Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Diversity of thermophilic populations during thermophilicaerobic digestion of potato peel slurryJ.O. Ugwuanyi, L.M. Harvey and B. McNeil
Strathclyde Fermentation Centre, University of Strathclyde, Glasgow, UK
Introduction
Thermophilic aerobic digestion (TAD) of waste arises as
a result of conservation of the heat generated during
microbial aerobic metabolism, as a result of which the tem-
perature of the waste undergoing digestion rises to thermo-
philic levels (‡45�C) (Surucu et al. 1975; Heinonen-Tanski
et al. 2005). It is a versatile process, which is increasingly
accepted for waste treatment, either in stand-alone reac-
tions, or as part of integrated two stage processes (Messen-
ger and Ekama 1993). Relative to classical waste process
such as activated sludge, TAD is a new process (Surucu
et al. 1975), and its advantages include the capacity for
rapid, cost-effective waste stabilization (Hawash et al.
1994) and pasteurization (Ugwuanyi et al. 1999). It is
encouraged in Europe and North America, where various
legislations seek to limit the levels of human and animal
pathogens in wastes intended for land or sea application
and disposal (EPA, 1992; Council of the European Union.
2001). Of particular acclaim is the potential for use of
TAD in upgrading, by protein enrichment, of a variety of
agricultural wastes for reuse in animal feed (Couillard and
Zhu 1990; Ugwuanyi et al. 2006). This application has con-
siderable implications for global food security; especially
in the tropics where on account of inadequate food supply,
animals and humans often compete directly for the same
sources of calories.
New studies are emerging that seek to properly charac-
terize TAD (Chu et al. 1997; Ugwuanyi et al. 2004,
2005a,b; Cibis et al. 2006). However, in spite of these, lit-
tle has been performed to properly understand the micro-
biology of the process (Cibis et al. 2002). This is due to
both the newness of the process and also because it has
tended to develop mainly along the lines of public health
Keywords
agricultural slurry, microbial diversity,
thermophilic aerobic digestion, thermophilic
Bacillus spp.
Correspondence
J. O. Ugwuanyi, Department of Microbiology,
University of Nigeria, Nsukka, Nigeria.
E-mail: [email protected]
2007 ⁄ 0036: received 10 January 2007,
revised 22 May 2007 and accepted 29 June
2007
doi:10.1111/j.1365-2672.2007.03557.x
Abstract
Aims: To study the diversity of thermophiles during thermophilic aerobic
digestion (TAD) of agro-food waste slurries under conditions similar to full-
scale processes.
Methods and Results: Population diversity and development in TAD were
studied by standard microbiological techniques and the processes monitored by
standard fermentation procedures. Facultative thermophiles were identified as
Bacillus coagulans and B. licheniformis, while obligate thermophiles were identi-
fied as B. stearothermophilus. They developed rapidly to peaks of 107 to 108 in
£48 h. Stability of obligate thermophiles increased with process temperatures.
Thermophiles were unstable at process pH above or below neutral, but devel-
oped rapidly at all aeration rates. Peak populations were higher in the median
than at extremes of aeration rates. Obligate thermophiles were unstable at low
aeration rates. Process self-heating was higher at lower than at higher aeration
rate. Beyond 96 h most thermophiles were present as spores.
Conclusions: Limited range of indigenous thermophiles drives TAD of slurry.
They develop rapidly and are stable at most digestion conditions.
Significance and Impact of the Study: Development and stability of thermo-
philes in TAD suggest that the process may be operated in a wide range of
conditions; and even at short HRT in continuous processes without compro-
mising efficiency.
Journal of Applied Microbiology ISSN 1364-5072
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 79

engineering (Mohaibes and Heinonen-Tanski 2004). In
addition, outside biogasification and use of caldoactive
species for pure culture high value applications (Krahe
et al. 1996), use of thermophiles in bioprocesses was
essentially unknown before the 1970s (Brock 1986).
Sonnleitner and Fiechter (1983a,b)studied the microbi-
ology of sewage sludge undergoing TAD at 50–67�C and
characterized at least 95% of the isolates as Bacillus spp.
with maximal growth temperatures in excess of 70�C.
They concluded that the active populations responsible
for heat generation during TAD consist entirely of mem-
bers of the extremely thermophilic B. stearothermophilus.
However, Loll (1976) reported that thermophilic and
thermotolerant Bacillus spp. were together responsible
for the stabilization of model wastewater. During TAD
of swine slurry, Beaudet et al. (1990) counted microbial
populations at 55�C, varying from 104 to 107 ml)1 and
identified them as Bacillus spp., including B. lichenifor-
mis. Burt et al. (1990) enumerated 65 and 55�C thermo-
philes in excess of 106 and 108 ml)1, respectively, during
TAD of sewage sludge, while Malladi and Ingham
(1993) reported thermophilic aerobic spore-former
counts of up to 109 ml)1 during TAD of potato process
wastewater. These populations, which develop from the
proliferation of thermophiles indigenous in the waste
(Burt et al. 1990), fluctuate by up to five orders of
magnitude, but are stable enough not to washout in
continuous processes even in short hydraulic retention
time (Sonnleitner and Fiechter 1983a), particularly at
the higher end of the digestion temperatures (Beaudet
et al. 1990).
The paucity of information on the microbiology of
TAD has left room for speculations on the diversity of
micro-organisms in the process. Even the limited infor-
mation that exists has been based on reactions that
employed sewage sludge (Sonnleitner and Fiechter,
1983a,b). This waste type has limitations as the basis for
projection to other more diverse and potentially reusable
(particularly in animal nutrition) wastes. The possibility
of upgrading and recycling of wastes by TAD (Couillard
and Zhu 1990; Ugwuanyi et al. 2006) requires that further
studies on the microbiology of the process be imple-
mented, particularly using such wastes that have potential
for reuse in animal nutrition. The importance of such
studies include the capacity to give a fair picture of the
microbiological safety and limitations of treated waste,
and also for making projections on process capabilities
and reproducibility in the production of materials re-
entering the animal and human food chain. It is impor-
tant for such studies to take account of variables likely to
obtain in full-scale operations. We have reported on the
efficiency of TAD in the treatment of potato peel waste
(Ugwuanyi et al. 2004, 2005a,b). It is the object of this
current work to address some of the microbiological
knowledge gaps in TAD.
