Embed Size (px)
Citation preview

803
J. Parasitol., 94(4), 2008, pp. 803–810� American Society of Parasitologists 2008
SEM AND TEM STUDY OF THE ARMED MALE TERMINAL GENITALIA OF THETAPEWORM PARAECHINOPHALLUS JAPONICUS (CESTODA: BOTHRIOCEPHALIDEA)
Celine Levron, Larisa G. Poddubnaya*, Roman Kuchta†, Mark Freeman‡, Yan-Hai Wang§, and Tomas Scholz†Institute of Parasitology, Academy of Sciences of the Czech Republic, Branisovska 31, 370 05 Ceske Budejovice, Czech Republic.e-mail: [email protected]
ABSTRACT: For the first time, the ultrastructure of the armed cirrus of an echinophallid cestode, Paraechinophallus japonicus(Yamaguti, 1934), has been studied with the use of scanning and transmission electron microscopy. Two sets of eversiblecopulatory organs (�300 �m in length and �130 �m in width) are present on the dorsal side of each segment near the lateralmargin of the strobila. Except for the terminal portion, the cirrus is covered with large spines (up to 40 �m long, measured fromSEM photomicrographs) composed of 2 parts. The basal portion contains a lobed electron-dense outer region that gives way toa reticular meshwork of electron-dense material. The apical region of the spines, composed of a homogeneous, moderatelyelectron-dense matrix, is slightly curved distally. Spines are covered with a cortical zone. Between the spines, the distal cytoplasmis covered with microvilli of about 1.2 �m in length. The wall of the cirrus sac, which is �500 �m long and �250 �m wide,is composed of 2 layers of muscles, i.e., an internal layer of circular muscles and external longitudinal muscles. Microvilli onthe cirrus of P. japonicus are reported for the first time in the Cestoda, whereas the spines on the cirrus may represent asynapomorphy of bothriocephalidean cestodes of the Echinophallidae.
Characters of the male reproductive system in tapeworms(Cestoda), especially the ultrastructure of spermiogenesis andspermatozoon, are considered as valuable tools in phylogeneticstudies (Hoberg et al., 1997; Justine, 1998; Hoberg et al., 2001;Olson et al., 2001). The shape and size of the cirrus sac andsurface structures (spines, microtriches) of the cirrus are alsocharacters used in data matrices of some groups of cestodes(Bray et al., 1999; Ivanov and Hoberg, 1999; Kuchta, 2007).
The male system of cestodes may reveal a great structuralvariation (Jones, 1998). Indeed, the cirrus, a male copulatoryorgan, is evaginated (by turning inside out) during copulationand may be or may not be armed with spines, microtriches, etc.(Caira and Littlewood, 2001). Ultrastructural studies of malegenitalia have been performed for a few cestode species (Cza-plinski et al., 1984; Beveridge and Smith, 1985; Jones, 1989;Cielecka et al., 1994; Davydov et al., 1994, 1997; Davydov andKorneva, 2002; Korneva, 2002; Poddubnaya, 2002, 2003; Pod-dubnaya et al., 2005).
The order Pseudophyllidea has recently been suppressed be-cause it contained 2 unrelated assemblages (Brabec et al.,2006), for which 2 new orders, Bothriocephalidea and Diphyl-lobothriidea, were erected (Kuchta et al., 2008). The formergroup contains 4 families, but the cirrus, observed with lightmicroscopy, is armed with spines only in species of Echino-phallidae (Bray et al., 1994; Kuchta, 2007).
The present study describes for the first time the ultrastruc-ture of the male copulatory organ and cirrus sac of an echino-phallid cestode, Paraechinophallus japonicus (Yamaguti,1934). The usage of scanning and transmission electron mi-croscopy also enabled us to examine in detail spines and mi-crovilli present on the surface of the cirrus.
Received 20 September 2007; revised 13 December 2007, 9 January2008; accepted 9 January 2008.
* Institute of Biology of Inland Waters, Russian Academy of Sciences,152742 Borok, Yaroslavl Province, Russia.
† Also at Faculty of Science, University of South Bohemia, Branisovska31, 370 05 Ceske Budejovice, Czech Republic.
‡ Institute of Aquaculture, University of Stirling, Stirling, FK9 4LA,United Kingdom.
§ Parasitology Research Laboratory, School of Life Sciences, XiamenUniversity, Xiamen, Fujian, 361005, People’s Republic of China.
MATERIALS AND METHODS
Adults of P. japonicus were obtained from the intestine of Psenopsisanomala (Temminck & Schlegel, 1844) (Perciformes: Centrolophidae),collected from an Inland Sea area, Japan, in 2004 by 1 of us (M.F.).Live worms were rinsed in 0.9% NaCl, their mature and gravid seg-ments were separated and fixed with 2.5% glutaraldehyde in cacodylatebuffer for 24 hr, washed overnight in 0.1 M sodium cacodylate bufferat pH 7.4, postfixed in cold (4 C) 1% OsO4 in the same buffer for 1 hr,dehydrated in a graded series of acetone, and embedded in Spurr’s ep-oxy resin within BEEM capsules. Ultrathin sections (60–90 nm in thick-ness) were cut on a Leica Ultracut UCT ultramicrotome, placed oncopper grids, and stained with uranyl acetate and lead citrate accordingto Reynolds (1963). The grids were examined with the use of a JEOLJEM 1010 transmission electron microscope operating at 80 kV.
