Embed Size (px)
Citation preview

Veterinary Microbiology 97 (2003) 123–133
Epidemiology ofMycobacterium bovisinfections ofpigs and wild boars using a molecular approach
A. Parraa,∗, P. Fernández-Llariob, A. Tatoc, J. Larrasad, A. Garcıae,J.M. Alonsoa, M. Hermoso de Mendozaa,
J. Hermoso de Mendozaa
a Cátedra de Patolog´ıa Infecciosa y Epidemiolog´ıa, Departamento de Medicina y Sanidad Animal,Facultad de Veterinaria de Cáceres, UEX, Avda. de la Universidad s/n, 10071 Cáceres, Spain
b Departamento de Biolog´ıa y Geolog´ıa, Instituto de Enseñanza Secundaria Santa Luc´ıa del Trampal,10160 Alcuéscar, Cáceres, Spain
c Servicios Veterinarios, Centro de Atención Primaria San Jorge, Servicio Extremeño de Salud (SES),Consejer´ıa de Sanidad y Consumo, Junta de Extremadura, Cáceres, Spain
d Laboratorios Larrasa S.L., C/Corredera Hernando de Soto 13-A, 06380 Jerez de los Caballeros, Spaine Bacteriology Division, Moredun Research Institute, Pentlands Science Park, EH26 OPZ Penicuik, UK
Received 14 January 2003; received in revised form 31 July 2003; accepted 18 August 2003
Abstract
A molecular epidemiological approach was applied to establishing a possible role for the wildboar as a natural reservoir ofMycobacterium bovisin Sierra de Villuercas, Western Spain; an areafree of farmed cattle and wild deer populations. Spoligo and VNTR typing were used over a threeyear period to study the epidemiological relationship between the occurrence of bovine tuberculosis(TB) in extensively bred Iberian pigs and indigenous wild boar. The 37 sampled wild boar showeddifferent degree of calcified granulomatous lesions in retropharyngeal, mediastinal and pulmonarylymph nodes. The 25 sampled Iberian pigs showed calcified lesions, mainly in the respiratory tract.Lesions located in the mesenteric lymph nodes appeared secondarily.M. boviswas isolated fromall affected animals. Twenty-five and 37 isolates ofM. boviswere obtained from domestic pigs andwild boar, respectively. Our findings provide evidence that supports the possibility of cross infectionbetween wild boar and domestic pig populations. This is contrary to the generally held belief thatswine represent an epidemiological dead end host and play no role in the epidemiology ofM. bovis.© 2003 Elsevier B.V. All rights reserved.
Keywords:Pig; Wild boar; Bovine tuberculosis; Spoligotyping; VNTR; Mediterranean basin
∗ Corresponding author. Tel.:+34-927257129; fax:+34-927257119.E-mail address:[email protected] (A. Parra).
0378-1135/$ – see front matter © 2003 Elsevier B.V. All rights reserved.doi:10.1016/j.vetmic.2003.08.007

124 A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133
1. Introduction
Feral and domestic swine are considered end hosts forMycobacterium bovisinfections.Evidence to support this belief has been provided by the observed decline in the prevalenceof M. bovison removal of the source of infection (McInerney et al., 1995; O’Reilly andDaborn, 1995). This decline has become apparent in countries where intensive pig breedingsystems are run in parallel with effective bovine TB control programs and the majority ofTB cases in swine are attributed to infection withMycobacterium avium.
In swine, most of the lesions are formed in the mesenteric or mandibular lymph nodeswith dissemination occasionally resulting in lesions in the liver, spleen and lungs. Theselesions are fibrotic, rarely calcified and scarcely caseous (Thoen et al., 1976; Corner et al.,1991; Feizabadi et al., 1996) supporting the theory that pigs are dead end hosts for TB.While this may be true for TB due toM. aviuminfection, this situation may not necessarilyapply in all types of swine tuberculosis. In countries such as Italy or Spain where bovine TBcontrol programs have been implemented relatively recently,M. bovisinfection in swineis not uncommon. Local enzootic situations, where the prevalence ofM. bovis is high,have been reported in these countries (Aranaz et al., 1996; Liebana et al., 1997; Serrainoet al., 1999; Bollo et al., 2000). In these areas a respiratory route of infection is recognisedand cattle are thought to represent the source of these outbreaks (McInerney et al., 1995;O’Reilly and Daborn, 1995; Bolin et al., 1997; Bollo et al., 2000).
