Embed Size (px)
Citation preview

RESEARCH ARTICLE
Developmental Potential of Gcn5�/� EmbryonicStem Cells In Vivo and In VitroWenchu Lin,† Geraldine Srajer,† Yvonne A. Evrard,‡ Huy M. Phan,§ Yas Furuta, andSharon Y.R. Dent*
Gcn5 is a prototypical histone acetyltransferase (HAT) that serves as a coactivator for multiple DNA-boundtranscription factors. We previously determined that deletion of Gcn512 (hereafter referred to as Gcn5)causes embryonic lethality in mice. Gcn5 null embryos undergo gastrulation but exhibit high levels ofapoptosis, leading to loss of mesodermal lineages. To further define the functions of Gcn5 duringdevelopment, we created Gcn5�/� mouse embryonic stem (ES) cells. These cells survived in vitro and formedembryoid bodies (EBs) that expressed markers for ectodermal, mesodermal, and endodermal lineages.Gcn5�/� EBs were misshapen and smaller than wild-type EBs by day 6, with an increased proportion of cellsin G2/M. Expression of Oct 4 and Nodal was prematurely curtailed in Gcn5�/� EBs, indicating early loss ofpluripotent ES cells. Gcn5�/� EBs differentiated efficiently into skeletal and cardiac muscle, which derivefrom mesoderm. High percentage Gcn5�/� chimeric embryos created by injection of Gcn5�/� ES cells intowild-type blastocysts were delayed in development and died early. Interestingly, elevated levels ofapoptosis were observed specifically in Gcn5 null cells within the chimeric embryos. Collectively, these dataindicate that Gcn5 may be required to maintain pluripotent states and that loss of Gcn5 invokes a cell-autonomous pathway of cell death in vivo. Developmental Dynamics 236:1547–1557, 2007.© 2007 Wiley-Liss, Inc.
Key words: chromatin; acetyltransferase; histone; stem cell; embryo; Gcn5; PCAF
Accepted 21 March 2007
INTRODUCTIONDifferentiation of pluripotent embry-onic stem cells into specialized celltypes and tissue lineages is driven byspecific transcription factors that or-chestrate changes in gene expressionprograms (Boiani and Scholer, 2005).During this process, chromatin is re-modeled into heritable states that ei-ther allow activation or maintain re-pression of tissue- and lineage-specific
genes (Bernstein et al., 2006). Theseepigenetic states are established andcontrolled largely by specific patternsof histone posttranslational modifica-tions (Li, 2002).
Histones are subject to lysine acet-ylation, lysine and arginine methyl-ation, serine/threonine phosphoryla-tion, and lysine ubiquitylation, amongother modifications (Zhang and Rein-berg, 2001). Each of these modifica-
tions alters chromatin structurethrough effects on histone: DNA, his-tone:histone, and nucleosome:nucleo-some interactions (Ura et al., 1997;Annunziato and Hansen, 2000). In ad-dition, histone modifications affect thebinding of nonhistone proteins tochromatin, including transcriptionfactors and additional chromatin re-modeling activities (Winston and Al-lis, 1999; Jenuwein and Allis, 2001;
Program in Genes and Development, Department of Biochemistry and Molecular Biology, University of Texas M.D. Anderson Cancer Center,Houston, TexasGrant sponsor: NIH; Grant number: GM067718.†Drs. Lin and Srajer contributed equally to this work.‡Dr. Evrard’s present address is Laboratory of Molecular Biology, National Institute of Diabetes and Digestive and Kidney Diseases,National Institutes of Health, Bethesda, MD 20892.§Dr. Phan’s present address is University of Arizona Health Sciences Center, Tuscon, Arizona 85721.*Correspondence to: Sharon Y.R. Dent (formerly known as Sharon Y. Roth), Department of Biochemistry and MolecularBiology, Unit 1000, U.T. M.D. Anderson Cancer Center, 1515 Holcombe Blvd., Houston, TX. E-mail: [email protected]
DOI 10.1002/dvdy.21160Published online 17 April 2007 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 236:1547–1557, 2007
© 2007 Wiley-Liss, Inc.

Kim et al., 2006). Different patterns ofmodifications have been proposed toconstitute an informational “code”that is read by other proteins and thatmay be passed from mother to daugh-ter cells as cells divide (Jenuwein andAllis, 2001).
Histone acetylation has long beenassociated with gene activation,whereas histone deacetylation is asso-ciated with gene repression. Thesestates are governed by a balance inthe activities of histone acetyltrans-ferases (HATs) and histone deacety-lases (HDACs). Although these en-zymes have been identified ascoactivators or corepressors, respec-tively, for several transcription factorsimportant to mammalian develop-ment, the functions of specific HATsand HDACs during embryogenesis arestill poorly defined. Only a few genedeletion studies have been done to de-fine HAT or HDAC functions duringmouse development. Data so far indi-cate that individual HATs andHDACs have specialized functions incell and tissue differentiation. Class IIHDACs (HDAC9 and HDAC5), for ex-ample, limit stress-induced cardiachypertrophy (McKinsey et al., 2001;Zhang et al., 2002; Song et al., 2006),whereas HDAC4 limits chondrocytehypertrophy and endochondral boneformation (Vega et al., 2004). Loss ofeither of p300 or CBP, two highly re-lated HATs, leads to mid-gestationembryonic death (Yao et al., 1998). Al-though these HATs share many func-tions, p300 is required for proper neu-rulation and heart development (Yaoet al., 1998), whereas CBP is involvedin hematopoiesis and skeletal devel-opment (Kung et al., 2000). Interest-ingly, p300 and CBP interact physi-cally with two other highly relatedHATs: Gcn5 and PCAF (Yang et al.,1996; Xu et al., 1998). Deletion ofPCAF causes no overt abnormalitiesin mouse embryos or adults, but dele-tion of Gcn5 causes early embryonicdeath (Xu et al., 2000; Yamauchi etal., 2000). Gcn5 null embryos exhibitincreased apoptosis relative to theirlittermates, leading to a loss of meso-dermal tissues (Xu et al., 2000). Miceheterozygous for the Gcn5 null alleleare viable, as are most mice heterozy-gous for a p300 deletion. However, asubstantial portion of mice heterozy-gous both Gcn5 and p300 null alleles
die in utero, indicating that a criticaldosage of these two HATs must bemaintained during development (Phanet al., 2005).
To further define Gcn5 functions indevelopmental processes, we createdmouse embryonic stem cells that lackthis HAT gene. We then determinedthe developmental potential of thesecells in vitro and in vivo. Our dataindicate that Gcn5 is not required forsurvival or differentiation of embry-onic stem (ES) cells into embryoidbodies (EBs) or muscle lineages invitro. However, Gcn5 null cells un-dergo cell-autonomous apoptosis inchimeric embryos, leading to earlyembryo death when present in highnumbers.
RESULTS
Creation of Gcn5 Null ESCells
To circumvent the early lethality ofGcn5 null embryos, we generated anallele of Gcn5 (Gcn5flox[neo]) in whichexons 3–19 were flanked by site-specificloxP recombination signal sequences(Fig. 1A) positioned in noncoding re-gions. Cre-mediated recombination be-tween the 5�-most and the 3�-most loxPsequences gives rise to a deletion allele(Gcn5�ex3-19) that lacks most of theGcn5 coding region, including the HATdomain and the bromodomain (Xu etal., 1998). We successfully introducedthe Gcn5flox(neo) allele into mouse em-bryonic stem (ES) cells, replacing onecopy of the Gcn5l2 (GCN5 general con-trol of amino acid synthesis-like 2; here-after called Gcn5) gene (Fig. 1B, leftpanel). Exons 3–19 and the neomycinmarker were removed from this alleleby transfection of the ES cells with aCre recombinase expression vector (Fig.1B, middle panel). We then used ouroriginal targeting vector (Xu et al.,2000) to delete the second allele of Gcn5in these cells, giving rise to the Gcn5�
allele (Fig. 1A,B). We confirmed the re-moval of both wild-type alleles of Gcn5by Southern blot (Fig. 1B, right panel).At least two independent Gcn5�/�ex3-19
null ES clones (hereafter referred to asGcn5 null or Gcn5�/�) were analyzed,and in all cases independent clonesgave identical results. Northern analy-ses confirmed that Gcn5 expression isabrogated in these cells (Fig. 1C). Im-
portantly, PCAF expression levels werenot affected by Gcn5 loss (data notshown).
