Embed Size (px)
Citation preview

www.elsevier.com/locate/ymcne
Mol. Cell. Neurosci. 28 (2005) 703–714
Experimental Charcot–Marie–Tooth type 1A: A cDNA
microarrays analysis
Tiziana Vigo,a,b,1 Lucilla Nobbio,a,b,1 Paul Van Hummelen,c Michele Abbruzzese,a,d
GianLuigi Mancardi,a,b Nathalie Verpoorten,e Kristien Verhoeven,e Michael W. Sereda,f
Klaus-Armin Nave,f Vincent Timmerman,e and Angelo Schenonea,b,*
aDepartment of Neurosciences, Ophthalmology and Genetics, University of Genova, Italy, via De Toni 5, 16132 Genova, ItalybCenter of Excellence for Biomedical Research, University of Genova, Italy, viale Benedetto XV, 16132 Genova, ItalycMicroArray Facility, Flanders Interuniversity Institute for Biotechnology, B-3000 Leuven, BelgiumdBioimaging and Molecular Physiology Institute, CNR, Genova, ItalyeDepartment of Molecular Genetics, Flanders Interuniversity Institute for Biotechnology, University of Antwerp, Antwerpen, B-2610, BelgiumfDepartment of Neurogenetics, Max-Planck Institute of Experimental Medicine, Hermann-Rein-Str. 3, D-37075 Gottingen, Germany
Received 11 June 2004; revised 25 November 2004; accepted 30 November 2004
To reveal the spectrum of genes that are modulated in Charcot–
Marie–Tooth neuropathy type 1A (CMT1A), which is due to
overexpression of the gene coding for the peripheral myelin protein
22 (pmp22), we performed a cDNA microarray experiment with
cDNA from sciatic nerves of a rat model of the disease. In
homozygous pmp22 overexpressing animals, we found a significant
down-regulation of 86 genes, while only 23 known genes were up-
regulated, suggesting that the increased dosage of pmp22 induces a
general down-regulation of gene expression in peripheral nerve
tissue. Classification of the modulated genes into functional categories
leads to the identification of some pathways altered by overexpression
of pmp22. In particular, a selective down-regulation of the ciliary
neurotrophic factor transcript and of genes coding for proteins
involved in cell cycle regulation, for cytoskeletal components and for
proteins of the extracellular matrix, was observed. Cntf expression
was further studied by real-time PCR and ELISA technique in
pmp22 transgenic sciatic nerves, human CMT1A sural nerve biopsies,
and primary cultures of transgenic Schwann cells. According to the
results of cDNA microarray analysis, a down-regulation of cntf, both
at the mRNA and protein level, was found in all the conditions
tested.
These results are relevant to reveal the molecular function of PMP22
and the pathogenic mechanism of CMT1A. In particular, finding a
specific reduction of cntf expression in CMT1A Schwann cells suggests
that overexpression of pmp22 significantly affects the ability of
Schwann cells to offer a trophic support to the axon, which could be
1044-7431/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.mcn.2004.11.016
* Corresponding author. Department of Neurosciences, Ophthalmology
and Genetics, University of Genova, Italy, via De Toni 5, 16132 Genova,
Italy. Fax: +39 010 3538639.
E-mail address: [email protected] (A. Schenone).1 The first two authors equally contributed to this project.
Available online on ScienceDirect (www.sciencedirect.com).
a factor, among other, responsible for the development of axonal
atrophy in human and experimental CMT1A.
D 2004 Elsevier Inc. All rights reserved.
Introduction
Charcot–Marie–Tooth (CMT) disease, with a prevalence of 1 in
2500, is the most common inherited peripheral neuropathy. CMT is
clinically and genetically heterogeneous, with autosomal dominant
(AD), recessive and X-linked transmission subtypes (Dyck et al.,
1993). Up to date more than 33 disease causing genes are known
for CMT and related peripheral neuropathies (http://www.molgen.
ia.ac.be/CMTMutations/).
Based on clinical, neuropathological, and genetic data, CMT
has been divided in different types. CMT type 1A (CMT1A) is an
AD demyelinating neuropathy normally due to a duplication of a
1.4 Mb region in chromosome 17p11.2–12, containing the gene
coding for the peripheral myelin protein 22 (PMP22) (Inoue et al.,
2001). Point mutations in the PMP22 gene may also cause
CMT1A (Roa et al., 1993; Valentijn et al., 1992).
PMP22, a member of an extended family of tetraspan membrane
proteins (Bolin et al., 1997; Magyar et al., 1997; Taylor et al., 1995),
is highly expressed in myelinating Schwann cells and in compact
myelin, where it represents 2–5% of total myelin proteins (Snipes et
al., 1999; Suter and Snipes, 1995). Pmp22 expression has also been
detected, during mouse development and in adulthood, in different
neural and non-neural tissues (Baechner et al., 1995). Even if
function and processing of PMP22 have been extensively studied,
the molecular mechanisms underlying CMT1A are still unclear
(Hanemann and Muller, 1998; Suter and Scherer, 2003).

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714704
Animal models of CMT1A have been developed (Huxley et al.,
1996; Magyar et al., 1996; Perea et al., 2001; Sereda et al., 1996).
Genetic characterization of pmp22 overexpressing nerves, in
CMT1A rats (Sereda et al., 1996), shows that Schwann cells
present abundant expression of genes encoding major structural
myelin proteins and aberrant co-expression of early Schwann cell
markers (Niemann et al., 2001).
As cDNA microarrays technology allows large-scale, compa-
rative gene expression profiling (Xiang et al., 2003) and has been
recently used to study gene expression in Schwann cells and sciatic
nerves of several animal models and in human nerves (Cameron et
al., 2003; Costigan et al., 2002; Kubo et al., 2002; Nagarajan et al.,
2001; Verheijen et al., 2003; Xiao et al., 2002), we performed a
cDNA microarrays experiment on sciatic nerves from a rat model
of CMT1A (Sereda et al., 1996) to reveal the complete spectrum of
genes that are modulated in the disease.
We found an altered expression level for 213 cDNA sequences,
among which several genes involved in specific pathways that may
be impaired in CMT1A. In particular, a selective down-regulation
of the ciliary neurotrophic factor (cntf) transcript was observed. As
cntf, which is produced by Schwann cells, specifically supports the
survival of motor and sensory neurons as well as the myelination
process (Sleeman et al., 2000; Stankoff et al., 2002), we further
studied its expression in human and experimental CMT1A and in
primary Schwann cells cultures from the CMT1A rat. We observed
a general down-regulation of cntf in CMT1A nerves and Schwann
cells. This result strongly suggests that pmp22 transgenic Schwann
cells are unable to offer an adequate trophic support to the axon,
leading to the late axonal atrophy observed in CMT1A nerves.
Results
Microarrays analysis and expression study on selected genes
In spite of the high sequence homology between mouse and rat
genomes, preliminary hybridization was conducted to test the
Fig. 1. (a) Scatter plot representing all the microarrays data. The duplicate spots we
The more red, the lower the P value. (b) Clones that were significant at the 5%
mouse cDNA array MouseV (VIB) with the rat RNA (data not
shown). Since these experiments showed that up to 80% of the
spotted sequences could be hybridized by rat probes, we proceeded
with testing gene expression in the pathological condition. We
performed two biological repeats, and every hybridization was
repeated in a dye swap. We considered up- or down-regulated
cDNA sequences in transgenic sciatic nerves when changes in
expression were greater than twofold compared to normal ones
(Fig. 1).
We found 213 cDNA sequences showing an altered expression
in transgenic nerves. Of these 213 cDNA sequences, 145 correlated
with 109 known genes and 68 with ESTs. The number of up-
regulated sequences was 55, referring to 23 known genes and 20
ESTs. Eighty-six known genes and 48 ESTs represented the group
of 158 down-regulated cDNA. In the array, more than a single
cDNA could represent a unique gene or an EST, so there was no
correspondence between the number of regulated cDNAs and the
genes. Considering the group of known genes, we observed that
only 22.5% were up-regulated, while most of the genes (77.5%)
were down-expressed. Furthermore, we classified the regulated
genes into functional categories on the basis of literature and using
Onto-Express database (Draghici et al., 2003) (Tables 1 and 2).
Among the up-regulated ones (Fig. 2a), we found a predominance
of genes coding for proteins involved in cell proliferation (16.7%),
transcription factors (12.5%), translation factors (8.3%), and signal
transducers (8.3%). The most representative categories in the
down-regulated group (Fig. 2b) were the proteins involved in
metabolic pathways (19.8%), integral membrane proteins (5.8%),
muscle (9.3%) and extracellular matrix components (10.4%), and
the cytoskeletal proteins (8.14%). Interestingly, we observed that
genes related to a specific functional category tended to cluster
among the up- or down-regulated ones (Fig. 3).
