Embed Size (px)
Citation preview

Journal of Structural Biology 164 (2008) 33–40
Contents lists available at ScienceDirect
Journal of Structural Biology
journal homepage: www.elsevier .com/locate /y jsbi
Immunolocalization of matrix proteins in nacre lamellae and their in vivo effectson aragonitic tablet growth
Ningping Gong a, Junlong Shangguan a, Xiaojun Liu a, Zhenguang Yan a, Zhuojun Ma a, Liping Xie a,b,*,Rongqing Zhang a,b,*
a Institute of Marine Biotechnology, Department of Biological Sciences and Biotechnology, Tsinghua University, Beijing 100084, Chinab Protein Science Laboratory of the Ministry of Education, Tsinghua University, Beijing 100084, China
a r t i c l e i n f o
Article history:Received 9 January 2008Received in revised form 23 May 2008Accepted 24 May 2008Available online 3 June 2008
Keywords:Mollusk shell nacreMatrix proteinBMP-2NacreinAntibodies
1047-8477/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.jsb.2008.05.009
* Corresponding authors. Fax: +86 10 62772899.E-mail addresses: [email protected] (L. Xi
du.cn (R. Zhang).
a b s t r a c t
How matrix proteins precisely control the growth of nacre lamellae is an open question in biominerali-zation research. Using the antibodies against matrix proteins for immunolabeling and in vivo experi-ments, we investigate the structural and functional roles of EDTA–soluble matrix (SM) and EDTA–insoluble matrix (ISM) proteins in nacre biomineralization of the pearl oyster Pinctada fucata. Immunola-beling reveals that a SM protein, nacrein, distributes within aragonitic tablets and intertabular matrix. AnISM protein, which we named P43, has been specifically recognized by polyclonal antibodies raisedagainst the recombinant protein of P. fucata bone morphogenetic protein 2 in immunoblot analysis.Immunolabeling indicates that P43 is localized to interlamellar sheet, and also embedded within arago-nitic tablets. Although nacrein and P43 both distribute within aragonitic tablets, they function differentlyin aragonitic tablet growth. When nacrein is suppressed by the antibodies against it in vivo, crystal over-growth occurs, indicating that this SM protein is a negative regulator in aragonitic tablet growth. WhenP43 is suppressed in vivo, the organo-mineral assemblage is disrupted, suggesting that P43 is a frame-work matrix. Taken together, SM and ISM proteins are indispensable factors for the growth of nacrelamellae, controlling crystal growth and constructing the framework of aragonitic tablets.
� 2008 Elsevier Inc. All rights reserved.
1. Introduction
Nacre (mother-of-pearl) is a biogenetic material widely distrib-uting in mollusks, and usually forms in the inner layer of shells.The structure of nacre from bivalves is characterized as an arrange-ment of continuous parallel lamellae (Fig. 1A), separated by sheetsof interlamellar matrix (Wada, 1968; Addadi et al., 2006). Each la-mella is composed of polygonal aragonitic tablets (Fig. 1B), whichis sealed to each other by intertabular matrix. An aragonitic tabletseems to be a single crystal, but actually is a coherent aggregationof crystalline nanograins (about 45 nm mean size) with the samecrystallographic orientation, connected by a continuous organicframework (Rousseau et al., 2005a; Stolarski and Mazur, 2005).
Nacre is an organo-mineral assemblage with dominant calciumcarbonate and a minor organic matrix complex, resulting in supe-rior mechanical properties and excellent osteoinductive activity(Silve et al., 1992; Lamghari et al., 1999; Weiner et al., 2003). Theorganic matrix complex includes proteins and polysaccharides,which is thought to direct the growth of calcium carbonate crystal
ll rights reserved.
and be responsible for the extraordinary properties of nacre (Wei-ner and Addadi, 1997; Mouriès et al., 2002; Zhang and Zhang,2006). Matrix proteins in nacre are usually classified into twogroups depending on their solubility in decalcifying solution (Cren-shaw and Ristedt, 1976). EDTA-based solution is often used fordecalcification. EDTA–soluble matrix (SM) proteins have been pro-posed to localize within calcium carbonate crystals as intracrystal-line matrix (Crenshaw, 1972) and control crystal morphology,crystal phase switching and orientation (Mann et al., 1993; Belcheret al., 1996; Zhang and Zhang, 2006). In contrast, EDTA–insolublematrix proteins (ISMs) are hypothesized to localize around calciumcarbonate crystallites and thus are supposed as intercrystallinematrices and framework macromolecules (Crenshaw, 1972; Sudoet al., 1997). Matrix proteins play vital roles in the formation ofnacre. But how matrix proteins interact with the mineral in theself-organization of nacre has not been fully understood yet.
Immunolocalization has been widely used to study the func-tions of matrix proteins in the growth of biomaterials, such assea urchin, coral skeletons and shell (Crenshaw and Ristedt,1976; Cho et al., 1996; Puverel et al., 2005; Nudelman et al.,2006; Marin et al., 2007). Antibody inhibition assay is proved tobe a reliable method for in vivo investigation of the functions ofmatrix proteins in the growth of nacre lamellae (Ma et al., 2007).

Fig. 1. Nacre structure of P. fucata. (A) SEM view shows the cross-section of nacre, indicating that nacre is composed of continuous parallel lamellae; bar, 2 lm. (B) Each nacrelamellae is composed of polygonal aragonitic tablets, which is sealed each other by intertabular matrix (arrow indicating); bar, 20 lm.
34 N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40
When injected into the extrapallial fluid between shell and mantletissue, where nacre calcification occurs, the antibodies against aspecial matrix protein could interrupt the action of this proteinin nacre calcification, resulting in aberrant growth of innermost la-mella. The functions of this protein in nacre calcification will beindicated from the aberrant growth pattern.
Nacrein is a major SM protein in the nacre of the pearl oysterPinctada fucata, and has carbonic anhydrase (CA) activity (Miyam-oto et al., 1996). CA activity is essential for rapid shell development(Wilbur and Jodrey, 1955; Freeman, 1960). Moreover, nacrein-likeproteins are conserved in bivalves and gastropods, and thus arethought to play an important role in nacre mineralization of thetwo species (Miyamoto et al., 2005). In this study, we conductimmunolocalization and antibody inhibition assays to examineits function in nacre mineralization. We also characterize an ISMprotein by the antibodies raised against the recombinant proteinof P. fucata bone morphogenetic protein 2 (Pf-BMP-2). A betterunderstanding of the structural and functional roles of SM andISM proteins in nacre biomineralization will provide essentialknowledge to mimic nacre.
