Embed Size (px)
Citation preview

UMEÅ PSYCHOLOGY SUPPLEMENT REPORTS
Supplement No. 4 2004
GAZE CONTROL IN EPISODIC MEMORY
Linus Holm

Umeå Psychology Supplement Reports
Acting EditorBo Molander
Associate EditorsAnders BöökEva SundinAnn-Louise Söderlund
Editorial BoardKerstin ArmeliusAnders BöökBo MolanderTimo MäntyläEva Sundin
This issue of Umeå Psychology Supplement Reports, and recent issues of other departmental reports areavailable as pdf-files. See the home page of Department of Psychology (http://www.psy.umu.se/forskning/publikationer/inst-rapportserie/UPSR.htm).
Department of PsychologyUmeå UniversitySE-901 87 Umeå, Sweden
ISSN 1651-565X

Holm, L.(2004). Gaze control in episodic memory. Department of Psychology, Umeå University, S-901 87, Sweden
Abstract
The role of gaze control in episodic recognition was investigated in two studies. In Study 1, participants encoded human faces inverted or upright, with or without eye movements (Experiment 1) and under sorting or rating tasks (Experiment 2) respectively. At test, participants indicated their recollective experience with R(emember) responses (explicit recollection) or K(now) responses (familiarity based recognition). Experiment 1 showed that face inversion and occlusion of eye movements reduced levels of explicit recollection as measured by R responses. In Experiment 2, the relation between recollective experience and perceptual reinstatement was examined. Whereas the study instructions produced no differences in terms of eye movements, R responses were associated with a higher proportion of refixations than K responses.
In Study 2, perceptual consistency was investigated in two experiments. In Experiment 1, participants studied scenes under different concurrent tasks. Subsequently, their recognition memory was examined in a R / K test. Executive load produced parallel effects on eye movements and R responses. Furthermore, R responses were associated with a higher proportion of refixations than K responses. However, number of fixations was correlated with refixations. Experiment 2 corroborated these results and controlled for number of fixations.
Together, these studies suggest that visual episodic representations are supported by perceptual detail, and that explicit recollection is a function of encoding and retrieving those details. To this end, active gaze control is an important factor in visual recognition. This thesis for the licentiate degree is based on the following studies: Holm, L, & Mäntylä, T.(2004). Gaze control in scene recognition: refixations reflect recollection.
Manuscript submitted for publication. Mäntylä, T, & Holm, L.(2004). Gaze control and recollective experience in face recognition.
Manuscript submitted for publication.


2
Acknowledgments
I am indebted to several people for support in producing this thesis. Firstly, my partner
Helena Pettersson encouraged me to apply for graduate school. Secondly, my supervisor Timo Mäntylä guided me through the work. Thirdly, the department of psychology at Umeå University including staff, faculty and fellow graduate students helped me in various matters. Lastly, I would like to thank Robert Bjork, cogfog and the people at UCLA for a perfectly splendid spring in a scientifically stimulating environment.


5
Contents INTRODUCTION ……………………………………….………………………………5 Vision……………………………………………….……....………………………….. 5 Eye movements and recognition memory………….…………………………………… 6
Dual components in recognition memory…………….…………………………………8 The concept of distinctiveness…………………….……………………………………. 9 Frontal lobe functions and elaborative processing ...…..…………………………………10 Attention and eye movements……………………………………………………….. ….10 Perceptual reinstatement……………………………………………………………….. 11 Research objectives………………………………………………………………………12
EMPIRICAL STUDIES………………………………………………………....................12 Study 1: Gaze control and recollective experience in face recognition……………………12 Study 2: Gaze control in scene recognition: Refixations reflect recollection…...…............14
GENERAL DISCUSSION……..…………………………………………………………. 16 REFERENCES……………………………………………………………………………..18


5
Gaze control in episodic memory Linus Holm
INTRODUCTION Episodic recognition memory refers to our ability to match consciously stored representations with current perceptual impressions. Therefore perception or perceptual behaviour profoundly constrains what is encoded and which information will serve as efficient retrieval cues in recollection. Previous experimental investigations of episodic memory have mostly been limited to inferences about perceptual behaviour, based on e.g. recognition performance and response times. Instead, an immediate investigation of perceptual behaviour should provide valuable information regarding episodic memory and its underlying mechanism. Within the field of vision, this perceptual behaviour amounts to eye movements and gaze control. Gaze control refers to the guidance of fixations in perceptual, cognitive and behavioural activity (Henderson, 2003). This thesis investigates the relation between gaze control and levels of awareness in recognition memory, a subject which has scarcely been studied before. Explicitly, the question under examination is whether eye movements reflect explicit memory in general and levels of awareness in recognition in particular. This thesis is guided by the hypothesis that the visual representation is based on the distribution of fixations across the scene, and that conscious recollection is achieved through the accurate reinstatement of that distribution. I will present empirical support for a functional relation between gaze control and levels of awareness in visual recognition memory, with emphasis on perceptual detail. Before I disclose the results, I will present the rationale for my position and summarize earlier research on eye movements and recognition memory.
Vision Although the visual field encompasses approximately 200° horizontally by 135° vertically, acuity is highly variable over that area (Wandell, 1995). The highest acuity is provided by the central 2 horizontal x 1vertical degrees of the visual field which corresponds to the fovea (Nelson & Loftus, 1980). Acuity rapidly decreases towards the visual periphery. For instance, at a distance of only 5 degrees from the fovea, visual acuity is reduced to 50%, and 10% at 40 degrees eccentricity (Hochberg 1978 in Irwin, 1992). Furthermore, this relationship seems at least in part to carry over to discrimination performance of complex objects. For instance, Nelson and Loftus showed that discrimination performance for category changed objects in a forced choice task decreased as a function of object distance from the central visual field. Specifically, a large decrease in performance was found from 0 to about 2 degrees eccentricity. While peripheral object identification might be worse than at the fovea, object identification can still be reasonably good. In an experiment adopting an artificial scotoma, Henderson and colleagues (Henderson, McClure, Pierce & Schrock, 1997) showed that identification of simple line drawings was relatively preserved outside the fovea. However, compensatory mechanisms might explain these results. For instance, the paradigm probably reflected a change in attention distribution towards the visual periphery. Hence, covert visual attention, rather than central vision might explain successful identification. Nevertheless, as long as attention can be assumed to be reflected in eye movements during normal, unconstrained vision, eye movements will serve as an important indicator of visual perception. In fact, there seems to be an obligatory coupling between saccade programming and visual attention (Deubel & Schneider, 1996).

6
In stable environments and observers, human vision is characterized by saccadic eye movements. Saccades occur with an approximate frequency of 3 to 5 Hz (Yarbus, 1967) and are fast (up to 900 degrees per second) ballistic movements. The ability of the oculomotor system to carry out saccades seems to solve the demand for high acuity access to the environment in the presence of a visual field with variable acuity.
Where we look depends on perceptual qualities in the scene like contrast and texture (Mannan, Ruddock & Wooding,1997, Reinagel & Zador, 1999), but also more abstract attributes like semantic relations (Friedman, 1979; Mores, Laiti & Chelazzi 2003; Yarbus 1967). Extended viewing of the same scene also alters the distribution of fixations (Mannan, Ruddock & Wooding, 1997; Yarbus, 1967), generally from highly salient regions to less salient regions (Antes 1974, see also Groner, Walder & Groner, 1983).
Long term memory has been shown to affect visual search in a variety of studies, including search for schema inconsistent objects in scenes (Friedman 1979; Henderson, Weeks & Hollingworth 1999), to effects of distractors semantically (but not visually) similar to targets in non scene displays (Mores, Laiti & Chelazzi 2003). Furthermore, familiarity with distractors or target (Tong & Nakayama, 1999) or distractor configuration (Chun & Jiang, 2003) facilitates search.
An important, underlying assumption in this thesis is that eye movements reflect visual attention. Undoubtedly, spatial attention can be directed away from the current fixation location. The question is to what extent this happens in normal visual perception. Specifically, how are overt changes of focus (i.e. eye movements) and covert allocation of attention related? Henderson provided an explanation (1992) based on a sequential attention model for eye movement control in complex visual tasks. According to the model, attention allocation always determines and precedes eye movements giving rise to a “rubber band” relation between attention and eye movements. Hence, fixations can be considered to reflect at least a subset of the deployment of spatial attention. More specifically, Henderson claimed that attention is initially allocated to the centre of fixation and is shifted away from that location when the content has been processed up to a criterion level. This view has received support from several subsequent studies (Deubel & Schneider, 1996; Kowler, Anderson, Dosher & Blaser, 1995; van Diepen & d'Ydewalle, 2003).
Alternatively, attention is not well characterized by a serial process, but instead a process with at least some parallel component (Wolfe, 2003). On this account of attention, the role of eye movements become less important, or in the least, would be expected to provide a rather noisy picture of the distribution of visual attention. Furthermore, given that visual processing during identification might be devoted to diagnostic scales of the stimulus, (e.g. Schyns & Oliva, 1997), large scales (low spatial frequency) might provide sufficient visual information to qualify as sufficient for scene recognition (see also Rousselet, Thorpe & Fabre-Thorpe, 2004 for a discussion). Such information should not be as dependent on foveal vision. As it stands, the role of focal attention in scene perception and recognition, as expressed in overt eye movements reflecting fovea size receptive fields, is an empirical question.
Eye movements and recognition memory Although eye movements seem an obvious thing to study in order to understand visual long term memory, surprisingly few studies have been published on the subject. An early attempt to establish a relation between scene memory and eye movements was made by Noton and Stark (1971a, 1971b). Following Hebb (1949), Noton and Stark claimed that visual inspection gives rise to a scanpath, a repetitive sequence of eye movements, specific for the observer watching a particular visual stimulus. In Noton and Starks’ own words, “Just as each person develop his own characteristic habits of behaviour for each situation with which he is familiar, so he may be expected to develop his own characteristic perceptual habits, his scanpath for each pattern he can recognize” (Noton & Stark, 1971a). The scanpath is stored as a sensory and motor connection called the feature ring. Recognition is characterized by the sequential reinstatement of the scan

