Embed Size (px)
Citation preview

Journal of Neuroimmunology
ELSEVIER Journal of Neuroimmunology 56 (1995) 53-63
Expression of vimentin increases in the hippocampus and cerebral cortex after entorhinal cortex lesioning and in response to transforming
growth factor pl
Knut Krohn, Nicholas J. Laping, Todd E. Morgan, Caleb E. Finch * Neurogerontology Division, Andrus Gerontology Center and Department of Biological Sciences, University of Southern California, Los Angeles, CA
90089-191, USA
Received 1 June 1994; revised 22 August 1994; accepted 22 August 1994
Abstract
Entorhinal cortex lesions (ECL) that damage the perforant path to the dentate gyrus of the hippocampal formation were used to model the regulation of vimentin (VIM) mRNA. ECL increased VIM mRNA in the ipsilateral hippocampus and in the ipsilateral cortex including the wound cavity within 1 day. By in situ hybridization, at 4 days post-ECL, VIM mRNA increased two-fold in the molecular layer of the dentate gyrus. VIM protein was co-localized by immunocytochemistry to astrocytes and microglia/macrophages. Transforming growth factor+?1 (TGF-/?l), which was previously shown to increase in microglia/macro- phages of the molecular layer after hippocampal deafferentation by ECL, was investigated as a regulator of VIM expression. Infusions of TGF-Pl into the lateral ventricle induced VIM mRNA with dose-dependence, e.g. infusion of 100 ng TGF-PI increased VIM mRNA three-fold. The increase in VIM mRNA was localized by in situ hybridization to astrocytes and microglia in the molecular layer of the dentate gyrus. These findings further implicate TGF-/?l as a regulator of cytoskeletal proteins during synaptic reorganization.
Keywords: Astrocytes; Entorhinal cortex lesion; Microglia/macrophages; TGF-Pl; Vimentin
1. Introduction
Vimentin (VIM), a type III intermediate filament, is expressed in astrocytes and neurons of the developing nervous system and is expressed in embryonic neurons prior to neurofilament proteins (Schnitzer et al., 1981; Bignami et al., 1982; Cochard and Paulin, 1984). VIM is also present in many types of different non-neuronal and neoplastic cells, in vivo and in vitro. In contrast, in the brains of healthy adult mammals, VIM expression is more restricted and is readily detected only in brain ependymal and subpial cells, and in certain astrocytes in white matter and in cerebellum (Bergmann glia) (Shaw et al., 1981; Mares et al., 1988; Yamada et al., 1992). However, VIM expression is increased in human neurodegenerative conditions that include Alzheimer’s disease, Pick’s disease, and amyotrophic lateral sclero-
* Corresponding author. Phone (213) 740 1758; Pax (213) 740 0853
01655728/95/$09.50 0 1995 Elsevier Science B.V. All rights reserved SSDl 0165-5728(94)00133-2
sis (Yamada et al., 1992). Experimental brain lesions that induce glial reactions also cause increased VIM immunoreactivity in both activated astrocytes and mi- croglia (Pixley and de Vellis, 1984; Schiffer et al., 1986; Graeber et al., 1988; Streit et al., 1988). Lesion-in- duced increases in VIM mRNA were found by North- ern blot hybridization in hippocampus (Day et al., 1992; Kindy et al., 1992) and in situ hybridization in brain stem astrocytes (Mikucki and Oblinger, 1991). Neural lesions also typically stimulate expression of glial fibrillary acidic protein (GFAP), another interme- diate filament protein of astrocytes (Brenner, 19941, that is associated with astrocytic maturation and reac- tions to brain injury (Rose et al., 1976; Schiffer et al., 1986; reviewed in Hatten et al., 1991; Eddlestone and Mucke, 1993).
Partial deafferentation of the hippocampus through entorhinal cortex lesions (ECL) is used as a mode1 for hippocampal deafferentation during Alzheimer’s dis- ease (Geddes et al., 1985). The entorhinal cortex con- tains the cell bodies of neurons that project to the
54 K. Krohn et ul. /Jolourmd “f Neuroimmunology 56 (1995) 53-63
hippocampus through the perforant path, where they contact dendrites of the granule neurons of the dentate gyrus. After electrolytic lesions of the entorhinal cortex terminals of the perforant path degenerate, as is found in Alzheimer’s disease. Subsequently, the deafferented granule cell dendrites degenerate as well (Matthews et al., 1976a). Within 10 days after lesioning, synaptic remodeling leads to the reappearance of morphologi- cally normal synaptic contacts in the deafferented hip- pocampus (Matthews et al, 1976b; Caceres and Stew- ard, 1983), with partial functional recovery (Myhrer 197.5; Scheff and Cotman, 1977). This synaptic reorga- nization begins several days after activation of mi- croglia and astrocytes (Gall et al., 1979; Gage et al., 1988; Fagan and Gage, 1990; Gehrmann et al., 1991; Morgan et al., 1993). Ongoing studies in this lab to clone mRNAs that are altered by entorhinal cortex lesions showed increased VIM mRNA sequences in hippocampal cDNA libraries (Day et al., 1992).
TGF-/31 is widely recognized for multiple roles dur- ing inflammatory responses to injury, but also during development and homeostasis (McCartney-Francis and Wahl, 1994). A review of evidence from this and other laboratories suggested that TGF+l has a broad role in organizing responses to brain injury (Finch et al., 1993). TGF-P1 mRNA and protein are present in the devel- oping rodent brain (Miller et al., 1989; Unsicker et al, 1991). Although TGF-Pl mRNA is easily detected in normal adult hippocampus or cortex by Northern blot hybridization (Nichols et al., 1991; Morgan et al., 19931, TGF-Pl protein is not found. Most agree that the occurrence of TGF+l protein within the brain re- quires neurodegenerative processes (Da Cunha et al., 1993; Morgan et al., 1994). TGF-Pl is also found in a subset of neuritic plaques of Alzheimer’s disease and Down syndrome (Van der Wal et al., 1993). TGF-PI mRNA is elevated by ECL and other experimental lesions (Nichols et al., 1991; Morgan et al., 1993; Pasinetti et al., 1993; Prehn et al., 1993) and in humans afflicted with HIV (Da Cunha et al., 1993).
