
1 3
J Biol Inorg ChemDOI 10.1007/s00775-015-1276-0
ORIGINAL PAPER
Lysozyme stability and amyloid fibrillization dependence on Hofmeister anions in acidic pH
Slavomíra Poniková1 · Andrea Antošová1 · Erna Demjén1 · Dagmar Sedláková1 · Jozef Marek1 · Rastislav Varhac2 · Zuzana Gažová1 · Erik Sedlák2,3
Received: 27 February 2015 / Accepted: 31 May 2015 © SBIC 2015
on fibrillization rate. In the presence of sodium chloride, lysozyme forms typical fibrils with elongated structure and with the secondary structure of the β-sheet. On the other hand, in the presence of both chaotropic perchlorate and kosmotropic sulfate anions, the fibrils form clusters with secondary structure of β-turn. Moreover, the acceleration of fibril formation is accompanied by decreased amount of the formed fibrils as indicated by ThT fluorescence. Taken together, our study shows Hofmeister effect of monovalent anions on: (1) lysozyme stability; (2) ability to accelerate nucleation phase of lysozyme fibrillization; (3) amount, and (4) conformational properties of the formed fibrils.
Keywords Protein stability · Acidic protein · Hofmeister series · Fibrillization kinetics
Introduction
Numerous proteins as well as peptides have been found to form amyloid aggregates accompanying an onset of amyloid-related diseases as Alzheimer’s disease or diabe-tes mellitus type II [1, 2]. Understanding of the conforma-tional transition to amyloid aggregates would bring benefit not only to medicine, but also to various biotechnological applications. In fact, formation of amyloid aggregates can be, from a mechanistic point of view, considered as a par-ticular case of aggregation, i.e., reaction that often com-plicates handling, storage, and production of peptides and proteins.
The process of amyloid fibrillization, aggregation and in general intermolecular protein–protein interaction of pro-teins depends on two factors: (1) intrinsic properties of pro-tein molecules such as net charge, hydrophobicity of their surfaces, conformational stability, and intrinsic propensity
Abstract We have explored an effect of Hofmeister ani-ons, Na2SO4, NaCl, NaBr, NaNO3, NaSCN and NaClO4, on stability and amyloid fibrillization of hen egg white lysozyme at pH 2.7. The stability of the protein was ana-lyzed by differential scanning calorimetry. The Hofmeister effect of the anions was assessed by the parameter dTtrs/d[anion] (Ttrs, transition temperature). We show that dTtrs/d[anion] correlates with anion surface tension effects and anion partition coefficients indicating direct interactions between anions and lysozyme. The kinetic of amyloid fibrillization of lysozyme was followed by Thioflavin T (ThT) fluorescence. Negative correlation between dTtrs/d[anion] and the nucleation rate of fibrillization in the presence of monovalent anions indicates specific effect of anions on fibrillization rate of lysozyme. The efficiency of monovalent anions to accelerate fibrillization corre-lates with inverse Hofmeister series. The far-UV circular dichroism spectroscopy and atomic force microscopy find-ings show that conformational properties of fibrils depend
Electronic supplementary material The online version of this article (doi:10.1007/s00775-015-1276-0) contains supplementary material, which is available to authorized users.
* Zuzana Gažová [email protected]
* Erik Sedlák [email protected]
1 Department of Biophysics, Institute of Experimental Physics Slovak Academy of Sciences, Watsonova 47, 04001 Košice, Slovakia
2 Department of Biochemistry, P.J. Šafárik University, Moyzesova 11, 04001 Košice, Slovakia
3 Centre for Interdisciplinary Biosciences, P.J. Šafárik University, Jesenná 5, 04001 Košice, Slovakia

J Biol Inorg Chem
1 3
of protein sequence to fibrillize/aggregate (form intermo-lecular complexes) and (2) environmental factors such as pH, temperature, type and concentration of salts, and the presence of additives.
Human lysozyme is one of the amyloidogenic proteins associated with serious disease—autosomal hereditary sys-temic amyloidosis [3]. This disease is triggered by fibrilliza-tion of mutated protein which forms amyloid deposits in the liver and kidney. Lysozyme amyloid aggregation was also observed in vitro at conditions in which wild-type protein is partially unfolded [4]. Hen egg white lysozyme (HEWL), important food protein, is homologous to human lysozyme. It has been shown that HEWL also forms amyloid aggregates in vitro at slightly modified conditions in comparison with those which fibrillizes human lysozyme [5, 6]. HEWL is very well characterized and intensively studied protein as regards its structural properties [7–9]. The protein forms amyloid aggre-gates at various conditions that have in common the fact that its native conformation is destabilized due to: (1) heat shock followed by incubation at pH 2.0 [5]; (2) presence of denatur-ing additives [10–12]; (3) continual incubation at 57 °C and pH 2.0 [6], and (4) full reduction of the protein [13].
HEWL is an α + β protein with ~35 % helical resi-dues, and ~10 % β-strands with four disulfide bonds. Its 129 amino acid residues fold into two structural domains: the α-domain consists of four α-helices and a 310 helix; the β-domain includes a long triple-stranded β-sheet, a short double stranded β-sheet, a 310 helix, and a long loop [14]. HEWL is basic protein with pI ~11.2 with net charge +19 at pH < 3 (11 from arginines, 6 from lysines, 1 from histi-dine, and 1 from N-terminal amine group) [15, 16]. Ther-mal transition of HEWL at neutral pH is a highly coopera-tive and reversible two-state transition with Ttrs ~ 73 °C [17, 18]. At acidic pH, at which the fibrillization of the protein is studied, its conformational properties as well as its stability will very likely strongly depend on the present anions. This dependence might be further modulated by the type of the salts, e.g., in dependence on their position in the Hofmeister series of salts. The Hofmeister series ranks the relative influ-ence of ions on the physical behavior of biomacromolecules as well as a wide variety of aqueous processes. Interestingly, although the effect of salt ions on protein aggregation known as the Hofmeister effect is known for more than a century [19, 20], its influence on the process of amyloid fibril for-mation is unclear. Hofmeister effect has been recently thor-oughly reviewed in several reviews [21–26]. This behavior is more pronounced for anions than cations. The typical order, the usually quoted, for the anion series is as follows [22]:
Anions on the left are called kosmotropes, which tend to precipitate proteins from solution and prevent protein unfolding, whereas anions on the right are chaotropes,
SO42−
> OH−
> F−
> Cl−
> Br−
> NO3−
> I−
> SCN−
> ClO−
4.
which increase solubility and promote the denaturation of proteins. Chloride is usually considered the dividing line between these two types of behavior.
Particular attention has been paid over the past decade to ion-partitioning behavior at the air/water interface. Molecu-lar simulations of air/electrolyte systems in the presence of salts suggest that larger, less hydrated anions have a pref-erence for the interfacial region [27, 28]. A salt ion-parti-tioning model has been used to interpret Hofmeister effect at the air/water and protein/water interfaces [29–31]. These studies further supported previous findings that more chao-tropic anions generally partition to the interface to a greater extent than other species with implication for solubility, crystallization and aggregation of proteins [21, 32–34].
