Embed Size (px)
Citation preview

Direct evidence of abca1-mediated efflux of cholesterolat the mouse blood–brain barrier
Tuan Minh Do • Melissa Ouellet • Frederic Calon •
Giovanna Chimini • Helene Chacun •
Robert Farinotti • Fanchon Bourasset
Received: 9 March 2011 / Accepted: 28 May 2011 / Published online: 10 June 2011
� Springer Science+Business Media, LLC. 2011
Abstract We investigated the expression and function
of Abca1 in wild-type C57BL/6, abca1(?/?), and
abca1(-/-) mice brain capillaries forming the blood–brain
barrier (BBB). We first demonstrated by quantitative RT-
PCR and Western immunoblot that Abca1 was expressed
and enriched in the wild-type mouse brain capillaries. In
abca1(-/-) mice, we reported that the lack of Abca1
resulted in an 1.6-fold increase of the Abcg4 expression
level compared to abca1(?/?) mice. Next, using the in situ
brain perfusion technique, we showed that the [3H]cho-
lesterol brain uptake clearance (Clup, ll/s/g brain), was
significantly increased (107%) in abca1(-/-) mice com-
pared to abca1(?/?) mice, meaning that the deficiency of
Abca1 conducted to a significant decrease of the choles-
terol efflux at the BBB level. In addition, the co-perfusion
of probucol (Abca1 inhibitor) with [3H]cholesterol resulted
in an increase of [3H]cholesterol Clup (115%) in abca1
(?/?) but not in abca1(-/-) mice, meaning that probucol
inhibited selectively the efflux function of Abca1. In
conclusion, our results demonstrated that Abca1 was
expressed in the mouse brain capillaries and that Abca1
functions as an efflux transporter through the mouse BBB.
Keywords Blood–brain barrier � Cholesterol � Abca1 �Abcg4 � Abcb1 � Abcg2 � Mouse � In situ brain perfusion
Abbreviations
ABC ATP-binding cassette
AD Alzheimer’s disease
BBB Blood–brain barrier
BCECs Brain capillary endothelial cells
Clup Brain uptake clearance
PBS Phosphate-buffered saline
Introduction
ABCA1, which belongs to the superfamily of the ATP-
binding cassette (ABC) proteins, is known for its efflux
function of cholesterol from intracellular compartment to
systemic and brain apolipoproteins [1]. ABCA1 is
expressed in several tissues, including brain, placenta, and
liver [2]. In the brain, ABCA1 is particularly expressed in
neurons and glial cells [3, 4]. ABCA1 mRNA has also been
detected in cultured human and rat brain capillaries endo-
thelial cells (BCECs) [5]. In addition, the Abca1 protein
has been identified in the brain capillaries of transgenic
mice lacking Abca1 selectively in neurons and glial cells
[4]. Associated with the basement membrane, pericytes and
astrocytes end-foot, the brain capillary endothelial cells
(BCECs) constitute the blood–brain barrier (BBB), which
represents the main interface separating the blood circu-
lation from the brain. The barrier effect is due to tight
T. M. Do � R. Farinotti � F. Bourasset (&)
Laboratory of Clinical Pharmacy, EA4123, Faculty of Pharmacy,
University of Paris Sud 11, 5 rue Jean-Baptiste Clement,
92296 Chatenay-Malabry, France
e-mail: [email protected]
M. Ouellet � F. Calon
Faculty of Pharmacy, Laval University, Quebec, QC, Canada
G. Chimini
Centre d’Immunologie de Marseille-Luminy, INSERM-CNRS-
Universite de La Mediterranee, Marseille, France
H. Chacun
Laboratory of Biopharmacy and Pharmaceutical Technology,
CNRS UMR 8612, Faculty of Pharmacy, University of Paris
Sud 11, Chatenay-Malabry, France
123
Mol Cell Biochem (2011) 357:397–404
DOI 10.1007/s11010-011-0910-6

junctions between BCECs as well as to the presence of
efflux transporters that limit the entry of their substrates
into the brain. Among these efflux transporters, ABC
proteins such as ABCB1 (P-glycoprotein, P-gp) and
ABCG2 (Breast Cancer Resistance Protein, BCRP) have
been shown to play prominent roles in brain homeostasis.
In addition to limiting the brain entrance of several drugs,
these proteins are also involved in a wide range of
pathologies, such as inflammatory or neurodegenerative
diseases [6–9].
Like other ABC proteins, ABCA1 has been suggested to
play a key role in Alzheimer’s disease (AD) [10, 11]. The
direct link between ABCA1 and AD has not been evi-
denced, but several studies showed relationships between
the brain level of ABCA1 and amyloid-beta peptide (Ab)
(see review [11]). Recently, Kim et al. [12] showed that
Abca1 was over-expressed in hippocampal neurons of
APP/PS1 mice. In addition, the link between cholesterol
metabolism and AD has been established by series of in
vivo and in vitro studies [13, 14].
So far, the exact mechanisms by which brain cholesterol
homeostasis, ABCA1 and AD are linked are unknown. A
hypothesis is that cholesterol could interfere with the BBB
efflux of Ab, by an ABCA1-mediated pathway. However,
the efflux function of ABCA1 in the BBB has not been
evidenced. Since the most commonly used animal models
of AD are developed in the mouse, it appeared necessary to
quantify the expression and efflux function of Abca1 in the
mouse BBB.
For this purpose, we performed quantitative RT-PCR
(qRT-PCR) and Western immunoblot to measure the
mRNA and protein expression of Abca1 in the C57BL/6
mouse BBB. We then performed in situ brain perfusion to
evaluate the Abca1 function at the mouse BBB by mea-
suring the brain uptake of [3H]cholesterol (used as a probe)
in the presence or absence of probucol, a known Abca1
inhibitor [15], in abca1(?/?), abca1(±), or abca1(-/-)
mice.