Several genes, known to be down- or up-regulated in CMT1A,
showed the expected pattern of expression. As previously observed
(Niemann et al., 2001), we found an up-regulation of the
transcription factor scip and of the mRNA coding for the low
affinity nerve growth factor receptor (p75ngfr), which is normally
re averaged. The color code in the picture is based on the significance level.
level.
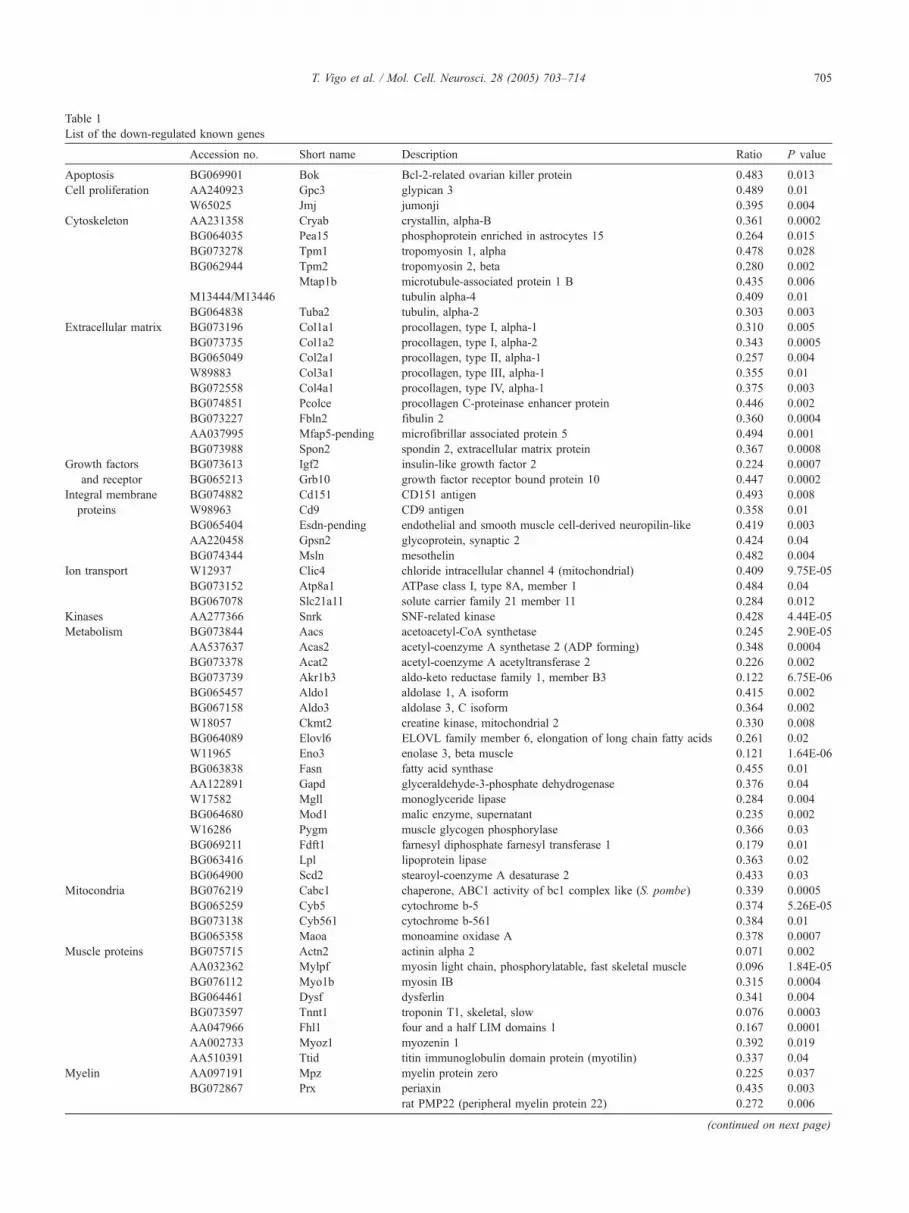
Table 1
List of the down-regulated known genes
Accession no. Short name Description Ratio P value
Apoptosis BG069901 Bok Bcl-2-related ovarian killer protein 0.483 0.013
Cell proliferation AA240923 Gpc3 glypican 3 0.489 0.01
W65025 Jmj jumonji 0.395 0.004
Cytoskeleton AA231358 Cryab crystallin, alpha-B 0.361 0.0002
BG064035 Pea15 phosphoprotein enriched in astrocytes 15 0.264 0.015
BG073278 Tpm1 tropomyosin 1, alpha 0.478 0.028
BG062944 Tpm2 tropomyosin 2, beta 0.280 0.002
Mtap1b microtubule-associated protein 1 B 0.435 0.006
M13444/M13446 tubulin alpha-4 0.409 0.01
BG064838 Tuba2 tubulin, alpha-2 0.303 0.003
Extracellular matrix BG073196 Col1a1 procollagen, type I, alpha-1 0.310 0.005
BG073735 Col1a2 procollagen, type I, alpha-2 0.343 0.0005
BG065049 Col2a1 procollagen, type II, alpha-1 0.257 0.004
W89883 Col3a1 procollagen, type III, alpha-1 0.355 0.01
BG072558 Col4a1 procollagen, type IV, alpha-1 0.375 0.003
BG074851 Pcolce procollagen C-proteinase enhancer protein 0.446 0.002
BG073227 Fbln2 fibulin 2 0.360 0.0004
AA037995 Mfap5-pending microfibrillar associated protein 5 0.494 0.001
BG073988 Spon2 spondin 2, extracellular matrix protein 0.367 0.0008
Growth factors
and receptor
BG073613 Igf2 insulin-like growth factor 2 0.224 0.0007
BG065213 Grb10 growth factor receptor bound protein 10 0.447 0.0002
Integral membrane
proteins
BG074882 Cd151 CD151 antigen 0.493 0.008
W98963 Cd9 CD9 antigen 0.358 0.01
BG065404 Esdn-pending endothelial and smooth muscle cell-derived neuropilin-like 0.419 0.003
AA220458 Gpsn2 glycoprotein, synaptic 2 0.424 0.04
BG074344 Msln mesothelin 0.482 0.004
Ion transport W12937 Clic4 chloride intracellular channel 4 (mitochondrial) 0.409 9.75E-05
BG073152 Atp8a1 ATPase class I, type 8A, member 1 0.484 0.04
BG067078 Slc21a11 solute carrier family 21 member 11 0.284 0.012
Kinases AA277366 Snrk SNF-related kinase 0.428 4.44E-05
Metabolism BG073844 Aacs acetoacetyl-CoA synthetase 0.245 2.90E-05
AA537637 Acas2 acetyl-coenzyme A synthetase 2 (ADP forming) 0.348 0.0004
BG073378 Acat2 acetyl-coenzyme A acetyltransferase 2 0.226 0.002
BG073739 Akr1b3 aldo-keto reductase family 1, member B3 0.122 6.75E-06
BG065457 Aldo1 aldolase 1, A isoform 0.415 0.002
BG067158 Aldo3 aldolase 3, C isoform 0.364 0.002
W18057 Ckmt2 creatine kinase, mitochondrial 2 0.330 0.008
BG064089 Elovl6 ELOVL family member 6, elongation of long chain fatty acids 0.261 0.02
W11965 Eno3 enolase 3, beta muscle 0.121 1.64E-06
BG063838 Fasn fatty acid synthase 0.455 0.01
AA122891 Gapd glyceraldehyde-3-phosphate dehydrogenase 0.376 0.04
W17582 Mgll monoglyceride lipase 0.284 0.004
BG064680 Mod1 malic enzyme, supernatant 0.235 0.002
W16286 Pygm muscle glycogen phosphorylase 0.366 0.03
BG069211 Fdft1 farnesyl diphosphate farnesyl transferase 1 0.179 0.01
BG063416 Lpl lipoprotein lipase 0.363 0.02
BG064900 Scd2 stearoyl-coenzyme A desaturase 2 0.433 0.03
Mitocondria BG076219 Cabc1 chaperone, ABC1 activity of bc1 complex like (S. pombe) 0.339 0.0005
BG065259 Cyb5 cytochrome b-5 0.374 5.26E-05
BG073138 Cyb561 cytochrome b-561 0.384 0.01
BG065358 Maoa monoamine oxidase A 0.378 0.0007
Muscle proteins BG075715 Actn2 actinin alpha 2 0.071 0.002
AA032362 Mylpf myosin light chain, phosphorylatable, fast skeletal muscle 0.096 1.84E-05
BG076112 Myo1b myosin IB 0.315 0.0004
BG064461 Dysf dysferlin 0.341 0.004
BG073597 Tnnt1 troponin T1, skeletal, slow 0.076 0.0003
AA047966 Fhl1 four and a half LIM domains 1 0.167 0.0001
AA002733 Myoz1 myozenin 1 0.392 0.019
AA510391 Ttid titin immunoglobulin domain protein (myotilin) 0.337 0.04
Myelin AA097191 Mpz myelin protein zero 0.225 0.037
BG072867 Prx periaxin 0.435 0.003
rat PMP22 (peripheral myelin protein 22) 0.272 0.006
(continued on next page)
T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714 705

Accession no. Short name Description Ratio P value
Neurotrophic factor AA543497 Cntf ciliary neurotrophic factor 0.075 6.15E-05
Transcription factors AA518455 Tcf4 transcription factor 4 0.440 0.001
W71604 Deaf1 deformed epidermal autoregulatory factor 1 (Drosophila) 0.256 0.01
Signal transduction AA217217 Itpkb inositol 1,4,5-trisphosphate 3-kinase B 0.498
AA020462 Rab2 RAB2, member RAS oncogene family 0.437 0.0006
BG063588 Rhoip3-pending rho interacting protein 3 0.412 0.002
BG076141 Chn1 chimerin 1 0.486
Others BG075128 Cetn2 centrin 2 0.441 0.016
BG063515 Fth ferritin heavy chain 0.258 0.019
AA049981 Gatm glycine amidinotransferase 0.383 0.008
AA474937 Epb4.1l2 erythrocyte protein band 4.1-like 2 0.474 0.004
BG063261 H19 H19 fetal liver mRNA 0.435 0.002
BG069748 Lims2 LIM and senescent cell antigen like domains 2 0.134 0.001
BG073671 Mfge8 milk fat globule-EGF factor 8 protein 0.434 0.0008
BG073463 Odf2 outer dense fiber of sperm tails 2 0.406 0.02
BG073096 Olfm1 olfactomedin 1 0.487 0.007
BG073341 Rsdr1-pending retinal short-chain dehydrogenase/reductase 1 0.496 0.003
AA461746 Ssg1-pending steroid-sensitive gene 1 0.458 0.001
AA030949 Necl1-pending nectin-lke 1 0.284 0.001
BG067852 Pros1 protein S (alpha) 0.211 0.02
BG075594 Pxp-pending peroxisomal protein 0.375 0.039
BG066411 Serpine2 serine (or cysteine) proteinase inhibitor, clade E, member 2 0.406 0.006
BG074009 Uchl1 ubiquitin carboxy-terminal hydrolase L1 0.461 0.0004
BG076040 Zdhhc2 zinc finger, DHHC domain containing 2 0.497 0.017
AA015155 S100a3 S100 calcium binding protein A3 0.322 0.0002
Several genes were represented in the array by more than one sequence; however, only one sequence is listed for each gene.
