Embed Size (px)
Citation preview

NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 393
A R T I C L E S
FAPPs control Golgi-to-cell-surface membrane trafficby binding to ARF and PtdIns(4)PAnna Godi1,4, Antonella Di Campli1,4, Athanasios Konstantakopoulos2, Giuseppe Di Tullio1, Dario R. Alessi3,Gursant S. Kular3, Tiziana Daniele1, Pierfrancesco Marra1, John M. Lucocq2 and M. Antonietta De Matteis1,5
The molecular mechanisms underlying the formation of carriers trafficking from the Golgi complex to the cell surface are stillill-defined; nevertheless, the involvement of a lipid-based machinery is well established. This includes phosphatidylinositol4-phosphate (PtdIns(4)P), the precursor for phosphatidylinositol 4,5-bisphosphate (PtdIns(4,5)P2). In yeast, PtdIns(4)Pexerts a direct role, however, its mechanism of action and its targets in mammalian cells remain uncharacterized. We haveidentified two effectors of PtdIns(4)P, the four-phosphate-adaptor protein 1 and 2 (FAPP1 and FAPP2). Both proteins localizeto the trans-Golgi network (TGN) on nascent carriers, and interact with PtdIns(4)P and the small GTPase ADP-ribosylationfactor (ARF) through their plekstrin homology (PH) domain. Displacement or knockdown of FAPPs inhibits cargo transfer tothe plasma membrane. Moreover, overexpression of FAPP-PH impairs carrier fission. Therefore, FAPPs are essentialcomponents of a PtdIns(4)P- and ARF-regulated machinery that controls generation of constitutive post-Golgi carriers.
The transport of lipids and proteins from the Golgi complex to the cellsurface is performed by carriers that are generated at the TGN, movecentrifugally along microtubules, and finally fuse with the plasmamembrane1–3. The molecular machinery responsible for the genera-tion of these constitutive post-Golgi carriers is only partially understood. However, the importance of a lipid-based machinery inTGN-to-plasma membrane transport is well established in both mam-mals and yeast. In yeast, the role of the phosphoinositide-transfer pro-tein Sec14 and diacylglycerol (DAG) in transport out of the Golgicomplex has been recognized since 1989 (ref. 4). In mammals, phos-phoinositide-transfer protein is required for vesicle budding at theTGN (refs 5, 6), and DAG levels control protein transport from theTGN to the cell surface7.
Other lipids with a recognized role in the Golgi complex are thephosphoinositides8. Among these, PtdIns(4)P — the product of phos-phatidylinositol 4-kinase (PI(4)K) activity and the precursor ofPtdIns(4,5)P2 — has been shown to be involved in membrane trans-port from the Golgi complex to the plasma membrane in yeast9–12. Inmammals, the Golgi complex contains PI(4)KIIα (ref. 13),PI(4)KIIIα (ref. 14) and PI(4)KIIIβ (ref. 15), with PI(4)KIIIβ beingrecruited to the Golgi complex in response to ARF activation16.Consequently, interfering with PI(4)KIIIβ induces structural changesin the Golgi complex16 and pleiotropic secretory defects in polarizedcells17, and PI(4)KIIα knock-out induces the redistribution of themannose 6-phosphate receptor and inhibition of protein transport tothe plasma membrane18.
Despite this recognized role for PI(4)K activity in the Golgi complex,key questions such as where and how PtdIns(4)P is involved in
TGN-to-plasma membrane transport remain unanswered and thePtdIns(4)P targets remain to be identified.
Here, we have cloned and defined the functional role of FAPP1 andFAPP2, two effectors of PtdIns(4)P at the TGN. The FAPPs can bindthrough their PH domain to PtdIns(4)P and ARF, and subsequentlylocalize to TGN exit sites, where they control the process of carrier for-mation. We propose that the main role of FAPPs is to ensure co-ordi-nation of the budding and fission reactions, which are required for thegeneration of mature carriers that are properly targeted and competentfor fusion with the plasma membrane.
RESULTSFAPP1 and FAPP2 localize to the TGNThe full-length cDNA of FAPP1 (GenBank accession number:AF286162) and FAPP2 (GenBank accession number: AF380162)was cloned. FAPP1 was originally identified through the screeningof PH-domain-containing proteins and named FAPP1, because itsPH domain can bind selectively to PtdIns(4)P (ref. 19). HumanFAPP1 (hFAPP1) is a 300-amino-acid protein with an amino-ter-minal PH domain and a putative proline-rich motif at the carboxylterminus; the hFAPP1 gene is localized on chromosome 2. hFAPP2is a 507-amino-acid protein, encoded by a gene located on chromo-some 7. The hFAPP2 protein is 80% identical and 90% similar tohFAPP1 at the N terminus, and differs in the C terminus due to aputative glycolipid transfer protein (GLTP) domain (ref. 20). FAPPsare ubiquitously expressed and their mRNA have been isolatedfrom many different sources and species, including Zebra fish(Danio rerio) and Xenopus laevis.
1Department of Cell Biology and Oncology, Consorzio Mario Negri Sud, via Nazionale, 66030 Santa Maria Imbaro (CH), Italy. 2MSI/WTB Complex, University ofDundee, Dundee, UK. 3Division of Cell Signalling, University of Dundee, Dundee DD1 5EH, UK. 4These authors contributed equally to this work. 5Correspondenceshould be addressed to M.A.D.M. (e-mail: [email protected]).
Published online: 25 April 2004; DOI: 10.1038/ncb1119
print ncb1119 15/4/04 5:18 PM Page 393
© 2004 Nature Publishing Group
A RT I C L E S
394 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
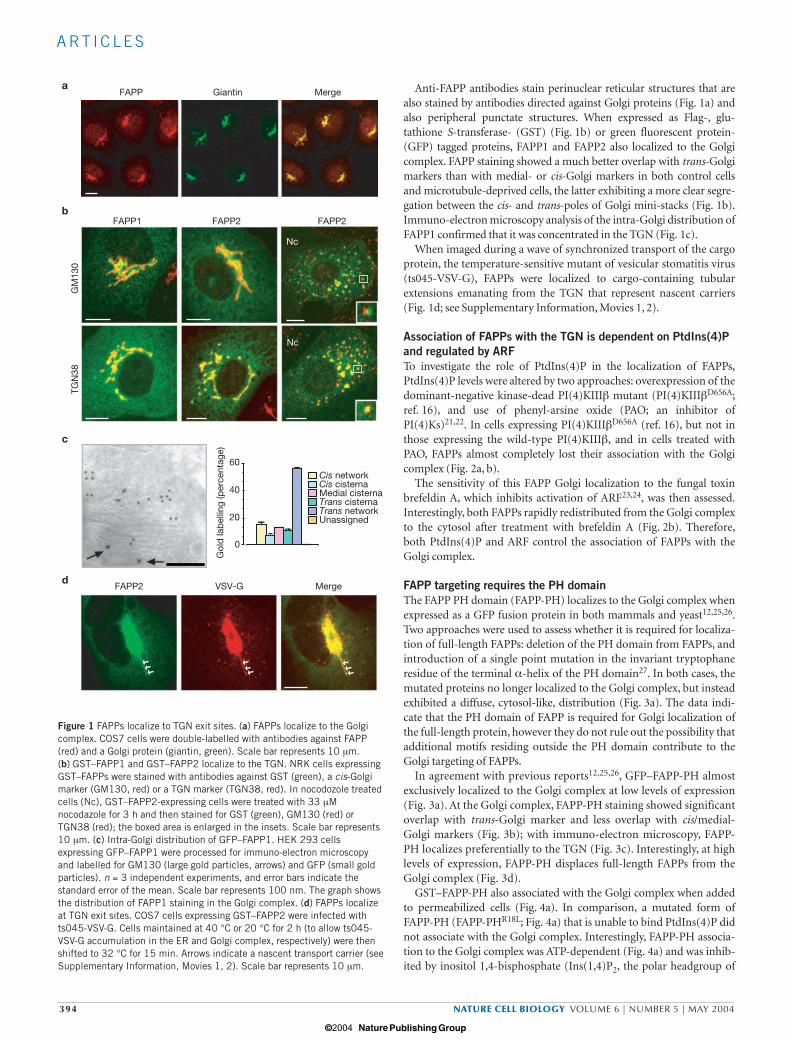
Anti-FAPP antibodies stain perinuclear reticular structures that arealso stained by antibodies directed against Golgi proteins (Fig. 1a) andalso peripheral punctate structures. When expressed as Flag-, glu-tathione S-transferase- (GST) (Fig. 1b) or green fluorescent protein-(GFP) tagged proteins, FAPP1 and FAPP2 also localized to the Golgicomplex. FAPP staining showed a much better overlap with trans-Golgimarkers than with medial- or cis-Golgi markers in both control cellsand microtubule-deprived cells, the latter exhibiting a more clear segre-gation between the cis- and trans-poles of Golgi mini-stacks (Fig. 1b).Immuno-electron microscopy analysis of the intra-Golgi distribution ofFAPP1 confirmed that it was concentrated in the TGN (Fig. 1c).
When imaged during a wave of synchronized transport of the cargoprotein, the temperature-sensitive mutant of vesicular stomatitis virus(ts045-VSV-G), FAPPs were localized to cargo-containing tubularextensions emanating from the TGN that represent nascent carriers(Fig. 1d; see Supplementary Information, Movies 1, 2).
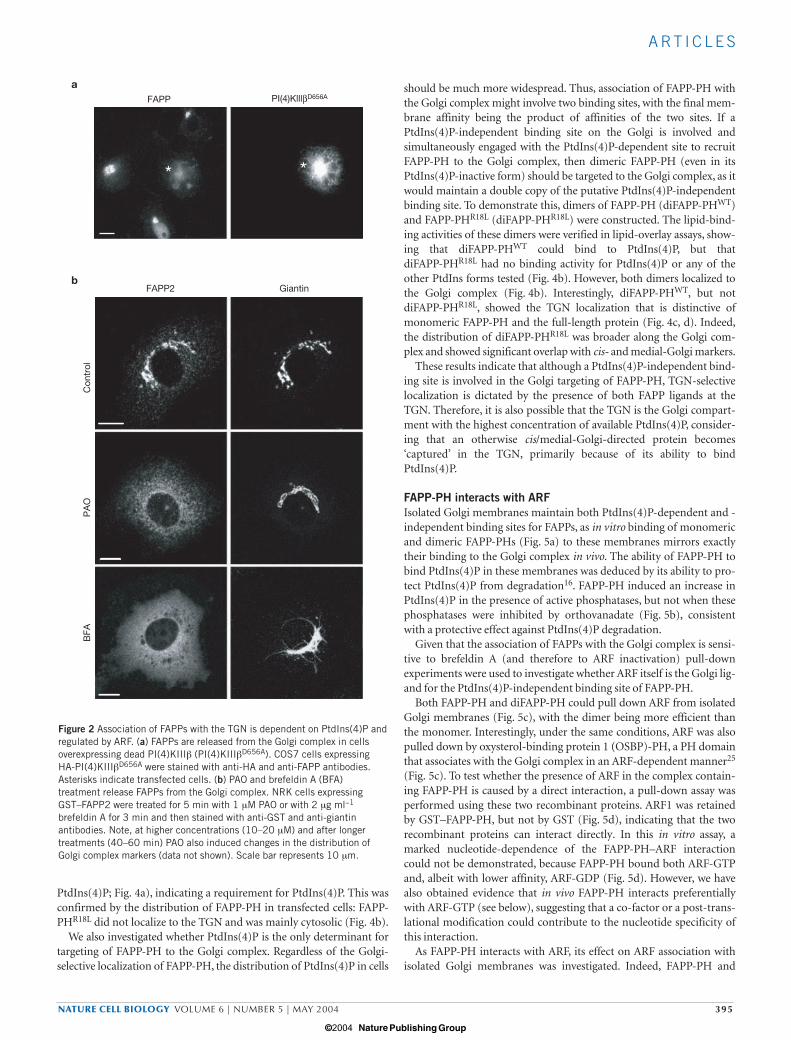
Association of FAPPs with the TGN is dependent on PtdIns(4)Pand regulated by ARFTo investigate the role of PtdIns(4)P in the localization of FAPPs,PtdIns(4)P levels were altered by two approaches: overexpression of thedominant-negative kinase-dead PI(4)KIIIβ mutant (PI(4)KIIIβD656A;ref. 16), and use of phenyl-arsine oxide (PAO; an inhibitor ofPI(4)Ks)21,22. In cells expressing PI(4)KIIIβD656A (ref. 16), but not inthose expressing the wild-type PI(4)KIIIβ, and in cells treated withPAO, FAPPs almost completely lost their association with the Golgicomplex (Fig. 2a, b).