2. Materials and methods
2.1. Animals
The pearl oyster Pinctada fucata with shells of about 4.5–6.0 cmin diameter and 32–40 g in wet weight were obtained from Beihai,China and maintained in aerated artificial seawater at 3% salinity inan aquarium.
2.2. Extraction and isolation of shell matrix proteins
The shell nacre from Pinctada fucata was ground and dissolvedin 0.5 M EDTA (pH 8.0) for 4 days at 4 �C. After centrifugation,the supernatant containing SM proteins was collected for immuno-blot analysis. The pellet containing ISM proteins was resuspendedin 20 mM Tris–HCl (pH 7.0) containing 1% SDS, 10 mM DTT andboiled for 20 min. After centrifugation, the supernatant was precip-itated in 85% ethanol at 4 �C. The precipitated proteins werewashed twice with 85% ethanol, and redissolved in 20 mM Tris–HCl (pH 7.0) containing 6 M urea. The resulting solution was ab-sorbed in a heparin agarose affinity column (Amersham Biosci-ences), washed thoroughly, and eluted with 20 mM Tris–HCl (pH7.0) containing 0.1 M or 0.2 M NaCl.
2.3. Preparation of antibodies
Rabbit polyclonal antibodies (P-anti-NCA) and a mouse mono-clonal antibody (M-anti-NCA) against CA domain of nacrein
(NCA) (GenBank Accession No: BAA11940) were developed inNew Zealand rabbits and BALB/c mice, respectively (Gong et al.,2008). The DNA encoding TGF-b domain (amino acids 346–447)of Pf-BMP-2 (GenBank Accession No. AB176952) was cloned intopET-28b expression vector (Novagen) and expressed in bacterialstrain of Escherichia coli BL21 (DE3). The recombinant proteinwas highly purified using a Hitrap chelating HP affinity column(Amersham Biosciences). Polyclonal antibodies against the recom-binant BMP-2 (anti-P43) or total SM proteins (anti-SM) were raisedin New Zealand rabbits using a protocol as described by Mayer andWalker (1987). The antisera were then purified by 33% saturationammonium sulfate precipitation and a protein A agarose column(Calbiochem). The purified antibodies were used for immunoblotanalysis, immunolabeling and antibody inhibition assay.
2.4. Immunoblot analysis
The proteins were electrophoretically transferred to nitrocellu-lose membranes using a Hoefer semidry blotter (Pharmacia Bio-tech). The nitrocellulose membranes were blocked with 5%skimmed milk and incubated with 5 lg/ml primary antibody for2 h. After washing and incubated with an appropriate second anti-body conjugated alkaline phosphatase (Santa Cruz), detection wascarried out using BCIP/NBT solution (Roche).
2.5. Shell decalcification
The complete decalcification of nacre lamellae was performedas described (Nudelman et al., 2006). Briefly, the nacreous layerswere mechanically isolated from the shell of P. fucata and decalci-fied at room temperature for 2 days in a solution containing 1 MEDTA (pH 8.0), 4% formaldehyde, and 0.5% cetylpyridinium chlo-ride. The decalcified fragments were thoroughly washed withMilli-Q water. The lamellar sheets were peeled off from the frag-ments, mounted on glass slides, air dried and ready forimmunolabeling.
Partial decalcification of nacre lamellae was performed as fol-lows. The fresh nacre was mechanically fractured into small frag-ments and cleaned in diluted sodium hypochlorite solution for2 min (0.2 wt% active chlorine; Marin et al., 2007). The fragmentswere then partially etched by 10 mM acetic acid (pH 3.5) for 1–3 min and neutralized with 10 mM sodium bicarbonate (Choet al., 1996). The partially etched nacre lamellae were thoroughlywashed with Milli-Q water and were ready for immunolabeling.
2.6. Immunolabeling of nacre lamellae
Completely decalcified lamellar sheets, as well as partiallyetched nacre lamellae, were blocked in 10% goat serum for 2 h,
N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40 35
and incubated with a primary antibody at concentration of 5 lg/ml or preimmune serum for 2 h. The samples were then incu-bated with an appropriate secondary antibody. Individual pri-mary antibodies used were rabbit polyclonal antibodies thatrecognized SM proteins, nacrein or P43 (anti-SM, P-anti-NCA oranti-P43) or a mouse monoclonal antibody that recognized nac-rein (M-anti-NCA). The secondary antibodies were goat anti-rab-bit/mouse coupled to rhodamine (Santa Cruz) and goat anti-rabbit coupled to 12 nm gold particles (Jackson). In each stepof immunolabeling, the samples were washed three times withPBS containing 0.05% Tween 20 for 5 min. At last, samples werewashed in water. The rhodamine labeled samples were exam-ined by a fluorescence microscope (Leica DIMR). The immuno-gold labeled samples were dried overnight, carbon coated andexamined using a scanning electron microscopy (SEM) (HitachiS-5500 UHR FE-SEM).
2.7. Antibody inhibition assay
The purified antibodies were injected into the center of theextrapallial space between the mantle and the shell through themantle with a microsyringe at the dosage of 0.5 lg (low dosage)and 1 lg (high dosage) per gram of wet weight per day. Each groupis consist of four specimens (Ma et al., 2007). The oysters were sac-rificed 3 days after antibody injection. The shells were separated,washed with Milli-Q water, and immersed in 5% NaOH for 8 h toremove organic components attached to the surfaces. The shellswere then thoroughly washed with Milli-Q water, air dried, goldcoated, and observed under a SEM (FEI Sirion2000).
