Embed Size (px)
Citation preview

Journal of Plankton Research Vol.19 no.3 pp.303-318, 1997
The abundance cycle and influence factors on red tide phenomenaotNoctiluca scintillans (Dinophyceae) in Dapeng Bay, the SouthChina Sea
C.Huang and Y.QiInstitute of Hydrobiology, Jinan University, Guangzhou 510632, China
Abstract. The seasonal occurrence of Noctiluca scintillans and the causative mechanisms of red tidephenomena were investigated in the northwestern part of Dapeng Bay, the South China Sea, fromMarch 1990 to June 1992. It occurred in the plankton usually from January through May/June withpeak abundance from March to early May. At least eight red tides of this organism, when the surfacewater was covered with the pink patches and slicks, were observed during the investigation period, andall of them took place between March and early May. Noctiluca occurred at temperatures from 15.8to 28.6°C, and the population density was the highest at temperatures between 19 and 25°C. It disap-peared from the plankton in summer and fall when the temperature wanned to lethal level of -30°Cand then the population probably was transported to outside of the bay. During the peak period ofabundance between March and May, the log transformed average density of Noctiluca at each stationshowed a significantly positive relationship with average water temperature, indicating that within itsoptimum temperature a higher temperature promotes the growth of Noctiluca population. Sharpdeclines of the population were associated with abrupt decreases in salinity due to heavy rain. Hence,rich food supply is basically necessary for Noctiluca to reproduce massively, but the suitable tempera-ture, stable muggy weather without heavy rain are considered to be the important factors for the Noc-tiluca blooms. The surface convergence, due to the geographical and tidal features, may also constitutea direct driving force leading to its frequent occurrences in the study area. The Noctiluca densityshowed a negative relationship with chlorophyll a concentration, indicating that this species gives apredation pressure on the phytoplankton.
Introduction
In recent years, there has been an increase in the frequency, magnitude and geo-graphic extent of red tides along the Chinese coast (Qi et ai, 1992,1995). In theYangtze River estuary, for example, Hong and Huang (1989) and Qi et al (1993)reported that a Noctiluca scintillans red tide occurred within a region of only10 km2 in August 1982, but the scale was enlarged to 6100 km2 in August 1988.Although Noctiluca is not as harmful as other toxic/noxious flagellates, the out-breaks of its bloom in China have been so severe that they have resulted inmassive fish and shrimp mortality, especially in cases of cage fish-culture. Since thedensity of viable cells, when red tides of Noctiluca occur, is always so high that thesurface water is covered with pink patches and slicks of Noctiluca, and stinks offish, the major consequence of red tides in Hong Kong and the South China Seais considered to be dissolved oxygen depletion (Ho and Hodgliss, 1992; Xie et al,1993).
In Dapeng Bay of the South China Sea, red tides caused by Noctiluca are fre-quently observed in spring and early summer, and their magnitude has beenexpanding (Qi et al, 1993). To make clear the causative mechanisms of the redtides, both field and laboratory investigations were performed from March 1990to June 1992 under the sponsorship of the National Natural Sciences Foundationof China. Some study results on the biology and ecology of Noctiluca in DapengBay have been published in various journals (Chen and Qi, 1991; Huang et al,
C Oxford University Press 303
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

C.Huang and Y.Qi
1992,1996; Huang and Qi, 1993; Qi and Li, 1994). For example, Chen and Qi (1991)found that the food of Noctiluca was mainly composed of phytoplankton withlimited debris, micro-zooplankton and their eggs. The mean digestion time ofvarious food particles is 4.5 ± 2.4 h, and the feeding showed remarkably diurnalrhythms with two peaks at night. Vegetative reproduction also showed two peaksa day, and occurred at night. Qi and Li (1994) reported that under experimentalconditions some mother cells of Noctiluca divided into two unequal daughter cells,and suggested that asexual reproduction may be a rapid way to increase individualnumbers for the population. Based on sampling data from 30 March to 10 June1990, Huang and Qi (1993) analysed the relationship between the density of Noc-tiluca and eight environmental factors, and stated that temperature, dissolvedinorganic nitrogen (DIN), dissolved oxygen (DO), chlorophyll a and salinity werethe major factors that determined the growth of Noctiluca.
In this paper, we describe the seasonal and geographical variations in occur-rence of Noctiluca in the inner part of Dapeng Bay, and discuss the effects ofenvironmental factors on the formation of its blooms.
Method
Study area
We established four stations in the northwestern part of Dapeng Bay, a semien-closed embayment east of Kowloon Peninsula, Hong Kong (Figure l).The averagedepth of the bay is 14.7 m, and bay mouth is -10 km wide. The depth of our studyarea was shallower than 12 m. Since there is no big river inflow into this bay, thewater movement is mainly generated by tide. Zhan (1989) demonstrated thatstrong tidal currents occur only in the southern part of the bay and no significantresidual current eddy is observed in the northern part. Because of this tidal currentfeature and the prevailing southeast wind, especially in spring and summer, thebuoyant materials tend to be accumulated along the northern coast (Zhan, 1990).
Materials and methods
Samplings were undertaken from a boat at three stations (S3/S0, SI, S2) fromMarch 1990 to June 1992 at 3 day intervals during March and June and ~2 weekintervals for the rest of the year (Figure 1). At each station, water samples weretaken at the surface and near the sea bed (only at the surface at SO due to shallowdepth) using a 5 1 water bottle. One litre of water sample from each depth wastaken into a sampling bottle and preserved with formalin at final concentration of4%, and brought back to the land laboratory. Later, it was concentrated to 20 mlby settling and vegetative cells of Noctiluca were counted. Simultaneously, a widerange of environmental parameters, i.e. weather, physical (temperature, salinityand pH), chemical (DIN, dissolved inorganic phosphate. Si, DO, chlorophyll a)and biological (phyto- and zooplankton) ones were determined, but only relatedparameters are presented in this paper.
