Embed Size (px)
Citation preview

FPE
DKJAa
Pb
Pc
Ld
e
Abisgeitrtutocppmdiisaowipaewnu
*EAbdimacNp
Neuroscience 166 (2010) 671–679
0d
UNCTIONAL AND IN SITU HYBRIDIZATION EVIDENCE THATREGANGLIONIC SYMPATHETIC VASOCONSTRICTOR NEURONS
XPRESS GHRELIN RECEPTORSrbbvr
Kg
GkgDtilam
ec(pbbCws(
icncprawcob(oipt
f
. M. FERENS,a L. YIN,a R. BRON,a B. HUNNE,a
. OHASHI-DOI,a P. D. KITCHENER,a G. J. SANGER,b,c
. WITHERINGTON,b Y. SHIMIZUa,d
ND J. B. FURNESSa*
Department of Anatomy and Cell Biology, University of Melbourne,arkville, Victoria 3010, Australia
Neurology and GI CEDD, GlaxoSmithKline, New Frontiers Scienceark, Harlow CM19 5AW, UK
Institute of Cell and Molecular Science, Queen Mary, University ofondon, London E1 2AJ, UK
Department of Basic Veterinary Science, School of Veterinary Sci-nces, Gifu University, Gifu, Japan
bstract—Agonists of ghrelin receptors can lower or elevatelood pressure, and it has been suggested that the increases
n blood pressure are caused by actions at receptors in thepinal cord. However, this has not been adequately investi-ated, and the locations of neurons in the spinal cord thatxpress ghrelin receptors, through which blood pressure
ncreases may be exerted, are not known. We investigatedhe effects within the spinal cord of two non-peptide ghrelineceptor agonists, GSK894490 and CP464709, and two pep-ide receptor agonists, ghrelin and des-acyl ghrelin, and wesed polymerase chain reaction (PCR) and in situ hybridiza-ion to examine ghrelin receptor expression. I.v. applicationf the non-peptide ghrelin receptor agonists caused biphasichanges in blood pressure, a brief drop followed by a bloodressure increase that lasted several minutes. The bloodressure rise, but not the fall, was antagonized by i.v. hexa-ethonium. Application of these agonists or ghrelin peptideirectly to the spinal cord caused only a blood pressure
ncrease. Des-acyl ghrelin had no significant action. The max-mum pressor effects of agonists occurred with application atpinal cord levels T9 to T12. Neither i.v. nor spinal cordpplication of the agonists had significant effect on heart rater the electrocardiogram. Ghrelin receptor gene expressionas detected by PCR and in situ hybridization. In situ hybrid-
zation localized expression to neurons, including autonomicreganglionic neurons of the intermediolateral cell columnst all levels from T3 to S2. The numbers of ghrelin receptorxpressing neurons in the intermediolateral cell columnsere similar to the numbers of nitric oxide synthase positiveeurons, but there was little overlap between these two pop-lations. We conclude that activation of excitatory ghrelin
Corresponding author. Tel: �61-3-83448859; fax: �61-3-93479619.-mail address: [email protected] (J. B. Furness).bbreviations: AP, area postrema; APhos, alkaline phosphatase; bpm,eats/min; CCD, cooled charge device; CSF, cerebrospinal fluid; DIG,igoxigenin; dms, dorsomedial sulcus; ECG, electrocardiogram; IML,
ntermediolateral cell column; i.t., intrathecal; L-NAME, L-nitro arginineethyl ester; MABT, maleic acid buffer with Tween20; NA, nucleusmbiguus; NBT/BCIP, nitro blue tetrazolium chloride/5-bromo-4-hloro-3-indolyl phosphate; nNOS, neuronal nitric oxide synthase;
iTS, nucleus tractus solitarius; PBS, phosphate-buffered saline; PCR,olymerase chain reaction; PFA, paraformaldehyde.
306-4522/10 $ - see front matter © 2010 IBRO. Published by Elsevier Ltd. All rightoi:10.1016/j.neuroscience.2010.01.001
671
eceptors on sympathetic preganglionic neurons increaseslood pressure, and that decreases in blood pressure causedy ghrelin agonists are mediated through receptors on bloodessels. © 2010 IBRO. Published by Elsevier Ltd. All rightseserved.
ey words: ghrelin receptors, spinal cord, autonomic pregan-lionic neurons, blood pressure.
hrelin, a 28-amino acid octanoylated peptide, is best-nown for its ability to increase food intake and stimulaterowth hormone release (Kojima et al., 1999; Inui, 2001;epoortere, 2009). Nevertheless, it has become evident
hat this endogenous peptide can mediate growth hormonendependent effects, including effects on the cardiovascu-ar system in animals (Matsumura et al., 2002; Shimizu etl., 2003, 2006) and humans (Nagaya et al., 2001; Oku-ura et al., 2002; Kleinz et al., 2006).
Ghrelin decreases blood pressure when given periph-rally (Nagaya et al., 2001; Okumura et al., 2002). Be-ause ghrelin causes dilatation of isolated blood vesselsWiley and Davenport, 2002; Kleinz et al., 2006), its bloodressure lowering effect has been concluded to be causedy a direct effect on arteries, even though it crosses thelood–brain barrier (Pan et al., 2006; Banks et al., 2008).onsistent with a peripheral site of action, when ghrelinas administered i.v. in rabbits, it decreased arterial pres-ure, but did not change renal sympathetic nerve activityMatsumura et al., 2002).
Ghrelin or ghrelin receptor agonists that are deliverednto the CNS also have cardiovascular effects. When mi-roinjected into the nucleus tractus solitarius (NTS), butot into the other brain regions of the rat, ghrelin de-reased mean arterial pressure and heart rate and sup-ressed the activity of sympathetic vasoconstrictor neu-ons, as shown by a reduction in renal sympathetic nervectivity (Lin et al., 2004). Renal sympathetic nerve activityas also inhibited when ghrelin was injected via an i.c.v.annula (Matsumura et al., 2002). Cardiovascular effectsf ghrelin receptor agonists applied to the spinal cord wereriefly reported in a study of effects on colonic motilityShimizu et al., 2006). In that study, intrathecal applicationf ghrelin receptor agonists to the spinal cord transiently
ncreased blood pressure, and a non-peptide agonist ap-lied i.v. also increased blood pressure, although quanti-ative data were not given (Shimizu et al., 2006).
Detectable levels of ghrelin receptor mRNA have beenound in various tissues relevant to cardiovascular control,
ncluding aorta, left ventricle and left atrium (Nagaya et al.,s reserved.