Materials and methods
Animals
C57BL/6 mice (6–8 weeks old) were purchased from
Janvier (Le Genest-St-Ile, France). The heterozygote
Abca1-deficient mice are on a C57BL/6 genetic back-
ground. These mice were bred to yield abca1(-/-),
abca1(?/-), and abca1(?/?) littermate mice [16]. The
genotype of offspring was checked by PCR analysis using
genomic DNA isolated from tail biopsies, with the fol-
lowing pairs of primers: forward, 50 TGG GAA CTC CTG
CTA AAA T 30, and reverse, 50 CCA TGT GGT GTG TAG
ACA 30 for the wild-type allele; forward, 50 TTT CTC
ATA GGG TTG GTC A 30, and reverse, 50 TGC AAT
CCA TCT TGT TCA AT 30 for the mutant allele. The wild-
type allele generated a 750 bp product and the mutant
allele a 500 bp product. Male and female mice, 6- to
8-week-old, had free access to standard laboratory food and
water and kept on a 12 h light–dark cycle at 22 ± 1�C.
They were housed under these conditions for at least
5 days before being used. Studies involving animals and
their care were performed according to the guidelines
issued by the European Economic Community for the care
and use of laboratory animals (Official Journal of the
European Community, 18/2/86-authorization L3600).
Reagents and antibodies
[3H]cholesterol (44.5 Ci/mmol), [14C]sucrose (588.0 mCi/
mmol), Soluene, and Ultima Gold scintillation cocktails
were purchased from PerkinElmer Life and Analytical
Sciences (Courtaboeuf, France). Probucol was obtained
from Sigma-Aldrich (St. Quentin Fallavier, France). Rat
monoclonal anti-mouse Abca1 antibody (clone NB400-
164) was purchased from Novus Biological (Littleton,
USA). Rat monoclonal anti-mouse Abcg2 antibody (clone
BXP-53) and mouse monoclonal anti-mouse Abcb1 anti-
body (clone C219) were obtained from Dako (Glostrup,
Denmark). Rabbit polyclonal anti-mouse Abcg4 antibody
was purchased from Alpha Diagnostic International (San
Antonio, TX, USA). Mouse monoclonal anti-mouse b-actin
antibody (clone AC74) was from Sigma-Aldrich (St Louis,
MO, USA). All the other chemicals were commercial
products of reagent grade.
Transporter expression studies
mRNA extraction and quantitative RT-PCR
The mRNA was extracted from cortex homogenate or from
mouse brain capillaries. The capillary depletion method of
Triguero et al. [17] was used with some modifications [18]
to obtain capillary-enriched fractions of mouse brains. To
remove the blood from brain capillaries, seven mice were
perfused with saline by in situ brain perfusion for 30 s.
After decapitation, the brain was removed and meninges
were discarded on ice. Brains were homogenized in 7 ml of
buffer (10 mM HEPES, 141 mM NaCl, 4 mM KCl, 1 mM
NaH2PO4, 2.8 mM CaCl2, 1 mM MgSO4, and 10 mM
D-Glucose (pH 7.4) at 4�C). Chilled 37% dextran solution
was then added to obtain a final dextran concentration of
18.5%. The homogenates were centrifuged at 54009g for
15 min at 4�C in a swinging-bucket rotor. In these condi-
tions, the supernatant corresponds to the brain parenchyma,
and the pellet constitutes the enriched-capillary fraction
398 Mol Cell Biochem (2011) 357:397–404
123

[17]. All these fractions were carefully separated. The
purity of the fractions has been controlled by measuring the
gamma-glutamyl transferase activity which is maximal in
the capillary fraction [19]. Total RNA was extracted from
the isolated mouse brain capillaries and brain cortex
homogenate by using the RNeasy Mini (Qiagen) extraction
kit, according to the manufacturer’s instructions. Random
Hexamers (pDN6) were used to synthesize cDNA from
total RNA (1 lg) using a SuperScript II cDNA synthesis
kit (Invitrogen, Carlsbarg, CA) according to the manufac-
turer’s instructions. The primers for the target gene abca1
were purchased from Applied Biosystems (Mm0044
2646_m1 and Mm00440736_m1, respectively). Mouse b-
actin (Applied Biosystems) was used as normalization
control as it was shown that b-actin was stably expressed in
the brain [20, 21]. The conditions were 95�C for 10 min,
followed by 42 cycles of 15 s at 95�C, and 1 min at 60�C,
as recommended by the primer’s provider. The method
used for the mRNA quantification was the comparative CT
method with the formula 2-DDCt, as described previously
[22, 23].
Protein extraction and Western immunoblot analysis
Total proteins were extracted from C57BL/6, abca1(?/?),
abca1(?/-), and abca1(-/-) mice brain capillaries and
brain cortex. To extract total protein from the capillary
fraction, mice brain microvessels were first isolated using a
density-gradient procedure [24]. The mouse brain cortex
and the pellets containing microvessels were homogenized
in a mixture of TENTS (containing 10 mM Tris–HCl at pH
7.4, 5 mM EDTA at pH 8, 126 mM NaCl, 1% (v/v) Triton
X-100, and 0.1% (v/v) SDS) supplemented with leupeptin,
aprotinin, pepstatin, and phenyl methane sulfonyl fluoride
(Sigma-Aldrich, France). The suspensions were agitated
gently during 1 h at 4�C and then centrifuged at
120009g at 4�C for 20 min. Protein content in the super-
natant was determined by using the bicinchoninic acid
protein assay kit (Sigma-Aldrich, France).
Samples containing 10–25 lg of total proteins were
subjected to electrophoresis on an 8% SDS-polyacrylamide
gel and transferred electrophoretically on nitrocellulose
membranes (Amersham Biosciences, UK). The blots were
blocked with nonfat dry milk/Tween Tris base solution
(TTBS) 10% for 1 h at 20-25�C. After washing with TTBS,
the blots were incubated for 2 h at 20–25�C with primary
antibody. The dilutions of primary antibodies used were:
1:1500 for Abca1, 1:100 for Abcb1, 1:80 for Abcg2,
1:1000 for Abcg4 and 1:5000 for b-actin. After five washes
in TTBS (10 min each), they were further incubated for 1 h
at 20–25�C with horseradish peroxidase-conjugated anti-
bodies diluted at 1:10000 (Dako, Glostrup, Danemark).
