
124 Monsalvo-SpencerP.et al.
Hidrobiológica
Hidrobiológica2013,23(1):124-129
Morfología de la membrana coriónica de los huevos de Octopus bimaculoides y Octopus hubbsorum (Cephalopoda: Octopodidae)
Chorionic membrane morphology of the eggs of Octopus bimaculoides and Octopus hubbsorum (Cephalopoda: Octopodidae)
PabloMonsalvo-Spencer,CésarAugustoSalinas-ZavalayTeodoroReynoso-Granados.
Centro de Investigaciones Biológicas del Noroeste, S.C. Apartado Postal 128, La Paz, B.C.S., 23080. México e-mail: [email protected]
Monsalvo-SpencerP.,C.A.Salinas-ZavalayT.Reynoso-Granados.2013.Morfologíade lamembranacoriónicade loshuevosdeOctopus bimaculoides yOctopus hubbsorum (Cephalopoda:Octopodidae).Hidrobiológica23(1):124-129.
RESUMENLoshuevosreciéndesovadosdeOctopus bimaculoides ydeO. hubbsorumsondeformaovocilíndrica,miden10.1±1.4y2.1±0.1mmdelongitud,respectivamente.Laenvolturadelhuevoconstadedospartesprincipales:lamembranaco-riónica(corion)yelpedúnculocoriónico.ElcoriondelhuevodeO.bimaculoidespresentaunasolacapa,deungrosorde8µm,conporosmenoresa1µmdediámetroyunaarquitecturainternaenconexiónconelpedúnculocoriónico.ElcoriondelhuevodeO. hubbsorumtieneungrosorde4µm,sinporosyunaarquitecturainternasininterconexiónconelpedúnculocoriónico.ElpedúnculocoriónicodelhuevodeO.hubbsorumpresentaunensanchamientoanulara300µmdelauniónconelcuerpoprincipaldelcorion.ElhuevodeO. hubbsorumtieneunmicrópiloenformadecavidadcircularprofundade15µmdediámetro,enelcentrodelpolomayordelcorion.EnesteestudiosedescribeporprimeravezquelamicroestructuramorfológicadelcorionesdistintaenloshuevosdeO. bimaculoidesyO. hubbsorum.
Palabras clave: Corion,morfología,Octopus bimaculoides,Octopus hubbsorum,ultraestructura.
ABSTRACTRecentlyspawnedeggsofOctopus bimaculoidesandO. hubbsorumareovo-cylindricalinshapeandhaveanaverageof10.1±1.4and2.1±0.1mminlength,respectively.Theeggenvelopeconsistsoftwomainparts:thechorionicmem-brane(Chorion)andthechorionicpeduncle.ThechorionofO.bimaculoideseggsshowsasinglelayerof8µminthick-ness,withporessmallerthan1µmindiameter,andtheinternalarchitectureofthechorionconnectstothechorionicpeduncle.ThechorionofO. hubbsorumeggshasathicknessof4µmwithoutpores,andtheinternalarchitectureofthechoriondoesnotinterconnectwiththechorionicpeduncle.ThechorionicpeduncleofO. hubbsorumeggsshowsanannularwidthof300µminthejunctionwiththemainbodyofthechorion.O. hubbsorumeggshaveamicropyleshapedasadeepcircularcavity,15µmindiameterinthecenterofthepoleofthechorion.Themicro-structuremorphologyofO. bimaculoidesandO.hubbsorumeggsshowsadifferentiationbetweenthechorionmembranes,whichisdescribedinthisstudyforthefirsttime.
Key words:Chorion,morphology, Octopus bimaculoides,Octopus hubbsorum, ultraestructure.