2eblas(thtcIgslc
P
A3wpEokitafw(mk(pwTolu
acTasbtsiiccr
CtEcwe
A
IdaensIppipevfT
P
Tur(wTabs(cbt
I
AgrpCdgpqlwAp(
ap7Pcs(p
sw1m72ws
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679672
001), hypothalamus and brainstem (Guan et al., 1997; Lint al., 2004; Zigman et al., 2006). The ghrelin receptor haseen localized to the media and intimal smooth muscle
ayers of several human and rat blood vessels, includingorta, saphenous vein, pulmonary artery, renal blood ves-els, internal mammary artery and small coronary arteriesKatugampola et al., 2001; Kleinz et al., 2006). However,he localization of receptor expression in the spinal cordas not been described. Thus, the possible neuronal siteshrough which activation of ghrelin receptors in the spinalord enhances blood pressure have not been determined.n the current work, we have investigated the effects ofhrelin receptor agonists applied at different sites in thepinal cord on the cardiovascular system and we haveocalized the expression of ghrelin receptor RNA to spinalord neurons.
EXPERIMENTAL PROCEDURES
hysiological studies
total of 98 male Sprague–Dawley rats with an average weight of50 g and average age of 60 days were supplied with food andater ad libitum prior to experiments. The procedures were ap-roved by the University of Melbourne Animal Experimentationthics Committee and the Animal Care Committee of the Schoolf Veterinary Sciences, Gifu University. Rats were sedated withetamine hydrochloride (50–60 mg/kg i.m.) and anesthesia wasnduced with �-chloralose (60 mg/kg i.v.). Following �-chloralose,he femoral artery was cannulated for the infusion of anestheticnd blood pressure recording. The femoral vein was cannulatedor delivery of drugs. Blood pressure and heart rate were recordedith a Power Laboratory recording system using Chart 5 software
both from ADInstruments, Sydney Australia). Anesthesia wasaintained by intra-arterial infusion of �-chloralose (12–20 mg/
g/h) plus ketamine (3–7 mg/kg/h) in phosphate-buffered salinePBS; 0.15 M NaCl containing 0.01 M sodium phosphate buffer,H 7.2). All rats were kept under a constant infusion of anesthesia,ith resting mean arterial pressure maintained at 70–100 mm Hg.he urinary bladder was cannulated to ensure continuous voidingf fluid. At the end of each experiment, the rat was killed with a
ethal dose of sodium pentobarbitone (300 mg/kg i.v.), while stillnder anesthesia.
In order to provide a positive control for experiments in whichgonists were applied to the spinal cord, rats were set up to recordolonic motility, as previously described (Shimizu et al., 2006).he distal colon was cannulated at the colonic flexure and at thenus. The colon was left in situ, and the muscle and skin weretitched closed. The oral cannula was connected to a Marriotteottle filled with warm PBS, and the distal cannula to a pressureransducer via a one-way valve. The baseline intralumenal pres-ure was maintained at 3–5 mm Hg. Expelled fluid was collectedn a cylinder distal to the one way valve, and measured by weigh-ng with a force transducer. Activity in the colon was used toalculate a motility index (integrated areas under pressurehanges, measured in mm Hg. min and scaled to 100�maximumesponse).
In five rats, the electrocardiogram (ECG) was recorded usinghart software (ADInstruments) with external electrodes on the
horax and the effects of i.v. ghrelin agonist were investigated.CG data were extracted using Copycat.pas (www.copycatsoftware.om) and the wave forms and intervals between p, r and t wavesere assessed by averaging over 1000 or more waves for differ-
nt conditions. 1pplication of drugs
.v. application of drugs was via the femoral vein. For intrathecalrug injection, spaces between vertebrae were cleared. The tip of27 gauge needle connected to a polyethylene tube, 0.5 mm
xternal diameter with 10 �l dead-space, was inserted so that theeedle tip was in the subarachnoid space. The cannula wasecured in place with silicone elastomer (Kwik-Sil; World Precisionnstruments, Hertfordshire, U.K.) which created a tight seal at theoint of cannulation. No cerebrospinal fluid (CSF) leak was ap-arent after the seal was in place. Compounds were injected
ntrathecally in volumes of 10–20 �L. In the text, the intrathecalosition is indicated relative to the more rostral vertebrum, forxample, drug application into the space between L1 and L2ertebral levels is referred to as L1. Intrathecal application wasrom vertebral levels T3 to L4, corresponding to spinal cord levels3–S3 (Padmanabhan and Singh, 1979).
olymerase chain reaction
otal RNA was extracted from freshly dissected rat spinal cordsing the RNeasy Mini Kit (Qiagen, Melbourne, Australia) and waseverse transcribed using Superscript III reverse transcriptaseInvitrogen, Melbourne, Australia). Rat ghrelin receptor mRNAas amplified by polymerase chain reaction (PCR) using Platinumaq (Invitrogen) with forward primer GTCCAGCATGGCCTTCTCnd reverse primer GAATGGGGTTGATGGCAG to produce a 716p product (Fig. 4). The identity of the product was confirmed byequencing using ABI PRISM Big Dye Terminator V3.1 reagentApplied Biosystems, Melbourne, Australia). Gel separation wasonducted by the Australian Genome Research Facility, Mel-ourne, Australia. Reactions were also conducted without reverseranscriptase to confirm the absence of genomic DNA.
n situ hybridization
plasmid to prepare antisense cRNA complimentary to the rathrelin receptor mRNA was constructed using a partial rat ghrelineceptor cDNA of 910 bp that was amplified by PCR using forwardrimer TGTGGTGGTGTTTGCTTTCATCC and reverse primer GGAC-TACTTTTCCATGCTCAAATT. The amplified product spans the pre-icted exon 2 and 352 bp of the 3= untranslated region of the rathrelin receptor gene. The PCR product was cloned into theCRII-TOPO vector (Invitrogen, Melbourne, Australia). The se-uence of the insert was confirmed and its orientation was estab-
ished using M13 forward and reverse primers. Linear templatesere prepared by PCR using M13 forward and reverse primers.ntisense and control sense digoxigenin (DIG)-labeled cRNA wasrepared by in vitro transcription with T7 and SP6 RNA polymerasesRoche Products, Dee Why, NSW, Australia), respectively.
Adult male Sprague–Dawley rats (240–380 g) were deeplynesthetized and subsequently transcardially perfused with 4%araformaldehyde in phosphate-buffered saline (PFA/PBS), pH.4. Spinal cords were removed and post-fixed overnight in 4%FA/PBS at 4 °C, then embedded in optimal cutting temperatureompound (Tissue Tek, Elkhart, IN, USA). Transverse cryostatections (30 �m) were cut and collected on Superfrost Plus slidesThermo Scientific, Braunschweig, Germany). Hybridization waserformed as previously described (Strahle et al., 1994).