Moreover, salt ions represent particularly important group of environmental factors ubiquitously present and tightly regulated in living cells. They may affect protein amyloid fibrillization processes through nonspecific effect by Debye–Hückel screening and/or through specific direct interaction with the protein [35]. Recent studies of the influence of salts on fibrillization provide complex pic-ture indicating that: (1) amyloid fibril formation of vari-ous proteins is accelerated by salts [36–42] with a very recently reported exception when in the presence of sul-fate anions fibrillization was decelerated [43]; (2) cati-ons have only minor impact on kinetics of fibrillization in comparison with anions [37, 41, 42], and (3) kinetics of fibrillization depend on salt ions in complex way both on type of ions and on its concentration [44]. Apparently, amyloid fibril formation is complex interplay among Debye–Hückel screening, ion selectivity, and Hofmeister effect [35, 39].
In the present work, we addressed an effect of various sodium salts, in a range of physiological concentrations of salts (80–200 mM), chosen according their position in Hofmeister series on HEWL stability and its propensity to form amyloid fibrils at acidic pH (pH 2.7) and high tem-perature (at 65 °C). Our results show clear dependence of thermal stability of HEWL on Hofmeister anions as well as fibrillization of HEWL on monovalent anions. We further show that amount of fibrils is negatively affected by fast kinetics of fibril formation in the presence of chaotropic salts.
Materials and methods
Chemicals and proteins
Lysozyme from hen egg white (lyophilized powder, lot number BCBB3704, ~10,0000 units mg−1 protein) were purchased from Fluka. Thioflavin T (ThT), 8-anilino-1-naphthalenesulfonic acid (ANS), glycine, NaCl, NaClO4,

J Biol Inorg Chem
1 3
NaBr, NaSCN, NaNO3 were obtained from the Sigma Chemical Company (St Louis, MO), Na2SO4 were obtained from Merck (Darmstadt, Germany). The lysozyme dis-solved in 70 mM glycine, pH 2.7 was dialyzed overnight, and its concentration was determined spectrophotometri-cally on Jasco V-630 spectrophotometer, using extinction coefficient at 280 nm equal to 38,940 L mol−1 cm−1.
Lysozyme amyloid aggregation in vitro
Lysozyme amyloid aggregates were prepared by incubation of the lysozyme (5 μM) in 70 mM glycine buffer, pH 2.7, containing salt (NaCl, Na2SO4, NaClO4, NaBr, NaSCN or NaNO3) at 65 °C and constant stirring (1200 rpm/min). In the absence of agitation, fibrillization is not observed even after incubation at given conditions for several hours. The final salt concentration in solution was 80, 140 or 200. For-mation of lysozyme aggregates was monitored with ThT assay and atomic force microscopy. The aliquots selected at given time intervals were monitored by ThT fluores-cence assay to determine the kinetics of lysozyme amyloid aggregation.
Fluorescence measurements
The lysozyme amyloid aggregation was detected by a characteristic increase in ThT fluorescence intensity reflecting the presence of amyloid fibrils. ThT was added to lysozyme samples (5 μM) to final concentration of 5 μM, and samples were then incubated at 37 °C for 45 min. The fluorescence intensity was measured using a 96-well plate by a Synergy Mx (BioTek) spectrofluorim-eter. The excitation was set at 440 nm, and the emission was recorded at 485 nm. The excitation and emission slits were adjusted to 9.0/9.0 nm, and the top probe vertical offset was 6 mm.
Thermally induced conformational change of lysozyme in the presence of ANS probe was monitored with a Var-ian Cary Eclipse fluorescence spectrophotometer equipped with a Peltier multicell holder. The temperature was changed from 25 to 95 °C with a heating rate of 1 °C/min. The excitation wavelength was 390 nm. The emission was collected at 520 nm. For determination of apparent transi-tion temperatures for the two-step thermal transition, the following equation for a two-step equilibrium transition was used [45]:
where F is the observed ellipticity, FN and FD are the fluo-rescence values of native and denatured states, respectively; mN and mD are slopes that describe the fluorescence sig-nal dependence of the native and denatured states with the
(1)F =
FN + mNT + (FD + mDT)·e−�H(1−T/Tm)/(RT)
1+ e−�H(1−T/Tm)/(RT),
temperature, respectively; ΔH is an apparent enthalpy, and Tm is an apparent temperature of the transitions.
Excitation and emission slits were set to 10 and 5 nm, respectively. The protein concentration was 5 μM. Con-centration of ANS was 0.2 mM. Measurements were per-formed in 70 mM glycine buffer (pH 2.7) in the absence and in the presence of 0.1, 0.2 and 0.3 M salts (Na2SO4, NaCl, NaNO3, NaBr, NaSCN, NaClO4).
Circular dichroism measurements
A Jasco J-810 spectropolarimeter (Tokyo, Japan) equipped with a Peltier-type thermostated single cell holder (PTC-423S) was used for circular dichroism (CD) measurements. Ellipticity was measured in the range 200–250 nm in a 1-mm pathlength quartz cuvette. The temperature was kept constant at 25 °C. Each spectrum is an average of 8 scans. All spectra were background-corrected and converted to mean residue ellipticity (deg cm2/dmol). Due to lower qual-ity of CD spectra obtained at 5 μM HEWL, we performed measurements of CD spectra also at 20 μM HEWL. Com-parison of mean residue ellipticity values of the CD spec-tra of HEWL fibrils formed at 5 and 20 μM of the protein point to an independence of the secondary structure of HEWL fibrils on protein concentration within the concen-tration range 5–20 μM in 70 mM glycine buffer, pH 2.7 (Fig. S1).
Differential scanning calorimetry measurements
Differential scanning calorimetry (DSC) experiments were performed on a VP-DSC differential scanning microcalo-rimeter (Microcal, Northampton, MA) at a scan rate of 1.5 °C/min. HEWL concentration was 105 μM in 70 mM glycine buffer, pH 2.7. Measurements were carried out under a constant overpressure of 1.5 atm. A background scan, collected with buffer in both cells, was subtracted from each sample scan. The reversibility of the transitions was assessed by the reproducibility of the calorimetric trace in a second heating cycle performed immediately after cooling from the first scan. Excess heat capacity curves were plotted using Origin software supplied by Microcal.
Atomic force microscopy (AFM)
Samples were deposited by drop casting on the freshly cleaved mica surface. After 10 min adsorption, the sam-ples were washed three times with ultrapure water and left to dry. The protein concentration of 5 μM was used. AFM images were taken by a scanning probe microscope (Veeco di Innova, Bruker AXS Inc., Madison, USA) in a tapping mode using uncoated silicon cantilevers TESPA, un-mounted with force constant 42 N/m and nominal

J Biol Inorg Chem
1 3
resonance frequency ~320 kHz, Al Reflective Coating (Bruker AFM Probes, Camarillo, USA). The resolution of image was 512 pixels per line (512 × 512 pixels/image) and scan rate 0.5–0.75 kHz. No smoothing or noise reduc-tion was applied.
