Embed Size (px)
Citation preview

doi:10.1006/jmbi.2000.3525 available online at http://www.idealibrary.com on J. Mol. Biol. (2000) 296, 1245±1256
Crystal Structure of Lysbbb182-Lysbbb282 CrosslinkedHemoglobin: A Possible Allosteric Intermediate
Elias J. Fernandez1, Cele Abad-Zapatero2 and Kenneth W. Olsen1*
1Loyola University ChicagoDepartment of ChemistryChicago, IL, 60626, USA2Protein CrystallographyLaboratory, Abbott LaboratoriesAbbott Park, IL, 60004, USA
Present address: E. J. Fernandez, YDepartment of Pharmacology, NewUSA.
Abbreviations used: a2bTmb, hemby trimesic acid between Valb1 1, Ly82; a2bCAb, hemoglobin crosslinkedcarboxycinnamic acid between Lysba2bNDb, hemoglobin crosslinked bydicarboxylic acid between Lysb1 82b82XLHbA, hemoglobin crosslinkedbetween Lysb182 and Lysb282; BMEhemoglobin crosslinked by bis(N-maHbA, hemoglobin A.
E-mail address of the [email protected]
0022-2836/00/051245±12 $35.00/0
The crystal structure of human hemoglobin crosslinked between theLysb82 residues has been determined at 2.30 AÊ resolution. The cross-linking reaction was performed under oxy conditions using bis(3,5-dibro-mosalicyl) fumarate; the modi®ed hemoglobin has increased oxygenaf®nity and lacks cooperativity. Since the crystallization occurred underdeoxy conditions, the resulting structure displays conformational charac-teristics of both the (oxy) R and the (deoxy) T-states. b82XLHbA does notfully reach its T-state conformation due to the presence of the crosslink.The R-state-like characteristics of deoxy b82XLHbA include the positionof the distal Hisb63 (E7) residue, indicating a possible reason for the highoxygen af®nity of this derivative. Other areas of the molecule, particu-larly those thought to be important in the allosteric transition, such asTyrb145 (HC2) and the switch region involving Proa144 (CD2), Thra141(C6) and Hisb297 (FG4), are in intermediate positions between the R andT-states. Thus, the structure may represent a stabilized intermediate inthe allosteric transition of hemoglobin.
# 2000 Academic Press
Keywords: hemoglobin; allosteric intermediates; cooperativity;crosslinking; X-ray crystallography
*Corresponding authorIntroduction
The allosteric transition of hemoglobin betweenthe deoxy T-state and the oxy R-state involvestransient structures with distinct structural confor-mations (Perutz et al., 1998.) The T-state and the R-state structures have been examined thoroughly byX-ray crystallography (Fermi et al., 1984; Shaanan,1983). Understanding how hemoglobin undergoesthis transition in molecular detail will requirethree-dimensional structures of all the intermediateconformations. However, the dif®culty in obtaining
ale University,Haven, CT 06510,
oglobin crosslinkedsb1 82 and Lysb2
by 4-
1 82 and Lysb2 82;2,6-napthalene
and Lysb2 82;by fumarate
-Hb, horseleimidoethyl) ether;
ng author:
stable intermediates has prevented a completeunderstanding of the allosteric mechanism.
Efforts have been made to capture these inter-mediate structures by the partial and step-wise lig-ation of hemoglobin crystals (Liddington et al.,1988, 1992; Paoli et al., 1996; Luisi et al., 1990;Mozzarelli et al., 1990; Brzowowski et al., 1984;Grabowski et al., 1978). The T-state structuredescribed by Paoli et al. (1996) has three out of fourheme molecules liganded with oxygen; the fourth,a b heme, showed only partial ligation. The result-ing molecule was observed to be largely in the T-state conformation but with a signi®cant deviationtowards the R-state con®guration. Partial R-state-like con®gurations were observed in the region ofthe liganded heme molecules, and the bindingpockets had noticeable expansion to accommodatethe bound oxygen atoms. There were also signi®-cant changes in the iron atom stereochemistry,which was closer to the heme geometry of R-statehemoglobin. The effect of ligation of the deoxyhemoglobin crystals was more pronounced in theb-subunits than in the a-subunits.
The other crystallographic approach that hasproduced potential intermediate structuresinvolves the use of crosslinking reagents. The con-cept behind these studies was to crosslink thedeoxy structure and then to grow the crystals in
# 2000 Academic Press

Figure 1. The 2Fo ÿ Fc map calculated at 3s in theregion of the crosslink. The two Lysb82 residues are dis-played ¯anking the fumarate crosslinker. The covalentbonds linking the crosslinker carbonyl carbon atoms tothe LysNz have been omitted deliberately. Also dis-played is the lone water molecule that appears to behydrogen-bonded to one of the crosslinker carbonylgroups. A similar water molecule was built in the non-crystallographic symmetry-related molecule.
1246 Structure of Hemoglobin Allosteric Intermediate
the presence of CO, so that the molecule attemptsto adopt the R-state. Schumacher et al. (1995) ®rstreported three crosslinked hemoglobin moleculesanalyzed in this manner. When deoxy HbA bindsto CO, the a2b2 dimer undergoes an approximately15.0 � rotation relative to the a1b1 dimer (Baldwin& Chothia, 1979). The same rotation was observedwhen the hemoglobin was crosslinked between thea-amino group of the b1-chain and the b282 lysineresidue using 3,30-stilbenedicarboxylic acid. How-ever, when trimesic acid was used to crosslink theb1 a-amino group to the b282 lysine residue or toboth the b182 lysine and the b282 lysine residues,this rotation was restricted to 7.4 � and 8.5 �,respectively. These derivatives appeared to beintermediates on the R! T pathway. Schumacheret al. (1997) applied the same technique to hemo-globin molecules crosslinked between the two b82lysine residues with bis(methylphosphate) deriva-tives of either 4-carboxycinnamic acid or 2,6-naphthalene dicarboxylic acid. These two struc-tures appeared to be intermediate between the R(Shaanan, 1983) and R2-states (Silva et al., 1992).Taken together, these crystal structures of cross-linked hemoglobin structures provide evidence fora pathway from the T-state to a2bTmb, to theR-state, to a2bCA (orND) to the R2-state.
Horse hemoglobin crosslinked with bis(N-malei-midoethyl) ether has been crystallized in the metform and then reduced by dithionite to obtain thestructure of deoxyhemoglobin trapped in the R-state (Wilson et al., 1996). This structure demon-strates shifts in the FG corners that are character-istic of the allosteric transition while remainingessentially in the R-state. Although the presumedcrosslinking sites are in the b-chain, the differencesbetween the crosslinked met and deoxy forms ofthe protein are more evident in the a-chain, poss-ibly indicating that the crosslink did not restrainthe movement of the protein.
We report here the crystal structure of ahemoglobin crosslinked between the two Lysb82residues. The reaction was performed under oxyge-nated conditions with bis(3,5-dibromosalicyl)fumarate (Walder et al., 1979), but the crystalliza-tion was achieved under deoxy conditions (thestereo code of the crosslinker is EF6). This product,b82XLHbA, had been examined previously by X-ray analysis at only 4.40 AÊ resolution (Walder et al.,1980). b82XLHBA exhibits high oxygen af®nity(Walder et al., 1979), high thermal stability (White& Olsen, 1987; Yang & Olsen, 1989), and normalautoxidation (Yang & Olsen, 1991). Crosslinkedhemoglobin molecules of this type have been con-sidered as potential blood substitutes (Winslow,1992). Due to the restraint imposed by the cross-link, the molecule is prevented from fully assum-ing the T-state conformation under the deoxycrystallization conditions. A signi®cant portion ofthe molecule appears to retain some R-state charac-teristics. Based on our crystal structure and on theunusual oxygen binding properties of b82XLHBA,we believe that this structure may be similar to an
intermediate close to the T-state on the R! T-statepathway.