7
pattern. In other words, the sequential reinstatement of eye movements is a prerequisite for recognition, scanpath and recognition are mutually dependent, because they are part of the same process. Noton and Stark (Noton & Stark, 1971a) observed the scanpath as often as 65 % of the cases within the first fixations on repeated scenes, hence they claimed that top-down gaze control is activated early. Their experiment was however based on very few participants and a very limited number of pictures presented. Furthermore, these pictures were somewhat blurred, in order to boost the role of foveal vision and hence increase the effect under investigation.
Although the scanpath hypothesis in its original formulation has failed to receive empirical support (Locher & Nodine, 1974; Walker-Smith, Gale & Findlay, 1977), it has not been succeeded by stronger and more consistent theories of the functional relationship between eye movements and scene recognition. In fact there is serious doubt about the meaningfulness in relating eye movements to visual long term memory (Fisher, Karsh, Breitenbach, & Barnette, 1983; Melcher & Kowler, 2001).
One problem with the scanpath hypothesis seems to be its exclusive reliance on central visual processing. Parker (1978), suggested an alternative account of eye movements in scene recognition, based on the idea that eye movements are successively planned according to peripheral information. Hence, the content of peripheral vision is compared to stored information, and the outcome of that comparison guides the subsequent saccade towards a diagnostic area. Based on the results of his experiments, he suggested that a conservative measure for an efficient visual field in terms of saccadic planning would correspond to approximately 6.5 degrees in his experimental setting. Although Parker defended a position where continuous peripheral information evaluation guides eye movements during scene recognition, he actually received some support for the scanpath hypothesis. His participants displayed rather consistent and sequentially constrained scanning patterns across repeated exposures of the stimuli. In fact, the most common sequence through the six objects (e.g. a,b,c,d,e,f) of his stimuli scenes explained 55% of all object transitions. When the opposite direction was included (e.g. f,e,d,c,b,a), these two directions accounted for an impressive 75% of object transitions. This should be considered in relation to the 120 possible paths connecting all objects in his stimuli. However, contrary to the prediction of the scanpath hypothesis, Parker found that in scenes with changed objects, participants consistently “jumped ahead” in their scan patterns, reflecting evaluation of peripheral information.
Another serious problem with the scanpath hypothesis seems to be its reliance on sequential order. Several subsequent studies found support for the presence of idiosyncratic sequences of eye movements, yet they all failed to support the relation between recognition and sequential reinstatement (Locher & Nodine, 1974; Walker-Smith, Gale & Findlay, 1977). Instead, a decrease in sequential eye movement dependency has been shown to relate to memory (Althoff, 1998; Althoff & Cohen, 1999). However, this eye movement based memory effect was shown in the context of implicit memory.
A general problem with assigning gaze control a key role in scene recognition is that humans are able to recognize scenes given very short presentation times. For instance, Intraub (1981), showed that participants were rather successful at discriminating previously seen pictures from their mirror reversals in a test following only 110 ms study time. Furthermore, the gist of a scene can be captured within the first fixation (Antes 1974; Antes, Penland & Metzger, 1981).
Although human scene recognition can be very good in the absence of extensive perceptual examination, recognition performance certainly benefits from extended viewing. For instance, Loftus suggested that distributing fixations across the scene increases the likelihood of fixating memorable features, explaining the correlation between number of study fixations and recognition performance (Loftus 1971; 1982, Nelson & Loftus, 1980). In addition, highly salient regions attract a lot of overt attention (Antes 1974; Friedman 1979; Loftus 1978, Parker, 1978). Investigating this finding further, Hollingworth, Williams and Henderson (2001) found support for Parker’s (1978) suggestion that fixations could explain detection performance for changed scene regions. In a forced choice task, scene items were presented simultaneously with items from

8
the same semantic category. Henderson’s participants performed well above chance, provided that they had fixated the object during scene inspection. Together, these results suggest that eye movements support visual representation by providing relatively detailed visual information and that informative features are extensively processed receiving more fixations than non-salient objects (Friedman, 1979; Henderson, Weeks, & Hollingworth, 1999).
The above mentioned studies imply that scene recognition might be based on rather holistic, unspecific representations of scenes (Intraub, 1981), whereas memory for scene objects seem to rely on fixations (i.e. high acuity information, Hollingworth, 2004).
One interpretation of these results is that they reflect different representational qualities, so that holistic information might correspond to a rather vague familiarity with the scene, whereas specific detail representations provide source information. This interpretation is consistent with most dual-component theories of recognition memory (Jacoby, 1991; Mandler 1980; Tulving, 1985; Yonelinas, 2002). Before this relationship is further developed, I will summarize past research on dual component perspectives on recognition memory.
Dual components in recognition memory According to dual-process models of recognition memory, overall recognition performance reflects different states of conscious awareness For instance, you might not immediately recognise your neighbour when meeting her in the city, although you may have a strong feeling that the woman is familiar. Instead, meeting her outside the door to your apartment, you instantly recognise the person and vividly recall who she is (cf. Mandler, 1980). One approach in studying differences in states of consciousness is to have participants introspect on the nature of the experience that accompanies a recognised event. Tulving (1985) introduced such a paradigm by distinguishing between two different descriptions of the subjective experience of recognition. At test, participants were instructed that if they had a vivid recollection of the previously studied event (typically a word), then the participant should reply “Remember.” Instead, if the test item was recognised in the absence of a detailed recollection from its prior presentation, but rather on the basis of a feeling of familiarity, the participant should reply “Know “ (see Gardiner & Java 1993, for a review). The underlying assumption is that conscious awareness reflects two distinct forms of recollective experience. This phenomenological distinction has since Tulving´s (1985) original work, been used to functionally dissociate recollection from assessment of familiarity on several different variables. For instance, divided vs. undivided attention (Gardiner & Parkin, 1990) and perceptual change in repeated presentation of faces (Mäntylä & Cornoldi, 2002) selectively influenced how well the items were consciously recollected whereas same vs. different modalities across study and test (Gregg & Gardiner, 1994) selectively affected assessment of familiarity. Other studies have shown opposing effects on “Remember” and “Know” responses (Gardiner & Java, 1990; Mäntylä, 1997). Apart from this behavioural evidence, the distinction between different states of awareness in recognition has been supported by fMRI studies (Eldridge, Knowlton, Furmanski, Bookheimer & Engel, 2000; Henson, Rugg, Shallice, Josephs, & Dolan, 1999) relating state specific activation to different brain regions and neuropsychological studies (Blaxton & Theodore, 1997; Knowlton & Squire, 1995; Levine et al., 1998) showing reduced memory functions related to focal brain lesions. Specifically, both patient lesion studies and fMRI studies suggest that hippocampus affect recollection but not, or to a lesser extent, familiarity. Instead, the temporal lobes have been suggested to affect familiarity but not recollection. Furthermore, damage to the frontal lobes, particularly the dorsolateraral and ventral prefrontal cortex seems to affect recollection and to a less extent familiarity (see Yonelinas 2002a for an overview). Although past research supports this distinction of states of awareness in recognition, the explanation of its underlying mechanisms is still open to debate. According to Tulving´s