Table 1 (continued)
T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714706
expressed by non-myelinating Schwann cells and immature
Schwann cells precursors. Some genes known to carry point
mutations responsible for other types of CMT, like the periaxin
(prx) and the laminin A (lmna) (De Sandre-Giovannoli et al., 2002;
Guilbot et al., 2001), were also differentially expressed. Moreover,
Table 2
List of the up-regulated known genes
Accession no. Short name Descrip
Basal lamina AA066180 Lmna lamin A
Cell proliferation BG070163 Ccnd1 cyclin D
BG066310 Ccnd3 cyclin D
AA272260 Csrp2 cysteine
B-cell t
BG072743 Btg1 anti-pro
Cytoskeleton BG074004 Vil2 villin 2
Extracellular matrix BG067727 Tnfrsf12a tumor n
Growth factors and receptor AA048449 Ngfr nerve g
Integral membrane protein W12889 Cdh3 cadherin
Myelin BG075879 Plp Proteoli
Poliamine catabolism BG072707 Sat spermid
Signal trasduction BG072288 Adcy9 adenyla
AA013851 Rap1ga1 Rap1, G
Transcription factors BG071421 Idb2 inhibito
BG065255 Scip POU do
W14398 Sox4 SRY-bo
Translation factors AA068436 Bzw2 basic le
BG069032 Gc20-pending translati
Others AA063753 Abca1 ATP-bin
BG069237 Abhd3 abhydro
BG072800 BC038058 transcri
W36002 Oraov1 oral can
BG066068 Zfp216 zinc fin
BG075959 Pcbp4 poly(rC
Several genes were represented in the array by more than one sequence, howeve
we identified a modulation of a few genes lying in chromosomal
regions associated with different forms of dominant and recessive
CMT (Table 3) (Berciano and Combarros, 2003). These genes may
be studied, in the future, as positional candidate in mutational
analysis.
tion Ratio P value
2.652 0.04
1 3.809 0.02
3 2.771 0.01
-rich protein 2 9.189 1.21E-05
ranslocation gene 1, 0.0005
liferative 2.842
2.371 0.002
ecrosis factor receptor superfamily, member 12a 5.256 0.005
rowth factor receptor 3.160 0.005
3 2.476 0.04
pid protein (myelin) 2.198 0.022
ine/spermine N1-acetyl transferase 3.837 0.001
te cyclase 9 2.027 0.045
TPase-activating protein 1 2.402 0.006
r of DNA binding 2 2.423 0.0005
main, class 3, transcription factor 1 9.077
x containing gene 4 2.526 0.02
ucine zipper and W2 domains 2 3.710 0.0005
on factor sui1 homolog 3.667 0.01
ding cassette, sub-family A (ABC1), member 1 3.999 0.004
lase domain containing 3 2.076 0.04
ption termination factor, mitochondrial-like 2.048 0.02
cer overexpressed 1 2.753 0.004
ger protein 216 2.686 0.001
) binding protein 4 2.094 0.012
r only one sequence is listed for each gene.

Fig. 2. Categorization of up-regulated genes (a) and down-regulated genes
(b) into functional categories.
T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714 707
Genes involved in cell cycle (cyclin D1 and cyclin D3), in cell
adhesion and motility (cd9-antigen), and trophic support (cntf)
were selected to confirm the microarray results by semiquantitative
RT-PCR reactions. We observed a full correspondence between
levels of gene expression in microarray hybridization and semi-
quantitative RT-PCR (Table 4). A significant (P b 0.05) up-
regulation of the cyclin (ccnd1 and ccnd3) transcripts was found in
Fig. 3. Categorization of modulated genes from the most representative functio
percentage was calculated by division of the number of down- or up-regulated g
both hemizygous and homozygous rats as compared to normal
controls. A significant (P b 0.05) down-regulation of the ciliary
neurotrophic factor (cntf) and the cd9-antigen transcripts was
observed only in the homozygous pmp22 overexpressing nerves,
but the hemizygous ones also showed lower levels of these genes
compared to normal controls. We also analyzed the expression
levels of genes coding for other important neurotrophic factors
using semiquantitative RT-PCR. We tested the expression of nerve
growth factor (ngf), brain derived neurotrophic factor (bdnf), glial
derived neurotrophic factor (gdnf), and neurotrophin 3 (nt3). The
cDNA corresponding to those transcripts was spotted on the mouse
gene chip but did not show altered expression in transgenic nerves.
The results of the RT-PCR experiments (Fig. 4) confirmed that cntf
is the only neurotrophic factor significantly down-regulated in
pmp22 overexpressing nerves.
Cntf expression in CMT1A
Considering the strong and specific reduction of cntf mRNA in
pmp22 overexpressing nerves, we further studied its expression in
experimental and human CMT1A and in cultures of pmp22
overexpressing Schwann cells.
In archived sural nerves, we could quantify by real-time PCR
the CNTF mRNA in all normal controls. Instead, the transcript was
not detectable in CMT1A nerves. To reliably compare cntf
expression in human and experimental CMT1A, we repeated the
transcript analysis, in rat sciatic nerves, by real-time PCR. Again, a
down-regulation of cntf was found in pmp22 transgenic hemi-
zygous (0.49 F 0.07) and homozygous (0.02 F 0.01) nerves
compared to normal controls.
As a reduced expression of cntf could be merely due to a loss of
Schwann cells in CMT1A nerves, we counted Schwann cells
number in pmp22 rat nerves and in normal controls. We did not
observe any difference between homozygous (13.4 F 0.4 cells/
mm2), hemizygous (13.4 F 0.65 cells/mm2), and normal nerves
(12.5 F 1.34 cells/mm2).
nal categories in the up (gray)- and down (white)-regulated groups. The
enes by the total number of genes in each category.

Table 3
Modulated genes lying in chromosomal regions associated with CMT
Accession no. Symbol Name Human chromosome Disease
BG073735 Col2a1 procollagen, type I, alpha-2 12q13 CMT2G
AA461746 Ssg1-pending steroid sensitive gene 1 3q13 HMSNP
AA047966 Fhl1 four and a half LIM domains 1 Xq26 CMT2X
AA240923 Gpc3 glypican 3 Xq26 CMT2X
BG073597 Tnnt1 troponin T1 19q13 AR-CMT2B2
AA002733 Myoz1 myozenin 1 10q22 HMSNR
AA220458 Gpsn2 glycoprotein, synaptic 2 19p13 DI-CMTB
AA013851 Rap1ga1 Rap1, GTPase-activating protein 1 19p13 DI-CMTB
BG073341 Rsdr1-pending retinal short-chain dehydrogenase/reductase 1 1p36 DI-CMTC
HMSN-P: hereditary motor and sensory neuropathy, proximal type; HMSN-R: hereditary motor and sensory neuropathy, russe type; DI-CMTB: Charcot–
Marie–Tooth neuropathy, dominant intermediate type B; DI-CMTC: Charcot–Marie–Tooth neuropathy, dominant intermediate type C.