Results
b82XLHbA was crystallized under deoxy con-ditions. The structure was re®ned to a resolution of2.3 AÊ . The crosslink is well de®ned in the electrondensity map, as are a number of changes fromthe normal T-state structure. The changes make thestructure slightly like the R-state in some areas,while generally assuming the T-state. Theb82XLHbA structure can be analyzed in terms ofthe R! T transition. Since oxygen is not presentin the crystals, the T-state will be highly favored inthe equilibrium between R and T-states (Monod,et al., 1965).
Conformation of the fumarate crosslink
Figure 1 is the un-averaged atomic model ofb82XLHbA superimposed on the 2Fo ÿ Fc electrondensity map in the region of the crosslink. Densitydue to the crosslink extends from one Lysb82 resi-due to the other and the fumarate crosslink was

Structure of Hemoglobin Allosteric Intermediate 1247
®tted into this difference density (stereo code EF6).This was in contrast to density observed in anothercrosslinked HbA species, where the crosslink wasbetween the two Lysa99 residues. In this lattercase, the electron density due to the crosslink wasvery poorly de®ned and this was attributed to thelarge mobility (temperature factors >40.0 AÊ 2) of thecrosslink and the Lysa99 atoms (E.J.F., C.A.-Z &K.W.O., unpublished results).
The introduction of the crosslink perturbs thesolvent in the region of the b-cleft. This leads toformation of new hydrogen bonds between resi-dues, and between residues and water molecules.There are eight water molecules in the b-cleftregion of b82XLHbA, compared with six in nativeT-state HbA. For example, the nitrogen atom ofValb11(NA1) hydrogen bonds to Leub178 (EF2) inthe T-state, while in b82XLHbA the same nitrogenatom is hydrogen bonded both to the leucine resi-due and to a water molecule. In both species thereis a water molecule bonded to the nitrogen atom ofAsnb1139(H17). However, while the carbonyl oxy-gen atom of Glub1136(H14) hydrogen bonds to thenitrogen atom of Alab1140(H18) in the T-state, thecorresponding Glub1136(H14) forms no noticeablehydrogen bonds in b82XLHbA.
Structural comparisons with R and T-statenative hemoglobin
Two reference frames were used for superposi-tion of T-state (Fermi et al., 1984), R-state (Shanaan,1983) and b82XLHbA. One reference frame wasobtained by superimposing only the a1b1 dimer ofeach of the three hemoglobins. The root-mean-square (rms) difference of b82XLHbA a1b1 dimerfrom the corresponding T-state dimer is 0.5 AÊ , andis 1.5 AÊ from the R-state. The second referenceframe was achieved by the overlay of the B, G andH helices (B-helix: a120-35, b119-34; G-helix: a194-112, b199-117; H-helix: a1118-138, b1123-143) of thethree molecules using the criteria prescribed byBaldwin & Chothia (1979). In each case only themain-chain atoms were used. The conclusionsobtained from both reference frames are the same.The BGH reference frame was used to generateFigures 2 and 3.
Upon release of the liganded oxygen atom, thea2b2 dimer undergoes an approximately 15.0 �rigid-body rotation from its corresponding positionin the T-state (Baldwin & Chothia, 1979). Thecorresponding rotation of the a2b2 dimer inb82XLHbA is about 3 � less than that of T-stateHbA (Figure 2). The rms difference of R-state,T-state and b82XLHbA can be quanti®ed andrepresented as a graphical plot (Figure 3). Despitethe nearly T-state structure of approximately 83 %of b82XLHbA, it still retains a signi®cantly detect-able R-state-like character. Although much smaller,the pattern of rms difference of b82XLHbA fromthe T-state closely resembles that of R-state fromthe T-state. Furthermore, inter-heme distancesmeasured between the Fe atoms of the four
subunits (Table 1A) are neither those of the T-statenor the R-state, but have values intermediatebetween the two conformations. These ®ndingssuggest that b82XLHbA is a snapshot of one of theintermediate structures in the R! T transition ofhemoglobin.
Tertiary conformation of bbb82XLHbA
The a1b1 dimers of the three hemoglobin mol-ecules are mostly similar (Figure 2). Closer inspec-tion indicates some signi®cant differences betweenb82XLHbA and the two native hemoglobin mol-ecules (Figure 3). The a chain of b82XLHbA ismostly in the T-state (Table 2). Signi®cant regionsof the b chain retain some R-state characteristics,and there are areas in b82XLHbA that are relatedto neither the T nor the R-state. At least 27 % of theresidues in b82XLHbA have completely switchedinto the T-state: 33 % of the a chain, and 21 % ofthe b chain.
a-Chain
The a chain of b82XLHbA displays most ofthe interactions which are characteristic of theT-state (Table 3). For example, Arga1141(HC3),Lysa2127(H10) and Vala11(NA1) appear to beinvolved in the same electrostatic interactions as inthe T-state. Another unique T-state interactioninvolving the H-bond between Arga1141(HC3) andthe carbonyl group of Aspa2126(H9) is alsoobserved.
The differences between the b82XLHbA and T-state a chains are minor (Figure 2). The largestdeviations are in the N-terminal Val residue and inthe region of the F helix closer to the FG loopwhose conformation in b82XLHbA is similar to theR-state. In both the R-state and b82XLHbA, the Fhelix is closer to the heme than in the T-state. Also,the R-state F helix is inclined relative to the T-stateF helix by an angle of 9.7 �; the correspondingincline of the b82XLHbA F helix to the T-state is4.0 �. This results in the b82XLHbA proximalHisa87(F8) being closer to the heme Fe atom by0.10 AÊ than in the T-state (Figure 4(a)). The aver-age rms difference of the b82XLHbA a chain fromthe T-state does not exceed 1.0 AÊ .
b-Chain
The crosslink affects the movement of the bchains signi®cantly. Normally the R! T transitionpulls the b chains approximately 4.0 AÊ furtherapart; in b82XLHbA the two chains are held closerby the crosslink.