Analysis of kinetics data of amyloid fibril formation
The kinetics of lysozyme fibrillization represented by ThT fluorescence were plotted as a function of time and fitted by a Boltzmann sigmoidal described by following equation:
where y is the fluorescence intensity, A1 and A2 are the initial and the final values of ThT fluorescence intensity, respectively, x is the time, x0 is the time at half-height of fluorescence (thalf), and τ is the time constant (the width of the range where the y value changes the most significantly). Equation (2) was used to extract a number of empiri-cal parameters including the lag time (tlag = x0 − 2τ), the apparent fibril growth rate (k = 1/τ) and span (span = A2 − A1) [46]. All calculations were performed in origin.
Results
To address Hofmeister effect in stability and amyloid fibrillization of HEWL, the effect of six sodium salts of Hofmeister series: sulfate, chloride, bromide, nitrate, thio-cyanate, and perchlorate have been studied. The fibril for-mation of HEWL was examined at acidic pH, pH 2.7, at 65 °C with agitation at 200 mM concentration of the salts. At such relatively high ion concentrations, Hofmeister effect dominates and is demonstrated by a linear depend-ence of the transition temperature of proteins as a function of ion concentration [47, 48]. On the other hand, at even higher concentrations of salt, the kinetic of fibrillization was too fast with problematic reproducibility for our set up of experiments.
Kinetics of amyloid fibril formation of HEWL
The kinetics of fibril formation of HEWL was followed by maximal fluorescence intensity of Thioflavin T (ThT) which is specific fluorescence probe for amyloid fibrils and its binding is independent of the primary structure of the protein [49, 50]. In the presence of amyloid fibrils, ThT gives rise to a new excitation maximum at 450 nm and enhanced emission at 482 nm, whereas unbound ThT is essentially nonfluorescent at these wavelengths. The
(2)y =A2 − A1
1+ exp[
x0−xτ
] + A1,
HEWL aggregation at acidic pH in all studied salts is characterized by three phases typical for amyloid nuclei-dependent kinetics of fibrillization: (1) lag phase or nucle-ation phase; (2) growth phase and (3) saturation phase (Fig. 1).
The kinetics of ThT fluorescence analyzed accord-ing the Boltzmann sigmoidal curve (Eq. 2) show that the fibril formation is dependent on the salt type. Obtained fit-ting parameters indicate the tendency in thalf and tlag values and in amplitude of ThT fluorescence on position of the anions in Hofmeister series (Table 1). For further analysis we chose three salts based on their positions in Hofmeister series: kosmotropic sodium sulfate, neutral (from the point of view of the Hofmeister effect) sodium chloride, and cha-otropic sodium perchlorate. Due to relatively good correla-tion between the values of thalf and tlag (Fig. S2), we used for further analysis only tlag values that contain also infor-mation about the growth phase (tlag = x0 − 2τ).
Characterization of HEWL amyloid fibrils formed in the presence of salts
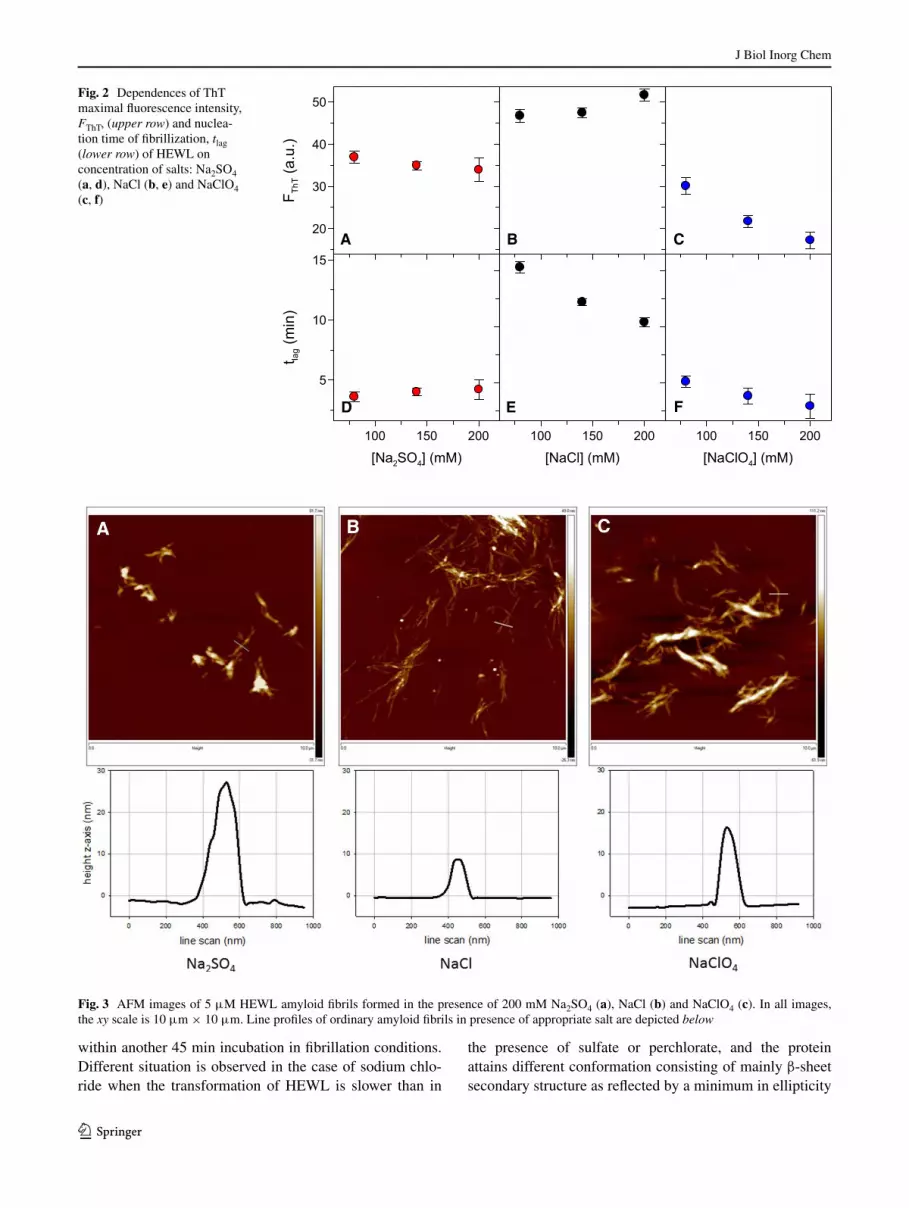
Closer analysis of fibril formation of HEWL on salt con-centration indicate strong dependence of ThT maximal fluorescence intensity FThT and tlag values on concentra-tion of sodium chloride and sodium perchlorate while these parameters are relatively independent of concentration of sodium sulfate (Fig. 2; Table S1). In the case of chloride and perchlorate salts, the values of tlag steeply decrease (by ~30–40 %) with increased salt concentration from 80 to 200 mM (Fig. 2e, f; Table S1). On the other hand, ThT maximal fluorescence has tendency to rise with chloride concentration (Fig. 2b), but decreases with increasing con-centration of sodium perchlorate (Fig. 2c).