Wild boar and extensively bred Iberian race pigs have important populations in westernand southern Spanish regions. In these areas young pigs are raised from 3 to 16 monthsin a free-range system, sharing natural resources with other wild and domestic animals.Topographically, the area selected for study comprises an abrupt mountainous area in Ex-tremadura, Western Spain, where TB has recently been detected in hunted wild boar andpigs slaughtered from October to February between 1998 and 2001. This area was selectedon the grounds that there is no local cattle farming and because it supports very low numbersof wild deer (Fernandez-Llario and Mateos-Quesada, 2003).
In this study, a molecular approach was applied to elucidating the epidemiological rela-tionship betweenM. bovisstrains isolated from both wild boar and domestic pigs. Spolig-otyping, a PCR-based method designed to detect the presence of spacer regions within theDR locus of theM. bovisgenome (Kamerbeek et al., 1997), has become a routine epidemi-ological tool. However, spoligotyping used as a stand-alone technique is considered to beonly moderately discriminatory forM. bovis isolates (Roring et al., 1998; Kremer et al.,1999). Recent studies have shown that in combination with other molecular typing meth-ods in a multiplexed approach, spoligotyping can be used to achieve much higher levelsof discrimination (Goguet de la Salmoniere et al., 1997; Aranaz et al., 1998; Roring et al.,1998; Kremer et al., 1999; Skuce, 2001; Skuce et al., 2002).
2. Material and methods
2.1. Geographic area of study
Sampling was carried out in the Sierra de las Villuercas, Extremadura, Western Spain,a wide area of about 100 km2. Las Villuercas is constituted by mountain ranges of greater

A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133 125
Fig. 1. Geographical situation of the estates inside the study area—v: valley area dedicated mainly to pig breeding(pg); m: mountain area, dedicated fundamentally to wild boar hunt (w). The properties B and J, located insidethe mountainous area, also support pig production. The numbers indicate spoligotyping-VNTR types detected ineach estate.
altitude (up to 1601 m) and an adjacent valley with the mountainous area (Fig. 1). Livestockin the region, mainly Iberian pigs and some goats, are kept in an extensive system, withusually low population densities. Several wild animal species, especially wild boar (Susscrofa) but also a low population of red deer (Cervus elaphus) share the same habitat withthe livestock (Fernandez-Llario and Mateos-Quesada, 2003).
2.2. Specimen collection
(i) Wild boar. During the hunting seasons 1998–2001 (October–February), retropharyn-geal lymph nodes and lung samples from 37 wild boar that showed tuberculosis-likelesions were collected. Every sample from each animal was identified, lesions weredescribed and the place where the animal was killed was identified. To avoid crosscontamination, different sets of sterile instruments and vials were used to collect andtransport the samples from each animal. The animals came from 11 game estates used

126 A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133
for hunting located mainly in the mountainous area within the area of study (Fig. 1),designates as A–C, E, F, H–J, L–N. The origin of the wild boars were 1 from A, 12from B, 6 from C, 2 from E, 1 from F, 1 from H, 1 from I, 7 from J, 1 from L, 4 fromM and 1 from N. The estates B and J also support domestic extensive pig breeding.
(ii) Domestic pigs. Retropharyngeal, mediastinal lymph nodes, and lung samples weretaken from 25 Iberian bred pigs slaughtered from October to December in 1999 and2000. These animals were less than 1-year-old and came from five estates mainlylocated in the valley adjacent to the mountainous area and inside the hunting area. Theorigin of 25 pigs was estates C and J inside the mountainous hunting area (2 from Cand 8 from J) and 15 were from three different estates in the valley, designed as D, Gand K (12 from D, 2 from G and 1 from K).
