Embed Size (px)
Citation preview

617
JPP 2003 55 617ndash630szlig 2003 The AuthorsReceived October 15 2002Accepted February 4 2003DOI 1012110022357021125ISSN 0022-3573
Department of PharmaceuticalSciences College of Pharmacyand Center for IntegratedBiotechnology WashingtonState University PullmanWashington USA
X W Teng N M Davies
Faculty of Pharmacy Universityof Sydney Sydney New SouthWales Australia
D J Cutler
Correspondence N M DaviesWashington State UniversityCollege of PharmacyDepartment of PharmaceuticalSciences PO Box 646534Pullman WA 99164-6534 USAE-mail ndavieswsuedu
Acknowledgements The authorswould like to thank Schering-Plough Australia Pty Ltd(Baulkham Hills NSW Australia)for a donation of mometasonefuroate and the assistance ofMr Bruce Tattam andDr A Cheung in chemicalanalysis
Kinetics of metabolism and degradation ofmometasone furoate in rat biological fluids and tissues
X W Teng D J Cutler and N M Davies
Abstract
Mometasone furoate (MF) is a potent glucocorticoid developed for the treatment of glucocorticoid-
responsive inflammatory disorders The in-vitro and ex-vivo kinetics of the degradation and meta-
bolism of MF were studied in selected biological fluids of rat and subcellular fractions of different rat
tissues In-vitro MF was found to degrade slowly into four products in serum and urine and
metabolized rapidly and extensively in rat liver minimally in extrahepatic tissues including intestine
stomach lung and kidney Further investigation found that the microsomal fraction was the major
intracellular site of MF 6shy -hydroxylation in rat liver Using chemical inhibitors CYP3A was found to
be the major enzyme involved in the in-vitro MF 6shy -hydroxylation in rat liver microsomes Enzyme
kinetic studies in rat liver microsomes showed that the overall metabolic process of MF followed
biphasic Michaelis-Menten kinetics while 6shy -hydroxylation obeyed monophasic Michaelis-Menten
kinetics The kinetic parameters derived from the kinetic models along with the enzyme inhibition
studies suggest that MF is mainly metabolized via 6shy -hydroxylation mediated by CYP3A primarily
and also biotransformed via other pathway(s) catalysed by other enzymes in rat liver in-vitro
Introduction
Mometasone furoate (MF) is a potent glucocorticoid with an improved ratio betweenlocal to systemic effects It has been marketed as a dermatological formulation(Elocon) an aqueous intranasal spray (Nasonex) and a dry-powder inhaler(Asmanex Twisthaler) for the treatment of glucocorticoid-responsible dermatoses(Prakash and Benfield 1998) and airway allergic inflammatory disorders (Onrust andLamb 1998) After oral (Wang et al 1992) dermatological (Prakash amp Benfield 1998)intranasal (Onrust amp Lamb 1998) or inhalational (Affrime et al 2000a) administrationof MF in humans its systemic exposure is negligible and the bioavailability is very low(lt1) Following the administration of MF via a metered-dose inhaler the deposi-tion pattern of MF is consistent with the 10plusmn20 pulmonary and 80 oropharynxdeposition demonstrated with other corticosteroids delivered in the same manner(Pickering et al 2000) It has been postulated that the extremely low bioavailability ofinhaled MF is owing to extensive metabolism of the drug in the liver following itsabsorption through respiratory and gastrointestinal tracts (Affrime et al 2000b)However to date there is limited information available on the metabolism of MFAn abstract reported that MF was metabolized extensively in the liver to some extentin the intestine but not in the lungs of different species in-vitro including rat mousedog and human with 6shy -hydroxy-MF suggested to be the major metabolite in ratwhile other moderately polar metabolites are also present in mouse dog and human(Zbaida et al 1997) It has not been known if metabolites of MF are active or inactiveThe most recent review on MF (Sharpe amp Jarvis 2001) quoted both published andunpublished results and stated that MF is metabolized extensively into polar com-pounds by cytochrome P450 (P450) 3A4 in the liver but no evidence of a single majormetabolite and no metabolites have been unequivocally identified Thus these pre-vious statements on metabolite(s) of this drug remain unclear
The purpose of the present studies was to qualitatively and quantitatively investi-gate the in-vitro and ex-vivo kinetics of degradation and metabolism of MF in a rat
model The specific objectives of the present studies were(i) to determine the major site(s) of MF metabolism inrats (ii) to investigate the contribution of microsomalenzymes towards the biotransformation of MF in rats(iii) to analyse the enzyme kinetics of MF in rat livermicrosomes and develop kinetic models describing themetabolism of MF in rats and (iv) to isolate and purifythe major degradation and metabolic products fromsuitable in-vitro systems in rats for its further pharma-cological study
Materials and Methods
Materials
Authentic MF was a gift from Schering-Plough Pty Ltd(Baulkham hills NSW Australia) Testosterone 17-acet-ate halothane Krebs-Henseleit buffer total proteinreagent protein standard solution monosodium glucose-6-phosphate shy -nicotinamide adenine dinucleotide phosphate(shy -NADP) sodium salt hydrate glucose-6-phosphatedehydrogenase HEPES (N-[2-hydroxyethyl] piperazine-N0-[2-ethane sulfonic acid]) 1-aminobenzotriazole fura-fylline coumarin sulfaphenazole diazepam quinidinediethyldithiocarbamate 4-methylpyrazole ketoconazoleand troleandomycin were purchased from SigmaChemical (St Louis MO USA) Medical carbogen gas(containing 5 carbon dioxide and 95 oxygen) andhigh purity nitrogen gas were purchased from BOCGases (Chatswood NSW Australia) Carbon monoxidewas obtained from Matheson Tri-Gas Inc (Newark CAUSA) All other chemicals and solvents were of analyticalor HPLC grade as appropriate
Animals and collection of biological fluidsand tissues
Adult male Sprague-Dawley rats 200plusmn250 g were sup-plied by the Laboratory Animal Services Center at theUniversity of Sydney The rats were allowed free accessto standard laboratory chow (Purina Rat Chow RalstonSydney NSW Australia) and tap water and weremaintained in a restricted access room with controlledtemperature (21 sect 1 macrC) humidity and a 12-h lightdarkcycle The experimental animal protocols were approvedby the Animal Ethics Committee of the University ofSydney
Rat urine was collected from individual rats housed inmetabolic cages (Technoplast Rochester NY USA) for0plusmn24 h After the rats were anaesthetized with 5 (vv)halothane the blood was collected via terminal cardiacpuncture Subsequently a vertical midline abdominal inci-sion was made and the tissues required were rapidlyexcised examined and immediately placed in ice-cold10 mM Tris-buffered saline (pH 75) Blood was allowedto clot and then centrifuged to yield serum which wasstored at iexcl20 macrC The pH of the pooled rat serum was736 measured before the kinetic study
Preparation of tissue subcellular fractions
Rat tissues were homogenized using a Polytron PTA 1035homogenizer (Kinematica GmbH CH-6010 KriensLuzerne Switzerland) and the 1000 g supernatant (S-1)9000 g supernatant (S-9) cytosolic and microsomal frac-tions of individual tissue were prepared by differentialcentrifugation (Ekins et al 1999) Pooled nuclear mito-chondrial cytosolic and microsomal fractions of rat livertissue (n ˆ 6) were also prepared by differential centrif-ugation (Pedersen et al 1978) All the homogenizationand fractionation processes were conducted in a coldroom at iexcl4 macrC The subcellular fractions of tissues werestored at iexcl80 macrC before use Total protein contents of thefractions of tissue homogenates were determined spectro-photometrically at 540 nm (Doumas et al 1981) Bovineserum albumin was used as the reference standard TheP450 content was determined as described previously(Ocuma amp Sato 1964)
Instrumentation and analytical methods
