrden nach den ublichen Methoden durchgefiihrt (s. LENKEIT-BECKER, Praktikum).
Die Nahrstoffzufuhr wurde in den beiden Phasen Tragaeit und Laktation jeweils konstant gehalten und ist aus Tabelle I zu ersehen.
Tabelle I
Futterration g/Tg
Gerste ............... 3500 Hafer . . . . . . . . . . . . . . . . 1000 Dorschmehl . . . . . . . . . . . . 600 Trockenhefe . . . . . . . . . . 150 Trodtenschnitzel . . . . . . . - Luzerne runmehl . . . . . . -
zus. Vit. D3 . . . . . . . . . . 1000 zus. Vit. A . . . . . . . . . . . . 10000 Kochsalz ............. -
- Molkenkchmehl .......
750 3500 2003 1000
150 600 100 150 - -
- 3050 300 450
200
1000
- -
- -
900 270 54 54 - - 162
1000 - -
1250 375 75 75 - - 225
1000 - -
649 551
20 700
80
1000
10
-
- -
3500 1000 600 150 - - -
1000 - -
Die Ration enthalt: Trockensubstanz . . . . . . . Rohprotein . . . . . . . . . . . N K , . . . . . . . . . . . . . . . . Gesamtnahrstoff . . . . . . .
Ca . . . . . . . . . . . . . . . . . . . P .................... N . . . . . . . . . . . . . . . . . . . Mg . . . . . . . . . . . . . . . . . . N a . . . . . . . . . . . . . . . . . . K . . . . . . . . . . . . . . . . . . . Carotin . . . . . . . . . . . . . . . Vit. A . . . . . . . . . . . . . . . .
4387 2521,2 4418,4 3388,4 1234,4 1676,2 1715,O 4369,O 977,2 410,5 878,s 662,4 260,7 353,7 192,7 831,O 8383 4337 8075' 5147 2202 3098 2651 7709
3673 1939 3533l 2357 965 1357 1280 3369 81492
3565 52,7 14,78 47,s 23,48 11,09 15,4 6,74 41,77 35,3 15,3 34,2 23,48 9,76 13,5 6,17 41,77
12,85 7,11 7,73 - - 2,63 - - 6,14 0,71 3,23 - - 1,86 - -
25,90 12,83 17,06 - - 11,13 - - - - -. 8,4mg - - 4,5 2,2
508 i.E. - - - 1027
- 156,4 65,7 140,6 - 41,71 56,6 -
- - -
'Aus den verdauten Nahrstoffen von 18 Trashtigkeitstagen errechnet. - *Aus den ver- dauten Nahrstoffen yon 59 Laktationstagen errechnet.
Aus den Versuchen sol1 aufgaeigt werden: 1. nochmals der Ablauf des Mineral- stoffumsanes wahrend der Lakration und die Moglichkeit der negativen Ca-P-Bilanz in den ersten Laktationstagen bei der Sau, 2. die Lage der Ausscheidung und der Re- tention wahrend der Graviditat. 3. war in weiteren Versudten (Rap. 11, Rotk. I, Si. 11) die Frage der Abhangigkeit dieses Bilanzverlaufes am Anfang der Laktation von der Grol3e der Ca- und P-Zufuhr wahrend der Tragaeit nu priifen.

Tabe
lle 2
Ca
- P - Umsatz
wPh
rend
der
Lak
tatio
n
Vers.
Snee
. II
(g/T
g.)
I P
ho
sp
ho
r
Ca
lciu
m
L.-T
.
gd:;. ~
-
Kot
Fu
tter
u.
Tra
nkw
. !
I
11 1 .-116.
1.-10.
11.-20.
21.-30.
31.40.
51.40.
1 .-lo.
n. d.
Lak
t.
~. 41.-50. -
52,221
52,771
52,759
52,797
52,746
52,618
52,541
52,087
~~ -
~
32,975
35,917
33,861
32,900
32,740
32,705
30,544
38,503
~~
._
. -
~ -~
1,105
0,407
0,540
0,368
0,567
1,014
0,720
1,863
-
__
__
_~
~~
11,043
15,796
17,333
17,852
14,047
13,770
11.-17.
Futt
erum
stel
lung
f 18,141
35,258
t 5,404
t 2,562
t 2,196
t 1,587
f 4,852
t 7,507
f 11,721
~.
~~
21,894
22,859
22,328
20,673
20,211
19,627
19,575
~~
4,639
5,154
4,754
4,047
4,930
5,489
6,318
23,907
5,900
-
~~
8,604
10,725
11,690
10,642
8,694
8,790
~~~
-
t 8,725
- 1,363
- 2,549
- 1,152
- 0,529
f 1,448
t 0,575
t 5,451
Futt
erum
stel
lung
18.-22.
18,013
12,471
0,629
t 4,913
19,394
13,394
5,087
f 0,913
23.-32.
13,157
0,603
t 4,253
12,736
3,969
t 2,689
33.-42.
12,805
0,779
t 4,429
12,931
4,103
f 2,360
53.-62.
13,316
0,729
f 3,968
11,520
4,840
f 3,034
63.-72.
13,990
0,588
f 3,435
13,112
4,606
t 1,676
43.-52.
11,997
0,530
t 5,486
1 1,772
4,049
t 3,573

- me.....
Z3
45
G7
lokr
f - W
artte
n
Abb
. 1.
Szu
gend
e Sa
u: C
a-,
P-B
ilanz
und
Aus
sche
idun
gen
in P
roze
nt d
er E
inna
hmen
(18
)

Zur Abhingigkeit des Mineralumsatzes 7
Die Versuchsergebnisse
I. Der Verlauf der Ca-P-Bilanz wahrend der Laktation
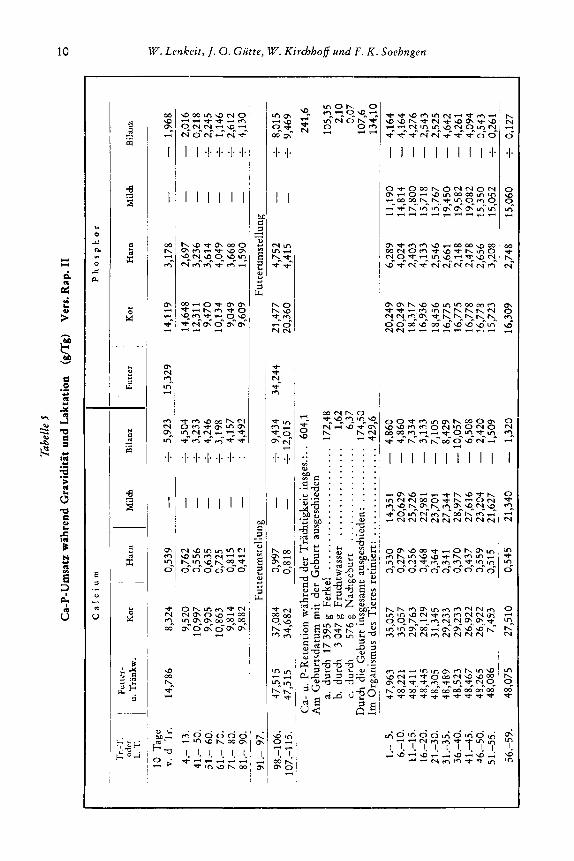
Der Versuch Snee. I1 (Tab. 2) begann am 6. Tage vor der Geburt und dauerte bis zum 72. Tage nach dem Absetzen der Ferkel am 60. Laktationstage. Bis zum Ende der Laktation wurden konstant 52,5 bis 52,7 g Ca (Futter + Trankwasser) und 35,3 g P zugefuhrt, 10 Tage nach der Laktation 18,O g Ca uad 19,4 g P. Der Versuch Rap. I1 (Tab. 5) setzte am 20. Tage vor der Tragae i t ein und dauerre bis zum 70. Tage nach der 59tagigen Laktation. Wahrend der Tragezeit bis zum 91. Tage erhielt das Tier 14,s g Ca und 15,3 g P, ab 91. Gravi,ditltstag bis zum Ende der Laktation 47,5 bis 48,3 g Ca, 34,2 g P, ab Laktationsende 12,9 g Ca und 14,2 g P (Tab. 5).