2.3. Decontamination procedure and identification
Tissue samples were divided into two similar sized portions, one of which was stored at−80◦C to allow re-testing, if needed. The hexadecylpyridinium chloride method (Cornerand Trajstman, 1988; Corner, 1993) was used for decontamination and Lowenstein–Jensenslants, containing pyruvate, were inoculated for culture. Isolates were identified on the basison their macroscopic features and standard biochemical tests (Casal, 1990). Identificationswere assessed by molecular tests using TB1-F and TB1-R primers that generate a DNAsegment of 372 bp specific for theM. tuberculosiscomplex MPB70 gene (Cousins et al.,1991; Liebana et al., 1996).
2.4. DNA extraction
Cells of eachM. bovisstrain were removed gently from the Lowenstein–Jensen slants,transferred into a microcentrifuge tube containing 100�l 10 mM Tris–HCl/1 m M EDTAbuffer, and incubated at 95◦C for 20 min. After centrifugation of the suspension, the super-natant, containing DNA, was harvest and stored at−20◦C until further use. DNA samplesfrom 36 wild-type strains ofM. bovis, andM. bovisBCG Pasteur II as reference strain wereused in this study.
2.5. Spoligotyping
Spoligotyping was performed as described previously byKamerbeek et al. (1997). TheDNA amplification was performed with 50�l volumes of reaction mixtures containing5�l of buffer reaction (335 mM Tris–HCl (pH 8.8 at 25◦C), 83 mM SO4(NH4)2), 3�l of25 mM MgCl2, 20 pmol of each oligonucleotide primers, 1.2 U ofTaqpolymerase enzyme(Biotech International, Bentley, Australia), and approximately 25 ng of template DNA. Theoligonucleotide primers DRa (5′-CCG AGA GGG GAC GCA AAC-3′) and DRb (5′-GGTTTT GGG TCT GAC GAC-3′) amplify the spacer sequences between the DRs. PrimerDRb was biotinylated at the 5′-end. The cycling temperatures were 1 min at 96◦C, 1 minat 55◦C and 30 s at 72◦C for 30 cycles. The first denaturation and the final extensionwere 10 min. The amplified DNA was tested hybridised with a set of 43 oligonucleotidesderived from the spacer sequences ofM. tuberculosisH37Rv and distilled water as negative

A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133 127
control. The multiple synthetic spacer oligonucleotides were covalently bound to a nylonmembrane (Biodyne C, Pall Biosupport, Postmouth, United Kingdom), in parallel lines.Hybridisation was performed in a miniblotter (MN45, Immunetics, Cambridge, MA). The20�l of PCR product was added in 150�l of 2× SSPE–SDS (1× SSPE is 0.15 M NaCl,10 mM NaH2PO4, and 1 mM EDTA (pH 7.4)–0.1% of sodium dodecyl sulphate) at 60◦Cfor 10 min. The membrane was washed twice in 250 ml of 2× SSPE–0.5% SDS for 10 minat 60◦C. Bound fragments were revealed by chemiluminescence after incubation withhorseradish peroxidase-streptavidin (Boehringer, Mannheim, German) for 45 min at 45◦Con exposure of the membrane to X-ray film (Hyperfilm, Amersham) for 10–12 min.
The spoligotyping profiles were expressed in a octal code format following publishedrecommendations (Dale et al., 2001). For the octal code system, the pattern was read ingroups of three, as a three-digit binary series 000, 001, 010, 011, 100, 101, 110, 111, read,respectively, as 0, 1, 2, 3, 4, 5, 6, 7; the 43rd spacer is read simply as 0 or 1.
2.6. VNTR typing
VNTR typing was essentially performed as described byFrothingham and Meeker-O’Connell (1998)with slight modifications. Briefly, PCR was performed with 20�l vol-umes of reaction mixtures containing 1 U ofTaq polymerase enzyme (Amersham), 10×PCR buffer (Amersham), 2 mM of MgCl2, 100 nM of each primer, 200�M of each of thefour dNTPs (Amersham) and 25 ng of DNA sample. The primers published previously (14)were designed to anneal upstream and downstream of each of five loci (ETR-A–ETR-E)containing exact tandem repeats (ETRs). The cycling temperatures were an initial denat-uration of 12 min at 96◦C was followed by 35 cycles of denaturation at 94◦C for 30 s,annealing at 60◦C for 1 min and extension at 72◦C for 2 min, followed by final extensionat 72◦C for 10 min. Negative controls (reagents only) were included. An “Allele CallingTable” was created based on the predicted PCR product size by the presence of differentnumbers of repeat units present at each locus (Filliol et al., 2000; McCarroll et al., 2001).The presence and size of each PCR product was determined by electrophoresis on a 2.5%agarose gel in Tris/boric acid/EDTA buffer running with molecular weight standards (50 bpladder Amersham) every 10 lines, followed by staining with ethidium bromide. A digitalcode could be calculated for each strain.
