Embed Size (px)
Citation preview

~ Pergamon 0306-4522(95)00255-3
Neuroscience Vol. 69, No. 2, pp. 601-618, 1995 Elsevier Science Ltd
Copyright © 1995 IBRO Printed in Great Britain. All rights reserved
0306-4522/95 $9.50 + 0.00
BAROSENSITIVE NEURONS IN THE ROSTRAL VENTROLATERAL MEDULLA OF THE RAT IN VIVO: MORPHOLOGICAL PROPERTIES AND RELATIONSHIP
TO CI ADRENERGIC NEURONS
J. LIPSKI,* R. K A N J H A N , B. K R U S Z E W S K A t and M. SMITH
Department of Physiology, School of Medicine, University of Auckland, Auckland, New Zealand
A~traet--The aim of this study, conducted in anaesthetized rats, was to examine the morphology of barosensitive neurons in the rostral ventrolateral medulla and their immunoreactivity for a catecholamine synthesizing enzyme, tyrosine hydroxylase. Thirty neurons displaying inhibitory postsynyptic potentials following stimulation of the aortic depressor nerve were intracellularly labelled with Lucifer Yellow or Neurobiotin. Some of these neurons could be excited antidromically from the second thoracic segment of the spinal cord, with conduction velocities of spinal axons ranging from 1.9 to 7.2 m/s. The filled somas were found immediately caudal to the facial nucleus and ventral or ventromedial to compact formation of the nucleus ambiguus. Some dendrites reached the ventral medullary surface. Axons usually projected dorsomedially and then made a sharp rostral and/or caudal turn. The caudally projecting axon could, in some cases, be followed to the first cervical segment of the spinal cord. Seven cells issued fine axon collaterals on the ipsilateral side. These were identified mainly in two areas: in the rostral ventrolateral medulla (or immediately dorsomedial to that region), and within the dorsal vagal complex. Seven of 27 examined cells (26%) were tyrosine hydroxylase-immunoreactive and were classified as CI adrenergic neurons. No clear relationship was found between the presence or absence of adrenergic phenotype and the morphology of filled cells. However, the amplitude of aortic nerve-evoked inhibitory postsynaptic potentials was significantly larger in tyrosine hydroxylase-positive neurons. Possible reasons for the low percentage of barosensitive cells with tyrosine hydroxylase immunoreactivity found in this study, in comparison with previously published estimates, are discussed.
This is the first study describing the morphology of neurons in this part of the medulla identified as barosensitive in vivo, and directly'demonstrating adrenergic phenotype in a subset of these neurons.
Key words: presympathetic (sympathoexcitatory) neurons, intracellular recording and labelling, tyrosine hydroxylase, central cardiovascular control.
The rostral and ventrolateral part of the medulla oblongata (the rostral ventrolateral medulla, RVLM) contains neurons that are believed to play a key role in controlling the sympathetic vasomotor tone and the level of arterial blood pressure (for review, see Refs 13, 21, 22, 38, 54). The main criterion for identification of these R V L M vasomotor neurons is a strong inhibition of their ongoing activity following stimulation of arterial baroreceptors or baroreceptor
*To whom correspondence should be addressed at: Department of Physiology, School of Medicine, University of Auckland, Private Bag 92-019, Auckland, New Zealand.
tPresent address: Department of Psychiatry, School of Medicine and Dentistry, University of Rochester Medi- cal Center, Rochester, NY 14642, U.S.A.
Abbreviations: DAB, 3,3'-diaminobenzidine tetrahydrochlo- ride; EPSP, excitatory postsynaptic potential; FITC fluorescein isothiocyanate; IPSP inhibitory postsynaptic potential; PNMT phenylethanolamine-N-methyltrans- ferase; RVLM, rostral ventrolateral medulla; TH tyro- sine hydroxylase.
afferents. As a large proport ion of these neurons project to thoracolumbar segments of the spinal cord, the other major criterion is the presence of antidromic excitation after spinal stimulation. The R V L M neur- ons which fulfil both criteria (i.e. are barosensitive and bulbospinal) are often referred to as presympa- thetic or sympathoexcitatory.
The electrophysiological properties of these neur- ons have been extensively studied extracellularly (e.g., Refs 6, 34, 39, 44, 47, 74), and more recently with sharp intracellular microelectrodes. 14'4°,41'69'79 On the
other hand, little is known about their morphology. An early analysis of the cytoarchitecture of the caudal port ion of the nucleus reticularis paragigantocellu- laris, which forms the major part of the RVLM, revealed that this region in the rat is heterogenous and contains at least eight types of densely-packed, small to medium-sized cells. 2 Two studies analysed the morphology of putative presympathetic neurons in the R V L M after their intracellular labelling in tissue slices. 29'7° However, in such studies identifi- cation of baroreceptor input is not possible, and
601

602 J. Lipski et al.
therefore some of these neurons may not be involved in cardiovascular control .
Recent studies indicate tha t R V L M presympa- thetic neurons use an exci tatory amino acid (gluta- mate) as the ma in neuro t ransmi t t e r (e.g., Refs 1, 11, 43, 46, 48, 71). A subset of R V L M neurons with a catecholaminergic phenotype , also k n o w n as the C1 adrenergic cell system, may release adrenal ine as a co- t ransmit ter . A l though these adrenergic neurons have been implicated in cont ro l of cardiovascular system (for review, see Refs 22, 54, 60 and Discus- sion), their involvement in exci tat ion of pregangl ionic sympathet ic neurons remains controvers ia l (e.g., Refs 11, 21,57,72). In addi t ion, there has been no direct evidence tha t these adrenergic neurons have an in- h ibi tory barorecep tor input.
One aim of our s tudy was to investigate the morpho logy of intracellularly labelled R V L M neur- ons identified as barosensi t ive in vivo. The o ther was to directly examine the relat ionship between these neurons and the adrenergic cells of the C i group.
EXPERIMENTAL PROCEDURES
Surgery and recording techniques
Experiments were conducted on 24 adult male Wistar rats which were also used in a separate electrophysiological study of RVLM neurons describing the surgical and record- ing techniques in more detail. 41 In brief, the rats were anaesthetized with sodium pentobarbitone (Nembutal, 80 mg/kg, i.p., supplementary doses 4qSmg/h, i.v.). Three nerves were dissected and prepared for standard bipolar stimulation or recording: the phrenic nerve, aortic depressor nerve and the mandibular branch of the facial nerve. The dorsal surfaces of the medulla oblongata and spinal cord (C 1 to Tz) were exposed. Two monopolar stimulating electrodes were placed in the lateral funiculus at T 2 segment ipsilateral to the recording side in the RVLM, and were used for antidromic identification of neurons projecting to this spinal level (0.2 ms pulses, up to 1.0nA). Another stimulating electrode was placed inside the oesophagus, as described previously. 37
The aortic nerve, which in the rat contains only barore- ceptor afferents, was stimulated at a frequency of 0.5 or 1 Hz, with trains of three stimuli (0.2 or 0.5 ms long) 5 ms apart. The stimulus intensity was five times the threshold for evoking a depressor response during stimulation of the nerve at 100 Hz (threshold, 0.1-0.5 V). In each experiment, low-impedance microelectrodes filled with 2 M NaC1 (resist- ance, 4-8 M~) were first used for mapping of the position of the facial nucleus as described by Brown and Guyenet. 6 This was followed by antidromic mapping of compact formation of nucleus ambiguus, as recently described in our laboratory. 37 The rostrocaudally oriented column of oesophageal motoneurons identified by such mapping served as a convenient reference area for subsequent intra- cellular recordings. Intracellular recordings were made with high-impedance microelectrodes containing either 1 or 1.5% Neurobiotin 35 in 0.4 M KC1 (resistance, 80 100 M[~), or 3 or 5% Lucifer Yellow CH 65 in 0.4 M lithium chloride (resist- ance, 75-85 MQ). Only somatic impalements with the mem- brane potential greater than - 4 0 m V (measured after a period of initial stabilization) were considered for further analysis. Lucifer Yellow was injected by using hyperpolarlz- ing current pulses and Neurobiotin with depolarizing pulses (400 ms long, 2 Hz, 2 ~ nA) applied for up to 20 min. The range of charge transfer was 10 to 20 nA x rain for Lucifer Yellow, and 15 to 50 nA x min for Neurobiotin. In most
experiments the animals were perfused with fixatives less than 3 h after the last intracellular injection (with one or two injections made per experiment). In some experiments (n = 5), an attempt was made to increase transport of Neurobiotin into distant parts of the axon by maintaining these animals under deep anaesthesia for (~12 h after injec- tion (see Ref. 42).
Histological analysis
Rats were perfused transcardially with 100-200ml of 0.9% NaC1 containing 10 mM sodium nitrite, followed by 500 ml of 0.1 M phosphate buffer (pH 7.4) containing 4% formaldehyde and 0.1% glutaraldehyde. The brainstem was then removed and stored for 12 18 h in the same fixative at room temperature. Sections (35 50/~m) were cut with a Vibratome and rinsed in 0.1 M phosphate buffer. Tissue blocks containing Lucifer Yellow-filled cells were cut in the transverse plane, while the blocks containing Neurobiotin- filled cells were cut in the parasagittal plane. Two different procedures were subsequently used, depending on whether neurons were injected with Lucifer Yellow or Neurobiotin.
For material containing cells injected with Lucifer Yel- low, sections were mounted serially on plain glass slides and coverslipped with glycerol-carbonate buffer (pH 8.6). Sec- tions containing Lucifer Yellow-labelled cell bodies and their processes were selected with the aid of a fluorescent microscope (Leitz Diaplan, filter block D). The section containing cell body, as well as two adjacent sections, was removed from the slides, washed in phosphate buffer and incubated for 48 72 h (4°C) with an antibody to tyrosine hydroxylase (Eugene Tech, polyclonal antibody raised in rabbit, 1:300; or a monoclonal antibody from Boehringer Mannheim, 1:200) in buffer containing Triton X-100 (0.3%, v/v), 10% blocking serum, and (in mM) NaCI 120, KCI 5, NaH2PO 4 1.5, Na2HPO 4 8.5, Tris base 10, sodium merthio- late 1 (pH 7.4). After several washes, they were incubated overnight in a biotinylated secondary antibody (sheep anti- rabbit, 1 : 500; or sheep anti-mouse, 1 : 500, Sigma), followed by washes and incubation (4~5 h) with Streptavidin Texas Red (1:200, Sigma). Incubations with secondary antibodies and Streptavidin-Texas Red were conducted in the same buffer which was used with the primary antibodies (but only with 1% blocking serum) at room temperature with con- tinuous agitation. Sections were then washed, coverslipped with glycerol-carbonate buffer, briefly viewed with a fluor- escent microscope (High Q Texas Red filter block, Chroma Technology Corporation; and filter block D) and analysed with a confocal microscope (see below). Subsequently, the sections were again removed from slides and incubated, together with all remaining sections which were not exam- ined for TH immunoreactivity, for 24-48 h in the mouse antibody to Lucifer Yellow (1:2000). 53 This was followed by an overnight incubation in a biotinylated sheep anti-mouse antibody (1 : 500, Sigma) and then for 4-12 h in ExtrAvidin peroxidase (1 : 1000, Sigma). Lucifer Yellow immunoreactiv- ity was revealed by incubation for 3 ~ min in Tris-HC1 buffer (pH 7.6) containing 3,Y-diaminobenzidine tetrahy- drochloride (DAB, 0.05%), nickel ammonium sulphate (0.6%) and hydrogen peroxide (0.005%). Finally, the sec- tions were mounted serially on to gelatin-coated slides, dehydrated and coverslipped. The permanently visualized cells ('hard copies') were used for standard camera lucida reconstructions which were made with 25 x or 40× dry microscope objectives, or for morphometric measurements with a 100 × oil immersion objective.
For material containing cells injected with Neurobiotin, sections were usually incubated (see below for the details of an abbreviated protocol used for three cells) with ExtrAvidin fluorescein isothiocyanate (FITC) conjugate (1:200, Sigma). They were then mounted, coverslipped with glycerol-carbonate buffer and viewed under the fluorescent microscope (filter block L3, Leitz). Free-floating sections containing Neurobiotin-labelled cell bodies (as well as two

