Embed Size (px)
Citation preview

A RECONSIDERATION OF THE OMO KIBISH REMAINS
AND THE ERECTUS-SAPIENS TRANSITION
par
Michael H. DAY and Chris B. STRINGER
Abstract : The Omo-Kibish remains have been restudied in
the light of new finds including those from Arago.
The dating of the Kibish remains is reviewed and a
new reconstruction of Omo 1 is presented.
The affinities of the Omo-Kibish crania have
been reassessed making use of the newer principles
of classification that have emerged in the last ten
years. Working definitions of Homo erectus and Homo
sapiens are offered, based on distinctive within
group characters that can be identified objectively.
These definitions are employed to assess the affini-
ties of the Omo 1 and 2 crania in terms of a scoring
system.
The conclusion reached is that the Omo 1 cranium
can .be clearly aligned with anatomically modern Rome
sapiens while Omo 2 is closer to Homo erectus than to
the Neanderthals or to anatomically modern Homo sapiens
on the characters considered.
The dating of the two specimens remains a problem
in terms of absolute age and contemporaneity.
Comparison with Arago 21 shows that Omo 2 has
more in common with this specimen than Omo 1, although
Arago 21 and Omo 2 do not appear to be closely related.
Their similarities are mainly symplesiomorphies retained
from earlier Middle Pleistocene ancestors rather than
synamorphies.
St. Thomas's Hospital Medical Scholl - Lambeth Palace Road -London SEl 7EH, Grande-Bretagne.

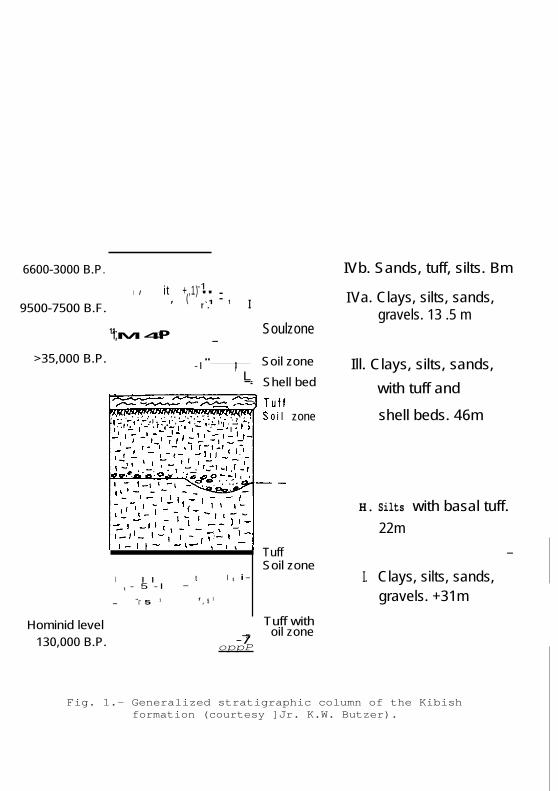
6600-3000 B.P.
9500-7500 B.F.
>35,000 B.P.
— I I — it — +,1)-1 — ": , ( r `.1 1 I
1,11-M4 '1CP _
" -I p L.
IVb. Sands, tuff, silts. Bm
I Va. Clays, silts, sands, gravels. 13 .5 m
Soulzone — — — — Soil zone Ill. Clays, silts, sands, Shell bed with tuff and
zone shell beds. 46m
H. Silts with basal tuff. 22m
Tuff — - Soil zone
I — I I t I t i- t - 5 -I -
- "-I 5 1 1 , 5 1 — Hominid level • Tuff with
130,000 B.P. -7 s oil zone — oppP
Fig. 1.- Generalized stratigraphic column of the Kibish formation (courtesy ]Jr. K.W. Butzer).
I. Clays, silts, sands, gravels. +31m

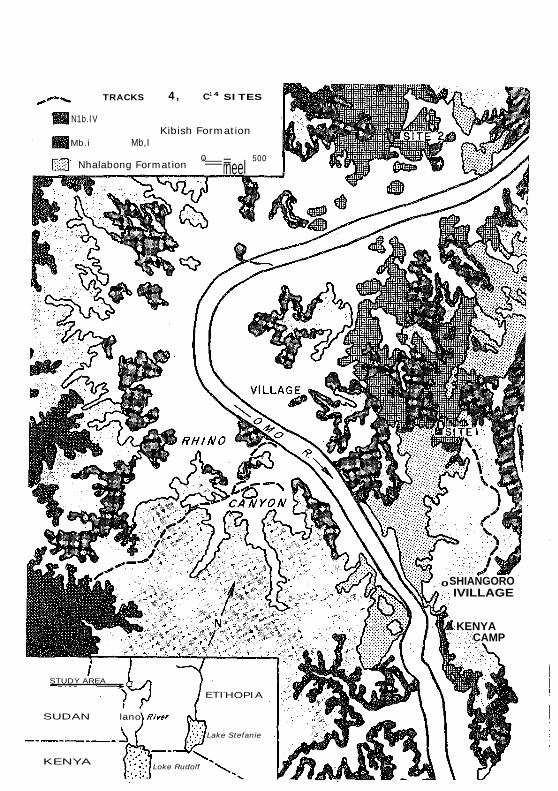
TRACKS 4, C1 4 SITES
N1b.IV Kibish Formation
Mb.i Mb,I
Nhalabong Formation 0 500 '=r=neel
0SHIANGORO IVILLAGE
KENYA CAMP
STUDY AREA
ETI-HOPIA
SUDAN lano
Lake Stefanie
KENYA Loke Rudolf

The only archaeological evidence is a few stone tools
found with Omo I and they are undiagnostic of a re-
cognised culture.
The Omo 2 calvaria which was discovered approxi-
mately 2.5 km from the site of Omo 1 ("Fig. 2) is less
certainly dated since it was essentially a surface
find with no clear stratigraphic, faunal or cultural
context although the sedimentary sequence is reported
to be the same on both sides of the Omo river in this
area.
The Omo 3 fragments are thought to come from
Member III of the Kibish formation and are thus
slightly higher in the sequence, but still older than
37,000 years b.p. according to radiocarbon determina-
tions on molluscs from Member III (Butter, 1976).
Some of the initial conclusions concerning the
dating of the Omo material remain essentially un-
changed in that:-
a) Omo 1 is most clearly asasociated with
Member T in that it was/partly in situ
find that was properly excavated.
b) The contemporaneity of Omo 1 and the
associated faunal remains seems secure.
c) The uncertainty concerning the precise
stratigraphic level of the Omo 2 remains
leaves some doubt as to its contempor-
aneity with Omo 1. Omo 3 certainly
appears to be younger than Omo 1.
d) The chronological date of 130,000 years
b.p. for Member I by the Uranium/Thorium
method remains unconfirmed by any other
technique, but it is not contradicted by
the limited available faunal and radio
carbon dating,
010

