Embed Size (px)
Citation preview

Vol. 33, No. 1INFECTION AND IMMUNITY, July 1981, p. 22-280019-9567/81/070022-07$02.00/0
Complement-Fixing Antibody to the AG-4 Antigen in HerpesSimplex Virus Type 2-Infected Patients
MINAS ARSENAKIS AND JOHN T. MAY*
Microbiology Department, La Trobe University, Bundoora, Victoria 3083 Australia
Received 18 December 1980/Accepted 2 April 1981
Sera collected from confirmed herpes simplex virus type 2 (HSV-2) patientswere found to be devoid of complement-fixing antibody to the AG-4 antigen atthe time of the herpes lesion outbreak in 10 out of 13 cases. However, 1 to 4weeks after HSV-2 lesion appearance, 28 out of 30 patients acquired complement-fixing antibody to the AG-4 antigen. The sera of these patients containedimmunoglobulin M antibody activity and the ability to immunoprecipitate a
160,000-molecular weight early HSV-2 antigen (the AG-4 antigen). Also, thesesera were used to show that a variety of anti-herpes virus compounds had anegligible effect on AG-4 production in HSV-2-infected HEp-2 cells. The majorityof the compounds tested (including acycloguanosine and phosphonoformate) are
known to inhibit late antigen production, suggesting that the AG-4 antigen is anearly antigen. It is probably an immediate early antigen (a) as it is formed in thepresence of cycloheximide and actinomycin D, a treatment which is used toaccumulate a proteins.
Several early antigens prepared from herpessimplex virus type 2 (HSV-2)-infected cells havebeen used in seroepidemiological studies in cer-vical carcinoma (CaCx) patients (12). One suchantigen, designated AG-4, has been found to betype specific and tumor specific (for CaCx; [2]).Since that initial study, several other studieshave supported the association of the presenceof AG-4 antibodies and CaCx (1, 5, 7, 10). More-over, AG-4 antibodies were found to persist inCaCx patients even after treatment (1, 5, 10).We have previously found that HSV-2-infectedpatients also possessed antibodies to the AG-4antigen (1). This fmding suggested that the AG-4 antigen is HSV-2-type specific but notuniquely associated with CaCx.
In the study described here we have furtherinvestigated the AG-4 antibody present in HSV-2-infected patients and identified when such an-tibody appears in the course ofHSV-2 infections.This antibody has also been used to evaluate theeffect of some anti-herpes virus compounds onthe production of the AG-4 antigen in HEp-2cells. To confirm the identity ofthe AG-4 proteindescribed by Strnad and Aurelian (15) with theprotein reacting with HSV-2 patients' sera, wehave used CaCx sera as positive controls in someparts of this study.
MATERIALS AND METHODSCells and viruse. Cells and viruses used were
described previously (1). The HSV-2G strain (15) wasobtained from L. Aurelian (School of Medicine, Johns
Hopkins University, Baltimore, Md.), who originallyobtained it from B. Roizman (15). HSV-2G was prop-agated and titrated by plaque assay on HEp-2 cells aspreviously described (16). HSV-2G was used through-out this study mainly for its ability to consistentlygenerate 20- to 50-fold higher virus titer (4 x 107 to 10X 107 plaque-forming units [PFU] per ml) than ourlocal strains of HSV-2 in HEp-2 cells (1).Sera and patient groups. The HSV-2 and HSV-
1 patients' sera included in this study were obtainedfrom patients attending the La Trobe UniversityHealth Centre or the Communicable Diseases Centre,Melbourne, Australia. All patients were confirmed asHSV-1 or HSV-2 infected by virus isolation and neu-tralization typing as previously described (8). Serawere collected from the patients at the time of lesionappearance or recurrence, and at various time inter-vals thereafter. Also included in this study were serafrom CaCx patients with confirmed invasive squa-mous-cell carcinoma of the uterine cervix attendingthe public wards of the Royal Women's Hospital inMelbourne (1). The normal control sera were collectedfrom similar age patients without HSV-2 infections orCaCx. Sera were stored at -20°C until needed.AG-4 antigen preparation. As described previ-
ously, HEp-2 cells were infected at a multiplicity ofinfection (MOI) of 0.7 PFU/cell. After 1 h for virusadsorption and 4 h of infection, the cells were scrapedoff, washed three times in Barbitone buffered saline(BBS), and suspended in 0.7 ml of BBS per 1 x 107cells. The cells were frozen and thawed four times, andthe supernatant was collected and kept at -70°C untilneeded. Control antigen (AG-H) was prepared in thesame way with mock-infected HEp-2 cells. Proteinconcentration in both AG-4 and AG-H preparationswas adjusted to 2.0 mg/ml for use in the complementfixation test (1). All antigen preparations were tested
22

VOL. 33, 1981
for AG-4 antigenicity with standard AG-4 positive andnegative sera before use.Complement fixation test. The complement fix-
ation test was previously described (1).Preparation of radiolabeled AG-4 antigen.