For combined in situ hybridization and immunohistochemistry,ections were incubated for 2 h after the probe hybridization stepith blocking solution consisting of 20% normal horse serum plus0% Roche Blocking Reagent (Roche Products) in MABT (150M NaCl containing 100 mM maleic acid and 0.1% Tween 20, pH.5). Sections were then incubated with sheep-anti-nNOS (1:000) overnight at 4 °C in the blocking solution. Sections wereashed 3�10 min in MABT before incubation with donkey-anti-heep-IgG, conjugated with Alexa647 or Alexa 488 (Invitrogen;
:500). After 3�10 min washes with MABT, sections were post-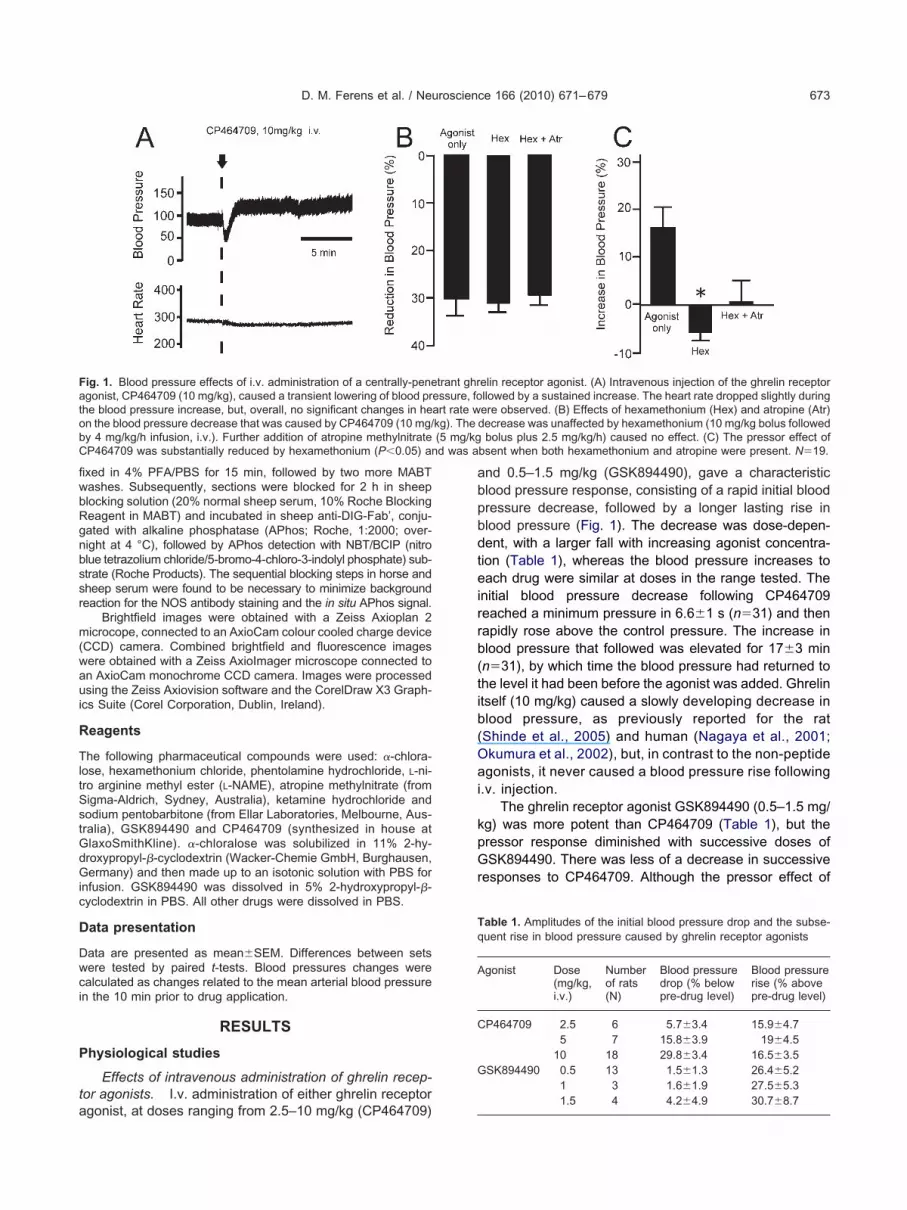
fiwbRgnbssr
m(waui
R
TltSstGdGic
D
Dwci
P
ta
abpbdteirrb(tib(Oai
kpGr
FatobC d was a
Tq
A
C
G
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679 673
xed in 4% PFA/PBS for 15 min, followed by two more MABTashes. Subsequently, sections were blocked for 2 h in sheeplocking solution (20% normal sheep serum, 10% Roche Blockingeagent in MABT) and incubated in sheep anti-DIG-Fab’, conju-ated with alkaline phosphatase (APhos; Roche, 1:2000; over-ight at 4 °C), followed by APhos detection with NBT/BCIP (nitrolue tetrazolium chloride/5-bromo-4-chloro-3-indolyl phosphate) sub-trate (Roche Products). The sequential blocking steps in horse andheep serum were found to be necessary to minimize backgroundeaction for the NOS antibody staining and the in situ APhos signal.
Brightfield images were obtained with a Zeiss Axioplan 2icrocope, connected to an AxioCam colour cooled charge device
CCD) camera. Combined brightfield and fluorescence imagesere obtained with a Zeiss AxioImager microscope connected ton AxioCam monochrome CCD camera. Images were processedsing the Zeiss Axiovision software and the CorelDraw X3 Graph-
cs Suite (Corel Corporation, Dublin, Ireland).
eagents
he following pharmaceutical compounds were used: �-chlora-ose, hexamethonium chloride, phentolamine hydrochloride, L-ni-ro arginine methyl ester (L-NAME), atropine methylnitrate (fromigma-Aldrich, Sydney, Australia), ketamine hydrochloride andodium pentobarbitone (from Ellar Laboratories, Melbourne, Aus-ralia), GSK894490 and CP464709 (synthesized in house atlaxoSmithKline). �-chloralose was solubilized in 11% 2-hy-roxypropyl-�-cyclodextrin (Wacker-Chemie GmbH, Burghausen,ermany) and then made up to an isotonic solution with PBS for
nfusion. GSK894490 was dissolved in 5% 2-hydroxypropyl-�-yclodextrin in PBS. All other drugs were dissolved in PBS.
ata presentation
ata are presented as mean�SEM. Differences between setsere tested by paired t-tests. Blood pressures changes werealculated as changes related to the mean arterial blood pressuren the 10 min prior to drug application.