Since the initial excavation, the site has not
been revisited and no further evidence as to its
dating has been published.
In terms of dating, therefore, several questions
can be posed. Is the relative dating evidence strong
enough to demand that the material be treated as a
single population sample? If the answer is yes, re-
construction and morphological, analysis can operate
within the bounds of variability of a local popula-
tion of a single taxon. If the answer is no, then
additional considerations arise in that the remains
may represent samples of time - successive populations
that are closely related to each other or they may
represent evidence of population replacement rather
than local in situ evolution.
• a. Previous work on the Omo-Kibish remains
The initial work on these remains included their
cleaning, restoration and description. The initial
assessment of the remains was based on anatomical
evidence and concluded that while all three specimens
should be attributed to Homo sapiens, specimens 1 and
2 show striking differences in skull form. The more
complete calvaria having features in common with the
Solo skulls, the Broken Hill skull and with Homo
erectus (Day, 1969). Later work including a discrim-
inant function analysis of metrical skull data
emphasised the differences between Omo 1 and Omo 2
and that both could be distinguished from at least
three populations of modern Homo sapiens (pay, 1973).
The Omo specimens were regarded as Neaderthals
(in the grade sense) by Brose & Wolpoff C1971), who
stated that no features of the specimens were outside
the range of Neanderthal morphology. Omo I "would
be identified as anatomically modern if found in
another context", but at the same time they made the

remarkable claim that the specimen was "extremely
similar to Amud". Overall, the Omo crania were
said to be Neanderthals (Sensu late), transitional
to "modern" H. sapiens, Howells (1974) strongly
disagreed with Brose & Wolpoff's (1971) general
approach and specifically criticised their conclu-
sions about the Omo specimens. We fully endorse
his criticisms.
Rightmire (Rightmire 1976; 1980; 1981)
favoured treating the specimens together as related
forms, while admitting that Omo 2 was more like
Broken Hill 1. As we also believe, Rightmire 01981)
felt that the unpublished Guomde specimen (KIN-ER
3884) from East Turkana might be related to the
Omo-Kibish fossils.
Stringer utilised multivariate analysis of
cranial measurements (1974) and analyses of size,
shape, angles and indices (1978) to compare the
Omo-Kibish specimens. The results tended to accen-
tuate the contrasts between the Omo crania, always
aligning Omo 1 much more closely to anatomically
modern (a.m.) H. sapiens. The multivariate analy-
ses (Stringer, 1974) were based on measurements of
Omo 2, and Omo 1 as reconstructed in.1972...,,.However
the new reconstruction presented here is only
marginally different in frontal dimensions and
cranial length from that of 1972, while parietal
and occipital regions have been retained. Thus the
1974 multivariate results comparing Omo 1 and 2 are
still relevant to the present discussion. The
analysis which employed most measurements for the
two specimens together utilised the following 13
measurements.
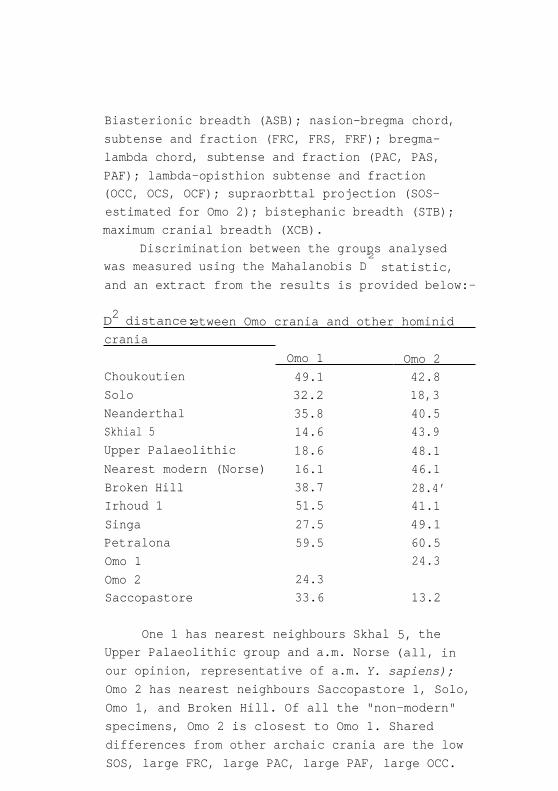
Biasterionic breadth (ASB); nasion-bregma chord,
subtense and fraction (FRC, FRS, FRF); bregma-
lambda chord, subtense and fraction (PAC, PAS,
PAF); lambda-opisthion subtense and fraction
(OCC, OCS, OCF); supraorbttal projection (SOS-
estimated for Omo 2); bistephanic breadth (STB);
maximum cranial breadth (XCB).
Discrimination between the groups analysed
was measured using the Mahalanobis D2
statistic,
and an extract from the results is provided below:-
2 D distance: etween Omo crania and other hominid
crania
Omo 1 Omo 2 Choukoutien 49.1 42.8
Solo 32.2 18,3
Neanderthal 35.8 40.5
Skhial 5 14.6 43.9
Upper Palaeolithic 18.6 48.1
Nearest modern (Norse) 16.1 46.1
Broken Hill 38.7 28.4' Irhoud 1 51.5 41.1
Singa 27.5 49.1
Petralona 59.5 60.5
Omo 1 24.3
Omo 2 24.3
Saccopastore 33.6 13.2
One 1 has nearest neighbours Skhal 5, the
Upper Palaeolithic group and a.m. Norse (all, in
our opinion, representative of a.m. Y. sapiens);
Omo 2 has nearest neighbours Saccopastore 1, Solo,
Omo 1, and Broken Hill. Of all the "non-modern"
specimens, Omo 2 is closest to Omo 1. Shared
differences from other archaic crania are the low
SOS, large FRC, large PAC, large PAF, large OCC.
.7"
Omo 2 calvaria