9
(1985) account, these states of awareness correspond to the episodic and semantic memory systems, where Remember responses are thought to reflect output from the former and Know responses to reflect output from the latter system. According to Tulvings view, episodic memory can only exist in the presence of semantic memory for the episode. Hence, remembering is knowing with an additional episodic character, and therefore redundant with respect to semantic memory (Knowlton, 1998; Tulving, 1985). Gardiner and Java (1990, 1993) extended Tulving´s original study, and proposed a modified systems account for the observed dissociations. They argued that Remember responses reflect output from an episodic memory system, characterised by conceptual factors, whereas Know responses reflect output from a procedural memory system, characterised by perceptual factors. This idea was based on the notion that several variables dissociating Remember and Know responses had parallel effects on performance in explicit and implicit memory tasks. For instance, only Remember responses (as well as explicit memory) were affected by levels of processing (Gardiner, 1988), word frequency (Gardiner & Java, 1990), divided attention (Gardiner & Parkin, 1990). On the other hand, only Know responses (as well as implicit memory) were affected by masked repetition priming (Rajaram, 1993) and modality match across study and test (Gregg & Gardiner, 1994). Knowlton and Squire (1995) questioned Gardiner and Java´s (1990) account and argued that the observed results reflected dissociations within the explicit memory system. Specifically, Knowlton and Squire showed that both Remember and Know responses were reduced in amnesics compared with normal controls. Gardiners (Gardiner & Java, 1990; 1993) conceptual/ perceptual account of the R / K distinction has also been refuted in studies demonstrating a reversed pattern of results. Specifically, conceptual manipulation has been found to influence Knowing (Mäntylä, 1997, Rajaram & Geraci 2000, Wagner, Gabrieli, & Verfaillie, 1997), and perceptual factors to influence Remembering (Mäntylä, 1997, Rajaram, 1996). To account for these findings, Rajaram (1996) proposed an alternative framework. She claimed that distinctiveness, rather than conceptual factors, affects Remembering, whereas fluency across study and test, rather than perceptual factors, affects Knowing. Rajaram stated that “an analysis of the distinctive or salient attributes of the information, be they conceptual or perceptual in nature, creates memories that are later accompanied by the subjective experience termed as Remember” (Rajaram, 1996, pp. 374). In support of this statement, Rajaram (1996) obtained a picture superiority effect for Remember responses (perceptual distinctiveness). Furthermore, Dewhurst and Hitch (1999) showed that effortful compared to shallow processing facilitated Remember responses (conceptual distinctiveness). Rajaram followed Luo´s (1993) notion that enhanced perceptibility (that is, how easily an item is perceived) is related to enhanced feelings of recognition. Rajaram suggested that fluency describes the necessary means for the establishment of feelings of familiarity. In Rajaram´s account, fluency means that similarities between stimuli at encoding and the subsequent recognition phase facilitate the experience of familiarity, measured by Know responses. The advantage of the fluency account over the perceptual processing explanation of Know responses is that the source of fluency can be either conceptual or perceptual. For instance modality match across study and test (perceptual fluency; Gregg & Gardiner, 1994) as well as semantically related primes (conceptual fluency; Rajaram & Geraci, 2000) facilitate Knowing. However, Rajaram did not provide an explanation for underlying memory systems related to the process categories. The concept of distinctiveness As noted above, the distinctiveness / fluency account seems to provide the most consistent explanation of the observed dissociations between Remembering and Knowing (Rajaram, 1996). However, instead of describing the concept of distinctiveness at any length, Rajaram (1996; see

10
also Rajaram & Geraci, 2000) refers to Hunt and his collaborator’s integrative framework on distinctiveness and organization (Hunt & Einstein, 1981, Hunt & Seta, 1984). According to Hunt (see Hunt & McDaniel, 1993, for a review), processing both distinct differences and relational similarities among the properties of the to the remembered item is crucial for optimal memory performance. Distinctiveness is a psychological resultant, not an inherent property of the stimulus. It seems that distinctiveness depends on the establishment of associations between the to-be-remembered item and its context, similar to source memory. Furthermore, the episodic distinction warrants that the to-be-remembered item is sufficiently deeply encoded so as to distinguish it from similar instances from the same category. Accordingly, the distinctiveness explanation of conscious recollection stresses the importance of elaborative or executive processing. For instance, levels of processing and divided attention selectively affects R responses (Gardiner & Parkin, 1990; Reinitz, Moorisey & Demb, 1994; see Yonelinas, 2002 for a review). In addition, executive processes are dependent on frontal lobe functioning (Fuster, 2000), a relationship which will be further discussed below.
Frontal lobe functions and elaborative processing Several studies support the notion that conscious recollection reflects elaborative processing involving frontal lobe functioning, particularly during encoding, but also in retrieval (Dewhurst & Hitch, 1999; Gardiner, 1988; Gardiner & Parkin, 1990, see also Hicks & Marsh, 2000). In addition, amnesic and non-amnesic patients with frontal-lobe lesions show selective problems in source recall and R/K judgments (Janowsky, Shimamura, & Squire, 1989; Levine et al., 1998; Levine, Freedman, Dawson, Black, & Stuss, 1999; Shimamura & Squire, 1987; but see also Thaiss & Petrides, 2003). While modularization of prefrontal functions seems somewhat dubious in light of findings showing overlapping networks within the prefrontal cortex (Duncan & Owen, 2000; Fuster, 2000), one prominent part seems to be the dorsolateral prefrontal cortex (DLPFC) in memory encoding and retrieval. Furthermore, the DLPFC is involved in voluntary gaze control (Pierrot-Deseilligny, Müri, Ploner, Gaymard, & Rivaud-Pechoux, 2003; Pierrot-Deseilligny, Milea, & Müri, 2004). Specifically, the DLPFC is activated in memory guided saccade tasks, suggesting that it has a role in spatial short-term memory. Furthermore, DLPFC is suggested to be involved in decision processes concerning eye movement targeting, inhibiting un-appropriate alternatives (i.e. in antisaccade tasks) and contributing to execution of predictive eye movements (learned saccade sequences). DLPFC exerts its control of eye movements by connecting to the frontal eye field (FEF), which also receives more direct visual input, suggested to reflect bottom-up visual control (Pierrot-Deseilligny, Milea, & Müri, 2004), from the occipital cortex via the intraparietal cortex (IP) (Pierrot-Deseilligny, Milea, & Müri, 2004). However, there is support for the processing of goal directed, internally motivated saccades in IP as well (Konen, Kleiser, Wittsack, Bremmer & Seitz, 2004). The FEF in turn projects to the superior colliculus. Although the DLPFC might not be the single structure responsible for top-down saccade control, it seems to constitute a common denominator for gaze control and levels of awareness in recognition. Generally, the prefrontal cortex (Fuster, 2000), and possibly specifically the DLPFC constitute a focal point at the top of the perception action cycle, and hence might be a good candidate for a structure responsible for continuous evaluation of long-term representations in relation to incoming perceptual information, contributing to appropriate eye guidance.
Attention and eye movements Processing the visual environment seems to be executively demanding, as the visual system has to react to new information with every fixation on a new area. Accordingly, there is an attentive cost inherit in saccade planning (Brockmole, Carlson & Irwin, 2002). Furthermore, Glenberg,