T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714708
Furthermore, to evaluate the ability of pmp22 overexpressing
Schwann cells to produce cntf in the absence of contaminating
cells and independently to the axon, we analyzed cntf expression in
short-term Schwann cells cultures. Again, real-time PCR was used
to quantify cntf mRNA in normal and transgenic Schwann cells
isolated from 30-day-old sciatic nerves (Nobbio et al., 2004). We
observed that mRNA levels of cntf were reduced in hemizygous
(0.7 F 0.06) and homozygous (0.53 F 0.07) transgenic cultures
compared to normal ones.
Using an ELISA method we also quantified the levels of cntf in
sciatic nerves and in Schwann cells cultures. We observed a
significantly (P b 0.01) lower concentration of protein in sciatic
nerves from homozygous (4.3 F 1.2 pg/Ag of total proteins) and
hemizygous (14.18 F 1.7 pg/Ag of total proteins) pmp22
overexpressing nerves compared to normal controls (64.4 F 19.2
pg/Ag of total proteins). Moreover, we found a significant (P b
0.01) decrease of cntf in homozygous (12.8 F 1.3 pg/Ag of total
proteins) and hemizygous (18.3 F 0.3 pg/Ag of total proteins)
purified Schwann cells compared to normal ones (47.9F 0.8 pg/Agof total proteins). We also quantified cntf protein in the culture
medium, observing that in homozygous (2.6 F 0.6 pg/100 Al) andhemizygous (2.15 F 0.4 pg/100 Al) Schwann cells cntf releasing
was significantly (P b 0.01) reduced compared to control cultures
(11.79 F 2.7 pg/100 Al). These results strongly support the
hypothesis that pmp22 overexpressing Schwann cells are primarily
unable to produce and release normal levels of cntf.
Discussion
We used cDNA microarrays technology to perform a complete
gene expression profiling in sciatic nerves of transgenic rats
overexpressing pmp22. In fact, although the genetic cause of
Table 4
Differential gene expression by cDNA microarrays (expressed as the average of log
intensity and ribosomal RNA 28S)
Gene Normal Heterozygous
cd9 1.268 F 0.23 0.442 F 0.09
cntf 1.138 F 0.32 0.57 F 0.19
ccnd1 0.16 F 0.02 0.39 F 0.11
ccnd3 0.11 F 0.007 0.26 F 0.06
Animals tested 4 4
RT-PCR tests were performed on four animals independently for each condition.
* Statistical significance ( P b 0.05) obtained with ANOVA test.
CMT1A is well known, and an abnormal expression of a few genes
coding for myelin proteins and Schwann cells differentiation
markers has been previously described in pmp22 overexpressing
rats (Niemann et al., 2001), little is known about the pathomechan-
isms underlying the disease and the effect of pmp22 over-
expression on the transcriptional activity in the peripheral
nervous system. We analyzed gene expression in nerves from
30-day-old rats, because at this age homozygous rats are easily
distinguishable from hemizygous ones (Sereda et al., 1996) and
their nerves show clear clinical, neuropathological, and neuro-
physiological abnormalities (Grandis et al., 2004).
Our microarray experiment first indicates that the increased
dosage of pmp22 induces a general down-regulation of gene
expression in sciatic nerves. This observation may represent a
generic damage to the nerve tissue by the genetic modification
more than a specific consequence of pmp22 overexpression. We
found only a minority of genes that were up-regulated in pmp22
transgenic rats. Among these, cyclin D1 and cyclin D3 mRNAs
seem to be particularly interesting. D-type cyclins are required for
the initial steps in cell division and nuclear import is crucial for the
function of cyclin D1 in promoting cell proliferation. Myelinating
Schwann cells express cyclin D1 in the perinuclear region, but after
axons are severed, cyclin D1 is strongly up-regulated in parallel
with Schwann cell proliferation and translocates into Schwann cell
nuclei (Atanasoski et al., 2001). In pmp22 overexpressing rats an
up-regulation in cyclin D1 expression was already found in the
Schwann cells nucleus (Atanasoski et al., 2002). A cyclin D1 up-
regulation was also reported in peripheral nerves after axotomy
(Kubo et al., 2002). Since the pmp22 gene shows homology to the
growth arrest-specific gene gas3, an effect of pmp22 over-
expression has been proposed on Schwann cells proliferation, but
results are contrasting in this regard. In vitro experiments on
PMP22 overexpressing human Schwann cells show a decreased
arithms) and semiquantitative RT-PCR (expressed as ratio between the band
Homozygous Microarray ratio
homozygous/norma
0.298 F 0.04* 0.36
0.084 F 0.01* 0.075
* 0.717 F 0.11* 3.8
* 0.45 F 0.15* 2.77
4
l

Fig. 4. RT-PCR analysis of neurotrophic factors expression. Representative gels from control (1), hemizygous (2), and homozygous (3) rat sciatic nerves. No
significant differences were found, between groups, in mRNA expression of (a) BDNF (0.54 F 0.12 n. 4 vs. 0.77 F 0.26 n. 4 vs. 0.62 F 0.028 n. 4; n.s.); (b)
GDNF (0.05 F 0.04 n. 4 vs. 0.34 F 0.18 n. 4 vs. 0.21 F 0.08 n. 4; n.s.); (c) NGF (0.31 F 0.12 n. 4 vs. 0.30 F 0.08 n. 4 vs. 0.28 F 0.12 n. 4; n.s.); (d) NT3
(0.75F 0.15 n. 4 vs. 0.88F 0.08 n. 4 vs. 0.5F 0.2 n. 4; n.s.). On the contrary, CNTF transcript levels (e) were significantly higher in control nerves compared
to homozygous ones (1.138 F 0.32 n. 4 vs. 0.57 F 0.19 n. 4 vs. 0.084 F 0.01 n. 4; P b 0.01).
T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714 709
proliferation (Hanemann et al., 1997). However, in a CMT1A
animal model, a continued Schwann cell proliferation into
adulthood was observed (Magyar et al., 1996). Our results,
although insufficient to make any conclusion, are in keeping with
the previous study (Atanasoski et al., 2002) showing that pmp22
overexpressing Schwann cells proliferate after demyelination.
Moreover, we found a deregulation of other genes involved in
negative modulation of cell proliferation, suggesting that in pmp22
overexpressing nerves impairment in the cell cycle regulation
affects the resident proliferating cellular populations. Finally,
among the up-regulated genes we identified several transcription
factors (Idb2, Sox4, Scip). Further studies are needed to elucidate a
possible role of these nuclear proteins in the Schwann cells
biology.
We found that a large group of down-regulated genes is
involved in lipidic and glucidic metabolism. Impairment in the
cholesterol biosynthesis pathways has been shown in tellurium-
induced neuropathy (Harry et al., 1989), and the consequent lack of
cholesterol destabilizes myelin (Wagner-Recio et al., 1991). Many
genes directly involved in lipid metabolism have been found
regulated by microarray analysis during myelination (Nagarajan et
al., 2002; Verheijen et al., 2003), underscoring the relevance of
Schwann cells cholesterol synthesis in myelination. Finding a
down-regulation of genes coding for enzymes involved in
cholesterol biosynthesis is consistent with the absence of myelin
in homozygous transgenic rats.
Genes coding for cytoskeleton components were also down-
regulated in homozygous transgenic nerves. It has been shown that
pmp22 overexpression affects the differentiation and the spreading/
adhesion properties of Schwann cells (Brancolini et al., 2000;
Magyar et al., 1996). Finding a diffuse down-regulation of
cytoskeleton elements further supports the idea that overexpression
of pmp22 induces changes in the ability of a Schwann cell to
change its shape and switch on the myelination process (Nobbio et
al., 2004). As Schwann cells are the highly predominant population
in peripheral nerves and morphological abnormalities of other
components of sciatic nerve, like axons, are not present at this age
(Grandis et al., 2004), changes in transcriptional profile have to be
considered mainly caused by the imbalance in Schwann cell
function.
PMP22 is a tetraspan membrane protein (Bolin et al., 1997;
Magyar et al., 1997; Taylor et al., 1995). Our microarray
experiment shows a down-regulation of Cd9 mRNA, another
tetraspan cell surface protein expressed in the peripheral nervous
system by Schwann cells (Banerjee and Patterson, 1995; Kaprie-
lian et al., 1995; Tole and Patterson, 1993). Down-regulation of
cd9 could represent a defensive mechanism carried out by
Schwann cells to counterbalance pmp22 overexpression. Gene
compensation mechanisms occur in different models (Bowe et al.,
2002; Groussin et al., 2000). We propose a similar mechanism to
explain Cd9 loss in pmp22 overexpressing nerves. Cd9 was shown
to be associated with h3, h6, and h1 integrins (Hadjiargyrou et al.,
1996), and perturbation of its expression alters Schwann cell
adhesion, proliferation, and migration as well as neurite outgrowth
in sympathetic neurons (Anton et al., 1995; Hadjiargyrou and
Patterson, 1995). Therefore, down-regulation of Cd9 could itself
account for some of the shaping defects previously observed in
pmp22 overexpressing Schwann cells (Brancolini et al., 2000;
Nobbio et al., 2004).
