COMMENTARY / COMMENTAIRE
Oligophosphoglyceroyl-ATP; a phosphate, purine, and energy resource for the cell?
JOHN MOWBRAY AND BRINDA PATEL Department of Biochemistry and Molecular Biology. University College London, Gower Street, London WClE 6BT, U.K.
Received May 20, 1991
Introduction Although in recent times the application of nmr spectro-
scopy to living tissues has tended to confirm the role of free ADP in regulating oxidative metabolism (Chance et al., Proc. Natl. Acad. Sci. U.S.A., 83, 9458-9462, 1986), this is only so provided the ADP concentration is below a relatively low threshold value ( - 50 pM). What is not clear are the mechanisms that restrain the ADP at this threshold, and nmr studies of the perfused rat heart (From et al., FEBS Lett., 206, 257-261, 1986) and intact dogs (Balaban et al., Science, 232, 1 121-1 123, 1986) have emphasized that respi- ratory control of heart mitochondria is not a function of free ADP concentration when this threshold is reached. Even more striking and difficult to explain is what limits the fall of ATP concentrations for up to tens of minutes when energy production is severely limited in hearts by ischaemia, while a "buffer" substrate like creatine phosphate disap- pears in tens of seconds; or again how the ATP pool is apparently replenished periodically during long-term ischaemia as nmr studies of frog muscle seem to show (Keidel et al., Res. Exp. Med., 84, 73-84, 1984). Similar experiments with perfused rat livers found nmr resonances for ATP declining faster than extracted ATP (Murphy et al., Biochemistry, 27, 526-528, 1988). The proposal that this missing nucleotide represented an nmr -invisible pool con- fined to the mitochondria1 matrix is challenged by the find- ings that all the ATP is nmr visible in isolated mitochon- dria (Hutson et al., Biochemistry, 28,4325-4332, 1989) and in the perfused heart (Humphrey and Garlick, Circulation, 80, 498, 1989). By contrast, on the related problem of cell inorganic phosphate concentrations, there is agreement that somewhat less than 50% of the extractable pool is ever nmr visible (see Humphrey et al., Eur. J. Biochem., 191, 755-759, 1990). The inference is that additional new infor- mation is required.
A new purine derivative One potentially apposite observation is our discovery in
rat heart of an entirely new oligomeric nucleoside tetra- phosphate derivative (Mowbray et al., Biochem. J., 223, 627-632, 1984). This discovery arose when we followed up observations that Langendorff perfused rat hearts, judged by a number of criteria to be in a steady state, showed synchronous time-based variations in the contents of their purine nucleotides. A disturbing feature of these results was that the sum of the adenine nucleotides was not constant but showed large regular and rapid changes in size so that within a 10-min period it could change by up to 1.3 pmol/g. There were no corresponding changes in any other purine nucleotides or nucleosides in either the tissue or the per- fusate, and the sizes of the changes ruled out de novo syn- thesis, known salvage pathways, and potential sources such Printed in Canada / Imprime au Canada
as RNA or pyridine nucleotides (see review, Lawson and Mowbray, Int. J. Biochem., 18, 407-413, 1986). Accord- ingly, we were driven to conclude that some previously unidentified major source of adenine nucleotide must exist in hearts. Very recent studies of adenine nucleotide deple- tion and purine degradation products in soleus muscle dur- ing brief tetanic contractions in ischaemic conditions led the authors to the same conclusions (Tullson et al., Am. J. Physiol., 258, C258-C265, 1990).
To test this hypothesis we selectively labelled heart nucleotides using 18-14~]adenosine and showed that all soluble radioactivity was essentially confined to ATP, ADP, AMP, GTP, and IMP, while about 15% of total tissue radioactivity had been incorporated into a trichloroacetic acid (TCA) - methanol insoluble form. This insoluble radioactivity could not be solubilized by repeated acid extraction, although it was readily dissolved by alkali to give species that chromatography showed to be slightly more polar than ATP, ADP, and AMP, as well as adenosine and a nonmigrating form (Mowbray et al., 1984 op.cit.). Fitt and his co-workers have confirmed this rapid incorporation of substantial amount of [14c]adenosine into TCA - methanol insoluble material in perfused hearts, although they report that the product resembles RNA or polyA (Fitt et al., Mol. Cell. Biochem., 78, 37-46, 1987).