11
Schroeder and Robertson (1998) showed that human subjects tend to avert their gaze while solving non visual tasks, and that aversion actually facilitated the subjects in solving the tasks. Glenberg and colleagues thus interpreted aversion as a means of disengagement from the visual environment during non-visual problem solving. That is, scanning the visual environment necessarily carries an attentive cost. In addition, eye movement behaviour is affected by dual task demands. For instance, Loftus (1972) showed that counting backwards while inspecting scenes, reduced fixation frequency. In a naturalistic setting, Recarte and Nunez (2000) showed that drivers fixate longer, make shorter saccades and distribute gaze to a limited field of view while driving and simultaneously carrying out various mental operations. Furthermore, several studies have shown that dual task demands specifically affect levels of conscious recollection (Parkin, Gardiner & Rosser, 1995, Yonelinas, 2001). Together, these findings suggest a relationship among gaze control, executive functioning and conscious recollection. Encoding and retrieval relations
The very concept of recognition hinges upon reinstatement: there must be a similarity between the current perception and a representation for recognition to occur. Generally, the relation between encoding and retrieval has long since been appreciated as essential in memory. For instance, memory theories such as the encoding specificity principle (Tulving, 1983) and the transfer appropriate processing account (see e.g., Roediger, Weldon & Challis, 1989) stress that recollection is related to the presence of information relating to the encoding event. In the context of eye movements, the transfer-appropriate processing in episodic retrieval might correspond to study-test consistency in terms of eye fixations. From that perspective, two identical pictures are perceived as similar insofar as they are viewed in the same way. Only those fixations that are reinstated during retrieval provide functional retrieval information for recognition. Perceptual reinstatement The outline sketched above seems to be consistent with Noton and Stark’s (1971a; 1971b) scanpath hypothesis, however the memory theories mentioned make no strong claims regarding sequential order. Instead, a more flexible proposition would be to focus on the importance of reinstatement of study fixations during recognition, without the restriction of a sequential order. This general approach is also more in line with a Popperian perspective, i.e., to first establish the phenomena (a relation between eye movements and recognition memory) and later on develop a more elaborate theoretical explanation (e.g. the scanpath hypothesis or hypothesis based gaze control). The relation could be expressed (in a recognition test) by the degree of refixations of areas fixated during encoding. Considering the rapidly decreasing acuity towards the eccentricity, refixations should be very accurate in order to convey fateful perceptual retrieval cues. Given that every fixation during encoding has a probability of supporting a visually detailed representation, then every refixation during test would have a probability of causing a match with that representation (see Deco & Schurmann, 2000, for a similar idea). This approach would also be more flexible with regard to the underlying mechanisms of gaze control in recognition. For instance, the unconstrained proportion of refixations across study and test could as well be an effect of an execution of a whole sequence of saccades, based on a procedural representation, as on continual hypothesis testing or bottom-up guidance based on the visual content.
Under the assumption that explicit recognition benefits from refixations, a scan pattern leading to successful recognition (Hit) should be distributed more effectively across prior fixation sites, compared to incorrect recognition (Miss). In contrast, given that the decision is developed throughout the visual inspection, a failure to re-fixate should produce a weaker match or even strengthen the viewer’s belief that the scene is new, and therefore lead to a miss. According to the dual-process framework suggested by Rajaram (1996), one would expect perceptual match to facilitate familiarity effects rather than conscious recollection due to fluency of processing across study and test. Therefore K responses should be related to study-test