3. Results
3.1. Immunoblot analysis of EDTA–soluble matrix (SM) proteins
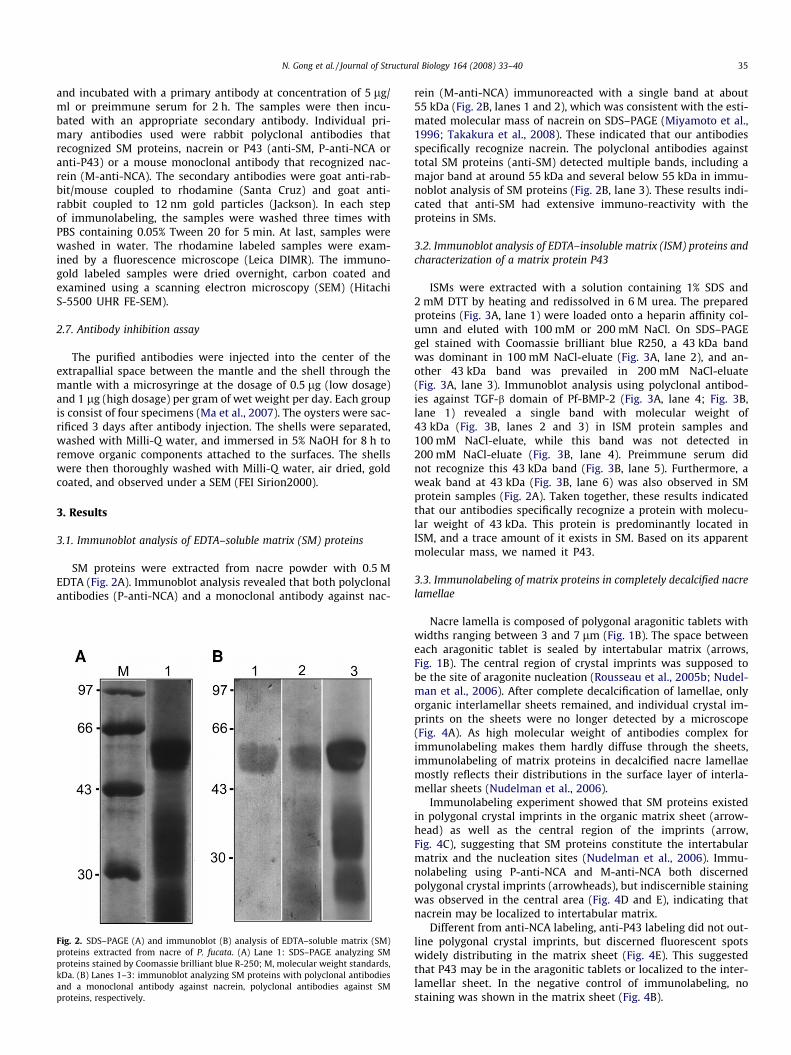
SM proteins were extracted from nacre powder with 0.5 MEDTA (Fig. 2A). Immunoblot analysis revealed that both polyclonalantibodies (P-anti-NCA) and a monoclonal antibody against nac-
Fig. 2. SDS–PAGE (A) and immunoblot (B) analysis of EDTA–soluble matrix (SM)proteins extracted from nacre of P. fucata. (A) Lane 1: SDS–PAGE analyzing SMproteins stained by Coomassie brilliant blue R-250; M, molecular weight standards,kDa. (B) Lanes 1–3: immunoblot analyzing SM proteins with polyclonal antibodiesand a monoclonal antibody against nacrein, polyclonal antibodies against SMproteins, respectively.
rein (M-anti-NCA) immunoreacted with a single band at about55 kDa (Fig. 2B, lanes 1 and 2), which was consistent with the esti-mated molecular mass of nacrein on SDS–PAGE (Miyamoto et al.,1996; Takakura et al., 2008). These indicated that our antibodiesspecifically recognize nacrein. The polyclonal antibodies againsttotal SM proteins (anti-SM) detected multiple bands, including amajor band at around 55 kDa and several below 55 kDa in immu-noblot analysis of SM proteins (Fig. 2B, lane 3). These results indi-cated that anti-SM had extensive immuno-reactivity with theproteins in SMs.
3.2. Immunoblot analysis of EDTA–insoluble matrix (ISM) proteins andcharacterization of a matrix protein P43
ISMs were extracted with a solution containing 1% SDS and2 mM DTT by heating and redissolved in 6 M urea. The preparedproteins (Fig. 3A, lane 1) were loaded onto a heparin affinity col-umn and eluted with 100 mM or 200 mM NaCl. On SDS–PAGEgel stained with Coomassie brilliant blue R250, a 43 kDa bandwas dominant in 100 mM NaCl-eluate (Fig. 3A, lane 2), and an-other 43 kDa band was prevailed in 200 mM NaCl-eluate(Fig. 3A, lane 3). Immunoblot analysis using polyclonal antibod-ies against TGF-b domain of Pf-BMP-2 (Fig. 3A, lane 4; Fig. 3B,lane 1) revealed a single band with molecular weight of43 kDa (Fig. 3B, lanes 2 and 3) in ISM protein samples and100 mM NaCl-eluate, while this band was not detected in200 mM NaCl-eluate (Fig. 3B, lane 4). Preimmune serum didnot recognize this 43 kDa band (Fig. 3B, lane 5). Furthermore, aweak band at 43 kDa (Fig. 3B, lane 6) was also observed in SMprotein samples (Fig. 2A). Taken together, these results indicatedthat our antibodies specifically recognize a protein with molecu-lar weight of 43 kDa. This protein is predominantly located inISM, and a trace amount of it exists in SM. Based on its apparentmolecular mass, we named it P43.
3.3. Immunolabeling of matrix proteins in completely decalcified nacrelamellae
Nacre lamella is composed of polygonal aragonitic tablets withwidths ranging between 3 and 7 lm (Fig. 1B). The space betweeneach aragonitic tablet is sealed by intertabular matrix (arrows,Fig. 1B). The central region of crystal imprints was supposed tobe the site of aragonite nucleation (Rousseau et al., 2005b; Nudel-man et al., 2006). After complete decalcification of lamellae, onlyorganic interlamellar sheets remained, and individual crystal im-prints on the sheets were no longer detected by a microscope(Fig. 4A). As high molecular weight of antibodies complex forimmunolabeling makes them hardly diffuse through the sheets,immunolabeling of matrix proteins in decalcified nacre lamellaemostly reflects their distributions in the surface layer of interla-mellar sheets (Nudelman et al., 2006).
Immunolabeling experiment showed that SM proteins existedin polygonal crystal imprints in the organic matrix sheet (arrow-head) as well as the central region of the imprints (arrow,Fig. 4C), suggesting that SM proteins constitute the intertabularmatrix and the nucleation sites (Nudelman et al., 2006). Immu-nolabeling using P-anti-NCA and M-anti-NCA both discernedpolygonal crystal imprints (arrowheads), but indiscernible stainingwas observed in the central area (Fig. 4D and E), indicating thatnacrein may be localized to intertabular matrix.