Since densities of Noctiluca were remarkably low at S3 (see the Results fordetails), the investigation at S3 was stopped and started at SO from January 1991.
304
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

The population dynamics and red tide phenomena of Nocttluca
15' 16' 19' El 14'
N
Fig. L Map of the study area in Dapeng Bay of the South China Sea with locations of samplingstations: S0/S3, SI and S2.
Results
Seasonal variation in environmental parameters
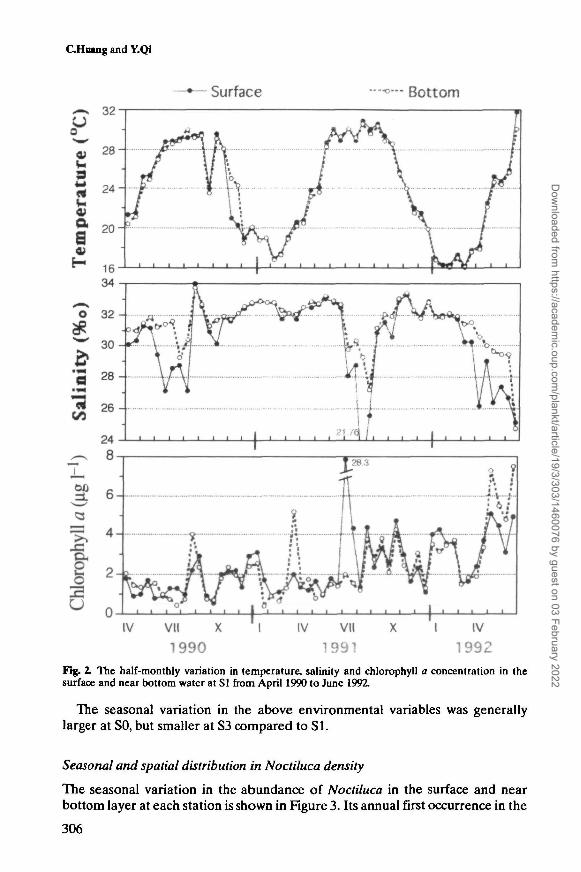
Figure 2 shows the half-monthly variations in temperature, salinity and chloro-phyll a concentration in the surface water at station SI from April 1990 to June1992. Throughout the investigation, the temperature varied between 16.1 and31.7°C. The lowest temperature was generally found in early February, and thenthe temperature tended to rise. In 1992, however, the lowest temperature occurredin early March. The temperature reached the annual maximum in June or July, andstayed warm until September or October. Usually, the temperature was almost thesame for the surface and near bottom layer (Figure 2).
The salinity fluctuated violently between 15.6 and 33.7%o during the presentstudy. Salinity was generally high in the fall and winter (October-February) andlow in the early spring and summer due to the rainy season and typhoon (Figure2). However, there was a great yearly difference in the variation depending on theweather of the year. Obviously, the salinity in the early half of 1992 was much lowerthan that of 1991. In the high salinity seasons, there was no large difference in salin-ity between the surface and near bottom layer, but in the low salinity seasons thedifference was larger. The salinity and temperature increased simultaneously inAugust of 1990, indicating that a warm offshore water flowed into the study area.
Chlorophyll a concentration also varied markedly between 0.39 and 28.3 y.g Mduring the investigation. In general, it was lower in spring and summer than in falland winter, but there was a remarkable yearly difference (Figure 2). The surfaceconcentrations were not always higher than those in the near bottom layer.
305
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
CHtang and Y.Qi
Surface —-O— Bot tom
y\f
Fig. 2. The half-monthly variation in temperature, salinity and chlorophyll a concentration in thesurface and near bottom water at SI from April 1990 to June 1992.
The seasonal variation in the above environmental variables was generallylarger at SO, but smaller at S3 compared to SI.
Seasonal and spatial distribution in Noctiluca density
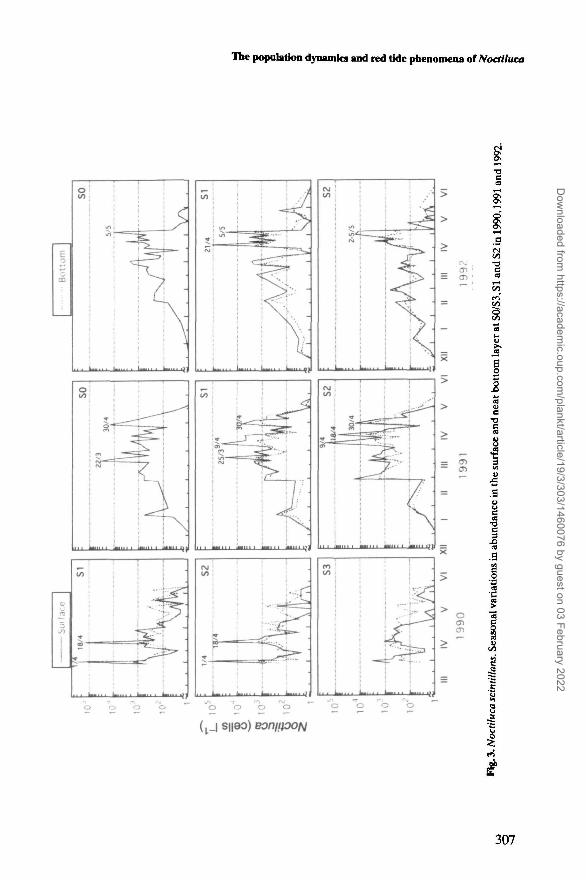
The seasonal variation in the abundance of Noctiluca in the surface and nearbottom layer at each station is shown in Figure 3. Its annual first occurrence in the
306
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
IV
V
VI
XII
I
|| II
I IV
V
V
I X
II
IIV
V
V
I
19
9?
t I n
F|g
,3. i
feer
i/uca
sci
ntil
lans
. Sea
sona
l var
iatio
ns in
abu
ndan
ce in
the
surf
ace
and
near
bot
tom
laye
r at
S0/
S3, S
I an
d S2
in 1
990,
1991
and
199
2.I
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

C.Huang and Y.Qi
plankton was observed in January for 1991 and December for 1992. The densityfluctuated remarkably, but increased to a peak between late March and earlyMay. The population density was generally >200 cells I"1 in March and April.Afterwards, the population declined rapidly to disappear completely from theplankton. The time of disappearance differed slightly depending on the year; it dis-appeared by 10 June in 1990,28 May in 1991 and 22 June in 1992.