12
consistency in terms of eye movements. However, every fixation is related to a probability of elaborative content processing, which implies conscious recollection to be related to study-test consistency. This hypothesis was based on the notions that explicit recollection (in terms of R judgments or source recall; see Johnson, Hashtroudi, & Lindsay, 1993, for a review) reflects encoding of distinctive event attributes (Mäntylä, 1997; Rajaram, 1996), and that encoding of specific details requires focal attention (i.e., fixations, see Hollingworth & Henderson, 2002). One interpretation of the R/K distinction states that R and K responses reflect different response criteria based on one common trace strength dimension (Donaldson, 1996). While this perspective have difficulties explaining specific effects of K responses and predicting differences in R and K responses with intact overall discrimination performance, at least K responses seem to be well modelled by signal detection theory (Dobbins, Kroll & Liu, 1998; Yonelinas 2002). One candidate to a trace strength dimension might be perceptual consistency across study and test. From this point of view, R responses should be related to higher study-test consistency than K-responses, which in turn would be predicted to reflect a higher study-test consistency than misses. Research objectives In the following experiments, I will investigate whether a functional relationship between episodic visual memory and gaze control exists, and if found, characterize that relationship following the theoretical outline presented above. According to Hunt and McDaniels notion (1993), distinctiveness should be accrued through the processing of differences in the presence of similarity. This might refer to finding and processing salient attributes in the visual environment (Loftus, 1972, Friedman 1979, Henderson, Weeks & Hollingworth, 1999). Hence, unconstrained (Loftus 1972) and explorative vision (Loftus 1972; Loftus, 1981) should be important for efficient visual encoding. In contrast, constraining eye movements during encoding should be detrimental to memory. Specifically, this should affect conscious recollection, conditional on its reliance on distinctive event attributes (Mäntylä 1997; Rajaram 1996; see also Johnson, Hashtroudi & Lindsay, 1993). The empirical take off for this thesis was to manipulate eye movements directly or indirectly, in order to assess a functional relationship between gaze control and episodic memory.
EMPIRICAL STUDIES
Study 1: Gaze control and recollective experience in face recognition In two face recognition experiments, eye movements were registered during study and test to provide insight in the role of gaze control in face recognition. Limited effects in earlier studies relating eye movements to visual memory might be due to assessment of overall recognition instead of its components of explicit recollection and familiarity respectively. Therefore, the R(emember) / K(now) procedure was applied in order to examine specific effects on recollective experience. Loftus (1972; 1981) suggested that active overt exploration of the scene should be important for scene encoding and hence subsequent recognition performance. To test this assumption, eye movements were eliminated during study, test or both in Experiment 1. Specifically, we reasoned that elimination of eye movements during either study or test would reduce levels of explicit recollection as measured by R responses.
Faces constitute a special visual category in that people in general demonstrate expertise effects in face processing (Diamond & Carey, 1986; Yin, 1969). Specifically, this expertise is considered to emanate from holistic (Farah, Tanaka & Drain, 1995) or configural (Searcy & Bartlett, 1996) processing. From this perspective, face processing is characterized by simultaneous processing of the relationship between facial components. Configural processing might constitute a powerful
13
mechanism in distinguishing items and should hence be related to explicit recollection. Half of the faces in Experiment 1 were presented upside down during study in order to reduce access to configural processing.
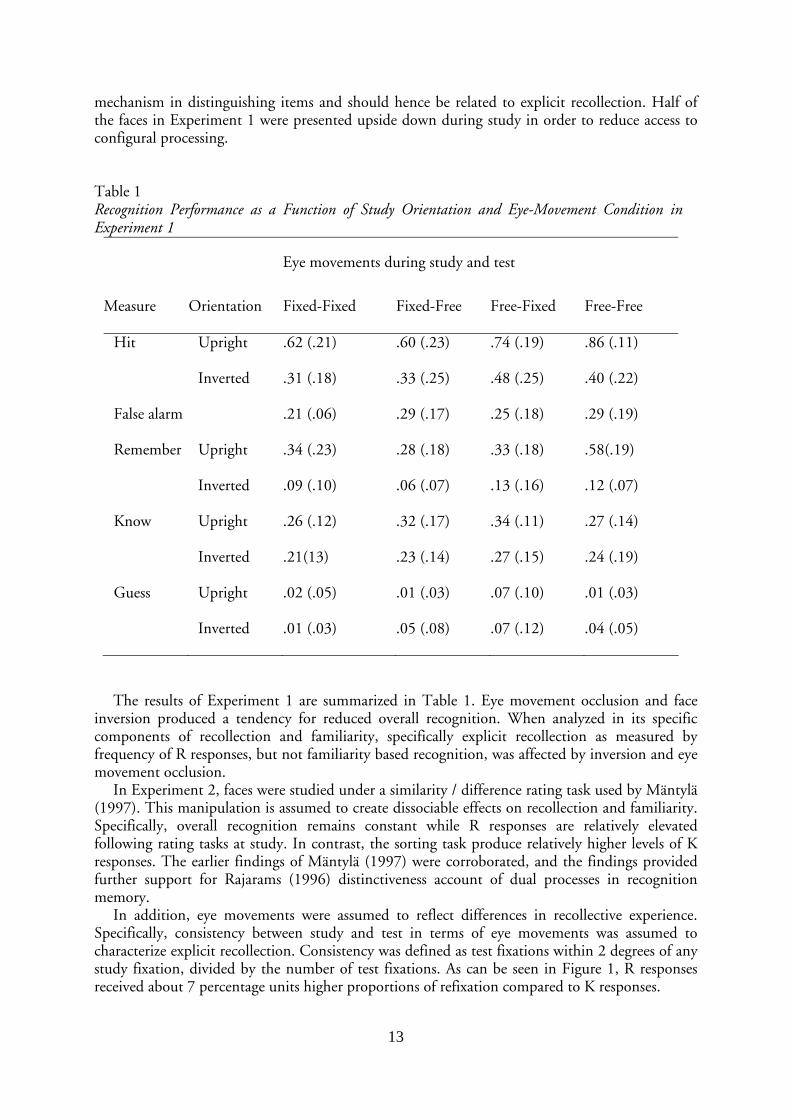
Table 1 Recognition Performance as a Function of Study Orientation and Eye-Movement Condition in Experiment 1
Eye movements during study and test
Measure Orientation Fixed-Fixed Fixed-Free Free-Fixed Free-Free
Hit Upright .62 (.21) .60 (.23) .74 (.19) .86 (.11)
Inverted .31 (.18) .33 (.25) .48 (.25) .40 (.22)
False alarm .21 (.06) .29 (.17) .25 (.18) .29 (.19)
Remember Upright .34 (.23) .28 (.18) .33 (.18) .58(.19)
Inverted .09 (.10) .06 (.07) .13 (.16) .12 (.07)
Know Upright .26 (.12) .32 (.17) .34 (.11) .27 (.14)
Inverted .21(13) .23 (.14) .27 (.15) .24 (.19)
Guess Upright .02 (.05) .01 (.03) .07 (.10) .01 (.03)
Inverted .01 (.03) .05 (.08) .07 (.12) .04 (.05)
The results of Experiment 1 are summarized in Table 1. Eye movement occlusion and face inversion produced a tendency for reduced overall recognition. When analyzed in its specific components of recollection and familiarity, specifically explicit recollection as measured by frequency of R responses, but not familiarity based recognition, was affected by inversion and eye movement occlusion.
In Experiment 2, faces were studied under a similarity / difference rating task used by Mäntylä (1997). This manipulation is assumed to create dissociable effects on recollection and familiarity. Specifically, overall recognition remains constant while R responses are relatively elevated following rating tasks at study. In contrast, the sorting task produce relatively higher levels of K responses. The earlier findings of Mäntylä (1997) were corroborated, and the findings provided further support for Rajarams (1996) distinctiveness account of dual processes in recognition memory.
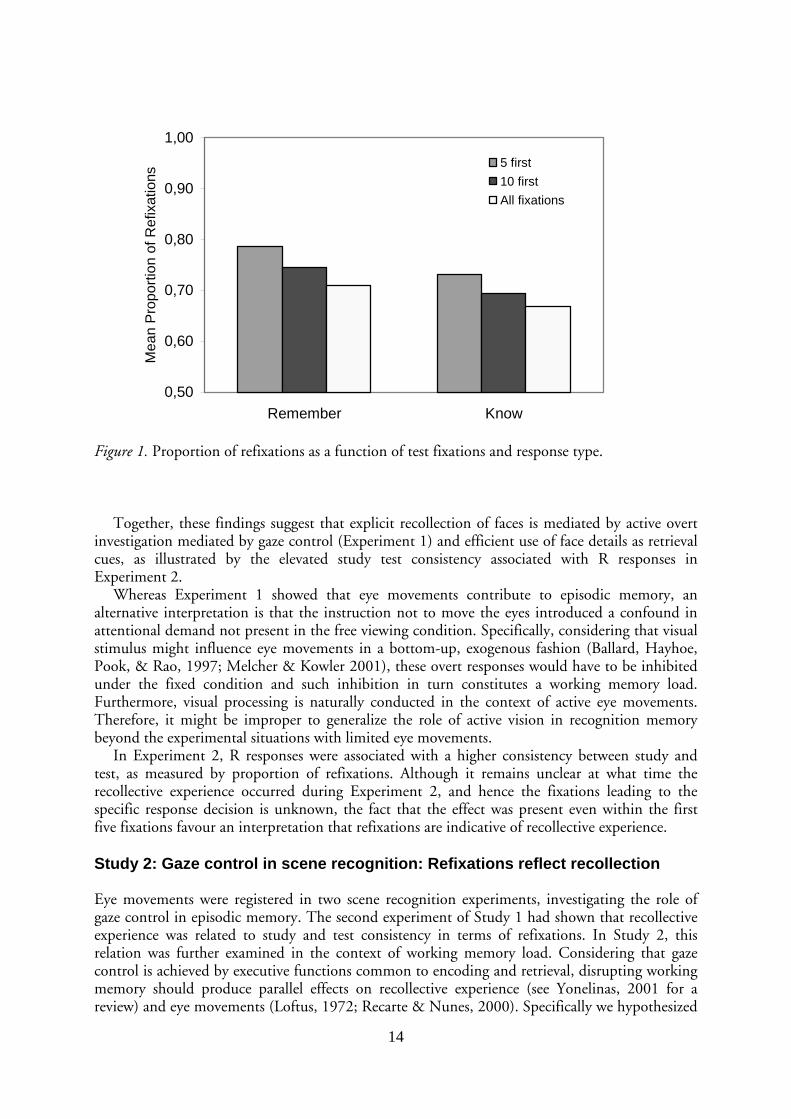
In addition, eye movements were assumed to reflect differences in recollective experience. Specifically, consistency between study and test in terms of eye movements was assumed to characterize explicit recollection. Consistency was defined as test fixations within 2 degrees of any study fixation, divided by the number of test fixations. As can be seen in Figure 1, R responses received about 7 percentage units higher proportions of refixation compared to K responses.
14
0,50
0,60
0,70
0,80
0,90
1,00
Remember Know
Mea
n P
ropo
rtion
of R
efix
atio
ns5 first10 firstAll fixations
Figure 1. Proportion of refixations as a function of test fixations and response type.
Together, these findings suggest that explicit recollection of faces is mediated by active overt investigation mediated by gaze control (Experiment 1) and efficient use of face details as retrieval cues, as illustrated by the elevated study test consistency associated with R responses in Experiment 2.
Whereas Experiment 1 showed that eye movements contribute to episodic memory, an alternative interpretation is that the instruction not to move the eyes introduced a confound in attentional demand not present in the free viewing condition. Specifically, considering that visual stimulus might influence eye movements in a bottom-up, exogenous fashion (Ballard, Hayhoe, Pook, & Rao, 1997; Melcher & Kowler 2001), these overt responses would have to be inhibited under the fixed condition and such inhibition in turn constitutes a working memory load. Furthermore, visual processing is naturally conducted in the context of active eye movements. Therefore, it might be improper to generalize the role of active vision in recognition memory beyond the experimental situations with limited eye movements.
In Experiment 2, R responses were associated with a higher consistency between study and test, as measured by proportion of refixations. Although it remains unclear at what time the recollective experience occurred during Experiment 2, and hence the fixations leading to the specific response decision is unknown, the fact that the effect was present even within the first five fixations favour an interpretation that refixations are indicative of recollective experience.
Study 2: Gaze control in scene recognition: Refixations reflect recollection Eye movements were registered in two scene recognition experiments, investigating the role of gaze control in episodic memory. The second experiment of Study 1 had shown that recollective experience was related to study and test consistency in terms of refixations. In Study 2, this relation was further examined in the context of working memory load. Considering that gaze control is achieved by executive functions common to encoding and retrieval, disrupting working memory should produce parallel effects on recollective experience (see Yonelinas, 2001 for a review) and eye movements (Loftus, 1972; Recarte & Nunes, 2000). Specifically we hypothesized
15
that tasks involving executive or visuo-spatial processing should affect both levels of explicit recollection and eye movement consistency between study and test as measured by proportion of refixations. This measure was identical to that used in Study 1.
Study Condition
Executive Visuo-Spatial Articulatory No-Load
Res
pons
e P
roba
bilit
y
0,0
0,1
0,2
0,3
0,4
0,5KnowRemember
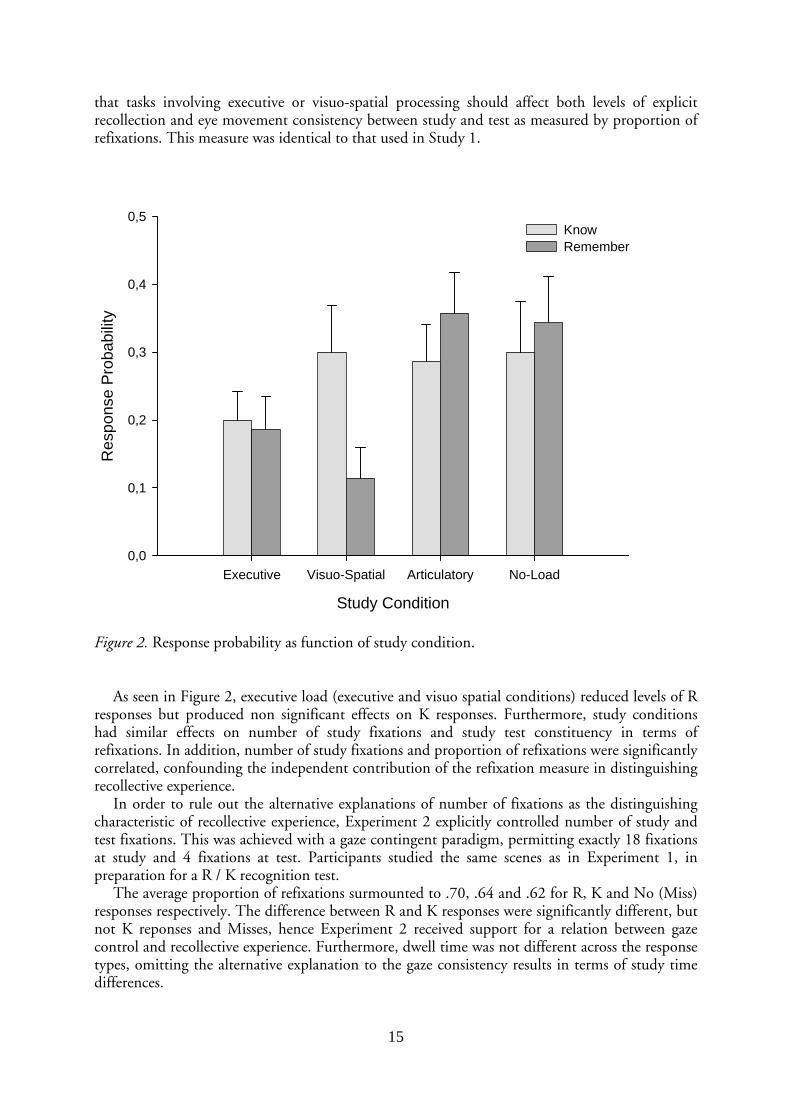
Figure 2. Response probability as function of study condition.
As seen in Figure 2, executive load (executive and visuo spatial conditions) reduced levels of R
responses but produced non significant effects on K responses. Furthermore, study conditions had similar effects on number of study fixations and study test constituency in terms of refixations. In addition, number of study fixations and proportion of refixations were significantly correlated, confounding the independent contribution of the refixation measure in distinguishing recollective experience.
In order to rule out the alternative explanations of number of fixations as the distinguishing characteristic of recollective experience, Experiment 2 explicitly controlled number of study and test fixations. This was achieved with a gaze contingent paradigm, permitting exactly 18 fixations at study and 4 fixations at test. Participants studied the same scenes as in Experiment 1, in preparation for a R / K recognition test.
The average proportion of refixations surmounted to .70, .64 and .62 for R, K and No (Miss) responses respectively. The difference between R and K responses were significantly different, but not K reponses and Misses, hence Experiment 2 received support for a relation between gaze control and recollective experience. Furthermore, dwell time was not different across the response types, omitting the alternative explanation to the gaze consistency results in terms of study time differences.