Absence of Growth Defectsin Undifferentiated Gcn5Null ES Cells
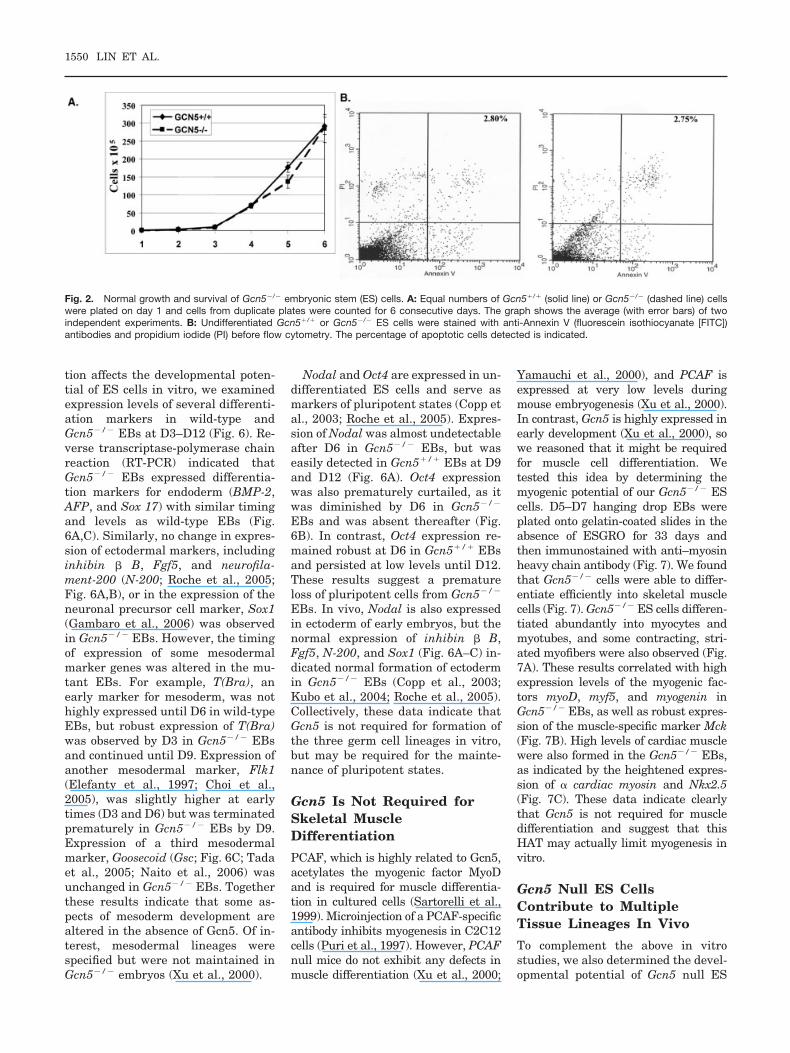
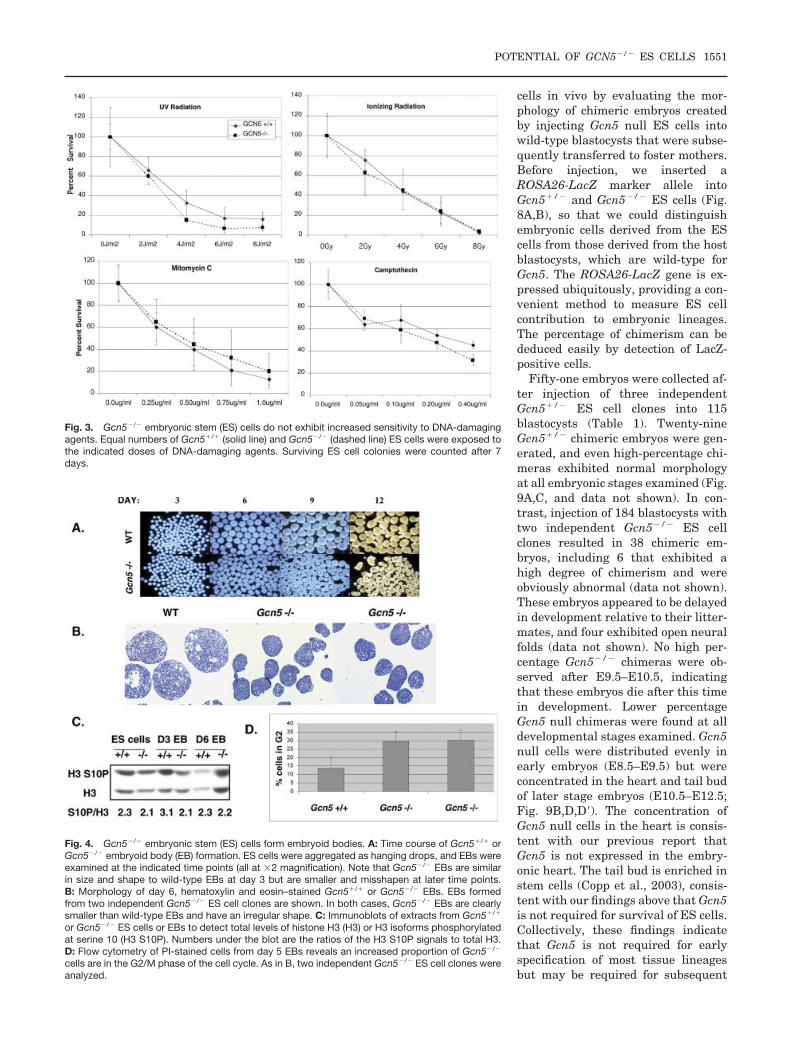
Gcn5 serves as a coactivator for sev-eral transcription factors in yeast andmammalian cells (Roth et al., 2001).However, the survival of Gcn5 null EScells indicates that the loss of Gcn5does not result in a global failure intranscription, consistent with our pre-vious studies (Xu et al., 2000) as wellas previous studies in yeast (Lee et al.,2000). Gcn5 null ES cells grow anddivide normally (Fig. 2A). No increasein apoptosis as measured by annexinV and propidium iodide (PI) stainingwas observed in undifferentiatedGcn5 null ES cells in vitro (Fig. 2B)relative to wild-type cells, in contrastto Gcn5 null embryos (Xu et al., 2000).The null ES cells were generally nomore sensitive to a variety of DNA-damaging agents than were wild-typecells (Fig. 3). The slight differences insensitivity to some doses of ultravioletor Camptothecin seen in these exper-iments are not statistically signifi-cant, as determined by unpairedt-tests. These data indicate that Gcn5is not required for cell division, cellsurvival, or DNA damage responses inundifferentiated ES cells. However,we cannot rule out that small differ-ences observed in vitro would have im-portant developmental consequencesin vivo.