The density was significantly lower in 1992 than in the other years, althoughthere were some differences among the stations (Figure 3). The mean densities ofNoctiluca at both SI and S2 during the peak period (March-May, but April andMay in 1990) were 8890,4600 and 2160 cells I"1 in 1990,1991, and 1992, respectively.
Before early May, the cell densities were usually higher in the surface than nearthe bottom (Figure 3). The greatest difference of 185 times was observed duringthe bloom occurring at SI on 1 April 1990. As the population declined, thedensities in the surface became less than the bottom. The population disappearedearlier in the surface than the bottom in 1991 and 1992 at S2, where the tempera-ture stratification was remarkable.
The patterns of the seasonal occurrence of Noctiluca were more or less similaramong stations, but the density was remarkably lower at S3 than the other stations(Figure 3). The two blooms were observed at SI and S2 in April 1990, whereas nobloom was found at S3 (Figure 3), where the mixing of the water was most intenseby a current from the Qinzhou Channel (Figure 1).
Noctiluca red tides
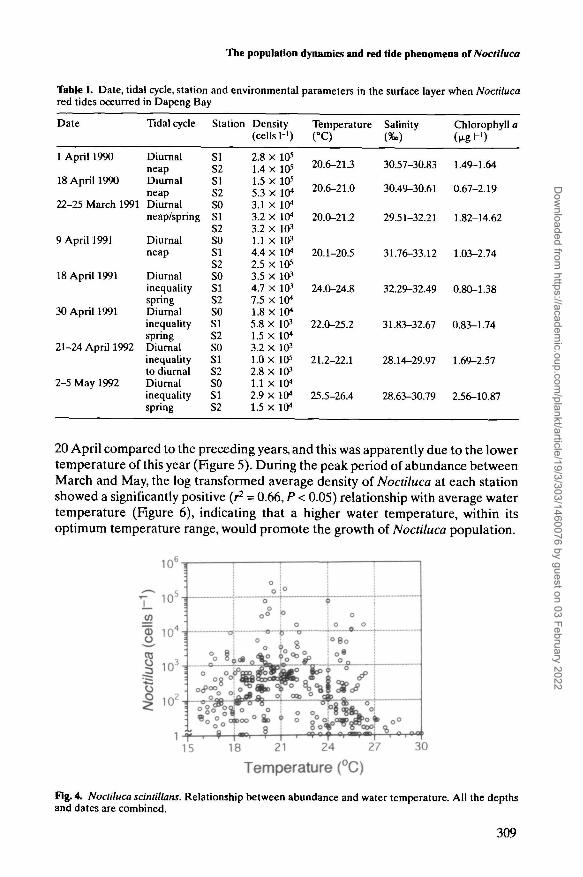
There were eight Noctiluca red tides, which represented the condition that thesurface water was covered with pink patches and slicks of Noctiluca and stank offish, in the study area throughout the investigation (Figure 3). When these redtides occurred, the cell densities in the surface were generally higher than 104 cellsI-1 (Table I). The highest density of 2.8 X 105 cells I"1 was observed at SI on 1 April1990. The red tides took place suddenly. For example, at S2 in 1991 the averagedensity in the water column increased from 25 cells I"1 on 28 February to 8900 cellsI"1 on 1 March, and at SI in 1990 it increased from 480 cells H on 30 March to1.4 X 105 cells H on 1 April. The duration of the blooms was short; in most casesthey did not last longer than 3 days.
The population density in relation to temperature
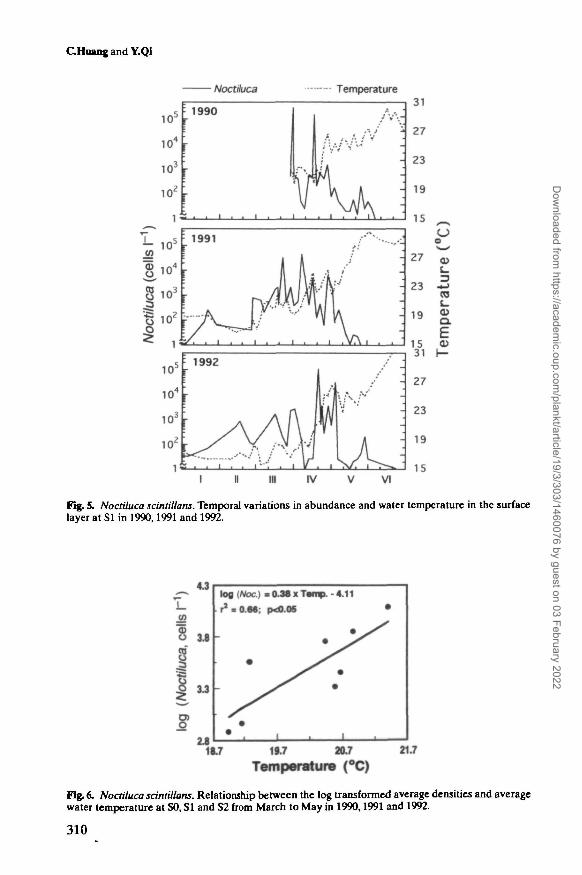
When Noctiluca occurred in the study area, the water temperature ranged from 15.8to 28.6°C (Figure 4), and most blooms occurred within a temperature range between19.8 and 22°C (Figure 4,Table I). The densities of Noctiluca increased with temper-ature up to ~20°C, and decreased rapidly when temperature exceeded ~25°C.