16
GENERAL DISCUSSION
This thesis departed in the question whether eye movements are related to recognition memory in general and explicit recollection in particular. The results presented above support the view that eye movements are related to recognition memory and distinguishes between recollective experience’s. For instance, the first experiment of Study 1 showed that precluding eye movements selectively reduced conscious recollection, implying that face memory is served by high resolved attribute information. Furthermore, in Experiment 2, Study 1, proportion of refixations was found to differentiate conscious recollection from familiarity based face recognition during free stimulus viewing. This finding was replicated and extended to scene recognition in both experiments of Study 2. In addition, the refixation measure successfully differentiated recollective experience following incidental (Study 1) as well as intentional (Study 2) study instructions. The first experiment of Study 2, also investigated the role of executive load on gaze control and recognition memory. Dual tasks involving executive or visuo-spatial processing produced parallel effects on eye movements and memory performance. Specifically, explicit recollection and number of fixations but not familiarity based recognition was reduced by these concurrent tasks.
Loftus (1972) showed that memory performance increased as a function of study fixations. In agreement with Loftus, number of study fixations could also differentiate recollective experience in Experiment 1 of Study 2. However, in view of Experiment 2 in Study 2, this alternative explanation seems less important. Specifically, the second experiment of Study 2 showed that when number of fixations was kept constant, proportion of refixations still differentiated between explicit recollection and familiarity. Furthermore, the effect size of this experiment was fairly similar to those found in Experiment 2 in Study 1 and those of Experiment 1 in Study 2.
Experiment 2 in Study 1 and Study 2 and both experiments of Study 2 provided support for study test consistency related to recollective experience, with effect sizes around 7 percentage units. Although this effect size might seem small, it should be considered that explicit recollection might also be achieved through other sources than high acuity perceptual reinstatement. For instance, low spatial frequency information (Schyns & Oliva, 1997) might be more diagnostic for explicit recollection in some cases, and should be less dependent on focal vision. In fact, people generally tend to fixate the centre of a picture (Maanan, Rudoch, & Wooding, 1997), irrespective of content. This should be a strategic choice given that peripheral vision provides sufficient visual detail for complex visual processing (But see also Reinagel, & Zador, 1999, for an alternative interpretation). Speculatively, the low contrast information provided by peripheral vision might be diagnostic in discrimination of difficult visual categories such as human faces, where configurations might provide the only discriminative cue. Furthermore, the data on eye movement consistency presented in this thesis show that some items were recollected in complete absence of refixations, whereas other items were not recognized despite fixation sequences entirely made up of refixations, thus refixations are neither necessary nor sufficient for recollection. However, precluding eye movements (Experiment 1 in Study 1) was detrimental to explicit recollection, supporting a necessary contribution of eye movements in explicit recollection. Alternatively, information provided under normal lightning conditions provides redundant information, reducing the demand for high study test consistency in recognition. Instead, visual processing under degraded conditions might require a higher consistency in order to support explicit recollection.
The studies presented in this thesis support a functional relation between eye movements and episodic memory, but the causal relations are yet unsolved. Specifically, to what extent does the long term representation of a scene affect gaze control? The elevated levels of refixations associated with conscious recollection might as well be explained by bottom-up processing (c.f. Melcher & Kowler, 2001), as by successive hypothesis testing based on iterative retrieval from episodic memory (c.f. Deco & Schurman, 2000) or the execution of a whole sequence of saccades reflecting a procedural memory for the scene instance (Noton & Stark, 1971).

17
There is an intrinsic problem in resolving the causal nature of gaze control in recognition memory. Manipulating eye movements (e.g. proportion of refixations) provides a strong avenue in assessing the causal role of episodic memory in gaze control but introduces artefact behaviour not present in ordinary situations (e.g. Experiment 1, Study 1). The alternative approach is more descriptive, however, certain cases might be found for which a causal relation might be inferred irrespectively. A possible solution on this account might be to investigate gaze control in recognition under visually degraded conditions. Considering that viewing in good lightning conditions provide redundant information, the cost of making errant saccades might be low, and eye guidance could hence be admitted to less processing costly bottom-up processes. Instead, under limited viewing conditions the visual information requires more extensive treatment, increasing processing time and effort invested in each fixation and hence increasing the appropriateness of investing top down resources on eye guidance.
An alternative approach would be to examine the fixation series more closely. If episodic memory intervened in gaze control during recognition, then one might expect that once a refixation was made and successfully matched with an instance representation, it would be followed by a higher degree of subsequent refixations, reflecting confirmatory eye movements. This should result in initially similar scan patterns, which eventually differs, depending on the memory outcome of the information sampling. Explicitly, one would expect all response types to produce the same proportion of refixations early during the test presentation, reflecting a general, bottom-up scanning strategy (see Henderson & Hollingworth, 1999), whereas later on, there should be an interaction of response type. Specifically, fixation series leading to misses should decrease in probability of refixations, whereas fixations series leading to explicit recollection should increase in proportion of refixations towards the end of the series.