RESULTS
hysiological studies
Effects of intravenous administration of ghrelin recep-or agonists. I.v. administration of either ghrelin receptor
ig. 1. Blood pressure effects of i.v. administration of a centrally-penegonist, CP464709 (10 mg/kg), caused a transient lowering of blood prhe blood pressure increase, but, overall, no significant changes in hen the blood pressure decrease that was caused by CP464709 (10 mg/y 4 mg/kg/h infusion, i.v.). Further addition of atropine methylnitrateP464709 was substantially reduced by hexamethonium (P�0.05) an
gonist, at doses ranging from 2.5–10 mg/kg (CP464709)
nd 0.5–1.5 mg/kg (GSK894490), gave a characteristiclood pressure response, consisting of a rapid initial bloodressure decrease, followed by a longer lasting rise inlood pressure (Fig. 1). The decrease was dose-depen-ent, with a larger fall with increasing agonist concentra-ion (Table 1), whereas the blood pressure increases toach drug were similar at doses in the range tested. The
nitial blood pressure decrease following CP464709eached a minimum pressure in 6.6�1 s (n�31) and thenapidly rose above the control pressure. The increase inlood pressure that followed was elevated for 17�3 minn�31), by which time the blood pressure had returned tohe level it had been before the agonist was added. Ghrelintself (10 mg/kg) caused a slowly developing decrease inlood pressure, as previously reported for the ratShinde et al., 2005) and human (Nagaya et al., 2001;kumura et al., 2002), but, in contrast to the non-peptidegonists, it never caused a blood pressure rise following
.v. injection.The ghrelin receptor agonist GSK894490 (0.5–1.5 mg/
g) was more potent than CP464709 (Table 1), but theressor response diminished with successive doses ofSK894490. There was less of a decrease in successive
esponses to CP464709. Although the pressor effect of
elin receptor agonist. (A) Intravenous injection of the ghrelin receptorllowed by a sustained increase. The heart rate dropped slightly duringere observed. (B) Effects of hexamethonium (Hex) and atropine (Atr)decrease was unaffected by hexamethonium (10 mg/kg bolus followed
bolus plus 2.5 mg/kg/h) caused no effect. (C) The pressor effect ofbsent when both hexamethonium and atropine were present. N�19.
able 1. Amplitudes of the initial blood pressure drop and the subse-uent rise in blood pressure caused by ghrelin receptor agonists
gonist Dose(mg/kg,i.v.)
Numberof rats(N)
Blood pressuredrop (% belowpre-drug level)
Blood pressurerise (% abovepre-drug level)
P464709 2.5 6 5.7�3.4 15.9�4.75 7 15.8�3.9 19�4.5
10 18 29.8�3.4 16.5�3.5SK894490 0.5 13 1.5�1.3 26.4�5.2
1 3 1.6�1.9 27.5�5.3
trant ghressure, foart rate wkg). The(5 mg/kg
1.5 4 4.2�4.9 30.7�8.7
GsOca
anhmd1posaHsobdmmcen
(saGs1ssb
uc(awrcrc�
pb
wbt(rih
G(iddaF(tprl
Fcri
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679674
SK894490 was prominent, this compound caused amaller drop in blood pressure, compared to CP464709.ut of 21 rats given i.v. GSK894490, blood pressure de-
reased in only four in response to 0.5–1.5 mg/kg ofgonist.
To test whether the initial blood pressure decrease wasconsequence of activation of autonomic nerve pathways,icotinic receptors of autonomic ganglia were blocked withexamethonium chloride (10 mg/kg bolus followed by 4g/kg/h infusion, i.v.). Hexamethonium caused a transientrop in blood pressure of about 30 mm Hg, that peaked at–2 min, following which the pressure returned towards there-hexamethonium level (Bogeski et al., 2005). At 10 minr more after addition of hexamethonium the blood pres-ure stabilized at 5–10 mm Hg below the original level andt this time the ghrelin receptor agonists were tested.examethonium did not attenuate the initial blood pres-ure decrease caused by either CP464709 (5–10 mg/kg)r GSK894490 (1.5 mg/kg), but blocked the increasedlood pressure elicited by these compounds (Fig. 1). Ad-ition of atropine methyl-nitrate (5 mg/kg bolus plus 2.5g/kg/h infusion, i.v.) in the continued presence of hexa-ethonium, had no effect on the blood pressure decrease
aused by the ghrelin receptor agonists, but in the pres-nce of both hexamethonium and atropine the agonists didot evoke a blood pressure increase (Fig. 1).
Addition of the nitric oxide synthase inhibitor, L-NAME10 mg/kg bolus followed by 4 mg/kg/h i.v.), caused aubstantial increase in blood pressure of 25.6�6.3%bove control blood pressure. In the presence of L-NAME,SK894490 (1 mg/kg intravenous bolus) elicited a tran-ient drop in blood pressure of 26.7�14 mm Hg, from02.6�13.6 to 75.9�22.3 mm Hg (n�3). Although thetarting blood pressures differ in the presence and ab-ence of L-NAME, it is clear that L-NAME did not reduce thelood pressure drop in response to GSK894490.