Different from anti-NCA labeling, anti-P43 labeling did not out-line polygonal crystal imprints, but discerned fluorescent spotswidely distributing in the matrix sheet (Fig. 4E). This suggestedthat P43 may be in the aragonitic tablets or localized to the inter-lamellar sheet. In the negative control of immunolabeling, nostaining was shown in the matrix sheet (Fig. 4B).

Fig. 3. SDS–PAGE (A) and immunoblot (B) analysis of EDTA–insoluble matrix (ISM) proteins extracted from nacre of P. fucata. (A): Lane 1, SDS–extracted ISM proteinsredissolved in 6 M urea; lane 2, 100 mM NaCl-eluate (E1) from a heparin affinity column; lane 3, 200 mM NaCl-eluate (E2) from the column; lane 4, the purified recombinantprotein encoding the TGF-b domain of Pf-BMP-2, indicated by arrow; M, molecular weight standards, kDa. (B): Lanes 1–4 and 6, immunoblot analyzing the recombinantprotein of Pf-BMP-2 (about 14 kDa), ISM proteins, E1, E2, and SM proteins (Fig. 2A, lane 1) with anti-P43 as primary antibody, respectively; lane 5, the negative control,analyzing ISM proteins with preimmune serum; arrows indicating the visualized bands.
Fig. 4. (A) Light micrograph of completely decalcified interlamellar sheets, in which only organic matrix remained; the lines in the picture are due to folds in the interlamellarsheet. (B–F) fluorescence micrographs of decalcified interlamellar sheets labeled by preimmune serum, anti-SM, P-anti-NCA, M-anti-NCA and anti-P43, respectively.Arrowheads indicate the stained intertabular matrix. Arrow in (C) indicates the stained area in the center of the crystal imprints. Bar, 20 lm.
36 N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40
3.4. Immunogold labeling of matrix proteins in partially etched nacrelamellae
Immunogold labeling partially etched nacre lamellae will revealthe distributions of matrix proteins in aragonitic tablets, which isobserved by a SEM in back-scattered electron mode (SEM-BSE).In this model, elements with higher atomic number scatter moreelectrons thus appearing lighter. Therefore, immunogold particlesare displayed as the brightest tiny spots; the mineral phase is light-er than the organic phase.
Fig. 5C and D (arrow indicating) revealed that nacrein actuallydistributes in intertabular matrix. Figs. 5C and D and 6C and D(arrowhead indicating) showed that this protein also widely existsin the aragonitic tablets and seems to attach to the mineral phase.As an ISM, P43 is rarely localized to the intertabular matrix (Fig. 5Eand F), but exists in aragonitic tablets (arrowhead indicating, Figs.5E and F and 6E and F). The negative control (Figs. 5A and B and 6Aand B) with preimmune serum suggested that the positive stainingis reliable.
Fig. 7 showed the distributions of nacrein and P43 in the cross-section of nacre lamellae (section parallel to the c-axis). The immu-nogold labeling confirmed that nacrein widely distributes in nacrelamellae and is embedded in the mineral phase as calcium carbon-
ate crystals growing (Fig. 7C and D). P43 distributes within thelamellae (Fig. 7E and F, black arrows), and also is localized to inter-lamellar matrix at interface with the mineral phase (Fig. 7E and F,white arrows). As expected, the negative controls displayed a lowbackground, which make the labeling reliable (Fig. 7A and B).
3.5. In vivo investigation of the actions of nacrein and P43 in thegrowth of aragonitic tablets
There is a space between nacre and mantle tissue, filled withbody fluid. Matrix proteins secreted by mantle tissue are releasedinto this space and act in the growth of nacre lamellae. The purifiedantibodies against P43 and nacrein were injected into this space todisrupt the functions of these proteins in nacre growth. Each anti-body-injected group contained four oysters. As a control, preim-mune rabbit serum was injected at same dosage. Three dayslater, the oysters were sacrificed. The inner surfaces of the nacrewere examined by a SEM.
3.5.1. Treatment of anti-P43In low dosage anti-P43 injected-group, the aragonite nanograins
failed to pack into a compact aggregation as the untreated ones(Fig. 8C), and became disordered (Fig.8C and D). As the injection dos-
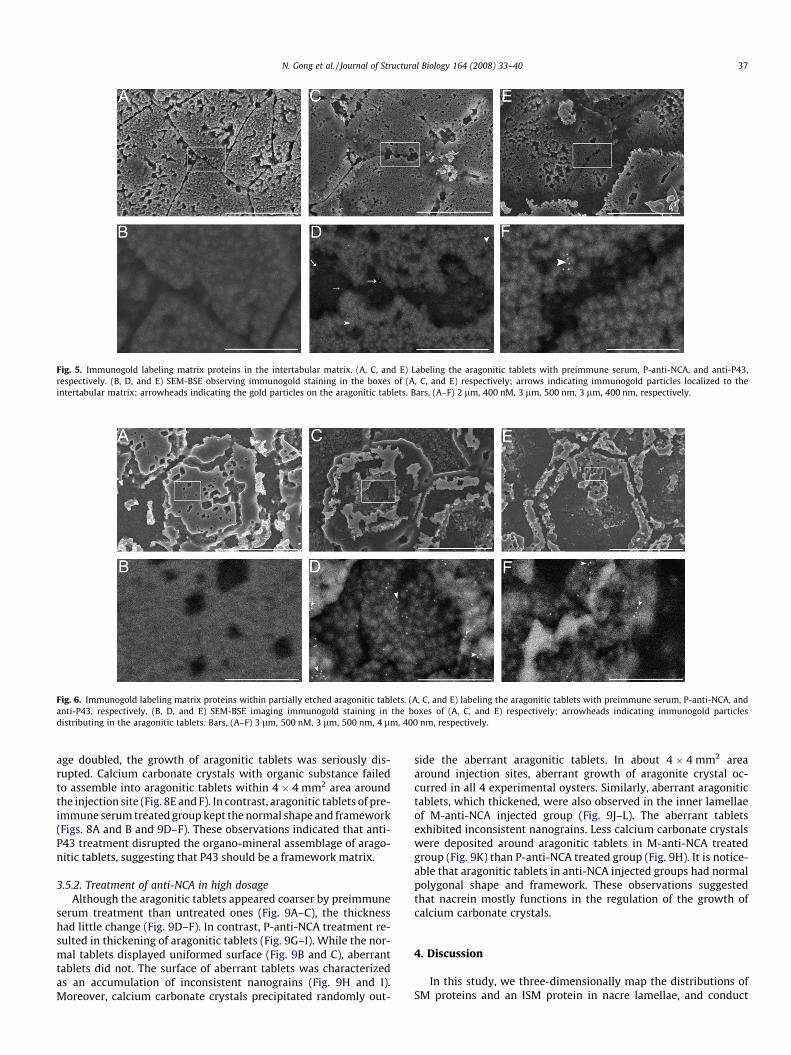
Fig. 6. Immunogold labeling matrix proteins within partially etched aragonitic tablets. (A, C, and E) labeling the aragonitic tablets with preimmune serum, P-anti-NCA, andanti-P43, respectively. (B, D, and E) SEM-BSE imaging immunogold staining in the boxes of (A, C, and E) respectively; arrowheads indicating immunogold particlesdistributing in the aragonitic tablets. Bars, (A–F) 3 lm, 500 nM, 3 lm, 500 nm, 4 lm, 400 nm, respectively.