The sensitivity of this FAPP Golgi localization to the fungal toxinbrefeldin A, which inhibits activation of ARF23,24, was then assessed.Interestingly, both FAPPs rapidly redistributed from the Golgi complexto the cytosol after treatment with brefeldin A (Fig. 2b). Therefore,both PtdIns(4)P and ARF control the association of FAPPs with theGolgi complex.
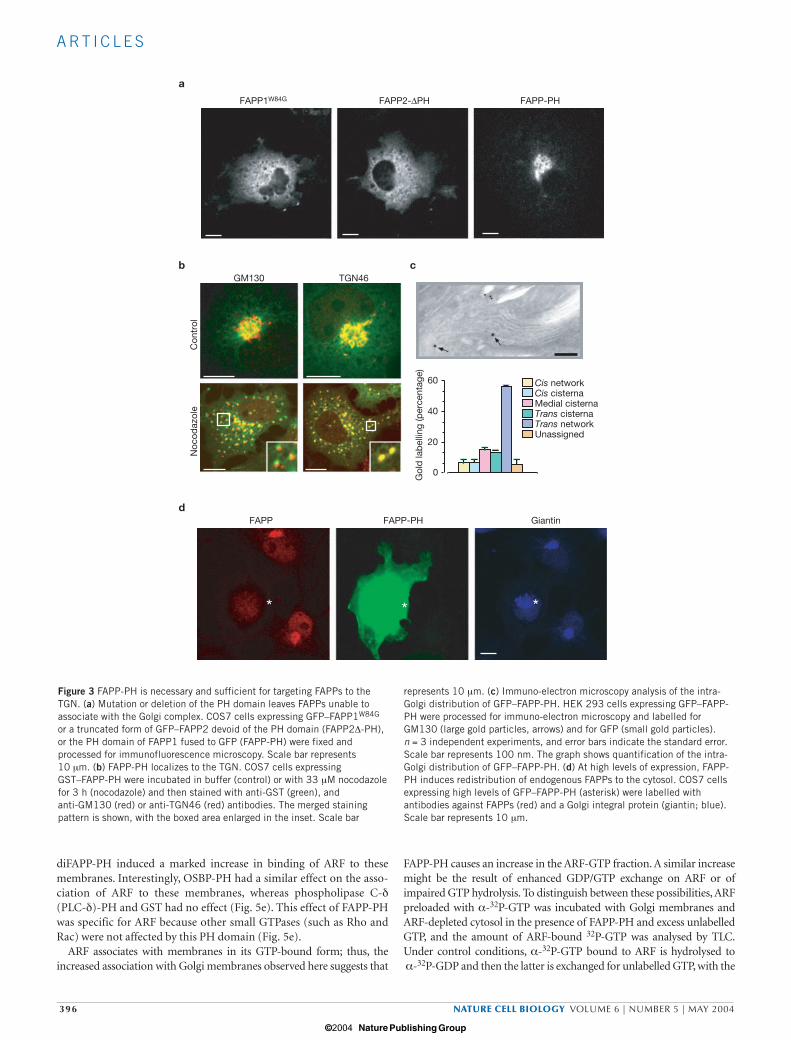
FAPP targeting requires the PH domainThe FAPP PH domain (FAPP-PH) localizes to the Golgi complex whenexpressed as a GFP fusion protein in both mammals and yeast12,25,26.Two approaches were used to assess whether it is required for localiza-tion of full-length FAPPs: deletion of the PH domain from FAPPs, andintroduction of a single point mutation in the invariant tryptophaneresidue of the terminal α-helix of the PH domain27. In both cases, themutated proteins no longer localized to the Golgi complex, but insteadexhibited a diffuse, cytosol-like, distribution (Fig. 3a). The data indi-cate that the PH domain of FAPP is required for Golgi localization ofthe full-length protein, however they do not rule out the possibility thatadditional motifs residing outside the PH domain contribute to theGolgi targeting of FAPPs.
In agreement with previous reports12,25,26, GFP–FAPP-PH almostexclusively localized to the Golgi complex at low levels of expression(Fig. 3a). At the Golgi complex, FAPP-PH staining showed significantoverlap with trans-Golgi marker and less overlap with cis/medial-Golgi markers (Fig. 3b); with immuno-electron microscopy, FAPP-PH localizes preferentially to the TGN (Fig. 3c). Interestingly, at highlevels of expression, FAPP-PH displaces full-length FAPPs from theGolgi complex (Fig. 3d).
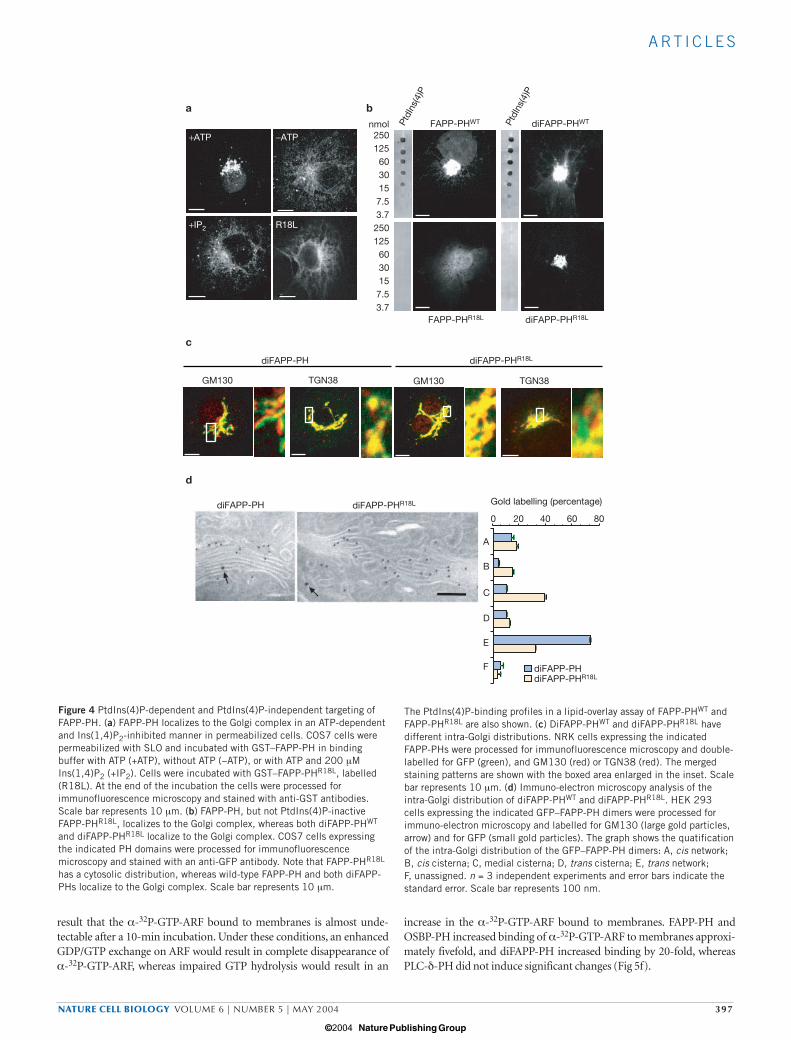
GST–FAPP-PH also associated with the Golgi complex when addedto permeabilized cells (Fig. 4a). In comparison, a mutated form ofFAPP-PH (FAPP-PHR18L; Fig. 4a) that is unable to bind PtdIns(4)P didnot associate with the Golgi complex. Interestingly, FAPP-PH associa-tion to the Golgi complex was ATP-dependent (Fig. 4a) and was inhib-ited by inositol 1,4-bisphosphate (Ins(1,4)P2, the polar headgroup of
a
b
c
d
FAPP Giantin Merge
FAPP2
TGN
38G
M13
0
FAPP1
Nc
FAPP2
Nc
60
40
20
0
Gol
d la
bel
ling
(per
cent
age)
Cis networkCis cisternaMedial cisterna Trans cisterna Trans networkUnassigned
FAPP2 VSV-G Merge
Figure 1 FAPPs localize to TGN exit sites. (a) FAPPs localize to the Golgicomplex. COS7 cells were double-labelled with antibodies against FAPP(red) and a Golgi protein (giantin, green). Scale bar represents 10 µm. (b) GST–FAPP1 and GST–FAPP2 localize to the TGN. NRK cells expressingGST–FAPPs were stained with antibodies against GST (green), a cis-Golgimarker (GM130, red) or a TGN marker (TGN38, red). In nocodozole treatedcells (Nc), GST–FAPP2-expressing cells were treated with 33 µMnocodazole for 3 h and then stained for GST (green), GM130 (red) orTGN38 (red); the boxed area is enlarged in the insets. Scale bar represents10 µm. (c) Intra-Golgi distribution of GFP–FAPP1. HEK 293 cellsexpressing GFP–FAPP1 were processed for immuno-electron microscopyand labelled for GM130 (large gold particles, arrows) and GFP (small goldparticles). n = 3 independent experiments, and error bars indicate thestandard error of the mean. Scale bar represents 100 nm. The graph showsthe distribution of FAPP1 staining in the Golgi complex. (d) FAPPs localizeat TGN exit sites. COS7 cells expressing GST–FAPP2 were infected withts045-VSV-G. Cells maintained at 40 °C or 20 °C for 2 h (to allow ts045-VSV-G accumulation in the ER and Golgi complex, respectively) were thenshifted to 32 °C for 15 min. Arrows indicate a nascent transport carrier (seeSupplementary Information, Movies 1, 2). Scale bar represents 10 µm.
print ncb1119 15/4/04 5:18 PM Page 394
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 395
PtdIns(4)P; Fig. 4a), indicating a requirement for PtdIns(4)P. This wasconfirmed by the distribution of FAPP-PH in transfected cells: FAPP-PHR18L did not localize to the TGN and was mainly cytosolic (Fig. 4b).
We also investigated whether PtdIns(4)P is the only determinant fortargeting of FAPP-PH to the Golgi complex. Regardless of the Golgi-selective localization of FAPP-PH, the distribution of PtdIns(4)P in cells
should be much more widespread. Thus, association of FAPP-PH withthe Golgi complex might involve two binding sites, with the final mem-brane affinity being the product of affinities of the two sites. If aPtdIns(4)P-independent binding site on the Golgi is involved andsimultaneously engaged with the PtdIns(4)P-dependent site to recruitFAPP-PH to the Golgi complex, then dimeric FAPP-PH (even in itsPtdIns(4)P-inactive form) should be targeted to the Golgi complex, as itwould maintain a double copy of the putative PtdIns(4)P-independentbinding site. To demonstrate this, dimers of FAPP-PH (diFAPP-PHWT)and FAPP-PHR18L (diFAPP-PHR18L) were constructed. The lipid-bind-ing activities of these dimers were verified in lipid-overlay assays, show-ing that diFAPP-PHWT could bind to PtdIns(4)P, but thatdiFAPP-PHR18L had no binding activity for PtdIns(4)P or any of theother PtdIns forms tested (Fig. 4b). However, both dimers localized tothe Golgi complex (Fig. 4b). Interestingly, diFAPP-PHWT, but notdiFAPP-PHR18L, showed the TGN localization that is distinctive ofmonomeric FAPP-PH and the full-length protein (Fig. 4c, d). Indeed,the distribution of diFAPP-PHR18L was broader along the Golgi com-plex and showed significant overlap with cis- and medial-Golgi markers.