Morfologíacoriónicadehuevosdepulpo 125
Vol.23No.1•2013
INTRODUCCIÓN
EnlasespeciesdepulposdelsubordenIncirrata(incluyendolospulposbentónicosde la familiaOctopodidae), loshuevosestánenvueltos por una membrana coriónica o corion (Villanueva &Norman,2008)yenelextremodistaltienenunpedúnculocorió-nico,conelcualse forman racimosdehuevosduranteeldes-ove (Boletzky, 1998; Olivares et al., 2001). Generalmente, en loshuevosdesovados,elcorionasumelafuncióndeproteccióndelembrión contra factores desfavorables del medio ambiente co-moladesecación,lainfecciónbacterianayladestrucciónfísica,además,aseguralaestabilidadenelinteriordelhuevoconres-pectoalmedioexternopermitiendoelintercambiodegasesyelmantenimientodelembriónenlíquidovitelogénico(Budelmannet al., 1997). Sin embargo, los mecanismos físicos y químicos quepromuevenyregulanlafuncióndelcorionnoestándescritos.Enalgunosgruposcomoeldelosinsectos,lasestructurascorióni-cascaracterísticasdeloshuevossondiferentesenlasdistintasespecies(Rogoet al.,1992)yporellopodríanserútilesparahacerconsideracionesfilogenéticas(Hinton,1981).Enloscefalópodos,elcoriondeloshuevosnohasidoclasificadotodavíadeacuer-doalascaracterísticasdesusestructuras,niasucomposiciónbioquímica.
Enestetrabajosepresentanlascaracterísticasmorfológi-casdelcorionde loshuevosde lasespeciesdepulpoOctopus bimaculoides Pickford y McConnaughey, 1949 y O. hubbso-rum Berry,1953, queformanpartedelapesqueríadepulpoenelPacificoMexicanoyGolfodeCalifornia(Sánchez,2003;López-Uriarteet al.,2005;Alejo-Plataet al.,2009).Sedescribenporpri-meravezparaestasespecieslosdetallesdelamicroestructurasuperficialexternaeinternadelcorion,laformadeuniónconelpedúnculo coriónico y la existencia de un micrópilo. Este estu-diofueútilparadeterminarlaexistenciadediferenciasentrelasmembranascoriónicasdeloshuevosdeOctopus bimaculoidesyO. hubbsorum,dichasdiferenciaspodríanserconsideradascomocaracterísticastaxonómicaseinclusoevolutivas.
MATERIALES Y MÉTODOS
SerecolectarondiezhembrasdeOctopus bimaculoidesenmayoyochodeO. hubbsorumennoviembrede2011enlaIslaMagdale-na,BajaCaliforniaSur,México(24°38´N;112°08´W).Enestosme-ses,lasdosespeciestienenlasgónadasmaduras(López-Peraza,2009;Pliego-Cárdenaset al.,2011).
LospulposfuerontransportadosalLaboratoriodeManteni-mientodeOrganismosAcuáticosdelCIBNOR,enLaPaz,B.C.S.,México.Sedistribuyeronenformaindividualenjaulasdeplásticode50x27x25cm,ydentrodeellassecolocóuntubodePVCHde4“comorefugio.Lospulpossemantuvieronenunsistemasemi-abiertoa17°Cysalinidadde35ups.Latasaderecambiodeaguafuede300%cada24h.Lospulposfueronalimentadosconjaibas
vivas (Callinectes bellicosusSimpson,1859),cada72h.Lasjaulasfueronlimpiadasextrayendolashecesutilizandounsifónyelrefugiofuerevisadoconstantementeparadetectareldesove.