Properties of
adjacent sections) were then incubated with one of the two primary antibodies to tyrosine hydroxylase (TH), as de- scribed above. Subsequently, sections were incubated with an anti-rabbit (or anti-mouse) antibody conjugated with Texas Red, raised in goat (1:500, Jackson Immuno- Research). They were then mounted in glycerol-carbonate buffer, and the tyrosine hydroxylase immunoreactivity of intracellularly filled neurons was briefly examined with a fluorescent microscope (Texas Red filter block, and filter block L3) followed by analysis with a confocal microscope. Subsequently, the sections were again removed from slides and incubated (together with all remaining sections not examined for TH immunoreactivity) for 4-12h in DAPA biotin-horseradish peroxidase (1:10000, Sigma). s After washes, they were reacted in Tris-HCl buffer contain- ing DAB, nickel ammonium sulphate and H202, mounted serially and reconstructed. For three neurons injected with Neurobiotin, the reaction step with ExtrAvidin-FITC was omitted and sections were incubated in ExtrAvidi~ peroxidase (l:1000, Sigma) dissolved in the same buffer which was used for incubations with primary antibodies. Following washes, the neurons were visualized with a DAB/H202/Ni protocol.
Following camera lucida reconstructions, photography and morphometric measurements, the coverslips were re- moved and the sections were counterstained using a stan- dard Nissl technique. Although this step decreased the intensity of staining of the cell bodies and processes belong- ing to injected neurons, it allowed them to be located with respect to major nuclei in the brainstem. For measurements of the size of the cell bodies, the values of the major and minor axes of the best-fit ellipse were taken. No correction was made for tissue shrinkage in morphometric measure- ments. Cell shrinkage, resulting from the histological pro- cedures, was estimated to be 9% (range, 7-11%) from comparison of cell diameter (n = 8) before tissue dehy- dration and DAB staining (in wet-mount fluorescent ma- terial), and after these procedures.
Confocal laser-scanned images were obtained with Leica TCS 4D system equipped with a Krypton/Argon Laser, and configured with a Leitz DMR microscope. Fluorochromes were viewed separately using single channel mode. FITC filter configuration (488 nm excitation) was used for viewing Lucifer Yellow or FITC. Rhodamine-Texas Red filter (568 nm excitation) was used for viewing Texas Red. Images were obtained with a 512 x 512 pixel resolution in the x-y mode, or 512 x 35 in z-series mode. Leica oil immersion objectives {40 x, or 63 x ; numerical aperture, 1.0 and 1.4, respectively) were used. Optical sectioning through intra- cellularly labelled neurons was always conducted at the level at which the cell's nucleus was most visible. The images were averaged (16-32 times), saved on 128 MB optical discs and processed (adjustments of the brightness and contrast) with the NIH Image software on a Quadra 950 computer. The hard copies (gray scale levels, 1~255) were obtained with a 430 Spectra Star printer, with 300 d.p.i, resolution. Control experiments were conducted to exclude the possibility that during excitation with 488 nm some fluorescence emitted by Lucifer Yellow or FITC could be detected through the Texas Red filter, and thus incorrectly taken as evidence of double-labelling. Two facial motoneurons (both excited antidromically from the mandibular branch of the facial nerve) were labelled intraceUularly; one with Lucifer Yellow and another with Neurobiotin and visualized as described above. In both cases, large charge transfers were used (50 and 60nA x min, respectively). The motoneurons were brightly fluorescent when excited with 488 nm and viewed with the FlTC/Lucifer Yellow filter configuration. With the 568 nm excitation and the Texas Red filter, both motoneu- rons showed no detectable fluorescence when conditions similar to those used to examine TH immunoreactivity were used (similar laser power, pinhole size and the gain of the photomultiplier).
RVLM neurons 603
RESULTS
Electrophysiological identification
All intracellularly injected neurons (n =34 ; 16 injected with Lucifer Yellow, 18 with Neurobiotin) met the following two criteria: (i) they were located near the caudal end or immediately behind the facial nucleus, and ventral or ventromedial to compact formation of the nucleus ambiguus; and (ii) they displayed short latency inhibitory postsynaptic potentials (IPSPs, onset latency, 30.3 + 9.5 ms; mean ___ S.D.) following stimulation of the ipsilateral or contralateral aortic depressor nerve with bursts of three pulses. Thirty neurons were recovered in histo- logical material (15 labelled with Lucifer Yellow and 15 with Neurobiotin) and were further analysed. Five of these neurons could be excited antidromically following near threshold stimulation of descending axons in the T2 segment of the spinal cord (conduc- tion velocity, 3.8 __+ 1.1 m/s; threshold, 216 +__ 178/~A) and therefore were positively identified as barosensi- tive and bulbospinal (presympathetic). One more neuron, which was not tested antidromically, was classified as bulbospinal on the basis of histology (Figs 8B, 10B). Spinal projection was not tested in 16 neurons in which spike inactivation occurred soon after impalement. In addition, the antidromic test was not conclusive in the other nine neurons which displayed short-latency ( < 10ms) suprathreshold excitatory postsynaptic potentials (EPSPs) after stimulation in the spinal cord, and therefore the lack of antidromic responses could be due to collision with synaptically induced action potentials (see Ref. 40). Eighteen neurons showed largely irregularly occur- ring action potentials, with no clear respiratory modulat ion of the firing frequency. All injected neur- ons displayed large synaptic activity (EPSPs and IPSPs, peak-to-peak amplitude > 5.0 mV) which, in spontaneously active neurons, could be particularly well seen when the firing was slowed down with a small continuous hyperpolarizing current (cf. Ref. 41). No evidence was found for the presence of gradual depolarizations (autodepolarizations) be- tween individual action potentials, except of conven- tional afterhyperpolarizations following each action potential. Examples of the firing pattern, synaptic responses recorded after stimulation of the aortic depressor nerve, and antidromic action potentials evoked after stimulation in T2 spinal segment are shown in Figures 1 and 9A, B.
Relationship to C 1 neurons
The next step of analysis aimed at assessing im- munoreactivity for TH. TH-immunoreact ivi ty was found within the cytoplasm of the cell bodies, and in the processes of numerous neurons in the RVLM. The distribution of immunoreactive cell bodies (irre- spective of the type of antibody used in our exper- iments) was similar to that reported in previous studies which described the location of TH or