These results indicate that although a PtdIns(4)P-independent bind-ing site is involved in the Golgi targeting of FAPP-PH, TGN-selectivelocalization is dictated by the presence of both FAPP ligands at theTGN. Therefore, it is also possible that the TGN is the Golgi compart-ment with the highest concentration of available PtdIns(4)P, consider-ing that an otherwise cis/medial-Golgi-directed protein becomes‘captured’ in the TGN, primarily because of its ability to bindPtdIns(4)P.
FAPP-PH interacts with ARFIsolated Golgi membranes maintain both PtdIns(4)P-dependent and -independent binding sites for FAPPs, as in vitro binding of monomericand dimeric FAPP-PHs (Fig. 5a) to these membranes mirrors exactlytheir binding to the Golgi complex in vivo. The ability of FAPP-PH tobind PtdIns(4)P in these membranes was deduced by its ability to pro-tect PtdIns(4)P from degradation16. FAPP-PH induced an increase inPtdIns(4)P in the presence of active phosphatases, but not when thesephosphatases were inhibited by orthovanadate (Fig. 5b), consistentwith a protective effect against PtdIns(4)P degradation.
Given that the association of FAPPs with the Golgi complex is sensi-tive to brefeldin A (and therefore to ARF inactivation) pull-downexperiments were used to investigate whether ARF itself is the Golgi lig-and for the PtdIns(4)P-independent binding site of FAPP-PH.
Both FAPP-PH and diFAPP-PH could pull down ARF from isolatedGolgi membranes (Fig. 5c), with the dimer being more efficient thanthe monomer. Interestingly, under the same conditions, ARF was alsopulled down by oxysterol-binding protein 1 (OSBP)-PH, a PH domainthat associates with the Golgi complex in an ARF-dependent manner25
(Fig. 5c). To test whether the presence of ARF in the complex contain-ing FAPP-PH is caused by a direct interaction, a pull-down assay wasperformed using these two recombinant proteins. ARF1 was retainedby GST–FAPP-PH, but not by GST (Fig. 5d), indicating that the tworecombinant proteins can interact directly. In this in vitro assay, amarked nucleotide-dependence of the FAPP-PH–ARF interactioncould not be demonstrated, because FAPP-PH bound both ARF-GTPand, albeit with lower affinity, ARF-GDP (Fig. 5d). However, we havealso obtained evidence that in vivo FAPP-PH interacts preferentiallywith ARF-GTP (see below), suggesting that a co-factor or a post-trans-lational modification could contribute to the nucleotide specificity ofthis interaction.
As FAPP-PH interacts with ARF, its effect on ARF association withisolated Golgi membranes was investigated. Indeed, FAPP-PH and
a
bGiantinFAPP2
FAPPC
ontr
olP
AO
BFA
* *
PI(4)KlllβD656A
Figure 2 Association of FAPPs with the TGN is dependent on PtdIns(4)P andregulated by ARF. (a) FAPPs are released from the Golgi complex in cellsoverexpressing dead PI(4)KIIIβ (PI(4)KIIIβD656A). COS7 cells expressingHA-PI(4)KIIIβD656A were stained with anti-HA and anti-FAPP antibodies.Asterisks indicate transfected cells. (b) PAO and brefeldin A (BFA)treatment release FAPPs from the Golgi complex. NRK cells expressingGST–FAPP2 were treated for 5 min with 1 µM PAO or with 2 µg ml−1
brefeldin A for 3 min and then stained with anti-GST and anti-giantinantibodies. Note, at higher concentrations (10–20 µM) and after longertreatments (40–60 min) PAO also induced changes in the distribution ofGolgi complex markers (data not shown). Scale bar represents 10 µm.
print ncb1119 15/4/04 5:19 PM Page 395
© 2004 Nature Publishing Group
A RT I C L E S
396 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
diFAPP-PH induced a marked increase in binding of ARF to thesemembranes. Interestingly, OSBP-PH had a similar effect on the asso-ciation of ARF to these membranes, whereas phospholipase C-δ(PLC-δ)-PH and GST had no effect (Fig. 5e). This effect of FAPP-PHwas specific for ARF because other small GTPases (such as Rho andRac) were not affected by this PH domain (Fig. 5e).
ARF associates with membranes in its GTP-bound form; thus, theincreased association with Golgi membranes observed here suggests that
FAPP-PH causes an increase in the ARF-GTP fraction. A similar increasemight be the result of enhanced GDP/GTP exchange on ARF or ofimpaired GTP hydrolysis. To distinguish between these possibilities, ARFpreloaded with α-32P-GTP was incubated with Golgi membranes andARF-depleted cytosol in the presence of FAPP-PH and excess unlabelledGTP, and the amount of ARF-bound 32P-GTP was analysed by TLC.Under control conditions, α-32P-GTP bound to ARF is hydrolysed toα-32P-GDP and then the latter is exchanged for unlabelled GTP, with the
c
Con
trol
Noc
odaz
ole
GM130b
a
TGN46
d
FAPP1W84G FAPP2-∆PH FAPP-PH
FAPP-PH
*
FAPP
*
Giantin
*
60
40
20
0Gol
d la
bel
ling
(per
cent
age)
Cis networkCis cisternaMedial cisterna Trans cisterna Trans networkUnassigned
Figure 3 FAPP-PH is necessary and sufficient for targeting FAPPs to theTGN. (a) Mutation or deletion of the PH domain leaves FAPPs unable toassociate with the Golgi complex. COS7 cells expressing GFP–FAPP1W84G
or a truncated form of GFP–FAPP2 devoid of the PH domain (FAPP2∆-PH),or the PH domain of FAPP1 fused to GFP (FAPP-PH) were fixed andprocessed for immunofluorescence microscopy. Scale bar represents10 µm. (b) FAPP-PH localizes to the TGN. COS7 cells expressingGST–FAPP-PH were incubated in buffer (control) or with 33 µM nocodazolefor 3 h (nocodazole) and then stained with anti-GST (green), and anti-GM130 (red) or anti-TGN46 (red) antibodies. The merged stainingpattern is shown, with the boxed area enlarged in the inset. Scale bar
represents 10 µm. (c) Immuno-electron microscopy analysis of the intra-Golgi distribution of GFP–FAPP-PH. HEK 293 cells expressing GFP–FAPP-PH were processed for immuno-electron microscopy and labelled forGM130 (large gold particles, arrows) and for GFP (small gold particles).n = 3 independent experiments, and error bars indicate the standard error.Scale bar represents 100 nm. The graph shows quantification of the intra-Golgi distribution of GFP–FAPP-PH. (d) At high levels of expression, FAPP-PH induces redistribution of endogenous FAPPs to the cytosol. COS7 cellsexpressing high levels of GFP–FAPP-PH (asterisk) were labelled withantibodies against FAPPs (red) and a Golgi integral protein (giantin; blue).Scale bar represents 10 µm.
print ncb1119 15/4/04 5:19 PM Page 396
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 397
result that the α-32P-GTP-ARF bound to membranes is almost unde-tectable after a 10-min incubation. Under these conditions, an enhancedGDP/GTP exchange on ARF would result in complete disappearance ofα-32P-GTP-ARF, whereas impaired GTP hydrolysis would result in an
increase in the α-32P-GTP-ARF bound to membranes. FAPP-PH andOSBP-PH increased binding of α-32P-GTP-ARF to membranes approxi-mately fivefold, and diFAPP-PH increased binding by 20-fold, whereasPLC-δ-PH did not induce significant changes (Fig 5f).
b
250125603015
7.53.7
250125603015
7.53.7
d
c
FAPP-PHWT
FAPP-PHR18L
diFAPP-PHWT
diFAPP-PHR18L
a
806040200
Gold labelling (percentage)
diFAPP-PHdiFAPP-PHR18L
A
B
C
D
E
F
diFAPP-PH diFAPP-PHR18L
nmol
TGN38GM130 TGN38GM130
diFAPP-PH diFAPP-PHR18L
+ATP −ATP
+IP2 R18L
PtdI
ns(4
)P
PtdI
ns(4
)P
Figure 4 PtdIns(4)P-dependent and PtdIns(4)P-independent targeting ofFAPP-PH. (a) FAPP-PH localizes to the Golgi complex in an ATP-dependentand Ins(1,4)P2-inhibited manner in permeabilized cells. COS7 cells werepermeabilized with SLO and incubated with GST–FAPP-PH in bindingbuffer with ATP (+ATP), without ATP (−ATP), or with ATP and 200 µMIns(1,4)P2 (+IP2). Cells were incubated with GST–FAPP-PHR18L, labelled(R18L). At the end of the incubation the cells were processed forimmunofluorescence microscopy and stained with anti-GST antibodies.Scale bar represents 10 µm. (b) FAPP-PH, but not PtdIns(4)P-inactiveFAPP-PHR18L, localizes to the Golgi complex, whereas both diFAPP-PHWT
and diFAPP-PHR18L localize to the Golgi complex. COS7 cells expressingthe indicated PH domains were processed for immunofluorescencemicroscopy and stained with an anti-GFP antibody. Note that FAPP-PHR18L
has a cytosolic distribution, whereas wild-type FAPP-PH and both diFAPP-PHs localize to the Golgi complex. Scale bar represents 10 µm.
The PtdIns(4)P-binding profiles in a lipid-overlay assay of FAPP-PHWT andFAPP-PHR18L are also shown. (c) DiFAPP-PHWT and diFAPP-PHR18L havedifferent intra-Golgi distributions. NRK cells expressing the indicatedFAPP-PHs were processed for immunofluorescence microscopy and double-labelled for GFP (green), and GM130 (red) or TGN38 (red). The mergedstaining patterns are shown with the boxed area enlarged in the inset. Scalebar represents 10 µm. (d) Immuno-electron microscopy analysis of theintra-Golgi distribution of diFAPP-PHWT and diFAPP-PHR18L. HEK 293cells expressing the indicated GFP–FAPP-PH dimers were processed forimmuno-electron microscopy and labelled for GM130 (large gold particles,arrow) and for GFP (small gold particles). The graph shows the quatificationof the intra-Golgi distribution of the GFP–FAPP-PH dimers: A, cis network;B, cis cisterna; C, medial cisterna; D, trans cisterna; E, trans network; F, unassigned. n = 3 independent experiments and error bars indicate thestandard error. Scale bar represents 100 nm.