2.7. Computer-assisted analysis of the patterns
Vision works (UVP) and gel works 1D advanced (UVP) were used to compare VNTRand hybridisation patterns. In spoligotyping, each positive spot was defined as a band.The software clustered strains with the same DNA fingerprint pattern, defined similaritycoefficient (DICE coefficient), cluster dendrograms (UPGMA) and recorded the picture ofthe typical pattern for each strain.
2.8. Discriminatory power index determination
To measure objectively the power of discrimination of each molecular technique thediscriminatory index (DI) was calculated as previously reported (Struelens, 1996).

128 A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133
Fig. 2. Dendrogram of genetic relationships among 62M. boviswild isolates from the area of study based onspoligotyping plus VNTR typing. Column I represents strain designation, species (pg: pig, w: wild board) estateof origin (A–I, K, M and N) and hunting season (8= 1998/1999, 9= 1999/2000, 0= 2000/2001). Column IIrepresents main cluster designation. Column III represents spoligotype allele designation and column VI representsVNTR allele profile.
A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133 129
3. Results
3.1. Wild boar and domestic Iberian pigs
The 37 sampled wild boar showed different degrees of calcified granulomatous lesionsin retropharyngeal, mediastinal and pulmonary lymph nodes. The 25 sampled Iberian pigsshowed calcified lesions, mainly in the respiratory tract. Lesions located in the mesentericlymph nodes appeared secondarily (one mesenteric lymph nodes lesion). All isolates (62isolates, 37M. bovisstrains from wild boar and 25 from Iberian pig) were biochemicallyidentified asM. bovis, their identity was confirmed by PCR (Cousins et al., 1991).
3.2. Genomic DNA analysis
Spoligotyping and VNTR typing analysis was performed for all the mycobacteria isolated(37 from wild boar, 25 from domestic pigs). All experiments were performed twice, giving
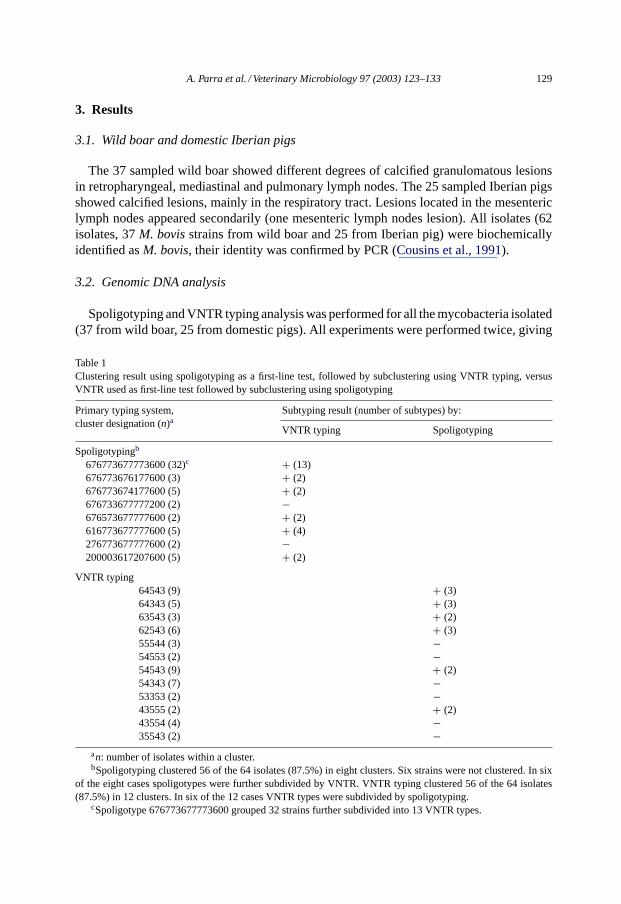
Table 1Clustering result using spoligotyping as a first-line test, followed by subclustering using VNTR typing, versusVNTR used as first-line test followed by subclustering using spoligotyping
Primary typing system,cluster designation (n)a
Subtyping result (number of subtypes) by:
VNTR typing Spoligotyping
Spoligotypingb
676773677773600 (32)c + (13)676773676177600 (3) + (2)676773674177600 (5) + (2)676733677777200 (2) −676573677777600 (2) + (2)616773677777600 (5) + (4)276773677777600 (2) −200003617207600 (5) + (2)
VNTR typing64543 (9) + (3)64343 (5) + (3)63543 (3) + (2)62543 (6) + (3)55544 (3) −54553 (2) −54543 (9) + (2)54343 (7) −53353 (2) −43555 (2) + (2)43554 (4) −35543 (2) −
an: number of isolates within a cluster.bSpoligotyping clustered 56 of the 64 isolates (87.5%) in eight clusters. Six strains were not clustered. In six