Cincoracimos dehuevos reciéndesovadosdecadaespe-cie,fuerondespegadosdelosrefugios.Delosracimossesepa-raron50huevosconpinzasysecolocaronencajasdePetride100mmdediámetro,conaguamarinafiltradaconuntamizde1µm,atemperaturade17°C.Sedeterminaronlassiguientesca-racterísticasmorfométricas:longitud(L)yancho(W)delhuevoylongituddelpedúnculocoriónico(LP).Lasmedidasserealizaronconunmicrómetroocularadaptadoaunmicroscopioestereos-cópico. La morfología macroscópica del huevo se registró conimágenes obtenidas con una cámara fotográfica conectada aun microscopio estereoscópico. Se estudió la microestructurasuperficialdelcorionde20huevosdecadaespecie,medianteelanálisisdelasimágenesobtenidasconunmicroscopioelectróni-codebarridomodeloHitachiS-300N(Murphy&Roomans,1984;Bozzola&Russell,1999).Lasmuestrasdehuevosanalizadasdecadaespeciefuerondelmismolotededesove.Elcorionfueobte-nidopordisección,separándolodelhuevo,abriendoesteúltimo,enformalongitudinalyventral,paraextraerelembrión.Elcorionaislado fue cortado en forma transversal en las regiones pola-resylongitudinalmenteenlazonacentral,quedandodivididoencuatropartesquefueronfijadas2horasenunasolución2.5%deglutaraldehidoa4°C.Después,fueronlavadostresvecesdurante1henaguademarsintéticaaunpHde7.4a4°C.Gradualmente,elaguademarfuesustituidaporunasoluciónamortiguadoradecacodilatodesodio,hastael100%,aunpHde7.4.Enseguida,sedeshidrataronenunaseriedeetanolal20%,40%,60%,80%y100%yposteriormentelasmuestrasfueronsecadasapuntocrí-ticoconunequipoSamdriPVT-3Byalfinalfueroncubiertosconpaladio,enunrecubridorDentonVacuumDeskII.
Apartirdelosdatosmorfométricosdeloshuevosseobtuvoelpromedioyladesviaciónestándar(n=50).
RESULTADOS
LashembrasdeOctopus bimaculoidesdesovaronentre15y20díasdespuésde larecolectay lasdeO. hubbsorum lohicieronentre20y24díasdespués.Lashembrasdelasdosespeciesmu-rieronalfinaldelcuidadoparentaldeloshuevos:O. bimaculoidesalos60díasyO. hubbsorumalos40díasdespuésdeldesove.
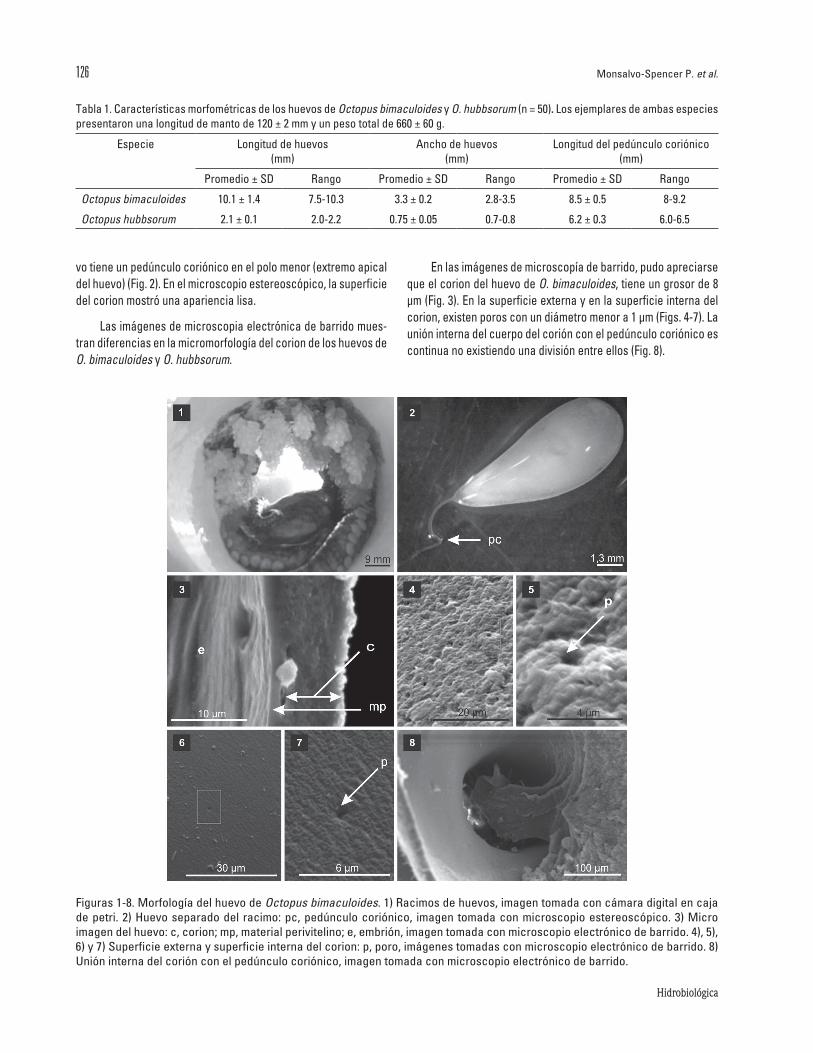
LascaracterísticasmorfométricasdeloshuevosdeO. bima-culoides yO. hubbsorumsemuestranenlaTabla1.LoshuevosdeO. bimaculoides tienen formaovocilíndricaysondesovadosenformaderacimos,unidosdirectamentealasuperficieinternadelrefugio(Fig.1).Loshuevosfrescossonsuaves,elembriónconvitelotieneuncolorblanquecino,inmersoenlíquidoperivitelinoydelimitadoporunamembranacoriónicadelgada,transparenteyflexible,quepermiteveralembriónquehayenelinterior.Elhue-