The membranes were washed five times for 10 min each in
TTBS and then probed with the Western lightning chemi-
luminescence reagent (PerkinElmer, Courtaboeuf, France).
The intensity of the bands was quantified by using Scion
Image (NIH, Scion Corporation, Bethesda, MD, USA).
In vivo transport studies
Surgical procedure and perfusion technique
The mouse brain transport of [3H]cholesterol was measured
using the in situ brain perfusion method [25]. Mice were
anesthetized by intra-peritoneal injection of a mixture of
xylazine (Merial, Lyon, France) and ketamine (Bayer,
Puteaux, France) at 8/140 mg/kg. Briefly, the right com-
mon carotid artery was catheterized with polyethylene
tubing (0.30 mm 9 0.70 mm; Folioplast, Sarcelles, France)
filled with heparin (25 IU/ml). Before inserting the cathe-
ter, the common carotid artery was ligated on the heart side
and the right external carotid artery was ligated rostral to
the occipital artery at the level of the bifurcation of the
common carotid. Before perfusion, the heart was cut and
the perfusion was started immediately at a flow rate of
2.5 ml/min. The perfusion fluid consisted of bicarbonate-
buffered physiological saline containing 128 mM NaCl,
24 mM NaHCO3, 4.2 mM KCl, 2.4 mM NaH2PO4,
1.5 mM CaCl2, 0.9 mM MgCl2, and 9 mM D-glucose. The
solution was gassed with 95% O2–5% CO2 for pH control
(7.4) and warmed to 37�C. The syringe containing the
perfusate was placed in an infusion pump (Harvard
Apparatus, Les Ulis, France) and connected to the catheter.
For the time course study, each mouse was perfused
with [3H]cholesterol (0.2 lCi/ml) and [14C]sucrose
(0.3 lCi/ml) for 15, 30, or 60 s. For all other single-time
point studies, each mouse was perfused for 60 s with
[3H]cholesterol and [14C]sucrose to check the physical
integrity of the blood–brain barrier. The brain uptake of
[3H]cholesterol was also measured in the presence of
unlabeled inhibitor, probucol (10 lM). The stock solution
of probucol was prepared in dimethyl sulfoxide (DMSO;
\0.25% in final perfusate) and then added to the perfusate.
Perfusion was terminated by decapitating the animal. The
right cerebral hemisphere and aliquots of the perfusate
were weighed. Samples were digested in 1 ml of Soluene at
50�C and mixed with 9 ml of Ultima Gold scintillation
cocktail. Both labels were counted simultaneously in a
scintillation counter (LS6000LL, Beckman, Galway,
Ireland).
Calculation of BBB transport parameters [25, 26]
The brain vascular volume of each animal (Vvasc, ll/g
brain) was estimated from the tissue distribution of
Mol Cell Biochem (2011) 357:397–404 399
123

[14C]sucrose which does not measurably cross the BBB
[27], using the following equation:
Vvasc ¼X�
C�perf
where X* (dpm/g) is the amount of [14C]sucrose in the right
brain hemisphere and C�perf (dpm/ll) is the concentration of
[14C]sucrose in the perfusate.
The apparent tissue distribution volume (Vbrain, ll/g
brain) was calculated from the amount of radioactivity in
the right brain hemisphere using the following equation:
Vbrain ¼Xbrain
Cperf
where Cperf (dpm/ll) is the concentration of [3H]cholesterol
in the perfusate, and Xbrain (dpm/g) is the amount of
[3H]cholesterol in the right cerebral parenchyma, corrected
for vascular contamination:
Xbrain ¼ Xtot � VvascCperf
where Xtot (dpm/g) is the total amount of [3H]cholesterol
measured in the brain tissue sample (vascular and
extravascular).
The brain uptake clearance, expressed as Clup (ll/s/g
brain), was calculated from:
Clup ¼Vbrain
Tperf
where Tperf is the perfusion time (s).
Efflux ratios were calculated as:
Clup(-/-)/Clup
(?/?) = Clup obtained in abca1(-/-) mice/Clup
obtained in abca1(?/?) mice,
Clup(prob?)/Clup
(prob-) = Clup obtained with probucol/Clup
obtained without probucol, in Abca1-competent or -defi-
cient mice.
Data and statistical analysis
Data were shown as means ± S.D. Student’s unpaired
t test was used to identify significant differences between
groups when appropriate. All the tests were two-tailed and
statistical significance was set at * P \ 0.05, ** P \ 0.01,
*** P \ 0.001.
Results
Mouse BBB expression of Abca1
Abca1 mRNA was detected in the brain cortex homogenate
as well as in the brain capillary fraction of C57BL/6 mice.