Figure 5 shows the seasonal variations of both Noctiluca abundance and tem-perature in the surface water at SI between January and June in each year. Thewater temperature always showed a rapid increase of ~4-7°C (from <20 to >25°C)during April due to consecutive sunny days and inflow of warm offshore water. Thepeaks of the population density were often associated with the rapid increase ofthe water temperature. In 1992, the densities of Noctiluca were much lower before
308
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
The population dynamics and red tide phenomena of Noctiluca
Table L Date, tidal cycle, station and environmental parameters in the surface layer when Noctilucared tides occurred in Dapeng Bay
Date Tidal cycle Station Density Temperature Salinity Chlorophyll a(cells!-') CC) («.) (u-gH)
20.6-213 30.57-30.83 1.49-1.64
20.6-21.0 30.49-30.61 0.67-2.19
20.0-212 29.51-32.21 1.82-14.62
20.1-20.5 31.76-33.12 1.03-2.74
24.0-24.8 32.29-32.49 0.80-1.38
22.0-25.2 31.83-32.67 0.83-1.74
21.2-22.1 28.14-29.97 1.69-2.57
25.5-26.4 28.63-30.79 2.56-10.87
1 April 1990
18 April 1990
22-25 March 1991
9 April 1991
18 April 1991
30 April 1991
21-24 April 1992
2-5 May 1992
DiurnalneapDiurnalneapDiumalneap/spring
Diurnalneap
DiurnalinequalityspringDiurnalinequalityspringDiurnalinequalityto diurnalDiurnalinequalityspring
SIS2SIS2SOSIS2SOSIS2SOSIS2SOSIS2SOSIS2SOSIS2
2.81.41.55.33.13.23.21.14.4153.54.77.51.85.81.53.21.02.81.12.91.5
XXXXXXXXXXXXXXXXXXXXXX
10s
10s
105
104
104
104
10-1
10-1104
\®1O1
lfr1
104
104
10-1
104
103
105
10-1
104
104
104
20 April compared to the preceding years, and this was apparently due to the lowertemperature of this year (Figure 5). During the peak period of abundance betweenMarch and May, the log transformed average density of Noctiluca at each stationshowed a significantly positive (r2 = 0.66, P < 0.05) relationship with average watertemperature (Figure 6), indicating that a higher water temperature, within itsoptimum temperature range, would promote the growth of Noctiluca population.
15
Temperature (°C)
Fig. 4. Noctiluca scintillans. Relationship between abundance and water temperature. All the depthsand dates are combined.
309
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
CHoang and Y.Qi
- NoctUuca Temperature
10
10
VI
Fig. 5. NoctUuca scintUlans. Temporal variations in abundance and water temperature in the surfacelayer at SI in 1990,1991 and 1992.
4.3
8 3,
2J
log (Woe.)
r* • 0.M;
• 0.38 x Tamp.
p<0.06
-4.11
18.7 19.7 20.7 21.7
Temperature (°C)
Fig. 6. NoctUuca sdntiUans. Relationship between the log transformed average densities and averagewater temperature at SO, SI and S2 from March to May in 1990,1991 and 1992.
310
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
The population dynamics and red tide phenomena of Noctiluca
19 23 27 31
Salinity (%o)
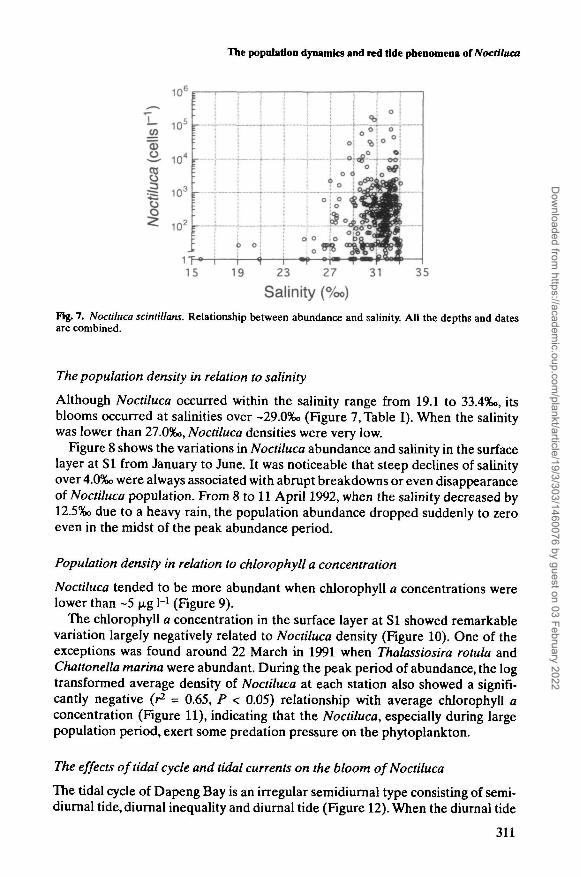
Fig. 7. Noctiluca scintillans. Relationship between abundance and salinity. All the depths and datesare combined.
The population density in relation to salinity
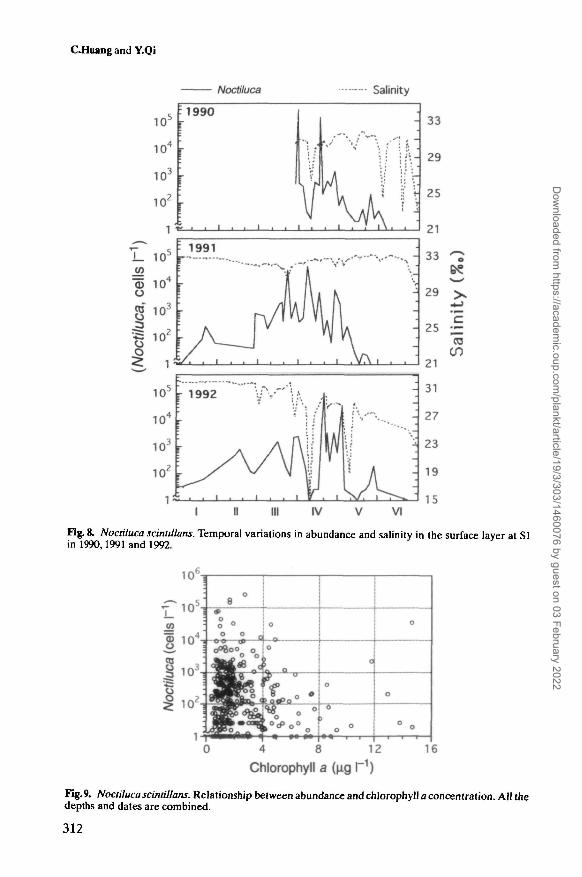
Although Noctiluca occurred within the salinity range from 19.1 to 33.4%o, itsblooms occurred at salinities over ~29.0%o (Figure 7, Table I). When the salinitywas lower than 27.0%o, Noctiluca densities were very low.