In beiden Versuchen ist die Bilanz sowohl des Ca wie des P, ofl auch des K und Na, zu Beginn der Laktation niedriger als gegen Ende der Laktation. Bei der Sau (Tab. 2, 5, 7, 9) kann danach die Ca/P-Bilanz, die K- und Na-Bilanz, ahnlich wie bei der Kuh, in den ersten Laktationstagen auch im negativen Bereich selbst bei reichlicher Ca-P-Zufuhr liegen, wie besonders der Versuch Rap. I1 zeigt. Das gleiche Bild er- gaben auch schon fruhere Untersuchungen (LENKEIT-BEINLICH 1953 [is], Abb. 1).
Dieser initiale Bilaiizverlauf ist nicht von einem entsprechenden Verlauf der mit der Milch ausgeschiedenen Ca- und P-Menge begleitet und ist auch ziemlich unabhan- gig von der Zufuhr in (dieser Zeit; an an'derer Stelie ist bereits dariiber berichtet wor- den (1954), (18). Gegenuberstellungen von Versuchsabschnitten mit gleichen Ca/P- Ausscheidungen in der Milch bei gleichbleibender Ca/P-Zufuhr (Tab. 7) ergeben, daf3 zu Laktationsbeginn die hoheren Ca- und P-Ausscheidungen mit dem Kot die Bilanz maf3geblich beeinflussen. Mit fortsdn-eitender Laktation nimmt in Bcstatigung fruhe- rer Untersuchungen an Sauen (Abb. 1) und n d Ergebnissen bei Ratten (FOURNIER), (8) die Ca-Ausscheidung im Kot ab, d. h. steigt die Ca/P-Ausnutzung. Bei Magnesium scheint der Verlauf umgekehrt zu sein (Abb. 6). Auf dieses Phanomen d e s hormonal gesteuerten zyklischen Ablaufs der Ausnutzung, auch wahrend der Tragaeit , bei konstanter Nahrstoffzufuhr sol1 an anderer Stelle eingegangen werden.
Diese Ergebnisse entsprechen den in den Mitteilungen 2, 3 und 4 aufgaeigten Be- fuiiden uber den Verlauf der N-Bilanz. Wie hierfur, so durfle au& fur die Mineral- bilanz am Anfang der Laktation der Umfang der Ernahrung wahrend der Tragae i t eine ursachliche Rolle spielen.
11. Die Bilanz wahrend der Graviditat
Daher seien zunachst die Untersuchungen uber die Ca- und 1'-Ausscheidung wie uber den Mineral-Bilanzverlauf wahrend der Graviditat mitgeteilt an Hand der Versuche Sie. 1 (Tab. 4), Rap. 11. (Tab. 5) und Sie. 11 (Tab. 7, Abb. 2, 3). Alle Versuche zeigen eine stark ansteigende Retention gegen Ende der Graviditat, etwa vom 75. bis 85. Tage ab, mit dem Maximum kurz vor der Geburt. So erreicht bei reichlicher Zufuhr schliei3- lich die Gesamteinlagerung z. B. bei Sie. I 690 g Ca, 518 g P (Tab. 3, 4). Die Reten- tion im Muttertier nach der Geburt kann bei reichlicher Ca-P-Zufuhr bis uber 70 O / o
der Gesamtretention betragen, bei niedriger Ca-P-Zufuhr (Tab. 3, 8, Abb. 4, 5) da- gegen nur bis um 25 O / o Die gesamten Mineraleinlagerungen konnen, wie beim N, in
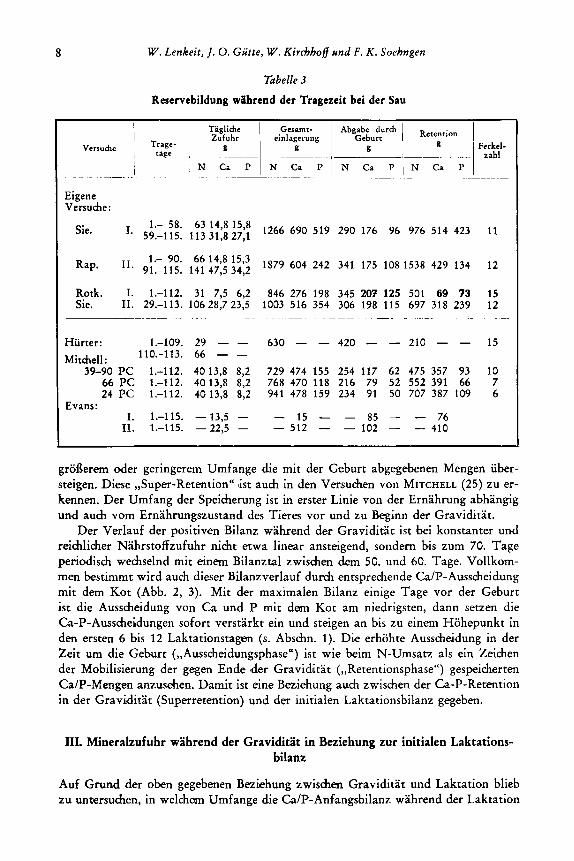
8 W. Lenkeit, J. 0. Giitte, W. Kirchhoff und F . K . Soehngen
Tabelle 3
Reservebildung wHhrend der Tragezeit bei der Sau
Eigene Versuche:
1.- 58. 63 14,s 15,8 59.-115. 113 31,s 27,l Sie. I.
Rap. 11. 1.- 90. 66 14,s 15,3 91.-115. 141 47,5 34,2
Gesamt- 1 Abgabe durdl 1 I g
einlageruiig Geburt
-~
1266 690 519 290 176 96 976 514 423 11
1579 604 242 341 175 1081538 429 134 12
Rotk. I. 1.-112. 31 7,5 6,2 846 276 198 345 201 125 501 69 '13 15 Sie. 11. 29.-113. 10628,723,5 1003 516 354 306 198 115 697 318 239 12
~ ~~~ -. ~
Hurter: 1.-109. 29 - - 630 - - 420 - - 210 - - 15 Mitchell: 110.-113. 66 - -
39-90 PC 1.-112. 4013,8 8,2 729 474 155 254 117 62 475 357 93 10 66 PC 1.-112. 4013,s 8,2 768 470 118 216 79 52 552 391 66 7 24 PC 1.-112. 4013,s 8,2 941 478 159 234 91 50 707 387 109 6
Evans: I. 1.-115. - 13,5 - - 15 - - 85 - - 76
rI. 1,115. -22,s - - 512 - - 102 - - 410
groderem oder geringereni Umfange die mit der Geburt abgegebenen Mengen iiber- steigen. Diese ,,Super-Retention" list auch in den Versuchen von MITCHELL (25) zu er- kennen. Der Umfang der Speicherung ist in erster Linie von der Ernahrung abhangig und auch vom Ernahrungszustand des Tieres vor und zu Beginn der Graviditat.