18
REFERENCES Althoff, R. R. (1998). Eye movement-based memory assessment: the use of eye movement
monitoring as an indirect measure of memory. Dissertation Abstracts International, (UMI No. 9912183)
Althoff, R. R., & Cohen, N. J. (1999). Eye-movement based memory effect: A reprocessing effect in face perception, Journal of Experimental Psychology: Learning, Memory, and Cognition, 25, 997-1010.
Antes, J. R. (1974). The time course of picture viewing. Journal of Experimental Psychology, 103, 62-70.
Antes, J. R., Penland, J. G., & Metzger, R. L (1981). Processing global information in briefly presented pictures. Psychological Research, 43, 277-292.
Ballard, D., Hayhoe, M. M., Pook, P. K., & Rao, R. P. N. (1997). Deictic codes for the embodiment of cognition. Behavioral and Brain Sciences, 20, 723-767.
Blaxton, T. A., & Theodore, W. H. (1997). The role of the temporal lobes in recognizing visuospatial materials: Remembering versus knowing. Brain and Cognition, 35, 5-25.
Brockmole, J. R., Carlson, L. A., & Irwin, D. E. (2002). Inhibition of attended processing during saccadic eye movements. Perception and Psychophysics, 64, 867-881.
Brooks, L. R. (1968). Spatial and verbal components of the act of recall. Canadian Journal of Psychology, 22, 349-368.
Chun, M. M., & Jiang, Y. (2003). Implicit long-term spatial contextual memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 29, 224-234.
Deubel, H., & Schneider, W. X., (1996). Saccade target selection and object recognition: Evidence for a common attentional mechanism. Vision Research, 36, 1827-1837.
Deco, G., & Schurmann, B. (2000). A neuro-cognitive visual system for object recognition based on testing of interactive attentional top-down hypotheses. Perception, 29, 1249-1264.
Dewhurst, S. A., & Hitch, G. J. (1999). Cognitive effort and recollective experience in recognition memory. Memory, 7, 129-146.
Diamond, R., & Carey, S. (1986). Why faces are and are not special: An effect of expertise. Journal of Experimental Psychology: General, 115, 107-117.
Dobbins, I. G., Kroll, N. E. A., & Liu, Q. (1998). Confidence-accuracy inversions in scene recognition: A remember-know analysis, Journal of Experimental Psychology: Learning, Memory, and Cognition, 24, 1306-1315.
Donaldson, W. (1996). The role of decision processes in remembering and knowing. Memory & Cognition, 24, 523-533.
Duncan, J., & Owen, A. D. (2000). Common regions of human frontal lobe recruited by diverse cognitive demands. Trends in Neurosciences, 23, 475-482.
Eldridge, L. L, Knowlton, B. J., Furmanski, C. S., Bookheimer, S. Y., & Engel, S. A. (2000). Remembering episodes: A selective role for the hippocampus. Nature Neuroscience, 3, 1149-1152.
Farah, M. J., Tanaka, J. W., & Drain, H. M. (1995). What causes the face inversion effect?, Journal of Experimental Psychology: Human Perception and Performance, 21, 628-634.
Fisher, D. F., Karsh, R., Breitenbach, F., Barnette, B. D. (1983). Eye movements and picture recognition: Contribution or embellishment, In R. Groner., C. Menz., D.F. Fisher & R. A. Monty (Eds.), Eye movements and psychological functions: International views (pp. 193-210). Hillsdale, NJ: Erlbaum.
Friedman, A. (1979). Framing pictures: The role of knowledge in automatized encoding and memory for gist. Journal of Experimental Psychology: General, 108, 316-355.
Fuster, J. M. (2000). Prefrontal neurons in networks of executive memory. Brain Research Bulletin, 56, 331-336.

19
Gardiner, J. M. (1988). Functional aspects of recollective experience. Memory & Cognition, 16, 309-313.
Gardiner, J. M., & Java, R. I. (1990). Recollective experience in word and nonword recognition, Memory & Cognition, 18, 23-30.
Gardiner, J. M., & Java, R. I. (1993). Recognizing and remembering. In A. F. Collins, M. A Conway, & P. E. Morris (Eds.), Theories of memory (pp. 163-188). Hillsdale, NJ: Erlbaum.
Gardiner, J. M., & Parkin, A. J. (1990). Attention and recollective experience in recognition memory. Memory & Cognition, 18, 579-583.
Glenberg, A. M., Schroeder, J. L., & Robertson, D. A. (1998). Averting the gaze disengages the environment and facilitates remembering. Memory & Cognition, 26, 651-658.
Hashtroudi, S., Johnson, M. K., & Chrosniak, L. D. (1989). Aging and source monitoring. Psychology and Aging, 4, 106-112.
Hebb, D. O. (1949). The organization of behavior. New York, Wiley. Henderson, J. (2003). Human gaze control during real-world scene perception. Trends in
Cognitive Sciences, 7, 498-504. Henderson, J. M., & Hollingworth. (1999). High-level scene perception. Annual Review of
Psychology, 50, 243-271. Henderson, J. M., McClure, K. K., Pierce, S., & Schrock, G. (1997). Object identification
without foveal vision: Evidence from an artificial scotoma paradigm, Perception and Psychophysics, 59, 323-346.
Henderson, J. M., Weeks, P. A., & Hollingworth, A. (1999). The effects of semantic consistency on eye movements during complex scene viewing. Journal of Experimental Psychology: Human Perception and Performance, 25, 210-228.
Henson, R. N. A., Rugg, M. D., Shallice, T., Josephs, O., & Dolan, R. J. (1999). Recollection and familiarity in recognition memory: An event-related functional magnetic resonance imaging study, Journal of Neuroscience, 19, 3962-3972.
Hicks, J. L., & Marsh, R. L. (2000). Toward specifying the attentional demands for recognition memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 26, 1483-1498.
Hollingworth, A. (2004). Constructing visual representations of natural scenes: the roles of short- and long term visual memory. Journal of Experimental Psychology: Human Perception and Performance, 30, 519-537.
Hollingworth, A., & Henderson, J. M. (2002). Accurate visual memory for previously attended
objects in natural scenes. Journal of Experimental Psychology: Human Perception and Performance, 28, 113-136.
Hollingworth, A., Williams, C. C., & Henderson, J. M. (2001). To see and remember: Visually specific information is retained in memory from previously attended objects in natural scenes. Psychonomic Bulletin and Review, 8, 761-768.
Hunt, R. R., & Einstein, G. O (1981). Relational and item-specific information in memory. Journal of Verbal Learning and Verbal Behaviour, 19, 497-514.
Hunt, R. R., & McDaniel, M. A. (1993). The enigma of organization and distinctiveness, Journal of Memory and Language, 32, 421-445.
Hunt, R. R., & Seta, C. E. (1984). Category size effects in recall: The roles of individual item and relational information, Journal of Experimental Psychology; Learning, Memory and Cognition, 10, 282-290.
Intraub, H. (1981). Identification and processing of briefly glimpsed visual scenes. In D. F. Fisher, R. A. Mony & J. W. Senders (Eds), Eye movements: Cognition and visual perception (pp 181-190), Erlbaum: Hillsdale New Jersey.