The mRNA level of abca1 was significantly 4.2-fold
enriched in the brain capillary fraction compared to the
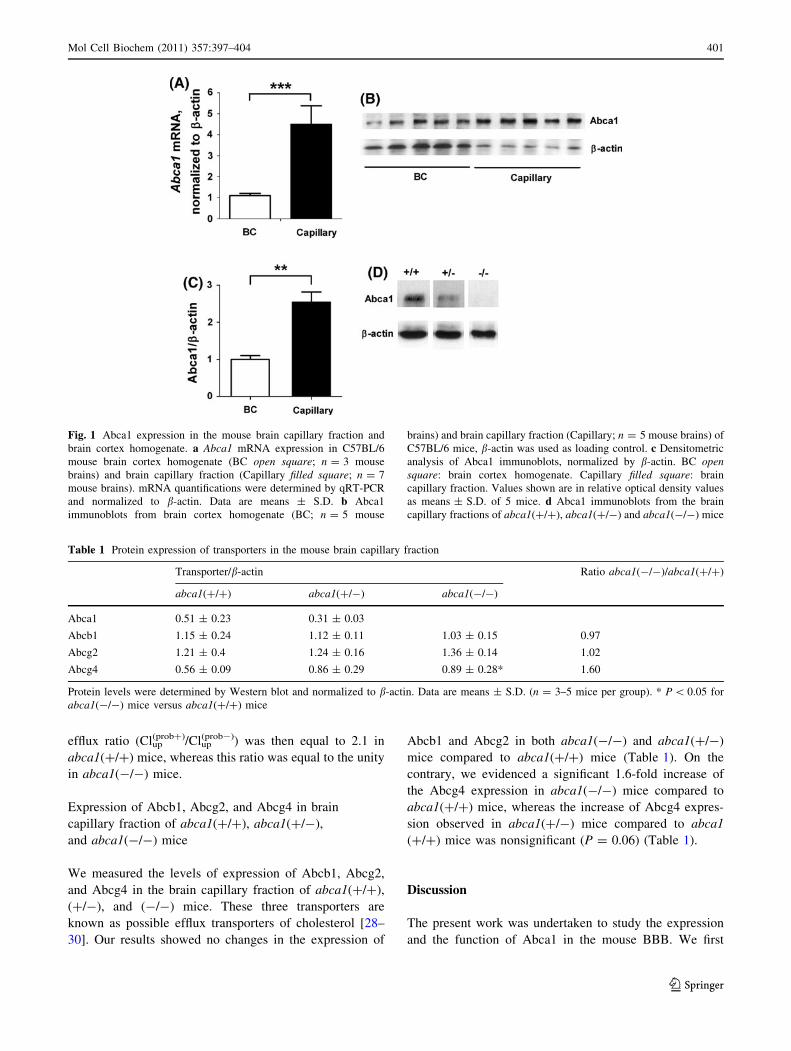
brain cortex homogenate (Fig. 1a).
The protein expression of Abca1 in C57BL/6 mice brain
capillary fraction was shown in Fig. 1b and c. The level of
Abca1 protein expression is 2.5-fold increased in the brain
capillary fraction compared to the brain cortex homogenate
(Fig. 1c).
To validate the model of Abca1-deficient mice, we
confirmed that Abca1 was expressed in the brain capillary
fraction of abca1(?/?) but not in that of abca1(-/-) mice
(Fig. 1d). We also observed a slight, but not significant
decrease of Abca1 expression in the brain capillary fraction
of abca1(?/-) mice, compared to abca1(?/?) mice
(Fig. 1d, Table 1).
Linearity of the cholesterol brain distribution over time
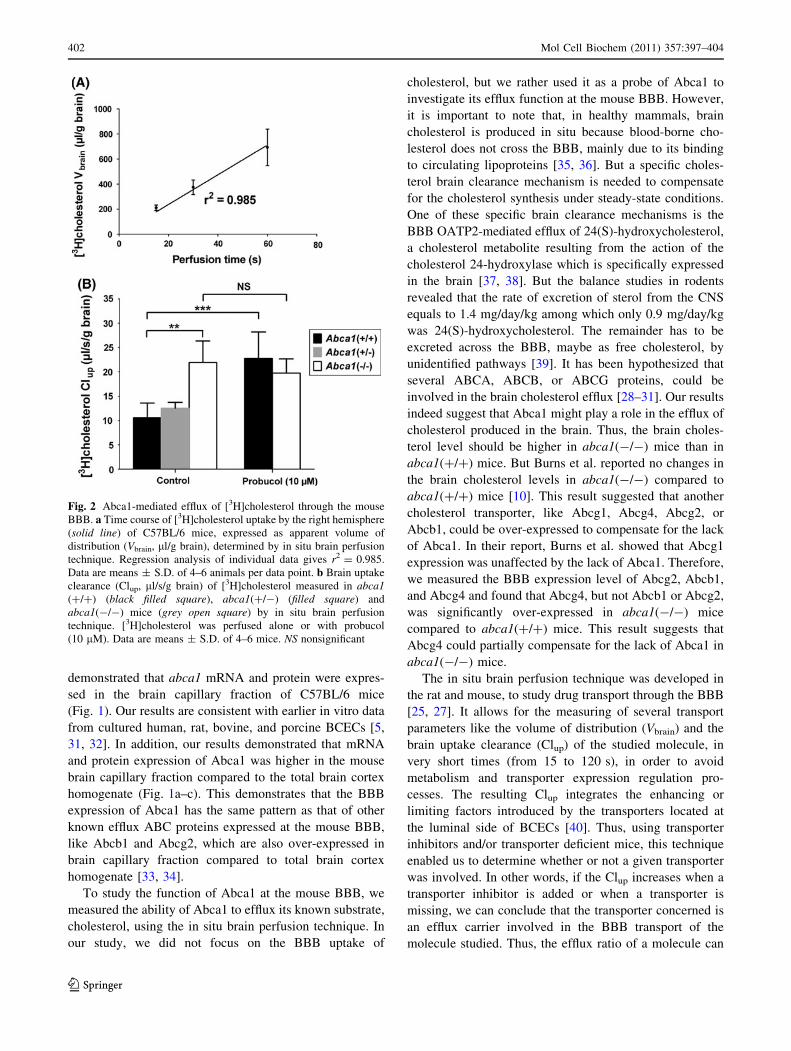
A first set of perfusion experiments was performed to study
the linearity of the brain volume of distribution (Vbrain, ll/g
brain) of [3H]cholesterol over time in wild-type C57BL/6
mice (Fig. 2a). The [3H]cholesterol Vbrain increased linearly
with the perfusion time suggesting that the transport rate
remained unchanged over time. A perfusion time of 60 s
was used for subsequent single-time point experiments.
Physical blood–brain barrier integrity
The brain vascular volume (Vvasc, ll/g brain) was measured
in each mouse by co-perfusing [14C]sucrose, a vascular
space marker, during the in situ brain perfusion experi-
ments. All the values were under 20 ll/g brain, indicating
that the BBB was intact during each experiment [25].