The characterisation of oligophosphoglyceroyl-ATP The TCA - methanol insoluble fractions of hearts previ-
ously labelled with [8-14~]adenosine were solubilized in detergent-containing buffer and subjected to phenol extrac- tion; the radioactive species in the nucleic acid rich fraction was purified by DEAE-cellulose chromatography under sterile conditions into one major and three minor species. The major species accounting for more than 70% of the total and two of the minor forms (together 16% of total) are closely related; while the fourth (10% of total) is a high molecular weight compound with a phosphate:purine ratio of 0.8, which comparison of [14c]uridylate and [14c]adenylate incorporation suggests is a rapidly syn- thesized species of RNA that represents about 4% of total heart RNA (Heyworth et al., Biochim. Biophys. Acta, 927, 158-162, 1987). While this latter would appear to be a can- didate for the rapidly labelled TCA insoluble material examined by Fitt et al. (op.cit.), this suggestion would be at variance with their report that it accounts for 70% of the original TCA insoluble radioactivity: based on the specific radioactivity of tissue adenylate, their data imply incorpora- tion of more than 0.4 pmol adenine per gram. Given that total RNA (mostly ribosomal) in a 1-g heart is estimated to contain around 1.1 pmol adenine, and polyA to have max- imally 40 nmol (see Heyworth et al., op.cit.), this would imply turnover of more than 30% of total tissue RNA in
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
SOU
TH
ER
N C
AL
IFO
RN
IA o
n 11
/09/
14Fo
r pe
rson
al u
se o
nly.

584 BIOCHEM. CELL BIOL. VOL. 69, 1991
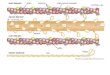
--I n FIG. 1. Structure of ~ligophosphoglycer~l-adenosine-triphosphate (oligo-PG-ATP).
a 20-min perfusion. The major radioactive species purified from our hearts proved to be at specific radioactivity equi- librium with tissue ATP and to have a phosphate:purine ratio of between 3.8 and 4.2:l (Mowbray et al., op.cit.). 3 1 ~ Nuclear magnetic resonance analysis showed that there were four distinct phosphate environments present in equal proportions: two correspond to the a and phosphate of ATP, while the third was near the Y, and the fourth appeared to be a typical phosphodiester (Hutchinson et al., Biochem. J. 234, 623-627, 1986). Selective digestion and chromatog- raphy of this compound allied to nmr showed that it con- tained equal proportions of adenosine and glyceric acid linked as a phosphodiester and we tentatively proposed a unit structure of ppp5 ' A3 ' 3pGri. Fast atom bombardment mass spectrometry produced a number of species in the high- field positive-ion spectrum. The largest corresponded exactly to a pentamer. Enzymic digestion supported the 3 ' diester link and we accordingly suggested the structure for the derivative (Fig. 1) and the name of oligophosphoglyceroy1- ATP (OPG-ATP). We have now synthesized by a chemically unambiguous route adenosyl-3 ' 3 phosphoglyceric acid, A3'3pGri (L. Wozniak, unpublished) and shown that it co-chromatographs on HPLC with the compound from rat heart (one of the minor species) that we had tentatively iden- tified as this. It also co-chromatographs with one of the species produced from purified OPG-ATP by mild alkaline hydrolysis (B. Patel, unpublished). Thus the proposed molecular form is on the way to being confirmed.
The problem of the extreme lability conferred by the mixed anhydride structure was overcome by our finding that it is remarkably stabilized in neutral solution by M ~ ~ + or ~ n ~ + above 2 mM (Basra and Mowbray, Biochem. Soc. Trans., 19,670-671, 1987). The monomer unit, 3-phosphoglyceroyl- Y-triphospho-5 '-adenosine, also appears to be stabilized by M ~ ~ + and exhibits unusual properties on ion-exchange chromatography (see below). The mechanism of this stabilization is at present unknown.
The distribution and cellular location of OPG-ATP In heart OPG-ATP appears to be present in both
myocytes and endothelial cells (see Lawson and Mowbray, op.cit.). Similar perfusion experiments to those with heart have revealed the presence of OPG-ATP in kidney (Hutchinson et al., Biochem. J., 240, 597-599, 1986) and in liver (Pate1 et al., Biochim. Biophys. Acta, 1074, 178-181,
1991). Liver appears to contain about 5-10 times more OPG- ATP per gram than heart, and cell fractionation experiments in 10 mM ~ g ~ + showed that 92% of the OPG-ATP co- sedimented with 89% of tissue succinate dehydrogenase; moreover, OPG-ATP, estimated to account for virtually all of the tissue pool, could be readily purified starting with crude liver mitochondrial fraction (Pate1 et al., 1991, op.cit.). While this still leaves uncertainty over a mitochon- drial location for OPG-ATP, the absence of significant amounts of 14c in the post-mitochondrial supernatant in incorporation experiments would argue against a lysosomal, peroxisomal, or endoplamic reticular site. However, in our search for enzymes metabolizing OPG-ATP, a specific exo-3 ' -phosphodiesterase has been partially purified from rat heart and liver mitochondria; it has been shown to be confined to the mitochondrial intermembrane space of liver (see below). This enzyme, as purified, contains tightly bound OPG-ATP, strongly suggesting that the oligomer is located also in the mitochondrial intermembrane space.