Fig. 5. Immunogold labeling matrix proteins in the intertabular matrix. (A, C, and E) Labeling the aragonitic tablets with preimmune serum, P-anti-NCA, and anti-P43,respectively. (B, D, and E) SEM-BSE observing immunogold staining in the boxes of (A, C, and E) respectively; arrows indicating immunogold particles localized to theintertabular matrix; arrowheads indicating the gold particles on the aragonitic tablets. Bars, (A–F) 2 lm, 400 nM, 3 lm, 500 nm, 3 lm, 400 nm, respectively.
N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40 37
age doubled, the growth of aragonitic tablets was seriously dis-rupted. Calcium carbonate crystals with organic substance failedto assemble into aragonitic tablets within 4 � 4 mm2 area aroundthe injection site (Fig. 8E and F). In contrast, aragonitic tablets of pre-immune serum treated group kept the normal shape and framework(Figs. 8A and B and 9D–F). These observations indicated that anti-P43 treatment disrupted the organo-mineral assemblage of arago-nitic tablets, suggesting that P43 should be a framework matrix.
3.5.2. Treatment of anti-NCA in high dosageAlthough the aragonitic tablets appeared coarser by preimmune
serum treatment than untreated ones (Fig. 9A–C), the thicknesshad little change (Fig. 9D–F). In contrast, P-anti-NCA treatment re-sulted in thickening of aragonitic tablets (Fig. 9G–I). While the nor-mal tablets displayed uniformed surface (Fig. 9B and C), aberranttablets did not. The surface of aberrant tablets was characterizedas an accumulation of inconsistent nanograins (Fig. 9H and I).Moreover, calcium carbonate crystals precipitated randomly out-
side the aberrant aragonitic tablets. In about 4 � 4 mm2 areaaround injection sites, aberrant growth of aragonite crystal oc-curred in all 4 experimental oysters. Similarly, aberrant aragonitictablets, which thickened, were also observed in the inner lamellaeof M-anti-NCA injected group (Fig. 9J–L). The aberrant tabletsexhibited inconsistent nanograins. Less calcium carbonate crystalswere deposited around aragonitic tablets in M-anti-NCA treatedgroup (Fig. 9K) than P-anti-NCA treated group (Fig. 9H). It is notice-able that aragonitic tablets in anti-NCA injected groups had normalpolygonal shape and framework. These observations suggestedthat nacrein mostly functions in the regulation of the growth ofcalcium carbonate crystals.
4. Discussion
In this study, we three-dimensionally map the distributions ofSM proteins and an ISM protein in nacre lamellae, and conduct
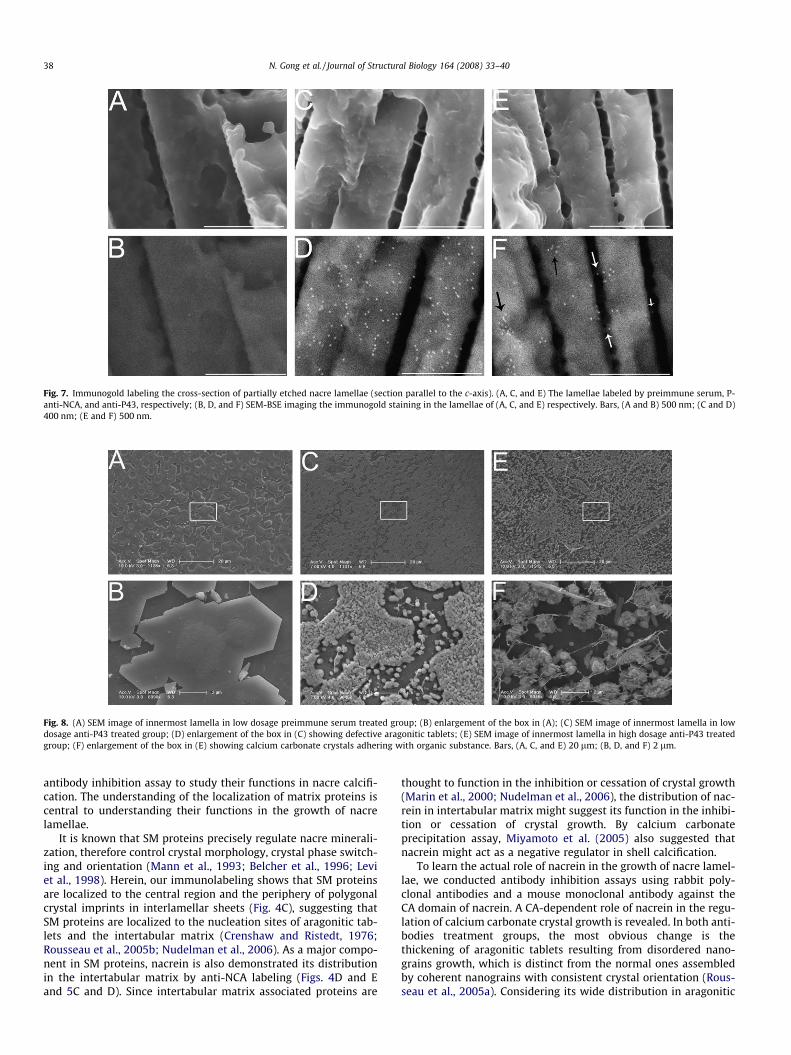
Fig. 7. Immunogold labeling the cross-section of partially etched nacre lamellae (section parallel to the c-axis). (A, C, and E) The lamellae labeled by preimmune serum, P-anti-NCA, and anti-P43, respectively; (B, D, and F) SEM-BSE imaging the immunogold staining in the lamellae of (A, C, and E) respectively. Bars, (A and B) 500 nm; (C and D)400 nm; (E and F) 500 nm.