HPLC analysis of MF and its degradation and metabolicproducts was performed as previously described (Tenget al 2001) The Shimadzu Class-LC10 HPLC (KyotoJapan) system consisted of an LC-10AT pump a SIL-10AXL autoinjector an FRC-10A fraction collector anSPD-M10A photodiode-array UVvis spectrophotometricdetector and a CBM-10A communication bus moduleData collection and integration were accomplished usingShimadzu Class-LC10 computer software version 164(Kyoto Japan) The analytical column used was aBeckman ultrasphere octyl column (150 mm 20 mm id5 middotm particle size Beckman Instruments Fullerton CAUSA) equipped with a pre-column (75 mm 20 mm id)of the same packing material The mobile phase consistedof methanol and water (5941 vv) filtered and degassedusing a Millipore filtration system through a 045-middotmHNWP nylon filter (Bedford MA USA) under reducedpressure before use Separation was carried out isocratic-ally at room temperature (22 sect 1 macrC) and with flow rate of028 mL miniexcl1 with UV detection at 248 nm and spectralscanning from 200 to 400 nm The injection volume was10 middotL
Kinetics of MF in rat serum and urine
MF (C0 ˆ 19 middotM added as a methanolic stock solution of20 mg mLiexcl1 at a volume of 05 in the final incubationmixtures) was incubated in rat urine and in rat serum at370 sect 01 macrC in a thermostatically controlled shakingwater bath as described previously (Teng et al 2001)
Kinetics of MF in rat tissue S-9 fractions
Studies of metabolic kinetics of MF in S-9 fractions oftissue homogenates were conducted in the presence ofcofactors that included 10 mM MgCl2 and an NADPH-generating system (75 mM glucose 6-phosphate 03 mM
shy -NADP and 042 units mLiexcl1 glucose-6-phosphate
618 X W Teng et al
dehydrogenase) in 100 mM phosphate buffer containing1 mM EDTA (pH 74) under carbogen gas at 370 sect 01 macrCin a shaking (75 rev miniexcl1) water bath Typically the S-9fraction (protein content 4 mg mLiexcl1 unless otherwiseindicated) and the parent drug (C0 ˆ 19 middotM added as amethanolic stock solution of 20 mg mLiexcl1 at a volume of05 in the final incubation mixtures) were pre-incubatedin the incubation buffer for 5 min at 37 sect 01 macrC Thereaction was initiated by adding the cofactors At pre-determined time intervals samples (05 mL each) werewithdrawn and the reaction was terminated immediatelyby adding 4 mL ice-cold dichloromethane Samples werethen extracted and analysed by HPLC (Teng et al 2001)Parallel incubations were performed in the same mannerin the absence of MF or the S-9 fraction as controlsExperiments with the tissue S-9 fractions were performedin four replicates
The metabolic kinetics of MF in pooled rat liver tissueS-9 fractions was investigated with respect to protein con-centration (range 1plusmn8 mg mLiexcl1) and substrate concentra-tion (range 1plusmn100 middotM)
Studies of degradation kinetics of MF (C0 ˆ 19 middotMadded as a methanolic stock solution of 20 mg mLiexcl1 ata volume of 05 in the final incubation mixtures) in eachS-9 fraction of the rat tissues were carried out withoutcofactors The reaction was initiated by adding the stockMF in the reaction media and then handled using thesame method
Kinetics of MF in rat liver microsomal cytosolicmitochondrial and nuclear fractions
Initially the microsomal incubation conditions wereinvestigated with respect to microsomal protein concen-tration (01plusmn5 mg mLiexcl1) incubation time (up to 60 min)substrate concentration (02plusmn100 middotM) and oxygenationusing pooled rat liver microsomes The optimizedmicrosomal incubation mixture was composed of livermicrosomes (02 mg protein mLiexcl1) 25 middotM MF and anNADPH-generating system (6 mM MgCl2 05 mM shy -NADP10 mM glucose 6-phosphate and 10 units mLiexcl1 glucose-6-phosphate dehydrogenase) in 100 mM phosphate buffercontaining 1 mM EDTA (pH 74) The reaction was car-ried out in open test tubes without extra oxygenation for8 min in a shaking (150 rev miniexcl1) water bath After themicrosomal fraction and the parent drug (added as amethanolic stock solution at a volume of 1 in the finalincubation mixtures) were pre-warmed in the incubationbuffer for 5 min at 37 sect 01 macrC the reaction was initiatedby the addition of the NADPH-generating system andterminated by addition of 4 mL ice-cold dichloromethaneSimilar incubations with the use of denatured microsomes(heating at 70plusmn80 macrC for 30 min) or without the addition ofmicrosomes or substrate were conducted in the same man-ner as controls In addition incubation in the absence ofthe NADPH-generating system was performed for theconfirmation of P450-dependent enzymatic biotransfor-mation Typically all the incubations of MF (C0 ˆ 4 middotM)with microsomal cytosolic mitochondrial and nuclear
fractions (protein content 4 mg mLiexcl1) were performedfollowing the same procedure Incubations for each indi-vidual condition with these subcellular fractions of ratliver tissue were conducted in duplicate
Kinetics in everted rat small intestine tissue
A modified version of the method of Esposito amp Csaky(1974) was employed in these studies After anaesthetiza-tion of the rat with halothane an abdominal incision wasmade and the whole small intestine was isolated excisedand examined After a section (sup110 cm) of intestine at theend of the ileocecal junction was taken for tissue sampleof zero time the intestine was immediately flushed withice-cold Krebs-Henseleit bicarbonate buffer containing 2 gLiexcl1 glucose (pH 74 K-H buffer) and then everted ona glass rod with one end tied with surgical thread After thesac was filled with warmed drug-free K-H buffer (37 macrC)the other end was tied and the middle part of the intestinewas tied every 20plusmn25 cm to give four isolated sections Thiseverted intestine was immediately placed in a freshly pre-pared warmed (37 macrC) and jacketed beaker containing25 mL of 19 middotM MF (10 middotg mLiexcl1 added as a methanolicstock solution of 20 mg mLiexcl1 at a volume of 05 in thefinal incubation mixtures) in K-H buffer and incubated at37 sect 01 macrC with continual carbogenation with carbogen ina shaking water bath (75 rev miniexcl1)
The zero-time buffer sample (the K-H buffer withdrug) was withdrawn before the tissue was placed in theK-H buffer and the drug-free K-H buffer was used as thezero-time buffer sample in the sac At 05 15 3 and 5 h3 mL of buffer in the bulk incubation medium and asection of the intestine (about 2 g) with the buffer in itslumen were sampled After the buffer in the lumen wastransferred into a beaker the tissue segment was weighedand then immediately added to ice-cold K-H buffer ofequal weight Subsequently the tissue was minced homo-genized extracted and analysed by HPLC
Buffer samples (05 mL) from both the bulk incubationmedium and the intestinal lumen were also collectedfor analysis of MF and its degradation and metabolicproducts as described for samples of tissue subcellularfractions The experiment was performed in triplicate
Inhibition studies
A number of potential inhibitors andor substrates ofCYP isoenzymes were tested at various concentrationsin pooled rat liver microsomes (protein content 02 mgmLiexcl1) with varying concentrations of MF (added as astock methanolic solution at a volume of 1 in the finalincubation mixtures) Incubation mixtures containingmechanism inhibitors furafylline diethyldithiocarba-mate troleandomycin or 1-aminobenzotriazole werepre-incubated in pooled rat liver microsomes in the pre-sence of the NADPH-generating system (6 mM MgCl205 mM shy -NADP 10 mM glucose-6-phosphate and 10units mLiexcl1 glucose-6-phosphate dehydrogenase) at37 sect 1 macrC for 15 min before the addition of MF Fornon-mechanism-based inhibitors the incubation mixtures
Rat metabolism and degradation kinetics of mometasone furoate 619
contained MF and inhibitors together with rat liver micro-somes without pre-incubation of inhibitors and thereactions were initiated by the addition of the NADPH-generating system and carried out as for rat liver micro-somes Potential inhibitors were dissolved in a mixture ofmethanol and phosphate buffer except for ketoconazolewhich was dissolved in acetone and added at a volume of1 in the final incubation mixtures Parallel controlincubations were conducted with 1 of the correspondingsolvent with no inhibitor present Each potential inhibitorwas also incubated with microsomes under the NADPH-generating system to examine if they caused any chro-matographic interference in the measurement of6shy -hydroxy-MF
Sample extraction and analysis