The metabolism of OPG-ATP Nonspecific nucleotidase-phosphatase digestion was
tackled by designing a crude assay that could be used to purify partially any cleavage activity with the aim of characterising the products and using this information to design a more specific assay. This assay relied on the ability of 14C-labelled oligomer to be precipitated in 50% aqueous ethanol and be collected on a Millipore filter, while degrada- tion products remain in solution. This has allowed the approximately 30-fold purification from washed liver mitochondria of a specific OPG-ATP exo-3 ' - phosphodiesterase with an M, of 85 000 (Pate1 et al., Biochem. J., 274, 275-279, 1991). The apparent K, for substrate (adenosine equivalents) is - 35 pM, consistent with the OPG-ATP content in cells, although the findings sug- gest that regulatory factors have been lost. Percoll gradient studies confirmed the activity as mitochondrial, and while the activity is latent in intact mitochondria, it is not associated with intact phosphorylating inner membrane vesicles, but is released during their preparation. Thus the enzyme is proposed to be located in the mitochondrial intermembrane space (Pate1 et al., 1991, op.cit.).
The single product of the reaction co-chromatographs on two-dimensional thin-layer chromatography with ATP, but was not recognised on enzymatic analysis as ATP. The
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
SOU
TH
ER
N C
AL
IFO
RN
IA o
n 11
/09/
14Fo
r pe
rson
al u
se o
nly.

COMMENTARY / COMMENTAIRE 585
phosphate:purine ratio of the chromatographically purified product was 3.8: 1 (ATP gave 2.7: 1). Somewhat surprisingly it elutes on ion-exchange HPLC between AMP and ADP. That it migrates in thin-layer chromatography in borate buffer suggests that the ribose 2' and 3' hydroxyls are free. The hypothesis that it might therefore be monomeric unit PG-ATP (p3Grilppp5 'A) was supported by the finding that digestion with snake venom phosphodiesterase (requires free 2' and 3 ' OH on ribose) yielded ADP.
Based on these findings, a sensitive enzyme assay could be devised for OPG-ATP by linking the 3'-phospho- diesterase product via snake venom diesterase to produce ADP and thence to ATP with phosphoenolpyruvate and pyruvate kinase; the ATP produced was detected using luciferin and luciferase. When this assay was attempted, a substantial blank value was observed suggesting that the OPG-ATP 3 ' -phosphodiesterase may be contaminated with substrate. This possibility was supported by the finding that dialysis of the enzyme in EDTA buffer at pH 8.5 produced substantial quantities of material absorbing light at 259 nm that crossed the membrane (B. Patel, unpublished).
Hence it would appear that OPG-ATP is an oligomer in mitochondrial intermembrane space, bound (at least) to a regulated enzyme that releases monomeric units of PG-ATP. All heart and liver extracts have hitherto unidentified purine derivatives that co-chromatograph on HPLC with PG-ATP (unpublished findings).
The possible function of OPG-ATP The true function of OPG-ATP remains conjectural. That
this oligomer shares mitochondrial intermembrane space, bound to an enzyme resistant to denaturation by trichloro- acetic acid at O°C (B. Patel, unpublished findings), with the similarly resistant enzymes adenylate kinase and creatine kinase suggests that it may have a central role in adenine nucleotide production or in homeostasis in oxidative cells.
It does not look suitable to be a high energy store, offering at best 5 s or so of contraction energy to a heart (about 50% of that available from creatine phosphate); indeed it increases in amount in very short-term cardiac ischaemia (Hutchinson et al., Biosci. Rep., 1,857-863, 1981; Fitt et al., Biosci. Rep., 5, 1061-1069, 1985), although it decreases as ischaemia continues (Fitt et al., Biosci. Rep., 5, 7-12, 1985).
With four phosphates per purine, it could provide a labile Pi resource; as noted above much of the acid extractable cell Pi is nmr invisible, consonant with an enzyme-bound intermembrane-space-located polyphosphate oligomer. As a source to replenish rapidly the relatively small amounts of adenosine sent as vasodilation signals to vascular smooth muscle in heart (see Berne, Circ. Res., 47, 807-813, 1980), it could prove an adequate store under physiological if not pathological conditions.
At variance with the concept of OPG-ATP as a store is its rapid and apparently complete turnover (to specific activ- ity equilibrium) in post-ischaemic hearts. It seems possible that this is an artifact of in vitro perfusion showing exten- sive perturbation of a regulatory system deprived of nor- mal in vivo controls. However, one nrnr study of hearts per- fused with glucose where the pulse-train was gated to the cardiac cycle allowed analysis of the major adenine nucleotides at set points during each contraction-relaxation. This suggested that four times more ATP became nmr invisible during systole to reappear during diastole than could be explained by oxidative phosphorylation (Fossel et al., Proc. Natl. Acad. Sci. U.S.A., 77, 3654-3658, 1980). Such a rapid exchange would be consistent with the radio- chemical equilibration that we observe and could imply some buffering role for OPG-ATP. Given that the requirement for life is the absolute maintenance, away from thermo- dynamic equilibrium of unidirectional metabolism by a high and nearly constant ATP:ADP ratio, some such mechanism would appear to be needed.
Bio
chem
. Cel
l Bio
l. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y U
NIV
OF
SOU
TH
ER
N C
AL
IFO
RN
IA o
n 11
/09/
14Fo
r pe
rson
al u
se o
nly.
Recommended