Fig. 8. (A) SEM image of innermost lamella in low dosage preimmune serum treated group; (B) enlargement of the box in (A); (C) SEM image of innermost lamella in lowdosage anti-P43 treated group; (D) enlargement of the box in (C) showing defective aragonitic tablets; (E) SEM image of innermost lamella in high dosage anti-P43 treatedgroup; (F) enlargement of the box in (E) showing calcium carbonate crystals adhering with organic substance. Bars, (A, C, and E) 20 lm; (B, D, and F) 2 lm.
38 N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40
antibody inhibition assay to study their functions in nacre calcifi-cation. The understanding of the localization of matrix proteins iscentral to understanding their functions in the growth of nacrelamellae.
It is known that SM proteins precisely regulate nacre minerali-zation, therefore control crystal morphology, crystal phase switch-ing and orientation (Mann et al., 1993; Belcher et al., 1996; Leviet al., 1998). Herein, our immunolabeling shows that SM proteinsare localized to the central region and the periphery of polygonalcrystal imprints in interlamellar sheets (Fig. 4C), suggesting thatSM proteins are localized to the nucleation sites of aragonitic tab-lets and the intertabular matrix (Crenshaw and Ristedt, 1976;Rousseau et al., 2005b; Nudelman et al., 2006). As a major compo-nent in SM proteins, nacrein is also demonstrated its distributionin the intertabular matrix by anti-NCA labeling (Figs. 4D and Eand 5C and D). Since intertabular matrix associated proteins are
thought to function in the inhibition or cessation of crystal growth(Marin et al., 2000; Nudelman et al., 2006), the distribution of nac-rein in intertabular matrix might suggest its function in the inhibi-tion or cessation of crystal growth. By calcium carbonateprecipitation assay, Miyamoto et al. (2005) also suggested thatnacrein might act as a negative regulator in shell calcification.
To learn the actual role of nacrein in the growth of nacre lamel-lae, we conducted antibody inhibition assays using rabbit poly-clonal antibodies and a mouse monoclonal antibody against theCA domain of nacrein. A CA-dependent role of nacrein in the regu-lation of calcium carbonate crystal growth is revealed. In both anti-bodies treatment groups, the most obvious change is thethickening of aragonitic tablets resulting from disordered nano-grains growth, which is distinct from the normal ones assembledby coherent nanograins with consistent crystal orientation (Rous-seau et al., 2005a). Considering its wide distribution in aragonitic
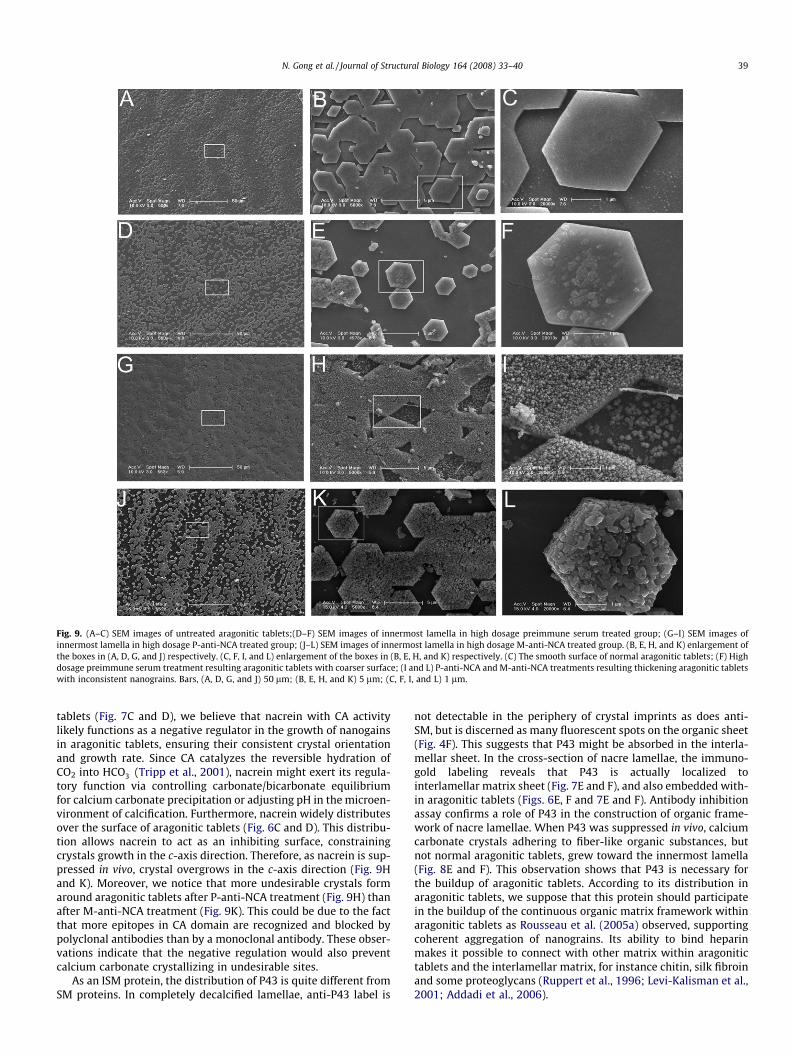
Fig. 9. (A–C) SEM images of untreated aragonitic tablets;(D–F) SEM images of innermost lamella in high dosage preimmune serum treated group; (G–I) SEM images ofinnermost lamella in high dosage P-anti-NCA treated group; (J–L) SEM images of innermost lamella in high dosage M-anti-NCA treated group. (B, E, H, and K) enlargement ofthe boxes in (A, D, G, and J) respectively. (C, F, I, and L) enlargement of the boxes in (B, E, H, and K) respectively. (C) The smooth surface of normal aragonitic tablets; (F) Highdosage preimmune serum treatment resulting aragonitic tablets with coarser surface; (I and L) P-anti-NCA and M-anti-NCA treatments resulting thickening aragonitic tabletswith inconsistent nanograins. Bars, (A, D, G, and J) 50 lm; (B, E, H, and K) 5 lm; (C, F, I, and L) 1 lm.