Abca1 function at the mouse BBB
The function of Abca1 at the BBB level was evaluated by
measuring the transport of [3H]cholesterol in abca1(?/?),
abca1(?/-), and abca1(-/-) mice, using the in situ brain
perfusion technique, with or without probucol. As shown in
Fig. 2b, the deficiency of Abca1 resulted in an increase of
107% in [3H]cholesterol Clup (P \ 0.01) in abca1(-/-)
mice (Clup = 21.93 ± 4.43 ll/s/g brain) as compared to
abca1(?/?) mice (Clup = 10.6 ± 2.98 ll/s/g brain). Thus,
the [3H]cholesterol efflux ratio (Clup(-/-)/Clup
(?/?)) was equal
to 2.1. The slight decrease of Abca1 expression in
abca1(?/-) mice resulted only in a nonsignificant increase
of the [3H]cholesterol Clup (Clup = 12.5 ± 1.2 ll/s/g
brain), compared to the abca1(?/?) mice. The addition of
probucol significantly increased the Clup of [3H]cholesterol
in abca1(?/?) mice by 115% (Clup = 22.74 ± 5.48 ll/s/g
brain) (P \ 0.01), but not in abca1(-/-) mice (Clup =
19.75 ± 2.90 ll/s/g brain) (Fig. 2b). The [3H]cholesterol
400 Mol Cell Biochem (2011) 357:397–404
123
efflux ratio (Clup(prob?)/Clup
(prob-)) was then equal to 2.1 in
abca1(?/?) mice, whereas this ratio was equal to the unity
in abca1(-/-) mice.
Expression of Abcb1, Abcg2, and Abcg4 in brain
capillary fraction of abca1(?/?), abca1(?/-),
and abca1(-/-) mice
We measured the levels of expression of Abcb1, Abcg2,
and Abcg4 in the brain capillary fraction of abca1(?/?),
(?/-), and (-/-) mice. These three transporters are
known as possible efflux transporters of cholesterol [28–
30]. Our results showed no changes in the expression of
Abcb1 and Abcg2 in both abca1(-/-) and abca1(?/-)
mice compared to abca1(?/?) mice (Table 1). On the
contrary, we evidenced a significant 1.6-fold increase of
the Abcg4 expression in abca1(-/-) mice compared to
abca1(?/?) mice, whereas the increase of Abcg4 expres-
sion observed in abca1(?/-) mice compared to abca1
(?/?) mice was nonsignificant (P = 0.06) (Table 1).
Discussion
The present work was undertaken to study the expression
and the function of Abca1 in the mouse BBB. We first
Fig. 1 Abca1 expression in the mouse brain capillary fraction and
brain cortex homogenate. a Abca1 mRNA expression in C57BL/6
mouse brain cortex homogenate (BC open square; n = 3 mouse
brains) and brain capillary fraction (Capillary filled square; n = 7
mouse brains). mRNA quantifications were determined by qRT-PCR
and normalized to b-actin. Data are means ± S.D. b Abca1
immunoblots from brain cortex homogenate (BC; n = 5 mouse
brains) and brain capillary fraction (Capillary; n = 5 mouse brains) of
C57BL/6 mice, b-actin was used as loading control. c Densitometric
analysis of Abca1 immunoblots, normalized by b-actin. BC opensquare: brain cortex homogenate. Capillary filled square: brain
capillary fraction. Values shown are in relative optical density values
as means ± S.D. of 5 mice. d Abca1 immunoblots from the brain
capillary fractions of abca1(?/?), abca1(?/-) and abca1(-/-) mice
Table 1 Protein expression of transporters in the mouse brain capillary fraction
Transporter/b-actin Ratio abca1(-/-)/abca1(?/?)
abca1(?/?) abca1(?/-) abca1(-/-)
Abca1 0.51 ± 0.23 0.31 ± 0.03
Abcb1 1.15 ± 0.24 1.12 ± 0.11 1.03 ± 0.15 0.97
Abcg2 1.21 ± 0.4 1.24 ± 0.16 1.36 ± 0.14 1.02
Abcg4 0.56 ± 0.09 0.86 ± 0.29 0.89 ± 0.28* 1.60
Protein levels were determined by Western blot and normalized to b-actin. Data are means ± S.D. (n = 3–5 mice per group). * P \ 0.05 for
abca1(-/-) mice versus abca1(?/?) mice
Mol Cell Biochem (2011) 357:397–404 401
123
demonstrated that abca1 mRNA and protein were expres-
sed in the brain capillary fraction of C57BL/6 mice
(Fig. 1). Our results are consistent with earlier in vitro data
from cultured human, rat, bovine, and porcine BCECs [5,
31, 32]. In addition, our results demonstrated that mRNA
and protein expression of Abca1 was higher in the mouse
brain capillary fraction compared to the total brain cortex
homogenate (Fig. 1a–c). This demonstrates that the BBB
expression of Abca1 has the same pattern as that of other
known efflux ABC proteins expressed at the mouse BBB,
like Abcb1 and Abcg2, which are also over-expressed in
brain capillary fraction compared to total brain cortex
homogenate [33, 34].
To study the function of Abca1 at the mouse BBB, we
measured the ability of Abca1 to efflux its known substrate,
cholesterol, using the in situ brain perfusion technique. In
our study, we did not focus on the BBB uptake of
cholesterol, but we rather used it as a probe of Abca1 to
investigate its efflux function at the mouse BBB. However,
it is important to note that, in healthy mammals, brain
cholesterol is produced in situ because blood-borne cho-
lesterol does not cross the BBB, mainly due to its binding
to circulating lipoproteins [35, 36]. But a specific choles-
terol brain clearance mechanism is needed to compensate
for the cholesterol synthesis under steady-state conditions.
