Embed Size (px)
Citation preview

A Palaeodietary Study of Stable IsotopeAnalysis from a High-status Burial in theCopper Age: The Montelirio MegalithicStructure at Valencina de la Concepción–Castilleja de Guzmán, SpainM. FONTANALS-COLL,a* M. DÍAZ-ZORITA BONILLAb AND M. E. SUBIRÀa
a GRAPAC Grup de Recerca Aplicada al Patrimoni Cultural, Unitat d’Antropologia Biològica, Departament deBiologia Animal, de Biologia Vegetal i d’Ecologia, Universitat Autònoma de Barcelona, Barcelona, Spainb Department of Geosciences, Biogeologie, University of Tübingen, Tübingen, Germany
ABSTRACT This palaeodietary study presents carbon and nitrogen stable isotope data from human and faunal skeletalremains from the Copper Age settlement of Valencina de la Concepción–Castilleja de Guzmán, located inSeville, Spain. Montelirio, the only Valencina–Castilleja human group from which we have obtained reliablepalaeodietary results, had a diet based on C3 terrestrial resources, including both plants and animals. Theprotein component of the diet consisted mainly of meat, milk and dairy products from livestock as well asC3 plant protein from cereals and pulses. This study compares data from Montelirio, the Copper Age groupfrom Valencina–Castilleja, with the published data from other Iberian Late Neolithic–Copper Age (LN-CA) so-cieties. This comparison reveals a homogeneous diet with some exceptions. Overall, the LN-CA diet in theIberian Peninsula consisted mainly of animal proteins from meat, milk and dairy products from livestock aswell as C3 plant sources from the characteristic agriculture of the societies of these periods. This study alsodemonstrates the minor use of aquatic resources from the Neolithic to the Copper Age period in the IberianPeninsula. Copyright © 2015 John Wiley & Sons, Ltd.
Key words: carbon; Copper Age; nitrogen; palaeodiet; stable isotopes; Valencina de la Concepción–Castilleja de Guzmán
Introduction
The reconstruction of prehistoric diets through stablecarbon and nitrogen isotope analysis from bone colla-gen has already been active for several years. However,because of difficulties with collagen preservation, thesestudies are not always easy, particularly in hot climates,such as those of southern European countries.Until now, few isotopic analyses have been per-
formed on the Iberian Peninsula. Most have focusedon understanding prehistoric populations from theMesolithic (e.g. Garcia-Guixé et al., 2006; Arias andSchulting, 2010; Fernández-López de Pablo et al.,2013; Fontanals-Coll et al., 2014) through the
Neolithic (e.g. Lubell et al., 1994) and Copper Age(Díaz-Zorita Bonilla, 2013) up to the Iron Age (e.g.Fuller et al., 2010; Salazar-García, 2011). Isotopicstudies in the southern area of the Iberian Peninsulaare rarer.The present isotopic study analyses data from hu-
man and faunal skeletal remains from the Copper Agesettlement of Valencina de la Concepción–Castillejade Guzmán (henceforth ‘Valencina–Castilleja’), Seville,Spain (Figure 1).The Iberian Copper Age spanned from ca.
3500–2250cal BC (Díaz del Río, 2004). This period isidentified mainly by the increase in the number andsize of sites, higher population densities andagricultural intensification (García-Sanjuán andMurillo-Barroso, 2012). However, these patterns havevaried throughout the Iberian Peninsula, and regionaldifferences exist (for detailed information on each area,see Gibaja et al., 2012). Burials in megalithic
* Correspondence to: Maria Fontanals-Coll, Unitat d’AntropologiaBiològica, Departament de Biologia Animal, Biologia Vegetal i Ecologia(BABVE), Facultat de Biociències (Edifici C), Universitat Autònoma deBarcelona (UAB), 08193 Cerdanyola del Vallès, Barcelona, Spain.e-mail: [email protected]
Copyright © 2015 John Wiley & Sons, Ltd. Received 30 January 2014Revised 3 December 2014
Accepted 23 December 2014
International Journal of OsteoarchaeologyInt. J. Osteoarchaeol. (2015)Published online in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/oa.2435

monuments were still prevalent in this period, althoughother structures such as pits and natural caves were alsoin use. The burial rite during the third millennium BCseems to be mainly collective; inhumation is predomi-nant, but cremations have also been documented(Agustí, 2002; Silva et al., 2012).During this period, evidence exists of long-distance
networks, craft specialisation and the use of certainraw materials as prestige objects in the funerary con-text. One of the best-studied Copper Age sites thatinclude all these indicators is Valencina–Castilleja,which is located in Seville, in the south-west of theIberian Peninsula.The Copper Age settlement of Valencina–Castilleja
is located 6km from the city of Seville, in south-westSpain. It is situated in the Aljarafe plateau, west ofthe Guadalquivir River, and it covers approximately400ha. This area is very rich in natural resources,and the ground is fertile. Freshwater resources areavailable because it was located next to marine andlake environments and near important forestry re-sources. According to the few published studies aboutthe site, the main two activities were cultivation andlivestock management in which cattle and suids werethe main animals exploited (e.g. Pajuelo and López,2013; Martínez, 2013). These results are consistentwith the exploitation of the landscape since the Neo-lithic period, based on the so-called ‘policultivoganadero’, an extensive pastoralist strategy (Harrison,
1985) along with dry forest management practices(a mixed exploitation of forest resources and livestockfarming). The modification of the landscape washighly visible during Late Prehistory in Iberia fromthe fourth millennium BC to the second millenniumAD (Stevenson and Harrison, 1992).Valencina–Castilleja was first investigated at the end
of the 19th century based on the study of the mega-lithic monuments of La Pastora, Matarrubilla andOntiveros (e.g. Carriazo, 1962; Collantes de Terán,1969). However, investigations have continued sincethen (for a complete review, see Vargas, 2004; Costaet al., 2010; García-Sanjuán, 2013).This site thus represents one of the most inten-
sively excavated Copper Age settlements in Iberia,and many different structures and monuments havebeen identified. The radiocarbon dates suggest thatsettlement at Valencina–Castilleja has a chronologi-cal range from the beginning of the third millennium(3000–2800 cal BC) until the first half of the secondmillennium BC (1710–1404 cal BC 2σ) (Noceteet al., 2011; García-Sanjuán, 2013).In 1998, a major megalithic monument, Montelirio,
similar to La Pastora and Matarrubilla, was discovered(Férnadez and Aycart, 2013). Montelirio representsone of the most recent excavations documented inValencina–Castilleja. The monument is a tholos witha double chamber, oriented on the east–west axis(Fernández and Aycart, 2013). Two individuals were
Figure 1. Location of Valencina–Castilleja, Seville (Spain), and the Late Neolithic–Copper Age sites from the Iberian Peninsula used for the study.
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

found buried in the corridor, and the rest (minimumnumber of individuals=24) were found in the mainchamber (Pecero et al., 2012). The funerary ritual ofthe main chamber was collective, and the burials inboth the corridor and main chamber were found tobe associated with a great diversity of grave goods, in-cluding ceramics, lithics, shells and metals that werefound along with exotic goods such as amber, ivoryand ostrich egg shells. Similar findings have previouslybeen documented at Valencina–Castilleja. For exam-ple, flint and flint objects (e.g. Morgado et al., 2011),ivory (e.g. Schuhmacher et al., 2012), amber (e.g.Murillo-Barroso and Martinón-Torres, 2012) and greenstones (Odriozola et al., 2010) have been found. Thehigh-status burial of Montelirio represents a good ex-ample of a specific segment of the society withexcellent preservation. Montelirio represents an excep-tional burial among the prehistoric monuments ofValencina–Castilleja and also within the Late Prehis-tory of the Spanish south-west. The excellent preserva-tion of the osteological record, the biological remainsand the burial materials make this site unique. In addi-tion, the funerary structure (one of the few large mega-lithic structures) itself is another element of socialdifferentiation. The different chronological phasesand the evolution of this deposit are explained in detailby Fernández and Aycart (2013). This monument rep-resents one of the few well-preserved Copper Ageburials in Iberia that contain high-status objects, in-cluding some allochthonous objects. Therefore, thissite is very significant for the study of social complexityand social differences during Late Prehistory in Iberia.The other studied sector from Valencina–Castilleja
is a surrounding area of the megalithic monument ofLa Pastora. Many negative structures with several pitsand a ditch were found in this sector (Vargas Jiménez,2013), including a circular structure where the individ-ual under study was found.This paper aims to analyse the diet and the subsis-
tence economy of the human individuals found in thehigh-status burial of Montelirio and the La Pastoracomplex from Valencina–Castilleja, by stable carbon
and nitrogen isotope analysis of human and faunalskeletal remains. To highlight similarities and differencesin diet between Late Neolithic and Copper Agesocieties, the Valencina–Castilleja results were comparedwith the published analyses of other contemporary sitesfrom the Iberian Peninsula (García et al., 2010; McClureet al., 2011;Waterman, 2012;Waterman et al., 2014). It isimportant to highlight that the megalithic constructionsreveal continuity between Portuguese and Spanish sites,which reinforces the validity of this comparison. ThePortuguese sites present a long occupational period thatincludes the Copper Age.