Shared differences compared with a.m. H. sapiens crania include larger XCB, STB, ASB, FRC, FRF and
PAF. Differences between Omo 1 and 2 mainly re-
late to contrasts in FRS (Omo 1 higher value -
more domed frontal), PAS (Omo 1 higher value -
more domed parietal), OCS (Omo 2 higher value -
more projecting occiput), OCF (Omo 1 higher value
- expanded occipital plane) and ASB (Omo 2 higher
value - broader occipital base), and in every one
of these differences Omo 1 is closer to the pattern
of a.m. H. sapiens crania.
Thus the multivariate results confirmed that
Omo 1 approximated to an anatomically modern
pattern in these cranial measurements, while Omo
2 did not. The two crania do share, however,
certain cranial characteristics suggesting that
they are related, though temporally separated,
members of an evolutionary lineage in north-east
Africa leading from erectus-like hominids to a
robust, but anatomically modern form. Alterna-
tively, if we accept the published dating, the .
Omo 1 cranium could represent a late Middle
Pleistocene/Upper Pleistocene spread of a.m.
R. sapiens into the area accompanied by some
degree of gene flow from a more archaic local
population represented by Omo 2.
Clark Howell (1978) was sceptical of the
Middle Pleistocene age estimates for the Omo-
Kibish crania, and questioned Day's (1973) con-
clusions regarding their affinities. He emphasized
their distinctiveness from R. erectus and H.
sapiens rhodesienses and concluded that they repre-
sented a late subspecies of H. sapiens closely
related to modern man. He quoted Stringer's (1974)
conclusions in support of this view, but these
results do not justify Howell's interpretation,
since Omo 2 is not closely related to recent H.
sapiens but is similar to various archaic specimens.
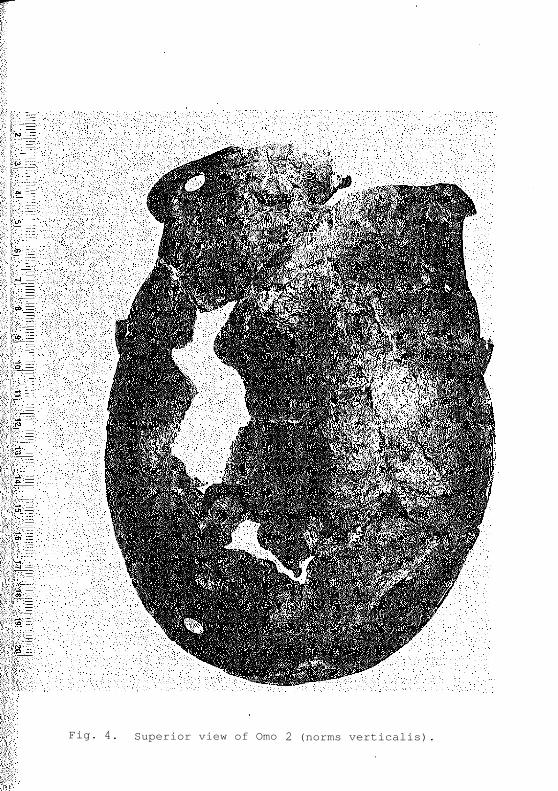
Fig. 4.— Superior view of Omo 2 (norms verticalis).

3. The Reconstruction of the Omo I cranium
The problems of the Kibish remains can also be
approached from the viewpoint of anatomical recon-
struction, governed by the internal evidence of
anatomical necessity rather than reconstruction by
the amalgamation of several species (vide Arago).
The reconstruction of undistorted cranial frag-
ments can proceed with confidence only if the frag-
ments concerned are adjacent and capable of a demon-
strable interlocking fit. Once those pieces that
have such a fit are identified and joined other
factors such as bilateral symmetry of skulls can aid
reasonable reconstruction.
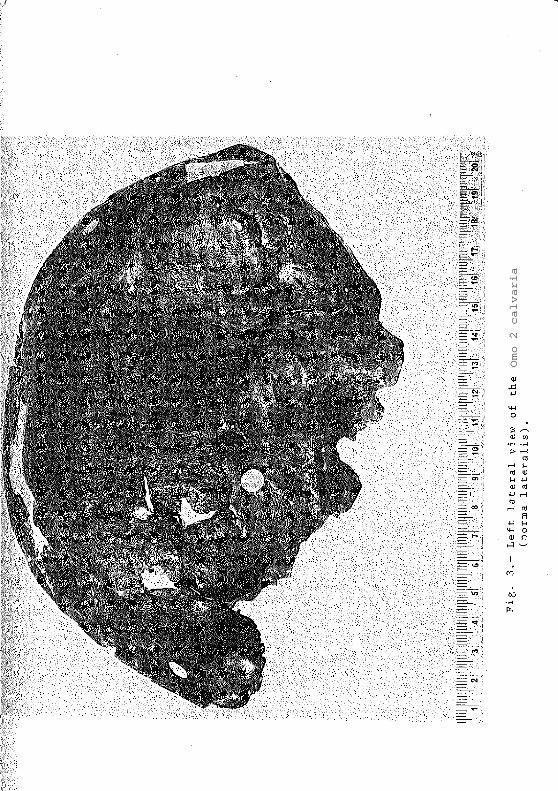
In the case of the Omo 2 calvaria (Figs. 3 & 4)
the four fragments fit perfectly and the specimen is
undistorted. Any attempt to reconstruct the face
would be purely speculative and of little or -no
scientific value. Similarly with Omo 3 the paucity
of fragments (one left fronto-parietal fragment and
a glabellar fragment) does not allow any reasonable
attempt at reconstruction to be made. It is the
purpose of this paper to publish a reconstruction of
Omo 1 and to discuss the problems encountered and to
compare the Omo I reconstruction with the other Omo
(Kibish) remains and with other specimens to compar-
able age from Africa and from Europe. This, in turn,
may throw some light on the problems of the ex,eetus -
sapiens transition and contribute to the current
debate on punctuated equilibrium as opposed to gradu-
alism in this period of human evolution.
Omo 1 has a posterior vault which can be recon-
structed with some confidence, giving a decidedly .
"modern" appearance. To what extent should the re-
construction of the anterior region reflect this?
We believe the robust but anatomically modern morpho-
logy of the posterior vault should be mirrored by an
appropriate reconstruction of the frontal, face and
mandible. Using early anatomically modern specimens
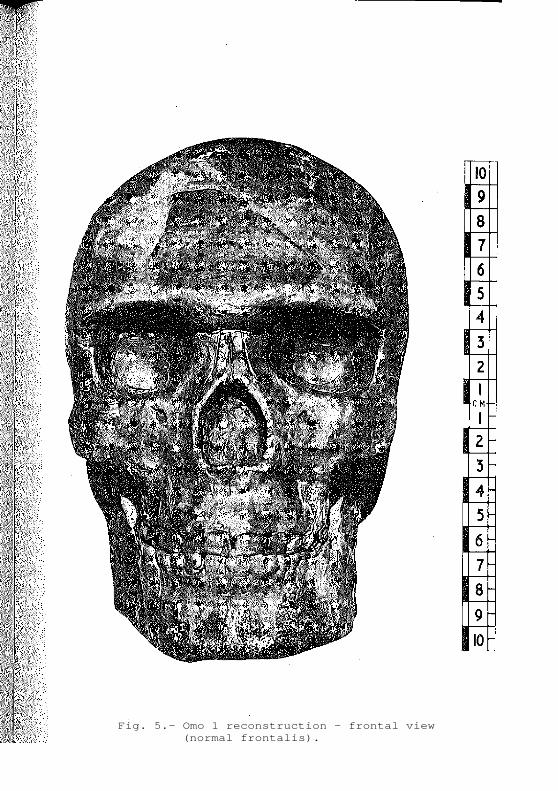
Fig. 5.- Omo 1 reconstruction - frontal view (normal frontalis).