of the eight cases spoligotypes were further subdivided by VNTR. VNTR typing clustered 56 of the 64 isolates(87.5%) in 12 clusters. In six of the 12 cases VNTR types were subdivided by spoligotyping.
cSpoligotype 676773677773600 grouped 32 strains further subdivided into 13 VNTR types.

130 A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133
reproducible fingerprints for each strain in both techniques. According to the genomicdifferentiation, using spoligotyping alone, a total of 56 strains (87.5% of the isolates) weregrouped in eight clusters, six isolates were unique and 32 strains (55%) were grouped inone cluster (type 676773677773600). Using VNTR typing alone, we found 13 clusterscontaining 56 strains (87.5% of the isolates) and six strains were considered unique. VNTRtyping was able to subdivide almost all the spoligotyping clusters in VNTR-defined subtypes(Fig. 2andTable 1). Using spoligotyping alone the DI was 0.72 and using VNTR alone theDI was 0.93. Although, when we used spoligotyping in combination with VNTR typing,the ID was 0.967, sufficient to be applied to epidemiological purposes (Struelens, 1996;Skuce, 2001). Spoligotyping plus VNTR typing results allowed identification of 43 (67.2%)strains grouped in 14 clusters and 21 individual non-clustered strains.
Spoligotyping plus VNTR typing as the more discriminatory system reveals, a complexepidemiological situation in the area of study. From the point of view of the species, fourclusters (denoted as 3, 7, 9 and 14,Fig. 2) were exclusively isolated from domestic pigs andseven clusters (6, 10, 23, 29, 30, 31 and 35) were exclusive of wild boars, while two clusters(1 and 4) were shared by domestic and wild species, grouping 14 strains. TheM. bovisstrainswere grouped with a good correlation with the geographical origin of strains (Fig. 2).
From the point of view of the estates, three showed a high epidemiological complexity.In the mountain wild boar state B, molecular markers identified nine types and the clusterdenoted as 4 was time stable over 2 years. In the same way, the estate J showed 10 types, bothestates were in the centre of the area of study and support both types of pigs, which sharefood and water resources. In the valley, estate D showed 10 types restricted to domestic pigs.
4. Discussion
M. boviswas the only mycobacterial species found in the tuberculosis-like lesions studied.This finding combined with the type and distribution of lesions, was in contrast to thethinking of authors describing the situation in the UK, USA and Northern EU, where itwas considered thatM. aviumwas the main agent of swine TB (Thoen et al., 1976; Esseyand Koller, 1994; Komijn et al., 1999). These authors describe the situation in areas wherebovine TB control has been ongoing for many years, and where pigs are reared intensively.