Materials and methods
The isotopical analysis performed in this study presentsdata from human and faunal remains collected in theMontelirio megalithic monument and from the LaPastora complex at the Valencina–Castilleja site. TheMontelirio sector dates from 3000 to 2200cal BC, ac-cording to the radiocarbon dating of the funerary con-text (Table 1).The remains include bone material from 15 humans
and five faunal bone samples of Sus sp., Ovis/Capra sp.and Bos taurus from Montelirio and material from onehuman and five faunal samples of Equus sp., Capra sp.,Cervus sp., Sus sp. and B. taurus from La Pastora.The age and sex diagnosis and the osteological
methods used are explained in detail by Pecero et al.(2012). The sampling process was conducted byselecting long bone fragments from lower extremities.All human and faunal individuals available for stableisotope analysis were sampled.The stable carbon and nitrogen isotope analyses
from bone collagen provide information about the die-tary content of the individuals, based on the protein in-take. In this sense, the stable carbon isotope value(δ13C) distinguishes the consumption of marine (13C-enriched) versus terrestrial C3-based foods, with theo-retical endpoints of approximately �21‰ to �20‰for a major terrestrial protein intake and approximately
Table 1. Radiocarbon-calibrated ages from Montelirio sector
Site Laboratory code Context 14C age BP14C age BC (calibrated 2σ) Source
Montelirio Ua-40804 Funerary 3862± 30 2470–2200 Fernández and Aycart, 2013Ua-40802 Funerary 4002± 31 2580–2460Ua-40805 Funerary 4086± 35 2862–2494CNA-588 Funerary 4060± 40 2851–2476CNA-585 Funerary 4250± 35 2923–2698Ua-40801 Funerary 4180± 30 2890–2660Ua-40803 Funerary 4165± 30 2880–2630CNA-586 Funerary 4140± 50 2880–2575
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

�12‰ for a purely marine protein consumption. Thisvalue also helps to discriminate between the input ofC3 (13C-depleted) and C4 (13C-enriched) plant sourcesinto the diet (Schoeninger and DeNiro, 1984;Schwarcz and Schoeninger, 1991; Richards, 2002;Lillie, 2003; Eriksson et al., 2008, Lillie, et al., 2011).However, when the consumption of both C4 plantsand marine resources is present in the individual’s diet,there could appear a difficulty in the interpretation. Butthis complication does not arise in this study because ofthe insignificant quantities of edible C4 plants duringthe Chalcolithic in Southern Iberia. The stable nitro-gen isotope value (δ15N) provides information aboutthe trophic level of each individual: the values of con-sumers present an increase of 3–5‰ over the valuesof their diets in the food chain (De Niro and Epstein,1981; Schoeninger and De Niro, 1984; Bocherens andDrucker, 2003; Richards et al., 2005; Lillie et al., 2011).However, it is always important to take into account thedifficulty of distinguishing animal-rich diets from plant-rich diets using δ15N values discussed in some studies suchas Hedges and Reynard (2007) and Fraser et al. (2013).The stable isotope signature of an adult human bone
reflects the average diet of at least the last 10–15yearsof life because of the rate of the bone turnover duringthe individual’s lifetime (Hedges et al., 2007).Established quality indicators are used to ensure thepreservation quality of the extracted collagen(De Niro, 1985; Ambrose, 1993; Van Klinken, 1999).The collagen extraction process for this study was per-
formed at the laboratories of the Unitat d’AntropologiaBiològica of theUniversitat Autònoma de Barcelona. Priorto analysis, mechanical abrasion with a multifunction toolDremel 3000 (Dremel Europe), with a diamond engravingtip, was used to remove visible contaminants. Collagenextraction proceeded following Richards and Hedges(1999), including Brown et al. (1988) and modificationsreferring to the ultrafiltration step. The measurementsfor the stable isotope analysis were performed at theInstitut de Ciència i Teconologia Ambientals using aThermo Flash 1112 (Thermo Scientific) elementalanalyser coupled to a Thermo Delta V Advantage massspectrometer (Thermo Scientific) with a Conflo III inter-face, which measures the ratios of 13C to 12C and 15Nto 14N relative to a standard (Vienna Pee Dee Belemnite(V-PDB) for carbon and air for nitrogen) and expressesthe stable isotope values in parts per thousand (‰) andin delta notation (δ). The international laboratory stan-dard, IAEA 600 (caffeine), is used here. An analyticalerror always below 0.2‰ (1σ) in all the repeated analyseswas determined for δ13C and δ15N. The descriptive statis-tics were calculated using the IBM Statistics Software SPSS20.0.0 (Armonk, NY, USA). Ta
ble
2.Stable
carbon
(δ13C)an
dnitrog
en(δ
15N)va
lues
obtained
from
faun
alsa
mplesfrom
Valen
cina
–Cas
tilleja:M
ontelirio
andLa
Pas
tora,ind
icatingco
llagen
control
indicators(yield,%
C,%
Nan
dC:N
)an
dsa
mpledbon
e
Site
Figures
code
Laboratory
code
Individua
lTa
xaδ1
3C
δ15N
Yield
(%)
%C
%N
C:N
Sam
pledbon
e
Mon
telirio
12MoF17
UE-253
Sus
sp.
�19.6
4.2
1.3
40.9
14.4
3.3
Pha
lanx
13MoF18
UE-253
Ovis/Cap
rasp
.�1
9.4
5.6
1.1
41.6
14.7
3.3
Long
bon
ediaphy
sis
14MoF19
UE-235
Ovis/Cap
rasp
.�1
8.7
5.6
0.9
45.3
16.1
3.3
Long
bon
ediaphy
sis
—MoF20
.1UE-17Ind.1
Bos
taurus
——
——
——
Lefttib
ia—
MoF20
.2UE-17Ind.2
Ovis/Cap
rasp
.—
——
——
—Lo
ngbon
ediaphy
sis
La Pas
tora
—Pa1
UC54
UD72
(63)
Equu
ssp
.—
——
——
—Lo
ngbon
ediaphy
sis
—Pa4
UC20
6UD20
7(101
)Cervu
ssp
.�2
0.8
4.7
0.8
35.1
12.7
2.8a
Long
bon
ediaphy
sis
15Pa5
UC54
UD72
(13)
Cap
rasp
.�2
0.4
5.9
1.1
39.8
13.7
2.9
Long
bon
ediaphy
sis
16Pa6
UC20
6UD23
4(23)
Bos
taurus
�18.7
6.2
1.4
38.9
12.7
2.9
Leftfemur
—Pa7
UC20
6UD23
4(36)
Sus
sp.
�20.6
9.0
0.5
38.9
14.1
2.8a
Long
bon
ediaphy
sis
aValue
sfalling
outsidetheac
ceptedrang
eforgoo
dco
llagen
prese
rvation.