The superimposed models in Figure 2 and therms difference plot in Figure 3 indicate speci®cregions in the b chain that retain R-state-likecharacteristics. As expected, the largest deviationsfrom the T-state are observed in the E helix andthe EF loop, which are close to the crosslinking site(Figure 5). There is a shift in the A helix relative to
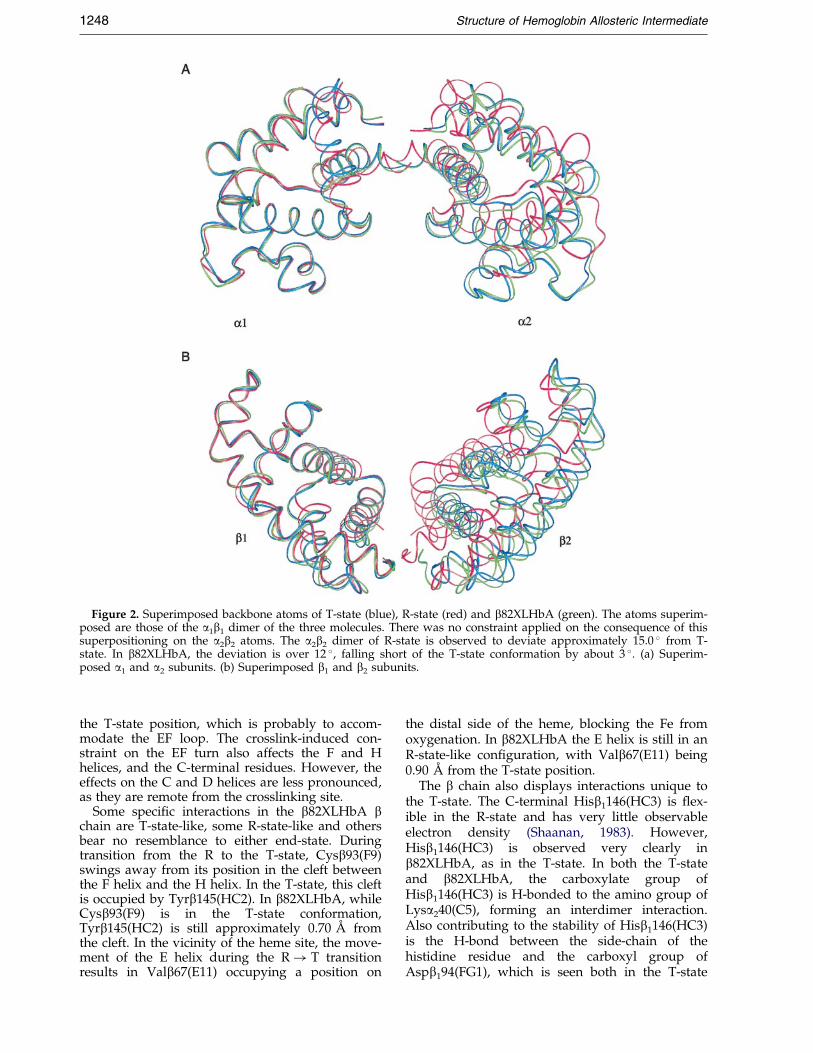
Figure 2. Superimposed backbone atoms of T-state (blue), R-state (red) and b82XLHbA (green). The atoms superim-posed are those of the a1b1 dimer of the three molecules. There was no constraint applied on the consequence of thissuperpositioning on the a2b2 atoms. The a2b2 dimer of R-state is observed to deviate approximately 15.0 � from T-state. In b82XLHbA, the deviation is over 12 �, falling short of the T-state conformation by about 3 �. (a) Superim-posed a1 and a2 subunits. (b) Superimposed b1 and b2 subunits.
1248 Structure of Hemoglobin Allosteric Intermediate
the T-state position, which is probably to accom-modate the EF loop. The crosslink-induced con-straint on the EF turn also affects the F and Hhelices, and the C-terminal residues. However, theeffects on the C and D helices are less pronounced,as they are remote from the crosslinking site.
Some speci®c interactions in the b82XLHbA bchain are T-state-like, some R-state-like and othersbear no resemblance to either end-state. Duringtransition from the R to the T-state, Cysb93(F9)swings away from its position in the cleft betweenthe F helix and the H helix. In the T-state, this cleftis occupied by Tyrb145(HC2). In b82XLHbA, whileCysb93(F9) is in the T-state conformation,Tyrb145(HC2) is still approximately 0.70 AÊ fromthe cleft. In the vicinity of the heme site, the move-ment of the E helix during the R! T transitionresults in Valb67(E11) occupying a position on
the distal side of the heme, blocking the Fe fromoxygenation. In b82XLHbA the E helix is still in anR-state-like con®guration, with Valb67(E11) being0.90 AÊ from the T-state position.
The b chain also displays interactions unique tothe T-state. The C-terminal Hisb1146(HC3) is ¯ex-ible in the R-state and has very little observableelectron density (Shaanan, 1983). However,Hisb1146(HC3) is observed very clearly inb82XLHbA, as in the T-state. In both the T-stateand b82XLHbA, the carboxylate group ofHisb1146(HC3) is H-bonded to the amino group ofLysa240(C5), forming an interdimer interaction.Also contributing to the stability of Hisb1146(HC3)is the H-bond between the side-chain of thehistidine residue and the carboxyl group ofAspb194(FG1), which is seen both in the T-state
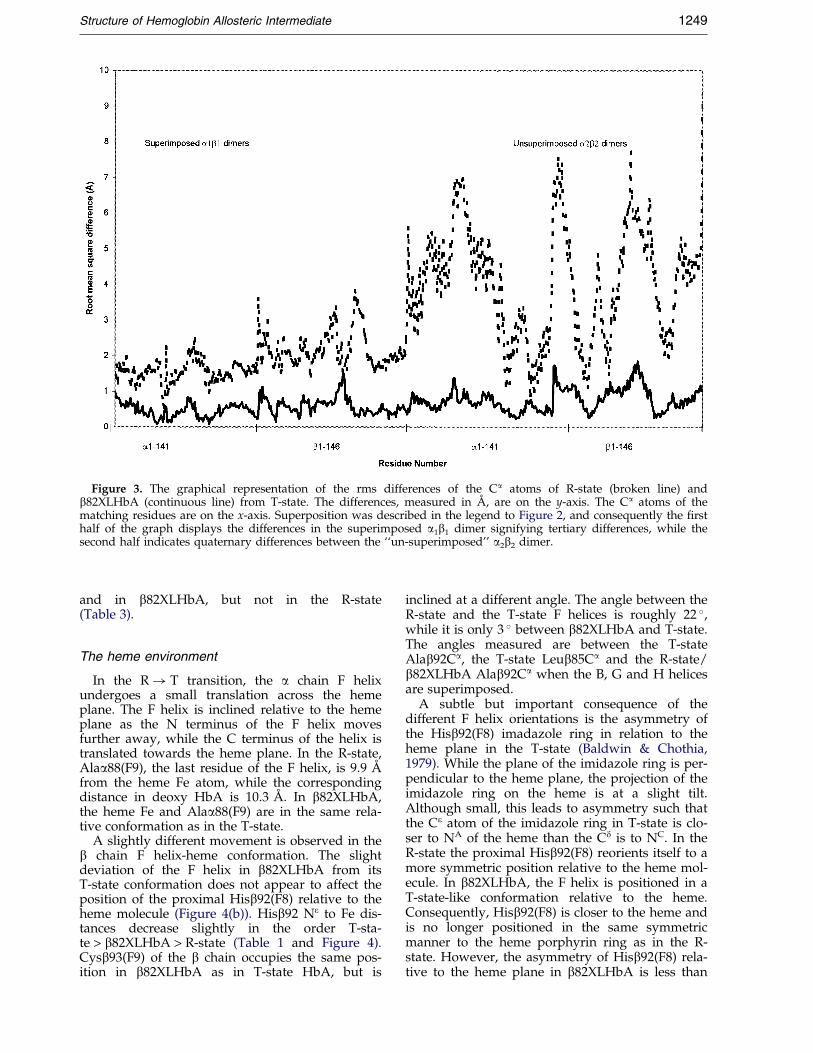
Figure 3. The graphical representation of the rms differences of the Ca atoms of R-state (broken line) andb82XLHbA (continuous line) from T-state. The differences, measured in AÊ , are on the y-axis. The Ca atoms of thematching residues are on the x-axis. Superposition was described in the legend to Figure 2, and consequently the ®rsthalf of the graph displays the differences in the superimposed a1b1 dimer signifying tertiary differences, while thesecond half indicates quaternary differences between the ``un-superimposed'' a2b2 dimer.
Structure of Hemoglobin Allosteric Intermediate 1249
and in b82XLHbA, but not in the R-state(Table 3).