Confluent monolayers of HEp-2 cells in plastic 150-cm2 tissue culture flasks (Corning Glass Works, Corn-ing, NY) were infected at an MOI of 20 PFU/cell inthe presence of 50 ,Lg of cycloheximide per ml. Afteradsorption of the virus for 1 h at 37°C, the virusinoculum was removed, and the cells were overlaidwith Eagle minimal essential medium (EMEM) con-taining 1% newborn calf serum, 100 jg of neomycin perml and 50 ltg of cycloheximide per ml. After 8 h ofincubation at 37°C, the cycloheximide-containing me-dia were removed, the cells were washed twice withphosphate-buffered saline, and then overlaid with 20ml of EMEM containing 1% dialyzed fetal calf serum,100 j&g of neomycin per ml, 1/20 the normal concen-tration of L-methionine, 10 ,uCi of L-[3S]methionineper ml (>600 Ci/mmol; Radiochemical Centre, Amer-sham, England) and 10 yg of actinomycin D per ml.The cells were then incubated for 6 h at 37°C, afterwhich time the media were discarded, and the cellswere harvested by scraping and washed three times inphosphate-buffered saline; the final cell pellet wasthen suspended in 0.7 ml of BBS per 1 x 107 cells. Thecells were then frozen and thawed four times, and celldebris was removed by centrifugation at 12,000 x g for20 min, and the supernatant was stored at -70°C untilneeded. Control uninfected antigen (AG-H) was pre-pared in the same way with mock-infected HEp-2cells. Although this method is essentially that used toprepare the immediate early (a) proteins (11), thepreparation contains the AG-4 antigen as detected bycomplement fixation.
Immunoprecipitations of radiolabeled anti-gens with HSV-2, CaCx sera, and normal controlsera. Essentially we followed the method of Strnadand Aurelian (15) with the following modifications.Sera to be used in immunoprecipitation experiments(with 1:8 complement-fixing antibody) were absorbedtwice with HEp-2 cells (5 x 106 cells/0.2 ml of serum)for 1 h at 37°C. After absorption they were adsorbedtwice with 0.2 ml of packed sheep erythrocytes per 0.2ml of serum at 37°C for 1 h, and finally they werecleared of any particulate matter by centrifugation at50,000 x g for 1 h in a A-100 rotor in an Airfuge(Beckman Instruments, Inc., Fullerton, Calif). Afterthis serum pretreatment, 0.15 ml of serum was mixedwith 0.15 ml of 3S-radiolabeled antigen, and the mix-ture was incubated at 4°C for 16 h. The immunopre-cipitates were sedimented by centrifugation at 130,000x g for 1 h in a A-100 rotor in an Airfuge (Beckman).The resulting pellets were washed twice with 0.15-mlvolumes of BBS, and the final pellets were stored at-70°C until needed for electrophoresis.Polyacrylamide gel electrophoresis. Samples
for polyacrylamide gel electrophoresis were denaturedin 1% sodium dodecyl sulfate-5% 2-mercaptoethanol-50 mM tris(hydroxymethyl)aminomethane-hydro-chloride buffer (pH 7.0) by heating at 80°C for 10 min.The denatured samples were loaded and run on a 7.5%continuous slab (1.2 mm thick by 12 cm deep by 13 cmwide) polyacrylamide gel (9) for 6 h at 5 V/cm (60
AG-4 ANTIBODY IN HSV-2 PATIENTS 23
mA). The gels were dried and autoradiographed usingKodak X-Omat RP X-ray film. The molecular weightsof labeled proteins were determined from their migra-tion relative to unlabeled molecular weight markers.