126 Monsalvo-SpencerP.et al.
Hidrobiológica
votieneunpedúnculocoriónicoenelpolomenor(extremoapicaldelhuevo)(Fig.2).Enelmicroscopioestereoscópico,lasuperficiedelcorionmostróunaapariencialisa.
Lasimágenesdemicroscopiaelectrónicadebarridomues-trandiferenciasenlamicromorfologíadelcoriondeloshuevosdeO. bimaculoidesyO. hubbsorum.
Enlasimágenesdemicroscopíadebarrido,pudoapreciarsequeelcoriondelhuevodeO.bimaculoides,tieneungrosorde8µm(Fig.3).Enlasuperficieexternayenlasuperficieinternadelcorion,existenporosconundiámetromenora1µm(Figs.4-7).Launióninternadelcuerpodelcoriónconelpedúnculocoriónicoescontinuanoexistiendounadivisiónentreellos(Fig.8).
Tabla1.CaracterísticasmorfométricasdeloshuevosdeOctopus bimaculoidesyO. hubbsorum (n=50). Losejemplaresdeambasespecies presentaronunalongituddemantode120±2mmyunpesototalde660±60g.
Especie Longituddehuevos(mm)
Anchodehuevos(mm)
Longituddelpedúnculocoriónico(mm)
Promedio±SD Rango Promedio±SD Rango Promedio±SD Rango
Octopus bimaculoides 10.1±1.4 7.5-10.3 3.3±0.2 2.8-3.5 8.5±0.5 8-9.2
Octopus hubbsorum 2.1±0.1 2.0-2.2 0.75±0.05 0.7-0.8 6.2±0.3 6.0-6.5
Figuras1-8.MorfologíadelhuevodeOctopus bimaculoides.1)Racimosdehuevos,imagentomadaconcámaradigitalencajadepetri.2)Huevoseparadodelracimo:pc,pedúnculocoriónico, imagentomadaconmicroscopioestereoscópico.3)Microimagendelhuevo:c,corion;mp,materialperivitelino;e,embrión,imagentomadaconmicroscopioelectrónicodebarrido.4),5),6)y7)Superficieexternaysuperficieinternadelcorion:p,poro,imágenestomadasconmicroscopioelectrónicodebarrido.8)Unióninternadelcoriónconelpedúnculocoriónico,imagentomadaconmicroscopioelectrónicodebarrido.