The heme environment
In the R! T transition, the a chain F helixundergoes a small translation across the hemeplane. The F helix is inclined relative to the hemeplane as the N terminus of the F helix movesfurther away, while the C terminus of the helix istranslated towards the heme plane. In the R-state,Alaa88(F9), the last residue of the F helix, is 9.9 AÊ
from the heme Fe atom, while the correspondingdistance in deoxy HbA is 10.3 AÊ . In b82XLHbA,the heme Fe and Alaa88(F9) are in the same rela-tive conformation as in the T-state.
A slightly different movement is observed in theb chain F helix-heme conformation. The slightdeviation of the F helix in b82XLHbA from itsT-state conformation does not appear to affect theposition of the proximal Hisb92(F8) relative to theheme molecule (Figure 4(b)). Hisb92 Ne to Fe dis-tances decrease slightly in the order T-sta-te > b82XLHbA > R-state (Table 1 and Figure 4).Cysb93(F9) of the b chain occupies the same pos-ition in b82XLHbA as in T-state HbA, but is
inclined at a different angle. The angle between theR-state and the T-state F helices is roughly 22 �,while it is only 3 � between b82XLHbA and T-state.The angles measured are between the T-stateAlab92Ca, the T-state Leub85Ca and the R-state/b82XLHbA Alab92Ca when the B, G and H helicesare superimposed.
A subtle but important consequence of thedifferent F helix orientations is the asymmetry ofthe Hisb92(F8) imadazole ring in relation to theheme plane in the T-state (Baldwin & Chothia,1979). While the plane of the imidazole ring is per-pendicular to the heme plane, the projection of theimidazole ring on the heme is at a slight tilt.Although small, this leads to asymmetry such thatthe Ce atom of the imidazole ring in T-state is clo-ser to NA of the heme than the Cd is to NC. In theR-state the proximal Hisb92(F8) reorients itself to amore symmetric position relative to the heme mol-ecule. In b82XLHbA, the F helix is positioned in aT-state-like conformation relative to the heme.Consequently, Hisb92(F8) is closer to the heme andis no longer positioned in the same symmetricmanner to the heme porphyrin ring as in the R-state. However, the asymmetry of Hisb92(F8) rela-tive to the heme plane in b82XLHbA is less than

Table 1. Comparison of hemoglobin structures
Subunit T-state b82XLHbA R-state
A. Intersubunit heme distances (AÊ )a1-a2 34.2 34.5 34.8a1-b1 36.5 35.8 34.7a1-b2 24.4 24.5 25.5a2-b1 24.3 24.4 25.5
B. Hisb92(F8) imadazole asymmetry relative to heme prophyrina b
Atoms T R b82 T R b82Ce ÐNA 3.2 3.1 3.1 3.5 3.2 3.0CdÐNC 3.8 3.4 3.8 4.2 3.2 3.7
C. Distances between iron and neighboring histidine residues (AÊ )Molecule Subunit Fe!HisF8Ne Fe!HisE7Ne
Deoxy a 2.2 4.4b 2.2 4.1
b82XLHbA a 2.0 4.3b 2.1 4.2
Oxy a 1.9 4.3b 2.1 4.2
D. Heme pucker angles NAÐFeÐNC
T-state (deoxy) b82XLHbA (deoxy) R-state (oxy)a subunit (deg.) 156.4 156.4 168.9b subunit (deg.) 158.3 164.3 176.6
Crystallography (A) Intersubunit heme distances in AÊ ; FeÐFe. (B) Hisb92(F8) imadazole asymmetry relative to heme porphyrin.(C) Distances (AÊ ) between iron and neighboring histidine residues. (D) Heme pucker angles NAÐFeÐNC.
1250 Structure of Hemoglobin Allosteric Intermediate
that in the T-state, as is evident from the corre-sponding distances (Table 1B), and may representan intermediate position in the R! T pathway.
In the T-state, the distal histidine E7 residue ispositioned such that it partially blocks the oxygen-binding site. During the R! T transition,Hisb63(E7) aligns itself with the heme Fe (Figure 4)and the Fe-His distances increase by a small butdetectable amount (Table 1C). The orientation ofthe distal HisE7 residue relative to the heme mol-ecule is different for the two subunits (Figure 4).Again, in b82XLHbA, these distances are inter-mediate between the two end-states, and in the bchain the distal HisE7 is closer to the R-state pos-ition than the T-state. A slight doming of the hemegroup is also observed in b82XLHbA, similar but
Table 2. The T-state-like, R-state-like, and intermediate-state
T-statea 40(C5)-59(E8)a 78(EF7)-96(G3)a 134(H17)-140(HC2)b 70(E14)-76(E20)b 87(F3)-105(G7)b 142(H20)-145(HC2)R-statea 14(A12)-18(A16)a 21(B2)-34(B15)a 37(C2)-39(C4)a 60(E9)-77(EF6)a 97(G4)-115(GH3)b 1(NA1)-21(B3)b 24(B6)-31(B13)b 36(C2)-69(E13)b 77(EF1)-86(F2)b 106(G8)-119(GH2)b 128(H6)-141(H19)
to a lesser extent, than in T-state HbA. This dom-ing, or pucker, arises as the Fe of the heme ispulled slightly out of the plane of the porphyrinring due to the asymmetric coordination of the Featom when there is no sixth ligand to counter thecoordinated proximal His(F8) residue. In theb82XLHbA a chains the pucker angle measuredbetween NA � � �Fe � � �NC (Table 1C) is similar to theT-state; in the b chains, however, this pucker is lessobvious, making the heme porphyrin rings moresimilar to the R-state.
The a1b2 (and a2b1) interface
b82XLHbA displays most of a1b2 subunit inter-face contacts associated with the T-state (Table 4).