print ncb1119 15/4/04 5:19 PM Page 397
© 2004 Nature Publishing Group
A RT I C L E S
398 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
a b c
d e
GAP1
GST
diFA
PP-P
HW
T
diFA
PP-P
HR
18L
FAPP
1-PH
WT
FAPP
-PH
R18
L
GST
diFA
PP-P
HW
T
diFA
PP-P
HR1
8L
PLCδ-
PHFA
PP-P
HR1
8L
OSB
P-PH
*
GST
FAPP
-PH
OSB
P-PH
diFA
PP-P
HW
T
diFA
PP-P
HR
18L
FAPP
-PH
WT
FAPP
-PH
R18
L
35
50
GDP GTP
GST FAPP-PH
GDP GTP
Rac
Rho
GST
FAPP
-PH
g
i
f hGSTFAPP-PHdiFAPP-PHWT
PLCδ-PH
1
0.75
0.50
0.25
00 2.5 10
GA
P a
ctiv
ity o
n A
RF
Time (min)
200100
0
400300
500600
2,000
32P
-GTP
-AR
F(p
erce
ntag
e of
con
trol
)
diFA
PP-P
HW
T
diFA
PP-P
HW
T
ARF
ARF1WT ARF1T31N
j
GST
diFAPP-PH
FAPP-PH
GST
0
50
100
150
500
600
ARF
ARF
GSTFAPP-PHPLCδ-PHVan+GSTVan+FAPP-PH
GSTFAPP-PHPLCδ-PHOSBP-PHdiFAPP-PHWT
FAPP-PHARF
*
*
*
*
* *
Mr(K)
32P
-Ptd
Ins(
4)P
(per
cent
age
of c
ontr
ol)
FAPP
-PH
WT
Figure 5 FAPP-PH interacts with the small GTPase ARF. (a) FAPP-PHbinds to isolated Golgi membranes. Golgi fractions were incubated withcytosol and 1.5 µM GST or the indicated GST–FAPP PH domains for 10min at 37 °C. Membrane-associated proteins were analysed by westernblotting. The level of FAPP-PHR18L bound to the Golgi fractions equalsthat of GST and is thus considered as background binding. The lower-molecular-weight bands in the presence of diFAPP-PHs (asterisks) aredegradation products. (b) FAPP-PH binds PtdIns(4)P. Golgi fractions wereincubated with cytosol, 1 µCi 32P-ATP, the indicated proteins, and thephosphatase inhibitor orthovanadate (Van, 500 µM). The lipids wereanalysed by thin layer chromatography (TLC). Quantification of 32P-PtdIns(4)P is shown. The data, expressed as percentage of control (GST),represent the mean ± SD of three experiments. (c) ARF1 is pulled down byGST–FAPP-PH and GST–OSBP-PH. Golgi fractions were incubated withthe indicated proteins immobilized on beads, and retained ARF1 wasassessed by western blotting. (d) FAPP-PH binds to ARF1. GTP- or GDP-loaded ARF1 retained on GST- or GST–FAPP-PH-coated beads wasassessed by western blotting. (e) FAPP-PHs enhance the association of
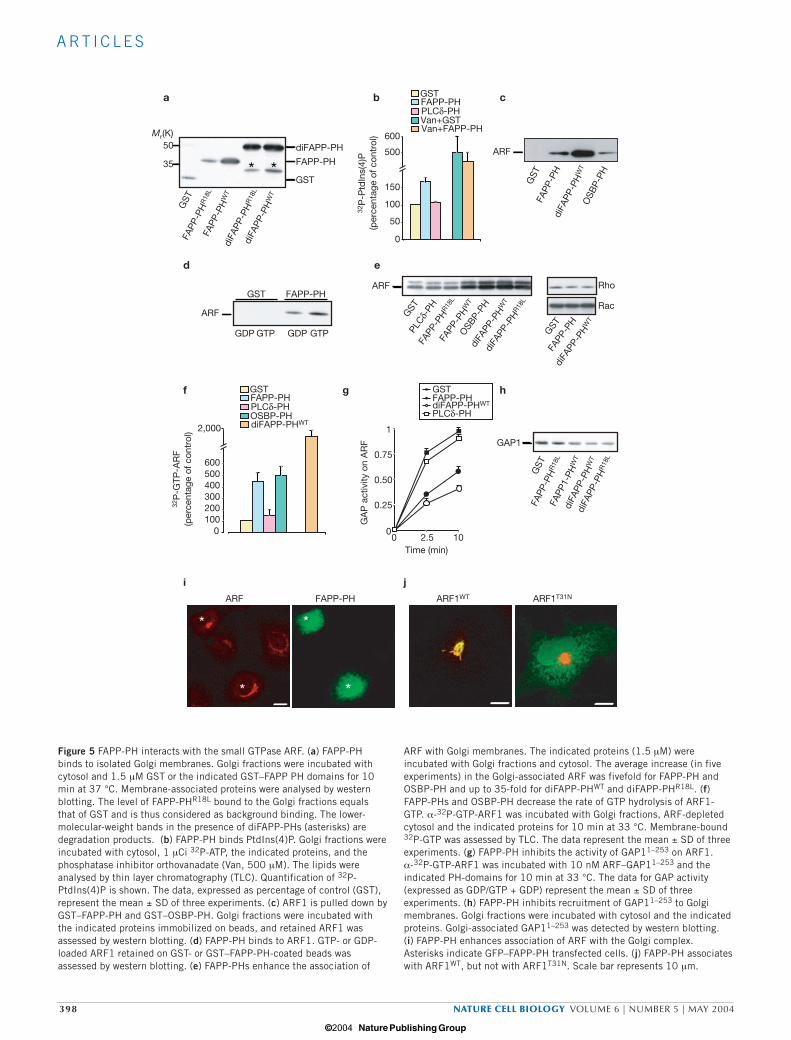
ARF with Golgi membranes. The indicated proteins (1.5 µM) wereincubated with Golgi fractions and cytosol. The average increase (in fiveexperiments) in the Golgi-associated ARF was fivefold for FAPP-PH andOSBP-PH and up to 35-fold for diFAPP-PHWT and diFAPP-PHR18L. (f)FAPP-PHs and OSBP-PH decrease the rate of GTP hydrolysis of ARF1-GTP. α-32P-GTP-ARF1 was incubated with Golgi fractions, ARF-depletedcytosol and the indicated proteins for 10 min at 33 °C. Membrane-bound32P-GTP was assessed by TLC. The data represent the mean ± SD of threeexperiments. (g) FAPP-PH inhibits the activity of GAP11–253 on ARF1.α-32P-GTP-ARF1 was incubated with 10 nM ARF–GAP11–253 and theindicated PH-domains for 10 min at 33 °C. The data for GAP activity(expressed as GDP/GTP + GDP) represent the mean ± SD of threeexperiments. (h) FAPP-PH inhibits recruitment of GAP11–253 to Golgimembranes. Golgi fractions were incubated with cytosol and the indicatedproteins. Golgi-associated GAP11–253 was detected by western blotting. (i) FAPP-PH enhances association of ARF with the Golgi complex.Asterisks indicate GFP–FAPP-PH transfected cells. (j) FAPP-PH associateswith ARF1WT, but not with ARF1T31N. Scale bar represents 10 µm.
print ncb1119 15/4/04 5:19 PM Page 398
© 2004 Nature Publishing Group
A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 399
ARF has no detectable intrinsic GTPase activity and so relies on theGTPase-activating proteins (GAPs )28 for this activity. To test whetherthe impairment of GTP hydrolysis induced by FAPP-PH is because ofits interference with the ARF–GAP interaction, the effects of FAPP-PHon the recruitment of ARF–GAP1, and on the GAP activity of recombi-nant GAP1 (amino acids 1–253, GAP11–253) toward ARF129, wasassessed. FAPP-PH both inhibited the recruitment of ARF–GAP11–253
to Golgi membranes (Fig. 5h) and reduced the GAP activity (Fig. 5g).A similar effect was observed with diFAPP-PH (Fig. 5g) and OSBP-PH(not shown), but not with PLC-δ-PH and GST (Fig. 5g).
These in vitro experiments indicate that FAPP-PH binds ARF-GTPand stabilizes its membrane-bound form by interfering with its inter-action with ARF–GAP. Similar effects on ARF have been reported for
other ARF-interacting proteins/domains, such as GGAs (Golgi-localiz-ing, γ adaptin ear homology domain, ARF-binding protein) and VHS-GAT (Vps27, HRS and STAM domain-GGA and TOM domain)30,31.Thus, these ARF-stabilizing effects of FAPP-PH could provide a usefulmeasure of the ARF–FAPP-PH interaction, and be used to verify thisinteraction in intact cells. Indeed, FAPP-PH enhanced the associationof ARF with the Golgi complex at the expense of the cytosolic pool ofARF (Fig. 5i), which is normally responsible for the high backgroundin the ARF staining pattern. As FAPP-PH can interact in vitro with bothGDP- and GTP-bound forms of ARF, we examined whether this alsooccurs in vivo. Thus, the co-localization and stabilization of ARF on theGolgi complex was evaluated in cells expressing FAPP-PH and ARF1 orARF1T31N (locked in the GDP-bound form) at very low levels (that is,
a
b c
d
Control siRNA
FAP
P
Control siRNA
siR
NA
Con
trol
FAPP267-
FAPP138-
ARF20-
Control siRNA
VS
V-G
VS
V-G
Time (min)
GA
G r
elea
se
(per
cent
age
of t
otal
)
siRNA
Control50
40
30
20
10
00 10 15 20 30
e f
Time (min)
Sur
face
VS
V-G
(per
cent
age
of t
otal
)
00
4
8
12
16
20
30 45
Control
siRNA
End
o-H
res
ista
nt(p
erce
ntag
e of
tot
al)
siRNACcontrol
100
0
20
40
60
80
Mr(K)
Figure 6 FAPPs are required for TGN-to-plasma membrane transport. (a) Effects of FAPP siRNA treatment on FAPP levels. COS7 cells weretreated with FAPP or control siRNAs for 72 h and processed forimmunofluorescence microscopy or western blotting with anti-FAPPantibodies. As controls, the levels of ARF protein are shown. (b) Transportof ts045-VSV-G to the plasma membrane is inhibited in FAPP siRNA-treated cells. Control and FAPP siRNA-treated COS7 cells were infectedwith ts045-VSV, maintained at 40 °C for 2 h, shifted to 32 °C for 60 min,and then fixed and processed for immunofluorescence microscopy with ananti-VSV-G antibody. (c) TGN-to-plasma membrane transport of ts045-VSV-G is inhibited in FAPP siRNA-treated cells. Control and FAPP siRNA-treatedCOS7 cells were infected with ts045-VSV, maintained at 40 °C for 2 h, at20 °C for 2 h, shifted to 32 °C for 60 min, and then fixed and processedfor immunofluorescence microscopy with an anti-VSV-G antibody. (d) ER-to-Golgi complex transport of ts045-VSV-G proceeds at similar rates incontrol and FAPP siRNA-treated cells. Control siRNA- and FAPP siRNA-treated COS7 cells were infected with ts045-VSV-G, pulsed with
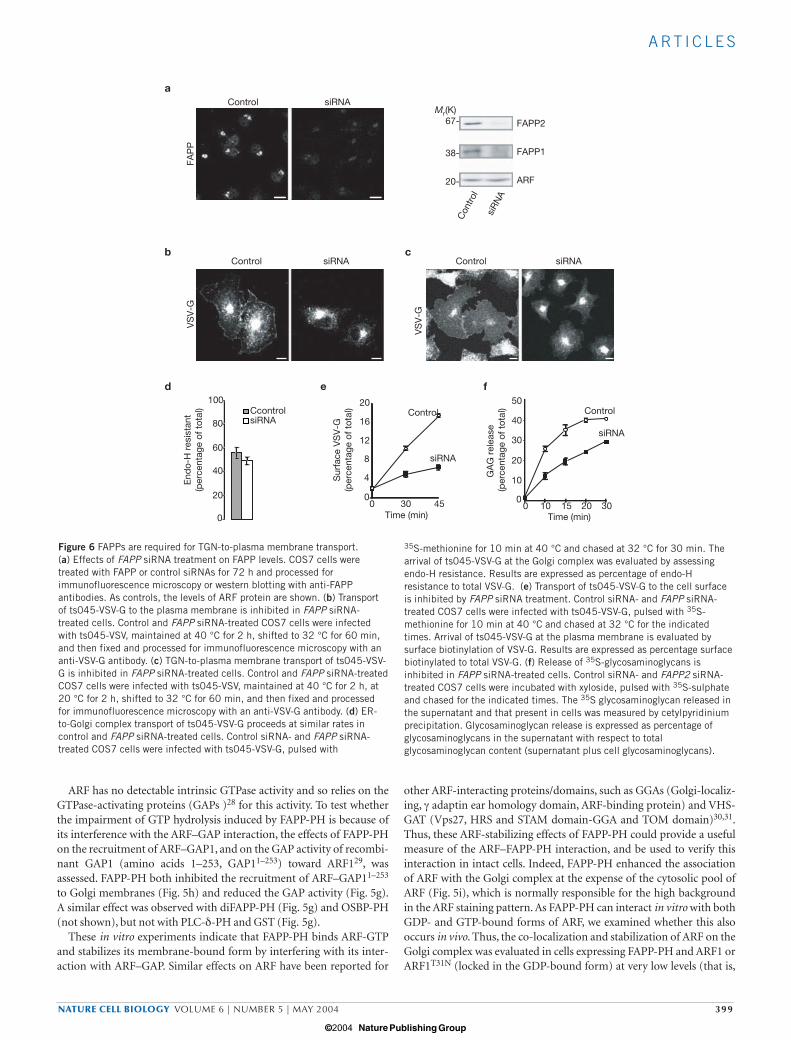
35S-methionine for 10 min at 40 °C and chased at 32 °C for 30 min. Thearrival of ts045-VSV-G at the Golgi complex was evaluated by assessingendo-H resistance. Results are expressed as percentage of endo-Hresistance to total VSV-G. (e) Transport of ts045-VSV-G to the cell surfaceis inhibited by FAPP siRNA treatment. Control siRNA- and FAPP siRNA-treated COS7 cells were infected with ts045-VSV-G, pulsed with 35S-methionine for 10 min at 40 °C and chased at 32 °C for the indicatedtimes. Arrival of ts045-VSV-G at the plasma membrane is evaluated bysurface biotinylation of VSV-G. Results are expressed as percentage surfacebiotinylated to total VSV-G. (f) Release of 35S-glycosaminoglycans isinhibited in FAPP siRNA-treated cells. Control siRNA- and FAPP2 siRNA-treated COS7 cells were incubated with xyloside, pulsed with 35S-sulphateand chased for the indicated times. The 35S glycosaminoglycan released inthe supernatant and that present in cells was measured by cetylpyridiniumprecipitation. Glycosaminoglycan release is expressed as percentage ofglycosaminoglycans in the supernatant with respect to totalglycosaminoglycan content (supernatant plus cell glycosaminoglycans).