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)
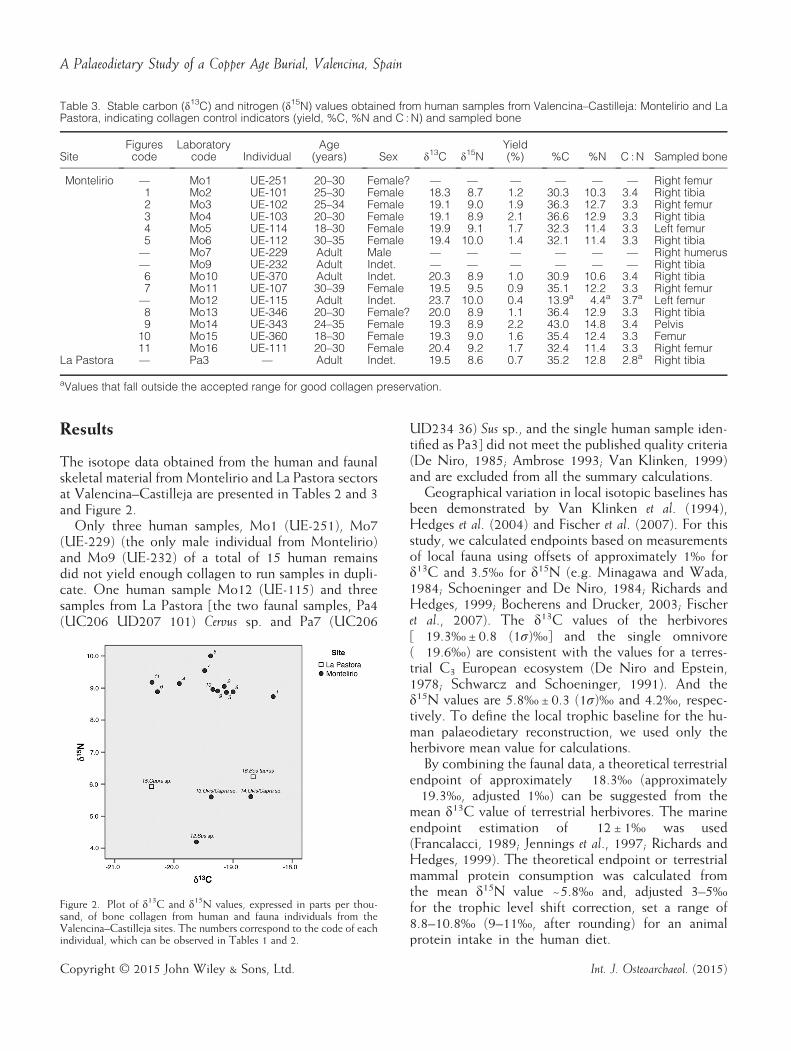
Results
The isotope data obtained from the human and faunalskeletal material from Montelirio and La Pastora sectorsat Valencina–Castilleja are presented in Tables 2 and 3and Figure 2.Only three human samples, Mo1 (UE-251), Mo7
(UE-229) (the only male individual from Montelirio)and Mo9 (UE-232) of a total of 15 human remainsdid not yield enough collagen to run samples in dupli-cate. One human sample Mo12 (UE-115) and threesamples from La Pastora [the two faunal samples, Pa4(UC206 UD207 101) Cervus sp. and Pa7 (UC206
UD234 36) Sus sp., and the single human sample iden-tified as Pa3] did not meet the published quality criteria(De Niro, 1985; Ambrose 1993; Van Klinken, 1999)and are excluded from all the summary calculations.Geographical variation in local isotopic baselines has
been demonstrated by Van Klinken et al. (1994),Hedges et al. (2004) and Fischer et al. (2007). For thisstudy, we calculated endpoints based on measurementsof local fauna using offsets of approximately 1‰ forδ13C and 3.5‰ for δ15N (e.g. Minagawa and Wada,1984; Schoeninger and De Niro, 1984; Richards andHedges, 1999; Bocherens and Drucker, 2003; Fischeret al., 2007). The δ13C values of the herbivores[�19.3‰±0.8 (1σ)‰] and the single omnivore(�19.6‰) are consistent with the values for a terres-trial C3 European ecosystem (De Niro and Epstein,1978; Schwarcz and Schoeninger, 1991). And theδ15N values are 5.8‰±0.3 (1σ)‰ and 4.2‰, respec-tively. To define the local trophic baseline for the hu-man palaeodietary reconstruction, we used only theherbivore mean value for calculations.By combining the faunal data, a theoretical terrestrial
endpoint of approximately �18.3‰ (approximately�19.3‰, adjusted 1‰) can be suggested from themean δ13C value of terrestrial herbivores. The marineendpoint estimation of �12±1‰ was used(Francalacci, 1989; Jennings et al., 1997; Richards andHedges, 1999). The theoretical endpoint or terrestrialmammal protein consumption was calculated fromthe mean δ15N value ~5.8‰ and, adjusted 3–5‰for the trophic level shift correction, set a range of8.8–10.8‰ (9–11‰, after rounding) for an animalprotein intake in the human diet.
Table 3. Stable carbon (δ13C) and nitrogen (δ15N) values obtained from human samples from Valencina–Castilleja: Montelirio and LaPastora, indicating collagen control indicators (yield, %C, %N and C :N) and sampled bone
SiteFigurescode
Laboratorycode Individual
Age(years) Sex δ13C δ15N
Yield(%) %C %N C :N Sampled bone
Montelirio ― Mo1 UE-251 20–30 Female? ― ― ― ― ― ― Right femur1 Mo2 UE-101 25–30 Female �18.3 8.7 1.2 30.3 10.3 3.4 Right tibia2 Mo3 UE-102 25–34 Female �19.1 9.0 1.9 36.3 12.7 3.3 Right femur3 Mo4 UE-103 20–30 Female �19.1 8.9 2.1 36.6 12.9 3.3 Right tibia4 Mo5 UE-114 18–30 Female �19.9 9.1 1.7 32.3 11.4 3.3 Left femur5 Mo6 UE-112 30–35 Female �19.4 10.0 1.4 32.1 11.4 3.3 Right tibia
― Mo7 UE-229 Adult Male ― ― ― ― ― — Right humerus― Mo9 UE-232 Adult Indet. ― ― ― ― ― — Right tibia6 Mo10 UE-370 Adult Indet. �20.3 8.9 1.0 30.9 10.6 3.4 Right tibia7 Mo11 UE-107 30–39 Female �19.5 9.5 0.9 35.1 12.2 3.3 Right femur
― Mo12 UE-115 Adult Indet. �23.7 10.0 0.4 13.9a 4.4a 3.7a Left femur8 Mo13 UE-346 20–30 Female? �20.0 8.9 1.1 36.4 12.9 3.3 Right tibia9 Mo14 UE-343 24–35 Female �19.3 8.9 2.2 43.0 14.8 3.4 Pelvis
10 Mo15 UE-360 18–30 Female �19.3 9.0 1.6 35.4 12.4 3.3 Femur11 Mo16 UE-111 20–30 Female �20.4 9.2 1.7 32.4 11.4 3.3 Right femur
La Pastora ― Pa3 ― Adult Indet. �19.5 8.6 0.7 35.2 12.8 2.8a Right tibia
aValues that fall outside the accepted range for good collagen preservation.
Figure 2. Plot of δ13C and δ15N values, expressed in parts per thou-sand, of bone collagen from human and fauna individuals from theValencina–Castilleja sites. The numbers correspond to the code of eachindividual, which can be observed in Tables 1 and 2.
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)
The mean δ13C value of the human group fromValencina–Castilleja is �19.5±0.6 (1σ)‰, and themean δ15N value is 9.1±0.4 (1σ)‰. These values areconsistent with a diet based mainly on C3 terrestrial re-sources and place all humans in a higher trophic levelthan the herbivores (3.3‰ higher). There is no traceof marine protein consumption in their diet, but theisotopic evidence presented here does not exclude theoccasional consumption of freshwater resources. How-ever, the δ15N values of human individuals reflect nosignificant consumption of these resources and high-light the necessity to increase the sample size inregional freshwater fauna to provide more informationon this subject.