Microneutralization. Microneutralization ofHSV-2G was performed with patients' sera essentiallyas described previously (13), except that 100 PFU ofHSV-2G and 5 x 103 HEp-2 cells were used per micro-titer plate well (Flow Laboratories, Sydney, Australia).Serum fractionation. Human sera were heat in-
activated at 56°C for 30 min, diluted 1:1 in BBS, and0.3-ml samples were layered on 10 to 40% (wt/vol)linear sucrose gradients in phosphate-buffered salineand centrifuged at 37,000 rpm for 16 h in a SpincoSW50.1 rotor. Gradients were fractionated from thebottom of the tube in 0.3-ml fractions and monitoredfor optical density at 280 nm. Fractions were testedfor immunoglobulin (IgM) and IgG content and quan-titated by immunodiffusion with S-Partigen-IgM andTri-Partigen-IgG plates (Behringwerke A. G. Mar-burg, West Germany). Samples were dialyzed againstBBS before they were tested for AG-4 complement-fixing activity.AG-4 production with antiviral compounds.
Phosphonoacetic acid was a gift from A. R. Geiszler(Abbott Laboratories, North Chicago, Il). Phosphon-oformate was the gift of B. Oberg (Astra-LakemedalA. D., Sodertalje, Sweden). Arildone was a gift fromF. Pancic (Sterling-Winthrop Research Institute,Rensselaer, N.Y.). Acycloguanosine (acyclovir) was agift from G. B. Elion (Burroughs Weilcome Co., Re-search Triangle Park, N.C.). 5'-Iododeoxyuridine, tri-fluorothymidine, 2'-deoxyglucose, adenine arabino-side, retinol, and retinoic acid were obtained fromSigma Chemical Co. (St. Louis, Mo.). Stock solutionsof all compounds were made in distilled water exceptfor Arildone, which was dissolved in 100% dimethylsulfoxide, and retinol and retinoic acid, which weredissolved in 95% ethanol. These solvents at the con-centrations used had no effect on AG-4 antigen pro-duction.
For AG-4 production the compounds were added atthe time of HSV-2G virus infection (and at the con-centration indicated in Table 1) and remained presentthroughout the 4-h infection period. AG-4 was pro-duced in HEp-2 cells as previously described.
RESULTSTime-course development of AG-4 anti-
body in HSV-2-infected patients. We previ-ously reported (1) the detection ofAG-4 comple-ment-fixing antibodies in 15 out of 17 HSV-2-infected-patients' sera. We have now extendedthose initial observations by following the de-velopment of the AG-4 antibody response inHSV-2 infections and particularly recurrent in-fections. Blood samples were taken, when pos-sible, from the patients, first at the time ofpresenting with genital herpes lesions and atintervals thereafter; virus samples were alsotaken from the lesion at presentation for confir-mation of virus type. The levels of AG-4 com-plement-fixing antibodies in four patients with

24 ARSENAKIS AND MAY