print ncb1119 15/4/04 5:19 PM Page 399
© 2004 Nature Publishing Group
A RT I C L E S
400 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
levels not affecting either organization of the Golgi complex or associa-tion of other ARF effectors with the Golgi complex). Whereas ARF1completely co-localized with FAPP-PH on the Golgi complex,ARF1T31N remained mainly cytosolic, even in the presence of Golgi-associated FAPP-PH (Fig. 5j). These results indicate that FAPP-PHinteracts mainly with ARF-GTP in vivo.
FAPPs are required for Golgi-to-plasma membrane transportTwo approaches were used to determine if FAPPs have a functional rolein Golgi-to-plasma membrane transport: knock-down of FAPP, andthe displacement of FAPPs from the Golgi complex.
COS7 cells were treated with FAPP-specific small-interfering RNAs(siRNAs) for 48–72 h. After this time, a reduction of 70–90% in protein
levels was achieved, as evaluated by immunofluorescence microscopyand western blotting (Fig. 6a).
The impact of FAPP knock-down was analysed by examining thetransport of ts045-VSV-G, assessing its arrival at the plasma membraneby immunofluorescence microscopy or cell-surface biotinylation. Cellswere infected, and then maintained at 40 °C for 2 h or pulsed with 35S-methionine for 10 min at 40 °C before immunofluorescencemicroscopy and biochemical studies, respectively, and then moved to32 °C for different times. Although ts045-VSV-G transport to the Golgicomplex (evaluated by the ts045-VSV-G fluorescence pattern andresistance to endoglycosidase-H treatment) proceeded at the same ratein control and FAPP-siRNA-treated cells (Fig. 6b, d), the arrival ofts045-VSV-G at the plasma membrane was inhibited in the latter
b cFAPP-PH VSV-G MergeFAPP-PH
d
a
**
60 m
in
*
*
*
*
0 m
in
VSV-G VSV-GFAPP-PH FAPP-PH
Domain Ligand
PH
GLTP
Oxysterol-binding motif
STAR-T domain
Glycolipid
Oxysterol
ARF
FAPP1 FAPP2
OSBP GPBP
FAPP2OSBP GPBP
FAPP1
Ceramide
PtdIns(4)P
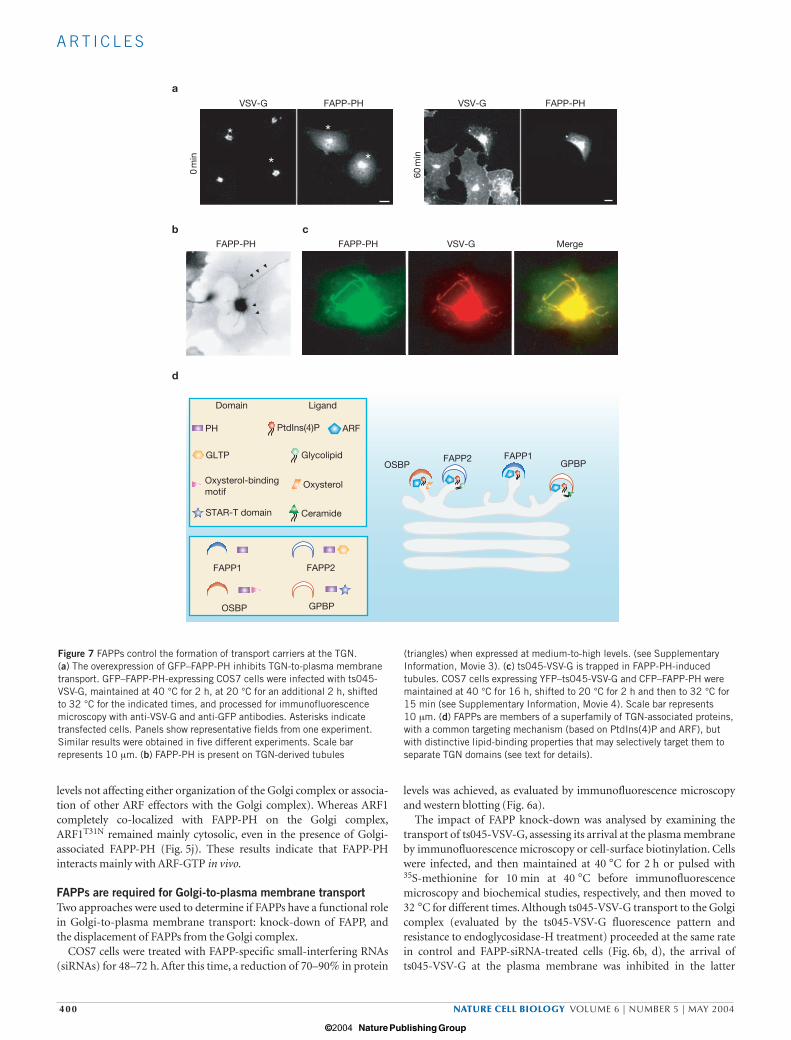
Figure 7 FAPPs control the formation of transport carriers at the TGN. (a) The overexpression of GFP–FAPP-PH inhibits TGN-to-plasma membranetransport. GFP–FAPP-PH-expressing COS7 cells were infected with ts045-VSV-G, maintained at 40 °C for 2 h, at 20 °C for an additional 2 h, shiftedto 32 °C for the indicated times, and processed for immunofluorescencemicroscopy with anti-VSV-G and anti-GFP antibodies. Asterisks indicatetransfected cells. Panels show representative fields from one experiment.Similar results were obtained in five different experiments. Scale barrepresents 10 µm. (b) FAPP-PH is present on TGN-derived tubules
(triangles) when expressed at medium-to-high levels. (see SupplementaryInformation, Movie 3). (c) ts045-VSV-G is trapped in FAPP-PH-inducedtubules. COS7 cells expressing YFP–ts045-VSV-G and CFP–FAPP-PH weremaintained at 40 °C for 16 h, shifted to 20 °C for 2 h and then to 32 °C for15 min (see Supplementary Information, Movie 4). Scale bar represents10 µm. (d) FAPPs are members of a superfamily of TGN-associated proteins,with a common targeting mechanism (based on PtdIns(4)P and ARF), butwith distinctive lipid-binding properties that may selectively target them toseparate TGN domains (see text for details).
print ncb1119 15/4/04 5:19 PM Page 400
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 401
(Fig. 6b, e). The ratio between ts045-VSV-G in the Golgi complex andat the plasma membrane was higher in FAPP-siRNA-treated cells thanin control cells at the different times analysed (Fig. 6e). The defectinduced by FAPP-siRNA was restricted to mainly the TGN-to-plasmamembrane transport step, as assessed by examining the arrival at theplasma membrane of ts045-VSV-G that had been accumulated in theTGN with a 20-°C block (Fig. 6c).
Next, we assessed whether interfering with FAPPs had an effect on thetransport of other cargo. Secretion of 35S-labelled glycosaminoglycans— markers for bulk-flow transport from the TGN to the plasma mem-brane44 — was evaluated in control and FAPP-siRNA-treated cells. Thissecretion was inhibited markedly in cells treated with FAPP siRNAs,compared with cells treated with control siRNAs (Fig 6f). This indicatesa general role for FAPPs in TGN-to-plasma membrane transport.
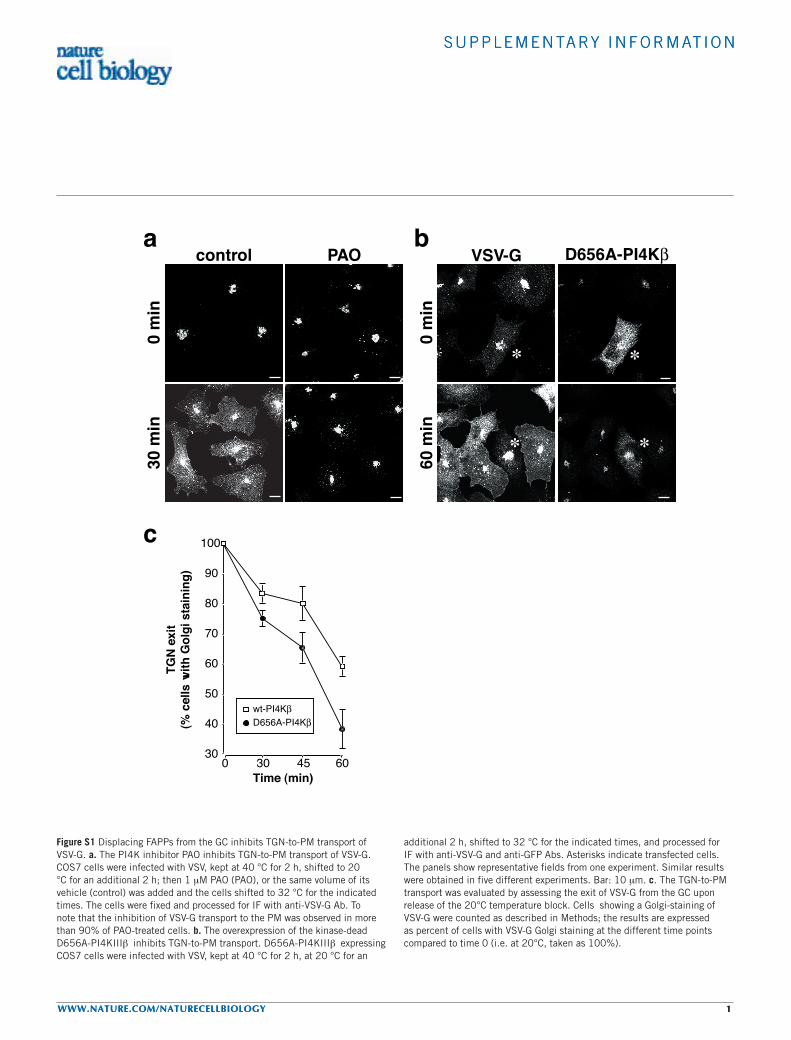
Consistently, this traffic step was also inhibited under conditionswhere FAPPs dissociate from the Golgi complex, such as treatmentwith PAO and overexpression of dead PI(4)KIIIβ (see SupplementaryInformation, Fig. S1a).