Morfologíacoriónicadehuevosdepulpo 127
Vol.23No.1•2013
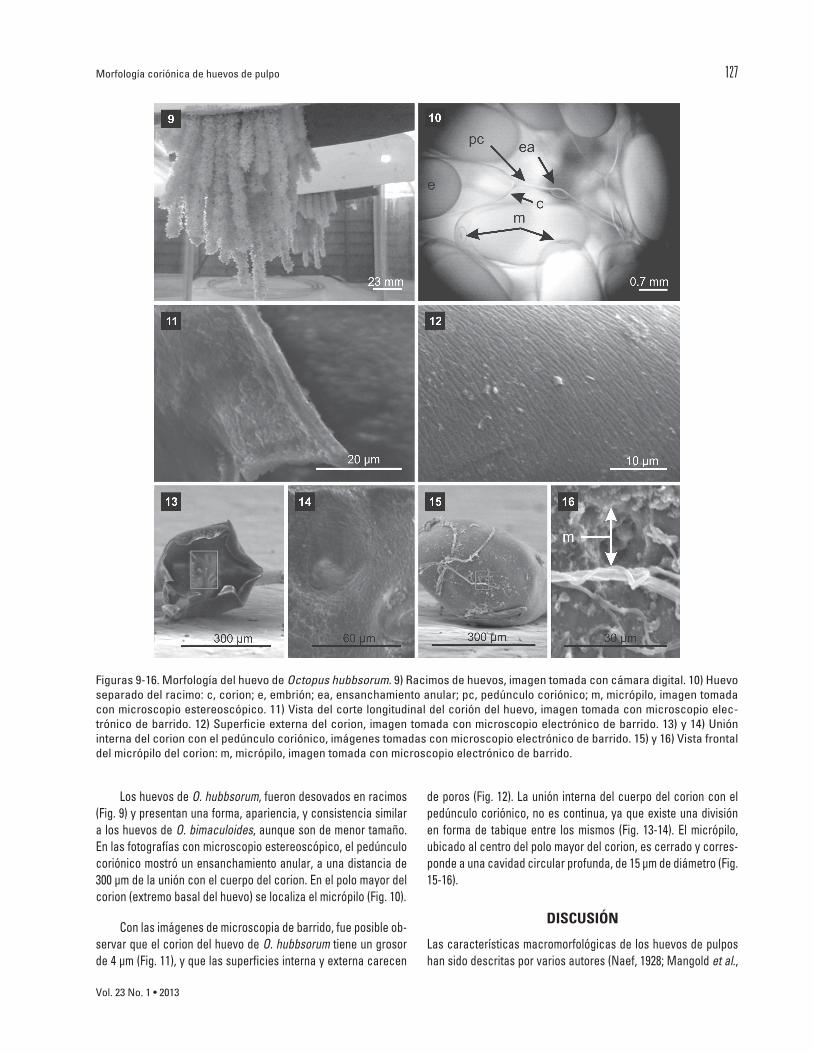
LoshuevosdeO. hubbsorum,fuerondesovadosenracimos(Fig.9)ypresentanunaforma,apariencia,yconsistenciasimilaraloshuevosdeO. bimaculoides,aunquesondemenortamaño.Enlasfotografíasconmicroscopioestereoscópico,elpedúnculocoriónicomostróunensanchamientoanular,aunadistanciade300µmdelauniónconelcuerpodelcorion.Enelpolomayordelcorion(extremobasaldelhuevo)selocalizaelmicrópilo(Fig.10).
Conlasimágenesdemicroscopiadebarrido,fueposibleob-servarqueelcoriondelhuevodeO.hubbsorumtieneungrosorde4µm(Fig.11),yquelassuperficiesinternayexternacarecen
deporos(Fig.12).Launióninternadelcuerpodelcorionconelpedúnculocoriónico,noescontinua,yaqueexisteunadivisiónen forma de tabique entre los mismos (Fig. 13-14). El micrópilo,ubicadoalcentrodelpolomayordelcorion,escerradoycorres-pondeaunacavidadcircularprofunda,de15µmdediámetro(Fig.15-16).
DISCUSIÓN
Lascaracterísticasmacromorfológicasdeloshuevosdepulposhansidodescritasporvariosautores(Naef,1928;Mangoldet al.,
Figuras9-16.MorfologíadelhuevodeOctopus hubbsorum.9)Racimosdehuevos,imagentomadaconcámaradigital.10)Huevoseparadodelracimo:c,corion;e,embrión;ea,ensanchamientoanular;pc,pedúnculocoriónico;m,micrópilo,imagentomadaconmicroscopioestereoscópico.11)Vistadelcortelongitudinaldelcorióndelhuevo,imagentomadaconmicroscopioelec-trónicodebarrido.12)Superficieexternadelcorion,imagentomadaconmicroscopioelectrónicodebarrido.13)y14)Unióninternadelcorionconelpedúnculocoriónico,imágenestomadasconmicroscopioelectrónicodebarrido.15)y16)Vistafrontaldelmicrópilodelcorion:m,micrópilo,imagentomadaconmicroscopioelectrónicodebarrido.