Der Verlauf der positiven Bilanz wahrend der Graviditat ist bei konstanter und reichlicher Nahrstoffzufuhr nicht etwa linear ansteigend, sondern bis zum 70. Tage periodisch wechselnd mit einem Bilanztal zwischen dom 5G. und 60. Tage. Vollkom- men bestimmt wird auch dieser Bilanzverlauf durch entsprechende Ca/P-Ausscheidung mit dem Kot (Abb. 2, 3). Mit der maximalen Bilanz einige Tage vor der Geburt ist die Ausscheiduiig von Ca und P mit dam Kot am niedrigsten, dann seaen die Ca-P-Ausscheidungen sofort verstarkt ein und steigen an bis zu einem Hohepunkt in den ersten 6 bis 12 Laktationstagsn (s. Abschn. 1). Die erhohte Ausscheidung in der Zeit um die Geburt (,,Ausscheidungsphase") ist wie beim N-Umsatz als ein Zeichen der Mobilisierung der gegen Ende lder Graviditat (,,Retentionsphase") gespeihrten Ca/P-Mengen anzusehen. Damit ist eine Beziehung auch zwischen der Ca-P-Retention in der Graviditat (Superretention) und der initialen Laktationsbilanz gegeben.
111. Mineralzufuhr wahrend der Graviditat in Beziehung zut initialen Laktations- bilanz
Auf G m d der oben gegebenen Bakhung zwischen Graviditat und Laktation blieb zu untersuchen, in weldiem Umfange die Ca/P-Anfangsbilanz wahrend der Laktation

Zur Abhangigkeit des Mineralumsatzes 9
Tabelle 4
Ca-P-Umsntz warend dcr Graviditat
Verr. Sic. I (g’Tg)
Calcium I Phosphor I -~ ...- . ~~-
Bilanz I Flitter 1 Kot 1 Harn 1 Bilanr Futtcr u.TrPnkw.
10 Tage v. d. Tr. 14,821 12,135 0,309 t 2,377 15,777 10,397 3,462 t 1,918
1.- 7. 9,783 0,125 - 4,913 9,657 1,750 - 4,370 29.- 38. 10,618 0,270 + 3,933 10,381 3,036 t 2,360 39.- 48. 10,229 0,251 t 4,341 9,669 1,821 t 4,287 49.- 58. 10,793 0,308 + 3,720 10,426 2,465 t 2,886
Futterumrtcllung ~~. ~ __ ~~ ~ - I Futterumsrellung
59.- 60. 61.- 65. 66.- 70. 71.- 75. 76.- 80. 81.- 90. 91 .-loo.
101 .-107. 108.-111. 1 12.-115.
31,783 13,610 26,151 26,577 25,314 24,614 25,169 22,742 23,022 22,441 19,435
0,455 0,520 0,374 0,375 0,444 0,433 0,342 0,200 0,261 0,168
t 17,718 t 5,112 t 4,832 t 6,094 t 6,725 t 6,181 i 8,699 f 8,561 f 9,081 t 12,180
27,054 11,882 19 029 19,633 18,760 18,346 17,255 16,045 15,433 15,175 13,199
3,382 4,602 5,387 4,379 4,776 3,199 5,196 5,350 4,229 5,528
f 11,790 t 3,423 t 2,034 t 3,915 t 3,932 t 6,600 t 5,813 t 6,271 t 7,650 f 8,327
Ca- und P-Retention wahrend der Trachtigkeit insgesamt: Ca 690,2
Am Geburtsdatum mit der Geburt ausgeschieden: a. durch 14 335 g Ferkel 172,47 b. durch 1106 g Fruchtwasser 0,55 E. durm 31 944 g Nachgeburc 2,88
Durch die Geburt insgesamt ausgedieden: 175,9 114,3 Im Organismus des Tieres retiniert:
P 518,6
-
93,24 0,19 2,36
95,8 422,8
von der Hohe der Mineralzufuhr wahrend der Graviditat abhangt. Hierzu sind die langfristigen Stoffwechselversuhe Sie. I1 und Rotk. I durhgefiihrt worden. Ober den N-Umsatz bei diesen V e r s u b wie iiber die allgemeinen Angaben ist in der Mittei- lung 4 berichtet worden (23).
1. Ernahrung wahrend der Graviditat reichlicb (Versuch Sie. 11, Rap. 11)
Die Versuchssau Sie. wurde erst vom 21. Graviditatstage vorgefuttert. Der Versuch begann am 29. GravidiGtsmg und dauerte ohne Unterbrdung bis zum 17. Tage nach Beendigung der 6Otagigen Laktation (Tab. 7). Wahrend der Tragae i t und hk ta t ion erhielt das Tier die gleihe Ration mit 530 g vadaulihem Rohprotein, mit 28,7 bis 29,6 g Ca, 23,5 g P, 5,s g Mg, 5,3 g Na und 22,5 g K, w m i t der Bedarf an Ca und P fur die Tragae i t iibershritten wird und in der Laktation fur eine mittlere Milchbil-
Tibe
lle 5
Ca-
P-U
msa
tz w
ahre
nd G
ravi
ditH
t und
Lak
tatio
n (g/Tg) Ve
rs.
Rap
. 11
Ph
os
ph
or
I
Ca
lciu
m
Futt
er
Kot
H
am
Mil&
B
ilanz
I
Tr-
T.
I F
utte
r-
oder
M
il&
Bila
nz
L,-
T,
1 U. T
rank
w.
I 0,
539
-
t
5,92
3 15
,329
14
,119
-
- 1
,968
~
-~
- _
__
_
3,17
8 ---
~~~~
-. .-
~ ~~
-
8,32
4 -~
v.
d
lr.
14,7
86
4.-
13.
41.-
50.
51.-
60.
61.-
70.
71.-
80.
81.-
90.
~ -_
- _
.
~
9,52
0 0,
762
-
f 4
,504
10
,997
0,
556
-
t
3,23
3 9,
905
0,63
5 -
t
4,24
6 10
,863
0,
725
-
f 3
,198
14,6
48
2,69
7 -
- 2
,016
12
,311
3,
236
-
- 0
,218
9,
470
3,61
4 -
t
2,24
5 10
,134
4,
049
-
f
1,14
6 9,
814
0,81
5 -
t
4,15
7 9,
882
0,41
2 -
f 4
,492
~~
~ ~
9,04
9 3,
668
-
+ 2
,612
9,
609
1,59
0 -
f 4
,130
~~
~
91.-
97.
Futte
rum
s:el
:ung
Fu
tteru
mst
ellu
ng
-~
~
~ _
_
98.-1
06.
47,5
15
37,0
84
0,99
7 -
f 9
,434
34
,244
21
,477
4,
752
-
f 8
,015
10
7.-1
15.
47,5
15
34,6
82
0,81
8 -
t 1
2,01
5 20
,360
4,
415
-
t
9,46
9 ~~
~~~
-~
~~
- .~
~
~-
Ca-
u. P
-Ret
entio
n w
ahre
nd d
er T
rach
tigke
it in
sges
.: . .
604,
l A
m G
ebur
tsda
tus
mit
der
Geb
urt
ausg
esch
iede
n 24
1,6
a. d
urh
17
395
g Fe
rkel
...
......
......
......
. 17
2,48
10
5,35
b.
dur
ch
30
47
g F
ruht
was
ser
....
....
....
....
1,
62
2,lO
134,
lO
c. d
urch
57
6g N
achg
ebur
t 6,
37
0,07
D
urch
die
Geb
urt i
nsge
sam
t aus
ges
174,
jO
107,
6 Im
Org
anis
mus
des
Tie
res
retin
iert
: ..
....
....
....
.. 42
9,6
-~
_
_ -
~~
~ ~
1.-
5.
47,9
63
35,0
57
0,53
0 14
,351
- 4
,860
20
,249
6,
289
11,1
90
- 4
,164
6.
-10.
48
,221
35
,057
0,
279
20,6
29
- 4
,860
20
,249
4,
024
14,8
14
- 4
,164
11
.-15.
48
,411
29
,763
0,
256
25,7
26
- 7
,334
18
,317
2,
403
17,8
00
- 4
,276
16
.-20.
48
,445
28
,129
0,
468
22,9
81
- 3
,173
16
,936
4,
133
15,7
18
- 2
,543
21
.-30.