Among its many effects on adult brain and cultured cells of neural origin (Finch et al., 19931, TGF-Pl has been shown to regulate cytoskeletal gene expression in many cell types. In cultured astrocytes, TGF-/31 in- duces the expression of GFAP and actin (Baghdas- sarian et al., 1993; Laping et al., 1994a). The effects on GFAP are transcriptional, as shown by nuclear run-on assays in hippocampus after infusions or in primary astrocyte cultures (Laping et al., 1994a). Moreover, TGF-/31 infusions increased levels of tubulin-al mRNA (Laping et al., 1994a), a developmental isoform that influences synaptic remodelling (Teichmann-Weinberg et al., 1988). In non-neural cells, TGF-/31 also regulates cytoskeletal proteins, e.g. in osteoblasts, it regulates VIM, but also (Y- and p-tubulin and actin (Lomri and Marie, 1990). These reports implicate TGF-01 as a
coordinator of cytoskeletal gene expression in diverse cell types. Because VIM mRNA was examined at only 14 days after entorhinal cortex lesioning (Day et al., 19921, we wished to resolve early phases of the re- sponse and to identify the cell types expressing VIM. We also tested the hypothesis that TGF-/31 regulates VIM gene expression, by infusing TGF-@I into the lateral ventricle.
2. Materials and methods
2.1. Entorhinal cortex lesion (ECL)
Male F344 rats, 2-3 months old, were kept in a light (12:12 h, L:D) and temperature (25”C)-controlled envi- ronment and given food and water ad libitum. Rats received unilateral electrolytic lesions of the entorhinal cortex under Nembutal anesthesia (40 mg/kg). Lesions were made applying a 1 mA current for 40 s through a unipolar electrode positioned consecutively at nine stereotaxic coordinates within the left temporal lobe of the cortex (Lynch et al., 1972; Geddes et al., 1985; Laping et al., 1991).
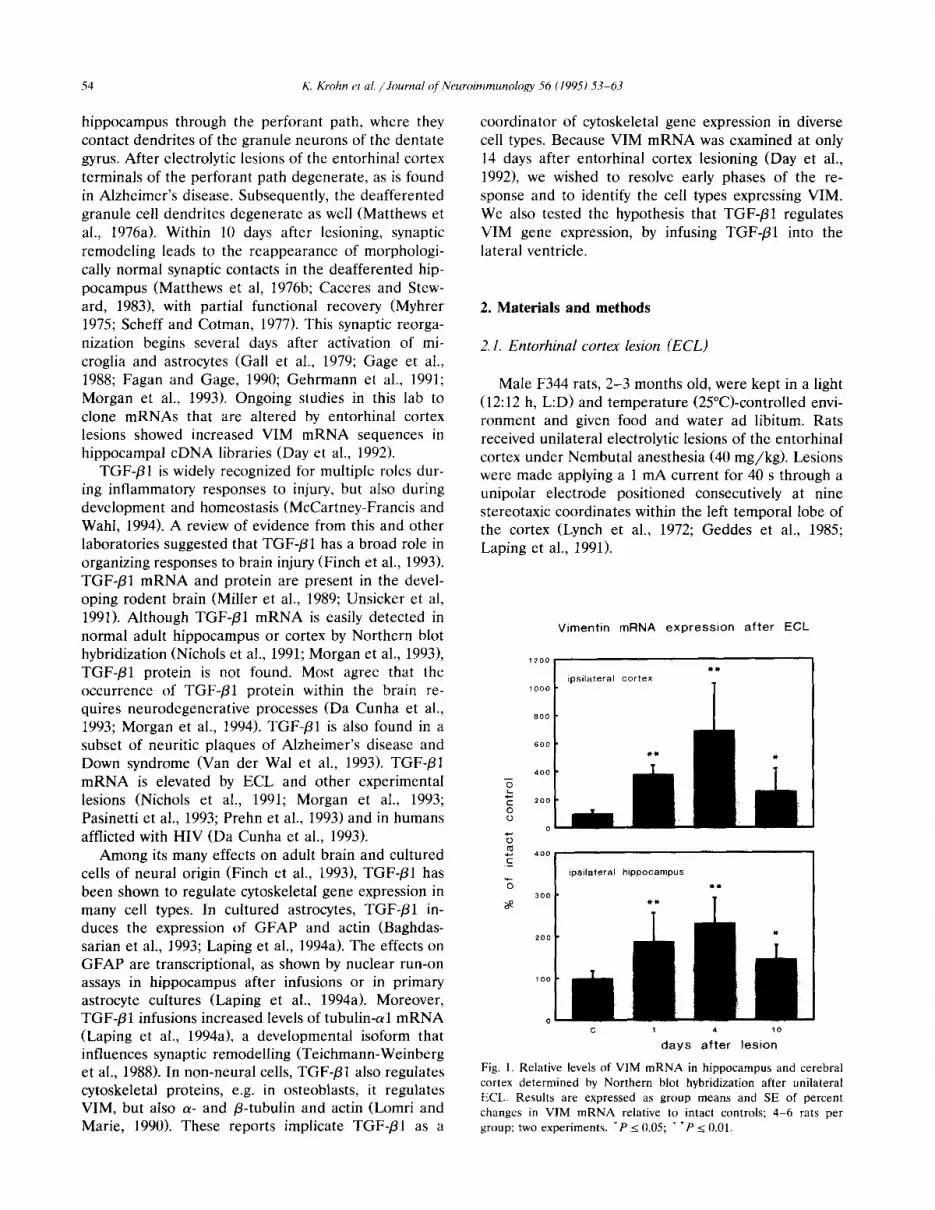
Vimentin mRNA expression after ECL
c 1 4 10
days after lesion
Fig. 1. Relative levels of VIM mRNA in hippocampus and cerebral
cortex determined by Northern blot hybridization after unilateral
ECL. Results are expressed as group means and SE of percent
changes in VIM mRNA relative to intact controls; 4-6 rats per
group; two experiments. ‘P < 0.05; * ‘P 2 0.01.