604 J. Lipski et al.
phenylethanolamine-N-methyltransferase (PNMT)- immunoreactive neurons in the medulla oblongata of the rat (e.g., Refs 25, 62, 76) As virtually identical numbers of R V L M neurons were reported to be immunoreactive for each of the two enzymes, positive staining for the T H in this part of the medulla oblongata was considered to be a marker of adrener- gic neurons. In the transverse sections, taken between the caudal end of the facial nucleus and 500 #m caudal to this point, TH-immunoreact ive cells ex- tended from below the compact formation of nucleus ambiguus to near the medial borders of the pyramidal tract and the inferior olive. In the parasagittal sec- tions, immunoreactive neurons formed a rostrocau- dally oriented column which terminated, at its rostral end, near the caudal end of the facial nucleus.
A total of 27 R V L M barosensitive neurons (15 labelled with Lucifer Yellow and 12 with Neurobi- otin) were examined under both the fluorescent and confocal microscopes for evidence of double- labelling. Lucifer Yellow-labelled neurons showed
fluorescence which was always stronger in the nucleus than in cytoplasm (Fig. 2E). The cell bodies of neurons injected with Neurobiot in and visualized with F ITC were more uniformly fluorescent, but the intensity of the staining of the nucleus was sometimes slightly lower than that of cytoplasm (Fig. 2A). TH-immunofluorescence was detected in seven of these neurons (26%; two labelled with Lucifer Yellow and five with Neurobiotin) (Fig. 2). Under the 568 nm excitation and the Texas Red filter configuration, the fluorescence was present in these neurons only within the cytoplasm and proximal processes, but not in cell nuclei. This observation confirmed that the fluor- escence was generated by the Texas Red-conjugated antibody, and not due to contamination from Lucifer Yellow or FITC.
TH-immunoreact ivi ty was not detected in 20 re- maining intracellularly labelled neurons (Fig. 3). Most of these neurons (as well as the TH-posit ive cells) were in close proximity to other immuno- reactive cell bodies and processes which were detected
A
-6o v
~ ~ ~ ~ ~ ~ I
ls
B . . . C
$ i 1 _ _
2 mV I ~
5 mV[ 10 ms
Fig. 1. Example of electrophysiologicai identification of a barosensitive RVLM neuron. The neuron was identified as TH positive (Fig. 2 C, D). (A) Synaptic activity and action potentials observed when a small continuous hyperpolarizing current ( - 0.05 nA) was used to slow down the firing. The traces from above: microelectrode signal, arterial blood pressure (calibration bar, 0-100 mmHg), activity of the phrenic nerve, and tracheal pressure (calibration bar, I~10cmH20 ). (B) The aortic nerve-evoked potentials (top, eight superimposed sweeps; middle, 26 averaged sweeps; bottom, extracellular control with 32 averaged sweeps). Asterisks mark stimulus artefacts. (C) Antidromic action potentials evoked by near threshold stimulation in T 2 segment (stimulus marked by an asterisk, six superimposed sweeps pre-triggered by 'spontaneous' action potentials). Note that spinal stimulation was followed by antidromic responses only in three sweeps, and that the level of the membrane potential at which the action potentials occurred after spinal stimulation was about 5 mV more negative than the threshold at which 'spontaneous' action potentials
were induced by ongoing synaptic activity. Action potentials are truncated in A and C.

Fig. 2. Confocal microscope images of three intracellularly filled neurons which were TH-immunoreactive. The panels on the left show intracellularly filled neurons (A and C with Neurobiotin, E with Lucifer Yellow) under excitation for Lucifer Yellow/FITC. The corresponding panels on the right show TH-immunofluorescence of the same neurons (arrows, Texas Red filter configuration). Electrophysiologi-
cal recordings from the neuron in panels C and D are shown in Fig. I. Scale bars = 20/~m.

606 J. Lipski et al.
C
4
Fig. 3. Example of confocal microscope images of a neuron filled with Neurobiotin which was TH negative (A, B), and of a confocal z-scan conducted through the same section (C). (A) Filled neuron visualized with the 488 nm excitation. (B) Excitation with 568 nm. The same field and z-depth as in A. (C) Confocal z-scan. The two borders indicated by arrows represent surfaces of a 35-#m-thick section. Scale
bar = 20 pm.
either in the confocal microscope (in optical sections used to image the intracellularly labelled neurons) or by using fluorescent microscope. As the histological sections were relatively thick (35-50 #m), one factor which could explain the lack of double-labelling in this group of neurons is incomplete penetrat ion of the primary antibody or other immunoreagents into the sections. This factor was excluded following the analysis of confocal z-scans conducted through some sections, which always revealed the presence of TH- immunoreactive cell bodies and processes not only near the surface, but also in the middle of the sections (Fig. 3C).
A clear relationship was found between the pres- ence or absence of TH-immunoreact ivi ty, and the amplitude of aortic nerve-evoked IPSPs. In TH- negative neurons, the amplitude was 1.2 + 0 .7mV (range, 0.5-3.2mV), and in TH-posit ive neurons 2 . 9 + 1.4mV (range, 1.0-5.2mV) (P <0.01). The onset latency of these IPSPs was similar in both groups (TH-positive, 33.7___ 4.3 ms; TH-negative, 28.0_+ 11.1; P <0.2) . The conduction velocities of spinal axons of three TH-posit ive neurons were 1.9, 4.1 and 5.0 m/s, and of two TH-negat ive neurons 4.0 and 7.2 m/s.
The numbers of examined cells are summarized in Table 1, and their location in Figure 4.
Morphological properties
A total of 17 intracellularly filled barosensitive neurons were processed according to one of the two protocols used to convert the Lucifer Yellow or F I T C fluorescent neurons into 'hard copies' (see Exper- imental Procedures). The quality of staining was considered sufficient for camera lucida reconstruc- tions of 14 neurons which showed well-stained cell body and dendrites (e.g., Figs 5B, 10A, B), with axons stained for a minimum distance of 1.0 mm from cell bodies. Four Lucifer Yellow-injected neurons were
Table 1. Intracellularly labelled and recovered barosensitive rostral ventrolateral medulla neurons
Lucifer Yellow Neurobiotin
Total number of neurons 15 (2) 15 (4*) Analysed for TH 15 (2) 12 (3) immunoreactivity Double-labelled 2 (0) 5 (3) (intracellularly labelled and TH + ) Converted to 'hard 4 (1) 10 (4*) copies' and reconstructed Cells with medullary 2 (1) 5 (3) axon collaterals
Number of neurons positively identified as bulbospinal given in brackets. Numbers with asterisk include a cell identified as bulbospinal using morphological criteria.