regions of b82XLHbA
Regions different from both T and R-statesa 1(NA2)-13(A11)a 19(AB1)-20(B1)a 35(B16)-36(C1)a 116(GH4)-133(H16)b 22(B4)-23(B5)b 32(B14)-35(C1)b 120(GH3)-127(H5)
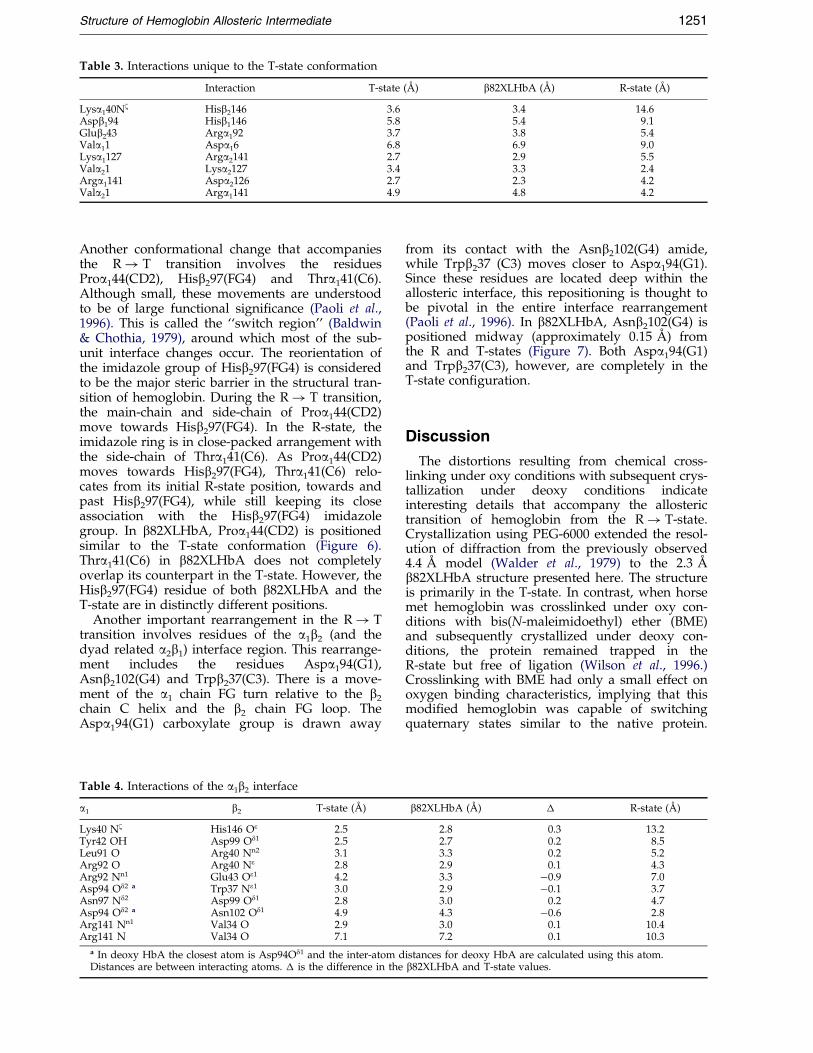
Table 3. Interactions unique to the T-state conformation
Interaction T-state (AÊ ) b82XLHbA (AÊ ) R-state (AÊ )
Lysa140Nz Hisb2146 3.6 3.4 14.6Aspb194 Hisb1146 5.8 5.4 9.1Glub243 Arga192 3.7 3.8 5.4Vala11 Aspa16 6.8 6.9 9.0Lysa1127 Arga2141 2.7 2.9 5.5Vala21 Lysa2127 3.4 3.3 2.4Arga1141 Aspa2126 2.7 2.3 4.2Vala21 Arga1141 4.9 4.8 4.2
Structure of Hemoglobin Allosteric Intermediate 1251
Another conformational change that accompaniesthe R! T transition involves the residuesProa144(CD2), Hisb297(FG4) and Thra141(C6).Although small, these movements are understoodto be of large functional signi®cance (Paoli et al.,1996). This is called the ``switch region'' (Baldwin& Chothia, 1979), around which most of the sub-unit interface changes occur. The reorientation ofthe imidazole group of Hisb297(FG4) is consideredto be the major steric barrier in the structural tran-sition of hemoglobin. During the R! T transition,the main-chain and side-chain of Proa144(CD2)move towards Hisb297(FG4). In the R-state, theimidazole ring is in close-packed arrangement withthe side-chain of Thra141(C6). As Proa144(CD2)moves towards Hisb297(FG4), Thra141(C6) relo-cates from its initial R-state position, towards andpast Hisb297(FG4), while still keeping its closeassociation with the Hisb297(FG4) imidazolegroup. In b82XLHbA, Proa144(CD2) is positionedsimilar to the T-state conformation (Figure 6).Thra141(C6) in b82XLHbA does not completelyoverlap its counterpart in the T-state. However, theHisb297(FG4) residue of both b82XLHbA and theT-state are in distinctly different positions.
Another important rearrangement in the R! Ttransition involves residues of the a1b2 (and thedyad related a2b1) interface region. This rearrange-ment includes the residues Aspa194(G1),Asnb2102(G4) and Trpb237(C3). There is a move-ment of the a1 chain FG turn relative to the b2
chain C helix and the b2 chain FG loop. TheAspa194(G1) carboxylate group is drawn away
Table 4. Interactions of the a1b2 interface
a1 b2 T-state (AÊ )
Lys40 Nz His146 Oe 2.5Tyr42 OH Asp99 Od1 2.5Leu91 O Arg40 Nn2 3.1Arg92 O Arg40 Ne 2.8Arg92 Nn1 Glu43 Oe1 4.2Asp94 Od2 a Trp37 Ne1 3.0Asn97 Nd2 Asp99 Od1 2.8Asp94 Od2 a Asn102 Od1 4.9Arg141 Nn1 Val34 O 2.9Arg141 N Val34 O 7.1
a In deoxy HbA the closest atom is Asp94Od1 and the inter-atom dDistances are between interacting atoms. � is the difference in the
from its contact with the Asnb2102(G4) amide,while Trpb237 (C3) moves closer to Aspa194(G1).Since these residues are located deep within theallosteric interface, this repositioning is thought tobe pivotal in the entire interface rearrangement(Paoli et al., 1996). In b82XLHbA, Asnb2102(G4) ispositioned midway (approximately 0.15 AÊ ) fromthe R and T-states (Figure 7). Both Aspa194(G1)and Trpb237(C3), however, are completely in theT-state con®guration.
Discussion
The distortions resulting from chemical cross-linking under oxy conditions with subsequent crys-tallization under deoxy conditions indicateinteresting details that accompany the allosterictransition of hemoglobin from the R! T-state.Crystallization using PEG-6000 extended the resol-ution of diffraction from the previously observed4.4 AÊ model (Walder et al., 1979) to the 2.3 AÊ
b82XLHbA structure presented here. The structureis primarily in the T-state. In contrast, when horsemet hemoglobin was crosslinked under oxy con-ditions with bis(N-maleimidoethyl) ether (BME)and subsequently crystallized under deoxy con-ditions, the protein remained trapped in theR-state but free of ligation (Wilson et al., 1996.)Crosslinking with BME had only a small effect onoxygen binding characteristics, implying that thismodi®ed hemoglobin was capable of switchingquaternary states similar to the native protein.
b82XLHbA (AÊ ) � R-state (AÊ )
2.8 0.3 13.22.7 0.2 8.53.3 0.2 5.22.9 0.1 4.33.3 ÿ0.9 7.02.9 ÿ0.1 3.73.0 0.2 4.74.3 ÿ0.6 2.83.0 0.1 10.47.2 0.1 10.3
istances for deoxy HbA are calculated using this atom.b82XLHbA and T-state values.
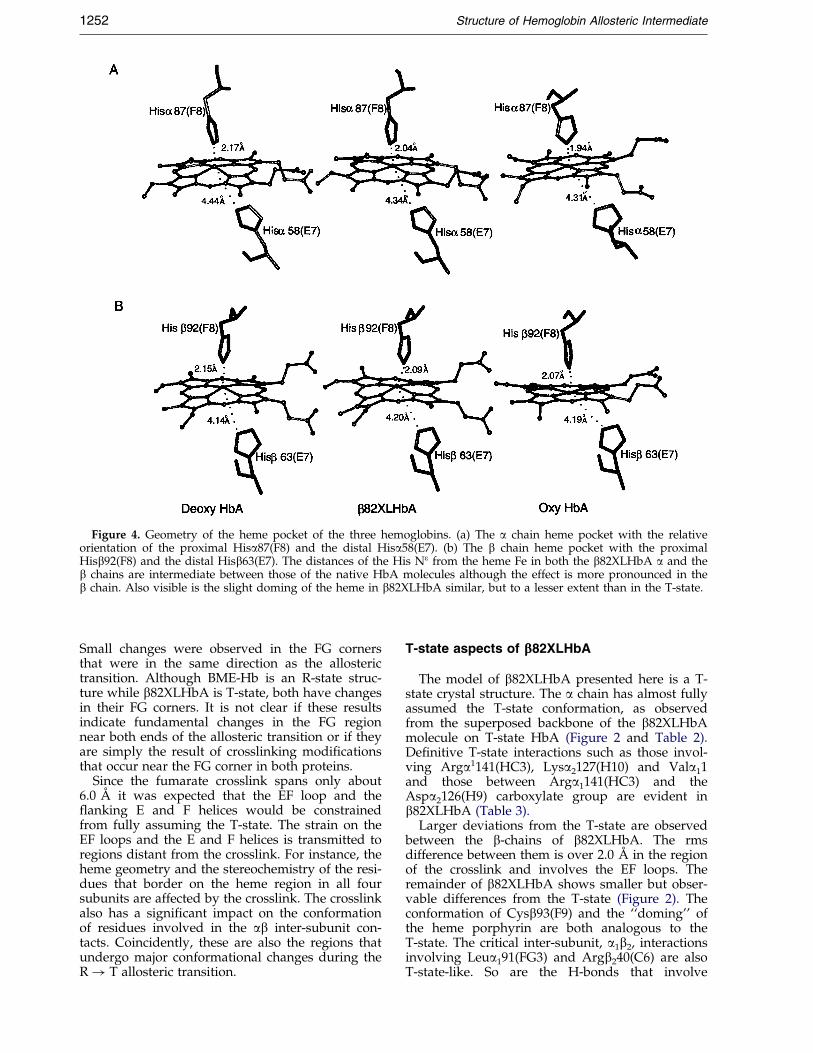
Figure 4. Geometry of the heme pocket of the three hemoglobins. (a) The a chain heme pocket with the relativeorientation of the proximal Hisa87(F8) and the distal Hisa58(E7). (b) The b chain heme pocket with the proximalHisb92(F8) and the distal Hisb63(E7). The distances of the His Ne from the heme Fe in both the b82XLHbA a and theb chains are intermediate between those of the native HbA molecules although the effect is more pronounced in theb chain. Also visible is the slight doming of the heme in b82XLHbA similar, but to a lesser extent than in the T-state.