Figure 8 shows the variations in Noctiluca abundance and salinity in the surfacelayer at SI from January to June. It was noticeable that steep declines of salinityover 4.0%o were always associated with abrupt breakdowns or even disappearanceof Noctiluca population. From 8 to 11 April 1992, when the salinity decreased by12.5%o due to a heavy rain, the population abundance dropped suddenly to zeroeven in the midst of the peak abundance period.
Population density in relation to chlorophyll a concentration
Noctiluca tended to be more abundant when chlorophyll a concentrations werelower than ~5 u.g I"1 (Figure 9).
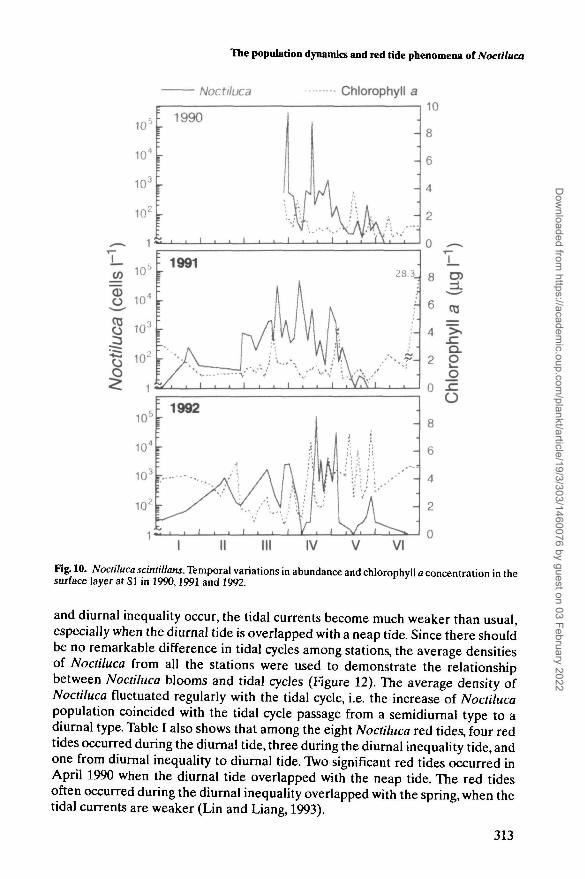
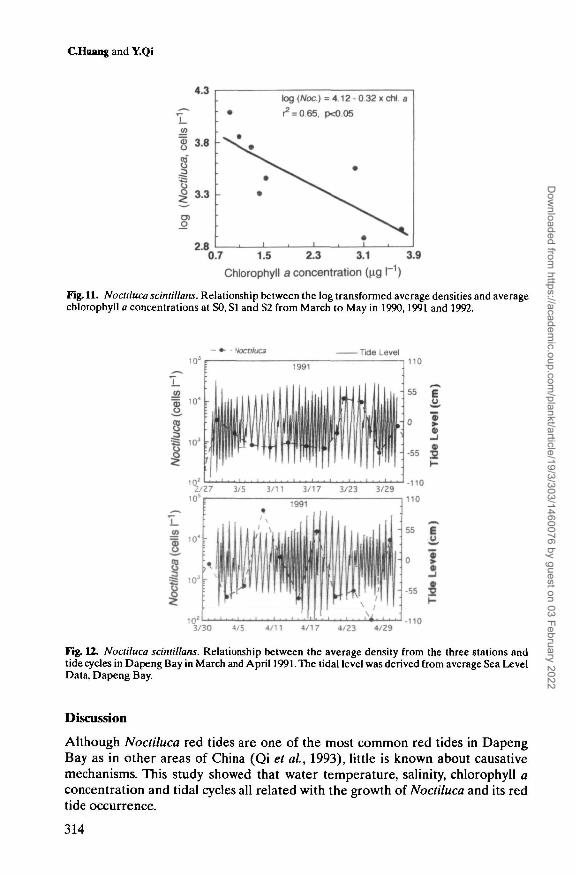
The chlorophyll a concentration in the surface layer at SI showed remarkablevariation largely negatively related to Noctiluca density (Figure 10). One of theexceptions was found around 22 March in 1991 when Thalassiosira rotula andChattonella marina were abundant. During the peak period of abundance, the logtransformed average density of Noctiluca at each station also showed a signifi-cantly negative (r2 = 0.65, P < 0.05) relationship with average chlorophyll aconcentration (Figure 11), indicating that the Noctiluca, especially during largepopulation period, exert some predation pressure on the phytoplankton.
The effects of tidal cycle and tidal currents on the bloom of Noctiluca
The tidal cycle of Dapeng Bay is an irregular semidiurnal type consisting of semi-diurnal tide, diurnal inequality and diurnal tide (Figure 12). When the diurnal tide
311
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
CJiuang and Y.Qi
10
10
10
10
J _ 1Ob
_co"o5 1 0 4
o
g 103
"•5 10
8
103 r
10' r
103
5
4
3
2
1 i
Noctiluca
1990
-
" • ; . -
Vi , . i . . i . . i .