recurrent HSV-2 infections are illustrated in Fig.1. In three of the four patients, the titer of thepresentation serum was less than 1:4. Subse-quent sera from the three patients taken atvarious times after lesion appearance exhibiteda pattern of rising AG-4 antibody titer as earlyas 4 days after lesion appearance. The fourthpatient had a titer of more than 1:4 at presen-tation and remained at the same level for 2weeks before declining; however, it is of interestto note that this particular patient experiencedrecurrent HSV-2 infections every 3 to 4 weeks.One ofthe four patients was followed for a periodof almost 1 year, and the AG-4 antibody titer,which was initially more than 1:4, fell to lessthan 1:4, during which period another HSV-2recurrence occurred in this same patient and theAG-4 antibody titer rose again 4 days after lesionrecurrence. The complement-fixing AG-4 anti-body was associated with the IgM fraction ofsera up to 4 weeks after lesion recurrence. In thecase of the high titer, 1:36 serum activity wasassociated with the IgG fraction also. Neutral-izing antibody to HSV-2 in three patients withrising antibody to AG-4 was monitored by mi-croneutralization. Over the same period of timethat the AG-4 complement-fixing antibody titer
rose, the HSV-2 neutalizing antibody also in-creased two- to fourfold in all three patients (13).Although we have only presented the AG-4
antibody response profiles of four patients inFig. 1, we also tested sera, at various times afterlesion appearance, from other patients in whichHSV-2 genital infection was confirmed. Serafrom 13 patients with HSV-2 infections (bothprimary and recurrent) were tested for AG-4antibody at the time ofpresentation with lesions,and only three were positive (titer >1:4) for AG-4 complement-fixing antibody. One of thesethree positive patients was a high-frequency-re-currence patient included in Fig. 1. Also, as theseblood samples were taken on presentation of thepatient (at the clinic or the Health Centre) withlesions (either primary or recurrent), delay inpresentation of 4 days or more after lesions firstappear would see the appearance of AG-4 anti-body in the sera. Thirty patients (both primaryand recurrent infections) were tested 1 to 4weeks after lesion appearance, and all but twoof these samples were positive (titer >1:4) forAG-4 complement-fixing antibody.Thus, it would appear that HSV-2-infected
patients can produce AG-4 complement-fixngantibody, and its detection by complement fix-
WEEKS AFTER HSV62 RECURRENCEFIG. 1. Appearance ofAG-4 complement-fixing antibody after HSV-2 recurrence. The complement-fixing
antibody was measured as described in the text with dilutions of sera up to 1:48.
INFECT. IMMUN.
AG-4 ANTIBODY IN HSV-2 PATIENTS 25
ation test depends on the time blood samplesare taken after lesion appearance. AG-4 comple-ment-fixing antibody titers in HSV-2-infectedpatients were usually in the range of 1:4 to 1:16,similar to those reported for CaCx patients (1),although one of our patients' samples (Fig. 1)did attain a titer of 1:36. Sera from patients withconfirmed HSV-1 genital infections were nega-tive for AG-4 complement-fixing antibody.Immunoprecipitation of AG-4 with HSV-