1252 Structure of Hemoglobin Allosteric Intermediate
Small changes were observed in the FG cornersthat were in the same direction as the allosterictransition. Although BME-Hb is an R-state struc-ture while b82XLHbA is T-state, both have changesin their FG corners. It is not clear if these resultsindicate fundamental changes in the FG regionnear both ends of the allosteric transition or if theyare simply the result of crosslinking modi®cationsthat occur near the FG corner in both proteins.
Since the fumarate crosslink spans only about6.0 AÊ it was expected that the EF loop and the¯anking E and F helices would be constrainedfrom fully assuming the T-state. The strain on theEF loops and the E and F helices is transmitted toregions distant from the crosslink. For instance, theheme geometry and the stereochemistry of the resi-dues that border on the heme region in all foursubunits are affected by the crosslink. The crosslinkalso has a signi®cant impact on the conformationof residues involved in the ab inter-subunit con-tacts. Coincidently, these are also the regions thatundergo major conformational changes during theR! T allosteric transition.
T-state aspects of bbb82XLHbA
The model of b82XLHbA presented here is a T-state crystal structure. The a chain has almost fullyassumed the T-state conformation, as observedfrom the superposed backbone of the b82XLHbAmolecule on T-state HbA (Figure 2 and Table 2).De®nitive T-state interactions such as those invol-ving Arga1141(HC3), Lysa2127(H10) and Vala11and those between Arga1141(HC3) and theAspa2126(H9) carboxylate group are evident inb82XLHbA (Table 3).
Larger deviations from the T-state are observedbetween the b-chains of b82XLHbA. The rmsdifference between them is over 2.0 AÊ in the regionof the crosslink and involves the EF loops. Theremainder of b82XLHbA shows smaller but obser-vable differences from the T-state (Figure 2). Theconformation of Cysb93(F9) and the ``doming'' ofthe heme porphyrin are both analogous to theT-state. The critical inter-subunit, a1b2, interactionsinvolving Leua191(FG3) and Argb240(C6) are alsoT-state-like. So are the H-bonds that involve
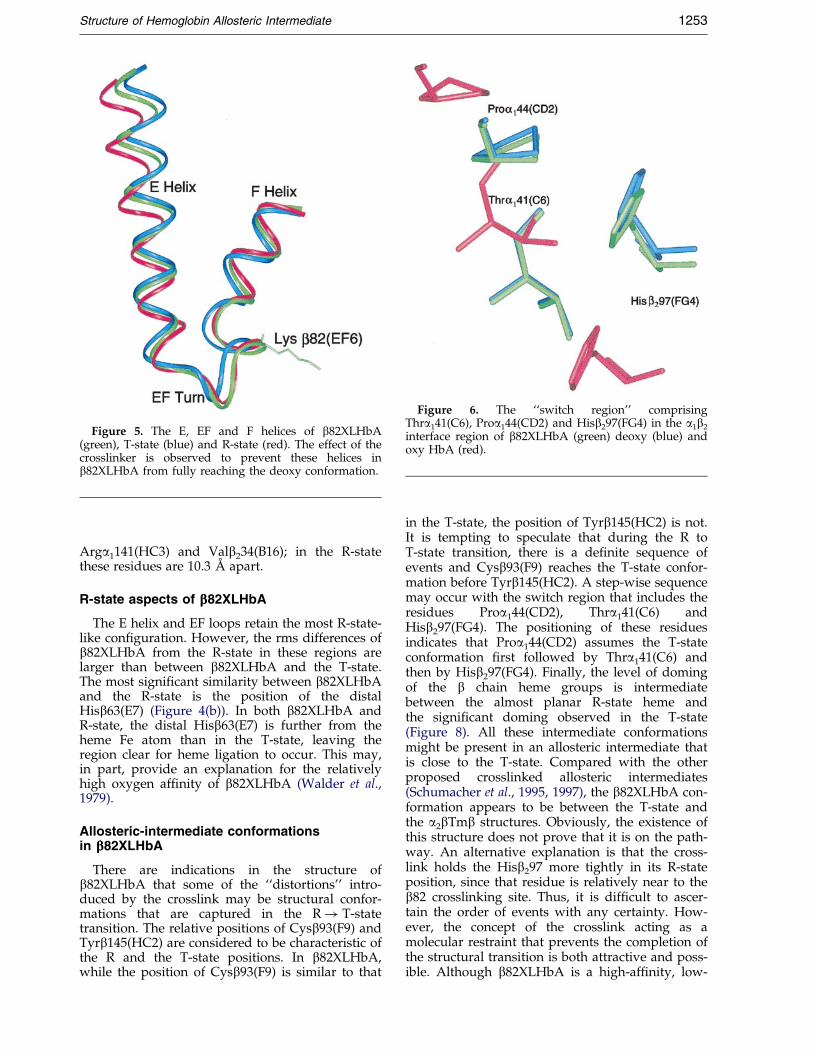
Figure 5. The E, EF and F helices of b82XLHbA(green), T-state (blue) and R-state (red). The effect of thecrosslinker is observed to prevent these helices inb82XLHbA from fully reaching the deoxy conformation.
Figure 6. The ``switch region'' comprisingThra141(C6), Proa144(CD2) and Hisb297(FG4) in the a1b2
interface region of b82XLHbA (green) deoxy (blue) andoxy HbA (red).
Structure of Hemoglobin Allosteric Intermediate 1253
Arga1141(HC3) and Valb234(B16); in the R-statethese residues are 10.3 AÊ apart.
R-state aspects of bbb82XLHbA
The E helix and EF loops retain the most R-state-like con®guration. However, the rms differences ofb82XLHbA from the R-state in these regions arelarger than between b82XLHbA and the T-state.The most signi®cant similarity between b82XLHbAand the R-state is the position of the distalHisb63(E7) (Figure 4(b)). In both b82XLHbA andR-state, the distal Hisb63(E7) is further from theheme Fe atom than in the T-state, leaving theregion clear for heme ligation to occur. This may,in part, provide an explanation for the relativelyhigh oxygen af®nity of b82XLHbA (Walder et al.,1979).