Discussion
The values of the Montelirio human group fromValencina–Castilleja are indicative of a homogeneousterrestrial C3 diet. There is no clear isotopic evidence
for C4 plant intake on the diet of this community.The contribution of domesticated C3 plants in the dietcould come from wheat and barley, the most com-monly used cereals during the Neolithic in the IberianPeninsula until the Middle Bronze Age where the earli-est millet (C4 pathway plant) was found as a crop(Rovira, 2007).The animal protein component of the diet consisted
mainly of meat, milk and dairy products from livestock.But some studies (e.g. Hedges and Reynard, 2007;O’Connell et al., 2012; Fraser et al., 2013) suggest thatthe interpretation of a very high animal protein intakein prehistoric farmers could have often beenoverestimated because of some limitations in the dis-tinction of animal-rich diets from plant-rich diets usingδ15N values.To resolve these limitations, we construct a model
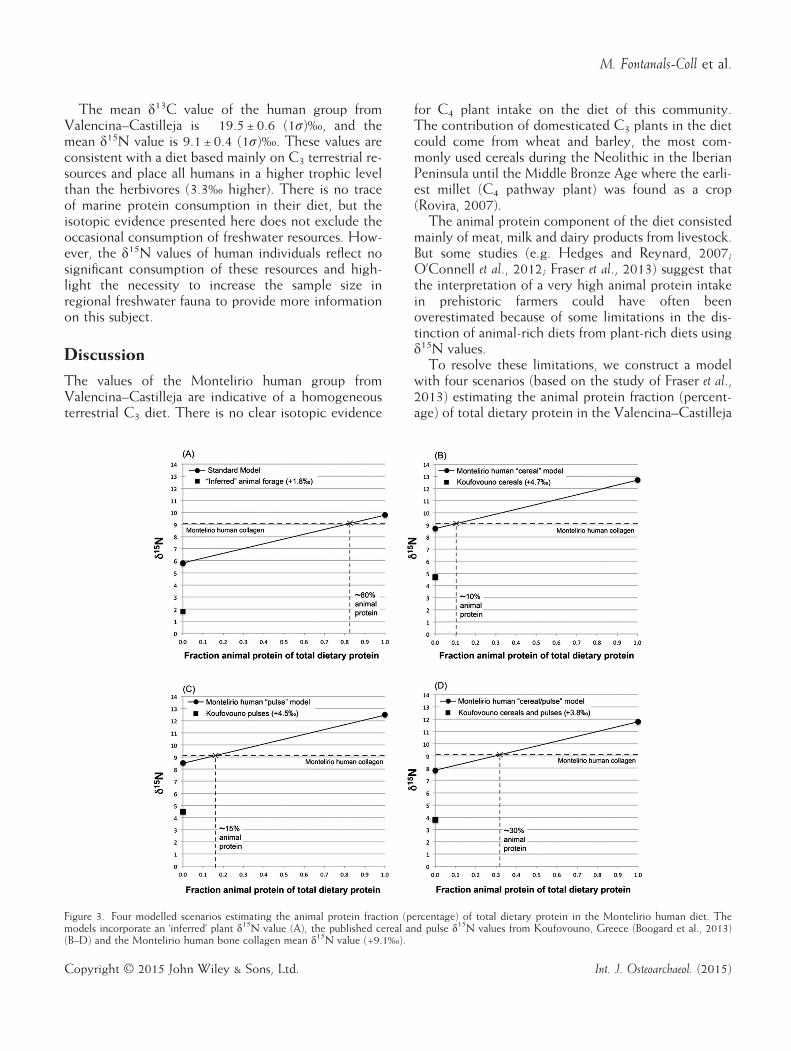
with four scenarios (based on the study of Fraser et al.,2013) estimating the animal protein fraction (percent-age) of total dietary protein in the Valencina–Castilleja
Figure 3. Four modelled scenarios estimating the animal protein fraction (percentage) of total dietary protein in the Montelirio human diet. Themodels incorporate an ‘inferred’ plant δ15N value (A), the published cereal and pulse δ15N values from Koufovouno, Greece (Boogard et al., 2013)(B–D) and the Montelirio human bone collagen mean δ15N value (+9.1‰).
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

human diet (Figure 3). And for it, we also consider thevalue of +4‰ (the midpoint of the usually δ15N offsets+3‰ to +5‰) to represent the ‘trophic level’ differ-ences for the δ15N values and the Δplant–herbivore offsetfor δ15N values.As can be seen in Figure 3, the first scenario (A)
represents the standard model (Hedges and Reynard,2007) in which we have calculated the ‘inferred’plant diet δ15N value by subtracting a fractionation(Δplant–herbivore) amount from the herbivore collagenδ15N value, assuming that humans and herbivores areconsuming plant resources of the same isotopic value.This standard model [Figure 3(A)] predicts that the meanδ15N value ofMontelirio human individuals (9.1‰) indi-cates a diet comprising ≥80% of animal protein. AsHedges and Reynard (2007) point out, in prehistoricagricultural societies, this high proportion of animalprotein is not impossible but requires enough cattle tosustain the community with enough meat and dairyproducts. This high amount of animal protein differsfrom all current agrarian subsistence economies and fitsbetter with known pastoralist economies where milk isa major item of nutrition.The three following dietary modelling scenarios [Fig-
ure 3(B–D)] are constructed using the crops and pulsesδ15N values and applies the same +4‰ enrichment to-wards higher trophic levels. Unfortunately, we couldnot analyse any remain of cereals or pulses fromValencina–Castilleja, and for this reason, we use thepublished δ15N values from the Middle–Late Neolithicsite of Koufovouno, Greece (Bogaard et al., 2013),which could represent the archaeobotanical data forthe Mediterranean environment. Using the Greecearchaeobotanical values, we are able to define the startand endpoint δ15N values for the prediction lines foreach model and, consequently, estimate the animal pro-tein fraction of the human diet. Their starting points arethe consumption of 100% plant protein: the B modelrepresents a human consumption of 100% cereal grain,the C model represents a 100% pulse consumer and theD model represents a 50:50 mixed cereal and pulse con-sumer. The estimated animal protein fractions of humandiet in these scenarios are ≥10%, ≥15% and ≥30%, res-pectively. These three values are not similar to the per-centage of animal protein obtained in the standardmodel, because their starting δ15N values (B: +4.7‰,C: +4.5‰ and D: +3.8‰) are not close to the ‘inferred’plant diet δ15N value (A: +1.8‰). In sum, the animalprotein fractions obtained suggest that these three sub-sequent scenarios estimate human diets with drasticallylower percentages of animal protein and that plants pro-vide most of the protein in the diet. In this sense, thehuman individuals from Montelirio would have a lower
intake of animal protein if a significant consumption ofcereals, pulses or a mixture of both crops had occurred.It is far more plausible that the protein component ofthe Montelirio human diet consisted mainly of thatmixture of cereals and pulses in a regular basis (almost,probably, 70%), as well as animal protein from meat,milk and dairy products (almost, probably, 30%), asthe D model predicted. However, it is important tonote the relevance of regional archaeobotanical datato conduct proper analysis on this subject and the con-sequent human dietary reconstruction. These modelshighlight the importance of the stable isotope analysisof the associated crop materials to not underestimatethe role of cereals and pulses in the human diet.Among the homogeneous human diet observed in
Montelirio, it is interesting to point two cases individu-ally that, despite not showing statistically significant dif-ferences with the rest of the group, present a higherconsumption of animal protein. These individuals identi-fied as 5.Mo6 (UE-112) and 7.Mo11 (UE-107), with10‰ and 9.5‰ δ15N values, respectively, are the twooldest women of the group studied. Our first hypothesissuggests that these two women have, probably, a highersocial status due to their age that could lead to a dietarydifferentiation. This hypothesis could be corroboratedby the lower intake of animal protein (1‰ lower δ15Nvalues) by the younger individuals of the group, allwomen with ages between 18 and 30years. Anotherhypothesis suggests that these lower δ15N values forthe younger women might be more related to their preg-nancies (they are the group most affected by pregnancy)that would produce a little reduction of δ15N values(0.3–1.1‰) from conception to birth (Fuller et al., 2004).Further zooarchaeological, archaeobotanical and
isotopical studies including more regional wild and do-mesticated fauna, and freshwater resources, should helpto clarify the specific proportion of animal protein anddomesticated crops in human diet and the social rolesthat could lead to an increase or a reduction of certainfoods in their diet.