Materials and methods
Preparation of model waste for digestion
Peel waste was manually generated in large batches from
the same type of locally procured white potatoes (Pent-
lands variety) for batch digestion. The potatoes were
washed in lukewarm tap water to remove dirt and air-
dried before peeling. Peel was stored in polythene bags in
batches of 2Æ0 kg at )20�C until required. Peel was
defrosted in water bath at 40�C immediately prior to use,
and blended using a commercial blender (Waring Co.,
MA, USA) to a fine paste capable of passing through a
250-lm mesh. The blended peel was used without any
further treatment unless otherwise specified. Minimal
mineral supplement was added to aid stability of thermo-
philic enzymes in the slurry (Tanner 1997). The composi-
tion of the mineral solution used to make up peel waste
volume was (g l)1); CaCl2, 0Æ1; MgSO4, 0Æ1; K2HPO4,
0Æ03; KH2PO4, 0Æ03 (Tanner 1997). The crude (Kjeldahl)
nitrogen content was determined to be approx. 2% (dry
weight basis). The total solids content of waste employed
in a digestion batch was determined by reference to the
dry matter content of peel and quantity of slurry digested
was then fixed at 0Æ2 kg l)1 of waste (wet weight, corre-
sponding to approx. 8Æ0 g l)1 soluble chemical oxygen
demand) (Ugwuanyi et al. 2004, 2005b).
Reactor
All digestion was carried out as batch processes in a fully
instrumented continuously stirred tank reactor CSTR,
IMCS 2000 [Multiple Bioreactor and Sterile Plants (MBR
AG) Zurich, Switzerland] of 15-l total volume (12-l work-
ing volume).
Digestion conditions
Digestion at different temperatures
Digestions were carried out at 45, 50, 55, 60 and 65�C to
study the effect of digestion temperatures on the develop-
ment of thermophilic populations in the slurry. Digestion
temperatures were selected to reflect what occurs in a
full-scale self-heating process (EPA, 1990) and what has
been variously considered optimal process temperatures
(Ginnivan et al. 1981; Messenger and Ekama 1993; LaPara
and Alleman 1999; Ugwuanyi et al. 2005a,b). The lower
temperature, 45�C was selected as the minimum at
which digestion may be considered thermophilic (Surucu
et al. 1975). The higher temperature 65�C was the first
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
80 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors

temperature at which the indigenous populations could
not start the digestion spontaneously. (Although thermo-
philic populations remained viable in low numbers in the
slurry, reaction failed to start as determined by DO and
other process variables). Microbiological data for this
reaction are therefore not presented. Digestions at differ-
ent temperatures were carried out without pH regulation,
while the aeration rate was fixed at 0Æ5-vvm (volume air
per volume slurry per minute). Changes in the pH of
slurry were recorded at 6- or 12-h intervals.
Control reactions in which in addition to the pH the
temperature was also not regulated were conducted at
0Æ5 and 1Æ0 vvm. Changes in temperature and pH of the
control reactions were recorded at 6- or 12-h intervals.
For these reactions, the external wall of the reactor was
insulated with 5-cm thick, dark PU foam to minimize
heat loss and ensure that the temperature of the slurry
rose to thermophilic levels without any heat input other
than from the metabolic heat and bioreactor moving
parts.
Digestion at different pH values
Digestions at the different pH values were conducted at
55�C based on superior performance and earlier studies
(Beaudet et al. 1990; Ugwuanyi et al. 2004, 2005a,b).
These reactions were implemented at 0Æ5 vvm. The pH
was automatically controlled at 6Æ0, 7Æ0, 8Æ0, 9Æ0 and 9Æ5using 2 mol l–1 H2SO4 and 2 mol l–1 NaOH. Above pH
9Æ5, TAD failed to start. Therefore, no digestions were
carried out beyond pH 9Æ5. Control reaction in which the
pH was not regulated was also included at the defined
temperature and aeration rate and the pH was monitored
at 6- and 12-h intervals.
Digestion at different aeration rates
To study the effect of aeration rates on the development
of thermophilic populations, digestions were carried out
at 0Æ1, 0Æ25, 0Æ5 and 1Æ0 vvm. All aeration rate studies
were carried out at 55�C and pH 7Æ0.
General digestion conditions
Digestions were carried out for 156 h (maximum of
240 h during temperature studies) and terminated when
on-line parameters [dissolved oxygen, oxygen uptake
and carbon dioxide evolution rates and pH (in unregula-
ted reactions)] did not change appreciably over a consec-
utive 24-h period. All digestions were carried out without
any inoculation, by using the indigenous microbial popu-
lations in the waste. Foaming was controlled manually
using sterile polypropylene glycol (BDH) antifoam,
pumped in as necessary. Agitation rate was fixed at
350 rev min–1 throughout all the studies. Any drop in the
volume of waste slurry was noted and the volume was
made up with sterile distilled water at the same tempera-
ture as the digesting slurry, at least 10 min before
sampling.
Sampling
Approximately 50-ml well mixed digestion slurry sample
was collected in a sterile sample bottle at regular 12-h
intervals for the first 36- and 24-h interval thereafter for
the duration of the reaction and processed for microbial
analysis as described below. Samples were analysed imme-
diately on collection (while being maintained in the water
bath at the same temperature as the reactor until inocula-
tion was completed). In all digestions the total volume of
slurry collected as sample for the duration of reaction
remained below 10% of total reaction volume.
On-line determinations
Temperature, pH and dissolved oxygen were measured by
using probes calibrated at the study temperature. Probes
were calibrated at pH 4Æ0, 7Æ0 and 10Æ0 for use in unregu-
lated pH reactions, and at 45�C during unregulated
temperature reaction. Dissolved oxygen and pH probes
were obtained from Ingold Ltd (Switzerland), while
temperature probe was as supplied with the bioreactor.
Oxygen in the exit gas was measured using a paramagnetic
analyser, Servomex, Model 500 (Sybron Taylor Ltd,
Crowborough, UK), while carbon dioxide was measured by
using an infrared analyser ADC, Model 7000 (Analytical
Development Corp., Cambridge, UK).
Enumeration of viable thermophilic populations
Samples were homogenized by mixing briskly in a wrist
action shaker (Gallenkamp, UK). Successive 1 : 10 dilu-
tions of samples were made in sterile peptone water
maintained at 45�C in a water bath and 0Æ1 ml of dilution
was plated by standard spread plate method (Collins et al.
1989) on predried nutrient agar (NA) and plate count
agar (PCA) plates maintained at 45�C before inoculation.
NA and PCA were prepared by fortifying with additional
1Æ5% bacteriological agar (No. 1) and wrapped in plastic
bags to minimize drying before incubation in a static air
incubator at 55 and 65�C for 24–48 h. Count of spores
were carried out after inactivating vegetative cells by heat-
ing samples at 80�C for 10 min in a water bath before
plating and incubating as for total count. All media were
procured from Oxoid Ltd (Basingstoke, UK). Total viable
counts were carried out in triplicate on each of the two
media for each dilution and incubation temperature. By
incubating at two temperatures, the active populations
were separated into obligate thermophiles, which are the
only ones that grow at 65�C, and facultative plus obligate
thermophiles (which together were enumerated at 55�C
J.O. Ugwuanyi et al. Diversity of thermophiles during thermophilic aerobic digestion
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 81

and constitute the total thermophilic population). The
difference in population obtained at both temperatures
was due to facultative thermophiles that do not grow at
65�C.