Fibrils formed in the presence of 200 mM sodium sul-fate, chloride, and perchlorate were characterized using AFM (Fig. 3). The salts have significant impact on the mor-phology of formed aggregates. In the presence of sodium chloride, lysozyme produces thin, long, and unbranched fibrils (Fig. 3b). The ordinary amyloid fibril (in Fig. 3a marked with white line perpendicular to the fibril) was cho-sen to determine the height of the fibrils. The fibril profile is shown in Fig. 3b (below AFM image) corresponding to the height of the fibril equal ~8 nm. Lysozyme fibrillization, in the presence of sodium sulfate and sodium perchlorate leads to formation of thicker and shorter fibrils with higher tendency to laterally associate into clusters (Fig. 3a, c). The heights of the fibrils obtained from line profiles were dif-ferent in comparison to fibrils formed in presence of NaCl. For fibrils formed in sodium perchlorate the height is twice (~16 nm) than in the case of sodium chloride fibrils. Even higher (~27 nm) were fibrils prepared in the presence of sulfate anions.

J Biol Inorg Chem
1 3
CD spectra in the far-UV region provide important information regarding the secondary structure of proteins. Unfortunately, due to high absorption in the far-UV region, the effect of anions such as thiocyanate, bromide, and nitrate could not be analyzed. CD spectrum of HEWL at pH 2.7 in the presence of all other sodium salts (sulfate, chlo-ride, and perchlorate) (at time 0 min) in the far-UV region indicates the presence of α-helical structure as reflected by pronounced ellipticity signals at 208 and 222 nm (Fig. 4).
After 15 min of incubation at 65 °C with agitation CD spectra indicates that HEWL in the presence of sulfate and perchlorate anions is fully transformed into conformation containing mainly β-turn as indicated by negative elliptic-ity at ~225 nm and positive signal at ~205 nm [51, 52]. However, the conformation of lysozyme in the presence of perchlorate is apparently slightly different from that in the presence of sulfate as indicated from more positive ellip-ticity at ~212 nm. The ellipticity signal is relatively stable
Fig. 1 Kinetics of amyloid fibril formation of HEWL incu-bated with 200 mM Hofmeister salts at pH 2.7 and temperature 65 °C with agitation detected by ThT assay. Error bars indicate standard deviation of three independent measurements
60
40
20
060
40
20
0
Na2SO4 NaCl
NaBr NaNO3
403020100
60
40
20
0
Time (min)
NaClO4
403020100
Time (min)
NaSCN
ThT
fluor
esce
nce
(a.u
. x10
3 )
Table 1 Effect of different salts at 200 mM concentration on aggregation kinetics of lysozyme at pH 2.7, at 65 °C with agitation. Parameters were extracted using Boltzmann fit (detailed description is provided in the section “Materials and methods”)
a Adjusted R-square quantifies how well a model fits the data
Salt Na2SO4 NaCl NaNO3 NaBr NaClO4 NaSCN
thalf(min) 7.0 ± 0.6 11.3 ± 0.2 4.3 ± 0.1 4.8 ± 0.2 5.9 ± 0.5 3.9 ± 0.4
tlag (min) 4.0 ± 0.5 10.3 ± 0.4 3.1 ± 0.2 3.6 ± 0.3 2.8 ± 0.4 1.8 ± 0.3
k (min−1) 0.63 ± 0.14 1.99 ± 0.53 1.67 ± 0.34 1.42 ± 0.44 0.63 ± 0.20 1.00 ± 0.21
Span 33,900 ± 2800 51,600 ± 1400 34,500 ± 1000 39,500 ± 2700 17,200 ± 2000 21,700 ± 2000
Adj. R2a 0.975 0.982 0.995 0.979 0.983 0.973
J Biol Inorg Chem
1 3
within another 45 min incubation in fibrillation conditions. Different situation is observed in the case of sodium chlo-ride when the transformation of HEWL is slower than in
the presence of sulfate or perchlorate, and the protein attains different conformation consisting of mainly β-sheet secondary structure as reflected by a minimum in ellipticity
Fig. 2 Dependences of ThT maximal fluorescence intensity, FThT, (upper row) and nuclea-tion time of fibrillization, tlag (lower row) of HEWL on concentration of salts: Na2SO4 (a, d), NaCl (b, e) and NaClO4 (c, f)
200150100
15
10
5
[Na2SO4] (mM)
t lag (
min
)
200150100
[NaCl] (mM)200150100
[NaClO4] (mM)
B CA
ED F
50
40
30
20
F ThT
(a.u
.)
Fig. 3 AFM images of 5 μM HEWL amyloid fibrils formed in the presence of 200 mM Na2SO4 (a), NaCl (b) and NaClO4 (c). In all images, the xy scale is 10 μm × 10 μm. Line profiles of ordinary amyloid fibrils in presence of appropriate salt are depicted below

J Biol Inorg Chem
1 3
observed at ~220 nm (Fig. 4). CD spectra strongly indicate that fibrillization process of HEWL in the presence of chlo-ride anions differs structurally in comparison with kosmo-tropic sulfate and chaotropic perchlorate anions.
Thermal stability of HEWL in the presence of salts
Hofmeister anions significantly affect stability of protein [18, 48]. An effect of anions on stability of HEWL at acidic pH was studied by DSC (Fig. 5) and ANS fluorescence (Fig. S3).
