Embed Size (px)
Citation preview

Pharmacology & Toxicology 1987, 60, 29-36
Effects of Methyl Mercury on Testicular Functions in Macaca fascicularis Monkeys
Mostafa K. Mohamedl], Thomas M. Burbacher" and N. Karle Mottetl,*)
Departments of ')Pathology and 2)Environmental Health of the Schools of Medicine and Public Health and Community Medicine, University of Washington, Seattle, Washington 98 195, U.S.A.
(Received May 22, 1986; Accepted August 6, 1986)
Abstract: These studies were performed to investigate the effects of MeHg on testicular function in Macaca fasciculuris monkeys. In an in vivo study involving oral treatment of adult males Macuca fasciculuris monkeys with MeHg for 20 weeks, changes in spermatozoa1 production, motility and morphology and in serum testosterone were followed before, during and after treatment. MeHg treatment significantly decreased YO motile spermatozoa and scores for sperm speed and forward progression and increased % abnormal sperm tail forms, at sub-neurotoxic levels. The MeHg-induced increase in semen abnormalities was not accompanied by any significant changes in serum levels of testosterone. No consistent histological abnormalities were detected in testicular biopsies from the treated animals at the end of the treatment period. A good recovery pattern was observed for the MeHg effects on sperm motility while this was unclear for the effects on sperm morphology.
Hypospermia and teratospermia (increased abnormal sper- matozoa) have been reported in workers occupationally exposed to methyl mercury (MeHg) and ethyl mercury (Po- pescu 1978). However, we found no information available on the MeHg effects on male fertility in the human epidemic poisoning due to MeHg (Piotrowski & Inskip 1981). In laboratory rodent experiments, MeHg was reported to in- duce disturbances of rat spermatogenesis (Sakai 1972) and to pass the blood testis barrier of mice (Lee & Dixon 1975). In treated male rats, MeHg reduced the number of fertile matings and mean litter size (Khera 1973), and impaired testosterone secretion (Burton & Meikle 1980). In addition, MeHg has been shown, in vitro, to be a potent microtubule assembly inhibitor (Vogel et al. 1985). Since microtubules are the motor apparatus for the sperm, the developing sperm is a highly possible target for the effects of MeHg.
The production of motile spermatozoa in semen is a very complex testicular function that requires the interplay of the hypothalamus, Pituitary, testicles and several other glandular secretions (Mann & Mann 1981). Any adverse effects of MeHg on the functions of these organs would be reflected on semen quality and/or testosterone secretion. Because of uncontrolled dose, duration of exposure and psychological variables, it will be difficult to obtain reliable data on testicular functions on humans. The effects of sub- neurotoxic doses of MeHg on male reproductive functions in non-human primates have not been reported. Such infor- mation would be of critical importance for evaluation of risk to human male testicular function. We have previously reported on the in vitro dose effects of MeHg on sperm motility and swimming speed (Mohamed et al. 1986a) and on the effects of MeHg on sperm oxygen consumption using semen of the Macaca fascicularis monkey (Mohamed et al. 1986b). This study was designed to identify the reproductive effects of MeHg exposure at sub-neurotoxic levels in male Macaca.fascicularis monkeys. Our goal was to identify and
characterize the effects of MeHg exposure on spermatozoal production, motility, and morphology and on testosterone secretion.
Materials and Methods
Healthy adult male Mucacufascicularis monkeys (9) were randomly assigned to a control group or one of two treatment groups, low (50 pg MeHg/kg/day), and high (70 Fg MeHg/kg/day) dose groups (3 each). All animals were individually housed at the Regional Primate Research Center at the University of Washington, Seattle, Washington, with 12 hr light cycle and controlled temperature (21").
Study plan. The experimental plan includes a base line, treatment, and recovery periods. Base line characteristics of semen quality and serum testosterone were established during a period of 8-16 weeks. During the base line period, semen samples were collected weekly and blood samples for testosterone assays were collected biweekly. During the treatment period, MeHg was given orally, mixed with apple juice, via a syringe on a daily basis over a period of 20 weeks. Semen samples were collected weekly for assessment of sperm con- centration, motility, and morphology and for analysis of semen mercury. Blood samples were collected weekly for mercury analysis and biweekly for serum testosterone assays during this period. A challenge test to measure the maximum ability of the Leydig cells to respond to pituitary stimulation by luteinizing hormone relasing hormone (LHRH) (Sigma Chemical Co., St. Louis, MO, U.S.A.) was performed twice during both the base line and the treatment periods. At the end of the treatment period, an excision testicular biopsy was performed for histologic and electron microscopic exam- ination as well as for mercury assay. Semen and blood samples were collected on the same schedule as before and analyzed throughout the recovery period. After a recovery period of 20 weeks, animals were sacrificed for histologic examination and mercury assay of all the organs and electron microscopy examination of the testicles.