Properties of RVLM neurons 607
TH - immunoreact ive
non TH - immunoreactive
f ~ . ~ / il not examJned for TH r CF j
500 ~m
Fig. 4. Location of intracellularly labelled and recovered barosensitive RVLM neurons established on the basis of recording coordinates, with the centre of the compact formation (defined during initial electrophysiological map- ping, see Experimental Procedures) used as a reference point. All recording points projected on a representative transverse section of the medulla taken immediately caudal to the facial nucleus. CF, compact formation; IO, inferior
olive; Pyr, pyramidal tract.
reconstructed in the transverse plane, and 10 injected with Neurobiotin in the parasagittal plane. An illus- tration of the stages leading to permanent visualiza- tion of a cell injected with Lucifer Yellow is shown in Figure 5.
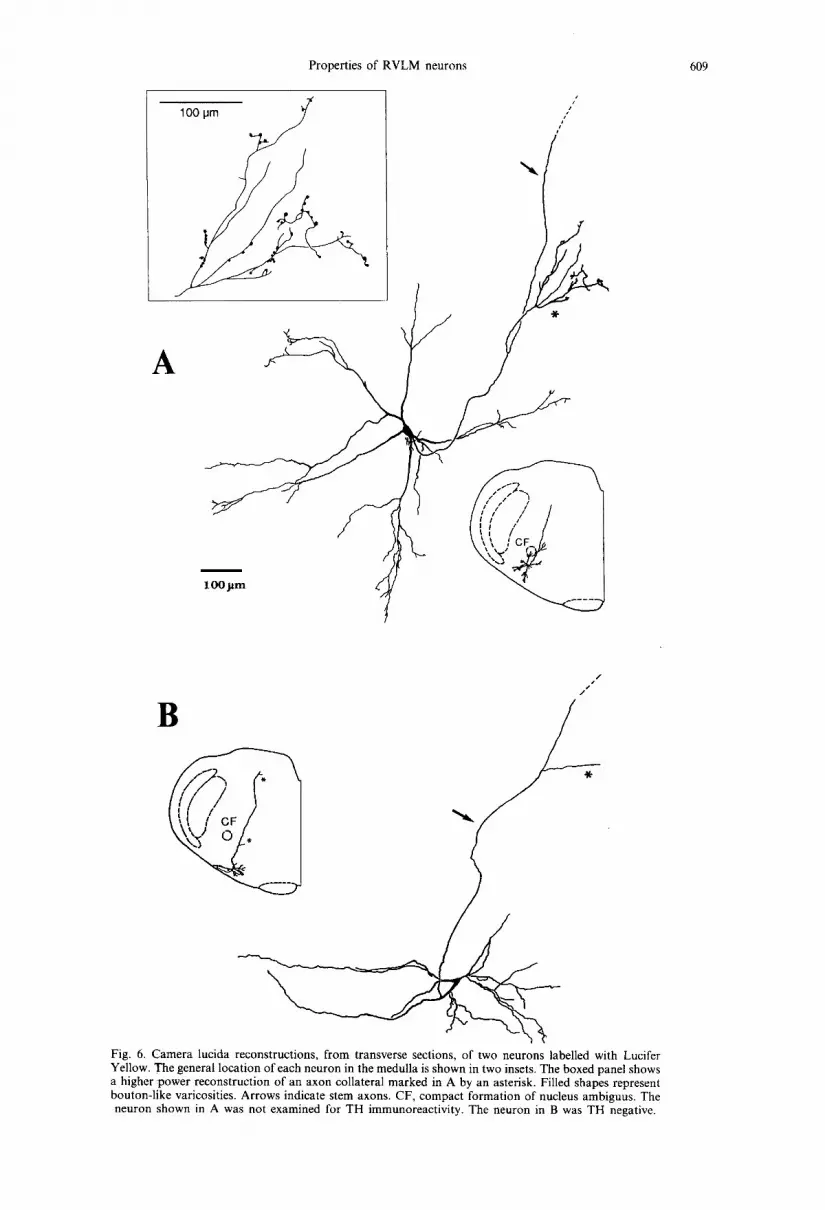
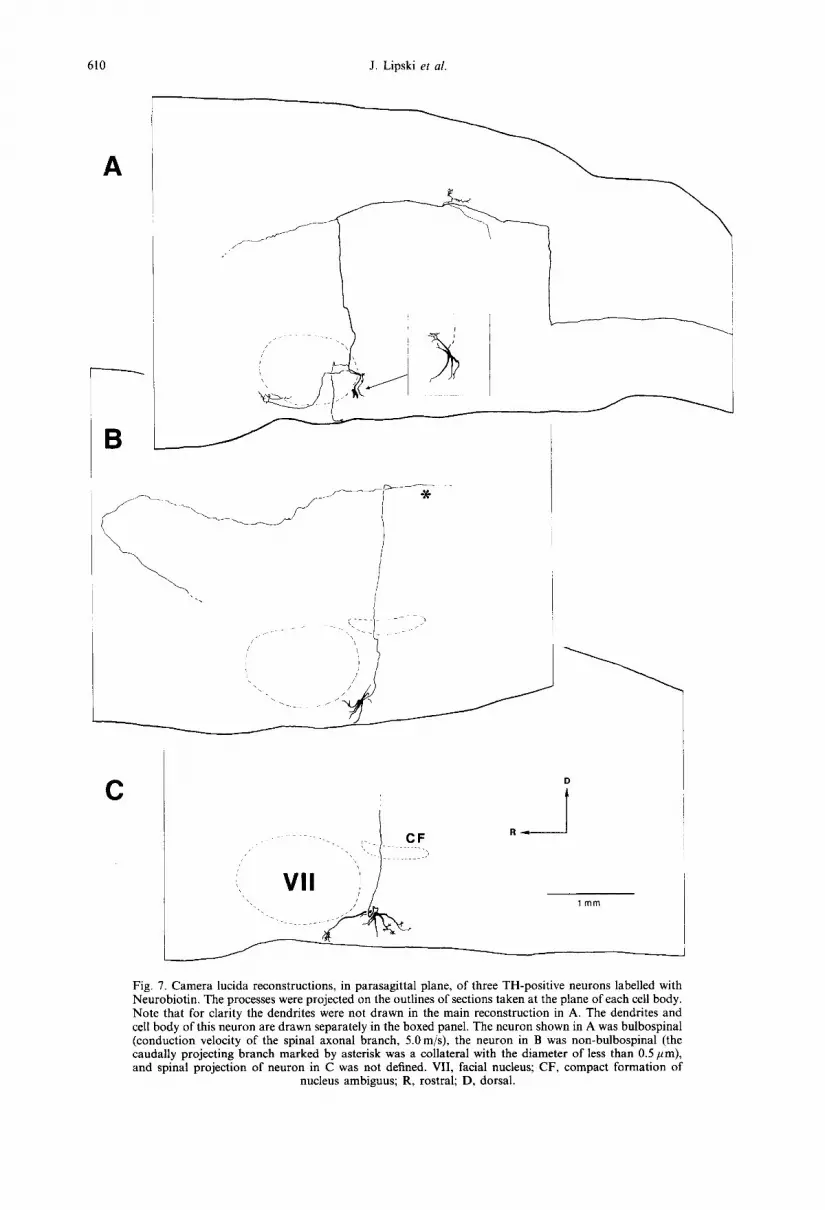
All cell bodies which were multipolar, oval or fusiform in shape, were found in the area identified during initial electrophysiological mapping (near the caudal end of the facial nucleus or immediately behind, and ventral or ventromedial to the compact formation). Their long axis, which had a variable orientation, ranged from 14 to 47pm (26.0 + 9.7/~m), and the short axis from 10 to 25/~m (14.8 -I- 4.8/~m), The size of the cell bodies, expressed as the mean value of the long and short axes, was 20.6 + 7.2/tm. This size was generally larger in bul- bospinal neurons (27.7 _+ 5.3/~m, n = 5) than in non- bulbospinal cells (17.1-I-1.5/~m, n =5) (P <0.01). Cell bodies issued three to eight primary dendrites (mean, 6.0) which branched two to four times and usually extended 300-650/~m. The longest dendrite, identified in the transverse plane, extended 780/~m from the cell body. No evidence for a predominant orientation in any plane was found for the ex- tension of the dendritic tree, with the exception of dendrites which in most cells (n =9 ) projected towards the ventral medullary surface and ter- minated immediately beneath the pia mater (Figs 6B, 7, 8C, 10B). Some dendrites had a beaded appearance resembling en passan t boutons of axon collaterals (not illustrated) and some issued, near the ends, several brush-like branches (Figs 6A, 7C, 10B). There was no significant difference between the number of primary dendrites belonging to bulbospinal neurons (6.5-t-1.3) and non-bulbospinal neurons (5.7 + 0.5).
Axons originated either from the cell body (n = 12) or from a primary dendrite (n = 2). In general, they could be traced for longer distances when neurons were filled with Neurobiotin. This was due to three factors: (i) generally larger charge transfer; (ii) often longer post-injection survival times; and (iii) recon- structions made in the parasagittal plane facilitated tracing of axons projecting in the rostral or caudal directions. After leaving the cell body, the axons usually projected dorsomedially towards dorsomedial medulla (Figs 6-8). The diameter of this portion of the axon, measured 100-200 #m from the cell body, was 1.22 _+ 0.35 #m (range, 0.7-1.8 #m). The diam- eter of axons of bulbospinal neurons (1.58 _+ 0.21/~m, n = 5) was larger than that of non-bulbospinal neur- ons (0.98 +_0.19/~m, n =4 ) (P < 0.01). Axons of three Neurobiotin-filled neurons were traced to the point in the dorsomedial medulla where they made a sharp turn towards the spinal cord, and could be followed until the caudal end of the C1 spinal seg- ment (Figs 7A, 8B, 9, 10E). The descending axon of one bulbospinal neuron showed a different trajectory; the descending spinal branch originated from a short (140 #m) stem axon which did not project towards the dorsomedial medulla (Fig. 9). In this, and one other neuron (Fig. 7A), a bifurcation of the stem axon was observed, approximately at the same ros- trocaudal level as the cell body, into major caudal and rostral branches. In five neurons, a rostrally pro- jecting axon could be followed for more than 1.0 mm from the point of bifurcation or sharp turn in dorso- medial medulla. In three of these neurons, the axon curved towards the region dorsomedial to the su- perior olivary nucleus (the area containing the A5 nora- drenergic cell group, e.g., Ref. 36) (Figs 7B, 8A, C).
Seven cells issued axon collaterals, which origi- nated directly from the dorsomedially projecting stem axon (Figs 5, 6A, 7A, 8B), or from the main axonal branches projecting in the rostral or caudal direction (Figs 7A, 9, 10C, D). These collaterals were always thin in diameter (approximately 0.3~).5/~m) and could be traced for only a few hundred micrometres from their origin. In some cases they divided and showed bouton-like varicosities. Such collaterals with terminal-like arborizations were identified in four regions (including three medullar regions): (i) in the general area of the RVLM or immediately dorsome- dial to it (five neurons; Figs 6A, 7A, 8B, 9). These collaterals originated from the stem axon at distances ranging from 90 to 440 #m from the cell body or, as illustrated in Fig. 9, from the caudally projecting axonal branch; (ii) in the dorsomedial medulla, within the dorsal vagal complex (three neurons; Figs 7A, 8A, 10G); (iii) in the A5 region (dorsomedial to the superior olivary nucleus, 1 neuron; Figs 7A, 10F); and (iv) in the general region of the caudal ventrolat- eral medulla (one neuron; Fig. 9).
No clear difference was found between the mor- phology of intracellularly labelled TH-immuno- reactive and non-immunoreactive neurons. Both