For tissue and sub-cellular fractions on terminating thereaction with 40 mL ice-cold dichloromethane 05 mL ofinternal standard (10 middotg mLiexcl1 of ethanolic testosterone17-acetate) was added to the sample (05 mL) The mixturewas extracted for 2 min on a vibrating mixer followed bycentrifugation at 1328 g (20 macrC) for 10 min The organiclayer was then collected evaporated to dryness undernitrogen at 35 macrC stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
To each sample (05 g) of tissue homogenate was added05 mL of internal standard (10 middotg mLiexcl1 ethanolic testo-sterone 17-acetate) The mixture was extracted twice with4 mL dichloromethane by vigorous shaking for 2 min ona vibrating mixer followed by centrifugation at 2602 g(20 macrC) for 10 min The dichloromethane phase was col-lected evaporated stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
Data analysis
Data are expressed as the mean sect sd of replicate deter-minations (n 5 5) Statistical analyses were performedusing Microsoft Excel (version 97) or SPSS (Spss IncIL USA) Significance of differences in means of pairedsamples was assessed using a paired-sample t-testAnalysis of variance of two or more groups of data wasperformed using analysis of variance and Tukeyrsquos post-hoc analysis All values of P were based on two-tailed testsand P values of less than 005 were considered statisticallysignificant
The observed rate constants (kobs) of parent drug dis-appearance were estimated from the initial slope of thelog-linear phase of declining concentration versus timeplots The initial half-lives were calculated using the fol-lowing equation t1curren2 ˆ 0693kobs The rate constant ofeach metabolic reaction was estimated according to thecorresponding model developed based on the experi-mental data to limit underestimation of the parametersdue to possible sequential transformation The rate con-stant of parent drug disappearance was correlated to thesum of those of its direct transformation reactions
Substrate concentrations ranging from 02 to 100 middotM
were employed for MF in view of its unknown in-vitro
metabolic kinetics in rat liver microsomes Kinetic analysisof the parent drug disappearance and product formationdata was undertaken by the least-square regression programusing SCIENTIST version 20 (Micromath ScientificSoftware Salt Lake City UT USA) Apparent enzymekinetics was determined by fitting reaction velocity (v) versussubstrate concentration [MF] Initially the data were exam-ined on Eadie-Hofstee Lineweaver-Burk and Hanes plots toassess the linearity of enzyme kinetics The kinetic data werethen fitted to expressions containing one or more Michaelis-Menten terms The correlation randomness of the residualsthe standard deviation of the parameter estimates (lt10)and the model selection were used as indicators of goodness-of-fit of the model to the experimental data
The nature of inhibition of the chemical inhibitors forP450 was initially assessed by plots from transformed datafrom the kinetic studies including Dixon plotLineweaver-Burk plot Eadie-Hofstee plot and Cornish-Bowden plot For competitive enzyme inhibitors theapparent inhibitory constant (Ki) was given by the inter-section point of linear regression lines in the Dixon plotwhere data sets of 1v were plotted against the initialconcentration of the inhibitor [I] Results were sub-sequently subjected to Equation 1 (Ito et al 1998) byleast-square regression using SCIENTIST
cedil ˆ Vmax cent permilMFŠ
Km cent 1 Dagger permilIŠKi
8gt
9gtDagger permilMFŠ
hellip1dagger
where Km represents the Michaelis constant of the enzy-matic reaction (cf Equation 5) [MF] is the initial drugconcentration and Vmax indicates the maximum velocityof the reaction
Kinetic parameters for mechanism-based enzyme inac-tivation were obtained as described by Ito et al (1998) Thelogarithm of the remaining enzymatic activity (formationrate of 6shy -hydroxy-MF) was plotted against the pre-incu-bation time in the presence of the inhibitor at variousconcentrations The apparent inactivation rate constant(kiobs) was determined from the slope of the initial linearphase at each inhibitor concentration The apparent kiobs
values were compared with those of control incubationscontaining the solvent for each inhibitor as appropriateThe values of kiobs were plotted against the inhibitorconcentrations and the parameters (kimax and K0
i) wereestimated from Equation 2 by a non-linear least squaremethod using SCIENTIST
kiobs ˆ kimax cent permilIŠK0
i Dagger permilIŠhellip2dagger
where kiobs is the apparent inactivation rate constant ofthe enzyme at initial inhibitor concentration [I] kimax
represents the maximum inactivation rate constantand K0
i represents the apparent dissociation constantbetween the enzyme and the inhibitor respectively(Ito et al 1998)
620 X W Teng et al
The IC50 (the concentration inhibiting 50 of enzymeactivity) was determined by linear regression (percentage ofcontrol) against the logarithmic inhibitor concentration
Results
Degradation in rat serum
MF (C0 ˆ 19 middotM) degraded in rat serum (pH 736) with ahalf-life of 80 sect 01 h (n ˆ 4) Four products namely AB C and D (Figure 1) were formed in rank order and arethe same as observed previously in human plasma and insimulated lung fluid (Teng et al 2001 products A B Cand D reported here correspond to products D1 D3D2 and D4 reported in Teng et al 2001) They were iden-tified as A 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione 17-(2-furoate) B 9not21shy -dichloro-11shy 21not-dihydroxy-16not-methylpregna-1417 20-tetraen-3-one 21-(2-furoate) C 21shy -chloro-21 not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-141720-tetraen-3-one 21-(2-furoate) and D 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione (Teng 2002)
Degradation in rat urine
MF (C0 ˆ 19 middotM) degraded in rat urine (pH 783) with a half-life of 111 sect 03 h (n ˆ 4) The degradation profile of MF inrat urine was qualitatively similar to that in rat serum
Metabolism of MF in S-9 fractions of rat tissues
MF (C0 ˆ 19 middotM) decomposed quickly with a half-life of233 sect 107 min (n ˆ 6) and rapid formation of 6shy -hydroxy-MF in the rat liver tissue S-9 fractions of proteincontent 4 mg mLiexcl1 The formation of 6shy -hydroxy-MFwas not observed in control samples in which the parentdrug or the S-9 fractions were absent In rat liver tissue S-9fraction the apparent rate of MF 6shy -hydroxylation in thepresence of cofactors was over 1000-times greater than thatin the absence of cofactors indicating that this reaction waslargely catalysed by enzymes dependent on cofactors
The apparent rate of MF (C0 ˆ 19 middotM) disappearanceand formation of 6shy -hydroxy-MF increased with anincrease in total protein concentration of rat liver tissueS-9 fractions from 1 to 8 mg mLiexcl1 On the other hand theapparent rates of MF disappearance and 6shy -hydroxy-MFformation were both enhanced with an increase in the initialconcentration of MF from 1 to 50 middotM but there was nofurther increase at 100 middotM when the total protein of rat livertissue S-9 fraction was kept constant at 4 mg mLiexcl1
In addition the relative activities of the S-9 fractions ofthe tested extrahepatic tissues catalysing MF 6shy -hydroxyla-tion were less than 1 of that of the rat liver tissue In theseS-9 fractions the apparent rates of MF 6shy -hydroxylationwere in the order of intestine 5 stomach gt spleen gt kid-ney gt lung In the rat lung tissue S-9 fraction 6shy -hydroxy-MF was detectable but not quantifiable in about half of thesamples using the HPLCUV assay
In the presence of cofactors the rate of degradation ofMF through the formation of products A B C and D wasmuch slower than in the corresponding tissue S-9 fractionswithout cofactors (P lt 001 paired-sample t-test) In factonly product A (not B C or D) was detectable duringincubation up to 72 h in all S-9 fractions of tissues testedwith cofactors
Metabolism in rat liver subcellular fractions
MF was metabolized rapidly in rat liver microsomes with6shy -hydroxy-MF as a primary metabolite (Figure 2) underan NADPH-generating system No significant transform-ation of MF to 6shy -hydroxy-MF was detected in thecontrols As in the S-9 fraction of rat liver tissue noneof the other potential metabolites listed by Affrime et al(2000a) were observed except for 6shy -hydroxy-MF