Allosteric-intermediate conformationsin bbb82XLHbA
There are indications in the structure ofb82XLHbA that some of the ``distortions'' intro-duced by the crosslink may be structural confor-mations that are captured in the R! T-statetransition. The relative positions of Cysb93(F9) andTyrb145(HC2) are considered to be characteristic ofthe R and the T-state positions. In b82XLHbA,while the position of Cysb93(F9) is similar to that
in the T-state, the position of Tyrb145(HC2) is not.It is tempting to speculate that during the R toT-state transition, there is a de®nite sequence ofevents and Cysb93(F9) reaches the T-state confor-mation before Tyrb145(HC2). A step-wise sequencemay occur with the switch region that includes theresidues Proa144(CD2), Thra141(C6) andHisb297(FG4). The positioning of these residuesindicates that Proa144(CD2) assumes the T-stateconformation ®rst followed by Thra141(C6) andthen by Hisb297(FG4). Finally, the level of domingof the b chain heme groups is intermediatebetween the almost planar R-state heme andthe signi®cant doming observed in the T-state(Figure 8). All these intermediate conformationsmight be present in an allosteric intermediate thatis close to the T-state. Compared with the otherproposed crosslinked allosteric intermediates(Schumacher et al., 1995, 1997), the b82XLHbA con-formation appears to be between the T-state andthe a2bTmb structures. Obviously, the existence ofthis structure does not prove that it is on the path-way. An alternative explanation is that the cross-link holds the Hisb297 more tightly in its R-stateposition, since that residue is relatively near to theb82 crosslinking site. Thus, it is dif®cult to ascer-tain the order of events with any certainty. How-ever, the concept of the crosslink acting as amolecular restraint that prevents the completion ofthe structural transition is both attractive and poss-ible. Although b82XLHbA is a high-af®nity, low-

Figure 7. The region of the a1b2 interface considered pivotal to the conformational R! T change. The residuesinvolved here are Aspa194(G1), Trpb237(C3) and Asnb2102(G4). Distances measured are AÊ between Asp Od2, Trp Ne1
and Asn Od1 atoms. It is evident from these distances that b82XLHbA lies in an intermediate state relative to the twonative hemoglobin molecules. In deoxy HbA, the closest atom is Asp94 Od1 and the interatom distances for deoxyHbA are calculated using this atom.
1254 Structure of Hemoglobin Allosteric Intermediate
cooperativity hemoglobin, our results clearlydemonstrate that it does accomplish most of theallosteric transition, since the structure is primarilyT-state. Thus, the fact that the cooperativity isreduced does not eliminate the possibility thatthis could be an intermediate along the allostericpathway.
Methods
Isolation and crosslinking of hemoglobin
HbA was isolated from whole blood by the methoddescribed by Dozy et al. (1968) and puri®ed by anion-exchange chromatography on a DEAE Sephadex A-50
Figure 8. The orientation of the proximal Hisb92(F8)and the F helix relative to the heme. The gradual dom-ing of the heme in b82XLHbA (green) is observedclearly. Also shown is T-state (blue) and R-state (red).
column. The reaction of HbA with bis(3,5-dibromosali-cyl) fumarate to crosslink the two Lys-b82 residues wasperformed under oxygenated conditions as described byWalder et al. (1979). The b82XLHbA product was alsoisolated using the anion-exchange chromatography onSephadex A-50.
Crystallization, data collection, structure solutionand refinement
b82XLHbA was crystallized in the deoxy state fromsolutions of PEG-6000 by the batch method. Protein,PEG-6000, and salt concentrations were as those pro-posed by Ward et al. (1975). The vials also contained4 mM sodium dithionite and were prepared in a nitro-gen-®lled glove-box.
Although the crystals of b82XLHbA were grown fromPEG solutions under conditions similar to those usedpreviously (Ward et al., 1975), the unit cell was differentfrom those previously observed. Precession photography(data not shown) and the auto-indexing routine of the R-axis software (RAXIS-II Data Processing Software, VMSversion 1.0, Rigaku Corporation 1990, Tokyo, Japan.)both supported a P21 unit cell that differed from otherhemoglobin crystal packings (Table 5). This crystal formhas two tetramers in the asymmetric unit of the crystal,with a total of four tetramers b82XLHbA in the unit cell.Data extended to 2.20 AÊ resolution.
Diffraction data were collected at the R-Axis II imageplate detector facility at Abbott Laboratories, AbbottPark, IL. Data collected were limited to re¯ections thatwere at least 2.5s (42,577 re¯ections). Unit cell par-ameters and data reduction statistics are listed in Table 5.
The asymmetric unit of the b82XLHbA unit cell con-tains two tetramers (P21, b-unique axis). The structurewas solved by molecular replacement (Rossmann, 1990)with the program package X-PLOR (Brunger, 1990a,b)using data between 8.0 AÊ and 4.5 AÊ . The search modelwas a 1.74 AÊ re®ned tetramer of deoxy, T-state, HbA(Fermi et al., 1984).
After determining the relative orientations of the twob82XLHbA tetramers in the asymmetric unit by a self-rotation function, the search model was oriented on eachof the target molecules by calculating a cross-rotationfunction. To position the correctly oriented search model,a translation function was calculated. Three independent

Table 5. Unit cell constants and data reduction statisticsfor b82XLHbA
Cell constantsSpace group P21Unit cell dimensions (AÊ ) a � 65.3, b � 96.2, c � 101.1,
b � 101.5 �Volume (AÊ 3) 6.2 � 105
Molecules/asymmetric unit 2 tetramersData collection statistics
Resolution range (AÊ ) 53.20-2.00Number of reflectionsmerging Bijvoets 53,187Ave. I/s(I) at 2.00 AÊ 2.02Ave. I/s(I) at 2.30 AÊ 2.60Number of observed
reflections >2.5 � s42,577
Number of theoreticalreflections
58,219
Rsym 3.97 %Refinement statistics
Refinement (AÊ ) 10.0-2.3R-factor/Rfree (%) 19.0/26.3Water molecules/tetramer 187
rms differencesTwo molecules in A.U. (AÊ ) 0.98Bond distances (AÊ ) 0.012Bond angles (deg.) 1.877
Structure of Hemoglobin Allosteric Intermediate 1255
translation searches were done. The ®rst search was per-formed in the x and z directions to position the searchmodel on one b82XLHbA molecule. A second search inthe x and z directions positioned the search model on thesecond b82XLHbA in the asymmetric unit. The thirdsearch determined the relative y coordinates of the twob82XLHbA molecules.
The b82XLHbA model was re®ned against theobserved data using the X-PLOR program package(Brunger, 1990a,b). The molecular replacement modelwas ®rst re®ned by rigid body re®nement by treatingeach molecule in the asymmetric unit as an independentrigid body. Re®nement was started with data between15.0 AÊ and 4.5 AÊ . The resolution was increased in ®vestages to 2.3 AÊ (3.50 AÊ , 3.00 AÊ , 2.70 AÊ , 2.50 AÊ , 2.30 AÊ ).The R-value dropped from an initial value of 0.48 to 0.32after rigid body minimization. The model was thenre®ned by the conjugate gradient method. After threecycles of re®nement, with the R-value at 0.27, differenceelectron density maps were calculated. These electrondensity maps were viewed using the program CHAIN(Sack, 1988) running on a Silicon Graphics Indigo. Errorsin the model were corrected manually. Also, water mol-ecules were added to the model at this stage. A total of374 water molecules were added to the asymmetric unitthat contained the two b82XLHbA molecules. Finally,the fumarate crosslink was built into difference electrondensity between the Nz atoms of the two Lysb82 resi-dues. Amide bonds were speci®ed between the Lysb82residues and the fumarate crosslink in the X-PLORparameter library. The R-value at this stage was 0.24.Re®nement of this model by simulated annealingresulted in an R-value of 0.20 and one round of re®ne-ment of the temperature factors brought the R-value to0.19. The Luzzati error for b82XLHbA is 0.25 AÊ .