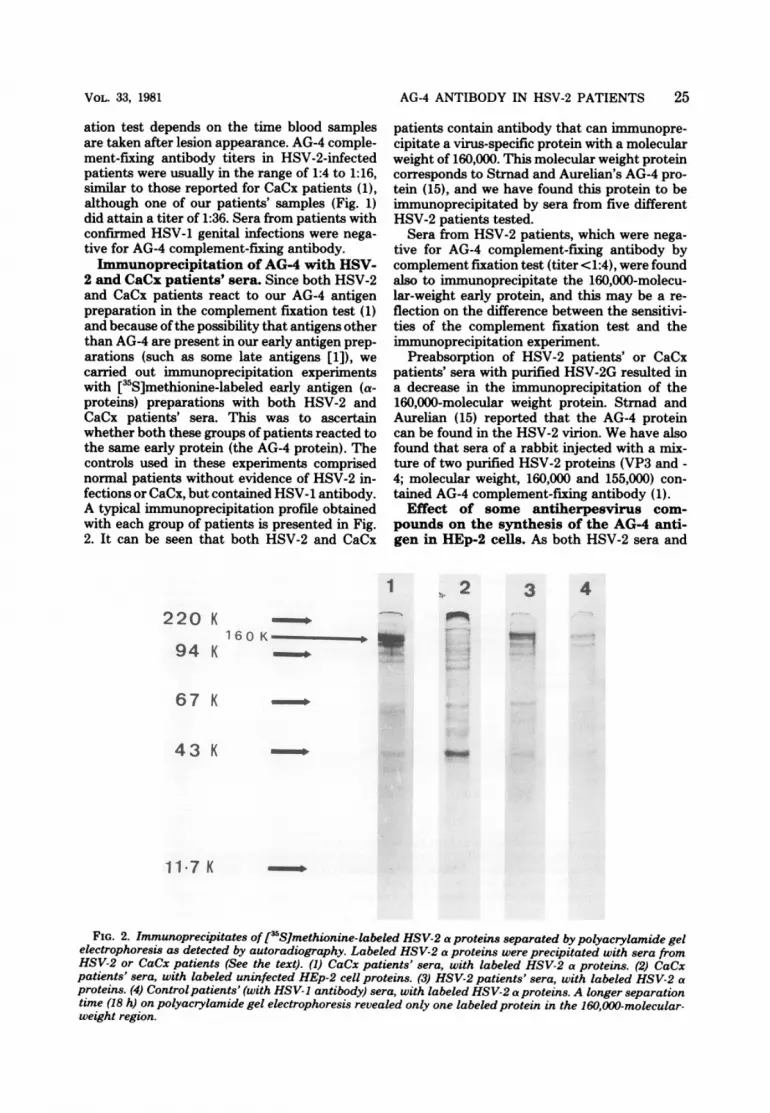
2 and CaCx patients' sera. Since both HSV-2and CaCx patients react to our AG-4 antigenpreparation in the complement fixation test (1)and because ofthe possibility that antigens otherthan AG-4 are present in our early antigen prep-arations (such as some late antigens [1]), wecarried out immunoprecipitation experimentswith [3S]methionine-labeled early antigen (a-proteins) preparations with both HSV-2 andCaCx patients' sera. This was to ascertainwhether both these groups of patients reacted tothe same early protein (the AG-4 protein). Thecontrols used in these experiments comprisednormal patients without evidence of HSV-2 in-fections or CaCx, but contained HSV-1 antibody.A typical immunoprecipitation profile obtainedwith each group of patients is presented in Fig.2. It can be seen that both HSV-2 and CaCx
1
220 K
94 K
patients contain antibody that can immunopre-cipitate a virus-specific protein with a molecularweight of 160,000. This molecular weight proteincorresponds to Strnad and Aurelian's AG-4 pro-tein (15), and we have found this protein to beimmunoprecipitated by sera from five differentHSV-2 patients tested.
Sera from HSV-2 patients, which were nega-tive for AG-4 complement-fixing antibody bycomplement fixation test (titer <1:4), were foundalso to immunoprecipitate the 160,000-molecu-lar-weight early protein, and this may be a re-flection on the difference between the sensitivi-ties of the complement fixation test and theimmunoprecipitation experiment.
Preabsorption of HSV-2 patients' or CaCxpatients' sera with purified HSV-2G resulted ina decrease in the immunoprecipitation of the160,000-molecular weight protein. Strnad andAurelian (15) reported that the AG-4 proteincan be found in the HSV-2 virion. We have alsofound that sera of a rabbit injected with a mix-ture of two purified HSV-2 proteins (VP3 and -4; molecular weight, 160,000 and 155,000) con-tained AG-4 complement-fixing antibody (1).
Effect of some antiherpesvirus com-pounds on the synthesis of the AG-4 anti-gen in HEp-2 cells. As both HSV-2 sera and
3 4
160 K_R
67 K
43 K
117 K
FIG. 2. Immunoprecipitates of [35S]methionine-labeled HSV-2 a proteins separated by polyacrylamide gelelectrophoresis as detected by autoradiography. Labeled HSV-2 a proteins were precipitated with sera fromHSV-2 or CaCx patients (See the text). (1) CaCx patients' sera, with labeled HSV-2 a proteins. (2) CaCxpatients' sera, with labeled uninfected HEp-2 cell proteins. (3) HSV-2 patients' sera, with labeled HSV-2 aproteins. (4) Control patients' (with HSV-1 antibody) sera, with labeled HSV-2 a proteins. A longer separationtime (18 h) on polyacrylamide gel electrophoresis revealed only one labeled protein in the 160,000-molecular-weight region.