from S.W. Asia (Skinal, Qafzeh) and more archaic specimens from Africa (Irhoud 1, Ngaloba L.H.18, Broken Hill
1) it is possible to reconstruct a face which is
"modern" but which nevertheless reflects the robust
morphology of other late Middlejearly Upper Pleisto-
cene hominids by displaying a wide upper face, large
interorbital breadth and strong alveolar prognathism
(Fig. 5).
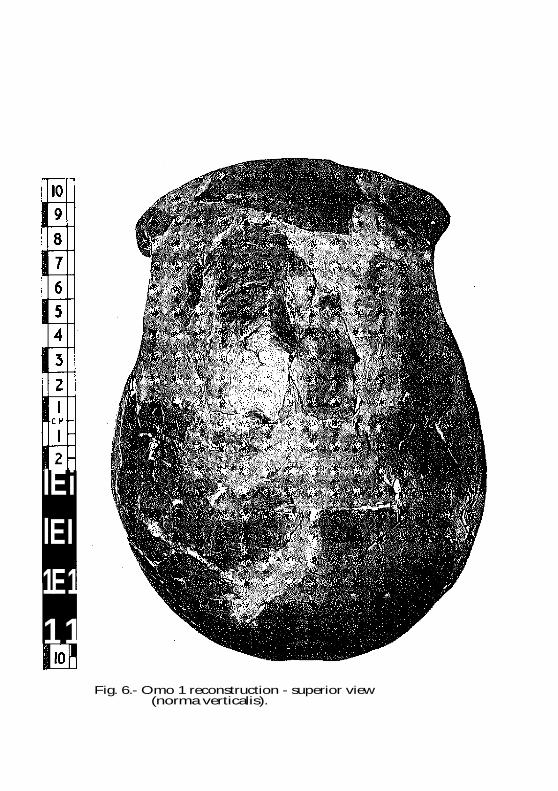
The frontal bone is very long, but some flexi-
bility is possible in the degree of flatness chosen
for the reconstruction (Fig. 6). The mandible has a
fairly narrow ascending ramus which would normally
indicate a short body and overall length. However,
given the constraints of the long frontal bone and
our preference for a face with some degree of alve-
olar prognathism, rather than an .orthognathic face,
the mandible has been reconstructed with a long body,
while avoiding the presence of retromolar spaces, not
otherwise found in African Pleistocene hominids.
4. New analyses and the classification of the Omo-
Kibish remains
Since the original studies of the Omo fossils,
much new material has been discovered and described
which is relevant to our interpretation of these
specimens. The characteristics of African H. erectus
are now better known, thanks to recent discoveries
and descriptions (Rightmire, 1980), while material
which throws light on the possible transition from
H. erectus and H. sapiens has been described from
sites such as Petralona, Arago and Bilzingsleben in
Europe (see e.g. Stringer et aZ., 1979) and Ndutu and
Bodo in Africa (Rightmire, 1980; 1981). Archaic
hominid material from Broken Hill and possible early
anatomically modern material from Florisbad may be
more ancient than had been supposed, probably dating
lEi lEI 1E1 1 1
I
Fig. 6.- Omo 1 reconstruction - superior view (norma verticalis).

from the Middle Pleistocene and early Upper Pleis-
tocene respectively (Rightmire, 1981), while other
African material which may document the latter
stages of hominid evolution has been described from
sites such as Djebel Trhoud (Howell, 1978) and Ngaloba (Day et al., 1980), From Border Cave and Klasies
Cave in Southern Africa, material which may represent
anatomically modern hominids from the early Upper
Pleistocene has been described CRightmire, 1981).
From all this additional knowledge we should now
be in a better position to assess the affinities of
the Omo-Kibish crania than was possible ten years ago,
but the intervening years have also seen reassessments
of the principles to be used in classifying fossil
hominids and a growing recognition of the problems we
face in interpreting a fragmented fossil record with
poor chronological control, against a background of
debate about the fundamental mechanisms of evolution-
ary change. Following some common palaeoanthropologi-
cal methods of classification we might use the "total
morphological pattern" approach of Le Gros Clark
(1964) or employ a multivariate approach (e.g. Stringer,
1974) to produce measures of overall similarity between
fossil hominids and recent populations. But such
approaches are primarily phenetic, often leading to the
creation of grade systems, disguised by the use of
subspecies names (Campbell, 1964; Howell, 1978) or
overt in the application of numbered grades (Stringer,
et al., 1979). When one of us suggested a grade system
for H. sapiens, it was because it was felt that the
application of subspecies names to fossils was hinder-
ing rather than helping the progress of palaeoanthro-
pology.
A numbered grade system should only be employed
pending the creation of valid taxonomic units, and