128 Monsalvo-SpencerP.et al.
Hidrobiológica
1971;Boletzky,1989).Entérminosgenerales,loshuevosdepulposolo tienenunamembranacoriónicaprotectoradelembrión in-mersoenlíquidoperivitelino.(Mangoldet al.,1971;Boletzky,1998;Villanueva&Norman,2008).Enloshuevosdecalamares,ellíqui-doperivitelinoesconsideradountranquilizantenaturalqueevitalarápidaeclosióndelasparalarvas(Marthyet al.,1976),sinem-bargo,supresenciaenloshuevosdepulponohasidoevaluadatodavía.Seconsideraquelospulposproductoresdeparalarvasplanctónicas,formanhuevospequeños(1.5-5mm),comoocurreen O. hubbsorum, y que los pulpos productores de paralarvasbentónicas forman huevos grandes (> 10 mm) (Hochberget al.,1992;Boletzkyet al., 2002),comosehadocumentadoparaO. bi-maculoides(Forsythe&Hanlon,1988).Segúnlaespeciedepulpo,loshuevosposeenunpedúnculocoriónicodeciertalongitud,quelosuneentresí,formandoracimos(Boletzky,1989;Huffard&Ho-chberg,2005).Estoseobservóenlosresultadosdelamorfometríadeloshuevosdeambasespeciesestudiadas:O. bimaculoidesyde O. hubbsorum. También se ha señalado que la variación delalongituddelospedúnculoscoriónicosdeloshuevosestámásrelacionadaconlacantidaddehuevosporracimoqueconelta-mañodeloshuevosdelosmismos(Villanuevaet al.,1995;Ortizet al.,2006).
Eneste trabajo,elexamende loshuevosconmicroscopioelectrónicodebarrido(MEB),mostróqueelcoriondeloshuevosde Octopus bimaculoides y de O.hubbsorumsondiferentes,enlamicromorfologíadelasuperficiedelcorionyenlasestructuraspresentesenlosdospolosdelmismo,especialmenteelpoloma-yor,dondesesitúaelmicrópilo.EnimágenesdeMEB,seobservóla existencia de poros en el corion del huevo de O. bimaculoi-des,locualnohabíasidoreportadoconanterioridad.Entrabajosacercadelcoriondehuevosdepeces teleósteos,comoelsal-món,Salmo salarLinnaeus,1758,sehasugeridoquelapresenciadeporosfacilitaladifusióndegasesyproductosmetabólicosdedesecho(Iconomidouet al.,2000)yademáspermitenlacomuni-caciónentrelascapasdelcorion(Jaramilloet al.,2009;2012).
Sehareportadoqueelcoriondeloshuevosdepulpo,tienecaracterísticasfísicasyquímicasquepermitenlaestabilidaddelhuevo (Budelmannet al., 1997)yevitanquemoléculasgrandesliberadas por el embrión y contenidas en el líquido perivitelinoabandonenelhuevo(Boletzky,2003).Sinembargo,elmecanismodeltipodepasodesustancias,yaseapasivo(O2,CO2,agua,urea,glucosa)y/oactivo(aminoácidos,polipéptidos,vitaminas),noes-tádeterminadotodavía.
LoshuevosdeOctopus vulgaris Cuvier,1797,carecendemicrópiloylasparalarvasaltiempodelaeclosiónrompenelpolomayordelcorion (extremobasaldelhuevo),con laglánduladeeclosiónuórganodeHoyle’s(Villanueva&Norman,2008).EnelhuevodeO. hubbsorum, elmicrópiloseencuentraprecisamenteenelpolomayordelcorionyaunquesufunciónnoesclarapo-dríaestarrelacionadoconlaeclosióndelasparalarvas,yaqueel
corionsealargayadelgaza,conformeavanzaeldesarrollodelaparalarva(Uriarteet al.,2009).Porotraparte,sufuncióntambiénpodríaestarrelacionadaparafacilitarelpasodelosespermato-zoidesparalafecundacióndelhuevo,antesdelapuesta,yaqueelcoriondelhuevoestáformado.