Thermal denaturation of HEWL at pH 2.7 is all-or-none transition in buffer as well as in the presence of all inves-tigated salts. This follows from the fact that value of the ratio of calorimetric and van’t Hoff enthalpies (obtained from DSC experiment) is close to ~1 (Table S2) what is prerequisite for two-state transition [17]. The thermal sta-bility of HEWL expressed by temperatures of transition, obtained by both above methods, are identical (within an
experimental error) indicating an independence of thermal transition on the protein concentration within the concentra-tion range 5–105 μM. This dependence of the temperatures of thermal transitions on salts follows an order of anions in Hofmeister series (from the least to the most stable): thio-cyanate < perchlorate < nitrate < bromide < chloride < sul-fate. Interestingly, stability of HEWL is decreased in all salts besides sodium sulfate in comparison with the protein stability in the buffer. More detailed analysis of the ther-mal stability of HEWL dependent on salt concentration fur-ther support the above findings. HEWL is destabilized with increasing concentration of all studied salts besides sulfate in a monotone way. This conclusion follows from the plots of dependence of transition temperatures expressed either as dependence on concentration (Fig. 6). Under assump-tion of two-state nature of thermal transitions, the transi-tion temperature dependence on anion concentration allows
Fig. 4 Dependence of mean residue ellipticities of HEWL on incubation time in the various salts (200 mM) of Hofmeister series at fibrilliza-tion conditions: 0 min (black curve), 15 min (blue), 30 min (green), and 60 min (red)
240220200
15
10
5
0
-5
-10[
] MR
W (x
103 d
eg.c
m2 .d
mol
-1)
240220200
15
10
5
0
-5
-10
240220200
15
10
5
0
-5
-10
W a v e l e n g t h (nm)
Na2SO4 NaCl NaClO4
Fig. 5 DSC scans of HEWL in the presence of sodium salts of: thiocyanate (cyan), perchlorate (blue), nitrate (green), bromide (magenta), chloride (black) and sulfate (red). DSC scan of HEWL in the buffer is shown in white circles. The measurements were per-formed in 70 mM glycine buffer, pH 2.7
Fig. 6 Dependence of transition temperatures of HEWL on salt concentration. Salt: NaSCN (cyan), NaClO4 (blue), NaNO3 (green), NaBr (magenta), NaCl (black) and Na2SO4 (red). The symbol rep-resented by white circle corresponds to transition temperature of lysozyme in the buffer. All data are obtained from DSC measure-ments with 105 μM HEWL in 70 mM glycine buffer, pH 2.7

J Biol Inorg Chem
1 3
to estimate the number of anions that are bound (negative value) or released (positive value) upon protein unfolding by applying the following equation [53]:
where Δνtrs is a number of bound or released anions, ΔHcal(Ttrs) is calorimetric enthalpy of the thermal transition, Ttrs is transition temperature (midpoint of thermal transi-tion) in Kelvin, and R is the gas constant. Denaturation of HEWL in the presence of salts (at 200 mM concentration) is found to be accompanied by: (1) release of ~1 sulfate anion; (2) neither release or binding of chloride, and (3) binding of ~1–2 anions in the case of chaotropic salts (Table S2).
With increasing concentration of salts, thermal denatur-ation of HEWL reaches a point when the reversibility of denaturation abruptly changes into irreversible process due to aggregation of its thermally denatured form. The aggre-gation is clearly indicated by steep decrease in a measured signal in DSC, typical feature of aggregation processes in DSC, as well as by eye inspection of DSC sample after the measurement. The aggregation of HEWL in the process of thermal denaturation is dependent both on salt concentra-tion and the type of anion. The order of efficiency of anions to trigger aggregation does not follow the Hofmeister series (order of efficiency of anions to trigger aggregation of ther-mally denatured state of HEWL): thiocyanate~perchlorate > sulfate~nitrate > bromide~chloride. The aggregation of thermally denatured state of HEWL is not accompanied by formation of amyloid fibrils as it is indicated by an absence of ThT fluorescence.
The slope of the linear dependence of Ttrs against the concentration of anions, i.e., dTtrs/d[anion], is a measure of the efficiency of each given anion to de/stabilize proteins [48]. This parameter has been used below in attempt to cor-relate various physical properties of the anions with their Hofmeister effect on HEWL.
Correlation of Hofmeister effect of anions on HEWL with the anion intrinsic properties
Detailed analysis of Hofmeister effect of anions on HEWL thermal denaturation with their inherent properties such as charge density, surface tension effects, partition coefficient, ion polarizability, hydration entropy, and viscosity B-coef-ficient, revealed strong correlation between the parameter dTtrs/d[anion] and the anions properties such as charge den-sity, surface tension and partition coefficient (Fig. 7) and weak or no obvious correlation with any of the other prop-erties of anions (Fig. S4). Interestingly, analogous findings were reported with two unrelated proteins such as horse heart cytochrome c and apoflavodoxin from Desulfovibrio desulfuricans [50].
(3)�νtrs =�Hcal(Ttrs)
RT2trs
dTtrs
d ln ([anion]),
Correlation of Hofmeister effect of anions and kinetics of fibril formation of HEWL
A relationship between Hofmeister effect of anions, rep-resented by the parameter dTtrs/d[anion], and the kinetics parameters of fibrillization of HEWL has been also exam-ined. In fact, it was found that the de/stabilization effect of anions, dTtrs/d[anion], was in good correlation with the parameters characterizing fibril formation such as 1/tlag
0.03
0.025
0.02
0.015
char
ge d
ensi
ty
2.5
2
1.5
1
0.5su
rface
tens
ion
100-10-20-30
2
1.5
1
0.5
0
dTtrs/d[ion] (oC/mol)
parti
tion
coef
ficie
nt
R=0.9957
R=0.9961
R=-0.9931
Fig. 7 Correlation between intrinsic properties of studied anions such as charge density, surface tension effect and partition coeffi-cient on parameter dTtrs/d[ion]. The values of parameters dTtrs/d[ion] were obtained from measurements described in this work and listed in Table S2. Parameters of intrinsic properties of anions were taken from Marcus (1997) [76]

J Biol Inorg Chem
1 3
that reflects rate of nucleation and ThT maximal fluores-cence intensity reflecting amount of formed fibrils (Fig. 8a, b). However, there are apparent outliers represented by parameters corresponding to the only divalent anion, sul-fate. Excluding sulfate anion from the correlations, there is good linear correlation (correlation coefficient R = 0.7453) between parameters 1/tlag and dTtrs/d[anion] and very strong linear correlation (R = 0.9848) between maximal ThT fluorescence and dTtrs/d[anion]. Interestingly, there is good linear correlation (R = 0.8263) between maxi-mal ThT fluorescence and 1/tlag values indicating that an amount of formed fibrils decreases with increasing rate of nucleation (Fig. 7c).
Discussion
Formation of amyloid fibrils is a complex process depend-ing on both intrinsic properties of proteins and peptides and properties of surrounding solvent. In the present work, we have explored the effect of Hofmeister anions on stability and fibrillization of small globular protein—hen egg white lysozyme (HEWL). Despite long-history of intensive stud-ies of Hofmeister effect on protein properties, its origin is still quite limited. Investigation of effect of Hofmeister ani-ons on fibrillization of HEWL is also complicated by the fact that anions influence both protein surface properties and protein stability [47, 48]. This is in contrast with pep-tides such as Aβ(1–40), glucagon, amylin, and insulin in which case the parameter of “protein stability” can be con-sidered as irrelevant [39, 40, 46, 54]. The present work is a contribution to understanding the effect of anions in pro-tein stability and protein fibrillization at acidic pH, i.e., at conditions at which numerous proteins readily form amy-loid fibrils. Moreover, at such pH protein surfaces contain only positively charged side chains that should facilitate to assess a role of anions. Our results show (1) clear depend-ence of thermodynamic stability of HEWL; (2) an apparent but more complex dependence of a parameter of kinetics of fibrillization—rate of nucleation, and (3) dependence of amount of formed fibrils on intrinsic properties of Hofmeister anions in the studied conditions.