Cells expressing high levels of FAPP-PH also showed a marked delayin transport of ts045-VSV-G from the TGN to the plasma membrane,compared with control cells (Fig. 7a). The same inhibitory effect onts045-VSV-G transport to the plasma membrane was exerted bymicroinjected FAPP-PH in ts045VSV-infected cells (data not shown).
Altogether, these results indicate that FAPPs are required at the TGNfor transport of cargo to the plasma membrane.
FAPP-PH inhibits fission of transport intermediates destined forthe plasma membraneTo determine the specific processes regulated by FAPPs, the dynamicsof ts045-VSV-G transport in living cells were examined, where endoge-nous FAPPs were displaced from the Golgi complex by high levels ofFAPP-PH. Here, FAPP-PH localized not only to the central Golgi com-plex, but also on very long tubules extending from the Golgi complextowards the cell periphery (Fig. 7b). These structures eventuallyretracted or bent and returned to the Golgi complex, but neitherdetached from the central Golgi complex nor fused with the plasmamembrane (see Supplementary Information, Movie 3). These tubulescontained cargo (ts045-VSV-G). Indeed, ts045-VSV-G remained in theGolgi complex and in the tubules after release of the 20 °C block(Fig. 7c; see Supplementary Information, Movie 4). Remarkably, thenumber of discrete carriers that detached from the Golgi complex wasvery low, compared with control cells. Finally, plasma membranelabelling of ts045-VSV-G was negligible in these cells. The tubules,induced by high levels of FAPP-PH in different cell lines, originatedfrom the TGN and did not contain resident Golgi proteins or endocytictracers (data not shown); the tubules ranged from 5–40 µm, elongatedat 0.7 µm s−1 (consistent with microtubule-based motility) and weremicrotubule-dependent.
DISCUSSIONHere we show that the ARF- and PtdIns(4)P-binding FAPPs arerequired for TGN-to-plasma membrane transport, and they controlthe formation of post-Golgi constitutive carriers.
FAPPs associate with cargo-containing tubular structures as theyemerge from the TGN. This localization of FAPPs relies on their PHdomain, which binds PtdIns(4)P with high selectivity19. Thedimeric FAPP-PH (diFAPP-PHR18L) which is inactive for bindingPtdIns(4)P, still localizes to the Golgi complex, but not specificallyto the TGN. This is consistent with the ability of the FAPP-PHdomain to bind to the small GTPase ARF, which has a broad intra-Golgi distribution.
The ability of FAPP-PH to interact with ARF is consistent withreports suggesting that other PH domains also interact with proteins aswell as with phosphoinositides. The list of PH-domain-interacting pro-teins includes protein kinase C32, the βγ subunits of heterotrimeric Gproteins33 and several small GTPases. For example, the PH domain ofPLC-β interacts with Rac34, and that of the PH–DH moduli of Rac gua-nine nucleotide-exchange factors (GEFs) can establish direct interac-tions with Rac during its activation35. Interestingly, we also found thatin addition to FAPP-PH, other Golgi-targeted PH domains, such asOSBP-PH, can interact with ARF. This is not surprising considering thehigh degree of similarity between FAPP-PH and OSBP-PH, and mayprovide the molecular explanation for the reported phosphoinositide-independent and ARF-dependent binding site of OSBP-PH in theGolgi complex25.
The formation of mature transport carriers relies on a co-ordinatedsequence of elementary processes, including cargo loading, priming forsubsequent transport steps, budding and fission. The precise mecha-nisms by which FAPPs intervene in this process remain to be defined.FAPPs may be involved either in cargo selection or in recruiting themachinery needed for subsequent transport steps (such as movementalong microtubules, and docking or fusion with the plasma mem-brane) to nascent carriers. The evidence obtained so far indicates thatan important action of FAPPs is to ensure that mature carriers that arereleased from the TGN are competent for fusion with the plasma mem-brane. FAPPs may exert this effect either by actively recruiting fusionmachinery components to nascent carriers, and/or by inhibiting fissionof immature transport carriers (that is, carriers not properly equippedwith fusion machinery components). Consistent with this hypothesis,overexpression of FAPP-PH promotes the formation of cargo-contain-ing tubules that are unable to undergo not only fission from the TGN,but also fusion with the plasma membrane. The impaired fission ofthese tubules would arise from a persistent inhibition of membrane fis-sion induced by the FAPP-PH domain at TGN exit sites (as this trun-cated protein is likely to be less sensitive to regulation and reside longeron the forming carriers, compared with the full-length proteins),whereas the inability of these tubules to fuse with the plasma mem-brane would reflect a defective recruitment of fusion machinery com-ponents induced by the displacement of the full-length FAPPs.
We envisage that FAPPs may ultimately regulate the number, size andcomposition of carriers budding from the TGN by functioning asmolecular switches between a fission-restrictive and a fission-permis-sive state of the nascent carriers. By binding PtdIns(4)P, FAPPs maymodulate availability of this lipid for enzymes such as PtdIns(4)P5Kand/or PLC, which generate lipids with intrinsic membrane-destabiliz-ing properties (for example, DAG and phosphatidic acid; ref. 36) orthat can recruit and/or activate fission-inducing proteins (for example,dynamin, protein kinase D and BARS50; ref. 37). By binding ARF,FAPPs may regulate the cycle of this GTPase through modulating itsinteraction with ARF–GAP.
Finally, why should there be two FAPPs? Both FAPPs might have syn-ergistic, redundant or antagonistic activities at the same site on theTGN that controls the formation of transport carriers. Alternatively,FAPP1 and FAPP2 could be recruited to separate domains at the TGNthat are involved in the generation of carriers destined for differentplasma membrane domains (Fig. 7d). Indeed, FAPP2 possesses a gly-colipid-transfer-protein-homology domain and is thus likely to bindglycolipids and potentially be targeted specifically to apical-sortingTGN domains38. It is intriguing that the other two proteins with PHdomains that also bind PtdIns(4)P and localize to the Golgi complex inmammals and yeast25 are OSBP1 and Goodpasture antigen-bindingprotein (GPBP)39. In addition, OSBP-PH may interact with ARF-GTP
print ncb1119 15/4/04 5:19 PM Page 401
© 2004 Nature Publishing Group

A RT I C L E S
402 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
(present study). As with FAPP2, they also possess lipid-binding mod-ules: the oxysterol-binding motif and the START domain40, respec-tively. Interestingly, it has recently been shown that a splice variant ofGPBP is targeted to the Golgi complex in a PtdIns(4)P-dependentmanner, and that this mediates intracellular trafficking of ceramide41.Thus, there might be a superfamily of TGN-associated proteins with acommon regulatory mechanism based on binding PtdIns(4)P andARF, but with distinctive lipid-transfer and/or lipid-binding propertiesthat may selectively target them to separate TGN domains (Fig. 7d).
It remains a challenge for future studies to further define the mecha-nisms of action of the PtdIns(4)P- and ARF-regulated FAPPs and therole of the putative glycolipid-transfer activity of FAPP2 in the genera-tion of transport intermediates at the TGN, as well as the nature oftheir molecular partners.
METHODSAntibodies and reagents. All chemical reagents were of analytical grade orhigher and purchased from Sigma (St Louis, MO) unless otherwise specified.Cell culture media were from Invitrogen (Carlsbad, CA). Polyclonal antibodiesto human FAPPs were raised in rabbits using GST–FAPP1-PH and GST–FAPP2as immunogens. Both were affinity-purified on their corresponding immuno-gens. The anti-ts045-VSV-G clone P5D4, Cy3-conjugated and the anti-Flagbiotinylated M5 monoclonal antibodies, and the anti-rabbit and anti-mouseIgG Cy3-conjugated antibodies were from Sigma. Rabbit polyclonal antibodiesagainst human TGN46 and the monoclonal anti-giantin antibody were giftsfrom V. Ponnambalan (University of Dundee, UK) and H. P. Hauri(Switzerland), respectively. The monoclonal 1D9 anti-ARF antibody was fromAlexis (Lausen, Switzerland). The anti-GM130, anti-Rac and anti-Rho mono-clonal antibodies were from Transduction Laboratories (Lexington, KY). TheAlexa 488 goat anti-mouse and anti-rabbit IgG (H+L) antibodies were fromMolecular Probes (Eugene, OR). The FluoroLink Cy 5 labelled goat anti-rabbitand anti-mouse IgG (H+L) antibodies were from Amersham Pharmacia Biotech(Piscataway, NJ).
Cloning of FAPP1 and FAPP2. To obtain the cDNA of FAPP1, a sequence cod-ing for the new PH domain (GenBank accession number W32183; ref. 19) wasused to screen the NCBI EST database. Two ESTs (GenBank accession numberAI337400, IMAGE number 2062490 and EST NCBI accession AI221252,IMAGE number 1842552) were ordered from IMAGE consortium, and used as templates in an overlap PCR strategy with the following N-terminal and C-terminal primers: 5′-AGATCTGCCACCATGGACTACAAGGACGACGAT-GACAAGGAGGGGGTGTTGTACAAGTGG-3′ and 5′-AGATCTTCAGGAAG-AGAAGGATGGAAG-3′. The resulting PCR product coding for the full-lengthFAPP1 protein with an N-terminal Flag tag was ligated into pCR2.1-TOPO vec-tor (Invitrogen), sequenced, then subcloned as a BglII–BglII fragment into theBamHI site of pEBG-2T vector59, and into the BglII site of pEGFP-C1 andpEYFP-C1 vectors (Clontech, Palo Alto, CA). To clone the cDNA of FAPP2,based on the sequence of NCBI AF308300, the region coding for amino acids1–192 was amplified from a brain cDNA library (Clontech). This partial cDNAand an EST (GenBank accession number AA449903, IMAGE number 788667)were used in an overlap PCR strategy to create the full-length coding region ofFAPP2 cDNA with the N-terminal and C-terminal primers: 5′-GGA-TCCGCCACCATGGACTACAAGGACGACGATGACAAGATGGAGGGGGT-GCTGTACAAGTGGACCAAC-3′ and 5′-GGATCCTTATAAAGCGTCCAGTA-TGGCCATCTGC-3′. The resulting PCR product was sequenced in pCR2.1-TOPO vector, then subcloned as a BamH1–BamH1 fragment into differentexpression vectors.