Specimens of P. japonicus for SEM and histological investigationswere obtained from the same species of fish collected in the fish marketat Xiamen, Formosa Strait, China, by 1 of us (Y.H.W.). For SEM, theworms were fixed with 3% glutaraldehyde in 0.1 M sodium cacodylatebuffer (pH 7.2) for 30 days at 5 C, rinsed for 20 min 4 times in thesame buffer, and postfixed in 1% osmium tetroxide for 1 hr. Fixedtapeworms were dehydrated in a graded series of ethanol and acetone,and were critical-point dried with liquid CO2. The specimens were sput-ter-coated with gold–palladium and examined on JEOL JSM 6700F(accelerating voltage 1.5 kV) and LEO-1420 (accelerating voltage 15kV). For histological investigations, cross and longitudinal sectionswere prepared by a standard procedure (staining with Heidenhain he-matoxylin-eosin and then mounting in Canada balsam; thickness of sec-tions was 12 �m).
RESULTS
General characters
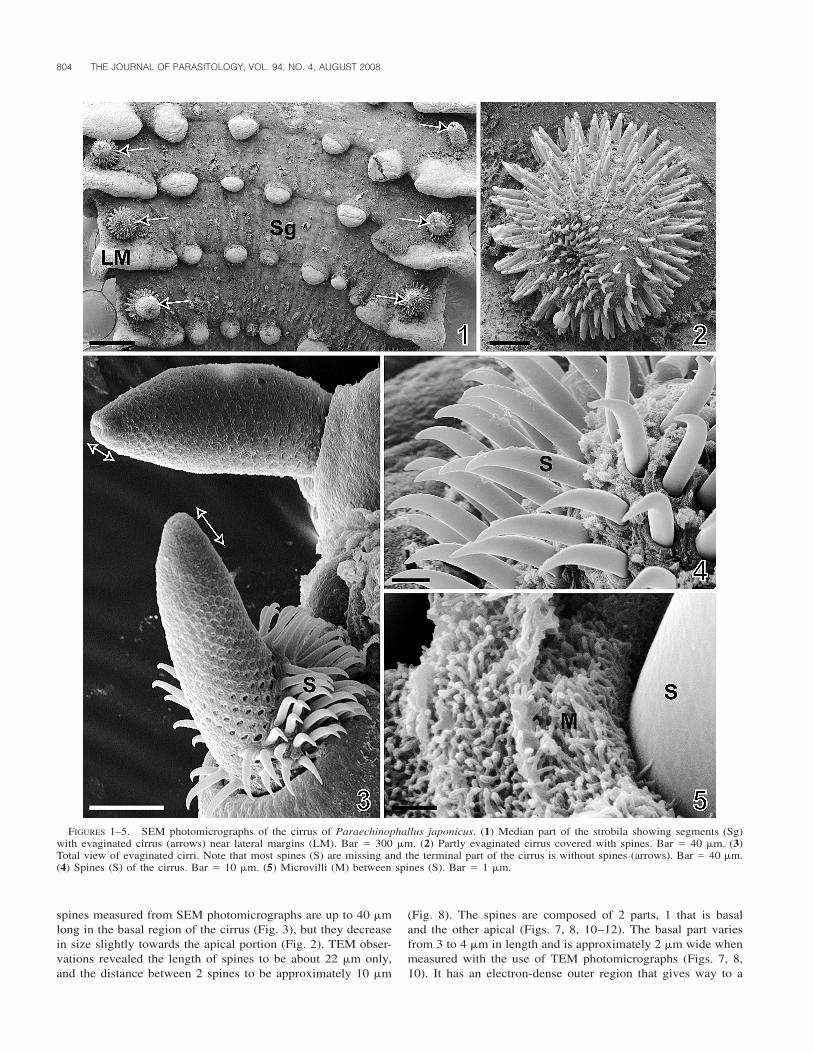
In P. japonicus, 2 eversible copulatory organs, each called acirrus, are present in each segment (Fig. 1). They are localizedon the dorsal side of the strobila, near the lateral margin in themiddle of the length of the segment (Fig. 1). They occur ap-proximately from the eighth segment. The cirrus is often evag-inated in specimens fixed for light and scanning microscopyobservations; it measures approximately 300 �m in length and130 �m in width (N � 40).
Ultrastructural characters of spines
Except for the most terminal part (approximately 20 �m), thecirrus is covered with large spines (Figs. 2–4, 7, 8). Spines aredistributed in spiral along the cirrus and their terminal part isslightly curved posteriorly towards the segment (Figs. 3, 4). The
804 THE JOURNAL OF PARASITOLOGY, VOL. 94, NO. 4, AUGUST 2008
FIGURES 1–5. SEM photomicrographs of the cirrus of Paraechinophallus japonicus. (1) Median part of the strobila showing segments (Sg)with evaginated cirrus (arrows) near lateral margins (LM). Bar � 300 �m. (2) Partly evaginated cirrus covered with spines. Bar � 40 �m. (3)Total view of evaginated cirri. Note that most spines (S) are missing and the terminal part of the cirrus is without spines (arrows). Bar � 40 �m.(4) Spines (S) of the cirrus. Bar � 10 �m. (5) Microvilli (M) between spines (S). Bar � 1 �m.