Hofmeister effect on HEWL stability at acidic pH
The Hofmeister effect of anions, assessed by the param-eter dTtrs/d[anion], on HEWL stability can be ordered as follows (from stabilization to destabilization effect): sul-fate > chloride > bromide > nitrate > thiocyanate > per-chlorate (Fig. 6; Table S2). Our results show the strong
0.6
0.4
0.2
0
1/t la
g (m
in-1)
100-10-20-30
50
40
30
20
10
dTtrs/d[ion] (oC/mol)
ThT
fluor
esce
nce
(a.u
.)
0.60.40.20
50
40
30
20
10
1/tlag (min-1)
ThT
fluor
esce
nce
(a.u
.)
NaClO4
NaSCN
NaNO3
NaBr
NaCl
Na2SO4
R=-0.8263
A
B
C
Fig. 8 Correlation between parameters obtained from kinetics of HEWL fibril formation in the presence of different salts, rate of nucleation 1/tlag (a) and maximal ThT fluorescence (b), and param-eters characterizing Hofmeister effect of the salts, dTtrs/d[ion]. Linear correlation in the box a and b (excluding parameters for sulfate) are 0.7453 and 0.9848, respectively. c Correlation between maximal ThT fluorescence and 1/tlag (R = 0.8263). The plotted parameters are listed in Tables 1 and S2

J Biol Inorg Chem
1 3
correlation between the parameter dTtrs/d[anion] and intrinsic properties of Hofmeister anions such as ion partition coefficients between surface and bulk water, ion surface tension effects and charge density of anions (Fig. 7) in agreement with previous study [48]. Although anions influence on protein stability might be exercised by direct interaction and/or indirect effects, recent stud-ies strongly indicate that the effects of ions arise from the closest hydration shell with negligible effects on the global hydrogen-bonding network of water [55–58]. These findings are in agreement with observed cor-relation of thermal stability of HEWL with ion parti-tion coefficient and ion surface tension effects (Fig. 7). Although the partition coefficient expresses the distribu-tion of ions in the first layer at the air/water interface in comparison with the bulk water, one can assume that dis-tribution of ions at protein/water will be similar. This is reasonable to assume for exposed hydrophobic patches on the protein with low polarity and dielectric constant. Such assumption is consistent with the observed effect of chaotropic ions at aqueous/oil interfaces, where more chaotropic ions such as iodide and thiocyanate decrease surface tension, while chloride increases it [59, 60]. Similar destabilization effect of anions on unrelated pro-teins with oppositely charged surfaces, cytochrome c and apoflavodoxin, led the authors to conclude that destabi-lization effect of anions is mediated by their interaction with peptide bond [48]. Comparison of dTtrs/d[anion] values obtained for those proteins for individual anions with the dTtrs/d[anion] values in the case of HEWL indi-cate stronger destabilization of all anions on HEWL. We believe that the more pronounced destabilization effect can be explained by two factors that differ HEWL from cytochrome c and apoflavodoxin: (1) HEWL contains only positive charges at pH 2.7 [15] while both pro-teins contain a mixture of positive and negative charges on their surfaces, even with high net positive charge, cytochrome c, or negative charge, apoflavodoxin, and (2) HEWL is partially unfolded at pH 2.7, i.e., its surface is apparently more hydrophobic providing thus larger area for interaction with chaotropic anions, than the proteins in their native compact folded conformations in studied conditions. In fact, some NMR studies as well as molec-ular dynamic simulations indicate that polarizable large chaotropic anions may interact with nonpolar groups on protein surfaces [28, 61–63]. At this point it is necessary to stress that in accordance with previous results [48] we did not observe a correlation between dTtrs/d[anion] and polarizability of ions (Fig. S4). Lack of correlation between analogous parameters at comparable anion con-centrations (<300 mM) was also observed in the study of anions effects on cloud-point temperature for the liquid–liquid phase transition of lysozyme [16].
Hofmeister anions effect on fibrillization of HEWL at acidic pH
Studies of physicochemical properties of an amyloidogenic variant of human lysozyme strongly indicate that its fibrilli-zation is dependent on its decreased stability [1, 64–67]. Ability of lysozyme as well as other globular proteins to form fibrils in vitro only at such harsh conditions strongly points out to the importance of proteins being (partially) unfolded to begin to form fibrils [66, 68]. The fact that pro-teins are able to form higher structures at conditions when high positive charge of protein surface forms high barrier for interaction points out to high affinity and specificity of proteins towards each other at “pre-fibril” conformation. Due to screening positive charges on the surface of pro-teins, presence of anions should significantly accelerate formation of fibrils, particularly to shorten its nucleation phase. In fact, in the case of HEWL the nucleation phase has been shortened in the presence of anions by several orders of magnitude from tens of hours to the minutes. This observation is in agreement with previous studies that show significant acceleration of fibrillization process of proteins and peptides in the presence of salts [36–42]. The pres-ence of chaotropic anions due to their destabilization effect should further accelerate the fibrillization process. Indeed, there is good correlation between the rate of nucleation and Hofmeister effect of chaotropic anions (Fig. 8a) which is in agreement with preferential accumulation of these ani-ons at lysozyme surface measured at pH 5.0 [69]. Although sulfate anions similarly accelerate fibrillization process and bind to lysozyme surface [66], obtained parameters do not match linear dependences formed by monovalent ani-ons as shown in Fig. 8a, b. From all studied salts, only in the presence of kosmotropic sulfate anions transition tem-perature of HEWL is higher (66.2 °C) than the tempera-ture (~65 °C) at which fibril formation has been followed (Table S2). This further indicates the important role of (par-tially) unfolded form of HEWL for its fibrillization. How-ever, one cannot exclude a possibility of other explanation of the effect of sulfate such as stronger screening effect due to its divalent nature, modification of protein–protein interaction [70], and/or modification of aggregation path-way [43]. Interestingly, a strong correlation between ThT fluorescence and the parameter dTtrs/d[anion] for monova-lent anions (Fig. 8b) indicates that more chaotropic anions decrease amount of fibrils detected by ThT fluorescence. Based on this as well as previous studies [47, 48, 69] one can reasonably assume that anions with more destabiliza-tion effect interact with proteins with higher affinity and/or more anion molecules per protein (Table S2) [33, 34]. Surprisingly, CD spectra show that the secondary structure of fibrils formed in the presence of chaotropic perchlorate as well as of kosmotropic sulfate differs from the secondary

J Biol Inorg Chem
1 3
structure of the fibrils formed in the presence of chloride anions. While in the presence of sodium chloride the sec-ondary structure represents type D (according Pedersen et al. classification of secondary structure of fibrils) [54], i.e., classic β-sheet CD, the fibrils formed in the presence of perchlorate and sulfate represents “new” (unclassified) secondary structure of fibrils resembling β-turn (Fig. 4) [51, 52]. Structures visualized by AFM (Fig. 3) also point out to different fibrils formed in the presence of chlo-ride on one side, and perchlorate and sulfate on the other side. These observations are summarized in the following scheme (Fig. 9).