Another group of down-regulated genes was represented by
extracellular matrix components such as collagens. In literature,
contrasting results have been reported about expression of
collagens in hereditary neuropathies of the CMT type. In human
and experimental CMT1A an up-regulation of collagen types I, III,
IV, V, and VI and an increased collagen deposition were,
respectively, observed (Palumbo et al., 2002; Robaglia-Schlupp
et al., 2002). Instead, in the sciatic nerves of Trembler-J mouse,
which carries a point mutation in the pmp22 gene, a reduction of
collagen IV was reported (Misko et al., 2002). Discrepancies in the
animal species and in the disease stage may account for these
differences. However, further studies are needed to elucidate the
role of collagens in the development of CMT1A.
The most interesting result of our cDNA microarrays experi-
ment is the profound down-regulation of the cntf gene observed in
homozygous CMT1A rats. Schwann cells are sources of cytokines
and neurotrophins that can affect the survival, differentiation, and

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714710
growth of neurons (Bunge, 1993). Cntf is one of the most
important neurotrophic factors produced by myelinating Schwann
cells (Sendtner et al., 1990) and is also able to enhance myelin
formation (Stankoff et al., 2002). Its levels dramatically fall in the
early stages of Wallerian degeneration (Sendtner et al., 1990). A
consistent reduction of the cntf transcript was also found in sciatic
nerves from the Trembler mouse (Friedman et al., 1992).
Accordingly, we found a dramatic decrease of the cntf mRNA
and protein in pmp22 overexpressing nerves. Interestingly, cntf is
the only neurotrophic factor to show a down-regulation, as
expression of bdnf, gdnf, ngf, and nt3 does not change, as
confirmed by RT-PCR analysis. We also found a complete absence
of the CNTF transcript in sural nerve biopsies from CMT1A
patients. Several reports described a reduction of CNTF expression
in different neuropathies (Ito et al., 2001; Lee et al., 1996;
Yamamoto et al., 2001, 2002), but a total absence of this
neurotrophic factor in peripheral nerves was never found. Finding
a normal number of Schwann cells in pmp22 overexpressing
sciatic nerves, as previously observed in human CMT1A sural
biopsies (Hanemann et al., 1997), excludes that the extreme
reduction of CNTF expression is due to a loss of Schwann cells.
Taken together, these results suggest that both in human and
experimental CMT1A there is a deficiency in CNTF support from
the Schwann cells to the axon, which could contribute to the
development of the axonal atrophy, observed in the late stages of
CMT1A (Sahenk et al., 1999; Sancho et al., 1999). In agreement
with this hypothesis, we also showed that pmp22 overexpressing
Schwann cells, besides expressing lower level of cntf transcript,
also produce and release low levels of the protein. This happens
both in the presence and absence of the axon, suggesting a primary
inability of transgenic Schwann cells in the production of this
specific neurotrophic factor.
In conclusion, our study provides the first comprehensive list of
genes showing altered expression levels in sciatic nerves of pmp22
transgenic rats. This gene expression profile suggests that pmp22
overexpression deeply alters the delicate balance regulating the
Schwann cells proliferation and differentiation, as showed by the
observation of a severe derangement in the expression of genes
involved in cell cycle support and in the cytoskeletal organization.
Moreover, finding reduced levels of cntf in CMT1A nerves and
Schwann cell cultures suggests that the trophic support offered by
pmp22 overexpressing Schwann cells to the axon is highly
insufficient.
Finally, since some of the modulated genes map into
chromosomal regions linked to other types of hereditary neuro-
pathies, our results might be of help in future mutational analysis to
reveal new disease responsible genes.
Experimental methods
Animal model
We used 30-day-old homozygous transgenic rats overexpress-
ing pmp22 (Sereda et al., 1996). Although the hemizygous
condition is a more appropriate model of CMT1A, we compared
homozygous rats with the normal controls, as we were interested in
studying the general consequences of pmp22 overexpression on
sciatic nerve mRNA profile. Next to homozygous animals, we also
used hemizygous ones, when we looked at selected genes by
semiquantitative RT-PCR, real-time PCR, and ELISA. Rearing
conditions were consistent with the guidelines of the Italian Health
Ministry relating to the use and storage of transgenic organisms.
RNA extraction
Total RNA was obtained from sciatic nerves using standard
methods to perform cDNA microarrays experiments. Eight sciatic
nerves from sex-matched homozygous animals and four sciatic
nerves from normal littermates were homogenized in TriPure
Isolation Reagent (Boehringer Mannheim, Germany), with a
Polytron homogenizer (Kinematica Srl, Italy) for 15 s. An equal
volume of 70% ethanol was added to the watering phases deriving
from chloroform extraction, and samples were transferred to
Qiagen Rneasy Mini Kit columns (Qiagen SpA, Germany). The
extraction proceeded according to the manufacturer instructions.
Digestion of contaminant DNA was performed in the columns,
using an RNAse-free DNAse (Qiagen SpA, Germany). Total RNA
extraction was repeated two times to perform the microarray
hybridization twice. The quality and the concentration of RNA
were checked with NanoDrop Spectophotometer ND-1000 (Nano
Drop Technologies Inc., Delaware, USA).
cDNA microarray
The mouse gene set consisted of five separate microarrays
containing a total of 21,492 cDNA fragments from the 6K
collection of Incyte (Mouse Gem I, Incyte, USA) and from the
15K collection of the National Institute of Aging (http://
lgsun.grc.nia.nih.gov). On each of the five slides, on average
4300 cDNAs were spotted in duplicate, distant from each other, on
type VIIstar silane-coated slides (Amersham BioSciences, Buck-
inghamshire, UK). The cDNA inserts were PCR amplified using
M13 primers, purified with MultiScreen-PCR plate (Millipore,
Belgium), and arrayed in 50% DMSO on Type VII silane-coated
slides (Amersham BioSciences, Buckinghamshire, UK) using a
Molecular Dynamics Generation III printer (Amersham BioScien-
ces). Slides were blocked in 2� SSPE, 0.2% SDS for 30 min at
258C.A minimum of 5 Ag total RNA was linearly amplified using in
vitro transcription as previously described (Puskas et al., 2002).
Briefly, RNA was reverse transcribed to double-stranded cDNA
using an anchored oligo-dT + T7 promoter (5V-GGCCAGT-GAATTGTAATACGACTCACTATAGGGAGGCGG-T24(ACG)-
3V) (Eurogentec, Belgium). From this cDNA, RNA was produced
via T7-in vitro transcriptase until an average yield of 10–30 Agamplified RNA (aRNA). From the aRNA, 5 Ag was labeled by
reverse transcription using random nonamer primers (Genset,
Paris, France), 0.1 mM d(G/T/A)TPs, 0.05 mM dCTP (Amersham
BioSciences, UK), 0.05 mM Cy3-dCTP or Cy5-dCTP (Amersham
BioSciences, UK), 1� first strand buffer, 10 mM DTT, and 200
Units of SuperScript II (Invitrogen, Belgium) in 20 Al total volume.
The RNA and primers were denatured at 758C for 5 min and
cooled on ice before adding the remaining reaction components.
After 2 h incubation at 428C, mRNA was hydrolyzed in 250 mM
NaOH for 15 min at 378C. The sample was neutralized with 10 Alof 2 M MOPS and purified with Qiaquick (Qiagen, Germany).
The probes were resuspended in 210 Al hybridization solution
containing 50% formamide, 1� hybridization buffer (Amersham
BioSciences, UK), 0.1% SDS, and 60 Ag/ml poly-dT. Hybrid-
ization and post-hybridization washing were performed at 458Cusing an automated slide processor (ASP; Amersham BioSciences,

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714 711
UK). Post-hybridization washing was performed in 1� SSC, 0.1%
SDS, followed by 0.1� SSC, 0.1% SDS, and 0.1� SSC. The
complete ASP program can be downloaded from www.
microarrays.be (/technology/protocols). Arrays were scanned at
532 and 635 nm using a Generation III scanner (Amersham
BioSciences, UK). Image analysis was performed with ArrayVi-
sion (Imaging Research Inc, Ontario, Canada). Two biological
repeats and each hybridization were repeated in a dye swap.