In pooled liver microsomes from six rats the apparentrate of both parent drug disappearance and 6shy -hydroxy-MF appearance increased with an increase in proteinconcentration over the range of 01 to 5 mg mLiexcl1 whenan initial concentration of MF was kept at 4 middotM
When total protein of rat liver microsomes was keptconstant at 02 mg mLiexcl1 the apparent rate of parent drugdisappearance (Figure 3) increased with an increase in theinitial drug concentration from 02 to 65 middotM The declineof MF followed first-order degradation kinetics in rat livermicrosomes of protein content 02 mg mLiexcl1 The rate of6shy -hydroxy-MF formation increased correspondinglywith increasing initial drug concentration from 02 to8 middotM However the formation was saturated at higherconcentrations reaching a plateau An optimal incubationtime of 8 min was selected as a convenient common incu-bation time for studying the kinetics of MF metabolism inrat liver microsomes of protein content 02 mg mLiexcl1
Little formation of 6shy -hydroxy-MF and disappearanceof the parent drug were observed following the incubationof MF (C0 ˆ 4 middotM) in pooled rat liver cytosolic mitochon-drial and nuclear fractions of protein content 4 mg mLiexcl1
at 37 macrC (Table 1) The enzyme activities of this cytosolicmitochondrial and nuclear fractions catalysing 6shy -hydro-xylation of MF were 04 43 and 16 of that of pooledrat liver microsomes respectively
Transformation in everted rat intestine
Following the incubation of MF (C0 ˆ 19 middotM) with evertedintestinal tissue in K-H buffer MF disappeared rapidlyfrom the incubation buffer with considerable formation of6shy -hydroxy-MF as the major metabolite (Figure 4)Meanwhile most of the parent drug and the metaboliteformed were taken up by the intestinal tissue during incu-bation
Inhibition of MF 6b-hydroxylation by potentialchemical inhibitors
A non-specific CYP inhibitor 1-aminobenzotriazole(Mathews et al 1985 Carcillo et al 1998) inhibited MF6shy -hydroxylation in pooled rat liver microsomes in a
Rat metabolism and degradation kinetics of mometasone furoate 621
concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

model The specific objectives of the present studies were(i) to determine the major site(s) of MF metabolism inrats (ii) to investigate the contribution of microsomalenzymes towards the biotransformation of MF in rats(iii) to analyse the enzyme kinetics of MF in rat livermicrosomes and develop kinetic models describing themetabolism of MF in rats and (iv) to isolate and purifythe major degradation and metabolic products fromsuitable in-vitro systems in rats for its further pharma-cological study
Materials and Methods
Materials
Authentic MF was a gift from Schering-Plough Pty Ltd(Baulkham hills NSW Australia) Testosterone 17-acet-ate halothane Krebs-Henseleit buffer total proteinreagent protein standard solution monosodium glucose-6-phosphate shy -nicotinamide adenine dinucleotide phosphate(shy -NADP) sodium salt hydrate glucose-6-phosphatedehydrogenase HEPES (N-[2-hydroxyethyl] piperazine-N0-[2-ethane sulfonic acid]) 1-aminobenzotriazole fura-fylline coumarin sulfaphenazole diazepam quinidinediethyldithiocarbamate 4-methylpyrazole ketoconazoleand troleandomycin were purchased from SigmaChemical (St Louis MO USA) Medical carbogen gas(containing 5 carbon dioxide and 95 oxygen) andhigh purity nitrogen gas were purchased from BOCGases (Chatswood NSW Australia) Carbon monoxidewas obtained from Matheson Tri-Gas Inc (Newark CAUSA) All other chemicals and solvents were of analyticalor HPLC grade as appropriate
Animals and collection of biological fluidsand tissues
Adult male Sprague-Dawley rats 200plusmn250 g were sup-plied by the Laboratory Animal Services Center at theUniversity of Sydney The rats were allowed free accessto standard laboratory chow (Purina Rat Chow RalstonSydney NSW Australia) and tap water and weremaintained in a restricted access room with controlledtemperature (21 sect 1 macrC) humidity and a 12-h lightdarkcycle The experimental animal protocols were approvedby the Animal Ethics Committee of the University ofSydney
Rat urine was collected from individual rats housed inmetabolic cages (Technoplast Rochester NY USA) for0plusmn24 h After the rats were anaesthetized with 5 (vv)halothane the blood was collected via terminal cardiacpuncture Subsequently a vertical midline abdominal inci-sion was made and the tissues required were rapidlyexcised examined and immediately placed in ice-cold10 mM Tris-buffered saline (pH 75) Blood was allowedto clot and then centrifuged to yield serum which wasstored at iexcl20 macrC The pH of the pooled rat serum was736 measured before the kinetic study
Preparation of tissue subcellular fractions
Rat tissues were homogenized using a Polytron PTA 1035homogenizer (Kinematica GmbH CH-6010 KriensLuzerne Switzerland) and the 1000 g supernatant (S-1)9000 g supernatant (S-9) cytosolic and microsomal frac-tions of individual tissue were prepared by differentialcentrifugation (Ekins et al 1999) Pooled nuclear mito-chondrial cytosolic and microsomal fractions of rat livertissue (n ˆ 6) were also prepared by differential centrif-ugation (Pedersen et al 1978) All the homogenizationand fractionation processes were conducted in a coldroom at iexcl4 macrC The subcellular fractions of tissues werestored at iexcl80 macrC before use Total protein contents of thefractions of tissue homogenates were determined spectro-photometrically at 540 nm (Doumas et al 1981) Bovineserum albumin was used as the reference standard TheP450 content was determined as described previously(Ocuma amp Sato 1964)
Instrumentation and analytical methods
HPLC analysis of MF and its degradation and metabolicproducts was performed as previously described (Tenget al 2001) The Shimadzu Class-LC10 HPLC (KyotoJapan) system consisted of an LC-10AT pump a SIL-10AXL autoinjector an FRC-10A fraction collector anSPD-M10A photodiode-array UVvis spectrophotometricdetector and a CBM-10A communication bus moduleData collection and integration were accomplished usingShimadzu Class-LC10 computer software version 164(Kyoto Japan) The analytical column used was aBeckman ultrasphere octyl column (150 mm 20 mm id5 middotm particle size Beckman Instruments Fullerton CAUSA) equipped with a pre-column (75 mm 20 mm id)of the same packing material The mobile phase consistedof methanol and water (5941 vv) filtered and degassedusing a Millipore filtration system through a 045-middotmHNWP nylon filter (Bedford MA USA) under reducedpressure before use Separation was carried out isocratic-ally at room temperature (22 sect 1 macrC) and with flow rate of028 mL miniexcl1 with UV detection at 248 nm and spectralscanning from 200 to 400 nm The injection volume was10 middotL
Kinetics of MF in rat serum and urine
MF (C0 ˆ 19 middotM added as a methanolic stock solution of20 mg mLiexcl1 at a volume of 05 in the final incubationmixtures) was incubated in rat urine and in rat serum at370 sect 01 macrC in a thermostatically controlled shakingwater bath as described previously (Teng et al 2001)
Kinetics of MF in rat tissue S-9 fractions
Studies of metabolic kinetics of MF in S-9 fractions oftissue homogenates were conducted in the presence ofcofactors that included 10 mM MgCl2 and an NADPH-generating system (75 mM glucose 6-phosphate 03 mM
shy -NADP and 042 units mLiexcl1 glucose-6-phosphate
618 X W Teng et al
dehydrogenase) in 100 mM phosphate buffer containing1 mM EDTA (pH 74) under carbogen gas at 370 sect 01 macrCin a shaking (75 rev miniexcl1) water bath Typically the S-9fraction (protein content 4 mg mLiexcl1 unless otherwiseindicated) and the parent drug (C0 ˆ 19 middotM added as amethanolic stock solution of 20 mg mLiexcl1 at a volume of05 in the final incubation mixtures) were pre-incubatedin the incubation buffer for 5 min at 37 sect 01 macrC Thereaction was initiated by adding the cofactors At pre-determined time intervals samples (05 mL each) werewithdrawn and the reaction was terminated immediatelyby adding 4 mL ice-cold dichloromethane Samples werethen extracted and analysed by HPLC (Teng et al 2001)Parallel incubations were performed in the same mannerin the absence of MF or the S-9 fraction as controlsExperiments with the tissue S-9 fractions were performedin four replicates