Examination of semen samples. Semen samples were collected weekly in a dry, clean glass beaker by anal electroejaculation (Roussel 62 Austin 1968). Sample volume could not be measured in most of the samples because of the very small volume, but sample weight was recorded. One ml of Biggers, Whiten, and Whittengham's (BWW) medium (Biggers et al. 1971), was added to each sample immediately
30 MOSTAFA K. MOHAMED ET AL.
after collection to provide enough volume for evaluation of all other parameters. Sperm concentration, percent of motile spermatozoa, and sperm speed and forward progression scores were determined by a Makler counting chamber (The Israel Electrooptical Industry Ltd., Rehovot, Israel). Motility evaluation was done at room tem- perature (21"). Air-dried smears on clean slides were stained using Bryan/Leisham stain for routine seminal fluid morphology. Trans- mission and scanning electron microscopy were used to examine the morphological abnormalities in semen samples.
Serum testosterone blood samples. Animals were lightly anaestheti- zed with ketamine HCI (10 mg/kg intramuscularly; Parke-Davis, Detroit, MI, U.S.A.), and blood samples were drawn from periph- eral veins by venipuncture. Because of the pulsatile nature of testos- terone hormone secretion (Steiner et al. 1980) five blood drawings were done on each animal during a two-hour period (9:OO-11:OO AM). Serum was separated and stored at - 70" until assayed. Serum testosterone was assayed by pooling the five serum samples as an average for the 2-hour period. For each LHRH challenge test a subcutaneous injection of LHRH (1 pg/kg body weight) was admin- istered to each animal after two blood drawings 10 min. apart. Blood samples were drawn 15, 30, 60 and 90 min. after the LHRH injection. All blood samples were analyzed to determine the maximum rise in serum testosterone. The challenge tests were run 6 weeks apart during the base line period and after 8 and 20 weeks from the start of treatment period. Serum testosterone was assayed using izsI radioimmunoassay test system (Liquisol, Damon Diagnos- tics, Needham Heights, MA, U.S.A.).
Mercury assay. Mercury was assayed in whole blood, semen and tissues by atomic absorption spectrophotometry (Mottet & Body 1974). Since the mercury was assayed as total mercury in all the samples we will refer to it as mercury (Hg).
Statistical analysis. The results of semen examination and serum testosterone analyses are presented as the mean of the samples collected for each animal during the base line, treatment and recov- ery periods. For each animal, comparisons were made between the means of each parameter during the base line period and both the
treatment and recovery periods separately using the Student's t-test. Group average was then calculated for the three animal means in each group. For each group, comparisons were made between the average of each parameter during the base line period and both the treatment and recovery periods using the paired Student's t-test. Across group comparisons of group averages were performed, dur- ing each period, using the Student's t-test. The frequency distri- bution of some parameters of interest were examined in a pool of the results of each group during each of the three periods. The level of significance was set as P < 0.05.
Results
None of the animals showed any significant changes in the general conditions, activity or weight during the treatment period. Observations for neurotoxic manifestations (Burba- cher et al. 1984), revealed no apparent change in sucking, biting, licking, visual follow or hand tremors at the end of the treatment period.
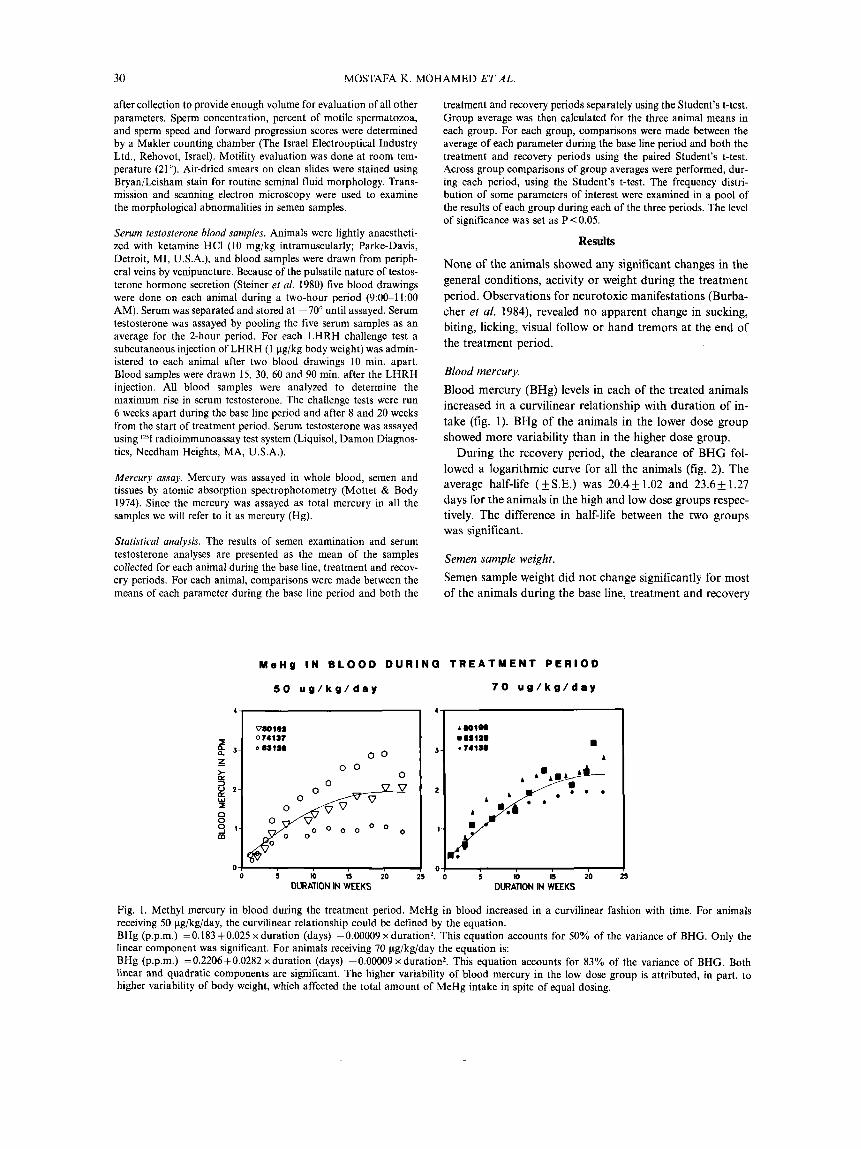
Blood mercury. Blood mercury (BHg) levels in each of the treated animals increased in a curvilinear relationship with duration of in- take (fig. 1). BHg of the animals in the lower dose group showed more variability than in the higher dose group.