N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40 39
tablets (Fig. 7C and D), we believe that nacrein with CA activitylikely functions as a negative regulator in the growth of nanogainsin aragonitic tablets, ensuring their consistent crystal orientationand growth rate. Since CA catalyzes the reversible hydration ofCO2 into HCO�3 (Tripp et al., 2001), nacrein might exert its regula-tory function via controlling carbonate/bicarbonate equilibriumfor calcium carbonate precipitation or adjusting pH in the microen-vironment of calcification. Furthermore, nacrein widely distributesover the surface of aragonitic tablets (Fig. 6C and D). This distribu-tion allows nacrein to act as an inhibiting surface, constrainingcrystals growth in the c-axis direction. Therefore, as nacrein is sup-pressed in vivo, crystal overgrows in the c-axis direction (Fig. 9Hand K). Moreover, we notice that more undesirable crystals formaround aragonitic tablets after P-anti-NCA treatment (Fig. 9H) thanafter M-anti-NCA treatment (Fig. 9K). This could be due to the factthat more epitopes in CA domain are recognized and blocked bypolyclonal antibodies than by a monoclonal antibody. These obser-vations indicate that the negative regulation would also preventcalcium carbonate crystallizing in undesirable sites.
As an ISM protein, the distribution of P43 is quite different fromSM proteins. In completely decalcified lamellae, anti-P43 label is
not detectable in the periphery of crystal imprints as does anti-SM, but is discerned as many fluorescent spots on the organic sheet(Fig. 4F). This suggests that P43 might be absorbed in the interla-mellar sheet. In the cross-section of nacre lamellae, the immuno-gold labeling reveals that P43 is actually localized tointerlamellar matrix sheet (Fig. 7E and F), and also embedded with-in aragonitic tablets (Figs. 6E, F and 7E and F). Antibody inhibitionassay confirms a role of P43 in the construction of organic frame-work of nacre lamellae. When P43 was suppressed in vivo, calciumcarbonate crystals adhering to fiber-like organic substances, butnot normal aragonitic tablets, grew toward the innermost lamella(Fig. 8E and F). This observation shows that P43 is necessary forthe buildup of aragonitic tablets. According to its distribution inaragonitic tablets, we suppose that this protein should participatein the buildup of the continuous organic matrix framework withinaragonitic tablets as Rousseau et al. (2005a) observed, supportingcoherent aggregation of nanograins. Its ability to bind heparinmakes it possible to connect with other matrix within aragonitictablets and the interlamellar matrix, for instance chitin, silk fibroinand some proteoglycans (Ruppert et al., 1996; Levi-Kalisman et al.,2001; Addadi et al., 2006).

40 N. Gong et al. / Journal of Structural Biology 164 (2008) 33–40
Since the discovery that nacre powder can induce bone forma-tion in vivo (Westbroek and Marin, 1998), BMP homologs arehypothesized to exist in the matrix of the nacreous shell layer.P43, which is a matrix protein, can be specifically recognized bythe antibodies against the recombinant protein encoding theTGF-b domain of Pf-BMP-2. This suggests that there are somehomological epitopes expressed in P43 and TGF-b domain of Pf-BMP-2. Similar to mammalian BMP-2 (Ruppert et al., 1996), P43exhibits an affinity for heparin in a heparin affinity chromatogra-phy. Immunoblot analysis and affinity chromatography revealsome features of P43. Future work will involve purification ofP43. When fully characterized, P43 could be proved importantnot only in biomineralogy but also in medicine and evolutionarybiology.
Our results show that SM and ISM proteins differentially dis-tribute in nacre lamellae and function differently in nacre miner-alization. Some regulating activities of matrix proteins in thegrowth of aragonitic tablets would be indicated by antibodyinhibition assays. The analysis of aberrant growth pattern of ara-gonitic tablets along with immunolocalization would further re-veal the regulatory mechanisms of matrix proteins in nacremineralization.
Acknowledgments
This work was financially supported by the National High Tech-nology Research and Development Program of China(2003AA603430, 2006AA09Z413) and the Natural Science Founda-tion of China (30221003, 30530600). We are grateful to Dr. Yan Ma,Dana-Farber Cancer Institute, Boston, for critical discussion.
References
Addadi, L., Joester, D., Nudelman, F., Weiner, S., 2006. Mollusk shell formation: asource of new concepts for understanding biomineralization processes. Chem.Eur. J. 12, 980–987.
Belcher, A.M., Wu, X.H., Christensen, R.J., Hansma, P.K., Stucky, G.D., Morse, D.E.,1996. Control of crystal phase switching and orientation by soluble mollusc-shell proteins. Nature 381, 56–58.
Cho, J.W., Partin, J.S., Lennnarz, W.J., 1996. A technique for detecting matrix proteinsin the crystalline spicules of the sea urchin embryo. Proc. Natl. Acad. Sci. USA 93,1282–1286.
Crenshaw, M.A., 1972. The soluble matrix from Mercenaria mercenaria shell.Biomineralization 6, 6–11.
Crenshaw, M.A., Ristedt, H., 1976. The histochemical localization of reactive groupsin septal nacre from Nautilus pompilius. In: Watabe, N., Wilbur, K.M. (Eds.), TheMechanisms of Mineralization in the Invertebrates and Plants. University ofSouth Carolina Press, Colombia, pp. 355–367.
Freeman, J.A., 1960. Influence of carbonic anhydrase inhibitors on shell growth offreshwater snail Physa heterostropha. Biol. Bull 118, 412–418.
Gong, N., Li, Q., Huang, J., Fang, Z., Xie, L., Zhang, R., 2008. Culture of outer epithelialcells from the mantle tissue to study matrix protein secretion forbiomineralization. Cell Tissue Res. doi:10.1007/s00441-008-0609-5.