48
,305
31
,345
0,
364
23,7
01
- 7
,105
18
,456
2,
546
15,7
67
- 2
,525
31
.-35.
48
,489
29
,233
0,
341
27,3
41
-
8,42
9 16
,775
2,
661
19,4
50
- 4
,642
36
.-40.
48
,523
29
,233
0,
370
28,9
77
- 10
,057
16
,775
2,
148
19,5
82
- 4
,261
4 1
.-45.
48
,467
26
,922
0,
437
27,6
16
- 6
,508
16
,778
2,
478
19,0
82
- 4
,094
46
.-50
48,2
65
26,9
22
0,55
9 23
,204
- 2
,420
16
,773
2,
656
15,3
50
-
0,54
3 7,
453
0,51
5 21
,627
- 1
,509
15
,723
~
-
3,20
8 15
,052
t
0,26
1 _
_
48,0
86
~ -.
~~
~
5 1 .-5
5.
56.-5
9.
48,0
75
27,5
10
0,54
5 21
,340
- 1
,320
16
,309
2,
748
15,0
60
t
0,12
7 ~~
Zur Abhartgigkeit des Mineralumratzrs 11
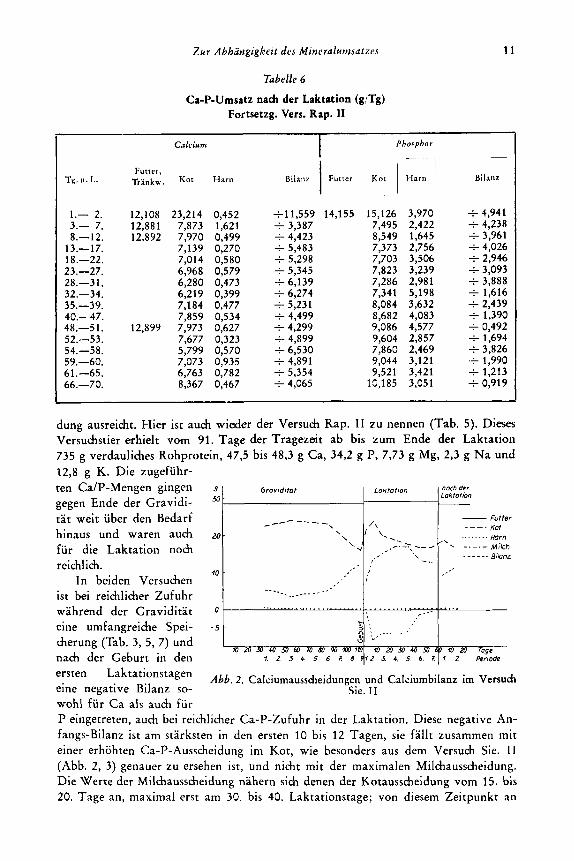
Tabelle 6
Ca-P-Umsatz nach der Laktation (g/Tg) Fortsetzg. Vers. Rap. JI
Tg. n. L.
1.- 2. 3.- 7. 8.--12.
13.--17. 18.-22. 23.-27. 28.-31. 32.-34. 35.-39. 40.- 47. 48.-51. 52.-53.
59.-60. 61.-65. 66.-70.
54.-58.
Calcium
l Futter, I Tri,,kw, Kot H a m
12,108 23,214 12,881 7,873 12,892 7,970
7,139 7,014 6,968 6,280 6,219 7,184 7,859
12,899 7,973 7,677 5,799 7,073 6,763 8,367
0,452 1,621 0,499 0,270 0,580 0,579 0,473 0,399 0,477 0,534 0,627 0,323 0,570 0,935 0,782 0,467
Phosphor
Biia iv Futter Kot ~ Harn ! -~ I I
t11,559 t 3,387 t 4,423 t 5,483 t 5,298 t 5,345 t 6,139 t 6,274 t 5,231 t 4,499 t 4,299 t 4,899 t 6,530 t 4,891 t 5,354 t 4,065
14,155 15,126 7,495 8,549 7,373 7,703 7,823 7,286 7,341 8,084 8,682 9,086 9,604 7,860 9,044 9,521
10,185
3,970 2,422 1,645 2,756 3,506 3,239 2,981 5,198 3,632 4,083 4,577 2,857 2,469 3,121 3,421 3,051
~
Bilanz
t 4,941 t 4,238 t 3,961 + 4,026 t 2,946 t 3,093 t 3,888 t 1,616 t 2,439 f 1,390 t 0,492 i 1,694 t 3,826 t 1,990 t 1,213 t 0,919
dung ausreicht. Hier ist auch wieder der Versuch Rap. I1 zu nennen (Tab. 5). Dieses Vemuchstier erhielt vom 91. Tage der Tragezeit a b bis zum En& der Laktation 735 g verdauliches Rohprotein, 473 bis 48,3 g Ca, 34,2 g P, 7,73 g Mg, 2,3 g Na und 12,8 g K. Die zugefuhr-
gegen Ende der Gravidi- ten Call’-Mengen gingen 3 Groviditot Lahfofion
t a t weit uber den Bedarf hinaus und waren auch LO-
fur die Laktation noch
In beiden Versuchen
_-----_
reichlich. - I 0
I I - Futter
- _ _ _ _ _ - - - - ist bei reichlicher Zufuhr wahrend der Graviditat * eine umfangreihe Spei- - 5
nach der Geburt in den ersten Laktationstagen eine negative Bilanz so- wohl fur Ca als auch fur P eingetreten, auch bei reichlicher Ca-P-Zufuhr in der Laktation. Diese negative An- fangs-Bilanz ist am starksten in den ersten 10 bis 12 Tagen, sie fallt zusammen mit einer erhohten Ca-P-Aussch’eidung im Kot, wie besonders aus dem Versuch Sie. I1 (Abb. 2, 3) genauer zu ersehen ist, und nicht mit der maximalen Milchausscheidung. Die Werte der Milchausscheidung nahern sich denen der Kotausscheidung vom 15. bis 20. Tage an, maximal erst am 30. bis 40. Laktationstage; von diesem Zeitpunkt an
herung (Tab* 3’ 5’’) und
Abb. 2. Calciumaussheidungen und Calciurnbilanz irn Versuch Sie. I1
12 W. Lenkeit, J . 0. Giitte, W . Kirchhoff und F . K . Soehngen
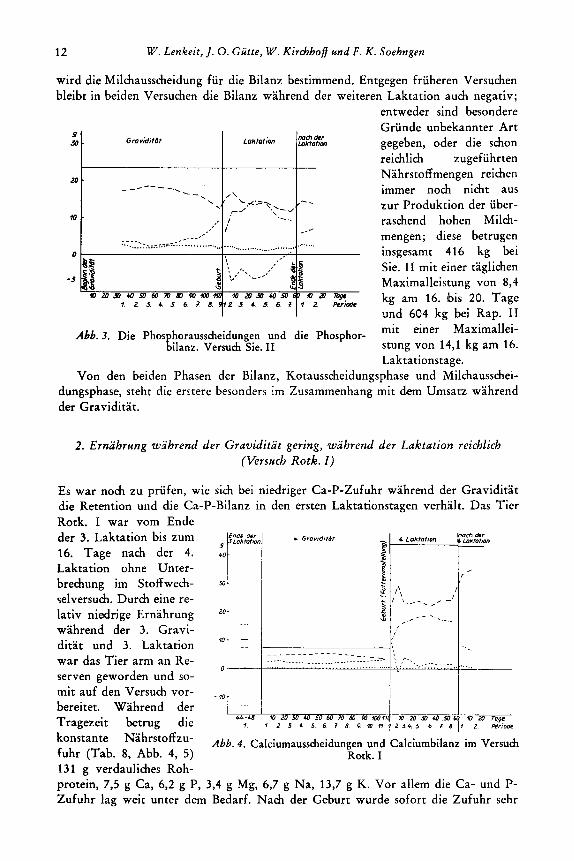
wird die Milchausscheidung fur die Bilanz bestimmend. Entgegen friiheren Versuchen bleibt in beiden Versuchen die Bilanz wahrend der weiteren Laktation auch negativ;
20 -
m -
4.. . ._ __-._ ... . . . ..._. , ,, , . . .. . . . ...