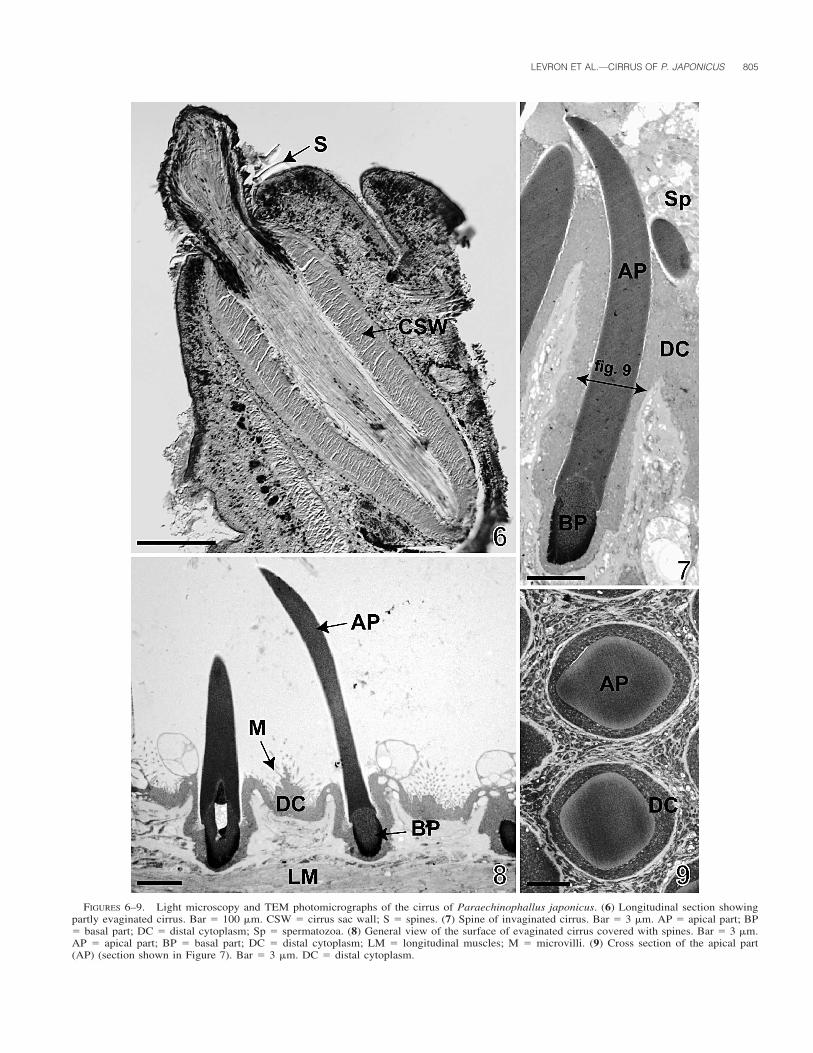
spines measured from SEM photomicrographs are up to 40 �mlong in the basal region of the cirrus (Fig. 3), but they decreasein size slightly towards the apical portion (Fig. 2). TEM obser-vations revealed the length of spines to be about 22 �m only,and the distance between 2 spines to be approximately 10 �m
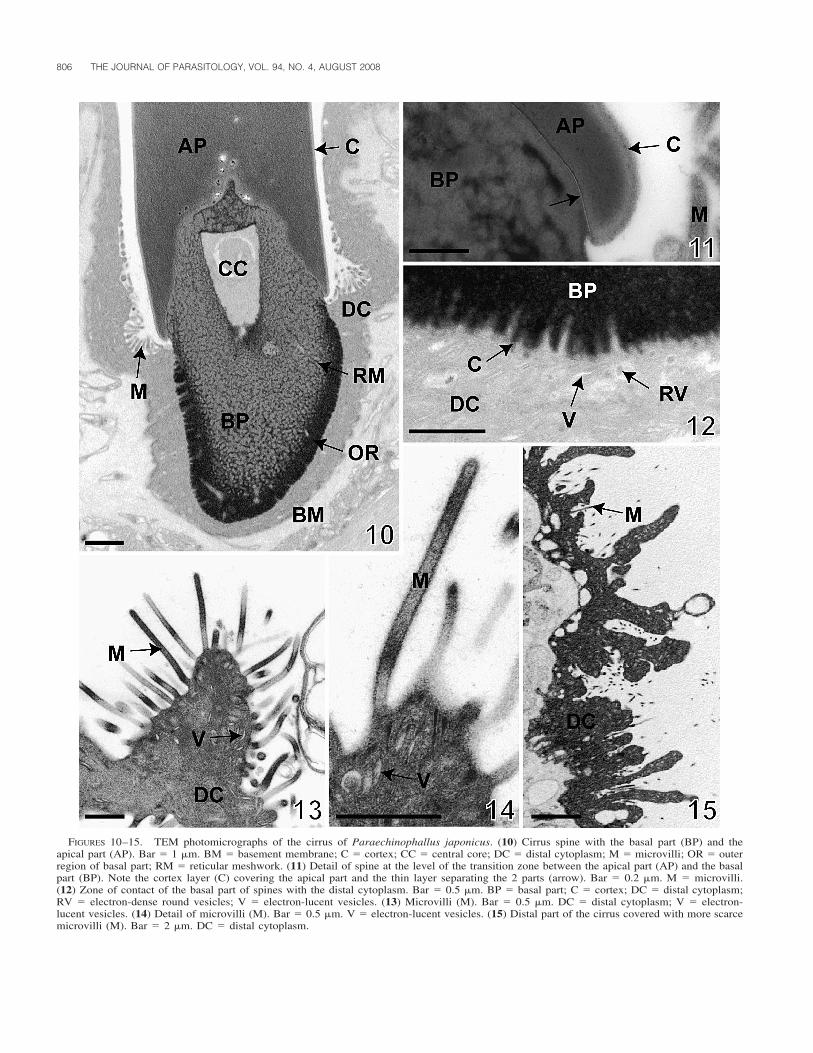
(Fig. 8). The spines are composed of 2 parts, 1 that is basaland the other apical (Figs. 7, 8, 10–12). The basal part variesfrom 3 to 4 �m in length and is approximately 2 �m wide whenmeasured with the use of TEM photomicrographs (Figs. 7, 8,10). It has an electron-dense outer region that gives way to a
LEVRON ET AL.—CIRRUS OF P. JAPONICUS 805
FIGURES 6–9. Light microscopy and TEM photomicrographs of the cirrus of Paraechinophallus japonicus. (6) Longitudinal section showingpartly evaginated cirrus. Bar � 100 �m. CSW � cirrus sac wall; S � spines. (7) Spine of invaginated cirrus. Bar � 3 �m. AP � apical part; BP� basal part; DC � distal cytoplasm; Sp � spermatozoa. (8) General view of the surface of evaginated cirrus covered with spines. Bar � 3 �m.AP � apical part; BP � basal part; DC � distal cytoplasm; LM � longitudinal muscles; M � microvilli. (9) Cross section of the apical part(AP) (section shown in Figure 7). Bar � 3 �m. DC � distal cytoplasm.