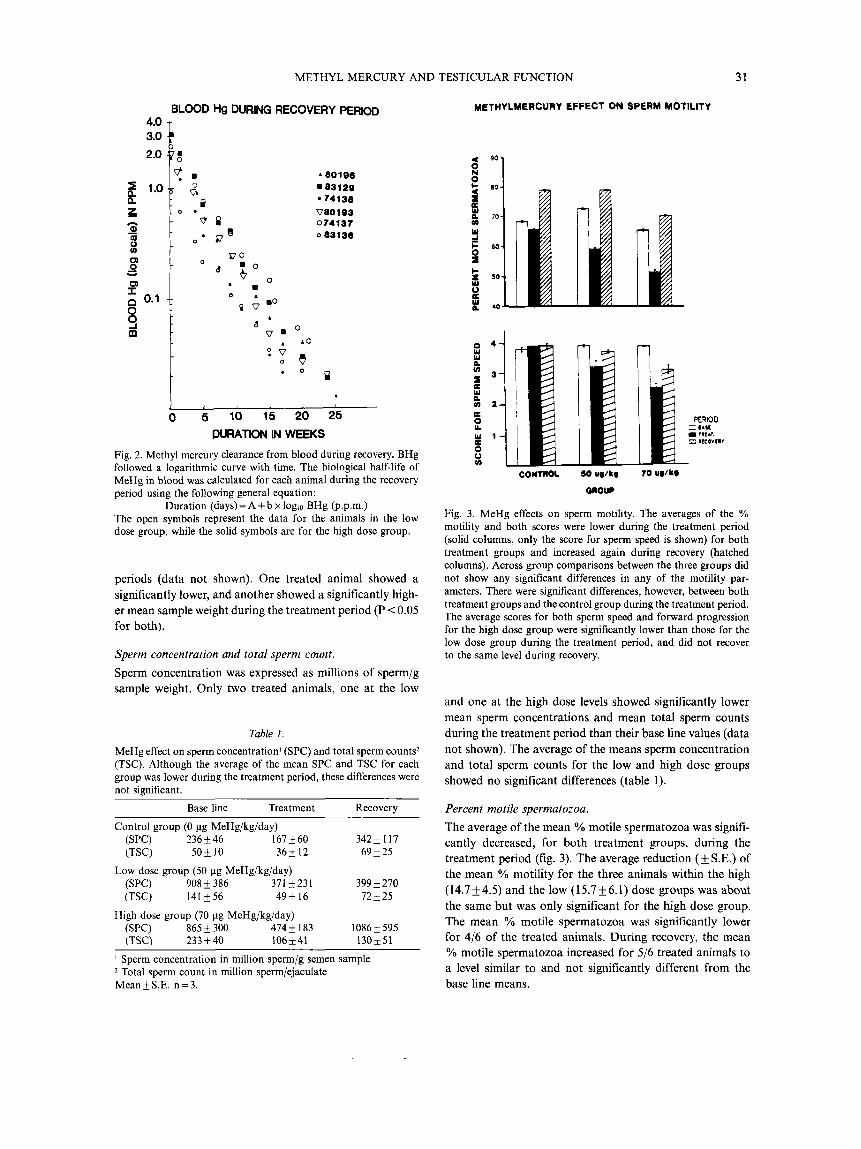
During the recovery period, the clearance of BHG fol- lowed a logarithmic curve for all the animals (fig. 2). The average half-life ( f S.E.) was 20.4 f 1.02 and 23.6 1.27 days for the animals in the high and low dose groups respec- tively. The difference in half-life between the two groups was significant.
Semen sample weight. Semen sample weight did not change significantly for most of the animals during the base line, treatment and recovery
M e H g I N B L O O D D U R I N G T R E A T M E N T P E R I O D
5 0 u g / k g / d a y
0 i m ;s 7 0 DURATION IN WEEKS
7 0 u g / k g / d a y
IS 20 5 m DURATION IN WEEKS
5
Fig. 1. Methyl mercury in blood during the treatment period. MeHg in blood increased in a curvilinear fashion with time. For animals receiving 50 pg/kg/day, the curvilinear relationship could be defined by the equation. BHg (p.p.m.) =0.183+0.025 x duration (days) -0.00009 x duration*. This equation accounts for 50% of the variance of BHG. Only the linear component was significant. For animals receiving 70 pg/kg/day the equation is: BHg (p.p.m.) =0.2206+0.0282 xduration (days) -0.00009 x duration2. This equation accounts for 83% of the variance of BHG. Both linear and quadratic components are significant. The higher variability of blood mercury in the low dose group is attributed, in part, to higher variability of body weight, which affected the total amount of MeHg intake in spite of equal dosing.