One of these specific brain clearance mechanisms is the
BBB OATP2-mediated efflux of 24(S)-hydroxycholesterol,
a cholesterol metabolite resulting from the action of the
cholesterol 24-hydroxylase which is specifically expressed
in the brain [37, 38]. But the balance studies in rodents
revealed that the rate of excretion of sterol from the CNS
equals to 1.4 mg/day/kg among which only 0.9 mg/day/kg
was 24(S)-hydroxycholesterol. The remainder has to be
excreted across the BBB, maybe as free cholesterol, by
unidentified pathways [39]. It has been hypothesized that
several ABCA, ABCB, or ABCG proteins, could be
involved in the brain cholesterol efflux [28–31]. Our results
indeed suggest that Abca1 might play a role in the efflux of
cholesterol produced in the brain. Thus, the brain choles-
terol level should be higher in abca1(-/-) mice than in
abca1(?/?) mice. But Burns et al. reported no changes in
the brain cholesterol levels in abca1(-/-) compared to
abca1(?/?) mice [10]. This result suggested that another
cholesterol transporter, like Abcg1, Abcg4, Abcg2, or
Abcb1, could be over-expressed to compensate for the lack
of Abca1. In their report, Burns et al. showed that Abcg1
expression was unaffected by the lack of Abca1. Therefore,
we measured the BBB expression level of Abcg2, Abcb1,
and Abcg4 and found that Abcg4, but not Abcb1 or Abcg2,
was significantly over-expressed in abca1(-/-) mice
compared to abca1(?/?) mice. This result suggests that
Abcg4 could partially compensate for the lack of Abca1 in
abca1(-/-) mice.
The in situ brain perfusion technique was developed in
the rat and mouse, to study drug transport through the BBB
[25, 27]. It allows for the measuring of several transport
parameters like the volume of distribution (Vbrain) and the
brain uptake clearance (Clup) of the studied molecule, in
very short times (from 15 to 120 s), in order to avoid
metabolism and transporter expression regulation pro-
cesses. The resulting Clup integrates the enhancing or
limiting factors introduced by the transporters located at
the luminal side of BCECs [40]. Thus, using transporter
inhibitors and/or transporter deficient mice, this technique
enabled us to determine whether or not a given transporter
was involved. In other words, if the Clup increases when a
transporter inhibitor is added or when a transporter is
missing, we can conclude that the transporter concerned is
an efflux carrier involved in the BBB transport of the
molecule studied. Thus, the efflux ratio of a molecule can
Fig. 2 Abca1-mediated efflux of [3H]cholesterol through the mouse
BBB. a Time course of [3H]cholesterol uptake by the right hemisphere
(solid line) of C57BL/6 mice, expressed as apparent volume of
distribution (Vbrain, ll/g brain), determined by in situ brain perfusion
technique. Regression analysis of individual data gives r2 = 0.985.
Data are means ± S.D. of 4–6 animals per data point. b Brain uptake
clearance (Clup, ll/s/g brain) of [3H]cholesterol measured in abca1(?/?) (black filled square), abca1(?/-) (filled square) and
abca1(-/-) mice (grey open square) by in situ brain perfusion
technique. [3H]cholesterol was perfused alone or with probucol
(10 lM). Data are means ± S.D. of 4–6 mice. NS nonsignificant
402 Mol Cell Biochem (2011) 357:397–404
123

be calculated [26]. In our case, we calculated the
[3H]cholesterol efflux ratio by comparing abca1(-/-) and
abca1(?/?) mice or by comparing mice perfused with or
without probucol.
In the current study, we demonstrated that the [3H]cho-
lesterol Clup was enhanced when probucol was co-perfused
with [3H]cholesterol in abca1(?/?) mice (Fig. 2b). The
[3H]cholesterol efflux ratio (Clup(prob?)/Clup
(prob-)) was calcu-
lated at 2.1, suggesting that probucol inhibited an efflux
transporter, likely to be Abca1. This result agrees with a
preliminary study, in which we showed that the brain cho-
lesterol Clup was enhanced when probucol was added [41].
To demonstrate that the probucol-sensitive transporter
evidenced here was Abca1, we measured the [3H]choles-
terol Clup in abca1(-/-) mice with and without probucol.
Without probucol, the [3H]cholesterol Clup was signifi-
cantly increased in abca1(-/-) mice compared to
abca1(?/?) mice (Fig. 2b), meaning that Abca1 was
actually involved in the BBB cholesterol efflux. The value
of the [3H]cholesterol efflux ratio (Clup(-/-)/Clup
(?/?)) was
equal to (Clup(prob?)/Clup
(prob-)) ratio obtained in abca1(?/?)
mice, suggesting that probucol inhibited specifically Abca1.
This is in agreement with the literature, since, to our
knowledge, probucol has been shown to inhibit solely
Abca1 [15, 42]. To confirm this point, we co-perfused
probucol with [3H]cholesterol in abca1(-/-) mice
(Fig. 2b). The lack of Abca1 led to the lack of the
probucol effect as illustrated by the value of the efflux ratio
(Clup(prob?)/Clup
(prob-)) obtained in abca1(-/-) mice, which
failed to unity. This result confirmed the specific effect of
probucol on Abca1-mediated transport of cholesterol at the
mouse BBB level. In addition, the higher [3H]cholesterol
Clup obtained in abca1(-/-) mice compared to abca1(?/
?) mice perfused without probucol suggests that the BBB
over-expression of Abcg4 is not sufficient to fully com-
pensate for the lack of Abca1.
Altogether, our results strongly support the hypothesis
that Abca1 is a cholesterol efflux transporter at the mouse
BBB. Our results are in agreement with two in vitro studies
in which Abca1 was shown to transport cholesterol through
cultures of bovine and porcine BCECs [31, 32]. But to our
knowledge, we have been the first to demonstrate, in vivo,
the efflux function of Abca1 at the mouse BBB level.
Conclusion
In summary, our results evidenced that Abca1 is enriched
in mouse brain capillaries compared to total brain cortex,
reminiscent to the expression pattern of other ABC trans-
porters ABCB1 and ABCG2. In addition, our results
underscore the role of Abca1 as an efflux transporter at the
mouse BBB. Finally, our data demonstrate that the lack of
Abca1 resulted in an over-expression of Abcg4, to partially
compensate for the lack of Abca1-mediated efflux of
cholesterol. Bearing in mind that several relationships were
established between cerebral cholesterol homeostasis and
AD, our results encourage further investigations to eluci-
date if Abca1- and/or Abcg4-mediated cholesterol efflux at
the mouse BBB could be one of the key segments of the
AD-cholesterol link.