could not be a workable solution in the long term.
As a first step towards a different approach to
hominid classification we would like to propose
simple working definitions of H. erectus and H.
sapiens based on distinctive within-group character-
istics which can be used to assess the affinities of
the Omo-Kibish fossils. Most of the characters have
previously been used in describing the two species
but we have attempted to provide clear i presence or
absence data which can be assessed objectively and
applied simply. Some of the metrical differences
recognised can be shown to lie behind multivariate
discrimination (see Stringer, 1974; 1978 and earlier
discussion of the Omo multivariate results), although
this is sometimes masked by amalgamation of size and
shape differences. Once broad groupings CH. erectus,
Neanderthals and a.m. H. sapiens) are established, it
should be possible to recognise further subdivisions
and identify other groupings for fossils which do not
fit well into any of these categories. More detailed
definitions based on more anatomical characters,
including those most generally used (e.g. see Le Gros
Clark, 1964; Howell, 1978;Howells„1980) would be de-
sirable, but without clear character weighting it is
difficult to classify fossil hominids which show only
some of these features, or which have characters only
partly developed.
It seems appropriate to begin with a definition
of a.m. E. sapiens based on derived characters which
can be assessed in the Omo specimens. There is a
large body of data available (Howells, 1973). Indeed
if we cannot provide a valid definition of the only
living human species there is little hope of our re-
cognising extinct taxa. Whether the species so recog-
nised should also form the basis for recognising the
species in the fossil record is a difficult problem,
but it does seem somewhat inappropriate to include in

the species fossil hominids which lack most of the
derived characters defining H. sapiens today. But
for the present we will equate our definition of H. sapiens (Sensu stricto) with the term a.m. H. sapiens,
and leave the question of whether various archaic
forms rightly belong within the species H. sapiens
to future research. The criteria used here to de-
fine a.m. H. sapiens will not all be present in all
modern specimens, but a majority should always be
present. Because of population variability, some
of these characters will also be found in fossil
hominids which are clearly not anatomically modern
forms. But if either of the Omo-Kibish specimens
can reasonably be classified as a.m. H. sapiens, we
should expect over 50% of the "modern" characters
to be present of those which can be tested.
Working definition of anatomically modern H. sapiens:
- Cranium short but high (ratio of basion-bregma
height or vertex radius to glabello-occipital length
should exceed 0.70 or 0.64 respectively);
Parietal arch is long and high, inferiorly
narrow, superiorly broad (parietal angle less than
1380, ratio of bregma-asterion chord to biasterionic
breadth greater than 1.19);
- Frontal bone high (frontal angle less than 133°);
- Supraorbital torus not continuous, but divided
into lateral and medial portions;
- Occipital bone well curved rather than angulated
(occipital angle more than 114°);
- Mental eminence on mandible;
- Gracile limb bones - thin walled with relatively
small articular surfaces.
For Homo erectus
Fossil material from Africa and Asia can simi-
larly be used to produce a working definition of H.

erectus. Material used here to formulate the mor-
phological and metrical definition includes KNM-ER
3733 and 3883; Choukoutien; Sangiran 2,4,17; and
Ngandong. The characters chosen exclude some of
the useful traditional characters (e.g. thick occi-
pital torus, presence of angular torus, presence of
mid-sagittal keeling and parasagittal depressions)
because these cannot be defined precisely or object-
ively. Such characters should also be assessed in
any overall study of the Omo-Kibish material and
have previously been investigated (e.g. Day, 1969;
1971). The characters chosen for Table 1 will all
be present in some H. eretus crania, and the majority should be present in all H. erectus crania. Other fossils will display some H. erectus characters even though they are not members of the species R. erectus, since our working definition is primarily created to separate H. erectus most effectively from anatomically modern H. sapiens. Other characters would
have to be utilised if we were trying to separate
Neanderthals from H. erectus, since they share some
of the characters which reflect the presence of a
long, low skull, low frontal and short flat parietal
arch, and this will have to be investigated in the
future. But Neanderthal crania do not show a
majority of the H. erectus characters, and a set of criteria differentiating Neanderthals from a.m. H. sapiens can also be created to test whether either
of the Omo-Kibish crania could be regarded as
"Neanderthal" gable 3). If specimens are found
that display combinations of characters typical of
the various groups (or intermediate values for cer-tain characters) this may indicate a) that they
represent genuine morphological intermediates between,
the forms, b) that they are hybrids if it can be
demonstrated that the morphologically distinct groups

overlap in time, or c) that the characters used in
the working definitions are inadequate and need
modification.
Working definition of 11. erectus:
- Cranium long and low (ratio of basion-bregma
height or vertex radius height to glabello-occipital
length is less than 0.62 or 0.58 respectively);
- Inion is coincident with opisthocranion;
- Thick cranial walls (for example parietal thick-
ness at bregma exceeds 8 mm, at asterion exceeds
14 mm, on occipital at inion exceeds 14 mm);
- Angulated occipital bone (occipital angle less
than 107°);
- Expanded nuchal portion of occipital bone
Cinion-opisthion chord exceeds lambda-inion chord);
- Inion more than 25 mm above internal occipital
protuberance);
- Parietal arch flat, short and low, superiorly
narrow, inferiorly broad (parietal angle exceeds
145.5°, ratio of bregma-asterion chord to biasterionic
breadth is less than 1.14);
Low frontal bone (frontal angle exceeds 136.5°);
- Cranial capacity generally below 1150 ml.
5. Conclusions: the affinities of the Omo-Kibish
fossils
In Tables 1,2 and 3 it is evident from the
characters considered that Omo 1 and 2 differ mark-
edly in their affinities, far more than would be
expected for one population. Omo 2 shows most resem-
blance to H. erectus, the main differences being related to larger cranial capacity and associated
vertical expansion of the cranial vault reflected by
a large vertex radius and bregma-asterion chord.