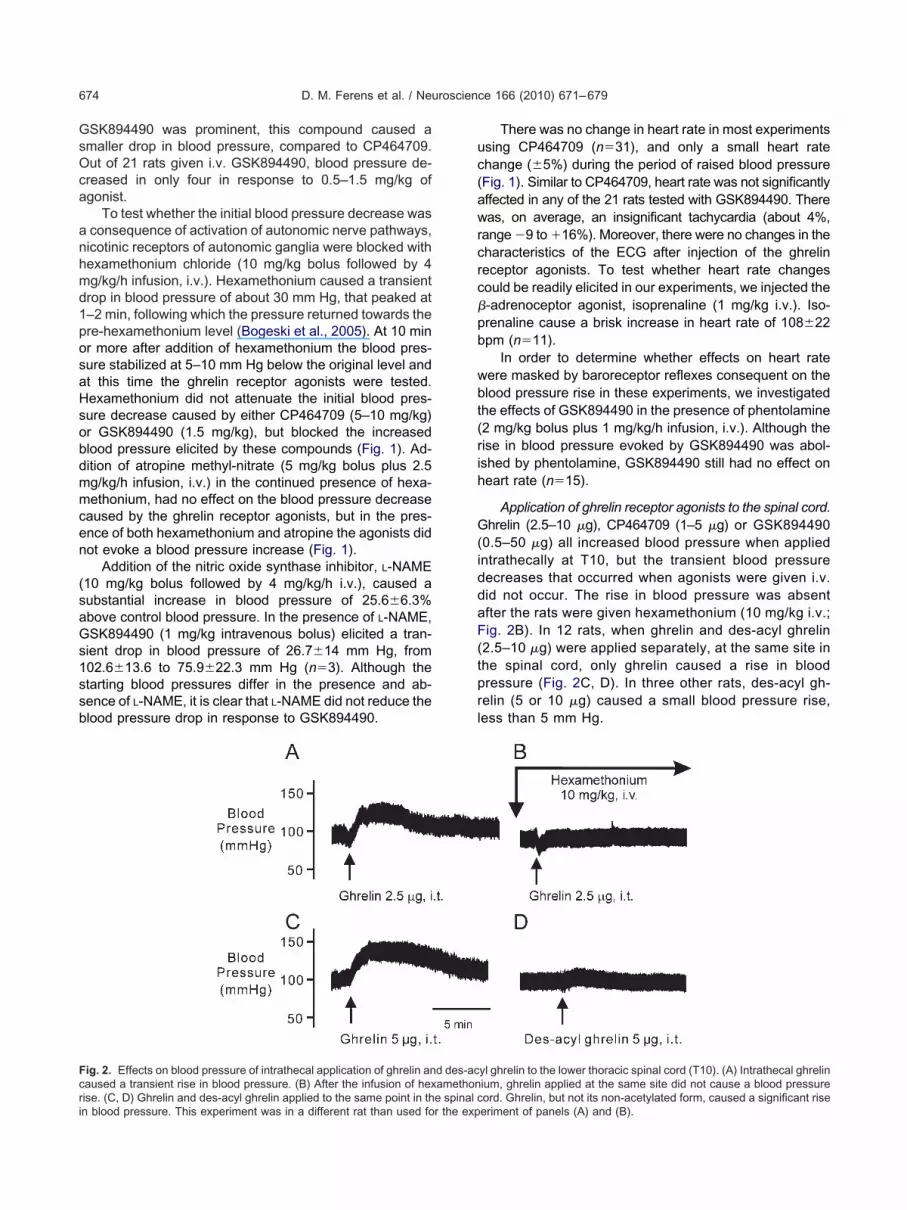
ig. 2. Effects on blood pressure of intrathecal application of ghrelin anaused a transient rise in blood pressure. (B) After the infusion of hex
ise. (C, D) Ghrelin and des-acyl ghrelin applied to the same point in the spinal cn blood pressure. This experiment was in a different rat than used for the expThere was no change in heart rate in most experimentssing CP464709 (n�31), and only a small heart ratehange (�5%) during the period of raised blood pressureFig. 1). Similar to CP464709, heart rate was not significantlyffected in any of the 21 rats tested with GSK894490. Thereas, on average, an insignificant tachycardia (about 4%,
ange �9 to �16%). Moreover, there were no changes in theharacteristics of the ECG after injection of the ghrelineceptor agonists. To test whether heart rate changesould be readily elicited in our experiments, we injected the-adrenoceptor agonist, isoprenaline (1 mg/kg i.v.). Iso-renaline cause a brisk increase in heart rate of 108�22pm (n�11).
In order to determine whether effects on heart rateere masked by baroreceptor reflexes consequent on thelood pressure rise in these experiments, we investigatedhe effects of GSK894490 in the presence of phentolamine2 mg/kg bolus plus 1 mg/kg/h infusion, i.v.). Although theise in blood pressure evoked by GSK894490 was abol-shed by phentolamine, GSK894490 still had no effect oneart rate (n�15).
Application of ghrelin receptor agonists to the spinal cord.hrelin (2.5–10 �g), CP464709 (1–5 �g) or GSK894490
0.5–50 �g) all increased blood pressure when appliedntrathecally at T10, but the transient blood pressureecreases that occurred when agonists were given i.v.id not occur. The rise in blood pressure was absentfter the rats were given hexamethonium (10 mg/kg i.v.;ig. 2B). In 12 rats, when ghrelin and des-acyl ghrelin
2.5–10 �g) were applied separately, at the same site inhe spinal cord, only ghrelin caused a rise in bloodressure (Fig. 2C, D). In three other rats, des-acyl gh-elin (5 or 10 �g) caused a small blood pressure rise,ess than 5 mm Hg.
yl ghrelin to the lower thoracic spinal cord (T10). (A) Intrathecal ghrelinium, ghrelin applied at the same site did not cause a blood pressure
d des-acamethon
ord. Ghrelin, but not its non-acetylated form, caused a significant riseeriment of panels (A) and (B).

ncmr2aab
pwetiw7oifpHmaCc
(Ct
D
mc(
at
riswrua
aihcctmr
rmnsn7Tuaga0twte
Tg
FampTtsata
Frct
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679 675
The pressor effect of non-peptide ghrelin receptor ago-ists was related to the spinal cord level (Fig. 3). Weonfirmed that the compounds were active in all experi-ents by comparing blood pressure and colonic motility
esponses to intrathecal agonist application (Shimizu et al.,006). Motility was increased when the agonists werepplied at T13–L4 vertebral levels (n�35), but not whenpplication was at low thoracic levels, where maximumlood pressure responses were obtained (n�13).
GSK894490 (5 �g), chosen because it is the moreotent agonist, was used to map the spinal cord sites thatere responsive. The greatest blood pressure increasingffects of GSK894490 (5 �g) were observed with applica-ions at T9 and T10 (Fig. 3). At T9, the blood pressurencreased by 75�33% over the value before GSK894490as applied, and at T10 there was a similar increase of3�15%. In comparison to T9 and T10, the hypertensionbserved at T8, and T11 to L4, ranged up to 24% in
ndividual experiments. At the most effective vertebral levelor eliciting colonic motility, L1, the effect on mean arterialressure was modest, an increase of 9.4�3.9% (n�14).eart rate fluctuations were observed in these experi-ents, but there was no consistent effect of the intrathecalpplication of GSK894490 at any level. Neither agonist,P464709 (10 �g) or GSK894490 (5 �g) applied intrathe-ally at T3–T5 affected heart rate.
Transection of the spinal cord at a mid thoracic levelT5–T6) did not affect the blood pressure rise caused byP464709 applied at T9–T10 (n�7; other compounds not
ested).
istribution of ghrelin receptor mRNA
Detection of mRNA by RT-PCR. Ghrelin receptorRNA was detected in extracts from the thoracic spinal
ord, the lumbar spinal cord and the medulla oblongata
ig. 3. Effects of the ghrelin receptor agonist, GSK894490 (5 �g),pplied at different vertebral levels, on blood pressure and colonicotility. Stimulation of ghrelin receptors caused increases in bloodressure (black bars), with maximum effects being observed at T9 and10 vertebral levels. Colonic motility was also increased (red) when
he agonist was applied at vertebral levels T13 to L3 (lower lumbar andacral spinal levels), but no motility responses were observed withpplication at T8 to T12. For interpretation of the references to color inhis figure legend, the reader is referred to the Web version of thisrticle.