The comparison of Montelirio data with other LateNeolithic–Copper Age series of the Iberian Peninsula
For a better understanding of Copper Age subsistencepatterns in the Iberian Peninsula, we compared theValencina–Castilleja results with other Iberian LateNeolithic–Copper Age series that have been published.The contextualised data and the mean values calculatedfor each site are displayed in Table 4.The Montelirio group was compared with nine
published studies of Late Neolithic–Copper Age sites
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)
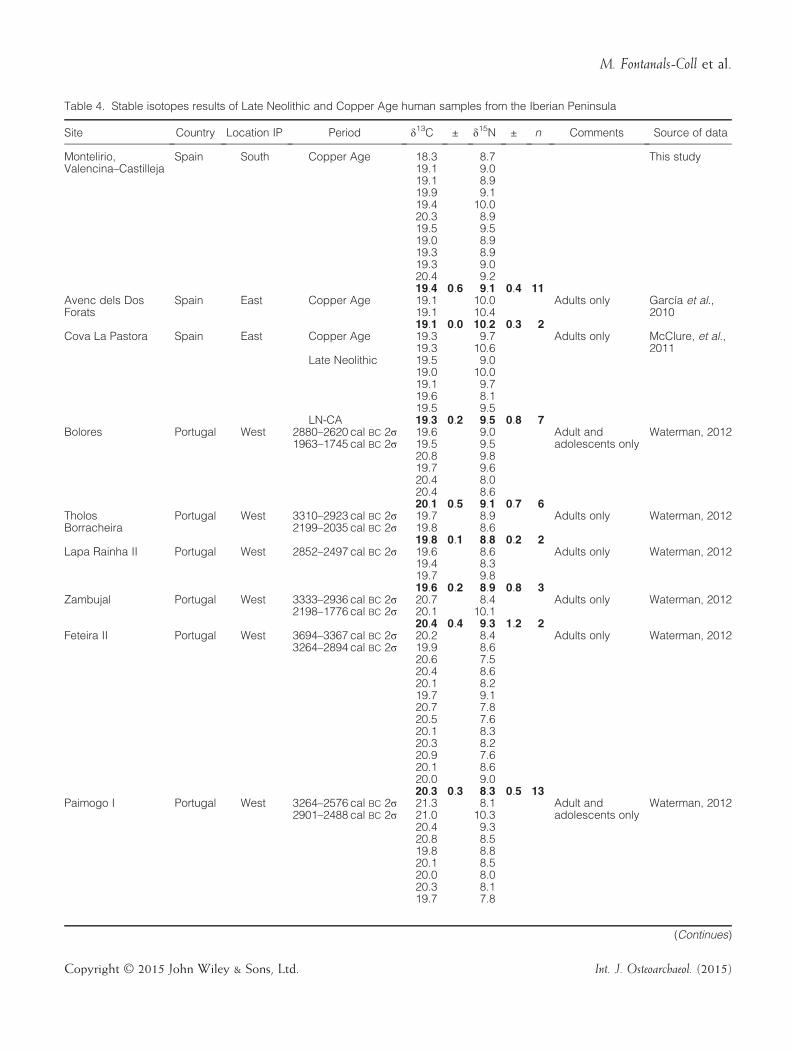
Table 4. Stable isotopes results of Late Neolithic and Copper Age human samples from the Iberian Peninsula
Site Country Location IP Period δ13C ± δ15N ± n Comments Source of data
Montelirio,Valencina–Castilleja
Spain South Copper Age �18.3 8.7 This study�19.1 9.0�19.1 8.9�19.9 9.1�19.4 10.0�20.3 8.9�19.5 9.5�19.0 8.9�19.3 8.9�19.3 9.0�20.4 9.2�19.4 0.6 9.1 0.4 11
Avenc dels DosForats
Spain East Copper Age �19.1 10.0 Adults only García et al.,2010�19.1 10.4
�19.1 0.0 10.2 0.3 2Cova La Pastora Spain East Copper Age �19.3 9.7 Adults only McClure, et al.,
2011�19.3 10.6Late Neolithic �19.5 9.0
�19.0 10.0�19.1 9.7�19.6 8.1�19.5 9.5
LN-CA �19.3 0.2 9.5 0.8 7Bolores Portugal West 2880–2620 cal BC 2σ
1963–1745 cal BC 2σ�19.6 9.0 Adult and
adolescents onlyWaterman, 2012
�19.5 9.5�20.8 9.8�19.7 9.6�20.4 8.0�20.4 8.6�20.1 0.5 9.1 0.7 6
TholosBorracheira
Portugal West 3310–2923 cal BC 2σ2199–2035 cal BC 2σ
�19.7 8.9 Adults only Waterman, 2012�19.8 8.6�19.8 0.1 8.8 0.2 2
Lapa Rainha II Portugal West 2852–2497 cal BC 2σ �19.6 8.6 Adults only Waterman, 2012�19.4 8.3�19.7 9.8�19.6 0.2 8.9 0.8 3
Zambujal Portugal West 3333–2936 cal BC 2σ2198–1776 cal BC 2σ
�20.7 8.4 Adults only Waterman, 2012�20.1 10.1�20.4 0.4 9.3 1.2 2
Feteira II Portugal West 3694–3367 cal BC 2σ3264–2894 cal BC 2σ
�20.2 8.4 Adults only Waterman, 2012�19.9 8.6�20.6 7.5�20.4 8.6�20.1 8.2�19.7 9.1�20.7 7.8�20.5 7.6�20.1 8.3�20.3 8.2�20.9 7.6�20.1 8.6�20.0 9.0�20.3 0.3 8.3 0.5 13
Paimogo I Portugal West 3264–2576 cal BC 2σ2901–2488 cal BC 2σ
�21.3 8.1 Adult andadolescents only
Waterman, 2012�21.0 10.3�20.4 9.3�20.8 8.5�19.8 8.8�20.1 8.5�20.0 8.0�20.3 8.1�19.7 7.8
(Continues)
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)
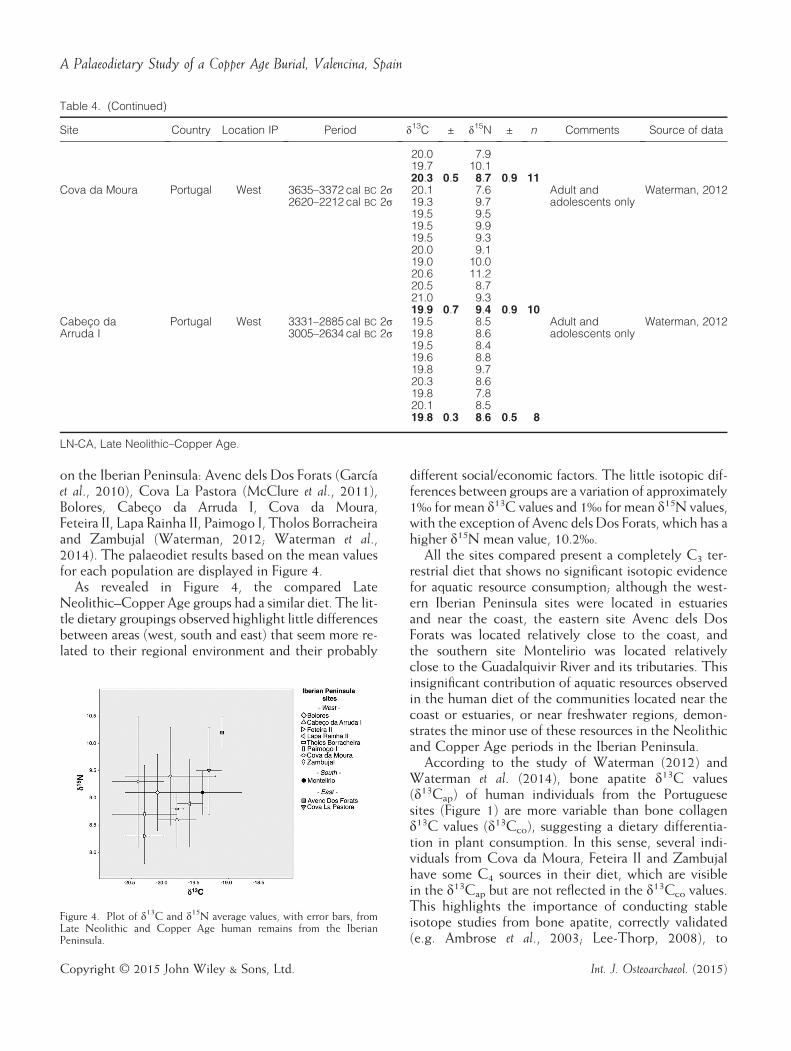
on the Iberian Peninsula: Avenc dels Dos Forats (Garcíaet al., 2010), Cova La Pastora (McClure et al., 2011),Bolores, Cabeço da Arruda I, Cova da Moura,Feteira II, Lapa Rainha II, Paimogo I, Tholos Borracheiraand Zambujal (Waterman, 2012; Waterman et al.,2014). The palaeodiet results based on the mean valuesfor each population are displayed in Figure 4.As revealed in Figure 4, the compared Late
Neolithic–Copper Age groups had a similar diet. The lit-tle dietary groupings observed highlight little differencesbetween areas (west, south and east) that seem more re-lated to their regional environment and their probably
different social/economic factors. The little isotopic dif-ferences between groups are a variation of approximately1‰ for mean δ13C values and 1‰ for mean δ15N values,with the exception of Avenc dels Dos Forats, which has ahigher δ15N mean value, 10.2‰.All the sites compared present a completely C3 ter-
restrial diet that shows no significant isotopic evidencefor aquatic resource consumption; although the west-ern Iberian Peninsula sites were located in estuariesand near the coast, the eastern site Avenc dels DosForats was located relatively close to the coast, andthe southern site Montelirio was located relativelyclose to the Guadalquivir River and its tributaries. Thisinsignificant contribution of aquatic resources observedin the human diet of the communities located near thecoast or estuaries, or near freshwater regions, demon-strates the minor use of these resources in the Neolithicand Copper Age periods in the Iberian Peninsula.According to the study of Waterman (2012) and
Waterman et al. (2014), bone apatite δ13C values(δ13Cap) of human individuals from the Portuguesesites (Figure 1) are more variable than bone collagenδ13C values (δ13Cco), suggesting a dietary differentia-tion in plant consumption. In this sense, several indi-viduals from Cova da Moura, Feteira II and Zambujalhave some C4 sources in their diet, which are visiblein the δ13Cap but are not reflected in the δ13Cco values.This highlights the importance of conducting stableisotope studies from bone apatite, correctly validated(e.g. Ambrose et al., 2003; Lee-Thorp, 2008), to
Figure 4. Plot of δ13C and δ15N average values, with error bars, fromLate Neolithic and Copper Age human remains from the IberianPeninsula.
Table 4. (Continued)
Site Country Location IP Period δ13C ± δ15N ± n Comments Source of data
�20.0 7.9�19.7 10.1�20.3 0.5 8.7 0.9 11
Cova da Moura Portugal West 3635–3372 cal BC 2σ2620–2212 cal BC 2σ