C
J. Lipski et al.
types included the subpopulations of bulbospinal and non-bulbospinal ceils, as well as cells with and with- out axon collaterals issued in the ipsilateral RVLM and the dorsal vagal complex (Figs 7, 8) The collat- eral arborizing in the A5 region was identified only in one TH-positive neuron, but two TH-negative cells had axonal branches which also appeared to project to that region (Fig. 8A, C). The collaterals in the caudal ventrolateral medulla could not be ascribed to any group as the neuron which issued such collaterals (Fig. 9) was not examined for TH-immunoreactivity.
DISCUSSION
Elec t rophys io logy and location
Electrophysiological properties of the rat RVLM barosensitive neurons have been described in detail in two recent studies conducted in our laboratory using intracellular microelectrodes in vivo. 4°'41 A low num- ber of stable intracellular recordings followed by successful labelling contrasted with the ease of recording the activity of such neurons with extracellu-
lar microelectrodes (e.g., Ref. 34). The difficulty was due not only to the small size of the cell bodies, but also to the relatively low density of barosensitive neurons in the examined part of the RVLM, esti- mated to be around 4°/o. 4t
The presence of IPSPs evoked by stimulation of the aortic depressor nerve was used as a way of identifying the baroreceptor input to labelled neurons. Interestingly, the mean amplitude of these synaptic potentials was significantly bigger in the TH-immunoreactive neurons than in TH-negative cells. This could be due to higher input resistance of adrenergic neurons, larger synaptic currents gener- ated by antecedent inhibitory interneurons, or simply to greater damage and depolarization of adrenergic neurons resulting from impalement. The contribution of these factors awaits future investigation. However, it should be noted that IPSP amplitudes could not be used as an unequivocal predictor of TH-immunoreac- tivity since some neurons responding with large IP- SPs (> 2.0 mV) were TH-negative, and some others with small IPSPs (< 2.0 mV) were TH-positive.
D ~ii! 11 i !
608
Fig. 5. An illustration of the three stages leading to permanent visualization of a Lucifer Yellow stained neuron. (A) Image photographed with the fluorescence microscope. (B) The same neuron viewed with bright-field microscopy after immunohistochemistry for Lucifer Yellow. (C) Camera lucida reconstruc- tion. Asterisk indicates a fine collateral originating from the stem axon at the distance 120 ttm from the cell body (interrupted lines). Scale bar = 110 pm. D, dorsal; M, medial. The neuron was TH negative.
Properties of RVLM neurons
I O0 pm ,~ i
t i
/ i
609
lO0~m
(,// '
B
/
/
Fig. 6. Camera lucida reconstructions, from transverse sections, of two neurons labelled with Lucifer Yellow. The general location of each neuron in the medulla is shown in two insets. The boxed panel shows a higher power reconstruction of an axon collateral marked in A by an asterisk. Filled shapes represent bouton-like varicosities. Arrows indicate stem axons. CF, compact formation of nucleus ambiguus. The neuron shown in A was not examined for TH immunoreactivity. The neuron in B was TH negative.
610 J. Lipski e t al.
A
.f_fy__~i B
//
" ~\ i lj
C D
l m m
Fig. 7. Camera lucida reconstructions, m parasagittal plane, of three TH-positive neurons labelled with Neurobiotin. The processes were projected on the outlines of sections taken at the plane of each cell body. Note that for clarity the dendrites were not drawn in the main reconstruction in A. The dendrites and cell body of this neuron are drawn separately in the boxed panel. The neuron shown in A was bulbospinal (conduction velocity of the spinal axonal branch, 5.0 m/s), the neuron in B was non-bulbospinal (the caudally projecting branch marked by asterisk was a collateral with the diameter of less than 0.5 ,um), and spinal projection of neuron in C was not defined. VII, facial nucleus; CF, compact formation of
nucleus ambiguus; R, rostral; D, dorsal.

Properties of RVLM neurons 611
A
, . . - . . . . . . , <~LL[ .[~ ]-L ~ i
I i i
\ ,
B
C
i i
R
CF
• \
" " o " " VIIs~ i i ~
Fig. 8. Examples of reconstruction, in the parasagittal plane, of three TH-negative neurons labelled with Neurobiotin. Note that the dendrites were not drawn in the main reconstruction in B. The dendrites and cell body of this neuron are drawn separately in the boxed panel. The neuron shown in B was identified as bulbospinal on the basis of morphology. VII, facial nucleus; CF, compact formation of nucleus
ambiguus; SO, superior olive; IO, inferior olive; R, rostral; D, dorsal.

612 J. Lipski et al.
Only a low proportion of RVLM barosensitive neurons (6/34, 18%) were identified as bulbospinal (presympathetic). This contrasts with results of pre- vious extracellular studies in the rat in which 63-82% of barosensitive neurons located in this medullary region could be excited antidromically from the tho- racic segments of the spinal c o r d . 6'27'32 However, the difference between these results is only apparent, as in the present study most neurons were either not tested for spinal projection by antidromic stimu- lation, or the test was not conclusive. Five out of nine neurons which were fully tested responded antidrom- ically, which is the proportion similar to that de- scribed previously. When morphological criteria were used, spinal projection could be identified in cells injected with Neurobiotin when a large charge trans- fer (> 20 nA x min) and long post-injection survival times ( > 6 h) were used. In such experiments three out of five injected cells had spinally projecting axons.
All recordings were made ventral or ventromedial to compact formation of the nucleus ambiguus. This location corresponds well with previously published
maps based on extracellular studies in the rat (e.g., Refs 6, 34, 47, 78).
Relat ionship to C1 adrenergic neurons
The adrenergic (TH and PNMT-immunoreactive) neurons in the rostral portion of the CI cell column have long been implicated in cardiovascular control. The main arguments are as follows: (i) the s.c. "pressor area' in the RVLM, usually identified with microinjections of excitatory amino acids, overlaps with the rostral portion of the C1 cell column (e.g., Refs 3, 59); (ii) many TH (and PNMT)-immuno- reactive neurons located in this pressor area (62-80%) 3]'6~ have axons projecting to the spinal cord, where they form synapses mainly with pregan- glionic sympathetic neurons (e.g., Refs 4, 45, 58); (iii) application of viral tracers to peripheral autonomic structures results in transneuronal labelling of some C1 cells; 15~66 (iv) extracellularly recorded barosensitive and spinally projecting RVLM neurons are found in the immediate vicinity of rostral C1 neurons (e.g., Ref. 47); and (v) the activity of some RVLM
R ~
. . . . i i , o j
l m m
Fig. 9. Reconstruction, in the parasagittal plane, of a Neurobiotin-filled neuron which had atypical axon trajectory. The neuron was not examined for TH-immunoreactivity. The insets illustrate electrophysiologi- cal identification of this neuron. Traces in A show aortic nerve evoked potentials (top, six superimposed sweeps; bottom, 30 averaged sweeps). Asterisks indicate stimulus artefacts. A continuous hyperpolarizing current (-0.1 nA) was used to stop firing. (B) antidromic action potentials evoked by near threshold stimulation in T 2 segment (stimulus marked by asterisk, five superimposed sweeps). Note that spinal
stimulation was followed by antifromic action potentials only in three sweeps.

Properties of RVLM neurons 613
Fig. 10. Micrographs illustrating details of cellular morphology. (A) Cell body and proximal processes of a Lucifer Yellow-filled neuron which was TH negative. (B) Cell body and processes of a Neurobiotin- filled neuron which was TH positive (the same neuron as in Fig. 7C). Arrow indicates brush-like dendritic arborizations. (C, D) Collaterals issued by descending axonal branches. The origins of collaterals are indicated by arrows. (E) Descending axonal branch traced to the caudal end of the parasagittal section (arrow, C1 spinal segment). The branch was part of the neuron reconstructed in Fig. 7A. (F) bouton-like varicosities in the A5 region. (G) Varicosities in the dorsal vagal complex. Scale bar = 250 #m (A, B, E);
50#m (C, D, F, G).
presympathet ic neurons can be inhibi ted by intra- venous or microelec t rophoret ic admin i s t ra t ion of ~2 receptor agonists, consis tent with the view tha t only
adrenergic neurons in this medul lary region respond to these agonists. ~'23'26 Two addi t ional findings suggest tha t at least some of these CI neurons are