METHYL MERCURY AND TESTICULAR FUNCTION 31
BLOOD Hg DURING RECOVERY PERIOD 4.0
4 80196 8 83129
74138 080193 074137 o 83138
0 5 10 15 20 25 DURATION IN WEEKS
Fig. 2. Methyl mercury clearance from blood during recovery. BHg followed a logarithmic curve with time. The biological half-life of MeHg in blood was calculated for each animal during the recovery period using the following general equation:
Duration (days)=A+b x loglo BHg (p.p.m.) The open symbols represent the data for the animals in the low dose group, while the solid symbols are for the high dose group.
periods (data not shown). One treated animal showed a significantly lower, and another showed a significantly high- er mean sample weight during the treatment period (P < 0.05 for both).
Sperm concentration and total sperm count. Sperm concentration was expressed as millions of sperm/g sample weight. Only two treated animals, one at the low
Table 1 .
MeHg effect on sperm concentration' (SPC) and total sperm counts2 (TSC). Although the average of the mean SPC and TSC for each group was lower during the treatment period, these differences were not significant.
Base line Treatment Recovery
Control group (0 pg MeHg/kg/day) (SPC) 236+46 167+60 342k117 (TSC) 50+ 10 36k 12 69 k 25
(SPC) 908 k 386 371f231 399 270 (TSC) 141 & 56 49+ 16 72 & 25
(SPC) 865 f 300 474+ 183 1086 k 595 ( T W 233 f 40 106k41 130f51
Low dose group (50 pg MeHg/kg/day)
High dose group (70 pg MeHg/kg/day)
' Sperm concentration in million sperm/g semen sample
MeanfS.E. n = 3 . Total sperm count in million sperm/ejaculate
METHYLMERCURY EFFECT ON SPERM MOTILITY
OllOUC
Fig. 3. MeHg effects on sperm motility. The averages of the % motility and both scores were lower during the treatment period (solid columns, only the score for sperm speed is shown) for both treatment groups and increased again during recovery (hatched columns). Across group comparisons between the three groups did not show any significant differences in any of the motility par- ameters. There were significant differences, however, between both treatment groups and the control group during the treatment period. The average scores for both sperm speed and forward progression for the high dose group were significantly lower than those for the low dose group during the treatment period, and did not recover to the same level during recovery.
and one at the high dose levels showed significantly lower mean sperm concentrations and mean total sperm counts during the treatment period than their base line values (data not shown). The average of the means sperm concentration and total sperm counts for the low and high dose groups showed no significant differences (table 1).
Percent motile spermatozoa. The average of the mean YO motile spermatozoa was signifi- cantly decreased, for both treatment groups, during the treatment period (fig. 3). The average reduction (f S.E.) of the mean % motility for the three animals within the high (14.7f4.5) and the low (15.7f6.1) dose groups was about the same but was only significant for the high dose group. The mean % motile spermatozoa was significantly lower for 4/6 of the treated animals. During recovery, the mean % motile spermatozoa increased for 5/6 treated animals to a level similar to and not significantly different from the base line means.

32 MOSTAFA K. MOHAMED ET A L
Fig. 4. Scanning electron-microscopy of semen spermatozoa. The bent tail (B) have a smooth bend at the middle of the tail. The kinked sperm (K) tails have a sharp angulation at the area of the middle piece and its connection with the head. Coiled sperm tails (C) have many coils especially at the terminal end. A peculia conti- nuity of the membrane was observed in spermatozoa with coils and kinks.
Scores for sperm speed and forward progression. The averages of the mean sperm speed and forward pro- gression scores for the three animals in each group were significantly lower during the treatment period for both treatment groups (fig. 3). All the treated animals (6/6) show- ed significantly lower mean scores for both sperm speed and forward progression during the treatment period (data not shown). However the control animals did not show any significant changes in either mean scores. During recovery, the scores for both paramters were improved to a level similar to the base line level in 4/6 treated animals (data not shown). For the other two animals (both from the high dose group), the scores improved but to a level that was still significantly lower than the base line means.
Morphological examination of semen smears. Almost all of the abnormal defects seen in semen samples during the whole experiment were tail defects. Amorphous, small and large sperm heads were occasionally seen. Total abnormal tail defects were divided into three main catego- ries: Bent, kinded and coiled tail defects. A scanning and transmission electronmicrograph of examples of these ab- normalities is shown in fig. 4 and 5.
During the base line period, average '/o total abnormal tail forms for each group, were not significantly different from each other (Fig. 6). However, during the treatment period, both treatment groups showed significantly and similarly higher averages of the mean total abnormal tail defects (fig. 6). The main component of the increase in the total abnormal tail defects during the treatment period was in the bent and kinked tail categories. Four of the treated animals (2 at each dose group) had significantly higher means of YO total abnormal forms, while none of the control animals showed any significant increase (data not shown).