806 THE JOURNAL OF PARASITOLOGY, VOL. 94, NO. 4, AUGUST 2008
FIGURES 10–15. TEM photomicrographs of the cirrus of Paraechinophallus japonicus. (10) Cirrus spine with the basal part (BP) and theapical part (AP). Bar � 1 �m. BM � basement membrane; C � cortex; CC � central core; DC � distal cytoplasm; M � microvilli; OR � outerregion of basal part; RM � reticular meshwork. (11) Detail of spine at the level of the transition zone between the apical part (AP) and the basalpart (BP). Note the cortex layer (C) covering the apical part and the thin layer separating the 2 parts (arrow). Bar � 0.2 �m. M � microvilli.(12) Zone of contact of the basal part of spines with the distal cytoplasm. Bar � 0.5 �m. BP � basal part; C � cortex; DC � distal cytoplasm;RV � electron-dense round vesicles; V � electron-lucent vesicles. (13) Microvilli (M). Bar � 0.5 �m. DC � distal cytoplasm; V � electron-lucent vesicles. (14) Detail of microvilli (M). Bar � 0.5 �m. V � electron-lucent vesicles. (15) Distal part of the cirrus covered with more scarcemicrovilli (M). Bar � 2 �m. DC � distal cytoplasm.

LEVRON ET AL.—CIRRUS OF P. JAPONICUS 807
reticular meshwork of electron-dense material more interiorly(Figs. 10, 12). The reticulum surrounds an area of moderatelyintense electron density. The zone of contact of the basal partwith the distal cytoplasm forms several irregular lobed digita-tions covered by a thin layer of cortex (Fig. 12). Some spinescontain an amorphous electron-lucent central core in the centerof the basal area (Figs. 8, 10). A thin line separates the basalregion from the apex (Fig. 11). The apical part of the spines,18–19 �m long and approximately 2.3 �m wide (TEM mea-surements), is slightly curved distally, near the pointed end, andis composed of homogeneous moderately electron-dense matrix(Figs. 4, 7, 8). Cross sections show that the apical region of thespines is more or less rectangular, with widely rounded corners(Fig. 9). A cortex, 60 nm thick, covers the apical region of thespines (Figs. 10, 11). When the cirrus is invaginated, the spinesare largely covered by epithelium and only the distal tips areseen in cirri with lumens filled with spermatozoa (Fig. 7).
Ultrastructural characters of microvilli
Between the spines (Fig. 5), the distal cytoplasm is coveredwith microvilli measuring approximately 1.2 �m in length and70 nm in width (Figs. 13–15). The microvilli are present duringthe entire length of the cirrus, but become scarce in the distalregion (Fig. 15).