Gcn5 Is Not Required forEmbryoid Body Formation
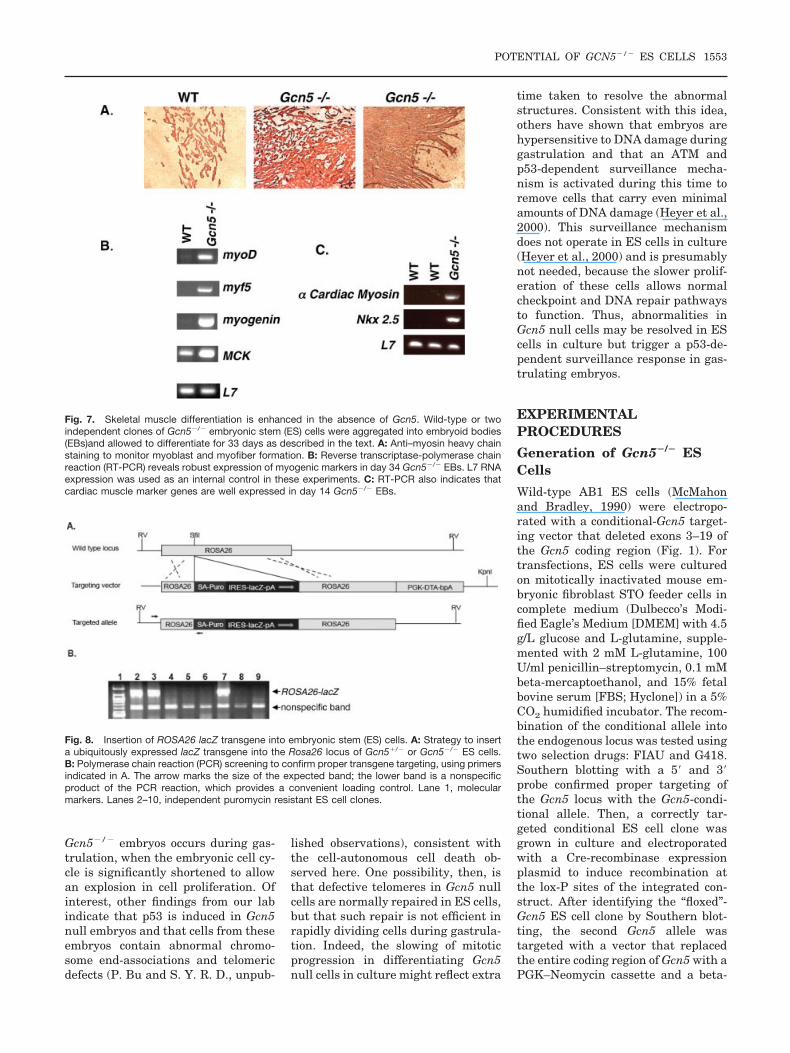
To study the in vitro differentiationpotential of Gcn5�/� ES cells, weplated wild-type or null ES cells ashanging drops in the absence of LIF sothat they would spontaneously formEBs. At day 3 (D3) of EB differentia-tion, Gcn5�/� EBs were morphologi-cally indistinguishable from Gcn5�/�
EBs (Fig. 4A). However, at D6Gcn5�/� EBs were noticeably smallerin size than Gcn5�/� EBs (Fig. 4A,B).Gcn5�/� EBs remained small at D9and D12 of EB differentiation (Fig.4A). In addition, Gcn5�/� EBs ap-peared more irregularly shaped andmore dense than Gcn5�/� EBs (Fig.
1548 LIN ET AL.

4B). To determine whether the smallsize and irregular shape of Gcn5�/�
EBs were due to increased apoptosis,D11 EBs were sectioned and subjectedto terminal deoxynucleotidyl trans-ferase–mediated deoxyuridinetriphos-phate nick end-labeling (TUNEL)analyses. Both Gcn5�/� and Gcn5�/�
EBs contained a large number of apo-ptotic cells, but no significant increasein apoptosis was detected in theGcn5�/� EBs (data not shown). Flowcytometry analyses of D4 and D5 EBsstained with PI and anti-annexin V
antisera also failed to reveal increasedlevels of apoptosis in Gcn5�/� EBs(data not shown). To monitor cell pro-liferation levels, sections of D11Gcn5�/� and Gcn5�/� EBs werestained with an antibody specific forphosphorylated serine 10 (S10) of his-tone H3, which is a marker for mitoticcells. Low levels of H3 S10 stainingwere observed in both Gcn5�/� andGcn5�/� EBs, but no significant de-crease in H3 S10 phosphorylation wasobserved in sections of Gcn5�/� EBs(data not shown), indicating that the
small size of these EBS is not due to afailure in mitotic entry. Western blotsof protein extracts from Gcn5�/� andGcn5�/� ES cells and EBs also indi-cated no major changes in H3 S10phosphorylation levels in the absenceof Gcn5, although H3 S10 phosphory-lation was slightly increased in wild-type, but not Gcn5�/�, EBs at D3(Fig. 4C). The significance of thissmall change is not clear, but it mightindicate fewer mitotic cells in the mu-tant EBs at this time point. Flow cy-tometry of PI-stained cells from D5EBs revealed an enrichment ofGcn5�/� cells with a G2 content ofDNA (Fig. 4D). Of interest, we previ-ously determined that gcn5� yeastcells accumulate in G2/M phase(Zhang et al., 1998). Together, theseresults indicate that Gcn5 is requiredfor efficient mitotic progression.
Decreased Acetylation inGcn5 Null ES Cells DuringDifferentiation
Gcn5 acetylates histones H3 and H2B(Grant et al., 1997, 1999). H3 acetyla-tion is decreased approximately two-fold in undifferentiated Gcn5�/� cells(data not shown), indicating thatGcn5 is a major H3 HAT activity inthese cells, but it is not the only one.To determine whether Gcn5 HATfunctions become more critical duringdifferentiation, H3 acetylation wasevaluated in wild-type and Gcn5�/�
EBs at D3 and D6. After normaliza-tion for the total amount of H3present, immunoblots revealed thatacetylation of H3 at K9 and K18 wasdecreased approximately threefold inboth D3 and D6 Gcn5�/� EBs relativeto wild-type EBs. H3 K9, 18 acetyla-tion dropped five- to sixfold in bothwild-type and Gcn5�/� null EBs be-tween D3 and D6 (Fig. 5). By D6, then,H3 acetylation at these sites was 17-fold lower in the mutant EBs than inwild-type D3 EBs (Fig. 5). These dataindicate that Gcn5 is a major H3 HATin these differentiating cells.
Gcn5 Null ES CellsDifferentiate Into Ectoderm,Mesoderm, and EndodermIn Vitro
To determine whether the loss of Gcn5and subsequent decreased H3 acetyla-
Fig. 1. Creation of Gcn5�/� embryonic stem (ES) cells. A: Targeting strategy to introduce loxPsites into the Gcn5 locus and resulting Gcn5 alleles. A PGK neo cassette flanked by loxP sites wasintroduced into the second intron, and a third loxP site was inserted downstream of exon 19 of theGcn5 gene. Integration of this vector at the Gcn5 locus creates the Gcn5flox(neo) allele. The locationsof restriction sites used for vector linearization (PmeI) and for screening (EcoRV) are indicated as arethe locations of 5� and 3� probes used in Southern blots. Sizes of EcoRV restriction productsdetected by these probes are shown to the right. Cre-mediated recombination generates theGcn5flox, Gcn5�ex3-19(neo), and the Gcn5�ex3-19 alleles. The Gcn5� was created as described previ-ously (Xu et al., 2000). B: Southern blots to confirm correct targeting of the vector shown in A andsubsequent generation of Gcn5�/� ES cells. Left panel: The 5� probe detects a 7-kb wild-type bandand a 9-kb band from the targeted allele, and the 3� probe detects a unique 5.5-kb band from thetargeted allele. Although multiple targeted ES cell lines were obtained, only one is shown here.Middle panel: ES cells carrying the Gcn5flox(neo) allele were exposed to a Cre-recombinase expres-sion vector to create the Gcn5�ex3-19 allele, identified by the creation of a 5.5-kb EcoRV digestionproduct detected by the 5� probe shown in A and loss of the 9-kb band. The clone marked �/*carries at least one wild-type allele and may carry either a second wild-type allele, the Gcn5flox
allele, or the Gcn5�ex3-19 (neo) allele, all of which give rise to a 7-kb band. Right panel: The Gcn5�
allele was introduced into ES cells carrying the Gcn5�ex3-19 allele to create Gcn5�/�ex3-19 cells,hereafter referred to as Gcn5�/�. The Gcn5� allele gives rise to a 4-kb band detected by the 5�probe and an 8.0-kb band detected by the 3� probe, as indicated in A. C: Northern blot to confirmthat Gcn5 is not expressed in Gcn5�/� ES cells. Cyclophilin was used as a loading control.