614 J. Lipski et al.
barosensitive: changes in blood pressure evoked by haemorrhage or pharmacological intervention influ- ence the catecholamine metabolism in the general area of the RVLM measured by in v ivo voltame- try,18'55 and such blood pressure changes induce c-Fos in a proportion of C1 neurons) 6'73
Our study provides a direct demonstration that some C1 neurons (including those which are spinally projecting) are inhibited by baroreceptor afferents. However, the percentage of double-labelled cells (26%) was lower than expected from the study by Allen and Guyenet, ~ who found that 380 of RVLM barosensitive neurons (mostly confirmed as bul- bospinal) were also inhibited by microelectrophoretic administration of clonidine or ~-methylnore- pinephrine. These authors concluded, on the basis of this and other evidence (low firing frequency and generally slow conduction velocity of spinal axons), that the neurons sensitive to these drugs may belong to the C1 adrenergic cell group.
The proportion of double-labelled cells found in our study was also lower than estimated based on counts of spinally projecting C1 cells. Previous ana- tomical studies showed that, in the rat, 45-72% of the neurons in the RVLM that were retrogradely labelled from the thoracic segments of the spinal cord also stained for TH o r P N M T . 27'29'61'73'76 Several factors can explain the discrepancy between the percentage of barosensitive TH-immunoreactive neurons found in our study, and previous estimates. First, a large proportion of all spinally projecting neurons in this region are non-adrenergic (28 to more than 500/0). 31'61
In spite of the fact that many putative non-adrenergic neurons receive an inhibitory baroreceptor input (a group corresponding to clonidine-insensitive neur- ons126), such cells were not included in estimates based on counts of spinal C1 cells. Second, some spinally projecting C1 neurons, even those located within the pressor area, may not be barosensitive. Consistent with this possibility is the observation that rostral C1 neurons are located not only in the pressor area, but also medial (or ventromedial) and caudal to this area (e.g., Refs 34, 61, 62), with anatomical boundaries not exactly overlapping the functionally- defined cardiovascular zone. 3~ In addition, some C1 neurons appear to project to non-cardiovascular re- gions of the spinal cord. 9'28 Third, as demonstrated in the present study, at least some barosensitive neurons which do not project to the spinal cord belong to the non-adrenergic group, and, therefore, would not be included in estimates based on anatomy and im- munocytochemistry. All these factors may have con- tributed to an overestimation of the proportion of C 1 neurons involved in control of vasomotor pregan- glionic sympathetic neurons and in mediating the baroreceptor reflex.
On the other hand, two methodological factors may have led to an underestimation of the proportion of adrenergic neurons found in our study. One possi- bility is that the impalements were biased towards the
cells which were non-adrenergic. Although the aver- age size of intracellularly labelled cell bodies (mean long axis, 26.0/~m) did not appear to be significantly different from the size of C1 neurons (long axis range, 18-32 pm62), another factor may have influenced the impalements. In this, and in a previous intracellular study in vivo, 41 n o neurons were found with spinal axons conducting less than 1.9 m/s. This contrasts with several extracellular studies which identified a subpopulation of RVLM presympathetic neurons with slower conducting, presumably unmyelinated (< 1.0 m/s;) axons (e.g., Refs 7, 47, 67). If all such neurons are adrenergic, then the failure of their impalement may have resulted in identifying fewer barosensitive neurons which are TH-positive. How- ever, in contrast to a previous study, 67 it has been demonstrated that most presympathetic neurons which are clonidine-sensitive and, therefore, pre- sumably belong to C1 group, have the range of conduction velocities approximately 1.0-4.0 m/s. In addition, Morrison e t al. 47 calculated, on the basis of the axonal diameter, that myelinated spinal PNMT- immtmoreactive axons conduct in the range 1.8 5.3 m/s. Such neurons are likely to be included in our sample.
Finally, the intracellular labels used in our previous study, particularly Lucifer Yellow, may have inter- fered with TH-immunoreactivity: '63 particularly that only two out of 15 Lucifer Yellow-filled neurons were double-labelled, while a higher proportion (5/12) was TH-positive when Neurobiotin was used. However, it has previously been demonstrated that Lucifer Yellow does not interfere with TH immunoreactivity in the locus coeruleus 68 or midbrain dopaminergic neurons) 9 In addition, Sun e t al. 68 reported that nine of 21 randomly sampled silent RVLM neurons, which were labelled with Lucifer Yellow, were also TH immunoreactive. Therefore, it is unlikely that interference by Lucifer Yellow affected our results.
Although our study has confirmed the presence of TH immunoreactivity in some RVLM barosensitive neurons, it is still not clear whether C1 neurons release adrenalin as the neurotransmitter, and if so, to what degree adrenaline controls the activity of preganglionic sympathetic neurons. The view that adrenaline may not be released from these neurons is supported by the study of Sved, 72 who failed to demonstrate the presence of this transmitter in the spina! cord, and by Rogers e t al., 57 who observed no change in blood pressure and no reduction in the magnitude of the pressor response to electrical stimu- lation within the RVLM despite reduction of central adrenalin content with a PNMT inhibitor (see also Ref. 12). In addition, it has been demonstrated that administration of catecholamine receptor antagonists produces relatively small reduction of the vasomotor tone, while blockade of excitatory amino acid recep- tors eliminates this tone entirely (e.g., Refs 36, 64). Further studies are necessary to elucidate the role of CI neurons in cardiovascular control.

Properties of RVLM neurons 615
Finally, these results should be discussed in the context of the study by Granata and Kitai, z° who intracellularly labelled RVLM neurons in anaes- thetized rats and attempted to assess their PNMT- immunoreactivity. No double-labelled cells were identified. However, their data cannot be compared with present results since no convincing evidence was presented that the neurons investigated in that study were barosensitive (see Ref. 40 for discussion).
M o r p h o l o g y
Several previous studies described morphology of putative presympathetic RVLM neurons in v i tro . In the early study by Sun et al. , 68 conducted with live tissue slices, the presence of regular, pacemaker-like activity was used as a criterion for identification of non-adrenergic neurons. However, it has been re- ported that adrenergic neurons also display similar type of activity. TM In addition, it remains to be demonstrated that pacemaker properties are ex- pressed in v i t ro only in the cardiovascular subset of RVLM neurons. In the study conducted by Huangfu et al. , 29 in fixed slices, the neurons analysed were shown to project to thoracic segments of the spinal cord. However, identification of spinal projection is not a sufficient criterion for classifying neurons as presympathetic (sympathoexcitatory) for two reasons. First, it has been reported that in in v ivo rats some RVLM neurons (particularly with faster con- ducting axons) could be excited antidromically from the spinal cord but did not respond to stimulation of baroreceptor afferents. 34 Second, in anaesthetized animals many RVLM bulbospinal neurons are si- lent, ~°'47 and therefore could not be tested for inhibi- tory baroreceptor input. It remains to be demonstrated whether such neurons are 'presympa- thetic'.
Our results demonstrate that RVLM neurons identified as barosensitive are heterogenous not only with respect to TH-immunoreactivity but also the morphology. The differences included the shape and size of the cell body, presence or absence of spinal projection (see above for discussion), rostral axonal projections within the brainstem, and the presence of axon collaterals within the medulla oblongata. How- ever, due to the small size of the sample analysed and large variability between individual cells, no firm conclusion could be drawn regarding the correlation between the morphology and the presence (or ab- sence) of adrenergic phenotype.
Most of the filled neurons had ventrally projecting dendrites that reached the ventral medullary surface. This feature was observed both in the TH-positive (see also Ref. 3) and TH-negative cells. Because the intracellular technique only allows a limited sampling, it is possible that there is an extensive dendritic plexus belonging to these neurons at and near the ventral medullary surface. Another promi- nent feature was the dorsomedial projection of stem axons (see also Ref. 70). Transtegmental projection of
axons towards the principal adrenergic bundle in the dorsomedial medulla was described in previous ana- tomical studies of C1 neurons. 58'6°'62 Benarroch et al., 3
reported that electrolytic lesions in the dorsomedial segmentum largely reduced pressor responses evoked by application of L-glutamate to the ipsilateral RVLM, and ascribed these effects to interruption of C1 axons (see also Ref. 17). However, our results show that some axons in the transegmental tract belong to TH-negative neurons and, therefore, the effects of these lesions can also be due to transections of axons of non-adrenergic neurons. Sun et al., 7°
hypothesized that transegmental axons of RVLM neurons bifurcate in the dorsomedial tegmentum into major rostral and caudal branches. Such bifurcations have indeed been demonstrated in our material. However, due to the small sample of neurons analysed, we were unable to establish whether the barosensitive RVLM neurons with such bifurcating axons mainly belong to the C1 group as proposed by Haselton and Guyenet. 27
Rostral projections of axons of functionally unidentified RVLM neurons have been described in a large number of anterograde and/or retrograde tracing studies. Most of these studies concentrated on projections of C1 neurons, a subset which could be easily identified immunocytochemically. The putative targets included the locus coeruleus (e.g., Refs 24, 52, 77), A5 region, 49 dorsolateral pons and periaqueduc- tal gray), 27"v7 hypothalamus 49'76 paraventricular thal- amic nucleus sl and forebrain limbic structures. 33's6'8° Many of these rostrally projecting C1 neurons, poss- ibly with the exception of cells projecting to the hypothalamus 76 and locus coeruleus, 3° also project to the spinal cord. 26'27 In addition, several extracellular antidromic stimulation studies have demonstrated the projection of neurons identified as barosensitive to some of these reg ions . 26"2v'3° In our study, the rostrally projecting branches could only be traced for a limited distance and, therefore, our data cannot be compared with these results (apart from confirming projection to the ventrolateral pontomedullary bor- der/A5 region). On the other hand, our intracellular approach demonstrated axons and axon collaterals within the medulla oblongata at relatively short distances from cell bodies, the projection which can- not be easily demonstrated with antidromic mapping due to the current spread. Medullary branches were identified in three main regions: in the rostral ventro- lateral medulla including the adjacent part of the lateral tegmental field, in the dorsomedial medulla (in the dorsal vagal complex) and in the caudal ventrolat- eral medulla. These projections are similar to those reported by Nicholas and Hancock 5° who made small deposits of an anterograde tracer in the RVLM. However, in contrast to that study, 5° no contralateral projections were found. One explanation for this discrepancy is that the anterograde tracer was also taken up by neurons which are not barosensitive, for example the expiratory neurons on the B6tzinger