Ultrastructural characters of the cirrus epithelium andcirrus sac
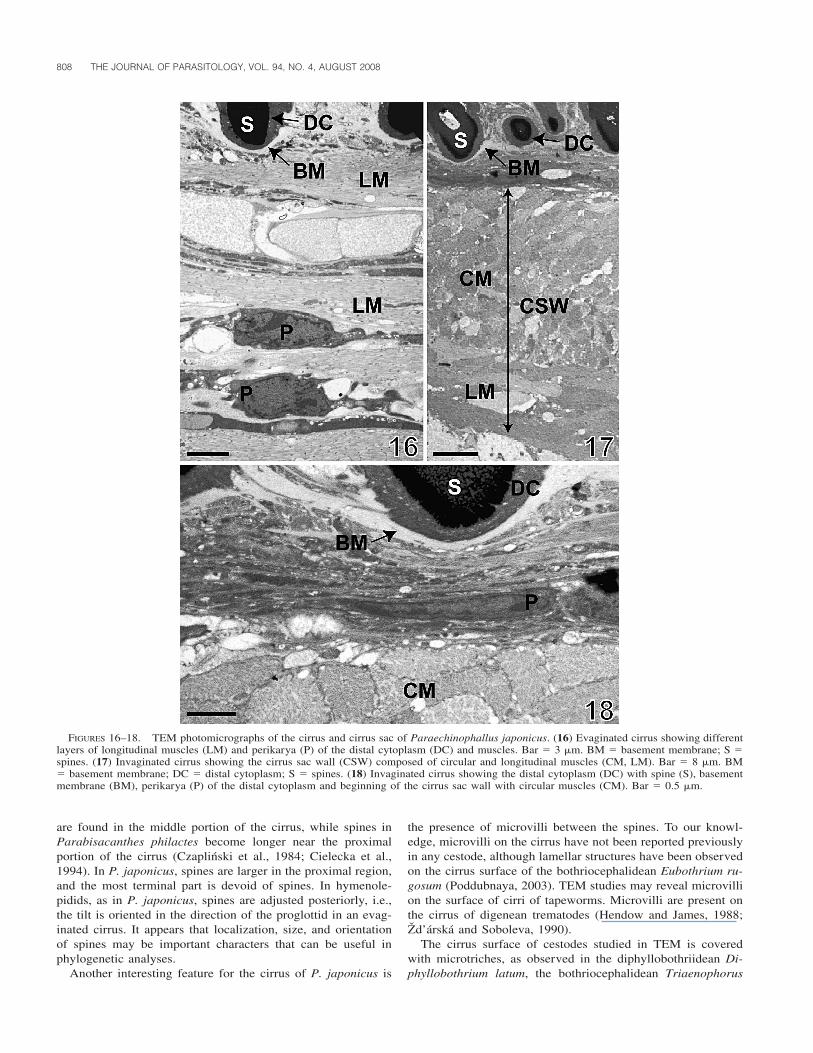
The epithelial wall of the cirrus is composed of an outer layerthat includes the distal cytoplasm with small, elongated elec-tron-lucent vesicles (Figs. 12–14). Toward the basal part of thespines, some round electron-dense vesicles are present in thedistal cytoplasm (Fig. 12). An electron-lucent basement mem-brane separates the distal cytoplasm from the cirrus sac (Figs.16–18). Beneath this membrane, longitudinal muscles and elon-gated cells of variable size corresponding to the perikarya ofthe distal cytoplasm and muscles are visible (Figs. 16, 18).
The invaginated cirrus is localized within the cirrus sac,which is an elongated, thick-walled structure. The size of thecirrus sac varies from 400 to 550 �m in length and from 180�m to 280 �m in width, as measured with the use of lightmicroscopy (Fig. 6). The cirrus sac wall is composed of 2 layersof muscles (Figs. 6, 17). The internal layer is about 30 �m inthickness (TEM observation), and is formed by circular muscle(Figs. 6, 17, 18). The external layer, considerably thinner, isonly about 4 �m thick (TEM observation) and consists of lon-gitudinal muscles (Fig. 17).
DISCUSSION
The present scanning and transmission electron microscopicobservations have revealed that the cirrus surface of the tape-worm P. japonicus is covered with large spines. Cirrus spineshave not been observed among members of the most basal or-ders, i.e., Amphilinidea, Gyrocotylidea, Spathebothriidea, andCaryophyllidea (Hoberg et al., 1999, 2001; Olson and Tkach,2005). They seem to be typical for more derived groups ofcestodes, especially cyclophyllideans (Khalil et al., 1994).Bothriocephalideans, which are probably the most derived‘‘lower’’ (‘‘bothriate’’) group of the Eucestoda and represent a
sister group of all ‘‘acetabulate’’ tapeworms (Brabec et al.,2006; Waeschenbach et al., 2007; Kuchta et al., 2008), are de-void of an armed cirrus, except for the Echinophallidae, mostspecies of which parasitize deep-sea or bathypelagic fish (Brayet al., 1994, 1999; Kuchta, 2007; Kuchta and Scholz, 2007).Only 1 echinophallid tapeworm, Neobothriocephalus aspinosus,possesses an unarmed cirrus (Mateo and Bullock, 1966; Kuchta,2007).
The solid structures present on the cirrus of cestodes havebeen referred to as ‘‘spines’’ in almost all morphological andtaxonomic studies (Yamaguti, 1934; Protasova, 1977; Schmidt,1986; Bray et al., 1994; Khalil et al., 1994; Bray et al., 1999;Kuchta, 2007). However, to our knowledge, no TEM ultrastruc-tural data exist for these sclerotized structures covering the ter-minal genitalia. Thus, it appears important to compare themwith other hard structures of cestodes that have been identifiedas hooks and spines. From an external structure point of view,the main difference lies in the overall shape, especially the tiltof the hard structure, i.e., spines are not curved or only slightlycurved at their terminal end, whereas hooks are stronglycurved. This definition may be problematic, because the degreeof tilt is sometimes difficult to evaluate. Studies on internalultrastructure should help in distinguishing between spines andhooks. However, few ultrastructural studies exist on this topic.
Externally, typical hooks, e.g., those of cyclophyllideans, arecomposed of 3 parts, the blade (apical part), guard, and handle.The internal structure of hooks has been described in the ros-tellum-like organ of the proteocephalidean cestode Silurotaeniasiluri, tentacles of the trypanorhynch Nybelinia queenslanden-sis, and rostellum of cyclophyllideans Fernandezia spinosissi-ma, Taenia crassiceps, and Taenia parva (Mount, 1970; Jonesand Beveridge, 1998; Stoitsova et al., 2001; Zd’arska and Ne-besarova, 2003; Swiderski et al., 2007). On the basis of thesestudies, it appears that there is no typical model for the internalstructure of hooks. For example, the electron density of thedifferent parts of hooks, the presence of a cortex layer, the pointof anchoring (deep or not in the distal cytoplasm), and theirrelation with musculature, are quite variable in hooks of differ-ent cestodes.