The occipital bone shows various typical erectus characters but it is actually less angulated than the
majority of other erectus crania. Omo 1 is clearly aligned with a.m. H. sapiens although the low and long
frontal bone of the reconstructed cranium is exceptional
and the robusticity of the specimen is indicated by
relatively thick cranial walls. It would be premature
to assign the Omo 2 fossil to the species H. erectus when some important anatomical areas are not represented
Ce.g. face, dentition, mandible, post-cranium) and there
are clear departures in supraorbital torus morphology,
cranial capacity and overall dimensions from the typical
and fairly consistent sample of H. erectus crania from
Africa and Asia. However the affinities of the specimen
are certainly closer to H. erectus than to the Neander-
thals or a.m. H. sapiens on the characters assessed here.
For Omo 1 we feel we have sufficient evidence from the
cranial parts, mandible and post-cranium to align the
specimen positively with a.m. H. sapiens, despite its
robusticity.
The main problems remaining are chronological. If
both specimens are of late Middle/early Upper Pleisto-.
cene age, then two distinct but approximately contem-
poraneous populations are being sampled. One of them
conformed to an anatomically modern pattern and would
represent one of the most ancient if not the most
ancient of such groups known. But a note of caution
must be injected at this point. Further confirmatory
absolute dating analyses have not been performed,
neither has subsequent field work been conducted in the
area to recover further material, including faunal re-
mains which might be more diagnostic of a Middle or
Upper Pleistocene age. Given the evidence from South-
ern Africa (Rightmire, 1981) it is no longer implausible
that anatomically modern hominids existed at the
beginning of the Upper Pleistocene. What would be

difficult to explain however is the occurrence of
such widely separated early anatomically modern
hominids when more archaic populations may still
have existed in North Africa and in the intervening
area of East Africa. Perhaps the dating of hominids
in East Africa such as Eyasi, Broken Hill and
Ngaloba may be significantly earlier than the more
"modern" specimens, or there may have been a rapid
population replacement in the early Upper Pleisto-
cene. However this cannot be demonstrated within
our present inadequate dating framework.
Alternatively the evolution of hominids resem-
bling anatomically modern man may have occurred more
or less independently in areas of Southern, Eastern
and Northern Africa, in which case the Omo-Kibish
remains may document a transition from an ercctus-like Cnot Neanderthal) ancestor to a robust but,
"modern" skeletal form. Whichever model eventually
proves most appropriate it is evident that relation-
ships between the contrasting Omo-Kibish specimens
will remain somewhat problematical until further
evidence is available about their relative chrono-
logical positions.
6. The Omo remains and the Arago 21 construction
One of the purposes of this symposium is to
discuss the new reconstruction of the Arago 21
specimen, and through comparative analyses, to place
the specimen in relation to various other fossil
hominids. Our reconstruction of the Gmo 1 cranium
was made 'using the available fragments, directly
where possible, or indirectly as indicators of the
probable morphology of missing parts. We did not use fragments of other fossil hominids in our re-
construction, but rather attempted to'produce a
reasonable reconstruction given the available frag-
ments and the morphology of other late Middlelearly
Upper Pleistocene crania. For the Arago reconstruc-

tion, modified parts of other fossils have been used
to create a composite cranium containing fragments
from specimens such as Sangiran 17, Swanscombe or
Vertesszbllbs to represent the missing parts of the
skull.
Direct comparisons between the Arago cranium
and the Omo fossils are difficult since the former
specimen consists primarily of parts not well repre-
sented in the latter (upper dentition, palate, face,
supraorbital region), and some of the most informa-
tive parts of the Omo crania (mastoid region occipital
bone, mandible and postcranium of Omo 1) are not
present in the Arago 21 cranium itself. Comparisons
can be made for the frontal bone and parietal region
and except for the common feature of a long and flat
frontal bone (accepting the Omo 1 reconstruction),
there are immediate contrasts between the specimens.
The Arago frontal certainly appears more archaic in
its massive supraorbital torus and narrow dimensions,
with a significant degree of postorbital constriction.
The latter character is no doubt related to the lower
cranial capacity of the Arago specimen.
In parietal morphology there are also contrasts
since Omo 1 displays a much more modern form than
Arago 47 (as reconstructed) while Omo 2 appears rather
archaic in comparison, with a midsagittal keel, para-
sagittal flattenings and a broad base to the parietal
arch. But some of the most important diagnostic
characters which separate the Omo crania are in the
occipital bone, which is unfortunately lacking in the
Arago specimen. Given the morphology of the face and
frontal bone, a more archaic occipital than that of
the Swanscombe specimen was probably present and there-
fore the VertesszbllUs, Petralona or Bilzingsleben
occipitals may be more appropriate models for the

Arago reconstruction. It is true that the Arago 47
parietal may appear rather "advanced" in certain
respects, but the arch has been reconstructed with
a low biasterionic breadth in comparison with all
other European Middle Pleistocene hominids except the
Steinheim cranium. In addition, the parietal form
of specimens such as Petralona, Biache, Steinheim and
Ehringsdorf 9 is very "progressive" in certain res-
pects, and this characteristic seems to be a signifi-
cant derived character for European Middle Pleisto-
cene hominids generally compared with H. erectus
fossils. It is not always matched by the morphology
of the occipital bone (vide Petralona). The strong
angular torus of the Arago 47 parietal compared with
the Petralona specimen may also be a clue to the
robusticity to be expected in the occipital bone.
Thus the Arago occipital bone would probably have
been much more archaic than that of Omo 1, perhaps
resembling that of Omo 2 more closely. The temporal
bone of European Middle Pleistocene hominids (such as
Petralona, Steinheim and perhaps Ehringsdorf 9) also
shows distinctive, more modern characters compared
with those of Asian H. erectus fossils, and therefore
the Arago temporal bones may have resembled those of
European hominids (or Broken Hill 1) rather than
Sangiran 17.
As we have already discussed, we believe that
Omo 1 must be classified as an a.m. H. sapiens, For
Omo 2 and Arago 21, the choice is not so easy. Under
the generally accepted system for hominid classifica-
tion, the choice of taxon would lie between H. erectus
and some form of "archaic" H. sapiens. In differen-
tiating between E. erectus and H. sapiens (sensu Zeta)
the ultimate classification depends entirely on which
characters are considered most significant. If endo-
cranial volume and supraorbital torus morphology are