POTENTIAL OF GCN5�/� ES CELLS 1549
tion affects the developmental poten-tial of ES cells in vitro, we examinedexpression levels of several differenti-ation markers in wild-type andGcn5�/� EBs at D3–D12 (Fig. 6). Re-verse transcriptase-polymerase chainreaction (RT-PCR) indicated thatGcn5�/� EBs expressed differentia-tion markers for endoderm (BMP-2,AFP, and Sox 17) with similar timingand levels as wild-type EBs (Fig.6A,C). Similarly, no change in expres-sion of ectodermal markers, includinginhibin � B, Fgf5, and neurofila-ment-200 (N-200; Roche et al., 2005;Fig. 6A,B), or in the expression of theneuronal precursor cell marker, Sox1(Gambaro et al., 2006) was observedin Gcn5�/� EBs. However, the timingof expression of some mesodermalmarker genes was altered in the mu-tant EBs. For example, T(Bra), anearly marker for mesoderm, was nothighly expressed until D6 in wild-typeEBs, but robust expression of T(Bra)was observed by D3 in Gcn5�/� EBsand continued until D9. Expression ofanother mesodermal marker, Flk1(Elefanty et al., 1997; Choi et al.,2005), was slightly higher at earlytimes (D3 and D6) but was terminatedprematurely in Gcn5�/� EBs by D9.Expression of a third mesodermalmarker, Goosecoid (Gsc; Fig. 6C; Tadaet al., 2005; Naito et al., 2006) wasunchanged in Gcn5�/� EBs. Togetherthese results indicate that some as-pects of mesoderm development arealtered in the absence of Gcn5. Of in-terest, mesodermal lineages werespecified but were not maintained inGcn5�/� embryos (Xu et al., 2000).
Nodal and Oct4 are expressed in un-differentiated ES cells and serve asmarkers of pluripotent states (Copp etal., 2003; Roche et al., 2005). Expres-sion of Nodal was almost undetectableafter D6 in Gcn5�/� EBs, but waseasily detected in Gcn5�/� EBs at D9and D12 (Fig. 6A). Oct4 expressionwas also prematurely curtailed, as itwas diminished by D6 in Gcn5�/�
EBs and was absent thereafter (Fig.6B). In contrast, Oct4 expression re-mained robust at D6 in Gcn5�/� EBsand persisted at low levels until D12.These results suggest a prematureloss of pluripotent cells from Gcn5�/�
EBs. In vivo, Nodal is also expressedin ectoderm of early embryos, but thenormal expression of inhibin � B,Fgf5, N-200, and Sox1 (Fig. 6A–C) in-dicated normal formation of ectodermin Gcn5�/� EBs (Copp et al., 2003;Kubo et al., 2004; Roche et al., 2005).Collectively, these data indicate thatGcn5 is not required for formation ofthe three germ cell lineages in vitro,but may be required for the mainte-nance of pluripotent states.
Gcn5 Is Not Required forSkeletal MuscleDifferentiation
PCAF, which is highly related to Gcn5,acetylates the myogenic factor MyoDand is required for muscle differentia-tion in cultured cells (Sartorelli et al.,1999). Microinjection of a PCAF-specificantibody inhibits myogenesis in C2C12cells (Puri et al., 1997). However, PCAFnull mice do not exhibit any defects inmuscle differentiation (Xu et al., 2000;
Yamauchi et al., 2000), and PCAF isexpressed at very low levels duringmouse embryogenesis (Xu et al., 2000).In contrast, Gcn5 is highly expressed inearly development (Xu et al., 2000), sowe reasoned that it might be requiredfor muscle cell differentiation. Wetested this idea by determining themyogenic potential of our Gcn5�/� EScells. D5–D7 hanging drop EBs wereplated onto gelatin-coated slides in theabsence of ESGRO for 33 days andthen immunostained with anti–myosinheavy chain antibody (Fig. 7). We foundthat Gcn5�/� cells were able to differ-entiate efficiently into skeletal musclecells (Fig. 7). Gcn5�/� ES cells differen-tiated abundantly into myocytes andmyotubes, and some contracting, stri-ated myofibers were also observed (Fig.7A). These results correlated with highexpression levels of the myogenic fac-tors myoD, myf5, and myogenin inGcn5�/� EBs, as well as robust expres-sion of the muscle-specific marker Mck(Fig. 7B). High levels of cardiac musclewere also formed in the Gcn5�/� EBs,as indicated by the heightened expres-sion of � cardiac myosin and Nkx2.5(Fig. 7C). These data indicate clearlythat Gcn5 is not required for muscledifferentiation and suggest that thisHAT may actually limit myogenesis invitro.
Gcn5 Null ES CellsContribute to MultipleTissue Lineages In Vivo
To complement the above in vitrostudies, we also determined the devel-opmental potential of Gcn5 null ES
Fig. 2. Normal growth and survival of Gcn5�/� embryonic stem (ES) cells. A: Equal numbers of Gcn5�/� (solid line) or Gcn5�/� (dashed line) cellswere plated on day 1 and cells from duplicate plates were counted for 6 consecutive days. The graph shows the average (with error bars) of twoindependent experiments. B: Undifferentiated Gcn5�/� or Gcn5�/� ES cells were stained with anti-Annexin V (fluorescein isothiocyanate [FITC])antibodies and propidium iodide (PI) before flow cytometry. The percentage of apoptotic cells detected is indicated.
1550 LIN ET AL.
cells in vivo by evaluating the mor-phology of chimeric embryos createdby injecting Gcn5 null ES cells intowild-type blastocysts that were subse-quently transferred to foster mothers.Before injection, we inserted aROSA26-LacZ marker allele intoGcn5�/� and Gcn5�/� ES cells (Fig.8A,B), so that we could distinguishembryonic cells derived from the EScells from those derived from the hostblastocysts, which are wild-type forGcn5. The ROSA26-LacZ gene is ex-pressed ubiquitously, providing a con-venient method to measure ES cellcontribution to embryonic lineages.The percentage of chimerism can bededuced easily by detection of LacZ-positive cells.