Acknowledgments We thank Chantal Guillemette for helping us
with the qRT-PCR technique. We thank Valerie Domergue-Dupont,
the head of the central animal house of the faculty of Pharmacy, for
taking care of animals. The authors thank the Banting Research
Foundation for their financial supports.
References
1. Krimbou L, Denis M, Haidar B, Carrier M, Marcil M, Genest J Jr
(2004) Molecular interactions between apoE and ABCA1: impact
on apoE lipidation. J Lipid Res 45(5):839–848
2. Klaassen CD, Aleksunes LM (2010) Xenobiotic, bile acid, and
cholesterol transporters: function and regulation. Pharmacol Rev
62(1):1–96
3. Wellington CL, Walker EK, Suarez A, Kwok A, Bissada N,
Singaraja R, Yang YZ, Zhang LH, James E, Wilson JE, Francone
O, McManus BM, Hayden MR (2002) ABCA1 mRNA and
protein distribution patterns predict multiple different roles and
levels of regulation. Lab Invest 82(3):273–283
4. Karasinska JM, Rinninger F, Lutjohann D, Ruddle P, Franciosi S,
Kruit JK, Singaraja RR, Hirsch-Reinshagen V, Fan J, Brunham
LR, Bissada N, Ramakrishnan R, Wellington CL, Parks JS,
Hayden MR (2009) Specific loss of brain ABCA1 increases brain
cholesterol uptake and influences neuronal structure and function.
J Neurosci 29(11):3579–3589
5. Ohtsuki S, Watanabe Y, Hori S, Suzuki H, Bhongsatiern J,
Fujiyoshi M, Kamoi M, Kamiya N, Takanaga H, Terasaki T
(2004) mRNA expression of the ATP-binding cassette transporter
subfamily A (ABCA) in rat and human brain capillary endothelial
cells. Biol Pharm Bull 27(9):1437–1440
6. Dutheil F, Jacob A, Dauchy S, Beaune P, Scherrmann JM,
Decleves X, Loriot MA (2010) ABC transporters and cyto-
chromes P450 in the human central nervous system: influence on
brain pharmacokinetics and contribution to neurodegenerative
disorders. Expert Opin Drug Metab Toxicol 6(10):1161–1174
7. Nicolazzo JA, Katneni K (2009) Drug transport across the blood–
brain barrier and the impact of breast cancer resistance protein
(ABCG2). Curr Top Med Chem 9(2):130–147
8. Ueno M, Nakagawa T, Wu B, Onodera M, Huang CL, Kusaka T,
Araki N, Sakamoto H (2010) Transporters in the brain endothelial
barrier. Curr Med Chem 17(12):1125–1138
9. Urquhart BL, Kim RB (2009) Blood–brain barrier transporters
and response to CNS-active drugs. Eur J Clin Pharmacol 65(11):
1063–1070
10. Burns MP, Vardanian L, Pajoohesh-Ganji A, Wang L, Cooper M,
Harris DC, Duff K, Rebeck GW (2006) The effects of ABCA1 on
cholesterol efflux and Abeta levels in vitro and in vivo. J Neu-
rochem 98(3):792–800
11. Koldamova R, Fitz NF, Lefterov I (2010) The role of ATP-
binding cassette transporter A1 in Alzheimer’s disease and neu-
rodegeneration. Biochim Biophys Acta 1801(8):824–830
12. Kim WS, Bhatia S, Elliott DA, Agholme L, Kagedal K, McCann
H, Halliday GM, Barnham KJ, Garner B (2010) Increased
Mol Cell Biochem (2011) 357:397–404 403
123

ATP-binding cassette transporter A1 expression in Alzheimer’s
disease hippocampal neurons. J Alzheimers Dis 21(1):193–205
13. Xiong H, Callaghan D, Jones A, Walker DG, Lue LF, Beach TG,
Sue LI, Woulfe J, Xu H, Stanimirovic DB, Zhang W (2008)
Cholesterol retention in Alzheimer’s brain is responsible for high
beta- and gamma-secretase activities and Abeta production.
Neurobiol Dis 29(3):422–437
14. Grimm MO, Grimm HS, Tomic I, Beyreuther K, Hartmann T,
Bergmann C (2008) Independent inhibition of Alzheimer disease
beta- and gamma-secretase cleavage by lowered cholesterol
levels. J Biol Chem 283(17):11302–11311
15. Favari E, Zanotti I, Zimetti F, Ronda N, Bernini F, Rothblat GH
(2004) Probucol inhibits ABCA1-mediated cellular lipid efflux.
Arterioscler Thromb Vasc Biol 24(12):2345–2350
16. McNeish J, Aiello RJ, Guyot D, Turi T, Gabel C, Aldinger C,
Hoppe KL, Roach ML, Royer LJ, de Wet J, Broccardo C, Chi-
mini G, Francone OL (2000) High density lipoprotein deficiency
and foam cell accumulation in mice with targeted disruption of
ATP-binding cassette transporter-1. Proc Natl Acad Sci U S A
97(8):4245–4250
17. Triguero D, Buciak J, Pardridge WM (1990) Capillary depletion
method for quantification of blood–brain barrier transport of
circulating peptides and plasma proteins. J Neurochem 54(6):
1882–1888
18. Rousselle C, Clair P, Lefauconnier JM, Kaczorek M, Scherrmann
JM, Temsamani J (2000) New advances in the transport of
doxorubicin through the blood–brain barrier by a peptide vector-
mediated strategy. Mol Pharmacol 57(4):679–686
19. Dallaire L, Tremblay L, Beliveau R (1991) Purification and
characterization of metabolically active capillaries of the blood–
brain barrier. Biochem J 276(Pt 3):745–752
20. Gebhardt FM, Scott HA, Dodd PR (2010) Housekeepers for
accurate transcript expression analysis in Alzheimer’s disease
autopsy brain tissue. Alzheimers Dement 6(6):465–474
21. Weickert CS, Sheedy D, Rothmond DA, Dedova I, Fung S,
Garrick T, Wong J, Harding AJ, Sivagnanansundaram S, Hunt C,
Duncan C, Sundqvist N, Tsai SY, Anand J, Draganic D, Harper C
(2010) Selection of reference gene expression in a schizophrenia
brain cohort. Aust N Z J Psychiatry 44(1):59–70
22. Kim WS, Guillemin GJ, Glaros EN, Lim CK, Garner B (2006)
Quantitation of ATP-binding cassette subfamily-A transporter
gene expression in primary human brain cells. Neuroreport 17(9):
891–896
23. Su YR, Linton MF, Fazio S (2002) Rapid quantification of
murine ABC mRNAs by real time reverse transcriptase-poly-
merase chain reaction. J Lipid Res 43(12):2180–2187
24. Milane A, Fernandez C, Vautier S, Bensimon G, Meininger V,
Farinotti R (2007) Minocycline and riluzole brain disposition:
interactions with p-glycoprotein at the blood–brain barrier.