K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63 55
2.2. infusion of TGF-PI
Male F344 rats were anesthetized with Nembutal. The lateral ventricle was infused with 2 ~1 vehicle
(Dulbecco’s medium with 2 mg/ml bovine serum albu- min, 4 mM HCl, pH 6.5) or l-500 ng human recombi- nant TGF-/31 (Genentech, San Francisco, CA) using a stereotaxically directed 22 gauge cannula and a peri-
Fig. 2. Micrographs (dark field, coronal sections) of in situ hybridization for VIM mRNA in ipsilateral intact hippocampus (A,B), 1 day (C) and 4 days (E,F) post-ECL. In intact and lesioned hippocampus, VIM mRNA was detected in ependymal cells (filled arrowheads in the left panels) and vascular endothelial cells (open arrowheads in B and F). 4 days after ECL (E,F) VIM mRNA is induced in the molecular layer (m) of the dentate gyrus (dg). (C) Two of four rats had increased VIM mRNA levels in the dentate gyrus hilar region (hi) at 1 day post-ECL. Increased VIM mRNA after ECL is also seen in white matter (wm) of the cerebral cortex (panels C,E). Panels B and F show high magnification micrographs of control hippocampus (B) and 4 days post-ECL (F). No hybridization is detectible with the sense-strand cRNA probe in a section (D) adjacent to the section in (E). Bar represents 50 pm and pertains to (B,F). Bar in (A) represents 1 mm and pertains to panels (A,C,D,E). Abbreviations: oml, outer molecular layer; gc, granule cell layer.

56 K. Krohn et ul. /Journal of Neuroimmunology 56 (1995) 53-63
staltic pump controlling the flow at 1 pl/min. Rats were killed 24 h later. Ipsilateral and contralateral hippocampus and cerebral cortex were frozen at - 70°C. Whole brains were frozen for in situ hybridiza- tion.
2.3. RNA analysis
Total RNA was extracted from frozen brain tissues by guanidinium-thiocyanate (Chomczynski and Sacchi, 1987). After electrophoresis on denaturing
Fig, 3. Micrographs (coronal sections) of bright field immunocytochemistry (ICC) for VIM (A,B); fluorescence immunocytochemistry (FIG) for
VIM (C-E); FIC for GSA I-B, (microglia/macrophage marker) (F); FIC for OX-42 immunoreactivity (microglia/macrophage marker) (G) and
FIC for GFAP (astroglia marker) (H) in ipsilateral rat hippocampus 4 days after unilateral ECL (B,C-H) and intact hippocampus (A). No
immunoreactivity in the intact hippocampus was detected (A). In the deafferented hippocampus (Bl, bar size represents 1 mm and pertains to
(A,Bl, strong VIM immunoreactivity is detectable in the molecular layer (m) of the dentate gyrus (dgl. Weaker signals are shown in the dentate
gyrus hilar region (hi). (C,F), (D,G) and (E,H) show the same field of view from identical sections using different filters. Double-labelled
microglia/macrophages (C,F and D,G) and a double-labelled astrocyte (E,F) are marked by the asterisk. Arrows indicate cells labeled only for
VIM (C-E) and arrowheads indicate cells labeled only for microglia by GSA I-B., (F) or by OX-42 immunoreactivity (G) and for astrocytes by
GFAP (H). Bar size in (Cl represents 10 Km and pertains to (C-H).

K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63 51
agarose/formaldehyde gels (Sambrook et al., 1989) and transfer to nylon membranes (Nylon 1, BRL) in 2 x standard saline citrate (2 x SSC), RNA was cross- linked by UV to the membranes and stained by Methy- lene blue. Blots were hybridized at 54°C with a 32P- anti-sense cRNA probe from linearized subclones in pBluescript (Stratagene, La Jolla, CA). The VIM clone (cDNA, 1.7 kb) was isolated in this laboratory (Day et al., 1992). The neurofilament-68 clone containing a 1.2-kb cDNA insert (Lewis and Cowan, 1985) was a gift from Nicholas J. Cowan; 0.6 kb of this clone were subcloned into Bluescript using an internal Sac1 re- striction site. Hybridization conditions were as follows: lo6 cpm/ml in 50% formamide, 1.5 X SSC, 1% SDS, 0.5% dry milk, 100 pg/ml yeast total RNA, and 300 pg/ml sonicated salmon sperm DNA. Blots were washed to a final stringency with 0.2 X SSC and 0.2% SDS at 74°C. Radioactivity was analyzed by phospho- rimaging using the computer program ImageQuant (Molecular Dynamics, Sunnyvale, CA).
2.4. In situ hybridization
Coronal or horizontal sections (30 pm> were mounted on polylysine-coated slides. Hybridization was done according to Morgan et al. (1993). Briefly, after rinsing with phosphate-buffered saline (PBS) and treat- ment with acetic anhydride in 0.1 M triethanolamine, sections were rinsed again in PBS and dehydrated. To avoid cross-hybridization with GFAP mRNA we used a probe containing 0.42 kb at the 3’ end of the VIM message that did not have sequence-homology to GFAP. The VIM clone (1.7 kb) digested with H&II was used as a template for cRNA synthesis. Sections were hybridized with a 33P-labeled sense- and anti- sense cRNA probe (0.3 ng cRNA probe/kl/kb of target sequence, 15 pi/section) at 55”C/3 h. Following hybridization, sections were treated with RNase A (25 pg/ml; Sigma, St. Louis, MO) and high stringency washed (63°C) in 50% formamide, 200 mM phosphate buffer, 500 mM NaCl and 100 mM &mercaptoethanol. Sections were dehydrated and exposed to XAR-5 film (Kodak; Rochester NY) for l-2 days before covering with NTB2 emulsion for cellular analysis. After devel- oping the emulsion, the sections were counterstained with Cresyl violet.
2.5. Bright field immunocytochemistry (ICC), fluores- cence immunocytochemistry, and lectin histochemistry
Coronal or horizontal sections were treated with normal serum before incubation with primary antibody (monoclonal anti-GFAP concentration 2.5 Fg/ml; monoclonal anti-VIM concentration 2 pg/ml, Boehringer Mannheim, Indianapolis, IN; monoclonal anti-CR3 complement receptor-MRC (clone 0X-42,
concentration 6 pg/ml, Serotec, Kidlington, UK)) for 1 h at room temperature. A peroxidase-labeled sec- ondary antibody (NA 931 sheep anti-mouse, dilution 1:1000, Amersham, Arlington Heights, IL) and di- aminobenzidine as the peroxidase-substrate was used for the detection of primary antibodies.