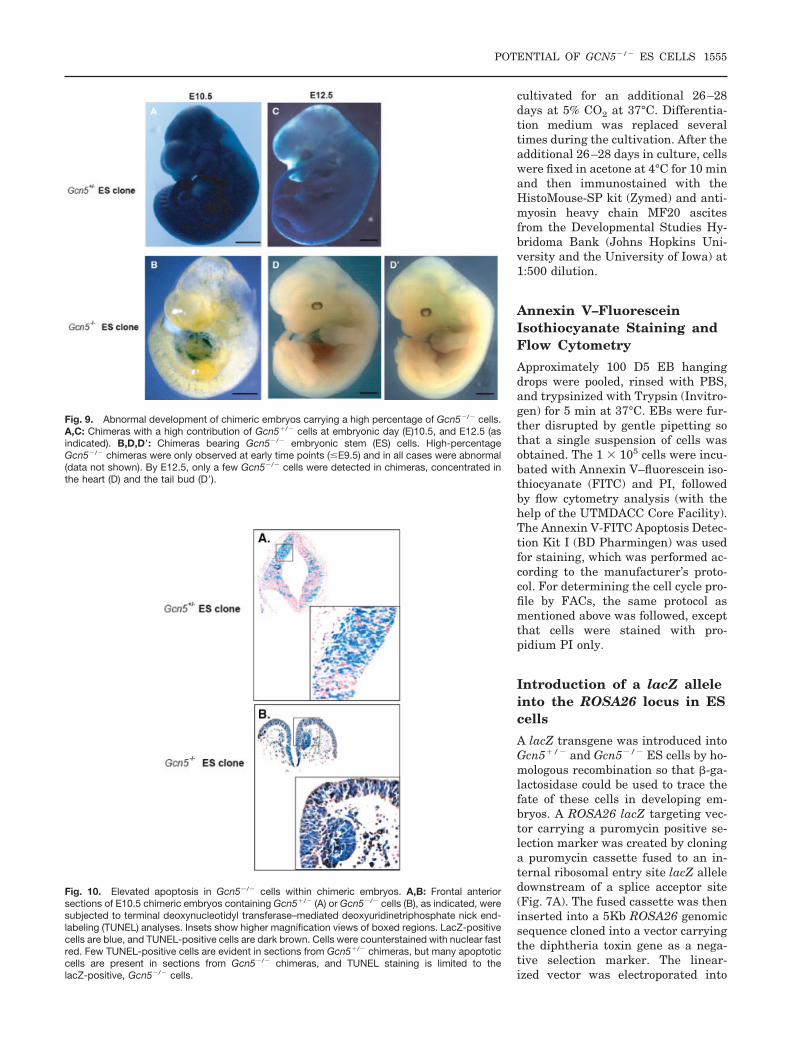
Fifty-one embryos were collected af-ter injection of three independentGcn5�/� ES cell clones into 115blastocysts (Table 1). Twenty-nineGcn5�/� chimeric embryos were gen-erated, and even high-percentage chi-meras exhibited normal morphologyat all embryonic stages examined (Fig.9A,C, and data not shown). In con-trast, injection of 184 blastocysts withtwo independent Gcn5�/� ES cellclones resulted in 38 chimeric em-bryos, including 6 that exhibited ahigh degree of chimerism and wereobviously abnormal (data not shown).These embryos appeared to be delayedin development relative to their litter-mates, and four exhibited open neuralfolds (data not shown). No high per-centage Gcn5�/� chimeras were ob-served after E9.5–E10.5, indicatingthat these embryos die after this timein development. Lower percentageGcn5 null chimeras were found at alldevelopmental stages examined. Gcn5null cells were distributed evenly inearly embryos (E8.5–E9.5) but wereconcentrated in the heart and tail budof later stage embryos (E10.5–E12.5;Fig. 9B,D,D�). The concentration ofGcn5 null cells in the heart is consis-tent with our previous report thatGcn5 is not expressed in the embry-onic heart. The tail bud is enriched instem cells (Copp et al., 2003), consis-tent with our findings above that Gcn5is not required for survival of ES cells.Collectively, these findings indicatethat Gcn5 is not required for earlyspecification of most tissue lineagesbut may be required for subsequent
Fig. 3. Gcn5�/� embryonic stem (ES) cells do not exhibit increased sensitivity to DNA-damagingagents. Equal numbers of Gcn5�/� (solid line) and Gcn5�/� (dashed line) ES cells were exposed tothe indicated doses of DNA-damaging agents. Surviving ES cell colonies were counted after 7days.
Fig. 4. Gcn5�/� embryonic stem (ES) cells form embryoid bodies. A: Time course of Gcn5�/� orGcn5�/� embryoid body (EB) formation. ES cells were aggregated as hanging drops, and EBs wereexamined at the indicated time points (all at �2 magnification). Note that Gcn5�/� EBs are similarin size and shape to wild-type EBs at day 3 but are smaller and misshapen at later time points.B: Morphology of day 6, hematoxylin and eosin–stained Gcn5�/� or Gcn5�/� EBs. EBs formedfrom two independent Gcn5�/� ES cell clones are shown. In both cases, Gcn5�/� EBs are clearlysmaller than wild-type EBs and have an irregular shape. C: Immunoblots of extracts from Gcn5�/�
or Gcn5�/� ES cells or EBs to detect total levels of histone H3 (H3) or H3 isoforms phosphorylatedat serine 10 (H3 S10P). Numbers under the blot are the ratios of the H3 S10P signals to total H3.D: Flow cytometry of PI-stained cells from day 5 EBs reveals an increased proportion of Gcn5�/�
cells are in the G2/M phase of the cell cycle. As in B, two independent Gcn5�/� ES cell clones wereanalyzed.
POTENTIAL OF GCN5�/� ES CELLS 1551

cell survival or for further organogen-esis.
Increased Apoptosis in Gcn5Null Chimeras
One striking phenotype of Gcn5 nullembryos is an increase in apoptosisrelative to wild-type or heterozygouslittermates as early as E7.5 (Xu et al.,2000). Consistent with those findings,we found that apoptosis was elevatedthroughout high-percentage Gcn5null chimeras (Fig. 10, and data notshown), whereas little or no apoptosiswas observed in Gcn5�/� chimeras.The even distribution of Gcn5�/� cellsin early stage chimeras and the ab-
sence of these cells in most regions oflater stage, lower percentage Gcn5null chimeras is consistent with wide-spread death of Gcn5�/� cells. Thesedata suggest that the apoptotic pheno-type is cell-autonomous. To further ex-amine this question and to determinewhether increased apoptosis occursspecifically in anterior regions whereneural tube closure is defective, weperformed TUNEL assays on sectionsof lower and higher percentageGcn5�/� or Gcn5�/� chimeras (Fig.10). Few to no TUNEL-positive cellswere observed in sections of anyGcn5�/� chimeras, but apoptotic cellswere obvious in the anterior regions ofGcn5�/� chimeras. Moreover, the
TUNEL signal was limited toGcn5�/� cells within the chimeras (asmarked by lacZ), further demonstrat-ing that this increased cell death isdue to Gcn5 loss.
DISCUSSION
Our studies provide several new in-sights to the functions of Gcn5 in cellsurvival and embryo development.First, we find that Gcn5 is not re-quired for ES cell survival before orafter differentiation in vitro. However,it is required for efficient progressionthrough G2/M phase in differentiatingcells and may be required for mainte-nance of pluripotent states withinEBs, as evidenced by premature ces-sation of Nodal and Oct4 expression.Second, Gcn5 null ES cells efficientlyform skeletal and cardiac muscle invitro. These findings, together withour previous observation that PCAFnull mice exhibit no abnormalities inmuscle formation, indicate that nei-ther Gcn5 nor PCAF are required formuscle development. These findingsare in contrast to previous reports byothers that indicate that PCAF canstimulate conversion of fibroblasts toskeletal myoblasts (Puri et al., 1997;Sartorelli et al., 1999). In fact, ourdata indicate that Gcn5 may limitmuscle formation, at least in vitro.Third, despite the ability of Gcn5 nullcells to differentiate and to survive invitro, these cells undergo apoptosis invivo, in the context of an embryo, evenwhen surrounded by wild-type cells.These findings indicate that loss ofGcn5 induces a cell-autonomous pro-gram of cell death.
Gcn5 is part of multiple large,multisubunit acetyltransferase com-plexes. The Gcn5 deletion allele usedhere, and the complete loss of Gcn5expression, might disrupt multiplefunctions within these complexes.Thus, the phenotypes we observe heremay not arise solely from the loss ofGcn5 HAT activity. Other studies on-going in our laboratory will addressthis possibility by comparing the ef-fects of catalytic site mutations inGcn5 with the effects of Gcn5 deletion.
Our findings raise the question ofwhy apoptosis is observed in Gcn5null cells in embryos but not in Gcn5null ES cells in vitro. The onset of theincreased apoptotic phenotype in
Fig. 5. H3 acetylation at K9 and K18 is diminished in Gcn5�/� embryoid bodies (EBs). Immuno-blots of increasing amounts of total protein extracted from pools of wild-type (WT) or Gcn5�/� EBsat the indicated time points using either an anti-H3 Ac K9, 18 antisera or an antibody to theC-terminus of H3. The ratios of acetylated H3 to total H3 are shown under the blots.
Fig. 6. Expression of developmental marker genes in Gcn5�/� embryonic stem (ES) cells andembryoid bodies (EBs). Expression levels of the indicated genes were examined by reversetranscriptase-polymerase chain reaction in ES cells or at the indicated times of EB formation.GAPDH expression was used to control for differences in sample input.