616 J. Lipski et al.
complex. 34 A recent intracellular- labell ing s tudy from our l abora to ry revealed an extensive axonal projec- t ion of B6tzinger neurons across the midline. 8 The pa thway f rom the R V L M to the dorsal vagal complex (presumably to the nucleus of the solitary tract) includes project ion from CI neurons, as demon- s t rated in our s tudy and the ana tomica l study by Thor and Helke 75 (see also Ref. 77). The exact synaptic targets in the medul la ob longa ta of barosen- sitive R V L M neurons remain to be identified, a l though some of these targets may include catechol- aminergic and serotoninergic n e u r o n s ) °
ons are adrenergic. However, the p ropor t ion of double- label led cells was lower than previous esti- mates and, therefore, our results are consis tent with the view that the non-catecholaminergic R V L M neurons also play an impor t an t role in vasomotor control . The use of intracel lular labels revealed sev- eral new features of these neurons, including the presence of axon collaterals and bouton- l ike varicosi- ties in several b ra ins tem regions. Therefore, the func- t ion of barosensi t ive R V L M neurons extends beyond their role in exciting pregangl ionic sympathet ic neur- ons (cf. also Ref. 27).
CONCLUSION
These experiments provide for the first t ime a direct demons t r a t i on tha t some barosensi t ive R V L M neur-
Acknowledgements This study was supported by grants from the New Zealand Health Research Council and the New Zealand Lottery Grants Board. We thank Lisa Christ- mas for secretarial assistance.
REFERENCES
1. Allen A. M. and Guyenet P. G. (1993) c¢2-Adrenoceptor-mediated inhibition of bulbospinal barosensitive cells of rat rostral medulla. Am. J. Physiol. 265, R1065 1075.
2. Andrezik T. A., Chan-Palay V. and Palay S. L. (1981) The nucleus paragigantocellularis lateralis in the rat. Confirmation and cytology. Anat. Embryol. 161, 355 371.
3. Benarroch E. E., Granata A. R., Ruggiero D. A., Park D. H. and Reis D. J. (1986) Neurons of CI area mediate cardiovascular responses initiated from ventral medullary surface. Am. J. Physiol. 250, R932 945.
4. Bernstein-Goral H. and Bohn M. C. (1989) Phenylethanolamine N-methyltransferase-immunoreactive terminals synapse on adrenal preganglionic neurons in the rat spinal cord. Neuroscience 32, 521 537.
5. Bornstein J. C., Costa M. Furness J. B. and Lees G. M. (1984) Electrophysiological and enkephalin immunoreactivity of identified myenteric plexus neurons of guinea-pig small intestine. J. Physiol. 351, 313 325.
6. Brown D. L. and Guyenet P. G. (1984) Cardiovascular neurons of brain stem with projections to the spinal cord. Am. J. Physiol. 247, R1009-I016.
7. Brown D. L. and Guyenet P. G. (1985) Electrophysiological study of cardiovascular neurons in the rostral ventrolateral medulla in rats. Circulation Res. 56, 359-369.
8. Bryant T. H., Yoshida S., de Castro D. and Lipski J. (1993) Expiratory neurons of the B6tzinger complex in the rat: a morphological study following intracellular labelling with biocytin. J. comp. Neurol. 335, 267 282.
9. Carlton S. M., Honda C. N., Willcockson W. S., Lacrampe M., Zhang D., Denoroy L., Chung J. M. and Willis W. D. (1991) Descending adrenergic input to the primate spinal cord and its possible role in modulation of spinothalamic cells. Brain Res. 543, 77-90.
10. Caverson M. M., Ciriello J. and Calaresu F. R. (1983) Direct pathway from cardiovascular neurons in the ventrolateral medulla to the region of the intermediolateral nucleus of the upper thoracic cord: an anatomical and electrophysiological investigation in the cat. J. auton, nerv. Syst. 9, 451475.
I1. Chalmers J., Arnolada L., Kapoor V., Llewellyn-Smith I., Minson J. and Pilowsky P. E. (1992) Amino acid neurotransmitters in the central control of blood pressure and in experimental hypertension. J. Hypertens. 10, $27-$37.
12. Connor H. E. and Drew G. M. (1987) Do adrenaline-containing neurons from the rostral ventrolateral medulla excite preganglionic sympathetic cell bodies? J. auton. Pharmac. 7, 87-96.
13. Dampney R. A. L. (1994) The subretrofacial vasomotor nucleus: anatomical, chemical and pharmacological properties, and role in cardiovascular regulation. Prog. Neurobiol. 42, 197~27.
14. Dembowsky K. and McAllen R. M. (1990). Baroreceptor inhibition of subretrofacial neurons: evidence from intracellular recordings in the cat. Neurosci. Lett. 111, 139-143.
15. Ding Z.-Q., Li Y.-W., Wesselingh S. L. and Blessing W. W. (1993) Transneuronal labelling of neurons in rabbit brain after injection of Herpes simplex virus type 1 into the renal nerve. J. auton, herr. Syst. 42, 23-32.
16. Dun N. J., Dun S. L. and Chiaia N. L. (1993) Hemorrhage induces Fos immunoreactivity in rat medullary catecholaminergic neurons. Brain Res. 608, 223-232.
17. Farlow D. M., Goodchild A. K. and Dampney R. A. L. (1984) Evidence that vasomotor neurons in the rostral ventrolateral medulla project to the spinal sympathetic outflow via the dorsomedial pressor area. Brain Res. 298, 313-320.
18. Gillon J.-Y., Richard F., Quintin L., Pujol J.-F. and Renaud B. (1990) Pharmacological and functional evidence for extracellular 3,4-dihydroxyphenylacetic acid as an index of metabolic activity of adrenergic neurons: an in vivo voltammetry study in the rat rostral ventrolateral medulla. Neuroscience 37, 421~430.
19. Grace A. A. and Onn S. P. (1989) Morphology and electrophysiological properties of immunocytochemically identified rat dopamine neurons recorded in vitro. J. Neurosci. 9, 3463 3481.
20. Granata A. R. and Kitai S. T. (1992) Intracellular analysis in vivo of different barosensitive bulbospinal neurons in the rat rostral ventrolateral medulla. J. Neurosci. 12, 1 20.
21. Guyenet P. G. (1990) Role of ventral medulla oblongata in blood pressure regulation. In Central Regulation of Autonomic Functions (eds Loewy A. D. and Spyer K. M.), pp. 145 167. Oxford University Press, New York.
22. Guyenet P. G., Koshiya N., Huangfu D., Baraban S. C., Stornetta R. L. and Li Y-W. (1995) Role of medulla oblongata in generation of sympathetic and vagal outflows. Prog. Brain Res. (in press).

Properties of RVLM neurons 617
23. Guyenet P. G., Stornetta R. L., Riley T., Norton F. R., Rosin D. L. and Lynch K. R. (1994) Alpha2A-adrenergic receptors are present in lower brainstem catecholaminergic and serotonergic neurons innervating spinal cord. Brain Res. 638, 285 294.
24. Guyenet P. G. and Young B. S. (1987) Projections of nucleus paragigantocellularis lateralis to locus coeruleus and other structures in the rat. Brain Res. 406, 171-184.
25. Halliday G. M. and McLachlan E. M. (1991) A comparative analysis of neurons containing catecholamine-synthesizing enzymes and neuropeptide Y in the ventrolateral medulla of rats, guinea-pigs and cats. Neuroscience 43, 531-550.
26. Haselton J. R. and Guyenet P. G. (1989) Electrophysiological characterization of putative CI adrenergic neurons in the rat. Neuroscience 30, 199-214.
27. Haselton J. R. and Guyenet P. G. (1990) Ascending collaterals of medullary barosensitive neurons and C1 cells in rats. Am. J. Physiol. 258, R1051-1063.
28. H6kfelt T., Fuxe K., Goldstein M. and Johansson O. (1974) Immunohistochemical evidence for the existence of adrenaline neurons in the rat brain. Brain Res. 66, 235-251.
29. Huangfu D., Goodwin W. B., Alheid G. F., Stornetta R. L. and Guyenet P. G. (1994) Morphology of identified bulbospinal neurons in the rostral ventrolateral medulla (RVLM) of the rat. Soc. Neurosci. Abstr. 20, 1182.
30. Huangfu D., Verberne A. J. M. and Guyenet P. G. (1992) Rostral ventrolateral medullary neurons projecting to locus coeruleus have cardiorespiratory inputs. Brain Res. 598, 67 75.
31. Jeske I. and McKenna K. E. (1992) Quantitative analysis of bulbospinal projections from the rostral ventrolateral medulla: contribution of CI adrenergic and nonadrenergic neurons. J. comp. Neurol. 324, 1 13.
32. Jeske I., Morrison S. F., Cravo S. L. and Reis D. J. (1993) Identification of baroreceptor reflex interneurons in the caudal ventrolateral medulla. Am. J. Physiol. 264, R169-178.
33. Jin G. R., Rao Z. R. and Shi J. W. (1994) Visceral noxious stimulation induced expression of Fos protein in medullary catecholaminergic neurons projecting to nucleus accumbens in the rat: a study with triple labeling method of HRP tracing combined with Fos and TH immunohistochemistry. Brain Res. 648, 196-202.
33a. Kangrga I. M. and Loewy A. D. (1995) Whole-cell recordings from visualized CI adrenergic bulbospinal neurons: ionic mechanisms underlying vasomotor tone. Brain Res. 670, 215-232.
34. Kanjhan R., Lipski J., Kruszewska B. and Rong W. A comparative study of presympathetic and B6tzinger respiratory neurons in the rostral ventrolateral medulla (RVLM) of the rat. Brain Res. (in press).
35. Kita H. and Amstrong W. (1991) A biotin containing compound N-(2-aminoethyl) biotinamide for intracellular labelling and neuronal tracing studies: Comparison with biocytin. J. Neurosci. Meth. 37, 141-150.
36. Koshiya N. and Guyenet P. G. (1994) A5 noradrenergic neurons and the carotid sympathetic chemoreflex. Am. J. Physiol. 267; R519-526.
37. Kruszewska B., Lipski J. and Kanjhan R. (1994) An electrophysiological and morphological study of esophageal motoneurons in rats. Am. J. Physiol. 266, R622-632.
38. Kumada M., Terui N. and Kuwaki T. (1990) Arterial baroreceptor reflex: Its central and peripheral neural mechanisms. Prog. Neurobiol. 35, 331-361.
39. Li Y.-W., Gieroba Z. J., McAllen R. M. and Blessing W. W. (1991) Neurons in rabbit caudal ventrolateral medulla inhibit bulbospinal barosensitive neurons in rostral medulla. Am. J. Physiol. 261, R44~51.
40. Lipski J., Kanjhan R., Kruszewska B. and Rong W. (1995) Criteria for intracellular identification of pre-sympathetic neurons in the rostral ventrolateral medulla in the rat. Clin. exp. Ityperten. 17, 51~5.
41. Lipski J., Kanjhan R., Kruszewska B. and Rong W. Properties of presympathetic neurons in the rostral ventrolateral medulla in the rat: an intracellular study in vivo. J. Physiol. (in press).
42. Lipski J., Zhang X., Kruszewska B. and Kanjhan R. (1994) Morphological study of long axonal projections of ventral medullary inspiratory neurons in the rat. Brain Res. 640, 171-184.
43. Matsumato M., Takayama K. and Miura M. (1994) Distribution of glutamate- and GABA- immunoreactive neurons projecting to the vasomotor center of the intermediolateral nucleus of the lower thoracic cord of Wistar rats: a double labelling study. Neurosci. Lett. 174, 165-168.
44. McAllen R. M. (1987) Central respiratory modulation of subretrofacial bulbospinal neurons in the cat. J. Physiol. 388, 533-545.
45. Milner T. A., Morrison S. F., Abate C. and Reis D. J. (1988) Phenylethanolamine N-methyltransferase containing terminals synapse directly on sympathetic preganglionic neurons in the rat. Brain Res. 448, 205-222.
46. Morrison S. F., Callaway J., Milner T. A. and Reis D. J. (199l) RVLM: A source of the glutamatergic innervation of the sympathetic IML nucleus. Brain Res. 562, 126 135.
47. Morrison, S. F., Milner, T. A. and Reis D. J. (1988). Reticulospinal vasomotor neurons of the rat rostral ventrolateral medulla: relationship to sympathetic nerve activity and C1 adrenergic cell group. J. Neurosci. 8, 1286-1301.
48. Morrison S. F. and Reis D. J. (1991) Responses of sympathetic preganglionic neurons to rostral ventrolateral medullary stimulation. Am. J. Physiol. 261, R1247-1256.
49. Nicholas A. P. and Hancock M. B. (1989) Evidence for substance P, serotonin and oxytocin input to medullary catecholamine neurons with diencephalic projections. Brain Res. Bull. 22, 213-223.
50. Nicholas A. P. and Hancock M. B. (1991) Projections from the rostral ventrolateral medulla to brainstem monoamine neurons in the rat. Neurosci. Lett. 122, 91 95.
51. Phillipson O. T. and Bohn M. C. (1994) CI-3 adrenergic medullary neurons project to the paraventricular thalamic nucleus in the rat. Neurosci. Lett. 176, 67 70.
52. Pieribone V. A., Aston-Jones G. and Bohn M. C. (1988) Adrenergic and non-adrenergic neurons in the C1 and C3 areas project to locus coeruleus: a fluorescent double labeling study. Neurosci. Lett. 85, 297-303.
53. Pilowsky P. M., Lipski J., Prestige R. and Jiang C. (1991) Dual fluorescence combined with a two-color immuno- peroxidase technique: a new way of visualizing diverse neuronal elements. J. Neurosei. Meth. 36, 185 193.
54. Reis D. J., Ruggiero D. A. and Morrison S. F. (1989) The C1 area of the rostral ventrolateral medulla oblongata. A critical brainstem region for control of resting and reflex integration of arterial pressure. Am. J. Hypertens. 2, 363S 374.
55. Rentero N., Kitahama K. and Quintin L. (1993) Baroreceptor reflex-linked changes in catechol metabolism in the rat rostral ventrolateral medulla. J. Physiol. 469, 717-737.
56. Roder S. and Ciriello J. (1993) Innervation of the amygdaloid complex by catecholaminergic cell groups of the ventrolateral medulla. J. comp. Neurol. 332, 105-122.