Although cattle are virtually non-existent in the “Sierra de las Villuercas”, the extensivedomestic pig breeding system and the presence of substantial, uncontrolled populations ofwild boar could be an important factor that favour the presence, maintenance and spread ofM. bovis. During the 3-year study period, the Official Veterinary Services reported that preva-lence in condemnation of hunted wild boar carcasses due to the presence of tubercle-likelesions in the area of study, ranged from 1.1 ± 0.47% in 1998 to 2.89 ± 0.66% in 2001analysing more than 3.000 wild boars carcasses.
The predominantly pulmonary lesions, miliary, caseous or calcified, are signs of a res-piratory route of infection in both species (Essey et al., 1981; Corner et al., 1991; Serrainoet al., 1999). This fact, together with their feeding and mobility habits, suggest that swineare not dead end hosts forM. bovis. The Iberian pigs are periodically slaughtered and re-placed by a new population, and the 13-month free-range period is clearly sufficient fortransmission from the infected wild boar population and development of severe generalised

A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133 131
conditions with open lesions, that allow spread of bacilli from these animals. Previous stud-ies demonstrated the existence of common molecular types between isolates from wild boar,red deer and cattle in Spain (Aranaz et al., 1996; Liebana et al., 1997) and other countriessuch as Italy (Serraino et al., 1999); and the epidemiological implications of wild boar inthe spread of bovine TB (Essey et al., 1981; Corner et al., 1991; McInerney et al., 1995)but never reported the epidemiological relationship between domestic pigs and wild boarin the Mediterranean basin.
Our result agree with others regarding to the low power of discrimination using spoligo-typing alone (DI= 0.72)(15). Following previous work (Filliol et al., 2000; Roring et al.,2002) VNTR typing provided reproducible results, an accurate level of discrimination andthis has the potential to increase our knowledge of bovine TB epidemiology. Spoligotypingplus VNTR typing results also provided evidence that some swine isolates are identical andspread freely between both swine (feral and domestic) populations. Clusters are clearly ge-ographically restricted to the estate area where the pigs were bred or wild boar were hunted(Fig. 2), but some estates in the centre of the area of study showed a high epidemiologicalcomplexity. The problematic states (B and J), because they maintain a production of swine,may possess better resources that the rest of properties of the mountainous area, so that thiswould induce the entrance of boars in search of food, allowing the transmission betweenboth types of pigs. Independent of this situation, our work supports the theory that boar canbe true host forM. bovisin an area without bovine livestock and a low density of deer.
Acknowledgements
The present study has been possible by funding of the Spanish Ministry of Science andTechnology grants 1FD97-0930 and AGL2001-0536, but also by the collaboration of theVeterinary Services of the Servicio Extremeño de Salud de la Consejerıa de Sanidad yConsumo, Servicio de Sanidad Animal and Servicio de Protección de la Naturaleza, Cazay Pesca de la Consejerıa de Agricultura y Medio Ambiente de la Junta de Extremadura.Special thanks to Alicia Aranaz and coworkers at the Universidad Complutense de Madridand Solving Roring, Robin Skuce and Sidney Neill at the University of Belfast and Neil F.Inglis at the Moredun Research Institute for their technical and personal help.
References
Aranaz, A., Liebana, E., Mateos, A., Dominguez, L., Vidal, D., Domingo, M., Gonzolez, O., Rodriguez-Ferri, E.F.,Bunschoten, A.E., Van Embden, J.D., Cousins, D., 1996. Spacer oligonucleotide typing ofMycobacteriumbovisstrains from cattle and other animals: a tool for studying epidemiology of tuberculosis. J. Clin. Microbiol.34, 2734–2740.
Aranaz, A., Liebana, E., Mateos, A., Dominguez, L., Cousins, D., 1998. Restriction fragment length polymorphismand spacer oligonucleotide typing: a comparative analysis of fingerprinting strategies forMycobacterium bovis.Vet. Microbiol. 61, 311–324.
Bolin, C.A., Whipple, D.L., Khanna, K.V., Risdahl, J.M., Peterson, P.K., Molitor, T.W., 1997. Infection of swinewith Mycobacterium bovisas a model of human tuberculosis. J. Infect. Dis. 176, 1559–1566.