Abb. 3. Die Phos horausscheidungen und die Phosphor- EiIanz. Versuch Sie. 11
entweder sind besondere Grunde unbekannter Art gegeben, oder die schon reichlich zugefuhrten Nahrstoffmengen reichen immer noch nicht aus zur Produktion der uber- raschend hohen Milch- mengen; diese betrugen insgesamt 416 kg bei Sie. 11 mit einer taglichen Maximalleistung von 8,4 kg am 16. bis 20. Tage und 604 kg bei Rap. 11 mit einer Maximallei- stung von 14,l kg am 16. Laktationstage.
Von den beiden Phasen der Bilanz, Kotausscheidungsphase und Milchausschei- dungsphase, steht die erstere besonders im Zusammenhang mit dem Umsatz wahrend der Graviditat.
2. Ernahrung wzhrend der Graviditat gering, wahrend der Laktation reichlich (Versuch Rotk. I )
Es war noch zu priifen, wie sich bei niedriger Ca-P-Zufuhr wahrend der Graviditat die Retention und die Ca-P-Bilanz in den ersten Laktationstagen verhalt. Das Tier Rotk. I war vom Ende der 3. Laktation bis zum 16. Tage nach der 4. Laktation ohne Unter- brechung im Stoffwech- selversuch. Durch eine re- lativ niedrige Ernahrung wahrend der 3. Gravi- ditat und 3. Laktation war das Tier arm an Re- serven geworden und so- mit auf den Versuch vor- bereitet. Wahrend der Tragezeit betrug die konstante Nahrstoffzu- fuhr (Tab. 8, Abb. 4, 5) 131 g verdauliches Roh-
L Gromdrlor 3Lohmtmn
40 i"" der I
a t -
I I 46-u x, ro u) 6o 6o f 7,' ro 3 9 ' w a rage
I f 2 3"4 :OiDg , w w m ~ ~ 2 J C M s 6@? 6 7 1 2 Mrrobe
Abb. 4. Calciumausscheidungen und Calciumbilanz im Versuch Rotk. I
- protein, 7,5 g Ca, 6,2 g P, 3,4 g Mg, 6,7 g Na, 13,7 g K. Vor allem die Ca- und P- Zufuhr lag weit unter dem Bedarf. Nach der Geburt wurde sofort die Zufuhr sehr
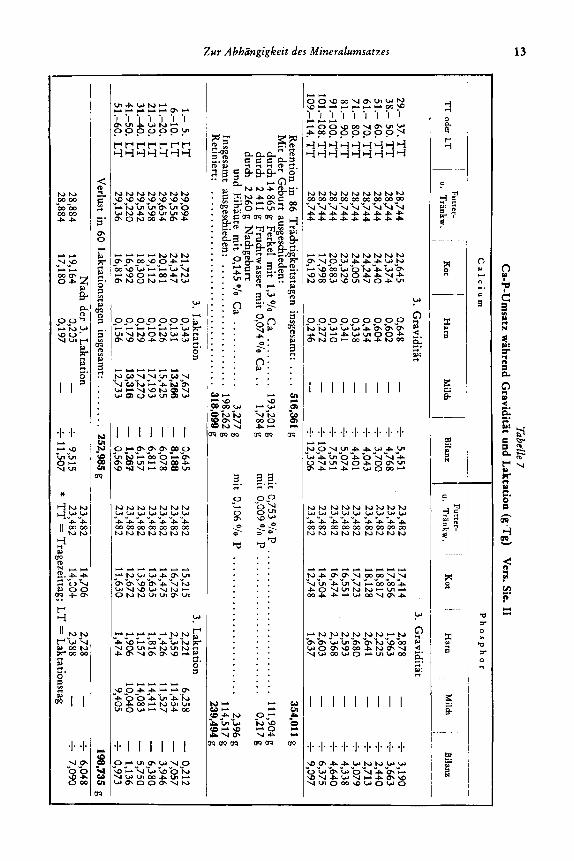
Tabe'lle 7 C
a-P-Um
satz wahrend G
raviditat und Laktation (g,T
g) V
ers. Sie. I1
Ca
lciu
m
I P
ho
sp
ho
r
~
I I
Mil&
,
Bilanz
Futter- I
u. Trankw.
I
Kot
~ H
arn M
il&
I TT O
der LT
29.- 37.T
T
28,744 38.-
50. TT
28,744
51.- 60. T
T
28,744 61.-
70. TT
28,744
71.- 80. T
T
28,744 81.-
90. TT
28,744
91.-100. TT
28,744
101.-108. TT
28,744
109.-114. TT
28,744
~
22,645 23,374 24,440 24,247 24,005 23,329 20,883 17,998 16,192
~ 3. Graviditat 0,648 0,602 0,604 0,454 0,338 0,341 0,310 0,272 0.2 46
f 5,451
t
4,768 t
3,700 +
4,043 t
4,401 f 5,074
f 7,551
t 10,474 f 12.306
3. Graviditat
23,482 17,414
2,878 -
23,482 17,856
1,963 -
23,482 18,817
2,225 -
23,482 18,128
2,641 -
23,482 17,723
2,680 -
23,482 16,551
2,593 -
23,482 16,474
2,368 -
23,482 14,504
2,603 -
23,482 12,748
1,637 -
t
3,190 f 3,663
f 2,440
f 2,713
t
3,079 t
4,338 t
4,640 t
6,375 f 9,097
Retention in 86 Trachtigkeitstagen
insgesamt: .... 516,361 g
354,011 g M
it der Geburt ausgedieden:
durch 14 865 g Ferkel mit 1,3 O/o
Ca ......... 193,201 g
mit 0,753 O
/o P .......................... 111,904 g
dur&
2 411 g Fruchtwasser m
it 0,074 O/o C
a . . 1,784 g
rnit 0,009°/o P ......................... 0,217g
durch 2 260 g N
achgeburt und E
ihaute mit 0,145 Olo C
a ............ 3,277 g
mit 0,106°/o P .........................
2.3
96
~
Insgesamt
ausgedieden: ....................... 198,262
114;517 g' R
etiniert: ...................................
318,099 g 239,494 g
~_
__
~~
~
~~~
3. Laktation
1.- 5. L
T
29,094 21,723
0,343 7,673
6.-10. L
T
29,556 24,347
0,131 13,266
11.-20. L
T
29,654 20.181
0,126 15,425
21.-30. L
T
29,598 19,112
0,104 17,193
31.40. LT
29,542
18,300 0,129
17,270 41.-50.
LT
29,220
16,992 0,179
13,316 0,156
12,733 51.40. L
T
29,136 16,816
~~ ~~~
Verlust in 60 L
aktationstagen insgesamt: .....
28,884 19,164
0,205 -
28,884 17,180
0,197 -
__
_-
Nach d
G 3. L
aktation
- 0,645
- 6,078
- 6,811
- 6,157
-
1,261 - 0,569
- 8,188
~_
__
_~
... 252,985 g - f 9,515
t 11,507
*
3. Laktation
23.482 15.215
2.221 6.258
23;482 16;726
2;359 11;454
23,482 14,475
1,426 11,527
23.482 13.635
1.816 14.411
23;482 13;992
1;157 14;083
23,482 12,672
1,906 10.040
23.482 11.630
1 :474 91405
--
23,482 14,706
2,728 -
23,482 14,004
2,388 -
TT
= T
ragezeittag; LT
= L
aktationstag
- 0,212
- 7,057
- 3,946
- 6,380
- 5,750
-
1,136 t
0,973 198,135 g t
6,048 t
7,090
14 W . Lenkert, J. 0. Gutte, W . Kirchhoff und F . K. Soehngen
erhoht, und zwar auf 681 g verdauliches Rohprotein, auf 44,O g Ca, 41,7 g P, 7,4 g Mg, entsprechend der Zufuhr in dem Versuch Rap. 11.