Fig. 4). The products had the predicted molecular sizespt
nd were not observed in control reactions with reverseranscriptase omitted.
Localization: in situ hybridization. In order to test theeliability of the ghrelin receptor mRNA probe that was usedn the present study, we examined the localization of expres-ion in the hypothalamus, pons and medulla. Localizationas identical to that described by Zigman et al., in these
egions (Fig. 5). In addition, we conducted localization studiessing the corresponding sense probe, and did not observeny reaction in neurons of the brain stem or spinal cord.
Ghrelin receptor expression was detected in neuronst spinal cord levels T3 to S2, but not in other cell types, by
n situ hybridization. Neurons showing a positive in situybridization signal were located in the intermediolateralell columns (IML, Figs. 6–8) at all levels of the spinalord. Positive reactions also occurred in ventral horn mo-oneurons and in neurons in the region of the central grayatter (lamina X). Small numbers of weakly reactive neu-
ons were sometimes located dorsal to the IML.Greater numbers of ghrelin receptor expressing neu-
ons were in the IML at T10, compared to upper andid-thoracic and upper lumbar levels (Fig. 7). The greatestumbers of cells were observed at L4 to S2. Nitric oxideynthase (NOS) is a marker of a high proportion of auto-omic preganglionic neurons and is contained in about5% of nerve cells in the IML in the lower thoracic, T9–11, region (Hinrichs and Llewellyn-Smith, 2009). Wesed NOS immunoreactivity as an identifier of the IML, ands in indicator of the relative frequency of occurrence ofhrelin receptor expressing neurons (Figs. 6 and 8). Onlyminority of ghrelin receptor expressing neurons, 6.6�
.8% (n�10 samples in five rats) were NOS immunoreac-ive. The numbers of ghrelin receptor expressing neuronsas about 50% of the number of NOS neurons, suggesting
hat about 30–40% of sympathetic preganglionic neuronsxpress the receptor mRNA.
DISCUSSION
he present study provides evidence that autonomic pregan-lionic neurons in the spinal cord express the ghrelin receptor
ig. 4. PCR detection of ghrelin receptor message. Detection of gh-elin receptor messenger RNA in extracts from the thoracic spinalord, lumbar spinal cord and medulla oblongata. The bands were ofhe sizes predicted by the primers used for amplification, 716 base
airs (bp). �RT, with reverse transcriptase; �RT, without reverseranscriptase.

anpp
D
Gblbrt
wfhtrcnrgsoe
Fsai er imaget D), 20 �
Fail
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679676
nd that ghrelin receptor agonists can activate some of theseeurons to increase blood pressure. By contrast, activation oferipheral ghrelin receptors caused blood pressure to decrease,robably by a direct action on blood vessels.
istribution of ghrelin receptor in the spinal cord
hrelin receptor gene expression was localized to the cellodies of sympathetic preganglionic neurons at spinal cord
evels where ghrelin receptor agonists caused rises inlood pressure. Thus the sympathetic preganglionic neu-ons of vasoconstrictor pathways are the likely sites wherehe effects that we observed were exerted. Other neurons
ig. 5. Localization of ghrelin receptor mRNA in the medulla oblongaulcus (dms) and the nucleus gracilis (N gracilis). Numerous small nembiguous (NA), showing strongly reactive nerve cells. (C) High power
n the region of the rough endoplasmic reticulum (arrow). (D) High powo the nucleus of the neuron (arrow). Scale bars: (A, B), 100 �m; (C,
ig. 6. Localization of ghrelin receptor expression and nitric oxide synt T10 (A, A=, A==) and L6 (B, B=, B==). To create the merged imag
mmunoreactivity and ghrelin receptor mRNA are in separate neurons. Scale begend, the reader is referred to the Web version of this article.
ith receptor expression were located in the spinal cord,or example nerve cells dorsal to the IML, in the ventralorns and close to the central canal. Moreover, the recep-or expression occurred in autonomic preganglionic neu-ons of levels where effects of the agonists on cardiovas-ular functions were not observed. For example, althougheurons expressing the receptor gene were present, gh-elin receptor agonists did not activate sympathetic pregan-lionic cardioaccelerator neurons at T3–T5 and had incon-istent effects on blood pressure at these levels. Thusther roles of autonomic neurons in the spinal cord thatxpress ghrelin receptors can be anticipated. At this time,
w power image of the area postrema (AP), between the dorsomedialthe AP express the receptor gene. (B) Low power image of nucleus
f a neuron in the area postrema. Reaction is seen around the nucleus,of a neuron of the nucleus ambiguus. Staining is prominent adjacent
m.
OS) immunoreactivity in neurons of the intermediolateral cell columnages of panels (A) and (B) were inverted and colored green. NOS
ta. (A) Lourons ofimage o
thase (Ne, the im
ar: 50 �m. For interpretation of the references to color in this figure

tdt
I
TvoCradcda
w
accirhsvanwntpacs
Frnl
Fratf
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679 677
he only other role of spinal cord ghrelin receptors to beescribed is the activation of pathways controlling defeca-
ion (Shimizu et al., 2006; Shafton et al., 2009).
ncreases in blood pressure
he evidence for activation of preganglionic neurons inasoconstrictor pathways is, first, that the pressor effectsf i.v. delivery of two centrally-penetrant ghrelin agonists,P464709 and GSK894490, were blocked by the nicotinic
eceptor antagonist, hexamethonium. Second, when wepplied either of the ghrelin receptor agonists intrathecally,irectly to the spinal cord, we consistently observed in-reases in blood pressure. Moreover, ghrelin itself, but notes-acyl ghrelin, increased blood pressure when it waspplied intrathecally.