The frequency distribution of total abnormal, bent, and kinked tail forms were calculated from a pool of all the sample results within each group during base line, treatment and recovery periods. The categories selected were based on general frequency distribution with emphasis on the upper 90th percentile of each tail abnormality in the base line period for all the groups. Samples with more than 30% total abnormal, 15% bent and 8% kinked tail forms were present in less than 10% of base line samples of every group. The frequency distribution of samples with more than 30% total abnormal tail forms (fig. 7), 15% bent tail forms and 8% kinked tail forms were significantly higher during the treatment period than during the base line period for both treatment groups.
During recovery, one control animal showed a significant increase in the mean % total abnormal forms, although none of the controls showed any increase during the treat- ment period. This could be due to an effect of the excision biopsy at the end of the treatment period. Four treated animals continued to have a high mean of % total abnormal forms during recovery, while the other two showed a slightly lower mean or no significant change from the treatment level.
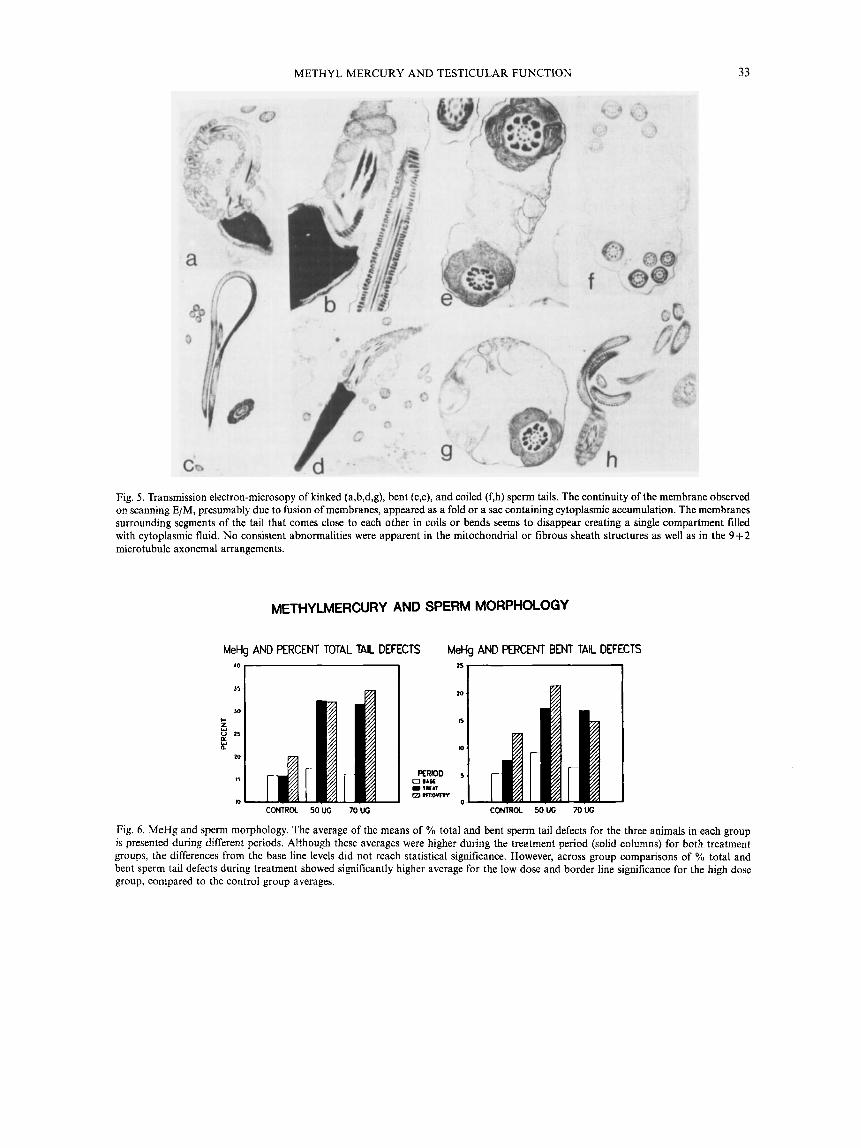
Scanning electron-microscopic examination of spermato- zoa revealed the details of the sperm tail abnormalities (fig. 4). Transmission E/M of the spermatozoa (fig. 5), showed cytoplasmic accumulation surrounding the axoneme in the bent tails or coiled tails and the site of the kinked tails. No consistent structural abnormality, in particular of the fibrous or the mitochondrial sheath and of the 9+2 micro- tubule arrangements were detected.
Histological and electron-microscopic features of the tes- ticular biopsy at the end of the treatment period and the autopsy after the recovery period, were consistent with the description by Dym & Cavicchia (1977) for the macaques. By light microscopy, no consistent differences were observed between the treated and the control animals. In particular, all the stages of spermatogenesis were observed, nearly all of the cells of the seminifrous tubules were normal with the occasional exception of degenerative and pyknotic cells (also observed in the controls). The basement membrane, the interstitial tissue, Sertoli cells and Leydig cells were all within the normal limits. Quantitation of different stages of spermatogenesis was not performed at this time. Trans- mission electron-microscopy of testicular biopsy of the treated animals did not show any significant alterations from the normal picture observed in the controls. We looked specifically for microtubule disturbances in the spermatid manchettes and axial filament arrangements, mitochondria1 swelling and Sertoli cell to spermatgeic cells borders.