618 J. Lipski et al.
57. Rogers P. F., Head G. A., Lungershausen Y. K. and Howe P. R. C. (1991) Effects of depleting central and peripheral adrenaline stores on blood pressure in stroke-prone spontaneously hypertensive rats. J. auton, nerv. Syst. 34, 9-16.
58. Ross C. A., Ruggiero D. A., Joh T. H., Park D. H. and Reis D. J. (1984) Rostral ventrolateral medulla: Selective projections to the thoracic autonomic cell column from the region containing CI adrenaline neurons. J. comp. Neurol. 228, 168 185.
59. Ross C. A., Ruggiero D. A., Park D. H., Joh T. H., Sved A. F., Fernandez-Fardal J., Saavedra J. M. and Reis D. J. (1984) Tonic vasomotor control by the rostral ventrolateral medulla: Effect of electrical or chemical stimulation of the area containing C1 adrenaline neurons on arterial pressure, heart rate, and plasma catecholamines and vasopressin. J. Neurosei. 4, 473-494.
60. Ruggiero D. A., Cravo S. L., Arango V. and Reis D. J. (1989) Central control of the circulation by the rostral ventrolateral reticular nucleus: anatomical substrates. Prog. Brain Res. 81, 49-79.
61. Ruggiero D. A., Cravo S. L., Golanov E., Gomez R., Anwar M. and Reis D. J. (1994) Adrenergic and non-adrenergic spinal projections of a cardiovascularly-active pressor area of medulla oblongata: quantitative topographic analysis. Brain Res. 663, 107 120,
62. Ruggiero D. A., Ross C. A., Anwar M., Park D. H., Joh T. H. and Reis D. J. (1985) Distribution of neurons containing phenylethanolamine N-methyltransferase in medulla and hypothalamus of rat. J. comp. Neurol. 239, 127±154.
63. Scharfman H. E., Kunkel D. D. and Schwartzkroin P. A. (1989) Intracellular dyes mask immunoreactivity of hippocampal interneurons. Neurosci. Lett. 96, 23-28.
64. Shi H., Lewis D. I. and Coote J. H. (1988) Effects of activating spinal alpha°adrenoreceptors on sympathetic nerve activity in the rat. J. auton, nerv. Syst. 23, 69-78.
65. Stewart W. W. (1981) Lucifer dyes--highly fluorescent dyes for biological tracing. Nature 292, 1751. 66. Strack A. M., Sawyer W. B., Platt K. B. and Loewy A. D. (1989) CNS cell groups regulating the sympathetic outflow
to adrenal gland as revealed by transneuronal cell body labelling with pseudorabies virus. Brain Res. 491,274-296. 67. Sun M.-K. and Guyenet P. G. (1986) Effect of clonidine and 7-aminobutyric acid on the discharge of medullo-spinal
sympathetic neurons in the rat. Brain Res. 368, 1-17. 68. Sun M.-K., Young B. S., Hackett J. T. and Guyenet P. G. (1988) Rostral ventrolateral medullary neurons with intrinsic
pacemaker properties are not catecholaminergic. Brain Res. 451, 345-349. 69. Sun M.-K. and Reis D. J. (1994) Hypoxia activates calcium currents in the reticulospinal vasomotor pacemaker neurons
of rat rostral ventrolateral medulla: An intracellular study in vivo. Soc. Neurosci. Abstr. 20, 842. 70. Sun M.-K., Stornetta R. L. and Guyenet P. G. (1991) Morphology of rostral medullary neurons with intrinsic
pacemaker activity in the rat. Brain Res. 556, 61-70. 71. Sundaram K. and Sapru H. (1991) NMDA receptors in the intermediolateral column of the spinal cord mediate
sympathoexcitatory cardiac responses elicited from the ventrolateral medullary pressor area. Brain Res. 544, 33-41. 72. Sved A. F. (1989) PNMT-containing catecholaminergic neurons are not necessarily adrenergic. Brain Res. 481, 113 118. 73. Sved A. F., Mancini D. L., Graham J. C., Schreihofer A. M. and Hoffman G. E. (1994) PNMT-containing neurons
of the C1 cell group express c-fos in response to changes in baroreceptor input. Am. J. Physiol. 266, R361 367. 74. Terui N., Saeki Y. and Kumada M. (1986) Barosensory neurons in the ventrolateral medulla in rabbits and their
responses to various afferent inputs from peripheral and central sources. Jap. J. Physiol. 36, 1141 1164. 75. Thor K. B. and Helke C. J. (1988) Catecholamine-synthesizing neuronal projections to the nucleus tractus solitarii of
the rat. J. comp. Neurol. 268, 264-280. 76. Tucker D. C., Saper C. B., Ruggiero D. A. and Reis D. J. (1987) Organization of central adrenergic pathways I.
Relationship ofventrolateral medullary projections to the hypothalamus and spinal cord. J. comp. Neurol. 259, 591q503. 77. Van Bockstaele E. J. and Aston-Jones G. (1992) Collateralized projections from neurons in the rostral ventrolateral
medulla to the nucleus of the solitary tract and the periaqueductal gray using double retrograde labeling technique in the rat. Neuroscience 49, 653~68.
78. Wang W. H. and Lovick T. A. (1992) Inhibitory serotoninergic effects on rostral ventrolateral medullary neurons. Pfliigers Arch. 422, 93 97.
79. Zagon A. and Spyer K. M. (1994) Projections from the nucleus of solitary tract towards identified and intracellularly labelled 'sympatho-excitatory' cells of the rostral ventrolateral medulla oblongata (RVLM) in rats. Soc. Neurosci. Abstr. 20, 302.
80. Zagon A., Totterdell S. and Jones R. S. G. (1994) Direct projections from the ventrolateral medulla oblongata to the limbic forebrain: anterograde and retrograde tract tracing studies in the rat. J. eomp. Neurol. 340, 445-468.
(Accepted 5 June 1995)





