Wahrend der ganzen 4. Graviditat findet eine konstante niedrige Retention an Ca und P statt. Das starke Ansteigen der Bilanz gegen Ende der Tragezeit, das bei hoher
noch der 14 Lohfotron
4 Grovrditot
-5 ... c4-(B TO m N 40 50 m 70 do Po TmT ?D 2G 3 40 50 TO 20 Toge
T ' 1 2 3 4 5 6 ? 8 9 TO Tf ' f234 5 6 ? 8"p? 2 Per;&
Abb. 5. Phosphorausscheidungen und Phosphorbilanz. Versuch Rotk. I
Zufuhr gefunden wurde, erfolgt hier nicht. Die mit der Geburt ausge- schiedenen Ca-Mengen von 207 g und P-Mengen von 125 g iibertreffen hier die im Mutterorganismus gering retinierten Men- gen von 68 g Ca und 73 5 P (Tab. 3,s). Eine deut- liche negative Bilanz iil den ersten Laktations- tagen und auch spater ist infolge Fehlens eines zu mobilisierenden umfang- reichen Depots nicht ein-
getreten. Die zunachst erhohte Kotauss&eidung von Ca und P hangt einmal mit der plotzlichen Futterumstellung, zum anderen mit der niedrigen Ausnutzung in dieser Zeit und nur in geringem MaBe vielleicht mit einer Mobilisierung zusammen. Bei den Tieren Rap. I1 und Rotk. I ist die Zufuhr wahrend der Laktation ziemlich gleich hoch, der Verlauf der Anfangsbilanz aber unterschiedlich, wie die Ernahrung besonders ge- gen Ende der Graviditat. Der Versuch Rotk. I mit niedriger Ernahrung weist rnit den vorhergehenden Versuchen den gleichen Zusammenhang zwischen Graviditats- und Laktationsbilanz auf, wie er bereits beim N-Umsatz gezeigt werden konnte.
Diskussion
Durch die vorliegenden Versuche wird erneut die Annahiiie bestatigt, da13 die trotz reichlicher Ernahrung auftretenden negativen Ca- und P-Bilanzen in den ersten Ta- gen der Laktation durch die Hohe der Mineralzufuhren und der davon abhangigen Groi3e der Speicherung wahrend der Graviditat bestimmt werden. Je hoher die Spei- cherung infolge reichlicher Ca-P-Zufuhr moglich ist, um so groi3er ist die Mobilisie- rung von Korper-Ca-P in der ersten Zeit der Laktation, in der ,,Ausscheidungsphase". Die Folge ist die bei Kuh und Sau beobachtete negative Bilanz zu Anfang der Lalita- tion. Negative Bilanzen dieser Art sind ein Ausklang einer guten Ernahrung wfhrend der Graviditat und daher auch bereits beim N-Umsatz (21, 23) als ,,negative Uber- schuflbilanz" bezeichnet worden; im Gegensatz zu der .,negativen Mangelbilanz" bei unzureichender Ernahrung
Als vorwiegender Or t der Speicherung von Ca und P, auch Mg, K und N a und damit auch der Mobilisierung zur Zeit der Geburt, ist das Skelett anzunehmen. Den Teil des aus dem Knochen stammenden Milchcalciums schatzt MITCHELL (26) auf 20 O/o des Mineraldepots. Bei Mausen konnte rnit Ca45 nachgewiesen werden, dai3 das
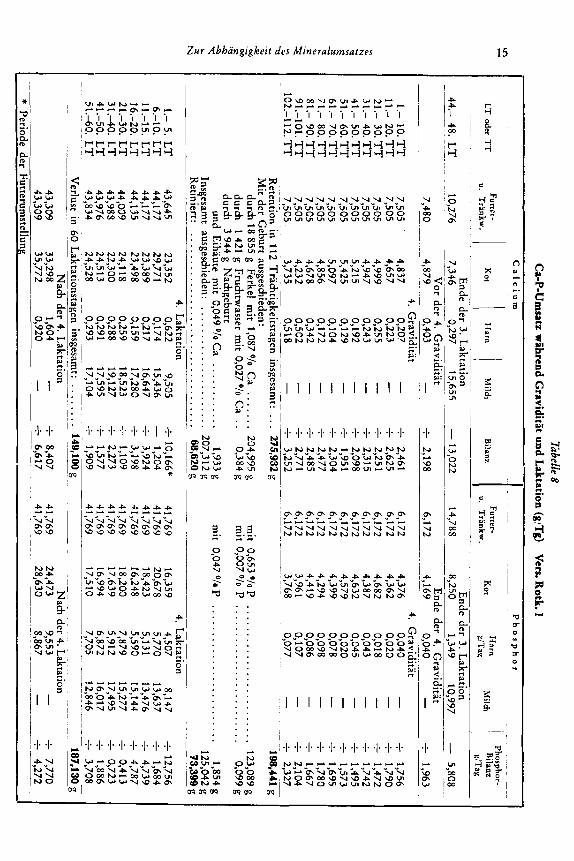
Zbelle 8 C
ia-P-Um
satz wjihrcnd G
raviditat und Laktation (g/Tg) Ven. R
otk. I
C a I c i u m
P
ho
sp
ho
r
-~
I-
~
1 P
hosphor- H
arn M
il&
Bilanz %
/Tag
Futter- K
ot d
Tag
Bi'anz
u.
Trankw
. I
Kot 1
Harn I
Mil&
Futter-
u. T
rankw.
LT oder T
T
10,276
7,480
7,505 7,505 7,505 7,505 7,505 7,505 7,505 7,505 7,505 7,505 7,505
.. ~
~~ ~
~~
Ende der 3. L
aktation 7,346
Vor der 4. G
raviditat 4,879
0,403 -
4. Graviditat
4,837 0,207
-
4,657 0,223
-
4,999 0,255
-
4,947 0,243
-
5,215 0,192
-
5,425 0,129
-
5,097 0,104
-
4,856 0,172
-
1,678 0,342
-
0,297 15,655 ~
-
__
~_
__
E
nde der 3. Laktation
8,250 1,349
10,997 E
nde der 4. Graviditat
4,169 0,040
-
4. Graviditat
4,376 0,040
-
4,362 0,020
-
4,682 0,018
-
4,387 0,043
-
4,632 0,045
-
4,579 0,020
-
4,399 0,078
-
4,294 0,098
-
4,419 0,086
-
--
~
44.- 48. L
T
____ - 13,022
i 2,198
i 2,461 i 2,625
+
2,251 i 2,315
+ 2,098 i 1,951 i 2,304
i 2,477 i 2,485
_____
14,788
6.1 72
.
5,808
1,963
1,756 1,790 1,472 1,742 1,495 1,573 1,695 1,780 1.667
~ ....
1.- 10. T
T
11.- 20. T
T
21.- 30. T
T
31.- 40. T
T
41.- 50. T
T
6,172 6,172 6,172 6,172 6.172
51.- 60. T
T
61.- 70. T
T
71.- 80. T
T
81.- 90. T
T
6;172 6,172 6,172 6,172
91.-101. TT
102.-112. T
T
4,232 0,502
-
i 2,771
6,172 3,961
Oil07
-
i 2,104
0,518 -
i 3,252
-
6,172 3,768
0,077 -
f 2,327
.-
__ ~
-~
~
3,735 R
etention in 112 TrachG
keitstagen insgesamt: ... 275,932 g
198.44G
Mit der G
eburt ausgeschieden: durch 18 855 g Ferkel m
it 1,087 O/o C
a ....... 204.995 g m
it 0.653 O/o P .........................