1552 LIN ET AL.
Gcn5�/� embryos occurs during gas-trulation, when the embryonic cell cy-cle is significantly shortened to allowan explosion in cell proliferation. Ofinterest, other findings from our labindicate that p53 is induced in Gcn5null embryos and that cells from theseembryos contain abnormal chromo-some end-associations and telomericdefects (P. Bu and S. Y. R. D., unpub-
lished observations), consistent withthe cell-autonomous cell death ob-served here. One possibility, then, isthat defective telomeres in Gcn5 nullcells are normally repaired in ES cells,but that such repair is not efficient inrapidly dividing cells during gastrula-tion. Indeed, the slowing of mitoticprogression in differentiating Gcn5null cells in culture might reflect extra
time taken to resolve the abnormalstructures. Consistent with this idea,others have shown that embryos arehypersensitive to DNA damage duringgastrulation and that an ATM andp53-dependent surveillance mecha-nism is activated during this time toremove cells that carry even minimalamounts of DNA damage (Heyer et al.,2000). This surveillance mechanismdoes not operate in ES cells in culture(Heyer et al., 2000) and is presumablynot needed, because the slower prolif-eration of these cells allows normalcheckpoint and DNA repair pathwaysto function. Thus, abnormalities inGcn5 null cells may be resolved in EScells in culture but trigger a p53-de-pendent surveillance response in gas-trulating embryos.
EXPERIMENTALPROCEDURES
Generation of Gcn5�/� ESCells
Wild-type AB1 ES cells (McMahonand Bradley, 1990) were electropo-rated with a conditional-Gcn5 target-ing vector that deleted exons 3–19 ofthe Gcn5 coding region (Fig. 1). Fortransfections, ES cells were culturedon mitotically inactivated mouse em-bryonic fibroblast STO feeder cells incomplete medium (Dulbecco’s Modi-fied Eagle’s Medium [DMEM] with 4.5g/L glucose and L-glutamine, supple-mented with 2 mM L-glutamine, 100U/ml penicillin–streptomycin, 0.1 mMbeta-mercaptoethanol, and 15% fetalbovine serum [FBS; Hyclone]) in a 5%CO2 humidified incubator. The recom-bination of the conditional allele intothe endogenous locus was tested usingtwo selection drugs: FIAU and G418.Southern blotting with a 5� and 3�probe confirmed proper targeting ofthe Gcn5 locus with the Gcn5-condi-tional allele. Then, a correctly tar-geted conditional ES cell clone wasgrown in culture and electroporatedwith a Cre-recombinase expressionplasmid to induce recombination atthe lox-P sites of the integrated con-struct. After identifying the “floxed”-Gcn5 ES cell clone by Southern blot-ting, the second Gcn5 allele wastargeted with a vector that replacedthe entire coding region of Gcn5 with aPGK–Neomycin cassette and a beta-
Fig. 7. Skeletal muscle differentiation is enhanced in the absence of Gcn5. Wild-type or twoindependent clones of Gcn5�/� embryonic stem (ES) cells were aggregated into embryoid bodies(EBs)and allowed to differentiate for 33 days as described in the text. A: Anti–myosin heavy chainstaining to monitor myoblast and myofiber formation. B: Reverse transcriptase-polymerase chainreaction (RT-PCR) reveals robust expression of myogenic markers in day 34 Gcn5�/� EBs. L7 RNAexpression was used as an internal control in these experiments. C: RT-PCR also indicates thatcardiac muscle marker genes are well expressed in day 14 Gcn5�/� EBs.
Fig. 8. Insertion of ROSA26 lacZ transgene into embryonic stem (ES) cells. A: Strategy to inserta ubiquitously expressed lacZ transgene into the Rosa26 locus of Gcn5�/� or Gcn5�/� ES cells.B: Polymerase chain reaction (PCR) screening to confirm proper transgene targeting, using primersindicated in A. The arrow marks the size of the expected band; the lower band is a nonspecificproduct of the PCR reaction, which provides a convenient loading control. Lane 1, molecularmarkers. Lanes 2–10, independent puromycin resistant ES cell clones.
POTENTIAL OF GCN5�/� ES CELLS 1553

galactosidase gene (Xu et al., 2000).Southern blotting confirmed the dele-tion of the second Gcn5 allele, result-ing in a Gcn5�/� ES cell clone.
General ES Cell Growth andDifferentiation Conditions
Undifferentiated ES cells were grownin the absence of feeder layers in com-plete medium as above plus 1,000U/ml of ESGRO (leukemia inhibitoryfactor; Sigma), and 15% FBS (Hy-clone). For differentiation, 30-�l dropscontaining 600 cells/drop in completemedium without ESGRO and 15%FBS from Sigma (F-2442) were platedas hanging drops on the inside of thelids of 10-cm bacterial Petri culturedishes. For skeletal muscle differenti-ation experiments, day 5 (D5) EBswere plated on 0.1% gelatin-coated,2-chamber well glass slides (LAB-TEK) for immunohistochemistry and24-well plates for RNA analysis andincubated an additional 26–28 days.
Analysis of ES Cell Growth
For growth curve analyses, ES cellswere seeded at 1 � 105 cells in 60-mmculture dishes. Duplicate plates ofcells were trypsinized and washed inPBS, and an equal volume of 0.4%trypan blue solution was added to thecells and incubated for 5 min. Cellswere counted daily by hemocytometer.Trypan blue-positive cells were con-sidered dead and trypan blue-negative cells alive. Data shown areaverages from two independent exper-iments with duplicate plates countedat each time point.
Immunoblots
ES cells or EBs were lysed by directlyadding TRIZOL reagent (Invitrogen; 6ml for one 10-cm culture dish or 2 mlfor 100–200 EBs). Cell lysates werepipetted several times and then incu-bated for 5 min at room temperature.Chloroform (0.2 ml per ml of TRIZOLreagent) was added, and the tubeswere shaken vigorously. After centrif-ugation, proteins were precipitatedfrom the supernatant with isopropylalcohol. Proteins were washed threetimes in 0.3 M guanidine hydrochlo-ride in 95% ethanol and dissolved in1% sodium dodecyl sulfate (SDS) solu-
tion. Proteins were separated by elec-trophoresis in 15% SDS-polyacryl-amide gels and then transferred ontoa polyvinylidene difluoride membraneusing a semi-dry transfer apparatus(Bio-Rad). The membrane was blockedwith 5% nonfat dry milk in 1� TBSTfor 1 hr and incubated with primaryantibody (anti–phospho-Histone H3[Ser10], [Cell signaling] at 1:1,000 di-lution; anti H3 Ac 9,18 [Edmondson etal., 1996] 1:1,000 dilution, or anti Hi-stone H3 C-terminus [Abcam] at1:2,000 dilution). The secondary anti-body used in these experiments wasdonkey anti-rabbit horseradish perox-idase (GE Healthcare; 1:2,000 dilution[for anti phospho-Histone H3 and an-ti-H3 Ac 9,18] or at 1:10,000 dilutionfor histone H3). Signals were detectedusing the Amersham ECL plus West-ern blotting detection system signalchemiluminescent solutions and expo-sure to X-ray film.
Expression Analyses ofDevelopmental MarkerGenes
Approximately 300 EBs were pooled,rinsed with PBS, and then homoge-nized using TRIzol reagent (Invitro-gen). RNA was isolated according tothe manufacturer’s protocol. TotalRNA (4 �g) isolated from EBs at var-ious time points was treated with 2 Uof Turbo DNA-free DNase I (Ambion)in a total volume of 10 �l for 1 hr at
37°C. DNase-treated RNA (600 ng)was used for reverse transcription us-ing the GeneAmp Gold RNA PCR CoreKit (PE Biosystems). The RT reactionscontained oligo d(T) as primer ratherthan specific sequence primers or ran-dom hexamers. The manufacturer’sprotocol was followed for the two-stepRT-PCR protocol, except that the vol-ume of all the reagents was halved.
Gene Expression Analysis ofMuscle-Specific Markers
Day 31–33 EBs grown on 24-wellplates were rinsed with phosphatebuffered saline (PBS) and then ho-mogenized with TRIzol reagent (In-vitrogen). RNA was isolated accordingto the manufacturer’s protocol. Thentotal RNA (2 �g) was treated withTurbo DNase (Ambion) as above. RNA(600 ng) was reverse transcribed us-ing the GeneAmp Gold RNA PCR CoreKit (PE Biosystems). Primers for anal-ysis of expression of the followinggenes were a kind gift from Dr. AnitaMeyer and Dr. William Klein: L7,Mck, Mrf4, Myf5, MyoD, and Myoge-nin.