To our knowledge, the ultrastructure of spines has been de-scribed only in the cephalic peduncle of the diphyllidean Echi-nobothrium chisholmae (Jones and Beveridge, 2001). Thespines have generally homogeneous electron-dense matrix andthe basal region is embedded deeply in the distal cytoplasm.
Cirrus spines of P. japonicus differ from all these descrip-tions of hard structures, because they are slightly curved andconsist of 2 distinct parts covered by a cortex, the basal partwith a reticular meshwork of electron-dense material surround-ing a moderately electron-dense matrix and the apical part withhomogeneous material that is moderately electron dense. A thinline separates the basal part from the apex. The presence of thisthin line allows us to compare the base plate of microtriches,from which spines probably evolved. However, only studies onthe formation of spines can help to answer questions regardingtheir evolution.
The cirrus surface has been studied with the use of SEM insome species of the Hymenolepididae (Czaplinski et al, 1984;Cielecka et al., 1994). These cestodes are characterized by acirrus covered with spines and by a stylet that protrudes fromthe cirrus canal. In Sobolevicanthus gracilis, the largest spines
808 THE JOURNAL OF PARASITOLOGY, VOL. 94, NO. 4, AUGUST 2008
FIGURES 16–18. TEM photomicrographs of the cirrus and cirrus sac of Paraechinophallus japonicus. (16) Evaginated cirrus showing differentlayers of longitudinal muscles (LM) and perikarya (P) of the distal cytoplasm (DC) and muscles. Bar � 3 �m. BM � basement membrane; S �spines. (17) Invaginated cirrus showing the cirrus sac wall (CSW) composed of circular and longitudinal muscles (CM, LM). Bar � 8 �m. BM� basement membrane; DC � distal cytoplasm; S � spines. (18) Invaginated cirrus showing the distal cytoplasm (DC) with spine (S), basementmembrane (BM), perikarya (P) of the distal cytoplasm and beginning of the cirrus sac wall with circular muscles (CM). Bar � 0.5 �m.
are found in the middle portion of the cirrus, while spines inParabisacanthes philactes become longer near the proximalportion of the cirrus (Czaplinski et al., 1984; Cielecka et al.,1994). In P. japonicus, spines are larger in the proximal region,and the most terminal part is devoid of spines. In hymenole-pidids, as in P. japonicus, spines are adjusted posteriorly, i.e.,the tilt is oriented in the direction of the proglottid in an evag-inated cirrus. It appears that localization, size, and orientationof spines may be important characters that can be useful inphylogenetic analyses.
Another interesting feature for the cirrus of P. japonicus is
the presence of microvilli between the spines. To our knowl-edge, microvilli on the cirrus have not been reported previouslyin any cestode, although lamellar structures have been observedon the cirrus surface of the bothriocephalidean Eubothrium ru-gosum (Poddubnaya, 2003). TEM studies may reveal microvillion the surface of cirri of tapeworms. Microvilli are present onthe cirrus of digenean trematodes (Hendow and James, 1988;Zd’arska and Soboleva, 1990).
The cirrus surface of cestodes studied in TEM is coveredwith microtriches, as observed in the diphyllobothriidean Di-phyllobothrium latum, the bothriocephalidean Triaenophorus

LEVRON ET AL.—CIRRUS OF P. JAPONICUS 809
nodulosus, the spathebothriidean Cyathocephalus truncatus, thecaryophyllideans Archigetes sieboldi and Caryophyllaeus lati-ceps, the tetraphyllidean Phyllobothrium vagans, and the cyclo-phyllideans Cylindrotaenia hickmani and Sobolevicanthus grac-ilis (Beveridge and Smith, 1985; Jones, 1989; Davydov et al.,1994; Davydov and Korneva, 2002; Korneva, 2002; Poddub-naya, 2002, 2003; Poddubnaya et al., 2005). It is interesting tonote that all these species possess an unarmed cirrus. The func-tion of these microtriches on the cirrus may be similar to thefunction of the spines, i.e., maintaining the cirrus in the vaginaduring insemination (Beveridge and Smith, 1985; Jones, 1989;Poddubnaya et al., 2005). The role of microvilli on the cirrusof P. japonicus is not known.
In a previous study, we described 2 types of microtrichescovering the strobila of P. japonicus, i.e., filiform and tusk-shaped microtriches (Levron et al., 2008). At the level of thecirrus, filiform microtriches are lost and are replaced by micro-villi and spines. An abrupt change in the surface of cestodeshas already been reported in numerous species, i.e., on the cir-rus of Cylindrotaenia sp. or on the scolex of Nybelinia sp.(Jones, 1989, 1998). Halton (1997) suggested that regional spe-cialization is possible in the syncytial epidermis, but no differ-ences in perikarya have been observed in P. japonicus.
ACKNOWLEDGMENTS
We are grateful to the staff of the Laboratory of Electron Microscopy,Institute of Parasitology, BC AS CR, Ceske Budejovice, Czech Repub-lic and the Centre of Electron Microscopy, Institute of Biology for In-land Waters, RAS Borok, for technical assistance. This study was sup-ported by the Grant Agency of the Czech Republic (Projects 524/04/0342, 524/07/P039, and 524/08/0885), Research Centre of Ichthyopar-asitology (LC 522), Institute of Parasitology (Z60220518), and theRussian Foundation of Fundamental Researches (Grant 05-04-48250 toL.G.P.).
LITERATURE CITED
BEVERIDGE, I., AND K. SMITH. 1985. An ultrastructural study of the cirrusand vagina of Phyllobothrium vagans (Cestoda: Tetraphyllidea).Zeitschrift fur Parasitenkunde 71: 609–616.