Bollo, E., Ferroglio, E., Dini, V., Mignone, W., Biolatti, B., Rossi, L., 2000. Detection of complex in lymph nodesof wild boar (Sus scrofa) by a target-amplified test system. J. Vet. Med. B 47, 337–342.

132 A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133
Casal, M., 1990. Microbiologıa clınica de las enfermedades por Micobacterias (Tuberculosis, Lepra yMicobacteriosis), Corboba, pp. 1–90.
Corner, L.A., 1993. Australian Standard Diagnostic Techniques for Animal Diseases. CSIRO, East Melbourne.Corner, L.A., Trajstman, A.C., 1988. An evaluation of 1-hexadecylpyridinium chloride as a decontaminant in the
primary isolation ofMycobacterium bovisfrom bovine lesions. Vet. Microbiol. 18, 127–134.Corner, L.A., Barret, R.H., Lepper, A.W.D., Pearson, C.W., 1991. A survey of mycobacteriosis of feral pigs in the
Northern Territory. Aust. Vet. J. 57, 537–542.Cousins, D.V., Wilton, S.D., Francis, B.R., 1991. Use of DNA amplification for the rapid identification of
Mycobacterium bovis. Vet. Microbiol. 27, 187–195.Dale, J.W., Brittain, D., Cataldi, A.A., Cousins, D., Crawford, J.T., Driscoll, J., Heersma, H., Lillebaek,
T., Quitugua, T., Rastogi, N., Skuce, R.A., Sola, C., Van Soolingen, D., Vincent, V., 2001. Spaceroligonucleotide typing of bacteria of theMycobacterium tuberculosiscomplex: recommendations forstandardised nomenclature. Int. J. Tuberc. Lung Dis. 5, 216–219.
Essey, M.A., Koller, M.A., 1994. Status of bovine tuberculosis in North America. Vet. Microbiol. 40, 15–22.Essey, M.A., Payne, R.L., Himes, E.M., Luchsinger, D., 1981. Bovine tuberculosis surveys of Axis deer and feral
swine on the Hawaiian island of Molakai. Proc. US Anim. Health Assoc. 85, 538–549.Feizabadi, M.M., Robertson, I.D., Cousins, D.V., Dawson, D., Chew, W., Gilbert, G.L., Hampson, D.J., 1996.
Genetic characterization ofMycobacterium aviumisolates recovered from humans and animals in Australia.Epidemiol. Infect. 116, 41–49.
Fernandez-Llario, P., Mateos-Quesada, P., 2003. Population structure of the wild boar (Sus scrofa) in twoMediterranean habitats in the western Iberian Peninsula. Folia Zool. 52, 143–148.
Filliol, I., Ferdinand, S., Negroni, L., Sola, C., Rastogi, N., 2000. Molecular typing ofMycobacterium tuberculosisbased on variable number of repeat tandem DNA repeats used alone and in association with spoligotyping. J.Clin. Microbiol. 38, 2520–2524.
Frothingham, R., Meeker-O’Connell, W.A., 1998. Genetic diversity in theMycobacterium tuberculosiscomplexbased on variable numbers of tandem DNA repeats. Microbiology 144, 1189–1196.
Goguet de la Salmoniere, Y.O., Li, H.M., Torrea, G., Bunschoten, A., van Embden, J., Gicquel, B., 1997. Evaluationof spoligotyping in a study of the transmission ofMycobacterium tuberculosis. J. Clin. Microbiol. 35, 2210–2214.
Kamerbeek, J., Schouls, L., Kolk, A., van Agterveld, M., van Soolingen, D., Kuijper, S., Bunschoten, A.,Molhuizen, H., Shaw, R., Goyal, M., van Embden, J., 1997. Simultaneous detection and strain differentiationof Mycobacterium tuberculosisfor diagnosis and epidemiology. J. Clin. Microbiol. 35, 907–914.
Komijn, R.E., de Haas, P.E., Schneider, M.M., Eger, T., Nieuwenhuijs, J.H., van den Hoek, R.J., Bakker, D.,van Zijd Erveld, F.G., van Soolingen, D., 1999. Prevalence ofMycobacterium aviumin slaughter pigs in TheNetherlands and comparison of IS1245 restriction fragment length polymorphism patterns of porcine andhuman isolates. J. Clin. Microbiol. 37, 1254–1259.