�20.1 7.6 Adult andadolescents only
Waterman, 2012�19.3 9.7�19.5 9.5�19.5 9.9�19.5 9.3�20.0 9.1�19.0 10.0�20.6 11.2�20.5 8.7�21.0 9.3�19.9 0.7 9.4 0.9 10
Cabeço daArruda I
Portugal West 3331–2885 cal BC 2σ3005–2634 cal BC 2σ
�19.5 8.5 Adult andadolescents only
Waterman, 2012�19.8 8.6�19.5 8.4�19.6 8.8�19.8 9.7�20.3 8.6�19.8 7.8�20.1 8.5�19.8 0.3 8.6 0.5 8
LN-CA, Late Neolithic–Copper Age.
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

employ linear dietary models such as the one proposedby Kellner and Schoeninger (2007).Overall, the similarities in δ13C values could demon-
strate that these populations practised a similar type ofagriculture and animal husbandry, where the C3 cropsselected for the cultivation grew successfully in the en-vironment of each region and the cattle grazed on aland with similar but not identical vegetation. Theseenvironmental differences between regions could havedirectly affected the diet of the fauna and, conse-quently, could also lead to these slight dietary differ-ences between human communities.It is important to note that Zambujal, Cova da
Moura and Paimogo I have a higher deviation in theirmean values. The intra-population heterogeneity ofthe diet of these societies, specially the variation inδ15N values among individuals, could be explained bythe existence of social roles that could have favoureda sector of the population with a differential protein in-take. However, it is important to highlight the smallsample size of Zambujal that could produce thesedifferences.Finally, the similarities in δ15N values, in conjunc-
tion with δ13C values, indicate that these humangroups present a significant consumption of animal pro-tein from meat, milk and dairy products from terrestrialC3 mammals, as well as C3 plant protein from cerealsand pulses. These animal resources came from thepopulation’s livestock, which possibly ate C3 plantsfrom pastures and meadows. The C3 plant protein came,probably, from a characteristic agriculture composed ofwheat, barley and some beans and possible C3 plantsources gathering in their surrounding areas.
Conclusions
The palaeodietary reconstruction of the human indi-viduals from the high-status burial from Montelirio(Valencina–Castilleja) demonstrates that this commu-nity has a homogeneous diet based on C3 terrestrial re-sources. The principal intake came from C3 animalsources, but the amount of animal protein may be re-duced to ≥30% or maybe less (≥15% or ≥10%) by asignificant consumption of C3 crops, highlighting thatmost of the protein come from C3 plant sources.The comparisons made with published data about
Late Neolithic–Copper Age societies on the IberianPeninsula indicate that the diet of these different com-munities was fairly homogeneous in these periods. Theprotein component of the diet consisted mainly ofmeat, milk and dairy products from livestock as wellas the C3 vegetable component from the characteristic
agriculture of the societies of these periods. This studyalso demonstrates the minor use of aquatic resourcesfrom the Neolithic to Copper Age periods in theIberian Peninsula.The present study is one of the first analyses of the
dietary content of Late Neolithic–Copper Age commu-nities in southern Spain, and more stable isotope datafrom different contexts such as isotopic data fromlower-status burials, including more faunal remains,are needed to clarify those issues commented earlierand to assert these assumptions about the LateNeolithic and Copper Age communities.
Acknowledgements
We would like to thank the directors of the archaeo-logical excavation of Montelirio, Álvaro FernándezFlores and Vicente Aycart, for entrusting us with thepresent analysis. Thanks also to J.C. Espín, J.M. Guijo,R. Lacalle, P. López and A. Pajuelo for providing thesamples. Finally, we would like to thank the two anon-ymous reviewers for their helpful contributions.This work has been supported by ‘Dirección General
de Bienes Culturales de la Consejería de Cultura de laJunta de Andalucía’, ‘Ayuntamiento de Valencina de laConcepción (Sevilla)’ and Generalitat de Catalunya(SGR2009-566).
References
Agustí B. 2002. Depósitos funerarios con cremación duranteel Calcolítico y el Bronce en el nordeste de Catalunya. InSobre el significado del fuego en los rituales funerarios del Neolítico,MA Rojo, M Kunst (ed.) Studia Archaeologica 91.Universidad de Valladolid: Valladolid; 65–82.
Ambrose SH. 1993. Isotopic analysis of palaeodiets:methodological and interpretive considerations. In In-vestigations of Ancient Human Tissue, Chemical Analyses in An-thropology, Sandford MK (ed.), Gordon and BreachScience Publishers, Langhorne, Pennsylvania: USA;59–130.
Ambrose SH, Buikstra J, Krueger HW. 2003. Status andgender differences in diet at Mound 72, Cahokia, revealedby isotopic analysis of bone. Journal of AnthropologicalArchaeology 22: 217–226.