Spot intensities were measured as artifact removed total
intensities, subtracted with the local background (sARVol), and
filtered based on two standard deviations above background. For
each gene, ratios of red (Cy-5) over green (Cy-3) intensities (I)
were calculated and normalized via a Lowess Fit of the log2 ratios
[log2(Icy-5 / Icy-3)] over the log2 total intensity [log2(Icy-5 � Icy-
3)]. Mean ratios were calculated from the duplicate spots, and only
values with covariance (CV) b0.5 were further taken into account.
Normalized ratios that were statistical significant using a two-tailed
t test (5% level) between the dye-swap repeat and higher than 1 or
lower than �1 (log2 scale) were considered differentially
expressed.
Semiquantitative reverse transcriptase PCR
To confirm the accuracy of cDNA microarrays, semiquantita-
tive RT-PCR was performed on selected genes, known to be
important in the biology of Schwann cells. Specific oligonucleotide
primer pairs (Table 5) were designed using Primer 3 software
(http://www-genome.wi.mit.edu/cgi-bin/primer/primer3_www.cgi)
in order to amplify fragments of 180–250 bp in length. To avoid
the amplification of contaminant genomic DNA, we selected
primers lying on distinct exons. We performed RT-PCR on sciatic
nerves from four different rats for genetic condition, each of them
Table 5
Oligonucleotide pairs used in semiquantitative RT-PCR experiments
Gene
Forward primers (5V–3V)Reverse primers (5V–3V)
ccnd1
gcgtaccctgacaccaatct
gaaccggtccaggtagttca
ccnd3
tgcatctatacggaccaagctat
aggtctgagcatgctttttga
cntf
gcaaacacctctgacccttc
acggtaagcctggaggttct
cd9
tgggattgttcttcggattc
gctatgccacagcagttcaa
bdnf
acttttgagcacgtgatcgaaga
ggtagttcggcattgcgagt
gdnf
ggacgggactctaagatgaagtt
cgtcatcaaactggtcaggata
ngf
cacaggagcaagcgctcatc
acacacacgcaggctgtatctatc
Trkb
cgacactcaggatttgtattgcc
tccgtgtgattggtgacgtgtatt
coming from a different breeding. Results were expressed as the
mean of the four animals.
First-strand cDNA was synthesized from 250 ng of total RNA
in a 30 Al reaction using the Superscript system (Invitrogen Srl,
Italy). Semiquantitative amplification was performed from 10 Al ofthe first strand reaction. The product of the endogenous 18S RNA
served as an internal standard. Amplification following hot start (5
min at 958C) was carried out for 20 cycles consisting of 1 min at
958C, 1 min at 568C, and 1 min at 728C; an additional extension
time of 10 min was added. Preliminary experiments were
conducted to ensure that measurements were performed in the
exponential phase of amplification process and the expression of
the reference gene was uniform in every condition. PCR products
were analyzed on 2% agarose gels, and band intensity was
measured on a Gel Doc 1000 image system (Bio-Rad, Hercules,
CA). Results were expressed as ratio between specific band
intensity and 18S RNA band.
Real-time PCR
cDNA was prepared from sciatic nerves of 30-day-old pmp22
overexpressing rats and their normal littermates and from primary
Schwann cell cultures. Moreover, we analyzed human sural nerve
biopsies from control subjects and patients with CMT1A. Total
RNA was extracted as previously described from an independent
pool of at least nine animals per genetic condition and from three
human subjects per group. Relative quantification of expression of
human and rat CNTF was performed using an ABI PRISM 7700
Sequence Detection System with SYBR green chemistry (Applied
Biosystem) as described (http://www.docs.appliedbiosystems.com/
pebiodocs/04303859.pdf). Dissociation curve analysis was per-
formed using Dissociation Curve 1.0 software (ABI) for each PCR
reaction to detect and eliminate possible primer–dimer artefacts.
Oligonucleotides were selected to amplify a fragment containing
sequences from two adjacent exons in order to avoid contaminating
genomic DNA amplification. To standardize the amount of cDNA
in each reaction, we measured the amount of 18S rRNA, which
showed no variation in expression in both human and animal
samples. The comparative cycle threshold (Ct) method (User
Bulletin 2, 1997; Applied Biosystems, Foster City, CA) was used
to analyze the data by generating relative values of the amount of
target cDNA. Relative quantification for cntf gene, expressed as
fold variation over control, was calculated by the DDCt method,
using control samples as calibrators.
Human sural nerves biopsy
Archived sural nerve biopsies, obtained for diagnostic purpose
when the genetic diagnosis of CMT1A was not yet available, were
used for this study. Negative controls are nerves that were biopsied
in the suspect of a peripheral neuropathy but showed to be normal
after morphological and morphometrical evaluation at the light and
electron microscopy levels. Sural nerves were biopsied under local
anesthesia at midcalf. Samples were snap frozen in liquid nitrogen
and stored at �808C until used.
Immunohistochemistry
Rat sciatic nerves were fixed in 4% paraformaldehyde in
sodium cacodylate 0.025 M for 18 h at room temperature and
embedded in paraffin. Sections of 5 Am were digested with trypsin

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714712
for 15 min at 378C, then incubated with 10% normal goat serum in
PBS for 15 min at room temperature. A mouse anti-rat S100
monoclonal antibody (Sigma-Aldrich, Saint Louis, Missouri, USA)
was used 1:400 in PBS containing 1% normal goat serum and
incubated over night at 48C in humid chamber. Sections were
washed in PBS and incubated with biotinylated anti-mouse
immunoglobulins (Biogenex Laboratories, San Ramon, CA) for
20 min at RT, then with peroxidase-conjugated streptavidin
(Biogenex Laboratories, San Ramon, CA). The peroxidase activity
was demonstrated using the DAB substrate (Biogenex Laborato-
ries, San Ramon, CA). For each nerve fascicle three frames,
randomly selected at a 20� magnification to cover at least 80% of
the fascicle, were digitized and stored, using the Pro Plus Imaging
System (Immagini e Computer, Rho, Italy). S100-positive cells
were counted. We analyzed three sciatic nerves per genetic
condition. Results are expressed as number of Schwann cells/mm2.
Primary Schwann cells cultures
Cell cultures were established from sciatic nerves of 30-day-old
pmp22 transgenic homozygous and hemizygous rats according to a
technique optimized for adult animals (Nobbio et al., 2004). Wild
type rats from the corresponding genetic background were used as
controls. Cntf expression and release were analyzed in Schwann
cell cultures treated with serum-free medium containing 0.1%
bovine serum albumin for 48 h. The supernatants were collected
and frozen and, at the same time, Schwann cells were carefully
rinsed with sterile phosphate-buffered saline (PBS), scraped from
the culture dish, recovered through centrifugation, and immediately
frozen.
ELISA quantification
Sciatic nerves and Schwann cells, scraped from the culture
dish, were mechanically disrupted in PBS containing 0.1 mM
PMSF. The extracts were obtained after two 15-min centrifuga-
tions at 100,000 � g. Protein concentration in the supernatants
was measured using Biorad Protein Detection kit (Bio-Rad
Laboratories, Srl, Milan). Cntf content was determined by ELISA
using a Rat Cntf DuoSet kit (R&D Systems, Inc, MN) according
to manufacturer instructions. Briefly, mouse-anti-rat cntf capture
antibody was coated at 2 Ag/ml into 96-well immunoassay plates
(Corning Incorporated) overnight at room temperature. Then,
plates were blocked with 1% BSA, 5% sucrose in phosphate-
buffered saline (PBS, pH 7.4) for at least 1 h at room
temperature, and washed with 0.01% Tween 20 in PBS (PBST).
Sciatic nerve and Schwann cells extracts (10 Ag/well) were added
to the plates and incubated at room temperature (RT) for 2 h. To
quantify the cntf released in cultures medium, we tested 100 Al ofthe serum-free medium from Schwann cell cultures. Plates were
washed three times with PBST between each assay step.
Biotynilated goat anti-rat cntf detection antibody, diluted to 200
ng/ml with 1% BSA in PBST, was incubated in wells for 1 h at
RT. The assay was developed with tetramethylbenzidine in
phosphate buffer pH 6.0 and 0.1% H2O2 (R&D Systems, Inc,
MN). Optical densities at 450 nm were measured using a
spectrophotometric plate reader (Metertech). For sciatic nerves
and Schwann cells extracts results were expressed as ratio
between cntf and total proteins concentration. For the quantifi-
cation of released cntf we normalized cntf concentration by the
number of Schwann cells.
Statistical analysis
Results were evaluated using a one-way analysis of variance
(ANOVA), followed by a Dunnet post-test to separately compare
the pathological conditions with the normal control.
Acknowledgments
We thank Dr. Giulio Palmisano for help in RNA extraction.
This work was financially supported by the European Science
Foundation (ESF) Integrated Aproaches for Functional Genomics
Program (L.N.); by Telethon contract GP02169 2002 (A.S.); by
FISM 2001/R/59 (A.S.); by FIRB RBAUO1KJE4/002 (M.A.);
and by COFIN-MIUR 2002 (M.A.). K.V. is a postdoctoral fellow
of the Fund for Scientific Research, FWO-Flanders, and N.V. is
received a PhD fellowship of the Institute for Science and
Technology, IWT, Belgium.