considered important, Arago 21 would probably be
classified as H. erectus and Omo 2 as H. sapiens.
If occipital morphology is emphasised, Omo 2 would
probably be classified as H. erectus while Arago 21
could not even be assessed. In details of parietal
form, the two specimens show different combinations of erectus-like and sapiens-like characters, but
share a flat parietal bone, typical of erectus,
with a parietal arch which is "advanced" in being
higher, diagonally longer and (as reconstructed for
Arago) relatively narrow inferiorly. If we define
H. erectus only by the shared derived characters of
the Asian and early African specimens then Arago 21
and Omo 2 both show significant, but distinct de-
partures from this condition. If however we adopt
a broader definition of H. erectus to include fossils
often regarded as "archaic" H. sapiens (e.g. Broken
Hill, Petralona) then both could be included in such
a group. Yet another alternative is to recognise a
third group for such specimens which display mosaic
or intermediate characters. We have both suggested
names for such a group in the past. Day (1973) pro-
posed that they be regarded as a "Pithecanthropoid
Intermediate" group, while Stringer et ai.,C1979)
suggested the term "H. sapiens grade I". Either of
these terms are still preferred by us to the alloca-
tion of sub-specific names which are of dubious
applicability and usefulness.
Eventually it may be possible to recognise sub-
sets within this grouping of "intermediate" fossils
based on derived characters probably linking the
Arago fossils with other European Middle Pleistocene
specimens, and perhaps ultimately, with the Neander-
thals. But convincing synapomorphies with the
Neanderthals are difficult to detect in the European

hominids until'the later Middle Pleistocene and the
search for derived characters linking together the
African fossils of this time is an even more diffi-
cult task. Ultimately the Arago and Omo 2 fossils
will probably not be classified together except in a
generalised grade framework. They do not appear to
be closely related specimens and their grade similar-
ities rest mainly on symplesiomorphies retained
from, earlier Middle Pleistocene ancestors rather than
on synapomorphies. But we are unable to propose a
definitive classification at the moment beyond suggest-
ing that neither belong in H. erectus sensu stricto
or H. sapiens sensu strict°.

REFERENCES
Brose, D.S. & Wolpoff, M.H. 1971. Early Upper Paleo-
lithic man and late Middle Paleolithic tools. American Anthropologist 73: 1156-1194.
Butzer, K.W. 1969. Geological Interpretation of Two Pleistocene Hominid ,Sites in the Lower Omo
Basin. Nature 222: 1133-1135.
Butzer, K.W. 1976. The Mursi, Nkalabong and Kibish
Formations, Lower Omo Basin, Ethiopia. In
Earliest Man and Environments in the Lake Rudolf
Basin Edited: Y. Coppens, F.C. Howell, G.L1.
Isaac & R.E.F. Leakey. Chicago: University of
Chicago Press. Pp. 12-23.
Butzer, K.W. & Thurber, D.L. 1969. Some Late Cenozoic
Sedimentary Formations of the Lower Omo Basin.
Nature 222: 1138-1143.
Butzer, K.W., Brown, F.H. & Thurber, D.L. 1969. Hori-
zontal sediments of the Lower Omo Valley: the
Kibish Formation. Quaternaria 11: 15-29.
Campbell, B.G. 1964. Quantitative taxonomy and human
evolution. In Classification and Human Evolution.
Edited: S.L. Washburn. Chicago: Aldine. Pp. 50-74.
Day, M.H. 1969. Omo Human Skeletal Remains. Nature 222: 1135-1138.
Day, M.H. 1972. The Omo human skeletal remains. In The origin of Homo sapiens. Edited: F. Bordes.
Paris: Unesco. Pp. 31-35.

Day, M.H. 1973. The Development of Homo sapiens. In
L'origine Dell 'Homo. (Darwin Centenary Symposium
on the origin of man). Roma: Accademia Nazionale
Del Lincei. Pp. 87-95.
Day, M.H., Leakey, M.D. & Magori, C. 1980. A new fossil
hominid skull CL.H.18) from the Ngaloba• Beds,
Laetoli, Northern Tanzania. Nature 284: 55-56
Howell, F.C. 1978. Hominidae. In Evolution of African
Mammals. Edited: V.J. Maglio & H.B.S. Cooke.
Cambridge: Harvard University Press. Pp. 154-248.
Howells, W.W. 1973. Cranial variation in Ban: a study
by multivariate analysis of patterns of difference
among recent human populations. Papers of the Pea-
body Museum. 67: 1-259 (whole volume).
P
Howells, W.W. 1974. Neanderthals: Names, Hypotheses and
Scientific Method. American Anthropologist 76: 24-38.
Howells, W.W. 1980. Homo erectus - Who When and Where: A
Survey. Yearbook of Physical Anthropology 23: 1-23.
Leakey, R.E.F. 1969. Early Homo sapiens remains from the
Omo River region of south-west Ethiopia: Faunal
Remains from the Omo Valley Nature 222: 1132-1133.
Le Gros Clark, W.E. 1964. The fossil evidence for Human
evidence. Chicago: University of Chicago Press.
Rightmire, G.P. 1976. Relationships of Middle and Upper
Pleistocene hominids from sub-Saharan Africa.
Nature 260: 238-240.

Rightmire, G.P. 1980. Homo erectus and human evolution
in the African Middle Pleistocene. In Current
Argument on Early Man. Edited: L.-K. Konigsson.
Oxford: Pergamon. Pp. 70-85.
ightmire, G.P. 1981. Later Pleistocene hominids of
eastern and southern Africa. Anthropologic Brno.
19: 15-26.
Stringer, C.B. 1974. Population relationships of later
Pleistocene hominids: a multivariate study of
available crania. Journal of Archaeological Science .
1: 317-342.
Stringer, C.B. 1978. Some problems in Middle and Upper
Pleistocene hominid relationships. In Recent
advances in Primatology. Vol. 3. Evolution.
Edited: D.J. Chivers & K.A. Joysey. London: Academic
Press. Pp. 395-418.
Stringer, C.B., Howell, F.C. & Melentis, J.K. 1979. The
Significance of the Fossil Hominid Skull from
Petralona, Greece. Journal of Archaeological Science
6: 235-253.
Stringer, C.B. & Trinkaus, E. 1981. The Shandar Neanderthal
Crania. In Aspects of Human Evolution. Edited: C.B.
Stringer. London: Taylor & Francis. Pp. 129-165.