Semen mercury. Semen mercury expressed as BWW-diluted mercury levels in p.p.m., pg mercury/g semen or as pg mercury/lO* sperma- tozoa were higher, in general, for the animals in the high dose group than in the low dose group (table 2). Mercury levels in semen, expressed as any of the above, did not have
METHYL MERCURY AND TESTICULAR FUNCTION 33
Fig, 5 . Transmission electron-microsopy of kinked (a,b,d,g), bent (c,e), and coiled (f,h) sperm tails. The continuity of the membrane observed on scanning E/M, presumably due to fusion of membranes, appeared as a fold or a sac containing cytoplasmic accumulation. The membranes surrounding segments of the tail that comes close to each other in coils or bends seems to disappear creating a single compartment filled with cytoplasmic fluid. No consistent abnormalities were apparent in the mitochondria1 or fibrous sheath structures as well as in the 9 + 2 microtubule axonemal arrangements.
METHYLMERCURY AND SPERM MORPHOLOGY
MeHg AND PERCENT TOTAL TAIL DEFECTS MeHg AND PERCENT BENT TAIL DEFECTS
J5 I JO
I-
15
e 20
13
10 CONTROL 50UC 70UG CONTROL SOW 70W
Fig. 6. MeHg and sperm morphology. The average of the means of % total and bent sperm tail defects for the three animals in each group is presented during different periods. Although these averages were higher during the treatment period (solid columns) for both treatment groups, the differences from the base line levels did not reach statistical significance. However, across group comparisons of YO total and bent sperm tail defects during treatment showed significantly higher average for the low dose and border line significance for the high dose group, compared to the control group averages.

34 MOSTAFA K. MOHAMED ET AL.
D~sTRIBUTION OF T O T A L SPERM TAIL DEFECTS
DURING EASE LINE PERIOD
DURING TREATMENT
Fig. 7. MeHg and frequency distribution of total sperm tail abnor- malities. The frequency distribution of samples with more than 30% total sperm tail defects (solid black) increased significantly for the two dose groups during treatment, while there was no significant changes in the control group. The frequencies of <20% total (blank), and 20-30% total (hatched) represented more than 90% of the samples in each of the groups during the base line period. During recovery, the frequency of total tail defects was not de- creased possibly requiring a longer time for recovery or delayed by adverse effects of testicular excision biopsy.
a clear relationship to duration of treatment, amount of mercury intake, or blood mercury for any of the treated animals. High levels of mercury alternated with low levels without any particular order.
Serum testosterone. The average serum testosterone for the animals in the two treatment groups were not significantly different from the average of the control group during the base line period. Serum testosterone levels did not change signifiantly for 5 / 6 of the treated and for 2/3 of the control animals (data not presented) during the treatment period. The results of the LHRH challenge test did not show any significant change
in the Leydig cell capacity to secrete testosterone for all the treated and the control animals during both the base line and treatment periods. Serum testosterone levels, 60 min. after LHRH injection was more than doubled for all the control and treated animals (data not shown).
Discussion
MeHg treatment at the two dose levels: 50 and 70 pg/kg/ day did not result in any apparent neurotoxic manifestations during the treatment period for any of the animals. How- ever, adverse effects were apparent on semen examination. All the animals showed lower % motile spermatozoa, lower scores for sperm speed and forward progression, and higher % of abnormal sperm tail forms especially bent and kinked tails. There were no significant differences between the ef- fects of MeHg treatment in the two dose groups.
Mercury levels in testicular biopsies at the end of the treatment period, were above 2 p.p.m. for 5/6 of the animals (table 2) and were positively correlated with the total amount of MeHg intake. Levels of MeHg above 2 p.p.m. were shown in vitro to induce a dose-related decrease of sperm motility and swimming speed (Mohamed et al. 1986a). The mechanism of in vitro MeHg effect on sperm motility was suggested to be primarily due to an interference of MeHg with the Dynein/microtubule sliding assembly (Mohamed et al. 1986b). Whether the mechanisn of action of MeHg observed in vivo is similar to that observed in vitro on the mature sperm or involve an adverse effect on the developing serum (or both) remains to be determined.
The increase in sperm tail abnormalities induced by MeHg treatment in our report confirms the earlier obser- vation by Popescu (1978), on teratospermia in occupational- ly MeHg-exposed workers. A similar sperm tail abnormality was induced in mice “spermatozoa folded on themselves” by vinblastine treatment (Wyrobek & Bruce 1975) and in monkeys by gossypol treatment (Shandilya et al. 1982). It is interesting to note that both vinblastine (Wilson el al. 1982; Luduena & Roach 1981), and MeHg (Vogel et al.
Table 2.
Mercury levels in semen and testicular biopsy during the treatment period. There was a high degree of varibility of semen mercury within the samples from the same animal and in between animals within the same group but the average for the low dose group was significantly lower than that for the high dose group.
BWW-diluted Semen @.p.m.)
Low dose group (50 pg/kg/day) #80193 0.048+0.006 #I4137 0.125k0.014 #83136 0.054k0.008
Average k S.E. 0.076+0.020
High dose group (70 pg/kg/day) #80196 0.391 *0.065 #83129 0.144_f0.030 #74138 0.067 k 0.009
Average+ S.E. 0.201 k 0.080
Semen Hg Pg/g
0.244k 0.031 0.749 k0.086 0.352k0.093
0.448k0.125
2.298k0.432 0.983k0.251 0.257 f 0.032
I . 179k0.488
Semen Hg pg/108sperm (testicular HE p.p.m.)