Immunofluorescence labeling was done with three reagents: a Texas red-conjugated secondary antibody, rabbit anti-mouse, 1:lOO (Jackson Immunoresearch Laboratories, West Grove, PA); a fluorescein isothio- cyanate (FIT0conjugated secondary antibody, goat anti-mouse, 1:lOO (Jackson Immunoresearch Laborato- ries); or a biotinylated secondary antibody (Vector, Burlingame, CA) together with rhodamine-conjugated streptavidin, 1:lOO (Jackson Immunoresearch Labora- tories). Sections were rinsed in PBS and coversliped, or processed for in situ hybridization or double-labeling. Sections treated with PBS, normal serum and sec- ondary antibody without primary antibody showed no specific staining.
FITC-labeled B,-isolectin from Grifioniu simplicifo- liu (GSA I-B,) (20 pg/ml, Vector, Burlingame, CA) was used for lectin histochemistry (Streit, 1990). After ICC sections were incubated for 1 h at 22°C rinsed, and coversliped.
2.6. Statistics
Significance of experimental results was evaluated with ANOVA in effects of TGF-Pl infusions (brain regions, mRNAs, dose, and time effects) and with two-tailed Student’s t-test for comparisons between single experimental and control groups (Sinclair, 1988).
3. Results
3.1. I/IM mRNA expression after entorhinal cortex lesions (ECL)
We analyzed VIM mRNA in major brain regions by Northern blot hybridization after ECL. In the ipsilat- era1 hippocampus, VIM mRNA was increased 180% at 1 day and continued to increase slightly to a maximum of 230% at 4 days (Fig. 1). The ipsilateral cortex including the wound cavity had larger increases, up to 700%, on the same schedule (Fig. 1). No changes in VIM mRNA were detected in the contralateral hip- pocampus and contralateral cortex up to 10 days post- ECL.
3.2. Regional and cellular localization of VIM mRNA and VIM protein
In situ hybridization was done with a smaller cRNA probe containing 0.45 kb at the 3’ end of VIM cDNA

58 K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63
that minimized cross-hybridization to GFAP mRNA (see Materials and methods). In unlesioned controls, VIM mRNA was detected in ependymal and endothe- ha1 cells but not in the hippocampal parenchyma (Fig. 2A). The ependymal cell layer gave strong signals (Fig. 2A, C, F), as did the pial layer (not shown).
ECL induces VIM mRNA in the outer molecular layer of the dentate gyrus at 4 days (Fig. 2E,F). These increases co-localize with the zone of degenerating terminals from the perforant path (see Introduction). A subgroup (2/4) of rats showed increased VIM mRNA at 1 day post-ECL, but only in the hilar region of the dentate gyrus (Fig. 20. These increases appear to be transient, because no elevations of VIM mRNA were found in this location at 4 days. In all rats, the cerebral cortex had strong VIM mRNA signals in white matter and around the wound cavity (not shown). No hybridization signal was found with a sense-strand cRNA probe (Fig. 2D).
with glia by each of these reagents (Fig. 31, which indicates the expression of VIM in subpopulations of both microglia and astrocytes. VIM was undetectable in neurons. In the outer molecular layer of the dentate gyrus, VIM was more frequently detected in astrocytes than in microglia/macrophages (ratio about 5:l). How- ever, most of the astrocytes (> 70%) and microglia (> 90%) in this area did not display VIM immuno- reactivity.
3.3. VIM mRNA and neurofilament-68 mRNA after TGF-_Pl infusion
After ECL, VIM protein was detected by immuno- cytochemistry in glial cells only in the ipsilateral hip- pocampal parenchyma and in the wound cavity at 4 days (Fig. 4B1, but not at 1 day (not shown). The localization of VIM immunoreactivity corresponds pre- cisely to the location of VIM mRNA by in situ hy- bridization. A more sensitive immunofluorescence technique detected VIM immunoreactivity in a small number of astrocytic processes in the inner molecular layer of the dentate gyrus in the intact and lesioned hippocampus (not shown). This localization agrees with findings of Schmidt-Kastner and Szymas (19901 that were based on acetone-fixed frozen sections.
The hypothesis that TGF-PI is a regulator of VIM expression during responses to lesions was examined by infusing TGF-@l into rats that were not given ECL. For comparison, we assayed neurofilament-68 mRNA, a neuronal intermediate filament that is not induced by ECL in the times studied here (C. Zarow and C.E. Finch, submitted for publication). Northern blots from total adult brain RNA showed a single band of 2.2 kb (VIM probe), as observed after ECL, and two bands of 4.0 and 2.5 kb (neurofilament probe). TGF-/31 in- creased VIM mRNA in the ipsilateral hippocampus and ipsilateral cortex 24 h after infusion into the lateral ventricle. The dose-dependent increase (Table 1) showed peak responses at 100 ng with suggestions of a biphasic reduction at higher doses. VIM mRNA levels in the contralateral hippocampus and contralateral cortex did not differ after TGF-/?l infusion from vehi- cle infusion. TGF-PI had no effect on neurofilament-68 mRNA.
The type of glia that contained increased VIM The induction of VIM mRNA by TGF-Pl (200 ng) mRNA and protein after ECL was determined by was localized to the outer molecular layer of the den- double-staining of brain sections using VIM immuno- tate gyrus and to the ependymal layer of the ventricles reactivity together with reagents that identify mi- adjacent to the hippocampus (Fig. 41. In contrast, VIM croglia/macrophages (the lectin, GSA I-B,; antibody mRNA did not increase in the surrounding zones of to OX-421 or astrocytes (antibody to GFAP). VIM pyramidal neurons. The increase in VIM mRNA in the immunoreactivity was co-localized by double-staining hippocampal parenchyma was co-localized to astro-
Table 1 Northern blot hybridization after TGF-fil infusion.
Dose of TGF-P 1 Vimentin
Hippocampus
Ipsi Contra
Cortex
Ipsi Contra
Neurofilament-68
Hippocampus
Ipsi Contra
Vehicle 100 + 25 100+26 100+39 100*33 100 + 16 lOOk
10 ng 113 + 36 71 f 11 214k I1 93 * 22 111* 15 137 10 *
50 ng 242 + 95 88 + 5 227 + 54 98 * 30 101 f 36 124+8
100 ng 296 + 122 86+ 10 3X1 + 46 130 54 f 91 38 f 114 21 +
500 ng 158 i 36 ND ND ND 114 30 * ND
P 0.05 NS 0.01 NS NS NS
Two to five rats per group; two independent experiments. Increases were signficant (P < 0.05) at doses of 50-500 ng. Relative mRNA levels 24 h
after human recombinant TGF-Pl infusion into the lateral ventricle of male rats. Results are expressed as group means and standard error
compared to the vehicle infused control group. ND. not determined; NS. not significant; P. level of significance for one-way ANOVA.