20
Irwin, D. E., & Zelinsky, D. J. (2002). Eye movements and scene perception: Memory for things observed. Perception & Psychophysics, 64, 882-895.
Jacoby, L. L. (1991). A process dissociation framework: Separating automatic from intentional uses of memory. Journal of Memory and Language, 30, 513-541.
Jacoby, L. L., & Dallas, M. (1981). On the relationship between autobiographical memory and perceptual learning. Journal of Experimental Psychology: General, 110, 306-340.
Janowsky, J. S., Shimamura, A. P., & Squire, L. R. (1989). Source memory impairment in patients with frontal lobe lesions. Neuropsychologia, 27, 8, 1043-1056.
Johnson, M. K., Hashtroudi, S., & Lindsay, D. S. (1993). Source monitoring. Psychological Bulletin, 114, 3-28.
Knowlton B. J., & Squire, L. R. (1995). Remembering and Knowing: Two different expressions of declarative memory, Journal of Experimental Psychology: Learning, Memory and Cognition, 3, 699-710.
Kowler, E., Anderson, E., Dosher, B., Blaser, E. (1995). The role of attention in the programming of saccades. Vision Research, 35, 1897-1916.
Konen, C. S., Kleiser, R., Wittsack, H. J., Bremmer, F., & Seitz, R. J. (2004). The encoding of saccadic eye movements within human posterior parietal cortex. Neuroimage, 22, 304-314.
Laeng, B., & Teodorescu, D-S. (2002). Eye scanpaths during visual imagery re-enact those of perception of the same visual scene. Cognitive Science, 26, 207-231.
Levine, B., Black, S. E., Cabeza, R., Sinden, M., McIntosh, A. R., Toth, J. P., Tulving, E., & Stuss, D. T. (1998). Episodic memory and the self in a case of isolated retrograde amnesia. Brain, 120, 1951-1973.
Levine, B., Freedman, M., Dawson, D., Black, S., & Stuss, D. T. (1999). Ventral frontal contribution to self-regulation; Convergence of episodic memory and inhibition. Neurocase, 5, 263-275.
Locher, P. J., & Nodine, C. F. (1974). The role of scanpaths in the recognition of random shapes. Perception & Psychophysics, 15, 308-314.
Loftus, G. R. (1972). Eye fixations and recognition memory for pictures. Cognitive Psychology, 3, 525-551.
Loftus, G. R. (1981). Tachistoscopic simulations of eye fixations on pictures. Journal of Experimental Psychology: Human Learning and Memory, 5, 369-376.
Luo, C. R. (1993). Enhanced feeling of recognition: Effects of identifying and manipulating test items on recognition memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 19, 405-413.
Maanan, S.K., Rudoch. K. H., & Wooding, D. S. (1997). Fixation patterns made during brief examination of two-dimensional images. Perception, 26, 1059-1072.
Mandler, G. (1980). Recognizing: The judgement of previous occurrence. Psychological Review, 87, 252-271.
McIntyre, J. S., & Craik, F. I. M. (1987). Age differences in memory for item and source information. Canadian Journal of Psychology, 41, 175-192.
Melcher, D., & Kowler, E. (2001). Visual scene memory and the guidance of saccadic eye movements. Vision Research, 41, 3597-3611.
Miyake, A., Friedman, N. P., Rettinger, D. A., Shah, P., Hegarty, M. (2001). How are working memory, executive functioning, and spatial abilities related? A latent variable analysis. Journal of Experimental Psychology: General, 130, 621-640.
Moores, E., Laiti, L., & Chelazz, L. (2003). Associative knowledge controls deployment of visual attention. Nature Neuroscience, 6, 182-186.
Mäntylä, T., & Raudsepp, J. (1996). Recollective experience following suppression of focal attention. European Journal of Cognitive Psychology, 8, 195-203.

21
Mäntylä, T. (1997). Recollection of faces: Remembering differences and knowing similarities. Journal of Experimental Psychology: Learning, Memory and Cognition, 23, 1203-1216.
Noton, D., & Stark, L. W. (1971a). Scanpaths in saccadic eye movements while viewing and recognizing patterns. Vision Research, 11, 929-942.
Noton, D., & Stark, L. W. (1971b). Scanpaths in eye movements during perception. Science, 171, 308-311.
Nelson, W. W., & Loftus, G. R. (1980). The functional visual field during picture viewing. . Journal of Experimental Psychology: Human Learning and Memory, 6, 391-399.
Parker, R. E. (1978). Picture processing during recognition. Journal of Experimental Psychology: General,4, 284-292.
Parkin, A. J., Gardiner, J. M., & Rosser, R. (1995). Functional aspects of recollective experience in face recognition. Consciousness and Cognition, 4, 387-398.
Pierrot-Deseilligny, C., Muri, R. M., Ploner, C. J., Gaymard, B., & Rivaud-Pechoux, S. (2003). Cortical control of ocular saccades in humans: A model for motricity. Progress in Brain Research, 142, 3-17.
Pierrot-Deseilligny, C., Milea, D., & Müri, R. M. (2004). Eye movement control by the cerebral cortex. Current Opinion in Neurology, 17, 17-25.
Rajaram, S. (1993). Remembering and knowing: Two means of access to the personal past. Memory& Cognition, 21, 89-102.
Rajaram, S. (1996). Perceptual effects on remembering: Recollective processes in picture recognition memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 22, 365-377.
Rajaram, S., & Geraci, L. (2000). Conceptual fluency selectively influences knowing. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 1070-1074.
Rajaram, S. & Roediger H. L. III (1996). Remembering and knowing as states of consciousness during retrieval. In J. D. Cohen & J. W. Schooler (Eds.), Scientific approaches to the question of consciousness. Hillsdale, NJ: Lawrence Erlbaum.
Recarte, M. A., & Nunes, L. M. (2000). Effects of verbal and spatial-imagery tasks on eye fixations while driving. Journal of Experimental Psychology: Applied, 6, 31-43.
Reinagel, P., & Zador, A. M. (1999). Natural scene statistics at the centre of gaze. Network: Computational Neural Systems, 10, 341-350.
Reinitz, M. T., Morrisey, J., & Demb, J. (1994). Role of attention in face encoding. Journal of Experimental Psychology: Learning, Memory, and Cognition, 20, 161-168.
Robbins, T. W., Anderson, E. J., Barker, D. R., Bradley, A. C., Fearnyhough, C., Henson, R., & Hudson, S. R. (1996). Working memory in chess. Memory & Cognition, 24, 83-93.
Roediger, H. L., Weldon, M. S., & Challis, B. H. (1989). Explaining dissociations between implicit and explicit measures of retention: A processing account. In H. L. Roediger III & F. I. M. Craik (Eds), Varities of memory and consciousness: Essays in honour of Endel Tulving. (pp. 3-41). Hillsdale, NJ: Lawrence Erlbaum Associates.
Rousselet, G. A., Thorpe, S. J., Fabre-Thorpe, M. (2004). How parallel is visual processing in the ventral pathway? Trends in Cognitive Sciences, 8, 363-370.
Rybak, I. A., Gusakova, V. I., Golovan, A. V., Podladchikova, L. N., & Shevtsova, N. A. (1998). A model of attention-guided visual perception and recognition. Vision Research, 38, 2387-2400.
Searcy, J. H., & Bartlett, J. C. (1996). Inversion and processing of components and spatial-relational information in faces, Journal of Experimental Psychology: Human Perception and Performance, 22, 904-915.
Schyns, P. G., & Oliva, A. (1997). Flexible, diagnosticity-driven, rather than fixed, perceptually determined scale selection in scene and face recognition. Perception, 26, 1027-1038.

22
Shimamura, A. P., & Squire, L. R. (1987). A neuropsychological study of fact memory and source amnesia. Journal of Experimental Psychology: Learning, Memory, and Cognition, 13, 464-473.
Shimamura, A. P., & Squire, L. R. (1991). The relationship between fact and source memory: Findings from amnesic patients and normal subjects. Psychobiology, 19, 1-10. Spencer, W. D., & Raz, N. (1994). Memory for facts, source, and context: Can frontal lobe
dysfunction explain age-related differences? Psychology and Aging, 9, 149-159. Thaiss, L., & Petrides, M. (2003). Source versus content memory in patients with a unilateral
frontal cortex or temporal lobe excision. Brain, 126, 1112-1126. Tong, F., & Nakayama, K. (1999). Robust representations for faces: Evidence from visual
search, Journal of Experimental Psychology: Human Perception and Performance, 25, 1016-1035.
Tulving, E. (1983). Elements of episodic memory. New York: Oxford University Press. Tulving, E. (1985). Memory and consciousness. Canadian Journal of Psychology, 26, 1-12. Wagner, A. D., Gabrieli, J. D. E., & Verfaellie, M. (1997). Dissociations between familiarity
processes in explicit recognition and implicit perceptual memory. Journal of Experimental Psychology: Learning, Memory and Cognition, 23, 305-323.
Walker-Smith, G. J., Gale, A. G., & Findlay, J. M. (1977). Eye movement strategies involved in face perception. Perception, 6, 313-326.
Wandell, B.A.(1995). Foundations of vision. Sunderland, MA: Sinauer Associates. Van Diepen, P. M. J., & d’Ydewalle, G. (2003). Early peripheral and foveal processing in
fixations during scene perception. Visual Cognition, 10, 79-100. Wolfe, J. M. (2003). Moving towards solutions on some long enduring controversies on visual
search. Trends in Cognitive Sciences, 7, 70-76. Yarbus, A. L. (1967). Eye movements and vision, New York: Plenum Yin, R. K. (1969). Looking at upside-down faces, Journal of Experimental Psychology, 81, 141-
145. Yonelinas, A. P. (2001). Consciousness, control and confidence: The 3 c’s of recognition
memory. Journal of Experimental Psychology: General, 130, 361-379. Yonelinas, A. P. (2002). The nature of recollection and familiarity: A review of 30 years of
research. Journal of Memory and Language, 46, 441-517.


Umeå University S-901 87 Umeå.Phone +46 90 786 50 00, Fax +46 90 786 99 95
ISSN 1651-565X
Department of Psychology
Umeå University
S-901 87 Umeå
Sweden
Phone +46 (0)90-786 50 00
Fax +46 (0)90-786 66 92
www.psy.umu.se





