Arias P, Schulting RJ. 2010. Análisis de isótopos establessobre los restos humanos de la Braña-Arintero.Aproximación a la dieta de los grupos Mesolíticos de laCordillera Cantábrica. In Los Hombres Mesolíticos de la Cuevade La Braña-Arintero (Valdelugueros, León), JM Vidal, MEPrada. (ed.) Museo de León, Estudios y Catálogos 18,Junta de Castilla y León: León; 130–137.
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

Bocherens H, Drucker D. 2003. Trophic level isotopic en-richments for carbon and nitrogen in bone collagen: Casestudies from recent and ancient terrestrial ecosystems.Journal of Osteoarchaeology 13: 46–53.
Boogard A, Fraser R, Heaton THE, Wallace M, Vaiglova P,Charles M, Jones G, Evershed RP, Styring AK, AndersenNH, Arbogast RM, Bartosiewicz L, Gardeisen A, KanstrupM, Maier U, Marinova E, Nivov L, Schäfer M, Stephan E.2013. Crop manuring and intensive land management byEurope’s first farmers. Proceedings of the National Academy ofSciences 110 (31): 12589–12594.
Brown TA, Nelson DE, Vogel SJ, Southon JR. 1988. Im-proved collagen extraction by modified Longin method.Radiocarbon 30: 171–178.
Carriazo, Arroquia JM. 1962. El dolmen de Ontiveros(Valencina de la Concepción, Sevilla). In Homenaje alProfesor Cayetano de Mergelina. Universidad deMurcia: Murcia;209–229
Collantes de Terán F. 1969. El dolmen de Matarrubilla. Actasdel V Symposium Internacional de Prehistoria Peninsular. Tartessosy sus Problemas (Jerez, 1968). Francisco Collantes de Terán:Barcelona; 47–61.
Costa ME, Díaz-Zorita M, García L, Wheatley D. 2010. TheCopper Age settlement of Valencina de la Concepción(Seville, Spain): Demography, metallurgy and spatial orga-nization. Trabajos de Prehistoria 67 (1): 87–118.
De Niro MJ, Epstein S. 1978. Influence of diet on the distri-bution of carbon isotopes in animals. Geochimica etCosmochimica Acta 49: 97–115.
De Niro MJ, Epstein S. 1981. Influence of diet on the distri-bution of nitrogen isotopes in animals. Geochimica etCosmochimica Acta 45: 341–351.
De Niro MJ. 1985. Postmortem preservation and alterationof in vivo bone collagen isotope ratios in relation topaleodietary reconstruction. Nature 317: 806–809.
Díaz del Río P. 2004. Copper Age ditched enclosures incentral Iberia. Oxford Journal of Archaeology 23 (2):107–121.
Díaz-Zorita Bonilla, M. 2013. The Copper Age in south-west Spain: a bioarchaeological approach to prehis-toric social organisation. PhD dissertation, DurhamUniversity.
Eriksson G, Linderholm A, Fornander E, Kanstrup M,Schoultz P, Olofsson H. 2008. Same island, different diet:Cultural evolution of food practice on Öland, Sweden,from the Mesolithic to the Roman Period. Journal ofAnthropological Archaeology 27: 520–543.
Fernández A, Aycart V. 2013. El dolmen de Montelirio. Unsepulcro clave para la comprensión del registro de losgrandes monumentos funerarios de Valencina–Castilleja.In Valencina Prehistórica. Actas del Congreso Conmemorativo delDescubrimiento de La Pastora (1860–2010). L García, VHurtado, JM Vargas, T Ruiz, R Cruz-Auñón (ed.)Universidad de Sevilla: Seville; 233–260.
Fernández-López de Pablo J, Salazar-García DC, Subirà ME,Roca de Togores C, Gomez-Puche M, Richards MP,Esquembre-Bebia A. 2013. Late Mesolithic burials at
Casa Corona (Villena, Spain): Direct radiocarbon andpaleodietary evidence of the last forager populations inEastern Iberia. Journal of Archaeological Science 40(1):671–680.
Fischer A, Olsen J, Richards MP, Heinemeier J,Sveinbjornsdottir A, Bennike P. 2007. Coast–inland mo-bility and diet in the Danish Mesolithic and Neolithic: Ev-idence from stable isotope values of humans and dogs.Journal of Archaeological Science 34: 2125–2150.
Fontanals-Coll M, Subirà ME, Marín-Moratalla N, Ruiz J,Gibaja JF. 2014. From Sado Valley to Europe: Mesolithicdietary practices through different geographic distribu-tions. Journal of Archaeological Science 50: 539–550.
Francalacci P. 1989. Comparison of archaeological, trace el-ement and stable isotope data from two Italian coastalsites. Rivista di Antropologia 66: 239–250.
Fraser RA, Bogaard A, Schäfer M, Arbogast R, Heaton THE.2013. Integrating botanical, faunal and human stable car-bon and nitrogen isotope values to reconstruct land useand palaeodiet at LBK Vaihingen an der Enz, Baden-Württemberg. World Archaeology 45 (3): 492–517.
Fuller BT, Fuller JL, Sage NE, Harris DA, O’Connell TC,Hedges REM. 2004. Nitrogen balance and δ15N: Whyyou’re not what you eat during pregnancy. Rapid Communi-cations in Mass Spectrometry 18: 2889–2896.
Garcia-Guixé E, Richards MP, Subirà ME. 2006. Paleodietsof humans and fauna at Spanish Mesolithic site of ElCollado. Current Anthropology 47(3): 549–556.
Fuller BT, Márquez-Grant N, Richards MP. 2010. Investiga-tion of diachronic dietary patterns on the islands of Ibizaand Formentera, Spain: Evidence from carbon and nitro-gen stable isotope ratio analysis. American Journal of PhysicalAnthropology 143: 512–522.
García O, Cotino F, Miret C, Pascual JL, McClure SB,Molina Ll, Alapont Ll, Carrión Y, Morales JV, Blasco J,Culleton B. 2010. Cavidades de uso funeario durante elNeolítico final/Calcolítico en el territorio valenciano:Trabajos arqueológicos en Avenc dels Dos Forats o Covadel Monedero (Carcaixent, Valencia). Archivo de PrehistoriaLevantina Valencia XXVIII: 139–206.
García Sanjuán L, Murillo-Barroso M. 2012. Social complex-ity in Copper Age Southern Iberia (c. 3200–2200 cal BC):reviewing the ‘state’ hypothesis at Valencina de laConcepción (Seville, Spain). In The Prehistory of Iberia:Debating Early Social Stratification and the State, M Cruz,L García, A Gilman. (ed.). Routledge: New York;119–140.
García-Sanjuán L. 2013. El asentamiento de la Edad delCobre de Valencina de la Concepción: estado actual dela investigación, debates y perspectivas. In ValencinaPrehistórica. Actas del Congreso Conmemorativo del Descubrimientode La Pastora (1860–2010), García L, V Hurtado, JM Vargas,T Ruiz, R Cruz-Auñón (ed.), Universidad de Sevilla: Se-ville; 21–60.
Gibaja JF, Carvalho F, Chambon P, (ed.) 2012. Funerary Prac-tices in the Iberian Peninsula from the Mesolithic to the Chalcolithic.BAR International Series 2417. Arachaeopress: Oxford.
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

Harrison RJ. 1985. The ‘policultivo ganadero’, or the second-ary products revolution in Spanish agriculture, 5000–1000bc. Proceedings of the Prehistoric Society 51: 75–102.
Hedges REM, Stevens RE, Richards MP. 2004. Bone as a sta-ble isotope archive for local climatic information. Quater-nary Science Reviews 23: 959–965.
Hedges REM, Reynard LM. 2007. Nitrogen isotopes and thetrophic level of humans in archaeology. Journal of Archaeo-logical Science 34: 1240–1251.
Hedges REM, Clement JG, Thomas CDL, O’Conell TC.2007. Collagen turnover in the adult femoral mid-shaft:Modelled from anthropogenic radiocarbon tracer mea-surements. American Journal of Physical Anthropology 133:808–816.