Isolation and purification of thermophiles
Isolates which showed distinct morphological features on
NA plates were isolated and purified on further NA
plates. To avoid duplication of populations, as 65�C pop-
ulations are capable of growth at the lower temperature,
isolates obtained at 55�C were tested for their ability to
grow at 65�C, and those that grew at 65�C were included
among the obligate thermophiles for identification.
Biochemical identification of thermophiles
Purified isolates were maintained on NA at 4�C. They
were classified into two groups depending on temperature
of their primary isolation, as well as on the maximum
temperature of growth, as facultative thermophiles enu-
merated at 55�C with a maximum growth temperature
not exceeding 60�C, or obligate thermophiles enumerated
at 65�C with growth temperature extending up to 70�C.
Within each temperature group, populations were
grouped on the basis of their colonial ⁄ cultural character-
istics and microscopic morphology including presence
and nature of endospores. Preliminary tests employed for
the initial characterization of the isolates were based on
the method described by Berkeley et al. (1984) and
Rowan (1996). Sporulation of isolates was determined,
after incubation for a minimum of 96 h in manganous
ion-enriched nutrient broth sporulation medium (Sonn-
leitner and Fiechter 1983a).
Identification was achieved by use of API 50CHB (Bio-
merieux, Basingstoke, UK) for Bacillus spp. supplemented
with the first 12 tests of API 20E (Berkeley et al. 1984;
Rowan 1996). The test isolate was prepared for identifica-
tion by growing it overnight on NA at the appropriate
temperature. A thick suspension of the isolate, prepared
by rubbing off the growth with sterile swab and emulsify-
ing it in sterile normal saline, was used to prepare the
inoculum as described by kit manufacturers, and all test
kits were incubated at 55�C in moist chambers.
The results obtained from the kits were interpreted
using the apilab plus software (Biomerieux, Basingstoke,
UK). In cases where a definitive result could not be
obtained from the APILAB system, because of colour fad-
ing arising from the high temperature of incubation, the
results of the biochemical tests were used to identify iso-
lates by reference to Bergey’s Manual of Systematic Bacte-
riology (Sneath 1984) and the diagnostic profile described
by Berkeley et al. (1984). Identification tests were re-
peated twice in all cases and the results compared with
eliminate ambiguities.
Analysis of results
Results obtained were subjected to anova (single classifi-
cation) using instat� statistical package (Statistical
Services Centre, University of Reading, Reading, UK).
Differences between treatment means were tested using
Tukey–Kramer’s multiple comparison test. Fermenter
experiments were conducted in duplicates. All assays were
carried out in triplicate unless otherwise stated and the
results are reported as the mean of the replicate determi-
nations.
Results
Effect of digestion temperature on development
of thermophiles
Thermophilic populations developed rapidly at all the
digestion temperatures (Fig. 1), reaching peak population
in approx. 48 h. During digestion at 45�C, the total ther-
mophiles (55�C populations) rose rapidly to a peak of
just under 9-log colony forming units (CFU) per millilitre
of slurry in 48 h (Fig. 1A). This population is five orders
of magnitude higher than in the raw waste. Following this
peak, the population decreased gradually for the duration
of reaction. The population of obligate thermophiles rose
from approx. 3-log CFU in the raw waste to peak of
5-log CFU per millilitre in 48 h. The peak count was
maintained for <48 h before dropping dramatically to
population similar to that of the raw waste. At this
temperature, obligate thermophiles accounted for only
0Æ05% of the total population at peak count.
The population of obligate thermophiles increased rap-
idly with temperature of digestion, and at 50�C it
accounted for over 90% of total thermophiles in 48 h.
However, as at 45�C obligate thermophiles declined sig-
nificantly after reaching the peak, and at end of the diges-
tion accounted for only approx. 10% of the total
population. As the digestion temperature increased the
population of obligate thermophiles also increased,
accounting for a higher proportion of the total popula-
tion. During digestion at 55�C, obligate thermophiles
accounted for over 95% of the total population at peak
count (Fig. 1C), and at 60�C for between 99% and 100%,
resulting in an essentially uniform population of obligate
thermophiles (Fig. 1D).
The decline in pH of the digesting slurry appeared to
be temperature-dependent. During digestion at 45 and
50�C, the pH dropped slightly in the first 12 h (remain-
ing above 5Æ0), before rising rapidly (Fig. 1A,B). The
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
82 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors
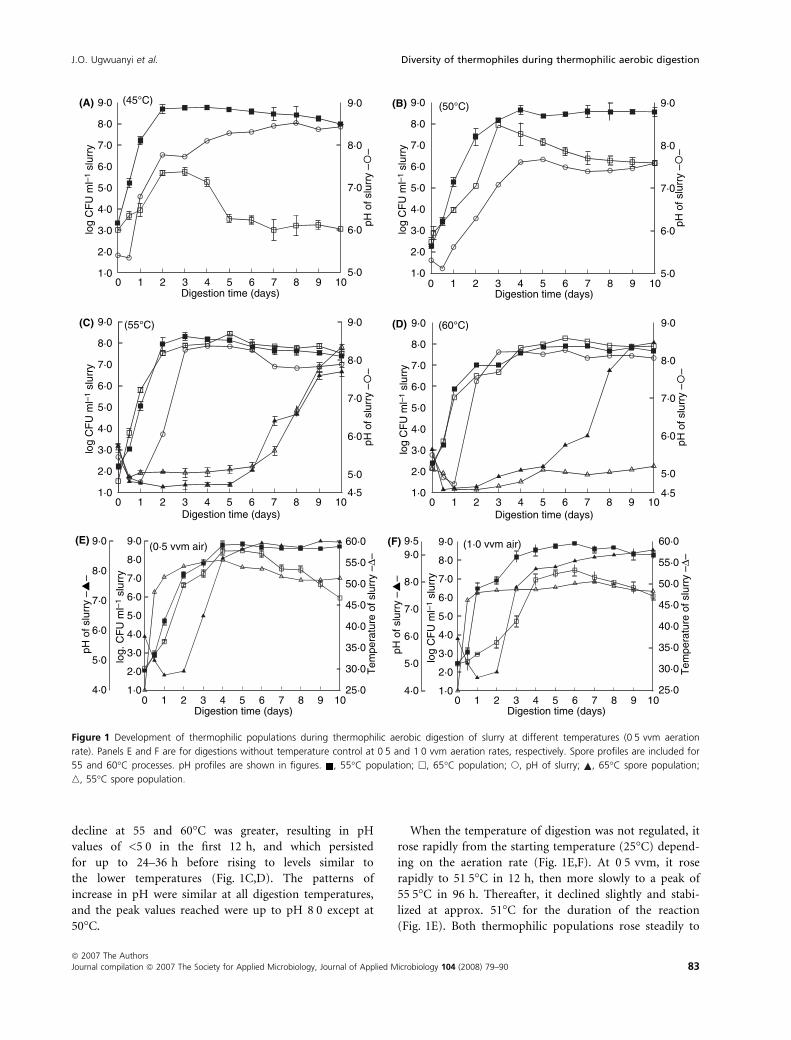
decline at 55 and 60�C was greater, resulting in pH
values of <5Æ0 in the first 12 h, and which persisted
for up to 24–36 h before rising to levels similar to
the lower temperatures (Fig. 1C,D). The patterns of
increase in pH were similar at all digestion temperatures,
and the peak values reached were up to pH 8Æ0 except at
50�C.