The metabolic kinetics of MF in pooled rat liver tissueS-9 fractions was investigated with respect to protein con-centration (range 1plusmn8 mg mLiexcl1) and substrate concentra-tion (range 1plusmn100 middotM)
Studies of degradation kinetics of MF (C0 ˆ 19 middotMadded as a methanolic stock solution of 20 mg mLiexcl1 ata volume of 05 in the final incubation mixtures) in eachS-9 fraction of the rat tissues were carried out withoutcofactors The reaction was initiated by adding the stockMF in the reaction media and then handled using thesame method
Kinetics of MF in rat liver microsomal cytosolicmitochondrial and nuclear fractions
Initially the microsomal incubation conditions wereinvestigated with respect to microsomal protein concen-tration (01plusmn5 mg mLiexcl1) incubation time (up to 60 min)substrate concentration (02plusmn100 middotM) and oxygenationusing pooled rat liver microsomes The optimizedmicrosomal incubation mixture was composed of livermicrosomes (02 mg protein mLiexcl1) 25 middotM MF and anNADPH-generating system (6 mM MgCl2 05 mM shy -NADP10 mM glucose 6-phosphate and 10 units mLiexcl1 glucose-6-phosphate dehydrogenase) in 100 mM phosphate buffercontaining 1 mM EDTA (pH 74) The reaction was car-ried out in open test tubes without extra oxygenation for8 min in a shaking (150 rev miniexcl1) water bath After themicrosomal fraction and the parent drug (added as amethanolic stock solution at a volume of 1 in the finalincubation mixtures) were pre-warmed in the incubationbuffer for 5 min at 37 sect 01 macrC the reaction was initiatedby the addition of the NADPH-generating system andterminated by addition of 4 mL ice-cold dichloromethaneSimilar incubations with the use of denatured microsomes(heating at 70plusmn80 macrC for 30 min) or without the addition ofmicrosomes or substrate were conducted in the same man-ner as controls In addition incubation in the absence ofthe NADPH-generating system was performed for theconfirmation of P450-dependent enzymatic biotransfor-mation Typically all the incubations of MF (C0 ˆ 4 middotM)with microsomal cytosolic mitochondrial and nuclear
fractions (protein content 4 mg mLiexcl1) were performedfollowing the same procedure Incubations for each indi-vidual condition with these subcellular fractions of ratliver tissue were conducted in duplicate
Kinetics in everted rat small intestine tissue
A modified version of the method of Esposito amp Csaky(1974) was employed in these studies After anaesthetiza-tion of the rat with halothane an abdominal incision wasmade and the whole small intestine was isolated excisedand examined After a section (sup110 cm) of intestine at theend of the ileocecal junction was taken for tissue sampleof zero time the intestine was immediately flushed withice-cold Krebs-Henseleit bicarbonate buffer containing 2 gLiexcl1 glucose (pH 74 K-H buffer) and then everted ona glass rod with one end tied with surgical thread After thesac was filled with warmed drug-free K-H buffer (37 macrC)the other end was tied and the middle part of the intestinewas tied every 20plusmn25 cm to give four isolated sections Thiseverted intestine was immediately placed in a freshly pre-pared warmed (37 macrC) and jacketed beaker containing25 mL of 19 middotM MF (10 middotg mLiexcl1 added as a methanolicstock solution of 20 mg mLiexcl1 at a volume of 05 in thefinal incubation mixtures) in K-H buffer and incubated at37 sect 01 macrC with continual carbogenation with carbogen ina shaking water bath (75 rev miniexcl1)
The zero-time buffer sample (the K-H buffer withdrug) was withdrawn before the tissue was placed in theK-H buffer and the drug-free K-H buffer was used as thezero-time buffer sample in the sac At 05 15 3 and 5 h3 mL of buffer in the bulk incubation medium and asection of the intestine (about 2 g) with the buffer in itslumen were sampled After the buffer in the lumen wastransferred into a beaker the tissue segment was weighedand then immediately added to ice-cold K-H buffer ofequal weight Subsequently the tissue was minced homo-genized extracted and analysed by HPLC
Buffer samples (05 mL) from both the bulk incubationmedium and the intestinal lumen were also collectedfor analysis of MF and its degradation and metabolicproducts as described for samples of tissue subcellularfractions The experiment was performed in triplicate
Inhibition studies
A number of potential inhibitors andor substrates ofCYP isoenzymes were tested at various concentrationsin pooled rat liver microsomes (protein content 02 mgmLiexcl1) with varying concentrations of MF (added as astock methanolic solution at a volume of 1 in the finalincubation mixtures) Incubation mixtures containingmechanism inhibitors furafylline diethyldithiocarba-mate troleandomycin or 1-aminobenzotriazole werepre-incubated in pooled rat liver microsomes in the pre-sence of the NADPH-generating system (6 mM MgCl205 mM shy -NADP 10 mM glucose-6-phosphate and 10units mLiexcl1 glucose-6-phosphate dehydrogenase) at37 sect 1 macrC for 15 min before the addition of MF Fornon-mechanism-based inhibitors the incubation mixtures
Rat metabolism and degradation kinetics of mometasone furoate 619
contained MF and inhibitors together with rat liver micro-somes without pre-incubation of inhibitors and thereactions were initiated by the addition of the NADPH-generating system and carried out as for rat liver micro-somes Potential inhibitors were dissolved in a mixture ofmethanol and phosphate buffer except for ketoconazolewhich was dissolved in acetone and added at a volume of1 in the final incubation mixtures Parallel controlincubations were conducted with 1 of the correspondingsolvent with no inhibitor present Each potential inhibitorwas also incubated with microsomes under the NADPH-generating system to examine if they caused any chro-matographic interference in the measurement of6shy -hydroxy-MF
Sample extraction and analysis
For tissue and sub-cellular fractions on terminating thereaction with 40 mL ice-cold dichloromethane 05 mL ofinternal standard (10 middotg mLiexcl1 of ethanolic testosterone17-acetate) was added to the sample (05 mL) The mixturewas extracted for 2 min on a vibrating mixer followed bycentrifugation at 1328 g (20 macrC) for 10 min The organiclayer was then collected evaporated to dryness undernitrogen at 35 macrC stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
To each sample (05 g) of tissue homogenate was added05 mL of internal standard (10 middotg mLiexcl1 ethanolic testo-sterone 17-acetate) The mixture was extracted twice with4 mL dichloromethane by vigorous shaking for 2 min ona vibrating mixer followed by centrifugation at 2602 g(20 macrC) for 10 min The dichloromethane phase was col-lected evaporated stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
Data analysis
Data are expressed as the mean sect sd of replicate deter-minations (n 5 5) Statistical analyses were performedusing Microsoft Excel (version 97) or SPSS (Spss IncIL USA) Significance of differences in means of pairedsamples was assessed using a paired-sample t-testAnalysis of variance of two or more groups of data wasperformed using analysis of variance and Tukeyrsquos post-hoc analysis All values of P were based on two-tailed testsand P values of less than 005 were considered statisticallysignificant
The observed rate constants (kobs) of parent drug dis-appearance were estimated from the initial slope of thelog-linear phase of declining concentration versus timeplots The initial half-lives were calculated using the fol-lowing equation t1curren2 ˆ 0693kobs The rate constant ofeach metabolic reaction was estimated according to thecorresponding model developed based on the experi-mental data to limit underestimation of the parametersdue to possible sequential transformation The rate con-stant of parent drug disappearance was correlated to thesum of those of its direct transformation reactions
Substrate concentrations ranging from 02 to 100 middotM
were employed for MF in view of its unknown in-vitro