Plotting of maximal ThT fluorescence vs nucleation rate leads to a good correlation (R = −0.8263). This correla-tion suggests that speed of fibril formation is reciprocally related to an amount of formed fibrils (Fig. 8c).
Apparently, aggregation and fibrillization are two com-peting processes [38]. Formation of fibrils at very acidic pH is not accompanied by formation of aggregates due to
strong repulsive electrostatic interaction between protein molecules. In such conditions, the high positive charge of HEWL functions as a filter that allows interacting only with proteins with properly matched conformation of which interactions counterbalance strong repulsive electrostatic interactions. The presence of salts effectively destroys “the filter” and significantly alleviates interactions between pro-tein molecules and allows to form also non-fibril aggre-gates. Facilitation of interaction between protein molecules is apparently proportional to the ability of anions to bind to protein surface and thus neutralize repulsive interac-tions [33, 34, 71]. CD spectra indicate that due to effect of chaotropic and kosmotropic anions in contrast to neutral anions may “lock” the fibril in pre-mature form. The effect of monovalent anions to accelerate fibrillization and at the same time, the aggregation of HEWL clearly follows the electroselectivity series which orders the salts according to their retention times on an anion exchange column [72, 73] that for the monovalent anions correspond to inverse
Fig. 9 Schematic overview of an effect of Hofmeister anions on CD spectra and shape of lysozyme fibrils. White bars in insets, which show pic-tures obtained from AFM experiments, correspond to length of 1 μm

J Biol Inorg Chem
1 3
Hofmeister series: perchlorate > thiocyanate > nitrate > bro-mide > chloride. The observation that the efficiency of anions to accelerate fibrillization of proteins and peptides follows the electroselectivity series is supported by conclu-sion of several previous studies [37, 39, 40]. Moreover, the inverse Hofmeister series observed in fibrillization/aggrega-tion of HEWL at pH 2.7 (present work) as well as in the liq-uid–liquid phase transition of HEWL at pH 9.4 [74] suggest its general nature for inter-lysozyme interaction at relatively low concentration (<300 mM) of salts.
Although our experimental setup did not allow us to analyze an effect of salts at higher concentrations than 200 mM, we hypothesize that fibrillization/aggregation of HEWL at salts concentrations (>500 mM) at which Hofmeister effect is usually examined will be analogous to the observed effects at the salt concentration ~200 mM. This hypothesis is based on: (1) the presented findings that modulation of protein–protein interaction by anions, at studied conditions, significantly depends on the posi-tion of the anion in the Hofmeister series which determines its interaction with HEWL surface and (2) an assumption, supported by the monotone dependences of Ttrs on salt con-centration (Fig. 6), that even high salt concentrations do not significantly affect conformation of HEWL that might affect protein–protein interaction. In fact, the latter assump-tion is supported by findings of recent works [39, 40].
Conclusion
Amyloid fibrillization of HEWL due to incubation at acidic pH, pH 2.7, high temperature, 65 °C, and agitation strongly depends on the type of anions present. The fact that effi-ciency of monovalent anions to accelerate fibrillization cor-relates with electroselectivity series as well as our analysis of effect of anions on thermal stability of HEWL strongly indicates that anion effect is mediated by direct interac-tion with protein and not due to modification of HEWL hydrodynamic properties [75]. Acceleration of nucleation phase of amyloid fibrillization is, however, accompanied by increased formation of amorphous aggregates and pre-mature fibrils pointing out that amyloid fibrillization is a thermodynamically driven process resembling more crys-tallization of molecular species than aggregation, in agree-ment with the idea originally brought up by Zurdo et al. [38].
Acknowledgments This work was supported by research grants from the Slovak Grant Agency VEGA (Projects No. 1/0521/12, 2/0181/13, 2/0175/14), from Slovak Research and Development Agency (Project APVV 0526-11), ESF 26220220005, and from CELIM (316310) funded by 7FP EU (REGPOT). We also thank Ivana Petrencáková for her editorial help in preparing the manuscript.
References
1. Booth DR, Sunde M, Bellotti V, Robinson CV, Hutchinson WL, Fraser PE, Hawkins PN, Dobson CM, Radford SE, Blake CC, Pepys MB (1997) Nature 385:787–793
2. Tan SY, Pepys MP (1994) Histopathology 25:403–414 3. Pepys MB, Hawkins PN, Booth DR, Vigushin DM, Tennent GA,
Soutar AK, Totty N, Nguyen O, Blake CC, Terry CJ, Feest TG, Zalin AM, Hsuan JJ (1963) Nature 362:553–557
4. Morozova-Roche LA, Zurdo J, Spencer A, Noppe W, Rece-veur V, Archer DB, Joniau M, Dobson CM (2000) J Struct Biol 130:339–351
5. Krebs MR, Wilkins DK, Chung EW, Pitkeathly MC, Chamber-lain AK, Zurdo J, Robinson CV, Dobson CM (2000) J Mol Biol 300:541–549
6. Arnaudov LN, de Vries R (2005) Biophys J 8:515–526 7. Redfield C, Dobson CM (1988) Biochemistry 27:122–136 8. Radford SE, Dobson CM, Evans PA (1992) Nature 358:302–307 9. Itzhaki LS, Evans PA, Dobson CM, Radford SE (1994) Bio-