123.089 e durch
1 421 Fruchtw
asser rnit 0,027 O/o C
a . . 01384
mit 0,'007 O/o P .........................
OIO99 durch
3 944 g Nachgeburt
und Eihaute m
it 0,049 OloCa ............
1,933 g rnit
O,O
47O/oP .........................
1,854g 125,042 g
Retiniert:
................................... 68.620 e
13.399 e Insgesam
t ausgeschieden: ...................... 207,312 g
__
_~
~
4. Laktation
4,507 5,770 5,131 5,590 7,879 5,912 6,872 7,705 .
-~
-0
_
_ ~
+ 12,756 i 1,684
f 4,739 i 4,787
i 0,413
i 0,723
+
1,886 f 3,708
187,130
~- -
Laktation
0,622 9,505
0,174 15,436
0,217 16,647
0,159 17,280
0,259 18,523
0,288 19,127
0,291 17.595
1.- 5. L
T
43,645 6.-10.
LT
44,177
4. 23,352 29,771 23,389 23,498 24,118 22,300 24.513
-+ 10,166* -
1,204 i 3,924
+
3,198 +
1,109
i 2,273
+
1,577
41,769 4 1,769 41,769 11,769 41,769 4 1,769 41,769 41,769
~~
8,147 13,637 13,476 15,144 15,277 17,495 16,017 12,846 -
16,359 20,678 18,423 16,248 18,200 17,639 16,994 17,510
11.-15. L
T
44;177 16.40. L
T
44,135
31.40. LT
43,985
41.-50. L
T
43.976
21.-30. L
T
44,009
0,293 17,104
+
1,909 _
_ ~
-
51.40. LT
43;834
24,528 V
erlust in 60 Laktationsiayen insgesam
t: ........ 14
9,lK
I
- -
.. ~~~
Nach der 4. L
aktation 43,309
33,298 1,604
-
+
8,407 43,309
35,772 0,920
-
t 6,617
* Periode der Futterurnstellung -
- N
ach der 4. Laktation 24,473
9,553 -
28,630 8,867
-
~~~ - .
-~
41,769 41,769 .
-
f 7,770 i 4,272
__
__
__
16 W. Lenkeit, J . 0. Giitte, W7. Kirchhoff und F. K. Soehngen
in den Knochen fixierte Calcium durch die Milch ausgeschieden wurde (PECHER und J. PECHER, 28). Sicher sind an dam zyk1,ischen Umsatz die verschiedenen Knochen unterschiedlich beteiligt. Bei Schafen waren noch unzuneichender Calciumzufuhr wah-
. . . . ... . . . . .. . . . . ., . . . . . , _. . . , . . , I , _.- ..
L I I n I 2 V 9 4 0 5 4 I O a T o g e
2 3 & 5 6 7 I 2 Penodc
Abb. 6 . Magnesiumausskeidungen und Magnesiumbilanz Versuch Sie. I1
rend der Trachtigkeit und der Laktation starker die Wirbel, 'die Rippen und die Schadelknochen be- teikigt (DUCKWORTH, 4). OR stehen die Bedren- knochen im Vordergrund des Umwtzes. So wurden bei legenden Hennen in 100 Tagen der Lege- periode in Beckenkno- chen von Calcium ein 900/oiger Austausch und von Phosphor ein 1000/0- iger gefunden (SHIRLEY). An Hennen konnte auch
gezeigt werden, dafl wahrend der Schalenbildung Resorption und Anlagerung im Kno- chen stufenweise nacheinander stattfinden (BLOOM und DOOM, 1). Auch die Milchdruse wird als Ca-Depot wieder diskutiert (34).
Die gesteigerten Ausscheidungen des Ca und des P mit dem Kot in der Ausschei- dungsphase durfien eine gewisse Anpassung darsoellen, dai3 mit Abbau des Mineral- depots auch eine erhohte Sekretion, vielleicht von verminderter Ca-P-Resorption be- gleitet, zu Beginn der Laktation stattfindet. Nach HANSARD (12) hat der Umfang des Mineralspeichers eine groflere Wirkung auf die Sekretion von endogenem Ca als die Zufuhr. Grundsatzlich liegen die Fragen der Beziehungen des Knochenumbaues in der Sexualsphare zur Ausscheidung und Resorption noch offen.
Tabelle 9
K- und Na-Bilanz in g/Tg
Zufuhr Vor der Geburt i Na& der Geburt bis 5. Lakt.-Tag
Na l K Na I K Na
Vers. Sul. 11 16,4 3,9 + 4,a -t 0,5 - 3,O - 1,5 Vers. Snee. I 10,5 4,o - 0,3 -I- 3,5 - 2,6 4- 4,8 Vers. Rap. I 21,o 5,3 - 2,4 -t 3,6 - 6,2 - 4 2
In einer dem retinierten N entsprechenden neu gebildeten Korpersubstanz wlre nur ein kleiner Teil des retinierten Ca und P enthalten, ein groflerer des Na und K ausgeschwennmt (32, Tab. 9). Die Betrachtung der Kationenaquivalente in den Ver- suchen laflc bei den Vorgangen auch die Beteiligung eines lonenaustausches miiglich erscheinen (2).
Auf die Steuerung dieses zyklischen Stoff wedmlablaufs durch das innersekreto- rische Sysxem ist schon an anderer Stelle ausfuhrlich eingegangen worden (23). Die

Zur Abhangigkeit des Mineralumsatzes 17
Oestrogene, das Wachstumshormon, 'durflen die regulierenden Momente in der Reten- tionsphase win, analog in der Ausbildung des Hirschgewsihes (GRUEER, lo), die Epi- thelkorperchen, Nebennierenrindenhorone (Laktationshormone?) in der Abbau- phase. Der Einflufl der Sexualhormone, also auch des Ovarialzyklus, auf den Ca- Haushalt und auf den Mineralumsatz im Knochensystem (Apposition und Resorption) ist bekannt (GARDENER, PFEIFFER, 9, HANAU, 35, KIRCHHOFF, 17).
Die Bildung eines Mineraldepots unter dem EinfluB der Oestrogene ist bei der Taube und beim Huhn in Form einer Ossifikation der Markraume der Knochen fest- gestellt (KYES u. POTTER, 31, IWANOFF u. MLADENOFF, 15, DORN, 3). Diese Knochen- bildung geht der Groflenzunahme der Follikel parallel (BLOOM, DOOM u. MCLEAN, l), wahrend der Abbau dieses als ,,Follikulinknochen" ,baeichneten Ossifikationsknochens mit der Eischalenbildung verbunden ist. Auch durch Injektion von Oestrogenen kann der ,,Follikulinknden" erzeugt werden (3, 15). Ob die oben beschriebene Depohil- dung an Ca uad P bei der Sau und a d beim Rind in ahnlicher Weise erfolgt, kann noch nicht aufgezeigt werden, jedoch ist analog die Depotbildung mit folgender Mobi- lisierung unter dem EinfluB der hormonalen Regulation auch bei wei&hen Sauge- tieren durchaus gegeben. Do& laflt sich allgemein bei den Haustieren uber den Wir- kungsmechanismus der einzelnen Hormone auf den Knochenumbau, auf Sekretion und Resorption wahrend Tragezeit und Laktation no& nichts aussagm.
Die Ergebnisse unterstreichen erneut, dafl die GroBe der Bilanz nicht nur durch Er- nahrung und Leistung, sondern entscheidend von der jeweiligen Regulationsphase des Organismus bestimmt wird.
Zusammenfassung
D u d weitere langfristige Stoffwechseluntersuchungen uber die Beziehung der Gravi- didt zur Laktation werden hier die Untersuchungen an 5 Sauen uber den Einflufl der Mineralstoffzufuhr wahrend der Graviditat auf den Mineralumsatz wahrend der Laktation mitgeteilt.