K. Krohn et al. /Journal of Neuroimmunology 56 (I 995) 53-63 59
Fig. 4. Micrographs (dark field, horizontal sections) of in situ hybridization for VIM mRNA (A,B) together with immunocytochemistry for GFAP (bright field, C; dark field, D) or OX-42 immunoreactivity (bright field, E; dark field, F) in the ipsilateral hippocampus 24 h after TGF-@l (B-F) or after vehicle infusion (A, the bar represents 500 Frn and pertains to A,B). VIM mRNA is induced 24 h after TGF-P1 infusion (B) in the molecular layer (m) of the dentate gyrus (dg), but not in vehicle-infused hippocampus. (C,D) Arrowheads indicate co-localization of VIM mRNA expression (D) with immunoreactivity for GFAP (C, bar represents 20 pm and pertains to C-F). (E,F) Open arrowheads indicate OX-42 immunreactive cells (E) co-localized with VIM mRNA (F).
cytes and macrophage/microglia, by immunostaining for GFAP and OX-42 IR, respectively (Fig. 4).
the perforant path terminals, that is downstream of the degenerating axons, in the ipsilateral hippocampus where synaptic reorganization is stimulated (see Intro-
4. Discussion
This study showed that electrolytic entorhinal cortex lesions induced VIM mRNA in glia that were near to
duction). In both the hippocampus and the cerebral cortex surrounding the electrolytic lesion, the induction of VIM mRNA was detected by 1 day and reached a peak by 4 days after lesioning. The increase in VIM

60 K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63
mRNA was localized to the molecular layer of the dentate gyrus, the white matter, and wound cavity of the cerebral cortex. Apparently transient increases of VIM mRNA 1 day postlesion in the hilar region of the dentate gyrus in a subgroup of rats probably were induced by seizures during electrolytic lesioning (Ste- ward et al., 1991). We emphasize that the detection of a signal for VIM mRNA or protein indicates relative prevalance and does not rule out lower levels of VIM that might be detected with more sensitive assays in other cells.
VIM protein was co-localized with markers for both microglia/macrophage and astrocytes. However, only a minority of OX-42 positive or GFAP positive cells showed VIM immunoreactivity. In microglia/macro- phages, the VIM immunoreactivity and either GSA I-B, staining or OX-42 immunoreactivity was co-local- ized to cells with a round-shape. This microglial mor- photype also contains increased levels of TGF-Pl mRNA after entorhinal cortex lesions and may repre- sent cells invading from the blood at the margin of the lesion (Morgan et al., 1993). VIM is also a putative marker for microglial activation (Schiffer et al., 1986; Gehrmann et al., 1991; Yamada et al., 1992). Entorhi- nal cortex lesions increase the numbers of activated astrocytes from several possible sources. While most activated astrocytes are thought to hypertrophy without DNA synthesis, a small subpopulation could be prolif- erating (reviewed in Fagan and Gage, 1990; Schiffer et al., 1993). Because both hypertrophic or hyperplastic astroglia express VIM (Janeczko 1993), the presence of VIM in adult astrocytes may be considered as a general marker for astroglial activation (Yamada et al., 19921, but does not indicate the recentness of proliferation. While neurons express VIM during development (Cochard and Paulin, 1984), we saw no VIM immuno- reactivity in hippocampal neurons.
Next we considered the role of TGF-/31 in hip- pocampal responses to lesioning. In the present study, lesions of the entorhinal cortex damage the perforant path neurons projecting to the dendrites in the outer molecular later of the dentate gyrus of the hippocam- pal formation. In the zone of degenerating perforant path afferents, the deafferented molecular layer of the dentate gyrus undergoes dendritic degeneration and synaptic remodeling (see Introduction). Activation of microglia/macrophages in the outer molecular layer of the dentate gyrus occurs within 1 day after lesioning (Gehrmann et al., 19911. By immunoassay, this activa- tion is associated with an increase of TGF-PI (Morgan et al., 1994). Because IL-1 increases TGF-/31 protein in cultured astrocytes (Da Cunha and Vitkovic 1992), a working hypothesis is that IL-1 is released in initial responses to ECL (Fagan and Gage, 1990); IL-1 may then cause the release of TGF-P1 from microglia and other local cells (Finch et al., 1993). Because TGF-Pl
Ent
Fig. 5. Diagram of the hippocampal region according to the brain
atlas of Paxinos and Watson (1986). The TGF-pl responsive glial
cells (VIM mRNA expression; dark grey area) in the molecular layer
(ml) of the dentate gyrus (dg) are distant from the highest TGF$l
concentrations in the lateral ventricle (Iv). Similarly, highly TGF-PI
responsive neurons (peak-increase of tubulin a-l mRNA (Laping et
al., 1994a); light grey area) are located in the CA3 and hilus region
(hi) as well as in granule cells (gc) of the dentate gyrus. CC, cerebral
cortex; Ent, entorhinal cortex; wm, white matter.
infusions also increase astrocyte mRNAs that encode GFAP, clusterin and fibronectin (Pasinetti et al., 1993; Laping et al., 1994a), TGF-Pl may be among the IL-l-triggered activators of astroglial reactions.