Salinity
-
A if iiVI A ! \\iV? A. . •
33
29
25
1991
J \
r 1992
r
V
A /
1
i .V\. . .
-
A / I : '••
e
- 29
III IV
25 aCO
CO21
31
27
23
19
15VI
Fig. 8. Noctiluca scinldlans. Temporal variations in abundance and salinity in the surface layer at SIin 1990,1991 and 1992.
4 8 12
Chlorophyll a (ng I"1)16
Fig. 9. Noctiluca scintillans. Relationship between abundance and chlorophyll a concentration. All thedepths and dates are combined.
312
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
The population dynamics and red tide phenomena of Noctiluca
Noctiluca Chlorophyll a
III IV V VI
Fig. 10. Noctiluca scintillans. Temporal variations in abundance and chlorophyll a concentration in thesurface layer at SI in 1990,1991 and 1992.
and diurnal inequality occur, the tidal currents become much weaker than usual,especially when the diurnal tide is overlapped with a neap tide. Since there shouldbe no remarkable difference in tidal cycles among stations, the average densitiesof Noctiluca from all the stations were used to demonstrate the relationshipbetween Noctiluca blooms and tidal cycles (Figure 12). The average density ofNoctiluca fluctuated regularly with the tidal cycle, i.e. the increase of Noctilucapopulation coincided with the tidal cycle passage from a semidiurnal type to adiurnal type. Table I also shows that among the eight Noctiluca red tides, four redtides occurred during the diurnal tide, three during the diurnal inequality tide, andone from diurnal inequality to diurnal tide. Two significant red tides occurred inApril 1990 when the diurnal tide overlapped with the neap tide. The red tidesoften occurred during the diurnal inequality overlapped with the spring, when thetidal currents are weaker (Lin and Liang, 1993).
313
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022
CHuang and Y.Qi
4.3log (Noc.) = 4 1 2 - 0 32 x cfH a
1^ = 0 65, [X0.05
0.7 1.5 2.3 3.1Chlorophyll a concentration (ng l~1)
Fig. 11. Noctiluca scintillans. Relationship between the log transformed average densities and averagechlorophyll a concentrations at SO, SI and S2 from March to May in 1990,1991 and 1992.
- Tide Level110
2/27 3/5 3/11 3/17 3/23 3/29
4/5 4/17 4/23 4/29-110
Fig. 12. Noctiluca scintillans. Relationship between the average density from the three stations andtide cycles in Dapeng Bay in March and April 1991. The tidal level was derived from average Sea LevelData, Dapeng Bay.
Discussion
Although Noctiluca red tides are one of the most common red tides in DapengBay as in other areas of China (Qi et al, 1993), little is known about causativemechanisms. This study showed that water temperature, salinity, chlorophyll aconcentration and tidal cycles all related with the growth of Noctiluca and its redtide occurrence.
314
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

The population dynamics and red tide phenomena of Noctiluca
The upper limit of water temperature for Noctiluca in the study area was 28.6°C.However, our culture experiments demonstrated that Noctiluca obtained fromthe study area could not survive at temperature over 26°C in the laboratory. Sinceno dormant stage of Noctiluca has been found, Noctiluca in Dapeng Bay may dieout by summer because of high temperature (~30°C). Ho (1993) suggested thatNoctiluca is generally absent from the waters around Hong Kong and in the PearlRiver Estuary from August to October because of high temperature, and theKuroshio Current is responsible for transporting the vegetative cells from otherareas to the South China Coast again. Since the Noctiluca blooms first appear inthe eastern waters of Hong Kong in late December every year (Ho, 1993), thevegetative cells are transported by wind or tidal forces to Dapeng Bay around theend of the year.
In the study area, Noctiluca was generally most abundant from early March tolate April or early May, when the water temperature ranged between 17.4 and24.5°C (Figure 2). The lowest temperature at which the Noctiluca red tide tookplace in the study area was 18.6°C. The Noctiluca population declined when thetemperature increased up to 25°C. Hence, the optimum temperatures for Noc-tiluca in Dapeng Bay are similar to those for Noctiluca around Hong Kong waters(20-24°C; Ho, 1993), and the culture experiments with cells from Helgolandwaters, Germany (22-24°C; Uhlig and Sahling, 1985).
At least eight Noctiluca red tides were detected during the present investi-gation. The developments of these red tides were very quick; 10- to 100-foldincreases of the population density were observed during an inter-samplingperiod (i.e. 3 days). However, such increases seemed to be impossible by thegrowth of the local population alone, since the cell division was reported as onlyonce per 12-24 h (Pratje, 1925) or every 3 days (Gross, 1934). Qi and Li (1994)reported an unequal division during the asexual reproductive process of Noctilucaunder an experimental condition (18-22°C; ~30%o), and suggested that this modeof division may lead to a rapid increase of the individual numbers since the smallcells may need fewer life materials. However, this hypothesis is not applicable tothe present results since the small cells occupied only a few percent of the totalpopulation. Moreover, during a red tide on 1 April 1990 (average cell density:1.4 X 105 cells I"1) almost the entire population was composed of normal veget-ative cells.
The passive accumulation of Noctiluca due to winds and/or tidal currents seemsto be one of the important mechanisms for the occurrence of the bloom (Chew,1955; Fevre and Grail, 1970). Wang et al. (1994) and Zhan (1989, 1990) havedemonstrated that the geographical and tidal features of Dapeng Bay makebuoyant matter accumulate in the study area, and especially during the periods ofdiurnal tide and remarkable diurnal inequality tide, buoyant materials tend to beconcentrated at the sites of convergence in the northwestern part of the bay. Thesephysical factors may explain the formation of slicks and patches of Noctiluca inthe present study (Liang and Qian, 1989; Xie et al, 1993).