The blood pressure increases elicited by the agonistsere greatest at lower thoracic and upper lumbar regions,
ig. 7. Positions of neurons in which ghrelin receptor mRNA was locaight side of each panel is a micrograph of the region, and on the left isucleus (IMM, circle). The intermediolateral nuclei (IML) are circled in b
egend, the reader is referred to the Web version of this article.
ig. 8. Ghrelin receptor and nitric oxide synthase expressing neuroneceptor gene transcripts or neuronal nitric oxide synthase (NOS) in st different spinal cord levels, revealed by in situ hybridization (left) or
hese populations, about 7% of ghrelin receptor expressing neurons were NOSrom five rats.nd the increase in blood pressure caused by direct appli-ation of the agonists to the lower thoracic or lumbar spinalord was not affected by severing the cord at T5–T6,
ndicating that blood pressure is elevated by activation ofeceptors in the cord. The agonists had little or no effect oneart rate, whether they were applied i.v. or directly to thepinal cord at levels of origin of cardio-accelerator inner-ation. Thus ghrelin agonists do not cause a generalizedctivation of sympathetic preganglionic neurons in the spi-al cord. This is consistent with the localization studies,hich showed that only about half of the preganglioniceurons expressed the receptor. It was previously foundhat ghrelin receptor activation in the spinal cord stimulatesarasympathetic outflows to the distal bowel (Shimizu etl., 2006). This response was used in the present work toontrol for drug effectiveness when no cardiovascular re-ponse was found at the level tested. Which other auto-
n situ hybridization at spinal cord levels T10 and L6 (red dots). On theowing spinal laminae (1–10) and the position of the intermediomedialbar: 200 �m. For interpretation of the references to color in this figure
rent spinal cord levels. Numbers of neurons that expressed ghrelin�m sections through the intermediolateral nucleus of the spinal cordnoreactivity for NOS (right). There was only a small overlap between
lized by ia map shlue. Scale
s at diffeingle 30by immu
immunoreactive. Data are mean�SEM for 10 sections, 30 �m thick,

nl
D
IrsesbomwopssvKa2wladvbaIrhism
ttp2eiIprtdcIdn
norGStrv
coa
M
TsaaDdohHdfabce
WpwTt
ANnstN
B
B
C
D
E
G
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679678
omic outflows from the spinal cord are activated by ghre-in receptor agonists has not yet been investigated.
ecreases in blood pressure
n addition to the central, pressor, action of the ghrelineceptor agonists that has been identified, we have ob-erved decreases in blood pressure. This is not an indirectffect through stimulation of preganglionic neurons of va-odilator pathways, because the decrease was unaffectedy hexamethonium, which blocks activation of postgangli-nic neurons in these pathways, and it was not inhibited byuscarinic receptor antagonism or NOS inhibition, whichould be expected to block transmission from postgangli-nic vasodilator neurons. Shinde et al. (2005) also re-orted that inhibition of NOS does not reduce the hypoten-ion caused by ghrelin. Our data are consistent with ob-ervations that ghrelin is a potent dilator of blood vessels initro (Wiley and Davenport, 2002; Shimizu et al., 2003;leinz et al., 2006). In vivo application of ghrelin peptidelso caused a decrease in blood pressure (Nagaya et al.,001; Matsumura et al., 2002; Okumura et al., 2002),hich we confirmed in the present study. It thus seems
ikely that when ghrelin, or ghrelin receptor agonists, aredministered into the systemic blood circulation, they actirectly on the ghrelin receptors on blood vessels, to causeasodilation. In agreement with this, we found that loweredlood pressure in response to peripherally injected ghrelingonists preceded the sympathetically-mediated increase.n contrast to their direct effect on the vasculature, ghrelineceptor agonists appear to have no direct effect on theeart, as shown by the lack of effect on ECG or heart rate
n the present work. Consistent with our study, in othertudies ghrelin peptide did not change heart rate in hu-ans or rats (Nagaya et al., 2001; Shinde et al., 2005).
In contrast to the blood pressure increases exertedhrough ghrelin receptors in the spinal cord, ghrelin appliedo the brain-stem, in the region of the NTS, lowers bloodressure and reduces sympathetic nerve activity (Lin et al.,004). This region, and other brainstem centres that influ-nce the cardiovascular system, contain ghrelin express-
ng neurons (Zigman et al., 2006, current observations).CV injection of ghrelin also inhibited activity of renal sym-athetic nerves (Matsumura et al., 2002). Thus ghrelineceptor activation affects the cardiovascular system athree levels at least, the brain stem, the spinal cord andirectly on resistance vessels. The physiological signifi-ance of each of these sites of action is not yet known.nvestigation of their significance will be facilitated by theevelopment of effective non-peptide receptor antago-ists.
An endogenous ligand for ghrelin receptors in the spi-al cord has not yet been identified. However, ghrelinccurs in CNS neurons (Cowley et al., 2003), and ghrelineleased from such neurons may be the natural ligand.hrelin receptors are constitutively active (Holst andchwartz, 2004; Holst et al., 2006; Pantel et al., 2006). It is
hus possible that ligand-independent activity of ghrelineceptors in sympathetic vasoconstrictor neurons is in-
olved in the maintenance of blood pressure. If this is thease, inverse agonists, that block constitutive activity with-ut inhibiting ligand-mediated activation, could be effectiventi-hypertensive agents.
eal associated cardiovascular changes
he pattern of release of ghrelin into the circulation istrongly linked to feeding (Kojima et al., 1999; Inui, 2001),nd feeding is linked to cardiovascular changes (Waalernd Eriksen, 1992; Gentilcore et al., 2009; Harthoorn andransfield, 2008). Blood pressure has been reported toecline leading up to a meal, to rise transiently at the timef the meal, and then to decline over the postprandial 1–2(Gentilcore et al., 2009; Harthoorn and Dransfield, 2008).owever, meal associated changes in plasma ghrelin areependent on macronutrient content, and may increase,or example after a high fat meal, or decrease, for examplefter carbohydrate (Erdmann et al., 2004). Causal relationsetween feeding state, ghrelin release and blood pressurehanges need to be investigated in carefully controlledxperiments.
CONCLUSION
e conclude that ghrelin receptors are expressed by sym-athetic preganglionic neurons of vasoconstrictor path-ays and that their activation increases blood pressure.hus it may be feasible to target ghrelin receptors to con-
rol blood pressure.
cknowledgments—This work was supported by a grant from theational Health and Medical Research Council of Australia (Grantumber 508947). We thank Zhengdong Qu for assistance with initu hybridization. Microscope facilities were provided by the Aus-ralian Phenomics Network Histopathology and Organ Pathologyode.