0.894 k0.017 (2.7) 0.298 k0.057 (2.1) 0.132 k0.035 (1.2) 0.441 k0.189
0.383 k0.059 (2.3) 0.342k0.092 (2.5) 0.999k0.218 (2.3)
0.575+0.213

METHYL MERCURY AND TESTICULAR FUNCTION 35
1985), inhibits microtubule assembly by interacting with sulfhydral groups on microtubules. Gossypol was also shown to disrupt the microtubule axoneme organization (Shandilya et ul. 1982). The mechanism of MeHg-induced sperm tail abnormalities could be an interference with the microtubule assembly during spermiogenesis, or an inter- ference with the dynein/microtubule sliding function in the mature sperm or both.
The effects of MeHg on sperm motility seems to be revers- ible during the recovery period (fig. 3). However, MeHg effects on sperm morphology did not show a similar recov- ery (fig. 6). An effect of the excision biopsy at the end of the treatment period may have caused an increase in abnormal sperm tails. This was suspected after detecting a significant increase of abnormal sperm tails in one of the control ani- mals during the recovery period. Thus it was difficult to interpret the recovery data of sperm morphology in our study. Lee & Dixon (1975), reported also a recovery of the decreased fertility of MeHg-treated males after cessation of treatment. Testicular effects may be a positive sign useful, in monitoring health effects in the workplace, for early detection of reversible adverse effects of MeHg.
We did not detect any decrease of testosterone secretion in the treated animals, even after pituitary stimulation by LHRH. This is in contrast to the report by Burton & Meikle (1980) in rats, where they detected a decrease in testosterone secretion after MeHg treatment. This could be due to a much higher dosage in their experiments (0.26 mg/60 g rat/ 48 hrs, >40 times our daily dose). Although we did not measure plasma LH levels during treatment and during LHRH challenge, a normal testosterone level and a normal increase after the challenge provide an evidence for normal pituitary LH release and normal Leydig cell function. Thus an indirect effect of MeHg on the developing sperm through an effect on the pituitary-testicular hormonal axis at this dose level seems to be unlikely.
Sperm concentration and total sperm counts were signifi- cantly reduced for only two treated animals. We expected a decrease in sperm production based on the known inhibitory effects of MeHg on cellular mitosis (Rozynkowa & Raczkie- wicz 1977; Miura et al. 1978; Prasad et al. 1979; Koerker 1980). We could not rule out such an effect in our exper- iments due to one or more of many uncontrolled variables. The first is related to the natural high degree of variability of sperm concentration (range < 1 million-4000 millions sperm/g semen), within and in between subjects. Secondly, is the uncontrolled mastubation behavour in monkeys that may affect abstinence time between sampling, and subse- quently, sperm concentration. Thirdly, is the small number of animals in each group due to high cost of niaintainance. This decreases the statistical power of detecting a significant difference in a parameter with such high degree of varia- bility. Examination of testicular biopsies did not show any apparent disruption of spermatogenesis, but further quanti- tation may reveal subtle differences at specific stages of the cycle.
The average biological half-life of mercury for the low
dose group (23.6+ 1.3) was significantly longer than the average for the high dose group (20.4+ 1.0). This may rep- resent a higher efficiency of execretion at the high dose level. Earlier reports on the Macaca mulatta (Finocchio et ul. 1980) showed a similar biological half-life of 27-34 days in the dose range of 50-90 pg/kg/day and 14 days at 100 pg/kg/day. A facilitated execretory mechanism that comes into play at a critical blood level was suggested to explain the shorter half life at higer dose levels.
The relationship between MeHg levels in semen and blood as well as the amount of MeHg intake to sperm motility and morphology was examined. A significant nega- tive correlation was found in all the animals between percent motile spermatozoa and one or more of the parameters of mercury levels in semen or blood. A lower percent motile spermatozoa was associated with higher blood mercury (R= -0.40 to -0.85) and semen mercury (R= -0.39 to -0.85). Semen mercury levels (table 2) did not correlate with the blood levels in the animals. That was probably due to the variable amounts of different secretions that contribute to semen volume. The final concentration in semen sample would vary independently from the level in the original secretion that contained mercury.
This is the first controlled study to report on testicular adverse effects of MeHg in non-human primates. MeHg treatment induced testicular toxicity at sub-neurotoxic levels which was significantly correlated with MeHg intake, blood, semen and testicular mercury levels. Percent motile sperma- tozoa, scores for sperm speed and forward progression were decreased and YO abnormal sperm tail forms were increased significantly during MeHg treatment. The MeHg-induced increase in semen abnormalities was not accompanied by any significant changes in serum levels of testosterone. As it is difficulat to predict the effects of these semen abnor- malities on fertility, mating studies involving male treatment with MeHg are necessary to answer this question.
References
Biggers, J. D., W. K. Whitten & D. F. Whittengham: The culture of mouse embryos in vitro. In: Methods of mammalian embry- ology. Ed.: C. J . Daniel. W. H. Freeman and Company, San Francisco. 1971, p. 101.