When the temperature of digestion was not regulated, it
rose rapidly from the starting temperature (25�C) depend-
ing on the aeration rate (Fig. 1E,F). At 0Æ5 vvm, it rose
rapidly to 51Æ5�C in 12 h, then more slowly to a peak of
55Æ5�C in 96 h. Thereafter, it declined slightly and stabi-
lized at approx. 51�C for the duration of the reaction
(Fig. 1E). Both thermophilic populations rose steadily to
1·0 1·0
1·0
1·0
1·0
0
(55°C) (60°C)
(50°C)
(0·5 vvm air) (1·0 vvm air)
(45°C)lo
g C
FU
ml–
1 sl
urry
log.
CF
U m
l–1
slur
ry
log
CF
U m
l–1
slur
ry
log
CF
U m
l–1
slur
ry
log
CF
U m
l–1
slur
ry
log
CF
U m
l–1
slur
ry
0
0
25·0
35·0
45·0
55·0
30·0
40·0
50·0
60·0
25·0
35·0
45·0
55·0
30·0
40·0
50·0
60·0
0
0
(A)
(C)
(E) (F)
(D)
(B)
Digestion time (days)
Digestion time (days) Digestion time (days)
Digestion time (days)
Digestion time (days)Digestion time (days)0
pH o
f slu
rry
–�–
pH o
f slu
rry
–�–
1 1
1
1
1
1
2 2
2
2
Tem
pera
ture
of s
lurr
y –Δ
–
Tem
pera
ture
of s
lurr
y –Δ
–
2
2
3 3
3
3
3
3
4 4
44
4
5 5
55
5
6 6
66
6
7 7
77
7
8 8
88
8
9 9
9
pH o
f slu
rry
–�–
pH o
f slu
rry
–�–
pH o
f slu
rry
–�–
pH o
f slu
rry
–�–
9
9
10 10
10
4 5 6 7 8 9 10
5·0 5·0
5·0
4·5 4·5
6·0 6·0
6·0
7·0 7·0
7·0
8·0 8·0
8·0
9·0 9·0
9·0
5·0
4·0
6·0
7·0
8·0
9·09·5
10
10
2·0 2·0
2·0
2·0
2·0
3·0 3·0
3·0
3·0
3·0
4·0 4·0
4·0
4·0
4·0
4·0
5·0 5·0
5·0
5·0
5·0
5·0
5·0
6·0 6·0
6·0
6·0
6·0
6·0
6·0
7·0 7·0
7·0
7·0
7·0
7·0
7·0
8·0 8·0
8·0
8·0
8·0
8·08·0
9·0 9·0
9·0
1·0
2·0
3·0
4·0
5·0
6·0
7·0
8·0
9·0 9·09·0
9·09·0
Figure 1 Development of thermophilic populations during thermophilic aerobic digestion of slurry at different temperatures (0Æ5 vvm aeration
rate). Panels E and F are for digestions without temperature control at 0Æ5 and 1Æ0 vvm aeration rates, respectively. Spore profiles are included for
55 and 60�C processes. pH profiles are shown in figures. , 55�C population; h, 65�C population; s, pH of slurry; , 65�C spore population;
n, 55�C spore population.
J.O. Ugwuanyi et al. Diversity of thermophiles during thermophilic aerobic digestion
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 83

peaks in 96 h during digestion at 0Æ5 vvm (Fig. 1E). During
digestion at 1Æ0 vvm, the temperature of the slurry rose to
48�C in 12 h and thereafter stabilized at approx. 49�C for
the duration of the digestion (Fig. 1F). At this aeration rate,
increase in thermophilic population was slower and peak
populations for both temperature groups were reached
after 6 days (Fig. 1F). During the unregulated reactions,
the contribution of obligate thermophiles to total popula-
tion was low at temperatures below 50�C (as at 1Æ0 vvm),
but was high when the temperature rose to 55�C (as at
0Æ5 vvm, during the first 96 h).
Effect of digestion pH on development of thermophiles
Thermophilic populations developed rapidly during diges-
tion at pH 6Æ0–9Æ0 (55�C), and peak populations were
reached in 12 h (earliest sampling time) to 24 h at pH
7Æ0–9Æ0 (Fig. 2B–D). This is significantly faster than at
comparable temperatures when the pH was not regulated
(see Fig. 1). At pH 6Æ0, 9Æ5 and in the unregulated
reaction, thermophilic populations increased more slowly
to reach peaks in 36 h or longer. Patterns of population
increase for both temperature groups were comparable
and the populations were stable in all the reactions. Dur-
ing digestion at pH 7Æ0–9Æ0 and in the unregulated reac-
tion obligate thermophiles accounted for up to 98% or
more of the total count, and in the unregulated reaction
declined only slightly after 60 h of digestion (Fig. 2F).
During digestion at pH 6Æ0, the obligate thermophiles
accounted for 85% of total count after 24 h and remained
so to the end of the reaction. A more pronounced drop
in obligate thermophiles contribution to total count was
obtained during digestion at pH 9Æ5, particularly after the
first 60 h of digestion.
The onset of reaction was followed by decline in the
spore population for both the total and obligate thermo-
philes to under 100 CFU ml)1 of slurry. The spore counts
remained low for up to 84 and 96 h for the total and obli-
gate populations, respectively, before increasing rapidly to
counts only slightly lower than the corresponding total
viable count (Fig. 2B). Endospore count followed the
development of respiration in the slurry. Periods when
spore counts were low corresponded to the periods of
most active metabolism, while increase in the spore count
coincided with rise in the DO (Ugwuanyi et al. 2005a,b).
Significantly, even during period of active metabolism,
considerable spore population remained in slurry.