BRABEC, J., R. KUCHTA, AND T. SCHOLZ. 2006. Paraphyly of the Pseu-dophyllidea (Platyhelminthes: Cestoda): Circumscription of mono-phyletic clades based on phylogenetic analysis of ribosomal RNA.International Journal for Parasitology 36: 1535–1541.
BRAY, R. A., A. JONES, AND K. I. ANDERSEN. 1994. Order Pseudophyl-lidea Carus, 1863. In Keys to the cestode parasites of vertebrates,L. F. Khalil, A. Jones, and R. A. Bray (eds.). CAB International,Wallingford, U.K., p. 205–247.
———, ———, AND E. P. HOBERG. 1999. Observations of the phylog-eny of the cestode order Pseudophyllidea Carus, 1863. SystematicParasitology 42: 13–20.
CAIRA, J. N., AND D. T. J. LITTLEWOOD. 2001. Worms, Platyhelminthes.In Encyclopedia of biodiversity, S. A. Levin (ed.). Academic Press,San Diego, California, p. 863–899.
CIELECKA, D., B. GRYTNER-ZIECINA, AND L. CHOMICZ. 1994. Studies onthe surface ultrastructure of Sobolevicanthus gracilis (Zeder, 1803)(Cestoda, Hymenolepididae). Acta Parasitologica 39: 131–137.
CZAPLINSKI, B., A. AESCHLIMANN, AND D. SZELENBAUM-CIELECKA. 1984.Scanning electron microscopy of the cirrus surface of some Hy-menolepididae (Cestoda). Acta Parasitologica Polonica 29: 59–62.
DAVYDOV, V. G., AND Z. KORNEVA. 2002. Structure of the copulativeapparatus of Sobolevicanthus gracilis (Cestoda: Cyclophyllidea).Parazitologiya 36: 224–230.
———, L. G. PODDUBNAYA, AND G. A. KOLESNIKOVA. 1994. Ultrastruc-ture of genital system ducts of Caryophyllaeus laticeps (Cestoda,Caryophyllidea). Parazitologiya 28: 501–509.
———, ———, AND B. I. KUPERMAN. 1997. An ultrastructure of some
systems of the Diplocotyle olrikii (Cestoda: Cyathocephalata) inrelation to peculiarities of its life cycle. Parazitologiya 31: 132–141.
HALTON, D. 1997. Nutritional adaptations to parasitism within the Platy-helminthes. International Journal for Parasitology 27: 693–704.
HENDOW, H. T., AND B. L. JAMES. 1988. The ultrastructure of the malereproductive ducts in Maritrema linguilla (Digenea: Microphalli-dae). International Journal for Parasitology 18: 221–229.
HOBERG, E. P., A. JONES, AND R. A. BRAY. 1999. Phylogenetic analysisamong the families of the Cyclophyllidea (Eucestoda) based oncomparative morphology, with new hypotheses for co-evolution invertebrates. Systematic Parasitology 42: 51–73.
———, J. MARIAUX, AND D. R. BROOKS. 2001. Phylogeny among ordersof the Eucestoda (Cercomeromorphae): Integrating morphology,molecules and total evidence. In Interrelationships of the Platyhel-minthes, D. T. J. Littlewood and R. A. Bray (eds.). The SystematicAssociation Special Volume Series 60, Taylor & Francis, London,U.K., p. 122–126.
———, ———, J. L. JUSTINE, D. R. BROOKS, AND P. J. WEEKES. 1997.Phylogeny of the orders of the Eucestoda (Cercomeromorphae)based on comparative morphology: Historical perspectives and anew working hypothesis. Journal of Parasitology 83: 1128–1147.
IVANOV, V. A., AND E. P. HOBERG. 1999. Preliminary comments on aphylogenetic study of the order Diphyllidea van Beneden in Carus,1863. Systematic Parasitology 42: 21–27.
JONES, M. K. 1989. Ultrastructure of the cirrus pouch of Cylindrotaeniahickmani (Jones, 1985) (Cestoda, Nematotaeniidae). InternationalJournal for Parasitology 19: 919–930.
———. 1998. Structure and diversity of cestode epithelia. InternationalJournal for Parasitology 28: 913–923.
———, AND I. BEVERIDGE. 1998. Nybelinia queenslandensis sp. n. (Ces-toda: Trypanorhyncha) parasitic in Carcharhinus melanopterus,from Australia, with observations on the fine structure of the scolexincluding the rhyncheal system. Folia Parasitologica 45: 295–311.
———, AND ———. 2001. Echinobothrium chisholmae n. sp. (Cesto-da, Diphyllidea) from the giant shovel-nose ray Rhinobatos typusfrom Australia, with observations on the ultrastructure of its scolexmusculature and peduncular spines. Systematic Parasitology 50:41–52.
JUSTINE, J. L. 1998. Spermatozoa as phylogenetic characters for theEucestoda. Journal of Parasitology 84: 385–408.
KHALIL, L. F., A. JONES, AND R. A. BRAY. 1994. Keys to the cestodeparasites of vertebrates. CAB International, Wallingford, U.K., 447 p.
KORNEVA, Z. 2002. Ultrastructural organization of reproductive systemin Triaenophorus nodulosus (Cestoda, Pseudophyllidea). Zoologi-scheskii Zhurnal 81: 1432–1438.