Comparison of crystallographic structures
The comparisons of b82XLHbA with deoxy (2HHB)and oxy (1HHO) hemoglobin molecules used structures
determined by Fermi et al. (1984) and Shaanan (1983),respectively. The percentage of T-state character was cal-culated by ®rst superimposing the three structures. Thenthe distance between the identical atom in the oxy andb82 crosslinked structures was divided by the same dis-tance between the oxy and deoxy structures to obtain anestimate of the fraction of the transition that atom hadundergone. These fractional values were averaged forthose residues being considered and converted to a per-centage.
RCSB Protein Data Bank accession numbers
The atomic coordinates of b82XLHbA have beendeposited with the RCSB Protein Data Bank under the®lename 1BIJ.
Acknowledgements
This work was supported, in part, by a grant from theResearch Corporation.
References
Baldwin, J. & Chothia, C. (1979). Haemoglobin: thestructural changes related to ligand binding and itsallosteric mechanism. J. Mol. Biol. 129, 175-220.
Brunger, A. T. (1990a). Extension of molecular replace-ment: a new search strategy based on Pattersoncorrelation re®nement. Acta Crystallog. sect. A, 46,46-57.
Brunger, A. T. (1990b). X-PLOR Manual, Version 2.1,Yale University, New Haven, CT.
Brzowowski, A., Derewenda, Z., Dodson, E., Dodson,G., Grabowoski, M. J., Liddington, R., Skarzynski,T. & Valley, D. (1984). Bonding of molecular oxygento T state human hemoglobin. Nature, 307, 74-76.
Dozy, A. M., Kleihauer, E. F. & Huisman, T. H. J.(1968). Studies on the heterogeneity of hemoglobin.13. Chromatography of various human and animalhemoglobin types on DEAE-Sephadex. J. Chromatog.32, 723-727.
Fermi, G., Perutz, M. F., Shaanan, B. & Forme, R. (1984).The crystal structure of human deoxyhaemoglobinat 1.74 AÊ resolution. J. Mol. Biol. 175, 159-174.
Grabowski, M. J., Brzowowski, A. M., Derewenda, Z. S.,Skarzynski, T., Glycer, M., Stepien, S. &Derewenda, A. (1978). Crystallization of humanoxyhaemoglobin from poly (ethylene glycol) sol-utions. Biochem. J. 171, 227-279.
Liddington, R. C., Derewenda, Z., Dodson, G. & Harris,G. (1988). Structure of the liganded T-state ofhaemoglobin identi®es the origins of cooperativeoxygen binding. Nature, 331, 725-728.
Liddington, R. C., Derewenda, Z., Dodson, G., Dodson,E., Hubbard, R. & Dodson, G. (1992). High resol-ution crystal structures and comparisons of T statedeoxy and two liganded T state haemoglobins: T(a-oxy) haemoglobin and T (met) haemoglobin.J. Mol. Biol. 228, 551-579.
Luisi, B., Liddington, R., Fermi, G. & Shibayama, N.(1990). Structure of deoxy quaternary haemoglobinwith liganded b subunits. J. Mol. Biol. 214, 7-14.
Monod, J., Wyman, J. & Changeux, J.-P. (1963). On thenature of allosteric transitions: a plausible model.J. Mol. Biol. 12, 88-118.

1256 Structure of Hemoglobin Allosteric Intermediate
Mozzarelli, A., Rivetti, C., Rossi, G. L., Henry, E. R. &Eaton, W. A. (1990). Crystals of haemoglobin withthe T quaternary structure bind oxygen noncoo-peratively with no Bohr effect. Nature, 351, 416-419.
Paoli, M., Liddington, R., Tame, J., Wilkinson, A.& Dodson, G. (1996). Crystal structure of T statehaemoglobin with oxygen bound at all four haems.J. Mol. Biol. 256, 775-792.
Perutz, M. F., Wilkinson, A. J., Paoli, M. & Dodson,G. G. (1998). The stereochemical mechanism of thecooperative effects in hemoglobin revisted. Annu.Rev. Biophys. Biomol. Struct. 27, 1-34.
Rossmann, M. J. (1990). The molecular replacementmethod. Acta Crystallog. sect. A, 46, 73-82.
Sack, J. S. (1988). CHAIN - a crystallographic modelingprogram. J. Mol. Graph. 6, 224-225.
Schumacher, M. A., Dixon, M. M., Kluger, R., Jones,R. T. & Brennan, R. G. (1995). Allosteric transitionintermediates modelled by crosslinked haemo-globins. Nature, 375, 84-87.
Schumacher, M. A., Zheleznova, E. E., Poundstone, K. S.,Kluger, R., Jones, R. T. & Brennan, R. G. (1997).Allosteric intermediates indicate R2 is the ligandedhemoglobin end state. Proc. Natl Acad. Sci. USA, 94,7841-7844.
Shaanan, B. (1983). Structure of oxyhaemoglobin at2.1 AÊ resolution. J. Mol. Biol. 171, 31-59.
Silva, M. M., Rogers, P. H. & Arnone, A. (1992). A thirdquaternary structure of human hemoglobin A at1.7 AÊ resolution. J. Biol. Chem. 267, 17248-17256.
Walder, J. A., Zaugg, R. H., Walder, R. Y., Steele, J. M.& Klotz, I. M. (1979). Diaspirins that cross-link b
chains of hemoglobin: bis(3,5-dibromosalicyl) succi-nate and bis(3,5-dibromosalicyl) fumarate. Biochem-istry, 18, 4265-4270.
Walder, J. A., Walder, R. Y. & Arnone, A. (1980). Devel-opment of antisickling compounds that chemicallymodify hemoglobin S speci®cally within the 2,3-diphosohoglycerate binding site. J. Mol. Biol. 141,195-216.
Ward, K. B., Wishner, B. C., Lattman, E. & Love, W. E.(1975). Structure of deoxyhaemoglobin A grownfrom polyethylene glycol solutions. J. Mol. Biol. 98,161-177.
White, F. L. & Olsen, K. W. (1987). Effects of cross-linking on the thermal stability of hemoglobin. I.The use of bis(3,5-dibromosalicyl) fumarate). Arch.Biochem. Biophys. 258, 51-57.
Wilson, J., Phillips, K. & Luisi, B. (1996). The crystalstructure of horse deoxyhaemoglobin trapped in thehigh-af®nity (R) state. J. Mol. Biol. 264, 743-756.
Winslow, R. (1992). The modi®cation of hemoglobin. InHemoglobin Based Blood Substitutes, pp. 72-97, TheJohns Hopkins Press, London.
Yang, T. & Olsen, K. W. (1991). Thermal stability ofhemoglobin crosslinked in the T-state by bis(3,5-dibromosalicyl) fumarate. Biochem. Biophys. Res.Commun. 174, 518-523.
Yang, T. & Olsen, K. W. (1989). The effect of cross-linking by bis(3,5-dibromosalicyl) fumarate on theautoxidation of hemoglobin. Biochem. Biophys. Res.Commun. 163, 733-738.
Edited by K. Nagai
(Received 20 September 1999; received in revised form 10 January 2000; accepted 18 January 2000)





