Kremer, K., van Soolingen, D., Frothingham, R., Haas, W.H., Hermans, P.W., Martin, C., Palittapongarnpim, P.,Plikaytis, B.B., Riley, L.W., Yakrus, M.A., Musser, J.M., van Embden, J.D., 1999. Comparison of methodsbased on different molecular epidemiological markers for typing ofMycobacterium tuberculosiscomplexstrains: interlaboratory study of discriminatory power and reproducibility. J. Clin. Microbiol. 37, 2607–2618.
Liebana, E., Aranaz, A., Francis, B., Cousins, D., 1996. Assessment of genetic markers for species differentiationwithin theMycobacterium tuberculosiscomplex. J. Clin. Microbiol. 34, 933–938.
Liebana, E., Aranaz, A., Dominguez, L., Mateos, A., Gonzalez-Llamazares, O., Rodriguez-Ferri, E.F., Domingo,M., Vidal, D., Cousins, D., 1997. The insertion element IS 6110 is a useful tool for DNA fingerprinting ofMycobacterium bovisisolates from cattle and goats in Spain. Vet. Microbiol. 54, 223–233.
McCarroll, J.F.J., McCorry, T.O., Brittainn, D., Smyth, M., Skuce, R., Neill, S., 2001. Evaluation of novelVNTR-PCR targets for typingMycobacterium bovisisolates. In: Proceedings of the Third InternationalConference onMycobacterium bovis, Cambridge.
McInerney, J., Small, K.J., Caley, P., 1995. Prevalence ofMycobacterium bovisinfection in feral pigs in theNorthern Territory. Aust. Vet. J. 72, 448–451.
O’Reilly, L.M.D., Daborn, C.J., 1995. The epidemiology ofMycobacterium bovisinfections in animals andhumans: a review. Tuberc. Lung Dis. 76 (Suppl. 1), 1–46.
Roring, S., Brittain, D., Bunschoten, A.E., Hughes, M.S., Skuce, R.A., van Embden, J.D., Neill, S.D., 1998. Spaceroligotyping ofMycobacterium bovisisolates compared to typing by restriction fragment length polymorphismusing PGRS, DR and IS6110 probes. Vet. Microbiol. 61, 111–120.

A. Parra et al. / Veterinary Microbiology 97 (2003) 123–133 133
Roring, S., Scott, A., Brittain, D., Walker, I., Hewinson, G., Neill, S., Skuce, R., 2002. Development ofvariable-number tandem repeat typing ofMycobacterium bovis: comparison of results with those obtainedby using existing exact tandem repeats and spoligotyping. J. Clin. Microbiol. 40, 2126–2133.
Serraino, A., Marchetti, G., Sanguinetti, V., Rossi, M.C., Zanoni, R.G., Catozzi, L., Bandera, A., Dini, W., Mignone,W., Franzetti, F., Gori, A., 1999. Monitoring of transmission of tuberculosis between wild boars and cattle:genotypical analysis of strains by molecular epidemiology techniques. J. Clin. Microbiol. 37, 2766–2771.
Skuce, R., 2001. Development of novel standardised methodology and nomenclature for identification ofMycobacterium bovisstrains. In: Proceedings of the Third International Conference onMycobacterium bovis,Cambridge.
Skuce, R.A., McCorry, T.P., McCarroll, J.F., Roring, S.M., Scott, A.N., Brittain, D., Hughes, S.L., Hewinson, R.G.,Neill, S.D., 2002. Discrimination ofMycobacterium tuberculosiscomplex bacteria using novel VNTR-PCRtargets. Microbiology 148, 519–528.
Struelens, M.J., 1996. Consensus guidelines for the appropriate use and evaluation of microbial epidemiologictyping systems. Clin. Microbiol. Infect. 2, 2–11.
Thoen, C.O., Himes, E.M., Weaver, D.E., Spangler, G.W., 1976. Tuberculosis in brood sows and pigs slaughteredin Iowa. Am. J. Vet. Res. 37, 775–778.





