Jennings S, Reñones O, Morales-Nin B, Polunin NVC,Moranta J, Coll J. 1997. Spatial variation in the 15N and13C stable isotope composition of plants, invertebrates,and fishes on Mediterranean reefs: Implications for thestudy of trophic pathways. Marine Ecology Progress Series146: 109–116.
Kellner CM, Schoeninger MJ. 2007. A simple carbon isotopemodel for reconstructing prehistoric human diet. AmericanJournal of Physical Anthropology 133(4): 1112–1127.
Lee-Thorp J. 2008. On isotopes and old bones. Archaeometry50: 925–950.
Lillie MC. 2003. Tasting the forbidden fruit: Gender baseddietary differences among prehistoric hunter-gatherers ofEastern Europe Before Farming 2 (3): 1–16.
Lillie MC, Budd Ch., Potekhina I. 2011. Stable isotope anal-ysis of prehistoric populations from the cemeteries of theMiddle and Lower Dnieper Basin, Ukraine. Journal of Ar-chaeological Science 38: 57–68.
Lubell D, Jackes M, Schwarcz H, Knyf M, Meiklejohn C.1994. The Mesolithic–Neolithic transition in Portugal:Isotopic and dental evidence of diet. Journal of Archaeologi-cal Science 21: 201–216.
Martínez RM. 2013. Cerdos, caprinos y náyades.Aproximación a la explotación ganadera y fluvial en elGuadalquivir entre el Neolítico y la Edad del Cobre(3500–2200 A.N.E.). Spal Universidad de Sevilla, Seville22: 29–46.
Minagawa M, Wada E. 1984. Stepwise enrichment of 15Nalong food chains: Further evidence and the relation be-tween δ15N and animal age. Geochimica et Cosmochimica Acta48: 1135–1140.
McClure SB, García O, Roca de Togores C, Culleton B,Kennett DJ. 2011. Osteological and paleodietary investi-gation of burials from Cova de la Pastora, Alicante, Spain.Journal of Archaeological Science 38: 420–428.
Morgado A, Lozano JA, Pelegrin J. 2011. Las explotacionesprehistóricas del sílex de la formación Milanos (Granada,España). Menga; Journal of Andalusian Prehistory 2: 135–156.
Murillo-Barroso M, Martinón-Torres M. 2012. Ambersources and trade in the prehistory of the Iberian Penin-sula. European Journal of Archaeology 15 (2): 1–30.
Nocete F, Sáez R, Rodríguez M, Péramo A, Inacio N, AbrilD. 2011. Direct chronometry (14C AMS) of the earliest
copper metallurgy in the Guadalquivir Basin (Spain) dur-ing the third millennium BC: First regional database. Jour-nal of Archaeological Science 38 (12): 3278–3329.3.
O’Connell TC, Kneale CJ, Tasevska N, Kuhnle GGC. 2012.The diet–body offset in human nitrogen isotopic values: Acontrolled dietary study. American Journal of Physical Anthro-pology 149: 426–434.
Odriozola CP, Linares JA, Hurtado V. 2010. Variscitesource and source analysis: Testing assumptions at PicoCenteno (Encinasola, Spain). Journal of Archaeological Science37: 3146–3157.
Pajuelo A, López P. 2013. Estudio arqueozoológico deestructuras significativas de c/Mariana de Pineda s/n(Valencina de la Concepción, Seville). In ValencinaPrehistórica. Actas del Congreso Conmemorativo del Descubrimientode La Pastora (1860–2010), L García, V Hurtado, JM Vargas,T Ruiz, R Cruz-Auñón, (ed.) Universidad de Sevilla: Se-ville; 454–448.
Pecero JC, Guijo JM, Lacalle R. 2012. CaracterizaciónBioarqueológica de los Restos Óseos de Montelirio.Memoria Antropológica Rovira N. 2007. Agricultura ygestión de los recursos vegetales en el sureste de laPenínsula Ibérica durante la prehistoria reciente. BarcelonaPhD Thesis. Universitat Pompeu Fabra.
Richards MP, Hedges REM. 1999. Stable isotope evidencefor similarities in the types of marine foods used by LateMesolithic humans at sites along the Atlantic coast ofEurope. Journal of Archaeological Science 26: 717–722.
Richards MP. 2002. A brief review of the archaeological ev-idence for Palaeolithic and Neolithic subsistence. EuropeanJournal of Clinical Nutrition 56: 1270–1278.
Richards MP, Jacobi R, Cook J, Pettitt PB, Stringer CB.2005. Isotope evidence for the intensive use of marinefoods by Late Upper Palaeolithic humans. Journal of HumanEvolution 49: 390–394.
Rovira N. 2007. Agricultura y gestión de los recursosvegetales del sureste de la Península Ibérica durante laprehistoria reciente. PhD dissertation, Universitat PompeuFabra, Barcelona.
Salazar-García DC. 2011. Patrón de dieta en la poblaciónpúnica de Can Marines (Ibiza) a través del anàlisis deisótopos estables (C y N) en colágeno óseo. Saguntum43: 95–102.
Schoeninger MJ, De Niro MJ. 1984. Nitrogen and carbonisotopic composition of bone collagen from marine and ter-restrial animals. Geochimica et Cosmochimca Acta 48: 625–639.
Schuhmacher TX, Banerjee A, Dinjorf W, Nocete Calco F,Vargas Jiménez JM. 2012. Los marfiles del yacimientode Valencina de la Concepción en el contexto delCalcolítico del Suroeste peninsular. In ValencinaPrehistórica. Actas del Congreso Conmemorativo del 150Aniversario del Descubrimiento de La Pastora (Sevilla), L García,JM Vargas, V Hurtado, R Cruz, (ed.). Universidad deSevilla: Seville.
Schwarcz, HP, Schoeninger, MJ. 1991. Stable isotope anal-yses in human nutritional ecology. Yearbook of Physical An-thropology 34: 283–321.
M. Fontanals-Coll et al.
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)

Silva AM, Leandro I, Pereira D, Valera A. 2012. Os ossoshumanos cremados da fossa 16 do recinto dos Perdigões:a quem pertenciam? Actas do VI Encontro de Arqueologiado Sudoeste Peninsular, Villafranca de los Barros, Portugal(in press)
Stevenson, A. C., R. J. Harrison. 1992. Ancient forests inSpain: A model for land-use and dry forest managementin south-west Spain from 4000 BC to 1900 AD. Proceedingsof the Prehistoric Society 58: 227–247.
Van Klinken GJ, Van der Plicht H, Hedges REM. 1994.Bone C-13/C-12 ratios reflect (Paleo)climatic variations.Geophysical Research Letters 21: 445–448.
Van Klinken GJ. 1999. Bone collagen quality indicators forpalaeodietary and radiocarbon measurements. Journal ofArchaeological Science 26: 687–695.
Vargas JM. 2004. Carta Arqueológica municipal de Valencina de laConcepción, Junta de Andalucía, Seville.
Vargas Jiménez, J.M. 2013. Indicadores arqueológicos parala planificación y gestión del yacimiento de Valencina.In Valencina Prehistórica. Actas del Congreso Conmemorativo delDescubrimiento de La Pastora (1860–2010), L García Sanjuán,V Hurtado Pérez, JM Vargas Jiménez, T Ruiz Moreno,R Cruz-Auñón Briones (eds.). Universidad de Sevilla:Sevilla; 535–558.
Waterman AJ. 2012. Marked in life and death: identifyingbiological markers of social differentiation in late prehis-toric Portugal. PhD Doctoral Dissertation, University of Iowa.http://ir.iowa.edu/etd/3007
Waterman AJ, Silva AM, Tykot RH. 2014. Stable isotopicindicators of diet from two Late Prehistoric burials sites inPortugal: an investigation of dietary evidence of social differ-entiation. In RH Tykot (ed.) Proceedings of the 38th Interna-tional Symposium on Archaeometry—May 10–14 2010,Tampa, Florida. Open Journal of Archaeometry 2: 22–27.
A Palaeodietary Study of a Copper Age Burial, Valencina, Spain
Copyright © 2015 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2015)





