Dadoqueenestetrabajo,sedescribendiferenciasenlami-cromorfologíadelcorionde loshuevosdeO. bimaculoidesyO. hubbsorum,seríaconvenienterealizarestudiosdemicroestruc-turadeloshuevosdeotrasespeciesdepulposparadeterminarsilamicromorfologíadelcorionpuedereflejarlafilogeníadees-pecies, géneros o familias. Además, sería relevante estudiar elmecanismode lapermeabilidaddelcorion,paracomprender lainfluenciadelmismoeneldesarrollodelembrión,considerandoelefectodefactoresambientales,comotemperaturaysalinidad,locualayudaríaaoptimizarlaobtencióndeparalarvasdepulposenlaboratoriosdeacuicultura.
AGRADECIMIENTOS
Los autores agradecen la asistencia de Ariel Arturo Cruz Villa-cortaenlaoperacióndelmicroscopioelectrónicodebarrido.ALauraIbarraporlatomadefotografíasconelmicroscopioeste-reoscópicoyaGerardoRafaelHernándezGarcíapor laedicióndelasfotografías.ADianaLeticiaDorantesSalasporlarevisióndelresumeneninglés.
REFERENCIAS
Alejo-PlAtA, M. C, j. l, GóMez-Márquez, S. rAMoS-CArrillo & j. e. HerrerA-GAlindo. 2009. Reproducción, dieta y pesquería del pulpo Octopus (Octopus) hubbsorum(Mollusca,Cephalopoda)enlacostadeOaxa-ca,México.Revista de BiologíaTropical 57:63-78.
Boletzky, S. V. 1989. Recent studies on spawning, embryonic develop-mentandhatchinginthecephalopoda.Advances in Marine Biology44:143-203.
Boletzky, S. V. 1998.Cephalopodeggsandeggmasses.Oceanography and Marine Biology: Annual Review 36:341-371.
Boletzky, S. V., M. FuenteS & n. oFFner.2002.DevelopmentalfeaturesofOctopusmacropusRissso,1826(Mollusca,Cephalopoda).Vie Milieu52(4):209-216.
Boletzky, S. V.2003.BiologyofearlylifestagesinCephalopodmolluscs.Advances in Marine Biology44:143-203.
BozzolA, j. j. & l. d. ruSSell.1999.Electron Microscopy: Principles and techniques for biologists.(2d.ed.).Jones&BartlettPublishers(Bos-ton),USA.670p.
BudelMAnn, B., r. SCHiPP, & S. V. Boletzky. 1997. Cephalopoda. In: Har-rison, F. W. & A. J. Kohn (Eds.). Microscopic Anatomy of Inverte-brates, Volume6A.NewYork:Wiley-Liss,pp.119-414.

Morfologíacoriónicadehuevosdepulpo 129
Vol.23No.1•2013
ForSytHe, j. W. & r. t. HAnlon.1988.Effectoftemperatureonlaboratorygrowth,reproductionandlifespanofOctopus bimaculoides.Marine Biology98:369-379.
Hinton, H. e.1981.Biology of insect eggs.PergamonPress.Oxford.Vol.3.1125p.
HoCHBerG, F. G., l. d. nixon & r. B. toll.1992.OrderOctopodaLeach1818.Smithsonian Contributions to Zoology513:213-280.
HuFFArd, C. l. & F. G. HoCHBerG.2005.Descriptionofanewspeciesofthegenus Amphioctopus (Mollusca: Octopodidae) from the HawaiianIslands.Molluscan Research 25:113-128.
iConoMidou V. A., d. G. CHrySSikoS, V. GioniS, M. A. PAVlidiS, S. j. HAMo-drAkAS.2000.Secondarystructureofchorionproteinsoftheteleos-teanfishDentex dentex byATRFT-IRandFT-Ramanspectroscopy.Journal of Structural Biology 132:112-122.