DNA constructs. The PH domain of FAPP1 coding for amino acids 1–100 wasamplified from the full-length FAPP1 clone using the following primers:5′-GGATCCGCCACCATGGAGCAGAAGCTGATCTCTGAAGAGGACTTG-GAGGGGGTGTTGTACAAGTGGACCAA-3′ and 5′-GGATCCTTAAGTCCTT-GTATCAGTCAAACATGC-3′. The resulting PCR product was sequenced inpCR2.1-TOPO vector, then subcloned as a BamHI–BamHI fragment intopEBG-2T, pEGFP-C1, pEYFP-C1 and pECFP-C1 expression vectors. The R18Lmutation in FAPP1-PH (as with the other mutants) was generated using the
Quickchange mutagenesis system (Stratagene, Cedar Creek, TX) following themanufacturer’s protocol. FAPP2∆PH was prepared by PCR and subcloned intoKpnI and SmaI sites of the pEGFP-C1/pECFP-C1 vectors (Clontech). The full-length FAPP1 and FAPP1W84G coding sequences were inserted into the BglII andXhoI sites of the pEGFP-C1/pEYFP-C1 vectors (Clontech). The full-length andtruncated version of FAPP2 were inserted into the KpnI and SmaI sites of thepEGFP-C1/pECFP-C1 vectors (Clontech). The FAPP1-PH dimers, diFAPP-PHand diFAPP-PHR18L, were obtained from the respective monomers linked by thepeptide LLCSLL. The tandem PHs were subcloned into the EcoRI and BamHIsites of the pEGFP-C1 vector (Clontech). The pEYFP and pECFP-ts045-VSV-Gwere prepared as described42.
Cell culture and transfection. COS7 and A549 cells were grown in DMEM sup-plemented with 10% foetal calf serum (FCS), and NRK cells in DMEM supple-mented with 5% FCS. The cells were plated on glass coverslips for 24 h andtransfected with the Trans-Fast transfection reagent (Promega, Madison, WI)according to the manufacturer’s instructions. Alternatively, resuspended cellswere transfected using the Gene Pulser Electroprotocol (Biorad, Hercules, CA),according to the manufacturer’s instructions. The cells were treated 16 h aftertransfection.
Recombinant protein purification. All the GST fusion constructs (cloned in p-GEX vectors) were produced and purified using the Bulk GST purificationmodule (Amersham Pharmacia Biotech), according to the manufacturer’sinstructions. OSBP-PH and PLC-δ-PH were prepared as described43.Myristoylated ARF1 was produced and purified as described44. ARF–GAP11–253
was produced as described29.
Binding to Golgi membranes and protein-lipid overlay assay. Binding assayswere performed as described16. Golgi membranes, cytosol and ARF-depletedcytosol were prepared as described43. Binding of FAPP-PH to PtdIns(4)P wasassessed evaluating the protection activity of FAPP-PH towards 32P-PtdIns(4)Pdegradation in Golgi membranes, as described16. The protein-lipid overlay assaywas performed as described19.
ARF pull-down and ARF-GAP assay. Golgi membranes were incubated withcytosol and GST–FAPP-PHs for 10 min at 37 °C. Membranes were collected bycentrifugation, resuspended in 20 mM Hepes-KOH at pH 7.2, 2.5 mMMg(OAc)2, 90 mM KOAc and 70 mM dithiobis(succinimidyl propionate)(Pierce, Rockford, IL), incubated for 30 min on ice, and quenched with 3 mMTris-HCl at pH 7.2. After incubation, Golgi membranes were centrifuged andthe pellet extracted with 20 mM Hepes-KOH at pH 7.4, 500 mM KCl, 1% TritonX-100, 2 mM EDTA, 2 mM dithiothreitol and protease inhibitors (30 min onice), diluted 1:1 with water and clarified for 10 min at 15,000g. Extracts wereincubated with 20 µl glutathione–Sepharose beads for 1 h at 4 °C. Beads werewashed extensively, resuspended in sample buffer and analysed by SDS–PAGEand western blotting. Recombinant myristoylated ARF1 was loaded with gua-nine nucleotides by incubation for 1 h at 32 °C in loading buffer (25 mM Hepesat pH 7.4, 100 mM NaCl, 1 mM dithiothreitol, 1 mM EDTA, 1 mM MgCl2, 0.5 gl−1 BSA and 0.1% Triton X-100) containing 100 µM GTP or GDP. Nucleotide-preloaded ARF mixture (50 µl; 15 µg per sample) was incubated with GST-PHdomains immobilized on glutathione–Sepharose beads for 1 h at room temper-ature. Beads were collected by centrifugation, washed and resuspended in sam-ple buffer. ARF associated with the beads was detected by SDS–PAGE andwestern blotting with anti-ARF1 antibodies. The ARF–GAP1 assay was per-formed as described29.
VSV-G transport assays. Analysis of transport of ts045-VSV-G was performedin cells infected with the ts045-VSV-G strain or transfected with GFP–VSV-G, asdescribed42. Immunofluorescence microscopy analysis was also performed asdescribed42. Quantitative evaluation of ts045-VSV-G transport was performedby analysing the immunofluorescence staining patterns of at least 200 cells inthree separate experiments.
Endoplasmic reticulum (ER)-to-Golgi and ER-to-plasma membrane trans-port of ts045-VSV-G was also assessed by evaluating acquisition of resistance toendoglycosidase-H treatment, and cell-surface biotinylation of 35S-metionine-labelled ts045-VSV-G, according to the procedures described in refs 43 and 45,respectively.
print ncb1119 15/4/04 5:19 PM Page 402
© 2004 Nature Publishing Group

A RT I C L E S
NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004 403
Analysis of glycosaminoglycan release. COS 7 cells in 12-well plates werewashed, incubated with xyloside, labelled with 35S-sulphate and incubated inrelease medium containing 0.1% BSA for the indicated times. Release mediawere collected and centrifuged (12,000g; 10 min), and the supernatant wasrecovered. Cell monolayers were extracted with 0.5 ml 100 mM NaOH for atleast 30 min at 37 °C. 35S-glycosaminoglycans were quantified by a cetylpyri-dinium chloride precipitation assay, as described45.
PH domain binding assay in permeabilized cells. COS7 cells were incubatedwith 1–2 U ml−1 SLO in SLO buffer (20 mM Hepes at pH 7.2, 110 mMMg(OAc)2 and 1 mM dithiothreitol) for 10 min at 4 °C. Cells were then washedtwice and incubated in permeabilization buffer (25 mM Hepes at pH 6.95, 125mM KOAc, 2.5 mM Mg(OAc)2, 10 mM glucose, 1 mM dithiothreitol, 1 mMATP, 5 mM creatine phosphate, 7.3 U ml−1 creatine phosphokinase and 1 mgml−1 rat brain cytosol) for 5 min at 37 °C, and then in permeabilization buffercontaining 1–3 µM GST–FAPP-PH for an additional 5 min. In selected experi-ments, permeabilized cells were incubated in permeabilization buffer in theabsence of ATP or in the presence of 200 µM Ins(1,4)P2. Cells were then fixedwith 4% paraformaldehyde and processed for immunofluorescence microscopy,as described above.
Time-lapse laser-scanning confocal microscopy and epifluorescence videomicroscopy. Time-lapse laser-scanning confocal microscopy was performed asdescribed42. Time-lapse epifluorescence video microscopy was performed usingan inverted microscope (Olympus IX70; Olympus, Hamburg, Germany)equipped with a polychrome II monochromator (TILL Photonics, Eugene,Oregon), a custom filter-block for visualization of GFP, YFP and CFP, a 63×1.4oil-immersion lens, and a 12-bit CCD IMAGO digital camera (TILL Photonics),controlled by TILLvisION v3.3 software (TILL Photonics). Switching of excita-tion wavelengths (430 nm and 500 nm for CFP and YFP, respectively) tookapproximately 2 ms, and data acquisition times typically ranged from 20 to100 ms per frame.
Immuno-electron microscopy. HEK 293 cells were grown to sub-confluence on10-cm plastic dishes and transfected with FAPP1 constructs using Fugene 6(Roche, Basel, Switzerland), according to the manufacturer’s instructions. After24 h, the medium was removed and replaced with 10 ml 0.5% glutaraldehyde inPIPES buffer at pH 7.2 and incubated for 30 min. Cells were washed in PBS,scraped using a rubber policeman and sedimented at 16,000g. The pellet wascryoprotected in 2.1 M sucrose in PBS and frozen on cryostubbs for cryomicro-tomy in a Leica EM ultracut UCT microtome (Leica; Microsystems Inc.,Bannockburn, IL). Ultrathin sections of nominal 80-nm thickness wereretrieved using a 1:1 mixture of 2% (w/v) methylcellulose/2.1 M sucrose in PBSand transferred to pioloform/carbon-coated 100-mesh copper electron micro-scope grids. The sections were labelled using anti-GFP antibodies (M. Lowe,Manchester, U.K.) followed by rabbit antisheep IgG (Southern Biotech,Birmingham, Alabama) and 8-nm Protein-A gold46. After washes in PBS andfixation in 0.5% glutaraldehyde, sections were further labelled using anti-GM130 antibodies and 12-nm protein-A gold. Fish-skin gelatin (0.5%) wasused as a blocking agent and 2% methyl cellulose/3% uranyl acetate (9/1.5) forcontrasting. Quantification was performed as described46. Cisternae weredefined as any profiles in the Golgi region with an axial ratio of ≥3. All cisternaethat were negative for GM130 were excluded from the study. Cis cisternae wereidentified as single cisternae that contained GM130 labelling or GM130-nega-tive cisternae that were in the same register in the stack as positive ones. Thetrans-Golgi cisterna was the most extreme cisterna on the opposite face of theGolgi stack to GM130-positive structures. Medial cisternae were all those cister-nal structures found between cis and trans cisternae. Cis vesiculotubular struc-tures and the TGN were identified as non-cisternal round or elongatedmembrane profiles closely associated with cis or trans cisternae, as definedabove.
RNA interference. siRNAs of human FAPP1 (AF286162) and FAPP2(AF380162) consisted of a mixture of four siRNA duplexes selected using theDharmacon SMARTselection process and SMARTpool algorithm, and theywere obtained from Dharmacon (Lafayette, CO). COS7 cells were plated at 30%confluence in 12-well plates and transfected with 120–150 pmol of FAPP1and/or FAPP2 siRNAs using Oligofectamine (Invitrogen) in accordance with
the manufacturer’s protocol. For the control, COS7 cells were treated with iden-tical concentrations of luciferase siRNA (Dharmacon). At 48/72 h after the ini-tial siRNA treatment, the cells were directly processed for immunofluorescencemicroscopy, infected with ts045-VSV, or labelled with 35S-sulphate, and assayedfor ts045-VSV-G and 35S-glycosaminoglycan transport. Cells treated with FAPPsiRNAs had a slower rate of growth and were less adherent to the substrate, com-pared with control cells (that is, cells treated with equivalent concentrations ofcontrol siRNAs). Of note, this FAPP knock-down did not affect the steady-statedistribution of mannose 6-phoshosphate receptor or the transferrin receptor.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGMENTSWe thank B. Antonny for providing reagents. We are also grateful to A. Luini, V.Malhotra, and to the members of the Golgi group of the Consorzio Mario NegriSud for discussions. We thank C. Berrie for reading the manuscript and E. Fontanafor artwork preparation. This work was supported in part by the ItalianAssociation for Cancer Research, Telethon Italia, the European Community andthe Italian Ministry of Education. A.D.C. and T.D. are supported by fellowshipsfrom the Italian Foundation of Cancer Research.
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Received 12 February 2004; accepted 8 March 2004Published online at http://www.nature.com/naturecellbiology
1. Hirschberg, K. et al. Kinetic analysis of secretory protein traffic and characterization ofGolgi to plasma membrane transport intermediates in living cells. J. Cell Biol. 143,1485–1503 (1998).
2. Kreitzer, G. et al. Three-dimensional analysis of post-Golgi carrier exocytosis in epithe-lial cells. Nature Cell Biol. 5, 126–136 (2003).
3. Polishchuk, R. S. et al. Correlative light-electron microscopy reveals the tubular-sac-cular ultrastructure of carriers operating between Golgi apparatus and plasma mem-brane. J. Cell Biol. 148, 45–58 (2000).
4. Bankaitis, V. A., Malehorn, D. E., Emr, S. D. & Greene, R. The Saccharomyces cere-visiae SEC14 gene encodes a cytosolic factor that is required for transport of secretoryproteins from the yeast Golgi complex. J. Cell Biol. 108, 1271–1281 (1989).