To further test this hypothesis, we studied the ex- pression of VIM, which is a putative marker for glial activation, after TGF-/!I1 infusion into the lateral ven- tricle. TGF-/?l induced expression of VIM mRNA in three distinct locations: (i) the molecular layer of the dentate gyrus in the ipsilateral hippocampus; (ii> ependymocytes lining the lateral ventricle into which the TGF-/31 was infused; (iii> ipsilateral cerebral cor- tex. The induction of VIM mRNA in the ipsilateral hippocampus was apparently restricted to glia in the molecular layer of the dentate gyrus. The increase in VIM mRNA was co-localized with two types of glia, astrocytes and microglia/macrophages in the molecu- lar layer of the dentate gyrus. Moreover, the responsive glia are distant from the highest concentrations of TGF-Pl in the lateral ventricle (Fig. 5). In contrast, we did not see increases of VIM mRNA in astrocytes or microglia that were nearer to the ventricle, e.g. in the dendritic fields of the CAl-CA2 pyramidal neurons. In a striking parallel to these findings, other studies from this laboratory showed that a neuron-specific cyto- skeletal mRNA, tubulin-al, was increased by TGF-/?I infusions in the granule neurons and nearby CA3 neu- rons to a greater extent than in the CAl-2 zone (Lap- ing et al., 1994a; see Fig. 51. Together, these findings indicate that TGF-j31 serves as an organizer of cytoar- chitectonic changes during responses to deafferenta-

K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63 61
tion in glia and neurons through effects on cytoskeletal mRNA. Because dose responses may be U-shaped (Table l), responsive subpopulations of glia and neu- rons may occupy particular positions in concentration gradients of TGF-Pl. However, the capacity of TGF-Pl for auto-induction of its own mRNA in brain (Young- Chan et al., 1993) might lead to mosaics of local concentrations. The issues raised by cell responses distant from the site of TGF-/31 infusion have counter- parts in development, where inducers of differentiation may cause cascades that influence differentiation in downstream targets.
Because autoregulation of TGF-@l is mediated by the transcription factors that bind to c&-acting AP-1 sites (Kim et al., 19901, it is interesting that the S-regu- latory regions of the GFAP (Masood et al., 1993) and VIM genes (Rittling et al., 1989) contain DNA binding sites for the JUN-FOS heteromeric complex. NF-1, another putative regulatory element that is found in the GFAP and VIM promoters (Laping et al., 1994b; Masood et al., 1993), can mediate activation of a colla- gen promoter by TGF-/31 (Rossi et al., 1988).
TGF-PI may function in three modes during re- sponses to lesions. (i) Autocrine regulation: TGF-/31 can induce its own mRNA in cultured cells (Lindholm et al., 1990; Morganti-Kossmann et al., 1992) and in the brain in vivo (Young-Chan et al., 1993), thereby amplifying initial microglial reactions. (ii> Astrocytic activation: TGF-Pl activates astrocytes, which are im- plicated in the regulation of synaptic remodelling in the deafferented hippocampus through their produc- tion of NGF and other growth factors (Gage et al., 1988; Johnson-Green et al., 1992). Moreover, TGF-/?l increases NGF production and synergizes with FGF and IL-lp to stimulate NGF secretion (Lindholm et al., 1990; Yoshida and Gage, 1992; Hahn et al., 1994). (iii) Neuronal remodelling: TGF-Pl might directly stimulate synaptic reorganization by promoting neurite sprouting and neurite elongation, as observed in cul- tured rat hippocampal neurons (Ishihara et al., 1994). Recall from above that TGF-Pl infusions induce tubu- lin-cwl mRNA &aping et al., 1994a), a developmental tubulin isoform that mediates neurite extension (Teich- mann-Weinberg et al., 1988). Thus, TGF-Pl, in con- junction with other cytokines and growth factors, may serve to organize glial and other responses to brain injury.
Acknowledgements
This work was supported by PHS Grant AG 07909 to C.E.F., a grant by the German Academic Exchange Service (DAAD) to K.K., NRSA Grant AG 05528 to N. J. L. and NRSA Grant to T. E. M.
References
Baghdassarian, D., Toru-Delbauffe, D., Gavaret, J.M. and Pierre, M. (1993) Effects of transforming growth factor-p1 on the extracellu- lar matrix and cytoskeleton of cultured astrocytes. Glia 7,193-202.
Bignami, A., Raju, T. and Dahl, D. (1982) Localization of vimentin, the nonspecific intermediate filament protein, in embryonal glia and in early differentiating neurons. Dev. Biol. 91, 286-295.
Brenner, M. (1994) Structure and regulation of the GFAP gene. Brain Pathol. 4, 245-257.
Caceres, A. and Steward, 0. (1983) Dendritic reorganization in the denervated dentate gyrus of the rat following entorhinal cortical lesions: a golgi and electron microscopic analysis. .I. Comp. Neu- rol. 214, 387-403.
Chomczynski, P. and Sacchi, N. (19871 Single-step method for RNA isolation by acid guanidinium thiocyanate-phenol-chloroform ex- traction. Anal. Biochem. 162, 156-159.
&chard, P. and Paulin, D. (1984) Initial expression of neurofila- ments and vimentin in the central and peripheral nervous system of the mouse embryo in vivo. J. Neurosci. 4, 2080-2094.
Da Cunha, A. and Vitkovic, L. (1992) Transforming growth factor- beta 1 (TGF-fill expression and regulation in rat cortical astro- cytes. J. Neuroimmunol. 36, 157-169.
Da Cunha, A., Jefferson, J.A., Jackson, R.W. and Vitkovic, L. (1993) Glial cell-specific mechanisms of TGF-Pl induction by IL-l in cerebral cortex. J. Neuroimmunol. 42, 71-86.
Day, J.R., Min, B.H., Laping, N.J., Martin III, G., Osterburg, H.H. and Finch, C.E. (19921 New mRNA probes for hippocampal responses to entorhinal cortex lesions in the adult male rat: a preliminary report. Exp. Neurol. 117, 97-99.
Eddlestone, M. and Mucke, L. (1993) Molecular profile of reactive astrocytes - implications for their role in neurologic disease. Neuroscience 54, 15-36.
Fagan, A.M. and Gage, F.H. (1990) Cholinergic sprouting in the hippocampus: a proposed role for IL-l. Exp. Neurol. 110, 105 120.
Finch, C.E., Laping, N.J., Morgan, T.E., Nichols, N.R. and Pasinetti, G.M. (1993) TGF-fll is an organizer of responses to neurodegen- eration. J. Cell. Biochem. 53, 314-322.
Gage, F.H., Olejniczak, P. and Armstrong, D.M. (1988) Astrocytes are important for sprouting in the septohippocampal circuit. Exp. Neurol. 102, 2-13.
Gall, C., Rose, G. and Lynch, G. (1979) Proliverative and migratory activity of glial cells in the partially deafferented hippocampus. J. Comp. Neurol. 183, 539-550.
Geddes, J.W., Monaghan, D.T., Cotman, C.W., Lott, IT., Kim, R.C. and and Chui, H.C. (1985) Plasticity of hippocampal circuitry in Alzheimer’s disease. Science 230, 1179-1181.