VOL. 33, 1981

26 ARSENAKIS AND MAY
CaCx sera cortain antibody to a 160,000-molec-ular-weight early HSV-2 protein, the AG-4 pro-
tein, we used these sera to investigate the effectof a number of antiherpesvirus compounds on
the synthesis of the AG-4 antigen in HSV-2-infected HEp-2 cells. Although the majority ofthe drugs tested block late functions ofthe HSV-2 replication, their effect on early antigens ofHSV-2 has not been investigated.The effect of the antiherpesvirus compounds
listed in Table 1 on AG-4 synthesis was investi-gated. Included in the table is their effect on
infectious HSV-2 virus yields, as well as theireffect on AG-4 synthesis. We evaluated the AG-4 antigen production by the AG-4 complementfixation test, using HSV-2 and CaCx patients'sera positive for AG-4 antibody. At the concen-
trations used (as indicated in Table 1), none ofthe compounds was toxic to the HEp-2 cells, andwith an MOI of 0.7 PFU/cell (which is that usedfor maximum AG-4 antigen production), a re-
duction of about 2 log values in infectious virusyields was generally obtained. Higher virus in-hibition levels could be achieved at a lower MOI(Table 1). At the MOI where AG-4 productionis maximal, none of the compounds tested had
any appreciable effect on the synthesis of theAG-4 antigen in HEp-2 cells.
DISCUSSIONWe previously reported the presence of AG-4
antibodies in HSV-2-infected patients (1). In thestudy described here, we extended those initialobservations by following the AG-4 antibodyresponse in the course of HSV-2 infections inpatients with confirmed primary or recurrentHSV-2 genital lesions. It was found that AG-4antibody was generally absent from sera col-lected at the time of lesion appearance; thus,only 3 of 13 patients tested were positive for AG-4 complement-fixing antibodies at the time oflesion appearance. Furthermore, as sera were
collected from the health clinics, it is possiblethat some sera were collected not at the timeherpes-like lesions first appeared, but a few dayslater and, as can be seen from Fig. 1, a delay inpresentation of more than 4 days can result inthose patients' reacting positive in the AG-4complement fixation test.Four patients were followed from the time of
presenting with HSV-2 genital lesions to varioustimes thereafter. As was shown in Fig. 1, in three
TABLE 1. Effect of antiherpesvirus compounds on HSV-2G virus titer and AG-4 early antigen production inHEp-2 cellsa
Amt of complement-fix-Log,o reduction in Log1o reduction in ing units of AG-4 antigen
Concn of antiher- virus titer at 0.7 virus titer at 0.1 per ml of antigen prepa-Compound pesvirus com- PFU/cell (MOIb c PFU/cell (MOIb c rationd
pound (,ug/ml) relative to un- relative to un-treated control) treated control) CaCxse- HSV-2 se-
rum' rum'
No antiherpesvirus compound 0 0.0 0.0 20 20Phosphonoformate 150 2.2 7.4 20 20Phosphonoacetic acid 100 2.3 7.4 20 20Adenine arabinoside 200 2.3 5.2 20 20Acycloguanosine 11.2 2.5 2.9 20 20Arildone 15 0.6 1.0 15 15Iododeoxyuridine 100 2.4 3.1 15 15Trifluorothymidine 100 2.2 5.7 20 202'-Deoxyglucose 10,000 2.0 3.7 15 15Retinol 7.5f 0.9 0.9 20 20Retinoic acid 12.0f 1.0 1.0 20 20
a All experiments were performed on at least two separate occasions.b HEp-2 cells were infected at an MOI of 0.7 or 0.1 PFU/cell in the presence of the particular antiherpesvirus
compound at the concentration indicated. Virus was harvested 24 h postinfection, and virus yields were assayedby plaque assay on HEp-2 cells as described in the text.