5. Jones, S. M., Alb, J. G., Phillips, S. E. Jr, Bankaitis, V. A. & Howell, K. E. A phos-phatidylinositol 3-kinase and phosphatidylinositol transfer protein act synergisticallyin formation of constitutive transport vesicles from the trans-Golgi network. J. Biol.Chem. 273, 10349–10354 (1998).
6. Simon, J. P. et al. An essential role for the phosphatidylinositol transfer protein in thescission of coatomer-coated vesicles from the trans-Golgi network. Proc. Natl Acad.Sci. USA 95, 11181–11186 (1998).
7. Baron, C. L. & Malhotra, V. Role of diacylglycerol in PKD recruitment to the TGN andprotein transport to the plasma membrane. Science 295, 325–328 (2002).
8. De Matteis, M., Godi, A. & Corda, D. Phosphoinositides and the Golgi complex. Curr.Opin. Cell Biol. 14, 434–447 (2002).
9. Audhya, A., Foti, M. & Emr, S. D. Distinct roles for the yeast phosphatidylinositol 4-kinases, Stt4p and Pik1p, in secretion, cell growth, and organelle membrane dynam-ics. Mol. Biol. Cell 11, 2673–2689 (2000).
10. Hama, H., Schnieders, E. A., Thorner, J., Takemoto, J. Y. & DeWald, D. B. Directinvolvement of phosphatidylinositol 4-phosphate in secretion in the yeastSaccharomyces cerevisiae. J. Biol. Chem. 274, 34294–34300 (1999).
11. Walch-Solimena, C. & Novick, P. The yeast phosphatidylinositol-4-OH kinase pik1 reg-ulates secretion at the Golgi. Nature Cell Biol. 1, 523–525 (1999).
12. Li, X. et al. Analysis of oxysterol binding protein homologue Kes1p function in regula-tion of Sec14p-dependent protein transport from the yeast Golgi complex. J. Cell Biol.157, 63–77 (2002).
13. Wei, Y. J. et al. Type II phosphatidylinositol 4-kinase beta is a cytosolic and peripheralmembrane protein that is recruited to the plasma membrane and activated by Rac-GTP. J. Biol. Chem. 277, 46586–46593 (2002).
14. Nakagawa, T., Goto, K. & Kondo, H. Cloning, expression, and localization of 230-kDaphosphatidylinositol 4-kinase. J. Biol. Chem. 271, 12088–12094 (1996).
15. Wong, K., Meyers, R. & Cantley, L. C. Subcellular locations of phosphatidylinositol 4-kinase isoforms. J. Biol. Chem. 272, 13236–13241 (1997).
16. Godi, A. et al. ARF mediates recruitment of PtdIns-4-OH kinase-beta and stimulatessynthesis of PtdIns(4,5)P2 on the Golgi complex. Nature Cell Biol. 1, 280–287(1999).
17. Bruns, J. R., Ellis, M. A., Jeromin, A. & Weisz, O. A. Multiple roles for phosphatidyli-nositol 4-kinase in biosynthetic transport in polarized Madin-Darby canine kidneycells. J. Biol. Chem. 277, 2012–2018 (2002).
18. Wang, Y. L. et al. Phosphotidylinositol 4 phosphate regulates tageting of clathrin adap-tor AP1 complexes to the Golgi. Cell 114, 299–310 (2003).
19. Dowler, S. et al. Identification of pleckstrin-homology-domain-containing proteins withnovel phosphoinositide binding specificities. Biochem. J. 351, 19–31 (2000).
20. Lin, X., Mattjus, P., Pike, H. M., Windebank, A. J. & Brown, R. E. Cloning and expres-sion of glycolipid transfer protein from bovine and porcine brain. J. Biol. Chem. 275,5104–5110 (2000).
21. Prescott, E. D. & Julius, D. A modular PIP2 binding site as a determinant of capsaicin
print ncb1119 15/4/04 5:19 PM Page 403
© 2004 Nature Publishing Group

A RT I C L E S
404 NATURE CELL BIOLOGY VOLUME 6 | NUMBER 5 | MAY 2004
receptor sensitivity. Science 300, 1284–1288 (2003).22. Wiedemann, C., Schafer, T. & Burger, M. M. Chromaffin granule-associated phos-
phatidylinositol 4-kinase activity is required for stimulated secretion. EMBO J. 15,2094–2101 (1996).
23. Donaldson, J. G., Finazzi, D. & Klausner, R. D. Brefeldin A inhibits Golgi membrane-catalysed exchange of guanine nucleotide onto ARF protein. Nature 360, 350–352(1992).
24. Helms, J. B. & Rothman, J. E. Inhibition by brefeldin A of a Golgi membrane enzymethat catalyses exchange of guanine nucleotide bound to ARF. Nature 360, 352–354(1992).
25. Levine, T. P. & Munro, S. Targeting of Golgi-specific pleckstrin homology domainsinvolves both PtdIns 4-kinase-dependent and -independent components. Curr. Biol.12, 695–704 (2002).
26. Stefan, C. J., Audhya, A. & Emr, S. D. The yeast synaptojanin-like proteins control thecellular distribution of phosphatidylinositol (4,5)-bisphosphate. Mol. Biol. Cell 13,542–557 (2002).
27. Cohen, G. B., Ren, R. & Baltimore, D. Modular binding domains in signal transductionproteins. Cell 80, 237–248 (1995).
28. Randazzo, P. A., Nie, Z., Miura, K. & Hsu, V. W. Molecular aspects of the cellular activ-ities of ADP-ribosylation factors. Sci. STKE 59, DOI: 10.1126/stke.2000.59.re1(2000).
29. Antonny, B., Huber, I., Paris, S., Chabre, M. & Cassel, D. Activation of ADP-ribosyla-tion factor 1 GTPase-activating protein by phosphatidylcholine-derived diacylglyc-erols. J. Biol. Chem. 272, 30848–30851 (1997).
30. Jacques, K. M. et al. Arf1 dissociates from the clathrin adaptor GGA prior to beinginactivated by Arf GTPase-activating proteins. J. Biol. Chem. 277, 47235–47241(2002).
31. Puertollano, R., Randazzo, P. A., Presley, J. F., Hartnell, L. M. & Bonifacino, J. S. TheGGAs promote ARF-dependent recruitment of clathrin to the TGN. Cell 105, 93–102(2001).
32. Yao, L., Kawakami, Y. & Kawakami, T. The pleckstrin homology domain of Bruton tyro-sine kinase interacts with protein kinase C. Proc. Natl Acad. Sci. USA 91,9175–9179 (1994).
33. Touhara, K., Inglese, J., Pitcher, J. A., Shaw, G. & Lefkowitz, R. J. Binding of G proteinbeta gamma-subunits to pleckstrin homology domains. J. Biol. Chem. 269,10217–10220 (1994).
34. Snyder, J. T., Singer, A. U., Wing, M. R., Harden, T. K. & Sondek, J. The pleckstrinhomology domain of phospholipase C-beta2 as an effector site for Rac. J. Biol. Chem.278, 21099–21104 (2003).
35. Rossman, K. L. et al. A crystallographic view of interactions between Dbs and Cdc42:PH domain-assisted guanine nucleotide exchange. EMBO J. 21, 1315–1326 (2002).
36. Burger, K. N. Greasing membrane fusion and fission machineries. Traffic 1, 605–613(2000).
37. Corda, D., Hidalgo-Carcedo, C., Bonazzi, M., Luini, A. & Spano, S. Molecular aspects ofmembrane fission in the secretory pathway. Cell. Mol. Life Sci. 59, 1819–1832(2002).
38. Ikonen, E. & Simons, K. Protein and lipid sorting from the trans-Golgi network to theplasma membrane in polarized cells. Semin. Cell Dev. Biol. 9, 503–509 (1998).
39. Raya, A. et al. Goodpasture antigen-binding protein, the kinase that aphosphorylatesthe goodpasture antigen, is an alternatively spliced variant implicated in autoimmunepathogenesis. J. Biol. Chem. 275, 40392–40399 (2000).
40. Soccio, R. E. & Breslow, J. L. StAR-related lipid transfer (START) proteins: mediators ofintracellular lipid metabolism. J. Biol. Chem. 278, 22183–22186 (2003).
41. Hanada, K. et al. Molecular machinery for non-vesicular trafficking of ceramide. Nature426, 803–809 (2003).
42. Marra, P. et al. The GM130 and GRASP65 Golgi proteins cycle through and define asubdomain of the intermediate compartment. Nature Cell Biol. 3, 1101–1113 (2001).
43. Godi, A. et al. ADP ribosylation factor regulates spectrin binding to the Golgi complex.Proc. Natl Acad. Sci. USA 95, 8607–8612 (1998).
44. Randazzo, P. A. & Kahn, R. A. Myristoylation and ADP-ribosylation factor function.Methods Enzymol. 250, 394–405 (1995).
45. Buccione, R. et al. Regulation of constitutive exocytic transport by membrane recep-tors. A biochemical and morphometric study. J. Biol. Chem. 271, 3523–3533 (1996).
46. Lucocq, J. M. in Fine Structure Immunocytochemistry (ed. Griffiths, G.) 279–302(Springer, Berlin, 1993).
print ncb1119 15/4/04 5:19 PM Page 404
© 2004 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
acontrol
0 m
in
PAO
30 m
in
b
*
*
*
VSV-G
0 m
in
*
60 m
in* *
D656A-PI4K�
c
wt-PI4K�D656A-PI4K�
�0 30 45 60Time (min)
TGN
exi
t(%
cel
ls w
ith G
olgi
sta
inin
g)
100
80
60
40
70
50
30
90
Figure S1 Displacing FAPPs from the GC inhibits TGN-to-PM transport of VSV-G. a. The PI4K inhibitor PAO inhibits TGN-to-PM transport of VSV-G. COS7 cells were infected with VSV, kept at 40 °C for 2 h, shifted to 20 °C for an additional 2 h; then 1 µM PAO (PAO), or the same volume of its vehicle (control) was added and the cells shifted to 32 °C for the indicated times. The cells were fixed and processed for IF with anti-VSV-G Ab. To note that the inhibition of VSV-G transport to the PM was observed in more than 90% of PAO-treated cells. b. The overexpression of the kinase-dead D656A-PI4KIIIβ inhibits TGN-to-PM transport. D656A-PI4KIIIβ expressing COS7 cells were infected with VSV, kept at 40 °C for 2 h, at 20 °C for an
additional 2 h, shifted to 32 °C for the indicated times, and processed for IF with anti-VSV-G and anti-GFP Abs. Asterisks indicate transfected cells. The panels show representative fields from one experiment. Similar results were obtained in five different experiments. Bar: 10 µm. c. The TGN-to-PM transport was evaluated by assessing the exit of VSV-G from the GC upon release of the 20°C temperature block. Cells showing a Golgi-staining of VSV-G were counted as described in Methods; the results are expressed as percent of cells with VSV-G Golgi staining at the different time points compared to time 0 (i.e. at 20°C, taken as 100%).

S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
Movie 1 Time laps images of GFP-FAPP1. Frame numbers and time elapsed in h.mm.ss are indicated in the bottom.Movie 2 Time laps images of YFP-FAPP1 (red) and CFP-VSV-G (green) upon release from 20 °C temperature block. Frame numbers and time elapsed in h.mm.ss are indicated in the bottom.
Movie 3 Time laps images of GFP-FAPP-PH. Frame numbers and time elapsed in h.mm.ss are indicated in the top.Movie 4 Time laps images of CFP-FAPP-PH (green) and YFP-VSV-G (red) upon release from 20 °C temperature block. Frame numbers and time elapsed in h.mm.ss are indicated in the top.





