Gehrmann, J., Schoen, S.W. and Kreutzberg, G.W. (1991) Lesion of the entorhinal cortex leads to rapid microglial reaction in the dentate gyrus. Acta Neuropathol. (Berl.1 82, 442-455.
Graeber, M.B., Streit, W.J. and Kreutzberg, G.W. (1988) The mi- croglial cytoskeleton: vimentin is localized within activated cells in situ. J. Neurocytol. 17, 573-580.
Hahn, M., Lorez, H. and Fischer, G. (1994) The immortalized astroglial cell-line RC7 is a new model system for the study of nerve growth-factor (NGF) regulation - stimulation by inter- leukin-l-beta and transforming growth-factor-beta-l is additive and affected differently by dibutyryl-cyclic-AMP. Glia 10, 286- 295.
Hatten, M., Liem, R.K.H., Shelanski, M.L. and Mason, C.A. (1991) Astroglia in CNS injury. Glia 4, 233-243.
Ishihara, A., Saito, H. and Abe, K. (1994) Transforming growth factor+1 and $32 promote neurite sprouting and elongation of cultured rat hippocampal neurons. Brain Res. 639, 21-25.
Janeczko, K. (19931 Co-expression of GFAP and vimentin in astro- cytes proliferating in response to injury in the mouse cerebral

62 K. Krohn et al. /Journal of Neuroimmunology 56 (1995) 53-63
hemisphere. A combined autoradiographic and double immuno-
cytochemical study. Int. J. Dev. Neurosci. 11, 139-147.
Johnson-Green, P.C., Dow, K.E. and Riopelle, R.J. (1992) Neurite
growth modulation associated with astrocyte proteoglycans: influ-
ence of activators of inflammation. Glia 5, 33-42.
Kim, S.J., Angel, P., Fafyatis, R., Hattori, K., Kim, K.Y., Sporn,
M.B., Karin, M. and Roberts, A.B. (1990) Autoinduction of
transforming growth factor pl is mediated by the AP-1 complex.
Mol. Cell. Biol. 10, 1492-1497.
Kindy, MS., Bhat, A.N. and Bhat, N.R. (1992) Transient &hernia
stimulates glial fibrillary acid protein and vimentin gene expres-
sion in the gerbil neocortex, striatum and hippocampus. Mol.
Brain Res. 13, 199-206.
Laping, N.J., Nichols, N.R., Day, J.R. and Finch, C.E. 11991)
Corticosterone differentially regulates the bilateral response of
astrocyte mRNAs in the hippocampus to entorhinal cortex le-
sions in male rats. Mol. Brain Res. 10, 291-297.
Laping, N.J., Morgan, T.E., Nichols, N.R., Rozovsky, I., Young-Chan,
C.S., Zarow, C. and Finch, C.E. (1994a) Transforming growth
factor;01 induces neuronal and astrocyte genes: Tubulin (~1, glial
fibrillary acidic protein, and clusterin. Neuroscience 58, 563-572.
Laping, N.J., Teter, B., Nichols, N.R., Rozovsky, I., and Finch, C.E.
(1994b) Glial fibrillary acidic protein: regulation of expression by
hormones, Cytokines, and growth factors. Brain Pathol. 4, 259-
275.
Lewis, S.A. and Cowan, N.J. (1985) Genetics, evolution and expres-
sion of the 68,000-mol-wt neurofilament protein: isolation of a
cloned cDNA probe. J. Cell Biol. 100, 843-850.
Lindholm, D., Hengerer, B., Zafra, F. and Thoenen, H. (1990)
Transforming growth factor-/31 stimulates expression of nerve
growth factor in the rat CNS. NeuroReport 1, 9-12.
Lomri, A. and Marie, P.J. (1990) Bone cell responsiveness to trans-
forming growth factor beta, parathyroid hormone, and prosta-
glandin E2 in normal and postmenopausal osteoporotic women.
J. Bone Miner. Res. 5, 1149-1155.
Lynch, G., Matthews, D.A., Mosko, S., Parks, T. and Cotman, C.
(1972) Induced acetylcholinesterase-rich layer in rat dentate gyrus
following entorhinal lesions. Brain Res. 42, 311-318.
Mares, V., Viklicky, V.. Gerstein, L.M., Draber, P. and Ciesielski-
Treska, J. (1988) Immunocytochemistry and heterogeneity of rat
brain vimentin. Histochemistry 88, 575-581.
Masood, K., Besnard, F., Su, Y. and Brenner, M. (1993) Analysis of a
segment of the human glial fibrillaty acidic protein gene that
directs astrocyte-specific transcription. J. Neurochem. 61, 160-
166.
Matthews, D., Cotman, C.W. and Lynch, G. (1976a) An electron
microscopic study of lesion-induced synaptogenesis in the dentate
gyrus of the adult rat. I. Magnitude and time course of degenera-
tion. Brain Res. 115, 1-21.
Matthews, D., Cotman, C.W. and Lynch, G. (1976bJ An electron
microscopic study of lesion-induced synaptogenesis in the dentate
gyrus ot the adult rat. II. Reappearance of morphologically
normal synaptic contacts. Brain Res. 115, 23-41.
McCartney-Francis, N.L. and Wahl, SM. (1994) Transforming growth
factor beta-a matter of life and death. J. Leukoc. Biol. 55,
401-409.
Mikucki, S.A. and Oblinger, M.M. (1991) Vimentin mRNA expres-
sion increases after corticospinal axotomy in the adult hamster.
Metab. Brain Dis. 6, 33-49.
Miller, D.A., Lee, A., Matsui, Y., Chen, E.Y., Moses, H.L. and
Derynck, R. (1989) Complementary DNA cloning of the murine
transforming growth factor-beta 3 (TGF beta 3) precursor and
the comparative expression of TGF beta 3 and TGF beta 1
messenger RNA in murine embryos and adult tissues. Mol.
Endocrinol. 3, 1926-1934. Morgan, T.E., Sarkar, D.K., Vale, W. and Finch, C.E. (1994) Trans-
forming growth factor (TGF)-pl regulates rat brain TGF-P type
II receptor mRNA levels. Neuroscience Sot. Abst. 690.16, 1589.