Effect of aeration rate on the development
of thermophiles
Thermophiles developed rapidly at all the aeration rates
(Fig. 3). However, slightly higher total populations were
obtained at 0Æ25 and 0Æ5 vvm than at 0Æ1 and 1Æ0 vvm
(Fig. 3) and these were not significantly different
(P > 0Æ05). Obligate thermophiles developed significantly
less rapidly at 0Æ1 vvm than at the other aeration rates,
unlike the total populations that developed equally rap-
idly at all aeration rates. They were also less stable at the
lower aeration rates (0Æ1 and 0Æ25 vvm) than the total
thermophiles and declined slightly after peak populations
(Fig. 3A,B). During digestion at the higher aeration rates
(0Æ5 and 1Æ0 vvm), obligate thermophiles accounted for
more than 97% of the total population, but <85% of
total population at 0Æ1 and 0Æ25 vvm.
The patterns of decline and rise in spore population
following onset of digestion were similar at all the aera-
tion rates (Fig. 3). At 1Æ0 vvm, sporulation was rapid in
both populations, starting after 36 and 48 h, respectively,
for the obligate and total populations, and by 120 h
nearly all the thermophiles were present as spores. As the
aeration rate declined to 0Æ5 vvm, the onset of sporulation
was delayed to 84 and 96 h, respectively, for the total and
obligate populations. With further decrease in aeration
rate, the onset of sporulation was delayed further and
during digestion at 0Æ1 vvm only a limited proportion of
the populations had sporulated even at time the digestion
was terminated. As metabolic rate had slowed consider-
ably at this time (Ugwuanyi et al. 2005a), the failure of
thermophiles to sporulate extensively may be related to
oxygen deficiency.
Identity of predominant thermophiles
In the course of the digestion processes, over 220 thermo-
philes, culturally and morphologically distinct on primary
isolation, were selected from both temperature groups.
Colonies were isolated to represent different morphologi-
cal types. Passage through different temperatures was used
to separate the isolates into those capable of growth at
55�C but not 65�C and those capable of growing at 65�C.
All isolates capable of growth at 65�C were included
among the obligate thermophiles. On subculture, most of
the isolates within the 65�C group were found to resemble
each other. Following preliminary tests, 12 isolates,
selected to represent the different cultural and microscopic
forms, were retained for definitive identification. These
were separated into four isolates capable of growth at
65�C and eight isolates that grew at 55�C but not 65�C.
All the isolates obtained at 65�C formed terminal, ellip-
soidal spores some of which distended the sporangium.
Considerable variations were noted in the length of the
vegetative cells and in the staining behaviour of cells par-
ticularly during primary isolation. Some of the 65�C iso-
lates lost the ability to sporulate, or sporulated poorly in
nutrient media and often required extended incubation
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
84 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors

(up to 96 h) in sporulation medium to be able to sporu-
late (if they did at all). Within the 55�C populations, var-
iation existed in the location, size and shape of spore,
and size of the vegetative cells both on primary isolation
and on subculture. For both temperature groups, the
microscopic morphology was the more consistent feature
9(A) (B)
8
7
6
5
4
3
log
CF
U m
l–1 s
lurr
y
2
10 12 24 36 48 60 72
Digestion time (h)
(pH 6·0 digestion)(pH 7·0 digestion)
84 96 108 120 132 144 156
9
8
7
6
5
4
3
log
CF
U m
l–1 s
lurr
y
2
1
00 12 24 36 48 60 72
Digestion time (h)
84 96 108 120 132 144 156
9(C) (D)
8
7
6
5
4
3
log
CF
U m
l–1 s
lurr
y
2
10 12 24 36 48 60 72
Digestion time (h)
(pH 8·0 digestion)(pH 9·0 digestion)
84 96 108 120 132 144 156
9
8
7
6
5
4
3
log
CF
U m
l–1 s
lurr
y
2
10 12 24 36 48 60 72
Digestion time (h)
84 96 108 120 132 144 156
9(E) (F)
8
7
6
5
4
3
log
CF
U m
l–1 s
lurr
y
2
10 12 24 36 48 60 72
Digestion time (h)
(pH 9·5 digestion) (Unregulated pH digestion)
84 96 108 120 132 144 156
9
8
10·0
9·0
8·0
7·0
Dig
estio
n pH
6·0
5·0
4·0
7
6
5
4
3log
CF
U m
l–1 s
lurr
y
2
10 12 24 36 48 60 72
Digestion time (h)
84 96 108120132144156
Figure 2 Effect of digestion pH on the development of thermophilic populations during thermophilic aerobic digestion of slurry at 55�C and
0Æ5 vvm aeration rate. Spore counts for both populations were determined at pH 7Æ0 and are shown in (B). Profile of pH in control reaction is
shown in panel F. s, 55�C population; d, 65�C population; h, 55�C spore population; , 65�C spore population.
J.O. Ugwuanyi et al. Diversity of thermophiles during thermophilic aerobic digestion
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 85

in primary isolation and in subculture. The biochemical
profile based on the use of API 50CHB for members of
the genus Bacillus is shown in Table 1. All the 65�C iso-
lates were identified as B. steraothermophilus while the
55�C isolates were identified as either B. coagulans or B.
licheniformis.
Discussion
The development and diversity of thermophilic popula-
tions during TAD were strongly influenced by the
digestion conditions, particularly the reaction temperature
(see Fig. 1). Burt et al. (1990) reported that obligate
thermophiles do not develop until the temperature of
digestion exceeds 54�C. Similar development is noted in
this study, as the 65�C thermophiles developed poorly
and fluctuated during digestion at 45 and 50�C but
developed rapidly and were stable during digestion at 55
and 60�C. At the higher end, the aeration rate does not
appear to be the principal component in the control of
population relations as counts did not vary significantly
between 0Æ5 and 1Æ0 vvm rates when the temperature was
fixed at 55�C (see Fig. 3C,D). However, it could become
important at the lower aeration rates (0Æ1 and 0Æ25 vvm;
see Fig. 3A,B).
The effect of digestion pH on the development of ther-
mophiles was not as pronounced as that of temperature.