metabolic kinetics in rat liver microsomes Kinetic analysisof the parent drug disappearance and product formationdata was undertaken by the least-square regression programusing SCIENTIST version 20 (Micromath ScientificSoftware Salt Lake City UT USA) Apparent enzymekinetics was determined by fitting reaction velocity (v) versussubstrate concentration [MF] Initially the data were exam-ined on Eadie-Hofstee Lineweaver-Burk and Hanes plots toassess the linearity of enzyme kinetics The kinetic data werethen fitted to expressions containing one or more Michaelis-Menten terms The correlation randomness of the residualsthe standard deviation of the parameter estimates (lt10)and the model selection were used as indicators of goodness-of-fit of the model to the experimental data
The nature of inhibition of the chemical inhibitors forP450 was initially assessed by plots from transformed datafrom the kinetic studies including Dixon plotLineweaver-Burk plot Eadie-Hofstee plot and Cornish-Bowden plot For competitive enzyme inhibitors theapparent inhibitory constant (Ki) was given by the inter-section point of linear regression lines in the Dixon plotwhere data sets of 1v were plotted against the initialconcentration of the inhibitor [I] Results were sub-sequently subjected to Equation 1 (Ito et al 1998) byleast-square regression using SCIENTIST
cedil ˆ Vmax cent permilMFŠ
Km cent 1 Dagger permilIŠKi
8gt
9gtDagger permilMFŠ
hellip1dagger
where Km represents the Michaelis constant of the enzy-matic reaction (cf Equation 5) [MF] is the initial drugconcentration and Vmax indicates the maximum velocityof the reaction
Kinetic parameters for mechanism-based enzyme inac-tivation were obtained as described by Ito et al (1998) Thelogarithm of the remaining enzymatic activity (formationrate of 6shy -hydroxy-MF) was plotted against the pre-incu-bation time in the presence of the inhibitor at variousconcentrations The apparent inactivation rate constant(kiobs) was determined from the slope of the initial linearphase at each inhibitor concentration The apparent kiobs
values were compared with those of control incubationscontaining the solvent for each inhibitor as appropriateThe values of kiobs were plotted against the inhibitorconcentrations and the parameters (kimax and K0
i) wereestimated from Equation 2 by a non-linear least squaremethod using SCIENTIST
kiobs ˆ kimax cent permilIŠK0
i Dagger permilIŠhellip2dagger
where kiobs is the apparent inactivation rate constant ofthe enzyme at initial inhibitor concentration [I] kimax
represents the maximum inactivation rate constantand K0
i represents the apparent dissociation constantbetween the enzyme and the inhibitor respectively(Ito et al 1998)
620 X W Teng et al
The IC50 (the concentration inhibiting 50 of enzymeactivity) was determined by linear regression (percentage ofcontrol) against the logarithmic inhibitor concentration
Results
Degradation in rat serum
MF (C0 ˆ 19 middotM) degraded in rat serum (pH 736) with ahalf-life of 80 sect 01 h (n ˆ 4) Four products namely AB C and D (Figure 1) were formed in rank order and arethe same as observed previously in human plasma and insimulated lung fluid (Teng et al 2001 products A B Cand D reported here correspond to products D1 D3D2 and D4 reported in Teng et al 2001) They were iden-tified as A 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione 17-(2-furoate) B 9not21shy -dichloro-11shy 21not-dihydroxy-16not-methylpregna-1417 20-tetraen-3-one 21-(2-furoate) C 21shy -chloro-21 not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-141720-tetraen-3-one 21-(2-furoate) and D 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione (Teng 2002)
Degradation in rat urine
MF (C0 ˆ 19 middotM) degraded in rat urine (pH 783) with a half-life of 111 sect 03 h (n ˆ 4) The degradation profile of MF inrat urine was qualitatively similar to that in rat serum
Metabolism of MF in S-9 fractions of rat tissues
MF (C0 ˆ 19 middotM) decomposed quickly with a half-life of233 sect 107 min (n ˆ 6) and rapid formation of 6shy -hydroxy-MF in the rat liver tissue S-9 fractions of proteincontent 4 mg mLiexcl1 The formation of 6shy -hydroxy-MFwas not observed in control samples in which the parentdrug or the S-9 fractions were absent In rat liver tissue S-9fraction the apparent rate of MF 6shy -hydroxylation in thepresence of cofactors was over 1000-times greater than thatin the absence of cofactors indicating that this reaction waslargely catalysed by enzymes dependent on cofactors
The apparent rate of MF (C0 ˆ 19 middotM) disappearanceand formation of 6shy -hydroxy-MF increased with anincrease in total protein concentration of rat liver tissueS-9 fractions from 1 to 8 mg mLiexcl1 On the other hand theapparent rates of MF disappearance and 6shy -hydroxy-MFformation were both enhanced with an increase in the initialconcentration of MF from 1 to 50 middotM but there was nofurther increase at 100 middotM when the total protein of rat livertissue S-9 fraction was kept constant at 4 mg mLiexcl1
In addition the relative activities of the S-9 fractions ofthe tested extrahepatic tissues catalysing MF 6shy -hydroxyla-tion were less than 1 of that of the rat liver tissue In theseS-9 fractions the apparent rates of MF 6shy -hydroxylationwere in the order of intestine 5 stomach gt spleen gt kid-ney gt lung In the rat lung tissue S-9 fraction 6shy -hydroxy-MF was detectable but not quantifiable in about half of thesamples using the HPLCUV assay
In the presence of cofactors the rate of degradation ofMF through the formation of products A B C and D wasmuch slower than in the corresponding tissue S-9 fractionswithout cofactors (P lt 001 paired-sample t-test) In factonly product A (not B C or D) was detectable duringincubation up to 72 h in all S-9 fractions of tissues testedwith cofactors
Metabolism in rat liver subcellular fractions
MF was metabolized rapidly in rat liver microsomes with6shy -hydroxy-MF as a primary metabolite (Figure 2) underan NADPH-generating system No significant transform-ation of MF to 6shy -hydroxy-MF was detected in thecontrols As in the S-9 fraction of rat liver tissue noneof the other potential metabolites listed by Affrime et al(2000a) were observed except for 6shy -hydroxy-MF
In pooled liver microsomes from six rats the apparentrate of both parent drug disappearance and 6shy -hydroxy-MF appearance increased with an increase in proteinconcentration over the range of 01 to 5 mg mLiexcl1 whenan initial concentration of MF was kept at 4 middotM
When total protein of rat liver microsomes was keptconstant at 02 mg mLiexcl1 the apparent rate of parent drugdisappearance (Figure 3) increased with an increase in theinitial drug concentration from 02 to 65 middotM The declineof MF followed first-order degradation kinetics in rat livermicrosomes of protein content 02 mg mLiexcl1 The rate of6shy -hydroxy-MF formation increased correspondinglywith increasing initial drug concentration from 02 to8 middotM However the formation was saturated at higherconcentrations reaching a plateau An optimal incubationtime of 8 min was selected as a convenient common incu-bation time for studying the kinetics of MF metabolism inrat liver microsomes of protein content 02 mg mLiexcl1
Little formation of 6shy -hydroxy-MF and disappearanceof the parent drug were observed following the incubationof MF (C0 ˆ 4 middotM) in pooled rat liver cytosolic mitochon-drial and nuclear fractions of protein content 4 mg mLiexcl1
at 37 macrC (Table 1) The enzyme activities of this cytosolicmitochondrial and nuclear fractions catalysing 6shy -hydro-xylation of MF were 04 43 and 16 of that of pooledrat liver microsomes respectively
Transformation in everted rat intestine
Following the incubation of MF (C0 ˆ 19 middotM) with evertedintestinal tissue in K-H buffer MF disappeared rapidlyfrom the incubation buffer with considerable formation of6shy -hydroxy-MF as the major metabolite (Figure 4)Meanwhile most of the parent drug and the metaboliteformed were taken up by the intestinal tissue during incu-bation
Inhibition of MF 6b-hydroxylation by potentialchemical inhibitors
A non-specific CYP inhibitor 1-aminobenzotriazole(Mathews et al 1985 Carcillo et al 1998) inhibited MF6shy -hydroxylation in pooled rat liver microsomes in a
Rat metabolism and degradation kinetics of mometasone furoate 621
concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

dehydrogenase) in 100 mM phosphate buffer containing1 mM EDTA (pH 74) under carbogen gas at 370 sect 01 macrCin a shaking (75 rev miniexcl1) water bath Typically the S-9fraction (protein content 4 mg mLiexcl1 unless otherwiseindicated) and the parent drug (C0 ˆ 19 middotM added as amethanolic stock solution of 20 mg mLiexcl1 at a volume of05 in the final incubation mixtures) were pre-incubatedin the incubation buffer for 5 min at 37 sect 01 macrC Thereaction was initiated by adding the cofactors At pre-determined time intervals samples (05 mL each) werewithdrawn and the reaction was terminated immediatelyby adding 4 mL ice-cold dichloromethane Samples werethen extracted and analysed by HPLC (Teng et al 2001)Parallel incubations were performed in the same mannerin the absence of MF or the S-9 fraction as controlsExperiments with the tissue S-9 fractions were performedin four replicates
The metabolic kinetics of MF in pooled rat liver tissueS-9 fractions was investigated with respect to protein con-centration (range 1plusmn8 mg mLiexcl1) and substrate concentra-tion (range 1plusmn100 middotM)
Studies of degradation kinetics of MF (C0 ˆ 19 middotMadded as a methanolic stock solution of 20 mg mLiexcl1 ata volume of 05 in the final incubation mixtures) in eachS-9 fraction of the rat tissues were carried out withoutcofactors The reaction was initiated by adding the stockMF in the reaction media and then handled using thesame method
Kinetics of MF in rat liver microsomal cytosolicmitochondrial and nuclear fractions
Initially the microsomal incubation conditions wereinvestigated with respect to microsomal protein concen-tration (01plusmn5 mg mLiexcl1) incubation time (up to 60 min)substrate concentration (02plusmn100 middotM) and oxygenationusing pooled rat liver microsomes The optimizedmicrosomal incubation mixture was composed of livermicrosomes (02 mg protein mLiexcl1) 25 middotM MF and anNADPH-generating system (6 mM MgCl2 05 mM shy -NADP10 mM glucose 6-phosphate and 10 units mLiexcl1 glucose-6-phosphate dehydrogenase) in 100 mM phosphate buffercontaining 1 mM EDTA (pH 74) The reaction was car-ried out in open test tubes without extra oxygenation for8 min in a shaking (150 rev miniexcl1) water bath After themicrosomal fraction and the parent drug (added as amethanolic stock solution at a volume of 1 in the finalincubation mixtures) were pre-warmed in the incubationbuffer for 5 min at 37 sect 01 macrC the reaction was initiatedby the addition of the NADPH-generating system andterminated by addition of 4 mL ice-cold dichloromethaneSimilar incubations with the use of denatured microsomes(heating at 70plusmn80 macrC for 30 min) or without the addition ofmicrosomes or substrate were conducted in the same man-ner as controls In addition incubation in the absence ofthe NADPH-generating system was performed for theconfirmation of P450-dependent enzymatic biotransfor-mation Typically all the incubations of MF (C0 ˆ 4 middotM)with microsomal cytosolic mitochondrial and nuclear
fractions (protein content 4 mg mLiexcl1) were performedfollowing the same procedure Incubations for each indi-vidual condition with these subcellular fractions of ratliver tissue were conducted in duplicate
Kinetics in everted rat small intestine tissue
A modified version of the method of Esposito amp Csaky(1974) was employed in these studies After anaesthetiza-tion of the rat with halothane an abdominal incision wasmade and the whole small intestine was isolated excisedand examined After a section (sup110 cm) of intestine at theend of the ileocecal junction was taken for tissue sampleof zero time the intestine was immediately flushed withice-cold Krebs-Henseleit bicarbonate buffer containing 2 gLiexcl1 glucose (pH 74 K-H buffer) and then everted ona glass rod with one end tied with surgical thread After thesac was filled with warmed drug-free K-H buffer (37 macrC)the other end was tied and the middle part of the intestinewas tied every 20plusmn25 cm to give four isolated sections Thiseverted intestine was immediately placed in a freshly pre-pared warmed (37 macrC) and jacketed beaker containing25 mL of 19 middotM MF (10 middotg mLiexcl1 added as a methanolicstock solution of 20 mg mLiexcl1 at a volume of 05 in thefinal incubation mixtures) in K-H buffer and incubated at37 sect 01 macrC with continual carbogenation with carbogen ina shaking water bath (75 rev miniexcl1)
The zero-time buffer sample (the K-H buffer withdrug) was withdrawn before the tissue was placed in theK-H buffer and the drug-free K-H buffer was used as thezero-time buffer sample in the sac At 05 15 3 and 5 h3 mL of buffer in the bulk incubation medium and asection of the intestine (about 2 g) with the buffer in itslumen were sampled After the buffer in the lumen wastransferred into a beaker the tissue segment was weighedand then immediately added to ice-cold K-H buffer ofequal weight Subsequently the tissue was minced homo-genized extracted and analysed by HPLC
Buffer samples (05 mL) from both the bulk incubationmedium and the intestinal lumen were also collectedfor analysis of MF and its degradation and metabolicproducts as described for samples of tissue subcellularfractions The experiment was performed in triplicate
Inhibition studies
A number of potential inhibitors andor substrates ofCYP isoenzymes were tested at various concentrationsin pooled rat liver microsomes (protein content 02 mgmLiexcl1) with varying concentrations of MF (added as astock methanolic solution at a volume of 1 in the finalincubation mixtures) Incubation mixtures containingmechanism inhibitors furafylline diethyldithiocarba-mate troleandomycin or 1-aminobenzotriazole werepre-incubated in pooled rat liver microsomes in the pre-sence of the NADPH-generating system (6 mM MgCl205 mM shy -NADP 10 mM glucose-6-phosphate and 10units mLiexcl1 glucose-6-phosphate dehydrogenase) at37 sect 1 macrC for 15 min before the addition of MF Fornon-mechanism-based inhibitors the incubation mixtures
Rat metabolism and degradation kinetics of mometasone furoate 619
contained MF and inhibitors together with rat liver micro-somes without pre-incubation of inhibitors and thereactions were initiated by the addition of the NADPH-generating system and carried out as for rat liver micro-somes Potential inhibitors were dissolved in a mixture ofmethanol and phosphate buffer except for ketoconazolewhich was dissolved in acetone and added at a volume of1 in the final incubation mixtures Parallel controlincubations were conducted with 1 of the correspondingsolvent with no inhibitor present Each potential inhibitorwas also incubated with microsomes under the NADPH-generating system to examine if they caused any chro-matographic interference in the measurement of6shy -hydroxy-MF
Sample extraction and analysis
For tissue and sub-cellular fractions on terminating thereaction with 40 mL ice-cold dichloromethane 05 mL ofinternal standard (10 middotg mLiexcl1 of ethanolic testosterone17-acetate) was added to the sample (05 mL) The mixturewas extracted for 2 min on a vibrating mixer followed bycentrifugation at 1328 g (20 macrC) for 10 min The organiclayer was then collected evaporated to dryness undernitrogen at 35 macrC stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
To each sample (05 g) of tissue homogenate was added05 mL of internal standard (10 middotg mLiexcl1 ethanolic testo-sterone 17-acetate) The mixture was extracted twice with4 mL dichloromethane by vigorous shaking for 2 min ona vibrating mixer followed by centrifugation at 2602 g(20 macrC) for 10 min The dichloromethane phase was col-lected evaporated stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
Data analysis
Data are expressed as the mean sect sd of replicate deter-minations (n 5 5) Statistical analyses were performedusing Microsoft Excel (version 97) or SPSS (Spss IncIL USA) Significance of differences in means of pairedsamples was assessed using a paired-sample t-testAnalysis of variance of two or more groups of data wasperformed using analysis of variance and Tukeyrsquos post-hoc analysis All values of P were based on two-tailed testsand P values of less than 005 were considered statisticallysignificant