The abrupt and big falls in salinity due to heavy rain caused sharp decline orbreakdown of the population in the surface layer. The culture experiments (S.Qi,personal communication) also demonstrated that an abrupt fall of salinity over
315
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

CHuang and Y.QI
4%o remarkably disturbed the growth of Noctiluca and occasionally resulted in celldeath. Although previous studies (Kuroda and Saga, 1978; Zhou et al., 1983; Uhligand Sahling, 1985; Ho, 1993) reported that Noctiluca can survive within a widerange of salinity between 14 and 34%o, stable weather condition without heavy rainis important for the formation and maintenance of the Noctiluca red tides.
According to our weather records, most Noctiluca red tides occurred in or justafter stable muggy weather without strong wind and waves. These weather con-ditions may not only promote the cell division of Noctiluca due to relatively hightemperature (Yentsch and Mague, 1980; Chisholm, 1981; Sweeney, 1987; Uhlig andSahling, 1990), but also make Noctiluca cells accumulate in the convergence zone.Based on their long-term study, Uhlig and Sahling (1990) found that there areannual oscillations of Noctiluca population developments in waters around Hel-goland at ~3-year intervals, i.e. a year with relatively high abundance is followedby 2 years of relative low abundance. In the present study, the population abun-dance of Noctiluca also exhibited yearly oscillation (high in 1990 and 1991, low in1992). Our weather records showed that it was remarkably rainier and colderduring the spring in 1992, and the log transformed density of Noctiluca alsoshowed a significantly positive relationship with water temperature (Figure 6).Although Uhlig and Sahling (1990) did not offer any interpretation for thephenomenon, the yearly difference in weather conditions should play an impor-tant role in the yearly oscillations of Noctiluca population.
Hada (1976), and Kuroda and Saga (1978) reported that the frequent occur-rences of Noctiluca red tides in the Inland Sea of Japan and the coastal waters ofJapan are mainly caused by the increase of phytoplankton due to eutrophication.In Dapeng Bay, the eutrophication may also be the main reason why Noctiluca redtides become more frequent and over wider areas (Qi et al., 1993). In addition,because of its extreme polyphagous heterotrophy (Enomoto, 1956; Sekiguchi,1977; Kimor 1979; Daan, 1987; Kirchner et al., 1996), food supply is considered tobe rich for Noctiluca to reproduce massively in Dapeng Bay. Lu and Qi (1995)reported that a spring peak of phytoplankton is observed in the study area everyyear, and the phytoplankton community is composed of diatoms and flagellates.As well as a Crnarina red tide on 22 March in 1991, a T.rotula bloom coincidedwith Noctiluca red tide between 22 and 25 March 1991. However, 12 other phyto-plankton red tides (one consisted of flagellates and the others of diatoms)observed during the present investigation did not coincide with the Noctiluca redtides (C.Huang, unpublished data). The log transformed density of Noctilucashowed a significantly negative relationship with chlorophyll a concentration(Figure 11). These findings suggested that Noctiluca, especially during a largepopulation period, may exert a predation pressure on the phytoplankton. Huanget al. (1997) also demonstrated that the large Noctiluca population exerts a greatpredation pressure on copepods, crustacean larvae, and fish eggs and larvae.
In conclusion, Noctiluca red tides occurred frequently in Dapeng Bay betweenMarch and early May. Stable and muggy weather conditions stimulate the growthof Noctiluca population as well cause accumulation in convergence regions in thenorthwestern part of the bay to form a significant red tide. Extremely weak tidalcurrents during the periods of diurnal tide and from semidiurnal tide to diurnal
316
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

The population dynamics and red tide phenomena of Noctiluca
inequality tide were associated with the formation of the blooms. Such physicalconvergence caused by weather and tidal cycle coupled with eutrophication of thewater and euryphagous dietary requirement of Noctiluca may be the reason whythe Noctiluca red tides constitute over 50% of the total red tides that occurred inDapeng Bay in recent years (C.Huang, unpublished data).
Acknowledgements
We sincerely thank Drs T.Onbe and S.Uye for valuable comments during prep-aration of the manuscript. Also, we are much grateful to Dr G.Uhlig for a carefulreview of manuscript and useful reprints on Noctiluca studies. This study was sup-ported by a grant from the National Natural Sciences Foundation of China.
ReferencesChen.H. and Qi,S. (1991) The feeding and vegetative reproduction diurnal rhythms of Noctiluca scin-
lillans. J. Jinan Univ. China, 12,104-107.Chew,F. (1955) On the offshore circulation and a convergence mechanism in the red tide region off
the west coast of Florida. Trans Am, Geophys. Un., 36,963—971.Chisholm,W.S. (1981) Temporal patterns of cell division in unicellular algae. Can. Bull Fish, Aquat.
ScL,210,150-181.Daan.R. (1987) Impact of eggs predation by Noctiluca miliaris on the summer development of
copepod population in the southern North Sea. Mar. Ecol. Prog. Ser.,37,9-17.Emomoto,Y. (1956) On the occurrence and the food of Noctiluca scmtillans (Macartney) in the waters
adjacent to the west coast of Kyushu, with special reference to the possibility of the damage causedto the fish eggs by that plankton. Bull Jpn Soc Fish., 22,82-88.
Gross F. (1934) Zur Biologie und Entwichlungsgeschichte von Noctiluca miliaris. Arch. Protistenkd.,83,178-196.
Hada.Y. (1976) The plankton of red tide. / Inst. Commercial Econ, Res.,Syudo Univ., Hiroshima, 13,23-48 (in Japanese).
HoJC.C. (1993) The dynamics of Noctiluca scintillans red tides in Hong Kong and the South China Sea:An overview, (a speech in the 6th International Conference on Toxic Phytoplankton, 1993, Nantes,France) in press.
Ho,K.C. and HodgkissJJ. (1992) Severe fishkill in Hong Kong caused by Noctiluca scintillans. Redride Newslett., 5,1-2.
HongJ. and Huang^X. (1989) Analysis and preliminary research of red tide frequent area in Changjiangestuary and adjacent sea area. J. Jinan Univ., Special Issue on Red Tides, 40-51 (in Chinese).