However, obligate thermophiles developed slowly,
declined rapidly, and the peak population obtained was
low at the pH extremes compared with the more neutral
reactions (see Fig. 2). Given this sensitivity of the obligate
thermophiles to extremes of pH, it appears that the drop
in the total population enumerated at pH 9Æ5 was a
reflection of the decline in the number of this subset, and
this may be due to their tendency to fail to sporulate and
so die out after the onset of stationary phase, as reported
by Sonnleitner and Fiechter (1983a). The behaviour of
obligate thermophiles in relation to digestion pH sup-
(0·1 vvm digestion) (0·25 vvm digestion)
(1·0 vvm digestion)(0·5 vvm digestion)
9·0
10·0(A) (B)
(C) (D)
8·0
7·0
6·0
5·0
4·0
3·0
2·0
1·0
00 12
Digestion time (h)
Digestion time (h)
Digestion time (h)
Digestion time (h)
24 36 48 60 72 84 96 108 120 132 144 156 0 12 24 36 48 60 72 84 96 108 120 132 144 156
0 12 24 36 48 60 72 84 96 108 120 132 144 1560 12 24 36 48 60 72 84 96 108 120 132 144 156
log
CF
U m
l–1 s
lurr
y
log
CF
U m
l–1 s
lurr
y
log
CF
U m
l–1 s
lurr
y
log
CF
U m
l–1 s
lurr
y
10·0
9·0
8·0
7·0
6·0
5·0
4·0
3·0
2·0
1·0
0
10·0
9·0
8·0
7·0
6·0
5·0
4·0
3·0
2·0
1·0
0
10·0
9·0
8·0
7·0
6·0
5·0
4·0
3·0
2·0
1·0
0
Figure 3 Effect of aeration rate on the development of thermophilic populations during thermophilic aerobic digestion of slurry at 55�C and pH
7Æ0. Spore profiles are also shown. s, 55�C population; d, 65�C population; h, 55�C spore population; , 65�C spore population.
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
86 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors

Table 1 Result of biochemical test for the identification of members of the genus Bacillus based on API 50CH B
Isolate Glycerol Erythritol D-Arabinose L-Arabinose Ribose D-Xylose L-Xylose Adonitol B-Methyl xyl Galactose D-Glucose D-Fructose
1V7 + ) ) + + + ) + ) + + +
1V8 + ) ) + + + ) + ) + + +
5V6 + ) ) + + + ) + ) + + +
P70 + ) ) + + + ) + ) + + +
1V3 + ) ) + + + ) ) ) + + +
2V4 + ) ) + + + ) ) ) + + +
2V5 + ) ) + + + ) ) ) + + +
5V2 + ) ) + + + - ) ) + + +
R5A + ) + + + + ) + ) + + +
R5E + ) + + + + ) + ) + + +
P63 + ) + + + + ) + ) + + +
P91 + ) + + + + ) + ) + + +
Isolate D-Mannose L-Sorbose Rhamnose Dulcitol Inositol Mannitol Sorbitol
a-methyl-
D-man
a-methyl-
D-gluco
Nacetyl
glucose Amygdalin Arbutin Aesculin
1V7 + + + ) + + + ) + + + + +
1V8 + + + ) + + + ) + + + + +
5V6 + + + ) + + + ) + + + + +
P70 + + + ) + + + ) + + + + +
1V3 + + + ) + + + ) + + + + +
2V4 + + + ) + + + ) + + + + +
2V5 + + + ) + + + ) + + + + +
5V2 + + + ) + + + ) + + + + +
R5A + + + ) + + + ) + + + + +
R5E + + + ) + + + ) + + + + +
P63 + + + ) + + + ) + + + + +
P91 + + + ) + + + ) + + + + +
Isolate Salicin Cellobiose Maltose Lactose Melibiose Sucrose Trehalose Inulin Melezitose D-Raffinose Starch Glycogen Xylitol
1V7 + + + + ) + + ) + + + + +
1V8 + + + + ) + + ) + + + + +
5V6 + + + + ) + + ) + + + + +
P70 + + + + ) + + ) + + + + +
1V3 + + + + + + + + ) + + + )2V4 + + + + + + + + ) + + + )2V5 + + + + + + + + ) + + + )5V2 + + + + + + + + ) + + + )R5A + + + + + + + ) ) + + + )R5E + + + + + + + ) ) + + + )P63 + + + + + + + ) ) + + + )P91 + + + + + + + ) ) + + + )
Isolate B-Gentiobiose D-Turanose D-Lyxose D-Tagatose D-fucose L-Fucose D-Arabitol L-Arabitol Gluconate 2Ketogluconate 5Ketogluconate
Identity of
Isolate
1V7 + + ) + ) + + ) ) ) + B.s
1V8 + + ) + ) + + ) ) ) + B.s
5V6 + + ) + ) + + ) ) ) + B.s
P70 + + ) + ) + + ) ) ) + B.s
1V3 + + ) + ) ) ) ) + ) ) B.l
2V4 + + ) + ) ) ) ) + ) ) B.l
2V5 + + ) + ) ) ) ) + ) ) B.l
5V2 + + ) + ) ) ) ) + ) ) B.l
R5A + + ) + ) ) + ) + + + B.c
R5E + + ) + ) ) + ) + + + B.c
P63 + + ) + ) ) + ) + + + B.c
P91 + + ) + ) ) + ) + + + B.c
Note: B. s is Bacillus stearothermophilus; B. l is B. licheniformis; B. c is Bacillus coagulans. Results were interpreted by reference to the table of Berkeley et al 31
and the APILAB software.
J.O. Ugwuanyi et al. Diversity of thermophiles during thermophilic aerobic digestion
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 87

ports earlier reports that highly thermophilic digestions
employing these organisms progress efficiently only at
neutral to slightly alkaline pH (Kosseva et al. (2001);
Ugwuanyi et al. 2005a). Although there is as yet no pub-
lished report of controlled pH digestion, Sonnleitner and
Fiechter (1983a) reported that TAD thermophiles are
neutrophilic in growth characteristics. Also Beaudet et al.
(1990) and Kosseva et al. (2001) reported that waste deg-
radation and population stability declined when the pH
of digesting slurry increased beyond neutral in a continu-
ous process, even though individual isolates were able to
grow up to pH 9Æ5. The enhanced stability of obligate
thermophiles obtained during the unregulated pH reac-
tion may be due to gradual adaptation of the populations
to the gradually increasing pH.
Obligate thermophiles developed poorly during diges-
tion under oxygen limitations such as obtained at
0Æ1 vvm in this study. It may be assumed that, during
digestion under conditions that select for obligate ther-
mophiles, a low aeration rate may result in poor process
performance (particularly in a batch process). This has
indeed been shown to be the case (Ugwuanyi et al.
2005b). As B. stearothermophilus (which predominated at
high temperatures in this study) does not grow in the
absence of oxygen (Sneath 1984), it is possible that they
were metabolically disadvantaged by the reduced avail-
ability of oxygen at 0Æ1 vvm. In summary, unless other
process requirements demand it, the high cost of strong
aeration may be avoided without compromising thermo-
philic activity by operating the process in median of air
supply. In fact, whereas comparable xylanase activities are
maintained at 0Æ5 and 1Æ0 vvm, protease activity at
0Æ5 vvm was over 20 times higher than at 1Æ0 vvm during
TAD of similar slurries (Ugwuanyi et al. 2004).