J Neurochem 103(1):164–173
25. Dagenais C, Rousselle C, Pollack GM, Scherrmann JM (2000)
Development of an in situ mouse brain perfusion model and its
application to mdr1a P-glycoprotein-deficient mice. J Cereb
Blood Flow Metab 20(2):381–386
26. Zhao R, Kalvass JC, Pollack GM (2009) Assessment of blood–
brain barrier permeability using the in situ mouse brain perfusion
technique. Pharm Res 26(7):1657–1664
27. Takasato Y, Rapoport SI, Smith QR (1984) An in situ brain
perfusion technique to study cerebrovascular transport in the rat.
Am J Physiol 247(3 Pt 2):H484–H493
28. Janvilisri T, Venter H, Shahi S, Reuter G, Balakrishnan L, van
Veen HW (2003) Sterol transport by the human breast cancer
resistance protein (ABCG2) expressed in Lactococcus lactis.
J Biol Chem 278(23):20645–20651
29. Koshiba S, An R, Saito H, Wakabayashi K, Tamura A, Ishikawa
T (2008) Human ABC transporters ABCG2 (BCRP) and ABCG4.
Xenobiotica 38(7–8):863–888
30. Le Goff W, Settle M, Greene DJ, Morton RE, Smith JD (2006)
Reevaluation of the role of the multidrug-resistant P-glycoprotein
in cellular cholesterol homeostasis. J Lipid Res 47(1):51–58
31. Gosselet F, Candela P, Sevin E, Berezowski V, Cecchelli R,
Fenart L (2009) Transcriptional profiles of receptors and trans-
porters involved in brain cholesterol homeostasis at the blood–
brain barrier: use of an in vitro model. Brain Res 1249:34–42
32. Panzenboeck U, Balazs Z, Sovic A, Hrzenjak A, Levak-Frank S,
Wintersperger A, Malle E, Sattler W (2002) ABCA1 and scav-
enger receptor class B, type I, are modulators of reverse sterol
transport at an in vitro blood–brain barrier constituted of porcine
brain capillary endothelial cells. J Biol Chem 277(45):
42781–42789
33. Cisternino S, Mercier C, Bourasset F, Roux F, Scherrmann JM
(2004) Expression, up-regulation, and transport activity of the
multidrug-resistance protein Abcg2 at the mouse blood–brain
barrier. Cancer Res 64(9):3296–3301
34. Warren MS, Zerangue N, Woodford K, Roberts LM, Tate EH,
Feng B, Li C, Feuerstein TJ, Gibbs J, Smith B, de Morais SM,
Dower WJ, Koller KJ (2009) Comparative gene expression pro-
files of ABC transporters in brain microvessel endothelial cells
and brain in five species including human. Pharmacol Res
59(6):404–413
35. Pardridge WM, Mietus LJ (1980) Palmitate and cholesterol
transport through the blood–brain barrier. J Neurochem 34(2):
463–466
36. Jurevics H, Morell P (1995) Cholesterol for synthesis of myelin is
made locally, not imported into brain. J Neurochem 64(2):
895–901
37. Lutjohann D, Breuer O, Ahlborg G, Nennesmo I, Siden A, Dic-
zfalusy U, Bjorkhem I (1996) Cholesterol homeostasis in human
brain: evidence for an age-dependent flux of 24S-hydroxycho-
lesterol from the brain into the circulation. Proc Natl Acad Sci
USA 93(18):9799–9804
38. Ohtsuki S, Ito S, Matsuda A, Hori S, Abe T, Terasaki T (2007)
Brain-to-blood elimination of 24S-hydroxycholesterol from rat
brain is mediated by organic anion transporting polypeptide 2
(oatp2) at the blood–brain barrier. J Neurochem 103(4):1430–
1438
39. Xie C, Lund EG, Turley SD, Russell DW, Dietschy JM (2003)
Quantitation of two pathways for cholesterol excretion from the
brain in normal mice and mice with neurodegeneration. J Lipid
Res 44(9):1780–1789
40. Cisternino S, Rousselle C, Lorico A, Rappa G, Scherrmann JM
(2003) Apparent lack of Mrp1-mediated efflux at the luminal side
of mouse blood–brain barrier endothelial cells. Pharm Res
20(6):904–909
41. Cattelotte J, Andre P, Ouellet M, Bourasset F, Scherrmann JM,
Cisternino S (2008) In situ mouse carotid perfusion model: glu-
cose and cholesterol transport in the eye and brain. J Cereb Blood
Flow Metab 28(8):1449–1459
42. Taylor JM, Borthwick F, Bartholomew C, Graham A (2010)
Overexpression of steroidogenic acute regulatory protein
increases macrophage cholesterol efflux to apolipoprotein AI.
Cardiovasc Res 86(3):526–534
404 Mol Cell Biochem (2011) 357:397–404
123





