'Antiviral compounds were added at the time of virus adsorption and were present throughout the infectionperiod.
d AG-4 was prepared as described in the text in either the presence or absence of the antiherpesviruscompounds. The protein concentration of each antigen preparation was 2.0 mg/ml. AG-4 product was assessedby complement fixation test with standard AG-4 +ve CaCx and HSV-2 patients' sera and various units ofantigen (0 to 2 units).'The titer of AG-4 complement-fixing antibody in these sera was 1:8. Similar results were found with rabbit
anti-VP3 and 4 (160,000 and 155,000 HSV-2 MI proteins) sera (1).f Found to destroy HSV-2 infectivity (14).
INFECT. IMMUN.

AG-4 ANTIBODY IN HSV-2 PATIENTS 27
of the four patients, the level of AG-4 antibodyrose from less than 1:4 at presentation to detect-able levels (more than 1:4) by day 4 after lesionappearance. One patient had a titer ofmore than1:4 at presentation, and she remained at thatlevel for 2 weeks before her AG-4 antibody titerdeclined; however, this patient was experiencingHSV-2 recurrences every 4 weeks. One patientwho was followed for nearly 1 year had a titer of>1:4 at 3 months after lesion appearance, whichfell below detectable levels (<1:4) 6 monthslater, when a lesion recurred in this same patient4 months later, the AG-4 antibody level wasagain <1:4 but rose to >1:4 within 5 days. There-fore, it appears that the AG-4 antibody (as de-tected by complement fixation) does not persistin HSV-2-infected patients and only appearsafter an outbreak of infection. This is in contrastto the situation in CaCx patients where the AG-4 antibody seems to persist even after apparentlysuccessful treatment. Antibody to cytomegalo-virus early antigens has been reported to persistin cytomegalovirus-infected patients also (4).However, in infectious mononucleosis patients,Epstein-Barr virus early antigens appear onlyafter an infection and decline with time (6).Similarly, in Varicella-Zoster virus infections,antibody to Varicella-Zoster early antigens ap-pear 3 to 5 days after symptoms, and as theinfection subsides, the early antibody declinesalso (3). Of 30 HSV-2 patients tested for AG-4antibody 1 to 4 weeks after lesion appearance,28 were positive. The antibody to AG-4 wasfound to be associated with the IgM fraction inthe sera tested during this period. In one patientwith unusually high AG-4 antibody titer (1:36),complement-fixing activity was associated withIgG as well as the IgM fraction. The rise in AG-4 antibody titer was paralleled by a two- tofourfold rise in HSV-2 neutralizing antibody inthe patients tested. Thus, the detection of AG-4antibody seems to depend on the time afterlesion appearance. Of the confirmed HSV-2 pa-tients we have tested for AG-4 complement-fix-ing antibody to date, the great majority arepositive, whereas when sera were negative at 1to 4 weeks postinfection, no HSV-2 virus couldbe isolated, and in some cases HSV-1 was iso-lated. We have not found AG-4 complement-fuixng antibody in confirmed HSV-1 patientsbeyond that found in normal control patients(1).To identify the protein(s) responsible for the
AG-4 complement-fixing reaction, we carried outimmunoprecipitation experiments with AG-4complement-fixing antibody positive HSV-2 andCaCx sera. The common protein immunoprecip-itated by both CaCx and HSV-2 sera, but notnormal control sera, was an 160,000-molecular-
weight protein absent from immunoprecipitateswith uninfected control preparations. This is inaccordance with the finding of Strnad and-Aurelian (15), who reported the immunoprecip-itation of a 160,000-molecular-weight protein byCaCx sera. Thus, it appears that both HSV-2and CaCx sera react to the same HSV-2 earlyprotein, the AG-4 protein, suggesting that theAG-4 antigen is an HSV-2-type-specific proteinand not exclusively a CaCx-tumor-specific anti-gen. As both CaCx and HSV-2 sera containedanti-AG-4 antibody, this was used to test theeffect on the production of AG-4 antigen inHSV-2-infected HEp-2 cells. Although the ma-jority of these compounds inhibited infectiousHSV-2 production by about 2 log values, theyhad no marked effect on the production of AG-4 antigen. All of these compounds tested (withthe exception of retinol, retinoic acid, and Aril-done) are known to inhibit HSV-2 replication byinhibiting late protein synthesis and presumablyhave no effect on early protein synthesis. Cer-tainly they have no effect on the AG-4 proteinwhich is an immediate early (a) HSV-2 inducedprotein (15).