Immunostaining
Approximately 92 EB hanging dropswere collected at day 5 of EB cultiva-tion and transferred to eight 0.1% gel-atin coated two-chamber glass slides.Briefly, six EBs in differentiation me-dium were plated per chamber and
TABLE 1. Generation of Chimeric Embryos Bearing Gcn5�/� and Gcn5�/�
Cells
Embryonic stemcell genotype Blastocystsa Embryosb
Chimeric embryosc
High(abnormal)
Low(abnormal)
Gcn5�/� 115 51 8 (0) 21 (0)Gcn5-/- 184 83 6 (6) 32 (0)
aData shown are compiled from results of blastocyst injections using threeindependent Gcn5�/� or Gcn5�/� embryonic stem cell clones.bNumbers indicate the number of embryos collected after injection of the indicatednumber of blastocysts.cNumbers represent the total number of embryonic day (E) 8.5–E12.5 chimericembryos obtained from all injections as scored by the presence of LacZ-positive cells.High refers to embryos with 50% or more LacZ-positive cells. Low refers to embryoswith less than 50% LacZ-positive cells. Numbers in parentheses indicate numbers ofembryos with abnormal phenotypes. All six high-percentage chimeras derived fromGcn5�/� embryonic stem cell clones exhibited abnormal morphologies. These embryoswere delayed in development, and four had open neural folds.
1554 LIN ET AL.
cultivated for an additional 26–28days at 5% CO2 at 37°C. Differentia-tion medium was replaced severaltimes during the cultivation. After theadditional 26–28 days in culture, cellswere fixed in acetone at 4°C for 10 minand then immunostained with theHistoMouse-SP kit (Zymed) and anti-myosin heavy chain MF20 ascitesfrom the Developmental Studies Hy-bridoma Bank (Johns Hopkins Uni-versity and the University of Iowa) at1:500 dilution.
Annexin V–FluoresceinIsothiocyanate Staining andFlow Cytometry
Approximately 100 D5 EB hangingdrops were pooled, rinsed with PBS,and trypsinized with Trypsin (Invitro-gen) for 5 min at 37°C. EBs were fur-ther disrupted by gentle pipetting sothat a single suspension of cells wasobtained. The 1 � 105 cells were incu-bated with Annexin V–fluorescein iso-thiocyanate (FITC) and PI, followedby flow cytometry analysis (with thehelp of the UTMDACC Core Facility).The Annexin V-FITC Apoptosis Detec-tion Kit I (BD Pharmingen) was usedfor staining, which was performed ac-cording to the manufacturer’s proto-col. For determining the cell cycle pro-file by FACs, the same protocol asmentioned above was followed, exceptthat cells were stained with pro-pidium PI only.
Introduction of a lacZ alleleinto the ROSA26 locus in EScells
A lacZ transgene was introduced intoGcn5�/� and Gcn5�/� ES cells by ho-mologous recombination so that �-ga-lactosidase could be used to trace thefate of these cells in developing em-bryos. A ROSA26 lacZ targeting vec-tor carrying a puromycin positive se-lection marker was created by cloninga puromycin cassette fused to an in-ternal ribosomal entry site lacZ alleledownstream of a splice acceptor site(Fig. 7A). The fused cassette was theninserted into a 5Kb ROSA26 genomicsequence cloned into a vector carryingthe diphtheria toxin gene as a nega-tive selection marker. The linear-ized vector was electroporated into
Fig. 9. Abnormal development of chimeric embryos carrying a high percentage of Gcn5�/� cells.A,C: Chimeras with a high contribution of Gcn5�/� cells at embryonic day (E)10.5, and E12.5 (asindicated). B,D,D�: Chimeras bearing Gcn5�/� embryonic stem (ES) cells. High-percentageGcn5�/� chimeras were only observed at early time points (�E9.5) and in all cases were abnormal(data not shown). By E12.5, only a few Gcn5�/� cells were detected in chimeras, concentrated inthe heart (D) and the tail bud (D�).
Fig. 10. Elevated apoptosis in Gcn5�/� cells within chimeric embryos. A,B: Frontal anteriorsections of E10.5 chimeric embryos containing Gcn5�/� (A) or Gcn5�/� cells (B), as indicated, weresubjected to terminal deoxynucleotidyl transferase–mediated deoxyuridinetriphosphate nick end-labeling (TUNEL) analyses. Insets show higher magnification views of boxed regions. LacZ-positivecells are blue, and TUNEL-positive cells are dark brown. Cells were counterstained with nuclear fastred. Few TUNEL-positive cells are evident in sections from Gcn5�/� chimeras, but many apoptoticcells are present in sections from Gcn5�/� chimeras, and TUNEL staining is limited to thelacZ-positive, Gcn5�/� cells.
POTENTIAL OF GCN5�/� ES CELLS 1555

Gcn5�/� and Gcn5�/� ES cells, andROSA26-lacZ–bearing colonies wereidentified by PCR after positive andnegative drug selections. Primer se-quences and PCR conditions will beprovided upon request.
Production and Analysis ofChimeric Embryos
Gcn5�/� or Gcn5�/� ES cells were in-jected into blastocysts from C57BL/6Jmice, which were then transferred intopseudopregnant Swiss foster mothers.Embryos were then isolated at specifictime points as indicated in the text. Em-bryos were immersed in fixative solu-tion (0.8% paraformaldehyde, 02% glu-taraldehyde, 0.1 M NaPO4 [pH7.5], 5mM EGTA [pH 8.0], 2 mM MgCl2) for45 min and then washed two times inrinse buffer (0.1 M NaPO4 (pH 7.5), 2mM MgCl2, 0.1% Na deoxycholate,0.2% NP40). Embryos were thenstained in 5-bromo-4-chloro-3-indolyl-�-D-galactopyranoside (X-gal) buffer (5mM potassium ferrocyanide, 5 mM po-tassium ferriyanide, 1 mg/ml of X-gal,0.1 M NaPO4 [pH 7.5], 2 mM MgCl2,0.1% Na deoxycholate, 0.2% NP40)overnight at 37°C. Stained sampleswere rinsed twice with cold PBS for30min.
TUNEL Assays
Embryos analyzed by X-gal stainingwere fixed in 4% paraformaldehyde in1� PBS at 4°C overnight and werethen embedded in paraffin and sec-tioned. Apoptotic cells were detectedusing a TUNEL assay kit (CALBIO-CHEM TdT-FragEL DNA fragmenta-tion Detection Kit) according to themanufacturer’s instructions. Sectionswere counterstained by nuclear FastRed after the TUNEL assay.
ACKNOWLEDGMENTSWe thank the Genetically EngineeredMouse Facility (supported by NCICA16672) for help in generating theGcn5�/� and Gcn5�/� ES cells.S.Y.R.D. was funded by a grant fromthe NIH (GM067718).
REFERENCES
Annunziato AT, Hansen JC. 2000. Role ofhistone acetylation in the assembly and
modulation of chromatin structures.Gene Expr 9:37–61.
Bernstein BE, Mikkelsen TS, Xie X, KamalM, Huebert DJ, Cuff J, Fry B, MeissnerA, Wernig M, Plath K, Jaenisch R, Wag-schal A, Feil R, Schreiber SL, Lander ES.2006. A bivalent chromatin structuremarks key developmental genes in em-bryonic stem cells. Cell 125:315–326.
Boiani M, Scholer HR. 2005. Regulatorynetworks in embryo-derived pluripotentstem cells. Nat Rev Mol Cell Biol 6:872–884.
Choi D, Lee HJ, Jee S, Jin S, Koo SK, PaikSS, Jung SC, Hwang SY, Lee KS, Oh B.2005. In vitro differentiation of mouseembryonic stem cells: enrichment ofendodermal cells in the embryoid body.Stem Cells 23:817–827.
Copp AJ, Greene ND, Murdoch JN. 2003.The genetic basis of mammalian neuru-lation. Nat Rev Genet 4:784–793.