Burbacher, T. M.. C. Monett, G. S. Kimberly & N. K. Mottet: Methylmercury exposure and reproductive dysfunction in the nonhuman primate. Toxicol. Appl. Pharmacol. 1984, 75, 18-24.
Burton, G. V. & A. W. Meikle: Acute and chronic methylmercury poisoning impairs rat adrenal and testicular function. J. Toxicol. Environ. Health. 1980, 6, 597-606.
Clarkson, T. W.: Mercury poisoning. In: Clinical chemistry and chemical toxicology of metals. Ed.: S . S. Brown. Elsevier/North- Holland Biomedical Press, Amsterdam, 1976, pp. 189-200.
Dym, M. & J. C. Cavicchia: Further observation on the bloodtestis barrier in monkeys. Biol. Reprod. 1977, 17, 39W.03.
Finocchio, D. V., E. S. Luschei, N. K. Mottet & R. Body: Effects of methylmercury on the visual system of rhesus macaque (Macacrr mulatta). I. Pharmacokinetics of chronic methylmercury related to changes in vision and behavior. In: Neurotoxiciry of the visual system. Eds.: W. H. Merigan and B. Weiss. Raven Press, New York, 1980, pp. 113-122.

36 MOSTAFA K. MOHAMED ET AL.
Khera, K. S.: Reproductive capability of male rats and mice treated with methymercury. Toxicol. Appl. Pharmacol. 1973,24, 167-177.
Koerker, R. L.: The cytotoxicity of methylmercuric hydroxide and colchicine in cultured mouse neuroblastoma cells. Toxicol. Appl. Pharmacol. 1980, 53, 458469.
Lee, I. P. & R. L. Dixon: Effects of mercury on spermatogenesis studied by velocity sedimentation cell separation and serial ma- ting. J . Pharmucol. Exp. Therap. 1975, 193, 171-181.
Luduena, R. F. & M. C. Roach Contrasting effects of maytansine and vinblastine on the alkylation of tubuline sulfhydrals. Arch. Biochem. Biophys. 1981, 210(2), 498-504.
Mann, T. & C. L. Mann: Male reproductive function and semen. Springer-verlag, Berlin, Heidelberg, New York. 1981, pp. 1-37.
Miura, K., K. Suzuki & N. Imura: Effects of methylmercury on mitotic mouse glioma cells. Environ. Res. 1978, 17, 453-471.
Mohamed, M. K., W. I. Lee, N. K. Mottet & T. M. Burbacher: Laser light-scattering study of the toxic effects of methylmercury on sperm motility. J . Androl. 1986a, 7, 11-15.
Mohamed M. K., T. Evans, N. K. Mottet & T. M. Burbacher: Effects of methyl mercury on sperm oxygen consumption. Acfa pharmacol. et foxicol. 1986b, 58, 219-224.
Mottet, N. K. & R. Body: Mercury burden of human autopsy organs and tissues. Arch. Environ. Health 1974, 29, 18-24.
Piotrowski, J. K. & M. J. Inskip: Health effects of methylmercury. MARC reporr No. 1981, 24, 1-8, 3848. Monitoring and Assess- ment Research Centre, Chelsea College, University of London.
Popescu, H. I.: Poisoning with alkylmercury compounds. Brit. Med. J . 1978, 1, 1347.
Prasad, K. N., E. Nobles & M. Ramanujam: Differential sensitivit- ies of glioma cells and neuroblastoma cells to methylmercury toxicity in cultures. Environ. Res. 1979, 19, 189-201.
Roussel, J. D. & C. R. Austin: Improved electro- ejaculation of primates. J . Instif. Animal Technicians 1968, 19(1), 22-32.
Rozynkowa, D. & B. Raczkiewcz: Destructive effect of methyl mercury chloride on human mitoses in living cells in vitro. Mutaf. Res. 1977, 56, 185-191.
Sakai, K.: Effects of methyl mercuric chloride on rat spermatogen- esis. Kumamoto Med. J . (Kumamoto, Japan). 1972,25(3), 94-100.
Shandilya, L., T. B. Clarkson, M. R. Adams & J. C. Lewis: Effects of gossypol on reproductive and endocrine function of male Cynomologus monkeys (Macaca fascicularis). Biol. Reprod. 1982,
Steiner, R. A., A. P. Peterson, Yu. Conner, H. JYL, M. Gilbert, B. Ter-Penning & W. J. Bremner: Ultradian LH and testosterone rhythms in the adult male monkey, Macaca fascicularis. Endocrin-
Vogel, D. G., R. L. Margolis & N. K. Mottet: Effects of methyl mercury binding to microtubules. Toxicol. Appl. Pharmacol. 1985, 80,473486.
Wilson, L., M. A. Jordan, A. Morse & R. L. Margolis: Interaction of vinblastine with steady-state microtubules in vitro. J . Mol. Biol. 1982, 159, 125-149.
Wyrobek, A. J. & W. R. Bruce: Chemical induction of sperm abnor- malities in mice. Proc. Natl. Acad. Sci. USA 1975, 72(11), 44254429.
27, 241-252.
ology 1980, 107(5), 1489-1493.





