Huang,C, Qi,S., Qi,Y. and Ling^X. (1997) The position and function of Noctiluca marina in its eco-logical community in Dapeng Bay, the South China Sea. Oceanogr. LimnoL Sin, 28, in press.
Huang,W. and Qi,Y. (1993) Environmental influence on growth of Noctiluca scintillans in Dapeng Bay,north of South China Sea. Mar. ScL Bull.,12,17-21 (in Chinese with English abstract).
Huang,W., Qi,Y. and Qiu,X. (1992) The identification of a mechanism model on the population dy-namics of Noctiluca scintillans in the Dapeng Bay, South China Sea. Ada Ecol Sin., 12,202-212 (inChinese with English abstract).
Huang,W., Qi,Y, Zhu.C. and Luo.Y. (1996) Model on change ratio of the population densities dynam-ics of Noctiluca scintillans (Ehr.) Macartney in Dapeng Bay, South China Sea. Oceanogr. LimnoLSin., 27,29-34 (in Chinese with English abstract).
Kimor,R (1979) Predation by Noctiluca miliaris Suriray on Acartia tonsa (Dana) eggs in the inshorewaters of southern California. LimnoL Oceanogr., 24,568-572.
Kirchner,M., Sahling,G., Uhlig,G, Gunkel.W. and Klings,K.W. (1996) Does the red tide-forming dino-flagellatc Noctiluca scintillans feed on bacteria? Sarsia, 81,45-55.
KurodaJ. and Saga,S. (1978) The distribution and ecology of Noctiluca scintillans in Ohsaka Bay. BullJpn Soc Fish. Oceanogr, 32,56-67 (in Japanese).
Le FevreJ. and GrallJ.R. (1970) On the relationship of Noctiluca swarming off the western coast ofBrittany with hydrological features and plankton characteristics of the environment. / Exp. Mar.BioLEcol, 4,287-306.
317
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022

dialing and Y.Qi
Liang,S. and Qian.H. (1989) The survey and discussion on the red tide occurrence from Yantian ofDapeng Bay to Coast Nanao. J. Jinan Univ., Special Issue on Red Tides, 90-92 (in Chinese).
Lin,Z. and Liang,S. (1993) Relationship between tide and Noctiluca scintillans bloom off YantianCoast, Dapeng Bay. Mar. Sci. Bull.,12,35-38 (in Chinese with English abstract).
Lu,S and Qi,Y. (1995) Seasonal succession of phytoplankton in Dapeng Bay, South China Sea. J.JinanUniv., 16,118-120 (in Chinese with English abstract).
Pratje,A. (1925) Noctiluca. In Die Tienvelt der Nord- und Ostsee. Grimpe and Wagler, Leipzig, Bd. 2dl,S.,pp. 1-12.
Qi,S and Li,D. (1994) Unequal cell division of Noctiluca scinlillans. Oceanogr. Limnol. Sin., 25,158-161 (in Chinese with English abstract).
Qi,Y, Hong,Y, Qian.H. and Lu,S. (1992) Problems caused by harmful algal blooms in China. BMTC-POLAMAR, 94,22-24.
Qi,Y, Zhang,Z. and Hong,Y. (1993) Occurrence of red tides on the coasts of China. In Smayda.TJ.and Shimizu,Y. (eds), Toxic Phytoplankton Bloom in the Sea. Elsevier Science Publishers B.V.,pp. 43-46.
Qi, Y, Lu,S. and Zhen,L. (1995) An overview of harmful algal blooms (red tides) occurrence on coastalChina and its research program in China. Proceedings, International Conference on the MarineBiology of the South China Sea, Guangzhou, China, 1993, pp. 17-21.
Sekiguchi.H. (1977) The ecology of Noctiluca, the luminous flagellate. Mar. So., 9,52-55 (in Japanesewith English abstract).
Sweeney.B.M. (1987) Bioluminescence and circadian rhythms. In Taylor.FJ.R. (ed.), The Biology ofDinoflagellates. Blackwell Scientific Publications, Oxford, pp. 269-281.
Uhlig,G. and Sahling,G. (1985) Blooming and red tide phenomenon in Noctiluca milians. Bull Mar.Sci., 37,780.
Uhlig,G. and Sahling,G. (1990) Long-term studies on Noctiluca scintillans in the German Bight: popu-lation dynamics and red tide phenomena 1968-1988. Neth. J. Sea Res, 25,101-112.
Wang,S., Feng,G., Xia,Z., ShiJ., Qi,Y. and Lu,S. (1994) Structural analysis of the essential factorsassociated with the Noctiluca scintillans red tide in Dapeng Bay. Oceanogr. Limnol. Sin., 25,146-151(in Chinese with English abstract).
XieJ., Li J., Lu,S., Cheng.Q. and Yang,L. (1993) Features of red tide caused by Noctiluca scintillans offYantian coast within Dapeng Bay. Mar. Sci. Bull., 12,1-6 (in Chinese with English abstract).
Yentsch.C.M. and Mague,F.C. (1980) Evidence of an apparent annual rhythm in the toxic red tidedinoflagellates Conyaulax excavata. Int. J. Chronobiol. ,1,77-84.
Zhan,X. (1989) Tidal modeling of Dapeng Bay, China. Oceanogr. Limnol Sin., 20,149-155 (in Chinesewith English abstract).
Zhan^X. (1990) Pollutant dispersion modeling of Dapeng Bay, China. Trop. Oceanol, 9, 82-86 (inChinese with English abstract).
ZhouJ., Dong,L. and Qin.B. (1983) Preliminary studies on the problem of eutrophication and red tidesin Bohai Bay. Mar. Environ. Sci., 2,41-54 (in Chinese with English abstract).
Received on June 16,1996; accepted on October 1, 1996
318
Dow
nloaded from https://academ
ic.oup.com/plankt/article/19/3/303/1460076 by guest on 03 February 2022





