From the result of this study, microbial succession will
take place in this process as digestion temperature
increases, with obligate thermophiles replacing the facul-
tative ones. As nonmetabolizing populations may be pres-
ent as viable spores, the total viable count may not be
relied upon as an index of active population unless steps
are taken, as in this study, to distinguish between active
vegetative and spore forms. Most of the thermophiles
enumerated in the last few hours of the reactions were
present as spores (see Fig. 3). This may be a stress
response to exhaustion of nutrients. Simplicity of assay
and the clear profile obtained with the spore count sug-
gest that it may be a revealing parameter for monitoring
the development of TAD.
To date, there are few studies on the microbiology of
TAD, because early and continuing research on the pro-
cess has been dominated by the engineering aspects of
process optimization (Vismara 1985). However, Sonnleit-
ner and Fiechter (1983a,b, 1985) reported that over 95%
of the populations associated with the TAD of sludge
were B. stearothermophilus, the balance not being so des-
ignated only because they could not sporulate. Malladi
and Ingham (1993) also identified thermophilic Bacillus
spp. as the predominant populations during the TAD of
potato slop. Although all the thermophiles identified in
the present study belong to the genus Bacillus, they were
not all extremely thermophilic B. stearothermophilus. On
the contrary when the digestion temperature declined to
50�C and below the combination of B. coagulans and
B. licheniformis predominated, comparable to the work of
Burt et al. (1990) with swine slurry, while B. stearother-
mophilus predominated only above 55�C. The use of wide
temperature ranges in the digestion process would have
contributed significantly to the variety of thermophiles
identified in this study. It is clear that when TAD is oper-
ated up to 60�C the majority of the active populations
are members of the B. stearothermophilus group, although
a variety of other species may be isolated, that only sur-
vive in the process by virtue of their ability to sporulate
(Grueninger et al. 1984; Bomio et al. 1989).
A major difficulty in comparing data from microbio-
logical studies of TAD arises from differences in waste
types and operational conditions used. However, it is seen
that a mixture of thermotolerant and thermophilic organ-
isms carry out TAD, with selection and succession
responding to local environments. The principal selection
pressure is the reaction temperature, rather than pH or
aeration rate although these also play a role in affecting
the biochemical potential of populations favoured from
among a variety capable of growing at a given tempera-
ture.
The isolation and identification protocol employed in
this study may have reduced the variety of species
encountered in this study with the possibility of omitting
some populations from particular treatment process.
However, such participating organisms need to be consis-
tent in the processes to be of significance in TAD, as
distinct from participation in a single reaction. It is con-
sidered that the omission of such organisms, if such
occurred, does not imperil the definition of the popula-
tions participating in TAD. (In any case, it is unlikely that
other than routine media and methods will be of any real
use in full scale waste treatment processes). Difficulties
associated with inconsistencies in colonial morphology of
isolates on primary isolation, and rapid changes in cul-
tural morphology of specific temperature groups within
the course of a single digestion constrained attempts to
count the different biochemical groups in the process
hence the recourse to temperature groups. It was also dif-
ficult, for the same reason to construct a profile of suc-
cession on the basis of specific species, particularly
between B. coagulans and B. licheniformis, while only one
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
88 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors

species, B. stearothermophilus, was responsible for activity
because of obligate thermophiles. Participation is likely to
remain most relevant at the level of individual tempera-
ture groups adapted to particular waste type.
In conclusion, a mixed population of facultative and
obligately thermophilic Bacillus spp. drive TAD of potato
slurry. Temperature of digestion is a principal selection
factor, and the starting indigenous mixed populations are
gradually replaced by a near uniform obligately thermo-
philic B. stearothermophilus as the temperature of diges-
tion increases to 55�C and above. The roles of digestion
pH and aeration rate are less pronounced in population
selection. Digesting populations are neutrophilic to
slightly alkaline in their pH preference, and developed
better at the median aeration rates of 0Æ25 and 0Æ5 vvm
than at the extremes of 0Æ1 or 1Æ0 vvm. It is unlikely that
in any waste process a homogeneous population will
carry out waste stabilization, and the variety may also be
influenced by the nature of the waste. However, if the
aim of the process is to use treated waste in animal feed,
there may be need for microbiological control on the
basis of temperature-dependent selection as obtained at
60�C, (besides the need for pasteurization and protein
enrichment, both of which benefit from temperature ele-
vation (Ugwuanyi et al. 1999, 2006).
Acknowledgement
The authors wish to thank the British Commonwealth
Commission for support in carrying out this work.
References.
Beaudet, R., Gagnon, C., Bisaillon, J.C. and Ishaque, M. (1990)
Microbiological aspects of aerobic thermophilic treatment
of swine waste. Applied and Environmental Microbiol 56,
971–976.
Berkeley, R.C.W., Logan, N.A., Shute, L.A. and Capey, A.G.
(1984) Identification of Bacillus spp. Methods Microbiol 16,
291–328.
Bomio, M., Sonnleitner, B. and Feichter, A. (1989) Growth and
biocatalytic activities of aerobic thermophilic populations
in sewage sludge. Appl Microbiol Biotechnol 32, 356–362.
Brock, T.D. (1986) Introduction: an overview of the thermo-
philes. In Thermophiles: General Molecular and Applied
Microbiology ed. Brock, T.D. pp. 1–16 New York: John
Wiley and Sons.
Burt, P., Morgan, S.F., Dancer, B.N. and Fry, J.C. (1990)
Microbial population and sludge characteristics in thermo-
philic aerobic sewage sludge digestion. Appl Microbiol Bio-
technol 33, 725–730.
Chu, A., Mavinic, D.S., Kelly, H.G. and Guarnaschelli, C.
(1997) The influence of aeration and solids retention time
on volatile fatty acid accumulation in thermophilic aerobic
digestion of sludge. Environ Technol 18, 731–738.
Cibis, E, Kent, C.A., Krzywonos, M, Garncarek, Z., Garncarek,
B. and Miskiewicz, T. (2002) Biodegradation of potato
slops from rural distillery by thermophilic aerobic bacteria.
Bioresour Technol 85, 57–61.
Cibis, E., Krzywonos, M. and Miskiewicz, T. (2006) Aerobic
biodegradation of potato slops under moderate thermo-
philic conditions: effect of pollution load. Bioresour Tech-
nol 97, 679–685.
Collins, C.H., Lyne, P.M. and Grange, J.M. (1989) Microbiolog-
ical Methods, 6th edn. Butherworth and Co., London.
Couillard, D. and Zhu, S. (1990) Thermophilic aerobic process
for the treatment of slaughterhouse effluents with protein
recovery. Environ Pollut 79, 121–126.