Morgan, T.E., Nichols, N.R., Pasinetti, G.M. and Finch, C.E. (1993)
TGF-/31 mRNA increases in macrophage/microgliaI cells of the
hippocampus in response to deafferentation and kainic acid-in-
duced neurodegeneration. Exp. Neurol. 120, 291-301.
Morganti-Kossmann, M.C., Kossmann, T., Brandes, M.E., Mergen-
hagen, S.E. and Wahl, S.M. (1992) Autocrine and paracrine
regulation of astrocyte function by transforming growth factor-p.
J. Neuroimmunol. 39, 163-174.
Myhrer, T. (1975) Maze performance in rats with hippocampal
perforant paths lesions: Some aspects of functional recovery.
Physiol. Behav. 15, 433-437.
Nichols, N.R., Laping, N.J., Day, J.R. and Finch, C.E. (1991) In-
creases in transforming growth factor-beta mRNA in hippocam-
pus during response to entorhinal cortex lesions in intact and
adrenalectomized rats. J. Neurosci. Res. 28, 134-139.
Pasinetti, G.M., Nichols, N.R., Tocco, G., Morgan, T., Laping, N.
and Finch, C.E. (1993) Transforming growth factor pl and fi-
bronectin messenger RNA in rat brain: responses to injury and
cell-type localization. Neuroscience 54, 893-907.
Paxinos, G. and Watson, C. (1986) The rat brain in stereotaxic
coordinates. Academic Press, Orlando, FL.
Pixley, S.K.R. and de Vellis, J. (1984) Transition between immature
radial glia and mature astrocytes studied with a monoclonal
antibody to vimentin. Dev. Brain Res. 15, 201-209.
Prehn, J.H.M., Backhauss, C. and Krieglstein, J. (1993) Transforming
growth factor;01 prevents glutamate neurotoxicity in rat neocorti- cal cultures and protects mouse neocortex from ischemic injury.
J. Cereb. Blood Flow Metab. 13, 521-525.
Rittling, S.R., Coutinho, L., Amram, T. and Kolbe, M. (1989) AP-
l/jun binding sites mediate serum inducibility of the human
vimentin promoter. Nucleic Acids Res. 17, 1619-1633.
Rose, G., Lynch, G. and Cotman, C.W. (1976) Hypertrophy and
redistribution of astrocytes in the deafferented dentate gyrus.
Brain Res. Bull. 1, 87-92.
Rossi, P., Karsenty, G., Roberts, A.B., Roche, N.S., Sporn, M.B. and
de Crombrugghe, B. (1988) A nuclear factor 1 binding site
mediates the transcriptional activation of type I collagen pro-
moter by transforming growth factor-p. Cell 52, 405-414.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular
Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY. Scheff, S.W. and Cotman, C.W. (1977) Recovery of spontaneous
alternation following lesions of the entorhinal cortex in adult rats:
Possible correlation to axon sprouting. Behav. Biol. 21, 268-293.
Schiffer, D., Giordana, M.T., Migheli, A., Giaccone, G., Pezzotta, S.
and Mauro, A. (1986) Glial fibrillary acidic protein and vimentin
in the experimental glial reaction of the rat brain. Brain Res. 374,
110-118.
Schiffer, D., Giordana, M.T., Cavalla, P., Vigliani, M.C. and Attana-
sio, A. (1993) Immunohistochemistry of glial reaction after injury
in the rat: double staining and markers of cell proliferation. Int.
J. Dev. Neurosci. 11, 269-280.
Schmidt-Kastner, R. and Szymas, J. (1990) Immunohistochemistry of
glial fibrillary acidic protein, vimentin and S-100 protein for study
of astrocytes in hippocampus of rat. J. Chem. Neuroanat. 3, 179-192.
Schnitzer, J., Franke, W.W. and Schachner, M. (1981) Immunocyto-
chemical demonstration of vimentin in astrocytes and ependymal
cells of the developing and adult mouse nervous system. J. Cell Biol. 90, 435-447.
Shaw, G., Osborn, M. and Weber, K. (1981) An immunofluorescence microscopical study of the neurofilament triplet proteins, vi-
mentin and glial fibrillary acidic protein within the adult brain.
Eur. J. Cell. Biol. 26, 68-82.

K. Krohn et al. /Journal of Neuroimmunology 56 (I 995) 53-63 63
Sinclair, J.D. (1988) Multiple t-tests are appropriate in science. Trends Pharmacol. Sci. 9, 12-13.
Steward, O., Torre, E.R., Tomasulo, R. and Lothman, E. (1991) Neuronal activity up-regulates astroglial gene expression. Proc. Natl. Acad. Sci. USA 88, 6819-6823.
Streit, W.J. (1990) An improved staining method for rat microghal cells using the lectin from Griffonia simplicifolia (GSA I-B4). J. Histochem. Cytochem. 38, 1683-1686.
Streit, W.J., Graeber, M.B. and Kreutzberg, G.W. (1988) Functional plasticity of microglia: a review. Glia 1, 301-307.
Teichmann-Weinberg, A., Littauer, U.Z. and Ginzburg, I. (1988) The inhibition of neurite outgrowth in PC12 cells by tubulin antisense oligodeoxyribonucleotides. Gene 72, 297-307.
Unsicker, K., Flanders, K.C., Cissel, D., Lafyatis, R. and Sporn, M. (1991) Transforming growth factor beta isofonns in the adult rat central and peripheral nervous system. Neuroscience 44, 613-625.
Van der Wal, E.A., Gomez-Pinilla, F. and Cotman, C.W. (1993) Transforming growth factor-p1 is in plaques in Alzheimer and Down pathologies. NeuroReport 4, 69-72.
Yamada, T., Kawamata, T., Walker, D.G. and McGeer, P.L. (1992) Vimentin immunoreactivity in normal and pathological human brain tissue. Acta Neuropathol. 84, 157-162.
Yoshida, K. and Gage, F.H. (1992) Cooperative regulation of nerve growth factor synthesis and secretion in fibroblasts and astrocytes by fibroblast growth factor and other cytokines. Brain Res. 569, 14-25.
Young-Chan, C.S., Laping, N.J., Morgan, T.E., Huang, C.J., Finch, C.E., and Nichols, N.R. (1993) Autoregulation of transforming growth factor+1 mRNA in the brain by intraventricular infusion in adult male rats. Sot. Neurosci. Abstr. 19. 1722.





