jArAMillo r., o. GoiCoeCHeA, o. GArrido & e. MolinAri.2009.Salmo salar: Morfologíaultraestructuraldelapareddelcorionenovasnormalesyconproblemasdeeclosión.Archivos de Medicina Veterinaria 41:67-71.
jArAMillo r., o. GoiCoeCHeA, o. GArrido & e. MolinAri.2012. Caracteriza-ciónelectroforéticadelasproteínasdelcorionnormalydelcoriondurodeSalmo salar. Archivos de Medicina Veterinaria 44:59-65.
lóPez-PerAzA, d. j.2009.Descripciónhistológicadelosestadiosdema-durezgonadalenlospulposOctopus bimaculoidesyOctopus rub-escens.TesisdeMaestría.CentrodeInvestigaciónCientíficaydeEducaciónSuperiordeEnsenada,México.81p.
lóPez-uriArte, e., e. ríoS-jArA & M. Pérez-PeñA.2005.RangeextensionforOctopus hubbsorum (Mollusca: Octopodidae) in the Mexican Pa-cific.Bulletin of Marine Science7(2):171-175.
MAnGold, k., S. V. Boletzky & d. FröSCH.1971.ReproductivebiologyandembryonicdevelopmentofEledone cirrhosa(Cephalopoda:Octopo-da).Marine Biology8:109-117.
MArtHy, H. j., r. HAuSer & A. SCHoo.1976.Naturaltranquillizerincephalo-podeggs.Nature 261:496-497.
MurPHy, j. A. & G. M. rooMAnS (edS.). 1984. Preparation of Biological specimens for scanning electron microscopy. Scanning Electron Microscopy,Inc.,AMFO’Hare,II.344p.
nAeF, A.1928.DieCephalopoden.FaunaFloraGolf.Neapel.1.Teil,II.Em-bryologie35:1-357.
oliVAreS, P. A., M. zAMorA-CoVArruBiAS, P. PortillA-reyeS & o. zuñiGA-ro-Mero.2001.Estudiohistológicodelaovogénesisymaduraciónová-ricaenOctopusmimus(Cephalopoda:Octopodidae)delaIIRegióndeChile.Estudios Oceanológicos20:13-22.
ortiz, n., M. e. ré & F. Márquez.2006.Firstdescriptionofeggs,hatchlingsandhatchlingbehaviourofEnteroctopus megalocyathus(Cephalop-oda:Octopodidae).Journal of Plankton Research28(10):881-890.
PlieGo-CárdenAS r., F. A. GArCíA-doMínGuez, B. P. CeBAlloS-Vázquez, M. Vil-lAlejo-Fuerte & M. ArellAno-MArtínez. 2011. Reproductive aspectsof Octopus hubbsorum (Cephalopoda: Octopodidae) from EspírituSantoIsland,southernGulfofCalifornia,Mexico.Ciencias Marinas37(1):23-32.
roGo, l. M, e. d. kokWAro, M. j. MutinGA & C. P. kHAMAlA.1992.Ladife-renciacióndelasespeciesdevectoresdePhlebotominae(Diptera:Psychodidae) en Kenia por la escultura coriónica de sus huevos.Journal of Medical Entomology29:1042-1044.
SánCHez, P.2003.CephalopodsfromoffthePacificcoastofMexico:Bio-logical aspects of the most abundant species. Scientia Marina 67(1):81-90.
uriArte, i., o. zuñiGA, A. oliVAreS, V. eSPinozA, V. CernA, A. FAriAS & C. ro-SAS.2009.MorphometricchangesandgrowthrateduringembryonicdevelopmentofRobsonella fontaniana.Vie et Milieu- Life and Envi-ronment59(3/4):315-323.
VillAnueVA, r., C. nozAiSAnd & S. V. Boletzky.1995.Theplanktoniclifeofoctopuses.Nature377:1-107.
VillAnueVA, r. & M. d. norMAn.2008.Biologyoftheplanktonicstagesofbentonicoctopuses.Oceanography and Marine Biology 46:105-202.
Recibido:29deagostodel2012.
Aceptado:29deenerodel2013.
Recommended

