Council of the European Union. (2001) Proposal for a regula-
tion of the European Parliament and of the Council laying
down health rules concerning animal by-products not
intended for human consumption. 8880 ⁄ 1 ⁄ 01. Agrileg 107
CODES 425.
EPA (1990) Autothermal Thermophilic Aerobic Digestion of
Municipal Wastewater Sludge. EPA ⁄ 625 ⁄ 10-90 ⁄ 007
September, Washington, DC: Office of Research and
Development, US Environmental Protection Agency.
EPA (1992) Control of Pathogens and Vector Attraction in Sew-
age Sludge. EPA ⁄ 625 ⁄ R-92 ⁄ 013 December, Washington,
DC: Office of Research and Development, US Environ-
mental Protection Agency.
Ginnivan, M.J., Woods, J.L. and O’Callaghan, J.R. (1981) The
thermophilic aerobic treatment of pig slurry. J Agric Eng
Res 26, 455–466.
Grueninger, H., Sonnleitner, B. and Fiechter, A. (1984) Bacterial
diversity in thermophilic aerobic sewage sludge, iii. A source
of organisms producing heat stable industrially useful enzy-
mes, e.g. a-amylase. Appl Microbiol Biotechnol 19, 414–421.
Hawash, S., EL Iibiari, N., Aly, F.H., EL Diwani, G. and
Hamad, M.A. (1994) Kinetic study of thermophilic aerobic
stabilization of sludge. Biomass Bioenergy 6, 283–286.
Heinonen-Tanski, H., Kiuru, T., Ruuskanen, J., Korhonen, K.,
Koivunen, J. and Ruokojarvi, A. (2005) Thermophilic aera-
tion of cattle slurry with whey and or jam wastes. Biore-
sour Technol 96, 247–252.
Kosseva, M.R., Kent, C.A. and Lloyd, D.R. (2001) Thermo-
philic bioremediation of whey: effect of physico-chemical
parameters on the efficiency of the process. Biotechnol Lett
23, 1675–1679.
Krahe, M., Antranikian, G. and Markl, H. (1996) Fermentation
of extremophilic microorganisms. FEMS Microbiol Rev 18,
271–285.
LaPara, T.M. and Alleman, J.E. (1999) Thermophilic aerobic
biological wastewater treatment. Water Research 33, 895–
908.
Loll, U. (1976) Purification of concentrated organic waste
waters from the foodstuffs industry by means of aerobic
J.O. Ugwuanyi et al. Diversity of thermophiles during thermophilic aerobic digestion
ª 2007 The Authors
Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90 89

thermophilic degradation process. Prog Water Technol 8,
373–379.
Malladi, B. and Ingham, S.C. (1993) Thrmophilic aerobic
treatment of potato processing wastewater. World J Micro-
biol Biotechnol 9, 45–49.
Messenger, J.R. and Ekama, G.A. (1993) Evaluation of dual
digestion system: part 4 Simulation of the temperature
profile in the batch fed aerobic reactor. Water SA 19,
209–215.
Mohaibes, M. and Heinonen-Tanski, H. (2004) Aerobic ther-
mophilic treatment of farm slurry and food wastes. Biore-
sour Technol 95, 245–254.
Rowan, N.J.B. (1996) Studies on the growth survival and detec-
tion of potentially pathogenic Listeria and Bacillus species in
infant formulae. PhD Thesis, University of Strathclyde,
Glasgow.
Sneath, P.H.A. (1984) Endospore forming Gram Positive rods
and cocci. In Bergeys Manual of Systematic Bacteriology,
Vol. 2 ed. Holt, J.G. pp. 1104–1139. Baltimore: William
Wilkins Co.
Sonnleitner, B. and Fiechter, A. (1983a) Bacterial diversity in
thermophilic aerobic sewage sludge: I. Active biomass
and its fluctuations. Eur J Appl Microbiol Biotechnol 18,
47–51.
Sonnleitner, B. and Fiechter, A. (1983b) Bacterial diversity in
thermophilic aerobic sewage sludge II. Types of organisms
and their capacities. Eur J Appl Microbiol Biotechnol 18,
174–180.
Sonnleitner, B. and Fiechter, A. (1985) Microbial flora studies
in thermophilic aerobic sludge treatment. Conserv Recycl 8,
303–313.
Surucu, G.A., Engelbrecht, R.S. and Chian, E.S.K. (1975) Ther-
mophilic microbiological treatment of high strength waste-
waters with simultaneous recovery of single cell protein.
Biotechnol Bioeng 17, 1639–1662.
Tanner, R.S. (1997) Cultivation of bacteria and fungi. In Man-
ual of Environmental Microbiology eds Hurst, C.J., Knud-
sen, G.R., McInerney, M.J., Stetzenbach, L.D. and Walter,
M.V. pp. 52–60. Washington, DC: ASM Press.
Ugwuanyi, J.O., Harvey, L.M. and McNeil, B. (1999) Effect of
temperature, pH and Suspended solids content upon
pasteurization of model agricultural waste during
thermophilic aerobic digestion. J Appl Microbiol 87, 387–
395.
Ugwuanyi, J.O., Harvey, L.M. and McNeil, B. (2004) Protease
and xylanase activities and thermophilic populations as
potential process monitoring tools during thermophilic
aerobic digestion. J Chem Technol Biotechnol 79, 30–38.
Ugwuanyi, J.O., Harvey, L.M. and McNeil, B. (2005a) Effect of
digestion temperature and pH on treatment efficiency and
evolution of volatile fatty acids during thermophilic aero-
bic digestion of model high strength agricultural waste.
Bioresour Technol 96, 707–719.
Ugwuanyi, J.O., Harvey, L.M. and McNeil, B. (2005b) Effect of
aeration rate and waste load on evolution of volatile fatty
acids and waste stabilization during thermophilic aerobic
digestion of a model high strength agricultural waste. Bio-
resource Techniology 96, 721–730.
Ugwuanyi, J.O., Harvey, L.M. and McNeil, B. (2006) Applica-
tion of Thermophilic Aerobic Digestion in Protein Enrich-
ment of High Strength Agricultural Waste Slurry for
Animal Feed Supplementation. J Chem Technol Biotechnol
81, 1641–1651.
Vismara, R. (1985) A model for autothermic aerobic digestion;
Effects of scale depending on aeration efficiency and sludge
concentration. Water Res 19, 441–447.
Diversity of thermophiles during thermophilic aerobic digestion J.O. Ugwuanyi et al.
90 Journal compilation ª 2007 The Society for Applied Microbiology, Journal of Applied Microbiology 104 (2008) 79–90
ª 2007 The Authors





