TABLES 1 & 2:
and anatomi-
Lk a) O
a) a)
Ca •P U U
f-F O
F~ u al 0
Ca
• g aS a)
Ca to
.r1 0) 4 hD U O - .0 0 X 1:3 0. to iK 0 a
tr)
- 60 g
) to
0 a Ca
• • H
C) Ca 0) a) a)
F, _Ca In • -P
g.
cally modern H.
•
4- )
m
are expected to be
Characters
remains.
Cn 0
C)▪ .
same criteria.
Aa
a)
Fes-, La
Arago 21 reconstruction is also tested
erectus (characters
Ca 0
0)
1-1
O bn Ca
•c(
O a O
0
0
Ca 41
differentiate from a.m. H.
nIN
rl 'to O
Cd CO 0 tO.
I a) Z 0.3 1.f1
Ctl
ttdrrs
r•I Ca Cd g g
I bl) bly
cd id g
II. g " • . , n , ■ , 0
rH I •..0
03 Cd t'r)
g 0 , .1-1 v.-1 taD c-... to I:10 •
0 0 i•-■ 0 0 0 0 0 0 0 1-1 U 0 Z Z ca Z 7 Z Z Z Z cd '-' Z
a a
Thick cranium:
Inion = opisthocranion
VRR/GOL <0.58
0
Cranial capacity .11150 ml
BBH/GOL <0.62
tol. ,e-4■
4--I ctS cC
• g [o to 0 g
•
to to 0 u) 0 a) z a) 0 Z
bo • k
U Cd cC
0 a-J O -1- i-t ..--1
a p.0 • o - as cIS 4--, P. if) rI Ln 4-1 4-) 4,1 0 0 0) 4.1 04 E.-. r0 N at C%.1
Lf) r•-1 V '0
• .-1 ..-4 ,--i 0 I u A 4 , ..i- gal to
Cdk k u -.-1 .1-4 1--1 Cr) r-I ctl IS V Ca Ca 0 S=1., A < A
0-. P.. 0 0 0 .ri -..... < .1-i. •r-t .,-1 u ..t
,^-, ,-, U g 0 0 U < < g '-cd".6 4:1 U 0 I-I .."1 [-I 0 04 P:1 f.11
above internal
cm e—, N. to') 00 0

O
r--
0 Z
co v, c, z z
>-■
Mental eminence
weakly developed lateral and
Clearly divided suptaorbital
CO 1.1 d ul
U) Vi Ul
ca.
CO 0 a) a) 0 0
Ct$ 0 rl
0 0 131) F-4 cd
0 0 z
I Ln.
wN
0 0 -.1" ai
•
V% .1% 1-4 0C.O.
0 CD
$2:1 1:4
° cO
ri V
‹, 1;41
n 0
rs ‹G' v
1,1 13:1 s
mall articular surfaces
Limb bones thin-walled with relatively
Arago 21 recon.
O E 0
0
0
td • 'tj A
• '71
E. sapiens (characters of modern
to differentiate from H.
erectus
Neanderthals)).

Arago 21 recon.
The Arago 21 reconstruction is also tested.
described as "Neanderthal" rather than as related
Selected metrical and morphological characters whish distinguish between
(Stringer & Trinkaus,
Relevant Neanderthal characters
to H. erectus or a.m. H.
Neanderthal and a.m.
either Omo
9) Limb bones thick walled with relatively
O z
large articular
ao
00
* Several of the more diagnostic Neanderthal characters of the face and post-cranium
cannot be used because of the incomplete nature of the Omo-Kibish finds. The Arago
21 specimendoes not show these' most distinctive Neanderthal characters of the face
~fi 01
D
ca
R
t3
0)
and hence would
classified as a Neanderthal using all available criteria.
-g I-, 4-, • •r-1 cd
4 r.) - H
cd
• ri ri
-H • cd
ra) xy
'717,71,71.1T, Le)
4) a) Ca cd
Anterior mastoid
erectus.
Cranial shape
Nasio-frontal angle
Large occipitomastoid crest
Suprainiac fossa
VI V/ 0 0 0 0 Z
cc! g 0 0 0 o o r
ZZZZ +DA
cd a)
'JD /7=1 cD
O H ri g 4-3
g rn u cc) cd cd cu 0 8
(1) P•
▪ tcr 0 0 0 0 O o O a)
er, er, rr, H) V 1-r) 00

L'HOMME DE PETRALONA :
COMPARAISONS AVEC L'HOMME DE TAUTAVEL
par
Louis de BONIS*et Jean MELENTIS"
}Mourne :
Malgre des differences, dont quelques unes sans doute liges an
sexe, le crane de Petralona et l'homme de Tautavel presentent plusieurs
points communs. Certains de ceux-ci correspondent A des caracteres que
l'on pent considerer come primitifs et gvoquent le stade archaique
d'Homo erectus. On peut relever la forme du torus sus-orbitaire, les
proportions du frontal, l'aplatissement du parietal ou la robustesse de
la denture. D'autres en revanche sont des synapomorphies, des caracteres
derives partages avec la sous-espece Homo sapiens neandertalensis ; ils
permettent de classer les deux fossiles dans cette sous-espece. Sur la
base des memos criteres on doit faire entret dans ce taxon les formes
africaines de Broken Hill et de Bodo. Un cladogramme resume cette hypo-
these phyletique et d'autres opinions sont discutges.
Cette ligngesemble s'etre developpee en Europe et avoir envahi
l'Afrique au morns A deux reprises. Wapres les datations effectuees A
Tautavel et A Petralona la divergence entre les deux sous-especes d'Homo
sapiens semble avoir ate tres precoce.
Abstract :
Petralona and Tautavel men show several differences, some of them
resulting from sexual differences. They show also a lot of similarities.
Some of these similarities are primitive or plesiomorphic characters shared
with the archaic man Homo erectus. They are the shape of the supraorbital
torus, the narrowness of the frontal bone, the flatnessof the parietal
bone and the robustness of the dentition. The other ones are synapomorphies
or derived characters shared with the sub-species Homo sapiens neandertalen-
sis. They are the shape of the maxillar without canine fosse, the low posi-
tion of the infra-orbital foramen, the shape of the malar bone and the
*Universite de Poitiers - Facultg des Sciences - Laboratoire de Paleontologie des Vertebras et de Paleontologie humaine -40 avenue du Recteur Pineau - 86022 Poitiers cedex.
"Universite de Thessalonique - Laboratoire de Geologie et Paleontologie - Thessalonique - Grace.





