Edmondson DG, Smith MM, Roth SY.1996. Repression domain of the yeastglobal repressor Tup1 interacts directlywith histones H3 and H4. Genes Dev10:1247–1259.
Elefanty AG, Robb L, Birner R, Begley CG.1997. Hematopoietic-specific genes arenot induced during in vitro differentia-tion of scl-null embryonic stem cells.Blood 90:1435–1447.
Gambaro K, Aberdam E, Virolle T, Aber-dam D, Rouleau M. 2006. BMP-4 inducesa Smad-dependent apoptotic cell death ofmouse embryonic stem cell-derived neu-ral precursors. Cell Death Differ 13:1075–1087.
Grant PA, Duggan L, Cote J, Roberts SM,Brownell JE, Candau R, Ohba R, Owen-Hughes T, Allis CD, Winston F, BergerSL, Workman JL. 1997. Yeast Gcn5functions in two multisubunit complexesto acetylate nucleosomal histones: char-acterization of an Ada complex and theSAGA (Spt/Ada) complex. Genes Dev 11:1640–1650.
Grant PA, Eberharter A, John S, Cook RG,Turner BM, Workman JL. 1999. Ex-panded lysine acetylation specificity ofGcn5 in native complexes. J Biol Chem274:5895–5900.
Heyer BS, MacAuley A, Behrendtsen O,Werb Z. 2000. Hypersensitivity to DNAdamage leads to increased apoptosis dur-ing early mouse development. Genes Dev14:2072–2084.
Jenuwein T, Allis CD. 2001. Translatingthe histone code. Science 293:1074–1080.
Kim J, Daniel J, Espejo A, Lake A, KrishnaM, Xia L, Zhang Y, Bedford M. 2006.Tudor, MBT, and chromodomains gaugethe degree of lysine methylation. EMBORep 4:397–403.
Kubo A, Shinozaki K, Shannon JM, Kous-koff V, Kennedy M, Woo S, Fehling HJ,Keller G. 2004. Development of defini-tive endoderm from embryonic stem cellsin culture. Development 131:1651–1662.
Kung AL, Rebel VI, Bronson RT, Ch’ng LE,Sieff CA, Livingston DM, Yao TP. 2000.Gene dose-dependent control of hemato-
poiesis and hematologic tumor suppres-sion by CBP. Genes Dev 14:272–277.
Lee TI, Causton HC, Holstege FC, ShenWC, Hannett N, Jennings EG, WinstonF, Green MR, Young RA. 2000. Redun-dant roles for the TFIID and SAGA com-plexesinglobaltranscription.Nature405:701–704.
Li E. 2002. Chromatin modification andepigenetic reprogramming in mamma-lian development. Nat Rev Genet 3:662–673.
McKinsey TA, Zhang CL, Olson EN. 2001.Control of muscle development by duel-ing HATs and HDACs. Curr Opin GenetDev 11:497–504.
McMahon AP, Bradley A. 1990. The Wnt-1(int-1) proto-oncogene is required for de-velopment of a large region of the mousebrain. Cell 62:1073–1085.
Naito AT, Shiojima I, Akazawa H, HidakaK, Morisaki T, Kikuchi A, Komuro I.2006. Developmental stage-specific bi-phasic roles of Wnt/beta-catenin signal-ing in cardiomyogenesis and hematopoi-esis. Proc Natl Acad Sci U S A 103:19812–19817.
Phan HM, Xu AW, Coco C, Srajer G,Wyszomierski S, Evrard YA, Eckner R,Dent SY. 2005. GCN5 and p300 shareessential functions during early embryo-genesis. Dev Dyn 233:1337–1347.
Puri PL, Sartorelli V, Yang XJ, HamamoriY, Ogryzko VV, Howard BH, Kedes L,Wang JY, Graessmann A, Nakatani Y,Levrero M. 1997. Differential roles ofp300 and PCAF acetyltransferases inmuscle differentiation. Mol Cell 1:35–45.
Roche E, Sepulcre P, Reig JA, Santana A,Soria B. 2005. Ectodermal commitmentof insulin-producing cells derived frommouse embryonic stem cells. FASEB J19:1341–1343.
Roth SY, Denu JM, Allis CD. 2001. HistoneAcetyltransferases. Annu Rev Biochem70:81–120.
Sartorelli V, Puri PL, Hamamori Y,Ogryzko V, Chung G, Nakatani Y, WangJY, Kedes L. 1999. Acetylation of MyoDdirected by PCAF is necessary for theexecution of the muscle program. MolCell 4:725–734.
Song K, Backs J, McAnally J, Qi X, GerardRD, Richardson JA, Hill JA, Bassel-Duby R, Olson EN. 2006. The transcrip-tional coactivator CAMTA2 stimulatescardiac growth by opposing class II his-tone deacetylases. Cell 125:453–466.
Tada S, Era T, Furusawa C, Sakurai H,Nishikawa S, Kinoshita M, Nakao K,Chiba T, Nishikawa S. 2005. Character-ization of mesendoderm: a divergingpoint of the definitive endoderm and me-soderm in embryonic stem cell differen-tiation culture. Development 132:4363–4374.
Ura K, Kurumizaka H, Dimitrov S, Al-mouzni G, Wolffe AP. 1997. Histone acet-ylation: influence on transcription, nu-cleosome mobility and positioning, andlinker histone-dependent transcriptionalrepression. EMBO J 16:2096–2107.
Vega RB, Matsuda K, Oh J, Barbosa AC,Yang X, Meadows E, McAnally J, Poma-
1556 LIN ET AL.

jzl C, Shelton JM, Richardson JA,Karsenty G, Olson EN. 2004. Histonedeacetylase 4 controls chondrocyte hy-pertrophyduringskeletogenesis.Cell119:555–566.
Winston F, Allis CD. 1999. The bromodo-main: a chromatin-targeting module?[news]. Nat Struct Biol 6:601–604.
Xu W, Edmondson DG, Evrard Y, Waka-miya M, Behringer RR, Roth SY. 2000.Loss of GCN5 leads to increased apopto-sis and mesodermal defects duringmouse development. Nat Genet 26:229–232.
Xu W, Edmondson DG, Roth SY. 1998.Mammalian GCN5 and P/CAF acetyl-transferases share homologous amino-terminal domains important for the rec-
ognition of nucleosomal substrates. MolCell Biol 18:5659–5669.
Yamauchi T, Yamauchi J, Kuwata T,Tamura T, Yamashita T, Bae N, West-phal H, Ozato K, Nakatani Y. 2000. Dis-tinct but overlapping roles of the histoneacetylase PCAF and of the closely re-lated PCAF-B/GCN5 in mouse embryo-genesis. Proc Natl Acad Sci U S A 97:11303–11306.
Yang X-J, Ogryzko VV, Nishikawa J-I,Howard BH, Nakatani Y. 1996. A p300/CBP associated factor that competeswith the adenoviral oncoprotein E1A.Nature 382:319–324.
Yao TP, Oh SP, Fuchs M, Zhou ND, Ch’ngLE, Newsome D, Bronson RT, Li E, Liv-ingston DM, Eckner R. 1998. Gene dos-age-dependent embryonic development
and proliferation defects in mice lackingthe transcriptional integrator p300. Cell93:361–372.
Zhang Y, Reinberg D. 2001. Transcriptionregulation by histone methylation: inter-play between different covalent modifi-cations of the core histone tails. GenesDev 15:2343–2360.
Zhang W, Bone JR, Edmondson DG,Turner BM, Roth SY. 1998. Essentialand redundant functions of histone acet-ylation revealed by mutation of targetlysines and loss of the Gcn5p acetyl-transferase. EMBO J 17:3155–3167.
Zhang C, McKinsey T, Chang S, Antos C,Hill J, Olson E. 2002. Class II histonedeacetylases act as signal-responsive re-pressors of cardiac hypertrophy. Cell110:479.
POTENTIAL OF GCN5�/� ES CELLS 1557





