References
Anton, E.S., Hadjiargyrou, M., Patterson, P.H., Matthew, W.D., 1995. CD9
plays a role in Schwann cell migration in vitro. J. Neurosci. 15 (1 Pt. 2),
584–595.
Atanasoski, S., Shumas, S., Dickson, C., Scherer, S.S., Suter, U., 2001.
Differential cyclin D1 requirements of proliferating Schwann cells
during development and after injury. Mol. Cell. Neurosci. 18 (6),
581–592.
Atanasoski, S., Scherer, S.S., Nave, K.A., Suter, U., 2002. Proliferation of
Schwann cells and regulation of cyclin D1 expression in an animal
model of Charcot–Marie–Tooth disease type 1A. J. Neurosci. Res. 67
(4), 443–449.
Baechner, D., Liehr, T., Hameister, H., Altenberger, H., Grehl, H., Suter, U.,
Rautenstrauss, B., 1995. Widespread expression of the peripheral
myelin protein-22 gene (PMP22) in neural and non-neural tissues
during murine development. J. Neurosci. Res. 42 (6), 733–741.
Banerjee, S.A., Patterson, P.H., 1995. Schwann cell CD9 expression is
regulated by axons. Mol. Cell. Neurosci. 6 (5), 462–473.
Berciano, J., Combarros, O., 2003. Hereditary neuropathies. Curr. Opin.
Neurol. 16 (5), 613–622.
Bolin, L.M., McNeil, T., Lucian, L.A., DeVaux, B., Franz-Bacon, K.,
Gorman, D.M., Zurawski, S., Murray, R., McClanahan, T.K., 1997.
HNMP-1: a novel hematopoietic and neural membrane protein differ-
entially regulated in neural development and injury. J. Neurosci. 17
(14), 5493–5502.
Bowe, D.B., Kenney, N.J., Adereth, Y., Maroulakou, I.G., 2002. Suppres-
sion of Neu-induced mammary tumor growth in cyclin D1 deficient mice
is compensated for by cyclin E. Oncogene 21 (2), 291–298.
Brancolini, C., Edomi, P., Marzinotto, S., Schneider, C., 2000. Exposure at
the cell surface is required for gas3/PMP22 To regulate both cell death
and cell spreading: implication for the Charcot–Marie–Tooth type 1A
and Dejerine–Sottas diseases. Mol. Biol. Cell 11 (9), 2901–2914.
Bunge, R.P., 1993. Expanding roles for the Schwann cell: ensheathment,
myelination, trophism and regeneration. Curr. Opin. Neurobiol. 3 (5),
805–809.
Cameron, A.A., Vansant, G., Wu, W., Carlo, D.J., Ill, C.R., 2003.
Identification of reciprocally regulated gene modules in regenerating
dorsal root ganglion neurons and activated peripheral or central nervous
system glia. J. Cell. Biochem. 88 (5), 970–985.
Costigan, M., Befort, K., Karchewski, L., Griffin, R.S., D’Urso, D.,
Allchorne, A., Sitarski, J., Mannion, J.W., Pratt, R.E., Woolf, C.J.,
2002. Replicate high-density rat genome oligonucleotide microarrays
reveal hundreds of regulated genes in the dorsal root ganglion after
peripheral nerve injury. BMC Neurosci. 3 (1), 16.

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714 713
De Sandre-Giovannoli, A., Chaouch, M., Kozlov, S., Vallat, J.M., Tazir, M.,
Kassouri, N., Szepetowski, P., Hammadouche, T., Vandenberghe, A.,
Stewart, C.L., Grid, D., Levy, N., 2002. Homozygous defects in
LMNA, encoding lamin A/C nuclear-envelope proteins, cause autoso-
mal recessive axonal neuropathy in human (Charcot–Marie–Tooth
disorder type 2) and mouse. Am. J. Hum. Genet. 70 (3), 726–736.
Draghici, S., Khatri, P., Bhavsar, P., Shah, A., Krawetz, S., Tainsky, M.A.,
2003. Onto-Tools, The toolkit of the modern biologist: Onto-Express,
Onto-Compare, Onto-Design and Onto-Translate. Nucleic Acids Res.
31 (13), 3775–3781.
Dyck, P.J., Chance, P., Lebo, R., Carney, J.A., 1993. Hereditary motor and
sensory neuropathies. In: Dyck, P.J., Thomas, P.K., Griffin, S.W., Low,
P.A., Poduslo, J.F. (Eds.), Peripheral Neuropathy, 3rd ed. WB Saunders,
Philadelphia, pp. 1094–1136.
Friedman, B., Scherer, S.S., Rudge, J.S., Helgren, M., Morrisey, D.,
McClain, J., Wang, D.Y., Wiegand, S.J., Furth, M.E., Lindsay, R.M., et
al., 1992. Regulation of ciliary neurotrophic factor expression in
myelin-related Schwann cells in vivo. Neuron 9 (2), 295–305.
Grandis, M., Leandri, M., Vigo, T., Cilli, M., Sereda, M.W., Gherardi, G.,
Benedetti, L., Mancardi, G., Abbruzzese, M., Nave, K.A., Nobbio, L.,
Schenone, A., 2004. Early abnormalities in sciatic nerve function and
structure in a rat model of Charcot–Marie–Tooth type 1A disease. Exp.
Neurol. 190 (1), 213–223.
Groussin, L., Massias, J.F., Bertagna, X., Bertherat, J., 2000. Loss of
expression of the ubiquitous transcription factor cAMP response
element-binding protein (CREB) and compensatory overexpression of
the activator CREMtau in the human adrenocortical cancer cell line
H295R. J. Clin. Endocrinol. Metab. 85 (1), 345–354.
Guilbot, A., Williams, A., Ravise, N., Verny, C., Brice, A., Sherman, D.L.,
Brophy, P.J., LeGuern, E., Delague, V., Bareil, C., Megarbane, A.,
Claustres, M., 2001. A mutation in periaxin is responsible for CMT4F,
an autosomal recessive form of Charcot-Marie-Tooth disease. Hum.
Mol. Genet. 10 (4), 415–421.
Hadjiargyrou, M., Patterson, P.H., 1995. An anti-CD9 monoclonal antibody
promotes adhesion and induces proliferation of Schwann cells in vitro.
J. Neurosci. 15, 574–583.
Hadjiargyrou, M., Kaprielian, Z., Kato, N., Patterson, P.H., 1996.
Association of the tetraspan protein CD9 with integrins on the surface
of S-16 Schwann cells. J. Neurochem. 67 (6), 2505–2513.
Hanemann, C.O., Muller, H.W., 1998. Pathogenesis of Charcot–Marie–
Tooth 1A (CMT1A) neuropathy. Trends Neurosci. 21 (7), 282–286.
Hanemann, C.O., Gabreels-Festen, A.A., Stoll, G., Muller, H.W., 1997.
Schwann cell differentiation in Charcot–Marie–Tooth disease type 1A
(CMT1A): normal number of myelinating Schwann cells in young
CMT1A patients and neural cell adhesion molecule expression in onion
bulbs. Acta Neuropathol. (Berl) 94 (4), 310–315.
Harry, G.J., Goodrum, J.F., Bouldin, T.W., Wagner-Recio, M., Toews,
A.D., Morell, P., 1989. Tellurium-induced neuropathy: metabolic
alterations associated with demyelination and remyelination in rat
sciatic nerve. J. Neurochem. 52 (3), 938–945.
Huxley, C., Passage, E., Manson, A., Putzu, G., Figarella-Branger, D.,
Pellissier, J.F., Fontes, M., 1996. Construction of a mouse model of
Charcot-Marie-Tooth disease type 1A by pronuclear injection of human
YAC DNA. Hum. Mol. Genet. 5 (5), 563–569.
Inoue, K., Dewar, K., Katsanis, N., Reiter, L.T., Lander, E.S., Devon, K.L.,
Wyman, D.W., Lupski, J.R., Birren, B., 2001. The 1.4-Mb CMT1A
duplication/HNPP deletion genomic region reveals unique genome
architectural features and provides insights into the recent evolution of
new genes. Genome Res. 11 (6), 1018–1033.
Ito, Y., Yamamoto, M., Mitsuma, N., Li, M., Hattori, N., Sobue, G., 2001.
Expression of mRNAs for ciliary neurotrophic factor (CNTF), leukemia
inhibitory factor (LIF), interleukin-6 (IL-6), and their receptors (CNTFR
alpha, LIFR beta, IL-6R alpha, and gp130) in human peripheral
neuropathies. Neurochem. Res. 26 (1), 51–58.
Kaprielian, Z., Cho, K.O., Hadjiargyrou, M., Patterson, P.H., 1995. CD9, a
major platelet cell surface glycoprotein, is a ROCA antigen and is
expressed in the nervous system. J. Neurosci. 15 (1 Pt. 2), 562–573.