Lamghari, M., Almeida, M.J., Berland, S., Huet, H., Laurent, A., Milet, C., Lopez, E.,1999. Stimulation of bone marrow cells and bone formation by nacre: in vivoand in vitro studies. Bone 25 (Suppl. 2), 91S–99S.
Levi, Y., Albeck, S., Brack, A., Weiner, S., Addadi, L., 1998. Control over aragonitecrystal nucleation and growth: an in vitro study of biomineralization. Chem.Eur. J. 4, 389–396.
Levi-Kalisman, Y., Falini, G., Addadi, L., Weiner, S., 2001. Structure of the nacreousorganic matrix of a bivalve mollusk shell examined in the hydrated state usingcryo-TEM. J. Struct. Biol. 135, 8–17.
Ma, Z., Huang, J., Sun, J., Wang, G., Li, C., Xie, L., Zhang, R., 2007. A novel extrapallialfluid protein controls the morphology of nacre lamellae in the pearl oyster,Pinctada fucata. J. Biol. Chem. 282, 23253–23263.
Mann, S., Archibald, D.D., Didymus, J.M., Douglas, T., Heywood, B.R., Meldrum, F.C.,Reeves, N.J., 1993. Crystallization at inorganic–organic interfaces: biomineralsand biomimetic synthesis. Science 261, 1286–1292.
Marin, F., de Corstjens, P., de Gaulejac, B., Vrind-De, Jong E., Westbroek, P., 2000.Mucins and molluscan calcification. Molecular characterization of mucoperlin, anovel mucin-like protein from the nacreous shell layer of the fan mussel Pinnanobilis (Bivalvia, pteriomorphia). J. Biol. Chem. 275, 20667–20675.
Marin, F., Pokroy, B., Luqueta, G., Layrollec, P., Groot, K.D., 2007. Protein mapping ofcalcium carbonate biominerals by immunogold. Biomaterials 28, 2368–2377.
Mayer, R.J., Walker, J.H., 1987. Immunochemical Methods in Cell and MolecularBiology. Academic Press, London.
Miyamoto, H., Miyashita, T., Okushima, M., Nakano, S., Morita, T., Matsushiro, A.,1996. A carbonic anhydrase from the nacreous layer in oyster pearls. Proc. Natl.Acad. Sci. USA 93, 9657–9660.
Miyamoto, H., Miyoshi, F., Kohno, J., 2005. The carbonic anhydrase domain proteinnacrein is expressed in the epithelial cells of the mantle and acts as a negativeregulator in calcification in the mollusk Pinctada fucata. Zool. Sci. 22, 311–315.
Mouriès, L.P., Almeida, M.J., Milet, C., Berland, S., Lopez, E., 2002. Bioactivity of nacrewater-soluble organic matrix from the bivalve mollusk Pinctada maxima inthree mammalian cell types: fibroblasts, bone marrow stromal cells andosteoblasts. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 132, 217–229.
Nudelman, F., Gotliv, B.A., Addadi, L., Weiner, S., 2006. Mollusk shell formation:mapping the distribution of organic matrix components underlying a singlearagonitic tablet in nacre. J. Struct. Biol. 153, 176–187.
Puverel, S., Tambutté, E., Zoccola, D., Domart-Coulon, I., Bouchot, A., Lotto, S.,Allemand, D., Tambutté, S., 2005. Antibodies against the organic matrix inscleractinians: a new tool to study coral biomineralization. Coral Reefs 24, 149–156.
Rousseau, M., Lopez, E., Stempflé, P., Brendlé, M., Franke, L., Guette, A., Naslain, R.,Bourrat, X., 2005a. Multiscale structure of sheet nacre. Biomaterials 26, 6254–6262.
Rousseau, M., Lopez, E., Couté, A., Mascarel, G., Smith, D.C., Naslain, R., Bourrat, X.,2005b. Sheet nacre growth mechanism: a Voronoi model. J. Struct. Biol. 149,149–157.
Ruppert, R., Hoffmann, E., Sebald, W., 1996. Human bone morphogenetic protein 2contains a heparin-binding site which modifies its biological activity. Eur. J.Biochem. 237, 295–302.
Silve, C., Lopez, E., Vidal, B., Smith, D.C., Camprasse, S., Camprasse, G., Couly, G.,1992. Nacre initiates biomineralization by human osteoblasts maintainedin vitro. Calcif. Tissue Int. 1, 363–369.
Stolarski, J., Mazur, M., 2005. Nanostructure of biogenic versus abiogenic calciumcarbonate crystals. Acta Palaeontol. Pol. 50, 847–865.
Sudo, S., Fujikawa, T., Nagakura, T., Ohkubo, T., Sakaguchi, K., Tanaka, M.,Nakashima, K., Takahashi, T., 1997. Structures of mollusc shell frameworkproteins. Nature 387, 563–564.
Takakura, D., Norizuki, M., Ishikawa, F., Samata, T., 2008. Isolation andcharacterization of the N-linked oligosaccharides in nacrein from Pinctadafucata. Mar. Biotechnol. (NY) 10, 290–296.
Tripp, B.C., Smith, K., Ferry, J.G., 2001. Carbonic anhydrase: new insights for anancient enzyme. J. Biol. Chem. 276, 48615–48618.
Wada, K., 1968. Mechanism of growth of nacre in bivalvia. Bull. Natl. Pearl. Res. Lab.13, 1561–1596.
Weiner, S., Addadi, L., 1997. Design strategies in mineralized biological materials. J.Mater. Chem. 7, 689–702.
Weiner, S., Gotliv, B., Levi-Kalisman, Y., Raz, S., Weiss, I. M., Addadi, L., 2003.Biomineralization (BIOM2001): formation, diversity, evolution and application.In: Kobayashi, I., Ozawa, H. (Eds.), Proceedings of the 8th InternationalSymposium on Biomineralization. Tokai University Press, Kanagawa, pp. 8-13.
Westbroek, P., Marin, F., 1998. A marriage of bone and nacre. Nature 392, 861–862.Wilbur, K.M., Jodrey, L., 1955. Studies of shell formation. V. The inhibition of shell
formation by carbonic anhydrase inhibitors.. Biol. Bull. (Woods Hole) 108, 82–112.
Zhang, C., Zhang, R., 2006. Matrix proteins in the outer shells of molluscs. Mar.Biotechnol. (NY) 8, 572–586.





