chemistry 33:5212–5520 10. Goda S, Takano K, Yamagata Y, Nagata R, Akutsu H, Maki S,
Namba K, Yutani K (2000) Protein Sci 9:369–375 11. Tanaka S, Oda Y, Ataka M, Onuma K, Fujiwara S, Yonezawa Y
(2001) Biopolymers 59:370–379 12. Yonezawa Y, Tanaka S, Kubota T, Wakabayashi K, Yutani K,
Fujiwara S (2002) J Mol Biol 323:237–251 13. Cao A, Hu D, Lai L (2004) Protein Sci 13:319–324 14. Blake CCF, Koenig DF, Mair GA, Sarma R (1965) Nature
206:757–761 15. Kuehner DE, Engmann J, Fergg F, Wernick M, Blanch HW,
Prausnitz JM (1999) J Phys Chem B 103:1368–1374 16. Zhang Y, Cremer PS (2009) Proc Natl Acad Sci USA
106:15249–15253 17. Privalov PL, Khechinashvili NN (1974) J Mol Biol 86:665–684 18. Bye JW, Falconer RJ (2013) Protein Sci 22:1563–1570 19. Hofmeister F (1888) Arch Exp Pathol Pharmacol 24:247–260 20. Kunz W, Henle J, Ninham BW (2004) Curr Opin Colloid Inter-
face Sci 9:19–37 21. Zhang Y, Cremer PS (2010) Annu Rev Phys Chem 61:63–83 22. Parsons DF, Boström M (2011) Lo Nostro P, Ninham BW. Phys
Chem Chem Phys 13:12352–12367 23. Lo Nostro P, Ninham BW (2012) Chem Rev 112:2286–2322 24. Collins KD (2012) Biophys Chem 167:43–59 25. Record MT Jr, Guinn E, Pegram L, Capp M (2013) Faraday Dis-
cuss 160:9–44 26. Salis A, Ninham BW (2014) Chem Soc Rev 43:7358–7377 27. Jungwirth P, Tobias DJ (2002) J Phys Chem B 106:6361–6373 28. Jungwirth P, Tobias DJ (2006) Chem Rev 109:1259–1281 29. Pegram LM, Record MT (2006) Proc Natl Acad Sci USA
103:14278–14281 30. Pegram LM, Record MT (2007) J Phys Chem B 111:5411–5417 31. Pegram LM, Record MT (2008) J Phys Chem B 112:9428–9436 32. Collins KD (2006) Biophys Chem 119:271–281 33. Flores SC, Kherb J, Cremer PS (2012) J Phys Chem C
116:11408–14413 34. Bogár F, Bartha F, Násztor Z, Fábián L, Leitgeb B, Dér A (2014)
J Phys Chem B 118:8496–8504 35. Buell AK, Hung P, Salvatella X, Welland ME, Dobson CM,
Knowles TP (2013) Biophys J 104:1116–1126 36. Jain S, Udgaonkar JB (2010) Biochemistry 49:7615–7624 37. Pedersen JS, Flink JM, Dikov D, Otzen DE (2006) Biophys J
90:4181–4194 38. Zurdo J, Guijarro JI, Jiménez JL, Saibil HR, Dobson CM (2001)
J Mol Biol 311:325–340

J Biol Inorg Chem
1 3
39. Marek PJ, Patsalo V, Green DF, Raleigh DP (2012) Biochemistry 51:8478–8490
40. Klement K, Wieligmann K, Meinhardt J, Hortschansky P, Rich-ter W, Fändrich M (2007) J Mol Biol 373:1321–1333
41. Campioni S, Mannini B, López-Alonso JP, Shalova IN, Penco A, Mulvihill E, Laurents DV, Relini A, Chiti F (2012) J Mol Biol 424:132–149
42. Ruzafa D, Conejero-Lara F, Morel B (2013) Phys Chem Chem Phys 15:15508–15517
43. Owczarz M, Arosio P (2014) Biophys J 107:197–207 44. Raman B, Chatani E, Kihara M, Ban T, Sakai M, Hasegawa K,
Naiki H, Rao CM, Goto Y (2005) Biochemistry 44:1288–1299 45. Campos LA, Sancho J (2006) Proteins 63:581–594 46. Nielsen L, Khurana R, Coats A, Frokjaer S, Brange J, Vyas S,
Uversky VN, Fink AL (2001) Biochemistry 40:6036–6046 47. Baldwin RL (1996) Biophys J 71:2056–2063 48. Sedlák E, Stagg L, Wittung-Stafshede P (2008) Arch Biochem
Biophys 479:69–73 49. LeVine H (1995) Int J Exp Clin Invest 2:1–6 50. Biancalana M, Koide S (2010) Biochim Biophys Acta
1804:1405–1412 51. Urry DW, Long MM, Ohnishi T, Jacobs M (1974) Biochem Bio-
phys Res Commun 61:1427–1433 52. Bush CA, Sarkar SK, Kopple KD (1978) Biochemistry
17:4951–4954 53. Privalov PL, Griko YV, Venyaminov SY, Kutyshenko VP (1986)
J Mol Biol 190:487–498 54. Pedersen JS, Dikov D, Flink JL, Hjuler HA, Christiansen G,
Otzen DE (2006) J Mol Biol 355:501–523 55. Omta AW, Kropman MF, Woutersen S, Bakker HJ (2003) Sci-
ence 301:347–349 56. Batchelor JD, Olteanu A, Tripathy A, Pielak GJ (2004) J Am
Chem Soc 126:1958–1961 57. Näslund LA, Edwards DC, Wernet P, Bergmann U, Ogasawara
H, Pettersson LG, Myneni S, Nilsson A (2005) J Phys Chem A 109:5995–6002
58. Chen X, Yang T, Kataoka S, Cremer PS (2007) J Am Chem Soc 129:12272–12279
59. Guest WL, Lewis WCM (1939) Proc R Soc Lond A 170:501–513 60. Aveyard R, Saleem SM (1976) J Chem Soc Farad Trans
72:1609–1617 61. Holz M, Grunder R, Sacco A, Meleleo A (1993) J Chem Soc,
Faraday Trans 89:1215–1222 62. Sacco A, De Cillis FM, Holz M (1998) J Chem Soc Faraday
Trans 94:2089–2092 63. Chang TM, Dang LX (2006) Chem Rev 106:1305–1322 64. Funahashi J, Takano K, Ogasahara K, Yamagata Y, Yutani K
(1996) J Biochem 120:1216–1223 65. Canet D, Last AM, Tito P, Sunde M, Spencer A, Archer DB,
Redfield C, Robinson CV, Dobson CM (2002) Nat Struct Biol 9:308–315
66. Buell AK, Dhulesia A, Mossuto MF, Cremades N, Kumita JR, Dumoulin M, Welland ME, Knowles TP, Salvatella X, Dobson CM (2011) J Am Chem Soc 133:7737–7743
67. Frare E, De Laureto PP, Zurdo J, Dobson CM, Fontana A (2004) J Mol Biol 340:1153–1165
68. Uversky VN, Fink AL (2004) Biochim Biophys Acta 1698:131–153
69. Gokarn YR, Fesinmeyer RM, Saluja A, Razinkov V, Chase SF, Laue TM, Brems DN (2011) Protein Sci 20:580–587
70. Möller J, Grobelny S, Schulze J, Steffen A, Bieder S, Paulus M, Tolan M, Winter R (2014) Phys Chem Chem Phys 16:7423–7429
71. Goto Y, Calciano LJ, Fink AL (1990) Proc Natl Acad Sci USA 87:573–577
72. Gjerde DT, Schmuckler G, Fritz JS (1980) J Chromatogr 187:35–45
73. Gregor HP, Belle J, Marcus RA (1955) J Am Chem Soc 77:2713–2719
74. Boström M, Parsons DF, Salis A, Ninham BW, Monduzzi M (2011) Langmuir 27:9504–9511
75. Parmar AS, Muschol M (2009) Biophys J 97:590–598 76. Marcus Y (1997) Ion Properties. Marcel Dekker, New York
Recommended

