REFERENCES
anks WA, Burney BO, Robinson SM (2008) Effects of triglycerides,obesity, and starvation on ghrelin transport across the blood–brainbarrier. Peptides 29:2061–2065.
ogeski G, Shafton AD, Kitchener PD, Ferens DM, Furness JB (2005)A quantitative approach to recording peristaltic activity from seg-ments of rat small intestine in vivo. Neurogastroenterol Motil17:262–272.
owley MA, Smith RG, Diano S, Tschöp M, Pronchuk N, Grove KL,Strasburger CJ, Bidlingmaier M, Esterman M, Heiman ML, Garcia-Segura LM, Nillni EA, Mendez P, Low MJ, Sotonyi P, Friedman JM,Liu H, Pinto S, Colmers WF, Cone RD, Horvath TL (2003) Thedistribution and mechanism of action of ghrelin in the CNS dem-onstrates a novel hypothalamic circuit regulating energy ho-meostasis. Neuron 37:649–661.
epoortere I (2009) Targeting the ghrelin receptor to regulate foodintake. Regul Pept 156:13–23.
rdmann J, Töpsch R, Lippl F, Gussmann P, Schusdziarra V (2004)Postprandial response of plasma ghrelin levels to various testmeals in relation to food intake, plasma insulin, and glucose. J ClinEndocrinol Metab 89:3048–3054.
entilcore D, Nair NS, Vanis L, Rayner CK, Meyer JH, Hausken T,Horowitz M, Jones KL (2009) Comparative effects of oral andintraduodenal glucose on blood pressure, heart rate, and splanch-nic blood flow in healthy older subjects. Am J Physiol 297:
R716–R722.
G
H
H
H
H
I
K
K
K
L
M
N
O
P
P
P
S
S
S
S
S
W
W
Z
D. M. Ferens et al. / Neuroscience 166 (2010) 671–679 679
uan XM, Yu H, Palyha OC, McKee KK, Feighner SD, SirinathsinghjiDJ, Smith RG, Van der Ploeg LH, Howard AD (1997) Distributionof mRNA encoding the growth hormone secretagogue receptor inbrain and peripheral tissues. Mol Brain Res 48:23–29.
arthoorn LF, Dransfield E (2008) Periprandial changes of the sym-pathetic–parasympathetic balance related to perceived satiety inhumans. Eur J Appl Physiol 102:601–608.
inrichs JM, Llewellyn-Smith IJ (2009) Variability in the occurrence ofnitric oxide synthase immunoreactivity in different populations ofrat sympathetic preganglionic neurons. J Comp Neurol 514:492–506.
olst B, Lang M, Brandt E, Bach A, Howard A, Frimurer TM, Beck-Sickinger A, Schwartz TW (2006) Ghrelin receptor inverse ago-nists: identification of an active peptide core and its interactionepitopes on the receptor. Mol Pharmacol 70:936–946.
olst B, Schwartz TW (2004) Constitutive ghrelin receptor activity as asignaling set-point in appetite regulation. Trends Pharmacol Sci25:113–117.
nui A (2001) Ghrelin: an orexigenic and somatotrophic signal from thestomach. Nat Rev Neurosci 2:1–10.
atugampola SD, Pallikaros Z, Davenport AP (2001) [125I-His(9)]-ghrelin, a novel radioligand for localizing GHS orphan receptors inhuman and rat tissue: up-regulation of receptors with atheroscle-rosis. Br J Pharmacol 134:143–149.
leinz MJ, Maguire JJ, Skepper JN, Davenport AP (2006) Functionaland immunocytochemical evidence for a role of ghrelin and des-octanoyl ghrelin in the regulation of vascular tone in man. Cardio-vasc Res 69:227–235.
ojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K(1999) Ghrelin is a growth-hormone-releasing acylated peptidefrom stomach. Nature 402:656–660.
in Y, Matsumura K, Fukuhara M, Kagiyama S, Fujii K, Iida M (2004)Ghrelin acts at the nucleus of the solitary tract to decrease arterialpressure in rats. Hypertension 43:977–982.
atsumura K, Tsuchihashi T, Fujii K, Abe I, Iida M (2002) Centralghrelin modulates sympathetic activity in conscious rabbits. Hyper-tension 40:694–699.
agaya N, Kojima M, Uematsu M, Yamagishi M, Hosoda H, Oya H,Hayashi Y, Kangawa K (2001) Hemodynamic and hormonal ef-fects of human ghrelin in healthy volunteers. Am J Physiol 280:
R1483–R1487.kumura H, Nagaya N, Enomoto M, Nakagawa E, Oya H, Kangawa K(2002) Vasodilatory effect of ghrelin, an endogenous peptide fromthe stomach. J Cardiovasc Pharmacol 39:779–783.
admanabhan R, Singh S (1979) Observations on the topographicalrelations of spinal nerve roots in the rat. Acta Anat 105:378–380.
an W, Tu H, Kastin AJ (2006) Differential BBB interactions of threeingestive peptides: obestatin, ghrelin, and adiponectin. Peptides27:911–916.
antel J, Legendre M, Cabrol S, Hilal L, Hajaji Y, Morisset S, Nivot S,Vie-Luton MP, Grouselle D, de Kerdanet M, Kadiri A, Epelbaum J,Le Bouc Y, Amselem S (2006) Loss of constitutive activity of thegrowth hormone secretagogue receptor in familial short stature.J Clin Invest 116:760–768.
hafton AD, Sanger GJ, Witherington J, Brown JD, Muir A, Butler S,Abberley L, Shimizu Y, Furness JB (2009) Oral administration of acentrally acting ghrelin receptor agonist to conscious rats triggersdefecation. Neurogastroenterol Motil 21:71–77.
himizu Y, Chang EC, Shafton AD, Ferens DM, Sanger GJ, Wither-ington J, Furness JB (2006) Evidence that stimulation of ghrelinreceptors in the spinal cord initiates propulsive activity in the colonof the rat. J Physiol (Lond) 576:329–338.
himizu Y, Nagaya N, Teranishi Y, Imazu M, Yamamoto H,Shokawa T, Kangawa K, Kohno N, Yoshizumi M (2003) Ghrelinimproves endothelial dysfunction through growth hormone-inde-pendent mechanisms in rats. Biochem Biophys Res Commun310:830 – 835.
hinde UA, Desai KM, Yu C, Gopalakrishnan V (2005) Nitric oxidesynthase inhibition exaggerates the hypotensive response to gh-relin: role of calcium-activated potassium channels. J Hypertens23:779–784.
trahle U, Blader P, Adam J, Ingham PW (1994) A simple and efficientprocedure for non-isotopic in situ hybridization to sectioned mate-rial. Trends Genet 10:75–76.
aaler BA, Eriksen M (1992) Post-prandial cardiovascular responsesin man after ingestion of carbohydrate, protein or fat. Acta PhysiolScand 146:321–327.
iley KE, Davenport AP (2002) Comparison of vasodilators in humaninternal mammary artery: ghrelin is a potent physiological antago-nist of endothelin-1. Br J Pharmacol 136:1146–1152.
igman JM, Jones JE, Lee CE, Saper CB, Elmquist JK (2006) Expres-sion of ghrelin receptor mRNA in the rat and the mouse brain.
J Comp Neurol 494:528–548.(Accepted 4 January 2010)(Available online 6 January 2010)





