Im Vorclergrund liegt die Prufung der Abhangigkeit auch der besonders beim Rind bekannten negativen Ca- und P-Bilanz im Anfang der Laktation von der Er- nahrung wahrend der Gravi'ditat.
Auch bei der Sau werden in der ersten Zeit der Laktation negative Ca- und P- Bilanzen gehmden, die abhangig von der Ernahrung und der Milchleistung sind.
Weiter wird gaeigt, daB der gravide Organismus befahigt ist, bei reichlicher Ca- P-Zufuhr groaere Ca-P-Mengen uber die Einlagerung in das Konzeptionsprodukt hinaus zu retinieren (Superretention). Nach der Geburt folgt wie beim Stickstoff eine Ausscheiidungsphase, in der diese umfangmkh gespeicherten Ca-P-Mengen ausgeschie- den wenden und somit zu einer negativen Ca-P-Bilanz in den ersten Laktationstagen fuhren, die als negative Ca-P- Uberschufibilanz bezeichnet wird, im Gegensatz zur negativen Mangelbilanz, die infolge unzureichender Ernahrung eintritt.
Durch niedrige Zufuhr von Mineralstoff en in der Retentionsphase (Graviditat) kann eine Speicherung von Ca und P verhindert werden; demzufolge bleibt in der fol- g d e n Ausscheidungsphase auch eine Abgabe von Ca und P aus, und eine ausge- glichene oder gar positive Bilanz wahrend der ersten Laktationscage ist die Folge.
Auch im Mineralumsatz wind die Groae der Bilanz nicht nur idurch die Ernahrung und Leistung, sondern entscheidend von der jeweiligen Regulationsphase des Organis- mus bestimmt.

18 18. Lenkeit, J. 0. Gutte, W . Kirchhoff wnd F. K. Soehngen
Summary
By further long term tests of metabolism with regard to the relation between gravidity and lactation there are published here examinations made with 5 sows dealing with the influence of mineral supply during gravidity upon mineral turnover during lactation.
Dominating is the test of the dependency of the negative Ca- and P-balance in the beginning of lactation upon nutrition during gravidity, which is known especially of cattle, too. Also in the case of sows during the first time of lactation negative Ca- and P-balances were found depending upon nutrition and milk production.
Further it is shown that the gravid organism is able to retain larger quantities of Ca/P when a surplus of Ca/P is offered by storing i t in the product of conception (superretention). Afier giving birth there follows a phase of elimination similar to that of nitrogen, when these Ca/P quantities stored in large degrees are eliminated and thus lead to a negative Ca/P balance during the first days of lactation, which is called a negative Ca/P surplus balance as compared to the negative lack balance which occurs in consequence of deficient nourishment. By a low supply of minerals during the retention phase (gravi'dity) a storage of Ca and P may be avoided, there- fore in the following phase of elimination and elimination of Ca/P does not occur and a level or even a positive balance during the first days of lactation is the consequence.
In the mineral turnover, too, the quantity of the balance is not only determined by nutrition and production, but decisively of the then dominant regulationphase of the organism.
Literatur
1. BLOOM, W., u. DOOM, L. V.: Anat. Rec. 81 (Amer. SOC. Zoo].), 91 (1941). - 2. BRONNER. F., HARRIS, R. S., MALETIKOS, C. J., und BENDA, C. E.: J. Clin. Invest. 35, 78 (1956). - 3. DORN, P.: D. T. W. 65. Jahrg., 563 (1958). - 4. DUCKWORTH, J.. u HILL- Nutr. Abstr. a. Rev. 23, 1 (1953). - 5. ELLENBERGER, H. B.: NEWLANDER and JONES: Vermont Agr. Expt. Sta. Bull. 342 (1932). - 6. EVANS und BISHOP: Science 56, 650 (1922). - 7. FORBES, E. B., SCHULZ, J. A., HUNT, C. H., WINTER, A. R., REMLER, R. F.: J. of Biol. Chern. 52, 281 (1922). - 8. FOURNIER, P., und SUSBIELLO, H.: J. Physiol. 44, 575-590, Paris 1952. - 9. GARDENER, W. U., und PFEIFFER, C. A.: Phys. Rev. 23, 139 (1943). - 10. GRUBER, G. B.: Zbl. f. allg. Pathol. u. pathol. Anat. 88, 336 (1952). - 1 1 . HANAU, A.: zitiert nach WERNLY. - 12. HAN- SARD, S. L., und PLUMLEE, M. P.: J. of Nutrit. 54, 17 (1954). - 13. HART, STEENBOCK und HOPPERT: J. of biol. Chem. 48, 33 (1951). - 14. HURTER, F.: Diss. agr. Bonn 1930. - 15. IWANOFF, X., u. MLADENOFF, Z.: Monatsschr. f . Vet. Med. 10, 175 (1955). - 16. KYES und POTTER: zit. nach SJOLTE. - 17. KIRCHHOFT, w.: Diss. sc. agr. Gottingen 1956. - 18. LENKEIT, W.: WissenschaRl. Abh. Dt. Akad. Landwirtschaftswissenschaften Berlin, Bd. V/2, S. 405 (1954). - 19. LENKEIT, W., und GUTTE, J. 0.: Z. Tierern. u. Futtermittelk. 10, 94 (1955). - 20. LENKEIT, W., GUTTE, J. O., und STREUTER-PETERMOLLER, A,: Z. Tierern. u. Futtermittelk. 10, 228 (1955). - 21. LENKEIT, W., GUTTE, J. O., WARNECKE, W., und KIRCHHOFF, W.: Z. Tierern. u. Futtermittelk. 10, 351 (1955). - 22. LENKEIT, W.: 6. Bericht der Ges.f. Ernah- rungsphysiologie der Haustiere. Z. Tierern. u. Futtermittelk. 11, 189 (1956) - 23. LENKEIT, W., GUTTE, J. O., KIRCHHOFF, W., SOEHNGEN, F. K., und FARRIES,, E.: Z. Tierern. u. Futter- mittelkunde 11, 323, (1956). - 24. MEIGS u. TURNER: J. of biol. Chem. 63. 29 (1925). - 25. MITCHELL, H. H., CARROLL W. E., HAMILTON, T. S., and HUNT, G. E.: University of Illinois, Agricultural Experiment Station, Bulletin 375 (1931). - 26. MITCHELL,H.H, u. CLURE, F. J.: Bulletin of the Nation. Res. Council Nr. 99 (1937). - 27. NEUMANN, W. F., und NEUMANN, M. W.: Chem. Rev. 53, 1 (19533). - 28. PECHER, C., und PECHER, J.: Proc. SOC. Exp. Biol. Med. 46, 91 (1941). - 29. RICHTER, K., und BECKER, M.: Arch. f . Tierern. 2, 338 (1952). - 30. SHIRLEY, R. L., DRIGGERS, J. C., MCCALL, J., DAVIS, G. K., und MEHRHOF, N. R.: Polutry Sci. 30, 730 (1951). - 31. SJOLTE, J. P.: Nordisk Veterinaermed. 1950. - 32. STREUTER- PETERMOLLER, A.: Diss. sc. agr. Gottingen 1955. - 33. SOEHNGEN, F. K.: Diss. sc. agr. Gottin- gen 1956. - 34. SWANSON, E. W., R. A. MONROE, D. B. ZILVERSMIT, W. J. VISEK und C. L. COMAR: J. Dairy Sci. 39, 1594 (1959). - 35. VISEK, W. J., BARNESI, L. u. LOOSLI, J. K.: J. Dairy Sci. 31, 783 (1952). - 36. WERNLY, M.: Die Osteomalacie, Thieme-Verlag, Stuttgart 1952.





