KUCHTA, R. 2007. Revision of the paraphyletic ‘‘Pseudophyllidea’’ (Eu-cestoda) with description of two new orders Bothriocephalidea andDiphyllobothriidea. Ph.D. Thesis. Faculty of Biological Sciences,University of South Bohemia, Ceske Budejovice, 97 p.
———, AND T. SCHOLZ. 2007. Diversity and distribution of fish tape-worms of the ‘‘Bothriocephalidea’’ (Eucestoda). Parassitologia 49:129–146.
———, ———, J. BRABEC, AND R. A. BRAY. 2008. Suppression of thetapeworm order Pseudophyllidea (Platyhelminthes: Eucestoda) andthe proposal of two new orders, Bothriocephalidea and Diphyllo-bothriidea. International Journal for Parasitology 38: 49–55.
LEVRON, C., L. G. PODDUBNAYA, R. KUCHTA, M. FREEMAN, Y.-H. WANG,AND T. SCHOLZ. 2008. Ultrastructure of the tegument of the cestodeParaechinophallus japonicus (Pseudophyllidea: Echinophallidae),a parasite of the bathypelagic fish Psenopsis anomala. InvertebrateBiology. 127: 153–161.
MATEO, E., AND W. L. BULLOCK. 1966. Neobothriocephalus aspinosusgen. et sp. n. (Pseudophyllidea: Parabothriocephalidea), from thePeruvian marine fish Neptomenus crassus. Journal of Parasitology52: 1070–1073.
MOUNT, P. M. 1970. Histogenesis of the rostellar hooks of Taenia cras-siceps (Zeder, 1800) (Cestoda). Parasitology 56: 947–961.
OLSON, P. D., D. T. J. LITTLEWOOD, R. A. BRAY, AND J. MARIAUX. 2001.Interrelationships and evolution of the tapeworms (Platyhelminthes:Cestoda). Molecular Phylogenetics and Evolution 19: 443–467.
———, AND V. V. TKACH. 2005. Advances and trends in the molecular

810 THE JOURNAL OF PARASITOLOGY, VOL. 94, NO. 4, AUGUST 2008
systematics of the parasitic Platyhelminthes. Advances in Parasi-tology 60: 165–243.
PODDUBNAYA, L. G. 2002. Ultrastructure of reproductive ducts in Di-phyllobothrium latum (Cestoda, Pseudophyllidea) males. Zoologi-scheskii Zhurnal 81: 394–405.
———. 2003. Structure of reproductive system of the amphicotylidecestode Eubothrium rugosum (Cestoda, Pseudophyllidea). Journalof Evolutionary Biochemistry and Physiology 39: 345–355.
———, J. S. MACKIEWICZ, M. BRUNANSKA AND B. S. DEZFULI. 2005.Fine structure of the male reproductive ducts, vagina and seminalreceptacle of Cyathocephalus truncatus (Cestoda: Spathebothri-idea). Folia Parasitologica 52: 241–250.
PROTASOVA, E. N. 1977. Bothriocephalata — Cestodes of fish. Princi-ples of cestodology, Volume VIII. Nauka, Akademia Nauk SSSR,Moscow, Russia, 298 p.
REYNOLDS, E. S. 1963. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. Journal of Cell Biology 17:208–212.
SCHMIDT, G. D. 1986. CRC handbook of tapeworm identification. CRCPress, Boca Raton, Florida, 675 p.
STOITSOVA, S. R., B. B. GEORGIEV, G. P. VASILEVA, AND M. JORDANOVA.
2001. Rostellar apparatus of Fernandezia spinosissima (von Lin-stow, 1894) (Cestoda, Cyclophyllidea, Davaineidae): Microanato-my and fine structure. Acta Zoologica 82: 241–250.
SWIDERSKI, Z., J. MIQUEL, D. MłOCICKI, B. B. GEORGIEV, C. EIRA, B.GRYTNER-ZIECINA, AND C. FELIU. 2007. Post-embryonic develop-ment and ultrastructural characteristics of the polycephalic larva ofTaenia parva (Baer, 1926) (Cyclophyllidea, Taeniidae). Acta Par-asitologica 52: 31–50.
WAESCHENBACH, A., B. L. WEBSTER, R. A. BRAY, AND D. T. J. LITTLE-WOOD. 2007. Added resolution among ordinal level relationships oftapeworms (Platyhelminthes: Cestoda) with complete small andlarge subunit nuclear ribosomal RNA genes. Molecular Phyloge-netics and Evolution 45: 311–325.
YAMAGUTI, S. 1934. Studies on the helminth fauna of Japan. Part. 4.Cestodes of fishes. Japenese Journal of Zoology 6: 1–112.
ZD’ARSKA, Z, AND J. NEBESAROVA. 2003. Ultrastructure of the early ros-tellum of Silurotaenia siluri (Batsch, 1786) (Cestoda: Proteoce-phalidae). Parasitology Research 89: 495–500.
———, AND T. N. SOBOLEVA. 1990. Scanning electron microscopic in-vestigation of the trematode Hasstilesia ochotonae Gvosdev, 1962.Folia Parasitologica 37: 347–348.









![[Compared parasitic infection of Ligula intestinalis (Cestoda: Diphyllobothridae) in Cyprinidae species: Rutilus rubilio and Scardinius erythrophthalmus in two dam reserves in Tunisia]](https://img.pdfslide.net/doc/110x75/634ec2490d6acb880b02f6c2/compared-parasitic-infection-of-ligula-intestinalis-cestoda-diphyllobothridae.jpg)



