ACKNOWLEDGMENTSWe acknowledge the assistance of D. Bradford of the Com-
municable Diseases Centre and the physicians at La TrobeUniversity's Health Centre for supplying sera from herpes-infected patients.
This work was supported in part by a grant from theAustralian Research Grant Committee.
LITERATURE CITD1. Arsenakis, M., G. M. Geogiou, J. K. Welsh, M. N.
Cauchi, and J. T. May. 1980. AG-4 complement fixingantibodies in cervical cancer and herpes-infected pa-tients using local Herpes simplex virus type 2. Int. J.Cancer 25:67-71.
2. Aurelian, L., B. Schumann, R. L. Marcus, and H. J.Davis. 1973. Antibody to HSV-2 induced tumor specificantigens in serums from patients with cervical carci-noma. Science 181:161-164.
3. Gerna, G., P. M. Cereda, E. Cattaneo, G. Achilli, andM. T. Gerna. 1979. Antibody to early antigens of vari-cella-zoster virus during varicella and zoster. J. Infect.Dis. 140:33-41.
4. Griffiths, P. D., K. J. Buie, and R. B. Heath. 1980.Persistence of high titre antibodies to the early antigensof cytomegalovirus in pregnant women. Arch. Virol. 64:303-309.
5. Heise, E. R., L. S. Kucera, M. Raben, and H. Homes-ley. 1979. Serological response pattern to herpes virustype 2 early and late antigens in cervical carcinomapatients. Cancer Res. 39:4022-4026.
6. Henle, W., G. Henle, and E. T. Lennette. 1979. TheEpstein-Barr virus. Sci. Am. 241:40-51.
7. Kawana, T., S. Sakamoto, T. Kocsamatsu, and L.Aurelian. 1978. Frequency of anti-AG-4 antibody inpatients with uterine cervical cancer and controls. Gann69:589-591.
8. Kennett, M., A. Donaldson, and F. A. Lewis. 1977.Herpes simplex, p. 13. In Annual Report of FairfieldHospital to the World Health Organisation, FairfieldHospital, Melbourne.
VOL. 33, 1981

28 ARSENAKIS AND MAY
9. Medappa, K. C., C. McLean, and R. R. Reuckert. 1971.On the structure of rhinovirus 1A. Virology 44:259-270.
10. Notter, M. F. D., and J. J. Docherty. 1976. Comparativediagnostic aspects of herpes simplex virus tumor-asso-ciated antigens. J. Natl. Cancer Inst. 57:483-488.
11. Pereira, L., M. H. Wolff, M. Fenwick, and B. Roizman.1977. Regulation of herpes virus macromolecular syn-thesis. V. Properties of a-polypeptides made in HSV-1and HSV-2 infected cells. Virology 77:733-749.
12. Rawls, W. E., S. Bacchetti, and F. L. Graham. 1977.Relation of herpes simplex viruses to human malignan-cies. Curr. Top. Microbiol. Immunol. 77:71-95.
13. Rawls, W. E., K. Iwamoto, E. Adam, and J. L. Mel-
INFECT. IMMUN.
nick. 1970. Measurement of antibodies to herpesvirustypes 1 and 2 in human sera. J. Immunol. 104:599-606.
14. Reinhardt, A., J. A. Sands, and W. Sniper. 1979.Virucidal activity of retinal. Antimicrob. Agents Chem-other. 16:421-423.
15. Strnad, B. C., and L. Aurelian. 1976. Proteins of her-pesvirus type 2. II. Srudies demonstrating a correlationbetween a tumor-associated antigen (AG-4) and a virionprotein. Virology 73:244-258.
16. Welsh, J. K., I. J. Skurrie, and J. T. May. 1978. Use ofSemliki forest virus to identify lipid-mediated antiviralactivity and anti-alphavirus immunoglobulin A in hu-man milk. Infect. Immun. 19:395-401.





