Kubo, T., Yamashita, T., Yamaguchi, A., Hosokawa, K., Tohyama, M.,
2002. Analysis of genes induced in peripheral nerve after axotomy
using cDNA microarrays. J. Neurochem. 82 (5), 1129–1136.
Lee, D.A., Gross, L., Wittrock, D.A., Windebank, A.J., 1996. Localization
and expression of ciliary neurotrophic factor (CNTF) in postmortem
sciatic nerve from patients with motor neuron disease and diabetic
neuropathy. J. Neuropathol. Exp. Neurol. 55 (8), 915–923.
Magyar, J.P., Martini, R., Ruelicke, T., Aguzzi, A., Adlkofer, K., Dembic,
Z., Zielasek, J., Toyka, K.V., Suter, U., 1996. Impaired differentiation
of Schwann cells in transgenic mice with increased PMP22 gene
dosage. J. Neurosci. 16 (17), 5351–5360.
Magyar, J.P., Ebensperger, C., Schaeren-Wiemers, N., Suter, U., 1997.
Myelin and lymphocyte protein (MAL/MVP17/VIP17) and plasmolipin
are members of an extended gene family. Gene 189 (2), 269–275.
Misko, A., Ferguson, T., Notterpek, L., 2002. Matrix metalloproteinase
mediated degradation of basement membrane proteins in Trembler
J neuropathy nerves. J. Neurochem. (83), 885–894.
Nagarajan, R., Svaren, J., Le, N., Araki, T., Watson, M., Milbrandt, J.,
2001. EGR2 mutations in inherited neuropathies dominant-negatively
inhibit myelin gene expression. Neuron 30 (2), 355–368.
Nagarajan, R., Le, N., Mahoney, H., Araki, T., Milbrandt, J., 2002.
Deciphering peripheral nerve myelination by using Schwann cell
expression profiling. Proc. Natl. Acad. Sci. U. S. A. 99 (13),
8998–9003.
Niemann, S., Sereda, M.W., Suter, U., Griffiths, I.R., Nave, K.A., 2001.
Uncoupling of myelin assembly and Schwann cell differentiation by
transgenic overexpression of peripheral myelin protein 22. J. Neurosci.
20 (11), 4120–4128.
Nobbio, L., Vigo, T., Abruzzese, M., Levi, G., Brancolini, C., Mantero, S.,
Grandis, M., Benedetti, L., Mancardi, G.L., Schenone, A., 2004.
Impairment of PMP22 transgenic Schwann cells differentiation in
culture: implications for Charcot-Marie-Tooth type 1A disease. Neuro-
biol. Dis. 16 (1), 263–273.
Palumbo, C., Massa, R., Panico, M.B., Di Muzio, A., Sinibaldi, P.,
Bernardi, G., Modesti, A., 2002. Peripheral nerve extracellular matrix
remodeling in Charcot-Marie-Tooth type I disease. Acta Neuropathol.
(Berl) 104 (3), 287–296.
Perea, J., Robertson, A., Tolmachova, T., Muddle, J., King, R.H., Ponsford,
S., Thomas, P.K., Huxley, C., 2001. Induced myelination and
demyelination in a conditional mouse model of Charcot–Marie–Tooth
disease type 1A. Hum. Mol. Genet. 10 (10), 1007–1018.
Puskas, L.G., Zvara, A., Hackler Jr., L., Micsik, T., van Hummelen, P.,
2002. Production of bulk amounts of universal RNA for DNA
microarrays. BioTechniques 33(4), 898-900, 902, 904.
Roa, B.B., Garcia, C.A., Pentao, L., Killian, J.M., Trask, B.J., Suter, U.,
Snipes, G.J., Ortiz-Lopez, R., Shooter, E.M., Patel, P.I., et al., 1993.
Evidence for a recessive PMP22 point mutation in Charcot–Marie–
Tooth disease type 1A. Nat. Genet. 5 (2), 189–194.
Robaglia-Schlupp, A., Pizant, J., Norreel, J.C., Passage, E., Saberan-
Djoneidi, D., Ansaldi, J.L., Vinay, L., Figarella-Branger, D., Levy, N.,
Clarac, F., Cau, P., Pellissier, J.F., Fontes, M., 2002. PMP22 over-
expression causes dysmyelination in mice. Brain (125), 2213–2221.
Sahenk, Z., Chen, L., Mendell, J.R., 1999. Effects of PMP22 duplication
and deletions on the axonal cytoskeleton. Ann. Neurol. 45 (1), 16–24.
Sancho, S., Magyar, J.P., Aguzzi, A., Suter1, U., 1999. Distal
axonopathy in peripheral nerves of PMP22-mutant mice. Brain 122
(Pt 8), 1563–1577.
Sendtner, M., Kreutzberg, G.W., Thoenen, H., 1990. Ciliary neurotrophic
factor prevents the degeneration of motor neurons after axotomy. Nature
345 (6274), 440–441.
Sereda, M., Griffiths, I., Puhlhofer, A., Stewart, H., Rossner, M.J.,
Zimmerman, F., Magyar, J.P., Schneider, A., Hund, E., Meinck,
H.M., Suter, U., Nave, K.A., 1996. A transgenic rat model of
Charcot-Marie-Tooth disease. Neuron 16 (5), 1049–1060.
Sleeman, M.W., Anderson, K.D., Lambert, P.D., Yancopoulos, G.D.,
Wiegand, S.J., 2000. The ciliary neurotrophic factor and its receptor,
CNTFR alpha. Pharm. Acta Helv. 74 (2-3), 265–272.

T. Vigo et al. / Mol. Cell. Neurosci. 28 (2005) 703–714714
Snipes, G.J., Orfali, W., Fraser, A., Dickson, K., Colby, J., 1999. The
anatomy and cell biology of peripheral myelin protein-22. Ann. N. Y.
Acad. Sci. 883, 143–151.
Stankoff, B., Aigrot, M.S., Noel, F., Wattilliaux, A., Zalc, B., Lubetzki, C.,
2002. Ciliary neurotrophic factor (CNTF) enhances myelin formation: a
novel role for CNTF and CNTF-related molecules. J. Neurosci. 22 (21),
9221–9227.
Suter, U., Scherer, S.S., 2003. Disease mechanisms in inherited neuro-
pathies. Nat. Rev., Neurosi. 4, 714–726.
Suter, U., Snipes, G.J., 1995. Peripheral myelin protein 22: facts and
hypotheses. J. Neurosci. Res. 40 (2), 145–151.
Taylor, V., Welcher, A.A., Program, A.E., Suter, U., 1995. Epithelial
membrane protein-1, peripheral myelin protein 22, and lens membrane
protein 20 define a novel gene family. J. Biol. Chem. 270 (48),
28824–28833.
Tole, S., Patterson, P.H., 1993. Distribution of CD9 in the developing and
mature rat nervous system. Dev. Dyn. 197 (2), 94–106.
Valentijn, L.J., Baas, F., Wolterman, R.A., Hoogendijk, J.E., van den
Bosch, N.H., Zorn, I., Gabreels-Festen, A.W., de Visser, M., Bolhuis,
P.A., 1992. Identical point mutations of PMP-22 in Trembler-J mouse
and Charcot-Marie-Tooth disease type 1A. Nat. Genet. 2 (4), 288–291.
Verheijen, M.H., Chrast, R., Burrola, P., Lemke, G., 2003. Local
regulation of fat metabolism in peripheral nerves. Genes Dev. 17
(19), 2450–2464.
Wagner-Recio, M., Toews, A.D., Morell, P., 1991. Tellurium blocks
cholesterol synthesis by inhibiting squalene metabolism: preferential
vulnerability to this metabolic block leads to Peripheral nervous system
demyelination. J. Neurochem. 57 (6), 1891–1901.
Xiang, Z., Yang, Y., Ma, X., Ding, W., 2003. Microarray expression
profiling: analysis and applications. Curr. Opin. Drug Discovery Devel.
6 (3), 384–395.
Xiao, Y., Segal, M.R., Rabert, D., Ahn, A.H., Anand, P., Sangameswaran,
L., Hu, D., Hunt, C.A., 2002. Assessment of differential gene
expression in human peripheral nerve injury. BMC Genomics 3 (1), 28.
Yamamoto, M., Ito, Y., Mitsuma, N., Li, M., Hattori, N., Sobue, G., 2001.
Pathology-related differential expression regulation of NGF, GDNF,
CNTF, and IL-6 mRNAs in human vasculitic neuropathy. Muscle Nerve
24 (6), 830–833.
Yamamoto, M., Ito, Y., Mitsuma, N., Li, M., Hattori, N., Sobue, G., 2002.
Parallel expression of neurotrophic factors and their receptors in chronic
inflammatory demyelinating polyneuropathy. Muscle Nerve 25 (4),
601–604.





