The observed rate constants (kobs) of parent drug dis-appearance were estimated from the initial slope of thelog-linear phase of declining concentration versus timeplots The initial half-lives were calculated using the fol-lowing equation t1curren2 ˆ 0693kobs The rate constant ofeach metabolic reaction was estimated according to thecorresponding model developed based on the experi-mental data to limit underestimation of the parametersdue to possible sequential transformation The rate con-stant of parent drug disappearance was correlated to thesum of those of its direct transformation reactions
Substrate concentrations ranging from 02 to 100 middotM
were employed for MF in view of its unknown in-vitro
metabolic kinetics in rat liver microsomes Kinetic analysisof the parent drug disappearance and product formationdata was undertaken by the least-square regression programusing SCIENTIST version 20 (Micromath ScientificSoftware Salt Lake City UT USA) Apparent enzymekinetics was determined by fitting reaction velocity (v) versussubstrate concentration [MF] Initially the data were exam-ined on Eadie-Hofstee Lineweaver-Burk and Hanes plots toassess the linearity of enzyme kinetics The kinetic data werethen fitted to expressions containing one or more Michaelis-Menten terms The correlation randomness of the residualsthe standard deviation of the parameter estimates (lt10)and the model selection were used as indicators of goodness-of-fit of the model to the experimental data
The nature of inhibition of the chemical inhibitors forP450 was initially assessed by plots from transformed datafrom the kinetic studies including Dixon plotLineweaver-Burk plot Eadie-Hofstee plot and Cornish-Bowden plot For competitive enzyme inhibitors theapparent inhibitory constant (Ki) was given by the inter-section point of linear regression lines in the Dixon plotwhere data sets of 1v were plotted against the initialconcentration of the inhibitor [I] Results were sub-sequently subjected to Equation 1 (Ito et al 1998) byleast-square regression using SCIENTIST
cedil ˆ Vmax cent permilMFŠ
Km cent 1 Dagger permilIŠKi
8gt
9gtDagger permilMFŠ
hellip1dagger
where Km represents the Michaelis constant of the enzy-matic reaction (cf Equation 5) [MF] is the initial drugconcentration and Vmax indicates the maximum velocityof the reaction
Kinetic parameters for mechanism-based enzyme inac-tivation were obtained as described by Ito et al (1998) Thelogarithm of the remaining enzymatic activity (formationrate of 6shy -hydroxy-MF) was plotted against the pre-incu-bation time in the presence of the inhibitor at variousconcentrations The apparent inactivation rate constant(kiobs) was determined from the slope of the initial linearphase at each inhibitor concentration The apparent kiobs
values were compared with those of control incubationscontaining the solvent for each inhibitor as appropriateThe values of kiobs were plotted against the inhibitorconcentrations and the parameters (kimax and K0
i) wereestimated from Equation 2 by a non-linear least squaremethod using SCIENTIST
kiobs ˆ kimax cent permilIŠK0
i Dagger permilIŠhellip2dagger
where kiobs is the apparent inactivation rate constant ofthe enzyme at initial inhibitor concentration [I] kimax
represents the maximum inactivation rate constantand K0
i represents the apparent dissociation constantbetween the enzyme and the inhibitor respectively(Ito et al 1998)
620 X W Teng et al
The IC50 (the concentration inhibiting 50 of enzymeactivity) was determined by linear regression (percentage ofcontrol) against the logarithmic inhibitor concentration
Results
Degradation in rat serum
MF (C0 ˆ 19 middotM) degraded in rat serum (pH 736) with ahalf-life of 80 sect 01 h (n ˆ 4) Four products namely AB C and D (Figure 1) were formed in rank order and arethe same as observed previously in human plasma and insimulated lung fluid (Teng et al 2001 products A B Cand D reported here correspond to products D1 D3D2 and D4 reported in Teng et al 2001) They were iden-tified as A 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione 17-(2-furoate) B 9not21shy -dichloro-11shy 21not-dihydroxy-16not-methylpregna-1417 20-tetraen-3-one 21-(2-furoate) C 21shy -chloro-21 not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-141720-tetraen-3-one 21-(2-furoate) and D 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione (Teng 2002)
Degradation in rat urine
MF (C0 ˆ 19 middotM) degraded in rat urine (pH 783) with a half-life of 111 sect 03 h (n ˆ 4) The degradation profile of MF inrat urine was qualitatively similar to that in rat serum
Metabolism of MF in S-9 fractions of rat tissues
MF (C0 ˆ 19 middotM) decomposed quickly with a half-life of233 sect 107 min (n ˆ 6) and rapid formation of 6shy -hydroxy-MF in the rat liver tissue S-9 fractions of proteincontent 4 mg mLiexcl1 The formation of 6shy -hydroxy-MFwas not observed in control samples in which the parentdrug or the S-9 fractions were absent In rat liver tissue S-9fraction the apparent rate of MF 6shy -hydroxylation in thepresence of cofactors was over 1000-times greater than thatin the absence of cofactors indicating that this reaction waslargely catalysed by enzymes dependent on cofactors
The apparent rate of MF (C0 ˆ 19 middotM) disappearanceand formation of 6shy -hydroxy-MF increased with anincrease in total protein concentration of rat liver tissueS-9 fractions from 1 to 8 mg mLiexcl1 On the other hand theapparent rates of MF disappearance and 6shy -hydroxy-MFformation were both enhanced with an increase in the initialconcentration of MF from 1 to 50 middotM but there was nofurther increase at 100 middotM when the total protein of rat livertissue S-9 fraction was kept constant at 4 mg mLiexcl1
In addition the relative activities of the S-9 fractions ofthe tested extrahepatic tissues catalysing MF 6shy -hydroxyla-tion were less than 1 of that of the rat liver tissue In theseS-9 fractions the apparent rates of MF 6shy -hydroxylationwere in the order of intestine 5 stomach gt spleen gt kid-ney gt lung In the rat lung tissue S-9 fraction 6shy -hydroxy-MF was detectable but not quantifiable in about half of thesamples using the HPLCUV assay
In the presence of cofactors the rate of degradation ofMF through the formation of products A B C and D wasmuch slower than in the corresponding tissue S-9 fractionswithout cofactors (P lt 001 paired-sample t-test) In factonly product A (not B C or D) was detectable duringincubation up to 72 h in all S-9 fractions of tissues testedwith cofactors
Metabolism in rat liver subcellular fractions
MF was metabolized rapidly in rat liver microsomes with6shy -hydroxy-MF as a primary metabolite (Figure 2) underan NADPH-generating system No significant transform-ation of MF to 6shy -hydroxy-MF was detected in thecontrols As in the S-9 fraction of rat liver tissue noneof the other potential metabolites listed by Affrime et al(2000a) were observed except for 6shy -hydroxy-MF
In pooled liver microsomes from six rats the apparentrate of both parent drug disappearance and 6shy -hydroxy-MF appearance increased with an increase in proteinconcentration over the range of 01 to 5 mg mLiexcl1 whenan initial concentration of MF was kept at 4 middotM
When total protein of rat liver microsomes was keptconstant at 02 mg mLiexcl1 the apparent rate of parent drugdisappearance (Figure 3) increased with an increase in theinitial drug concentration from 02 to 65 middotM The declineof MF followed first-order degradation kinetics in rat livermicrosomes of protein content 02 mg mLiexcl1 The rate of6shy -hydroxy-MF formation increased correspondinglywith increasing initial drug concentration from 02 to8 middotM However the formation was saturated at higherconcentrations reaching a plateau An optimal incubationtime of 8 min was selected as a convenient common incu-bation time for studying the kinetics of MF metabolism inrat liver microsomes of protein content 02 mg mLiexcl1
Little formation of 6shy -hydroxy-MF and disappearanceof the parent drug were observed following the incubationof MF (C0 ˆ 4 middotM) in pooled rat liver cytosolic mitochon-drial and nuclear fractions of protein content 4 mg mLiexcl1
at 37 macrC (Table 1) The enzyme activities of this cytosolicmitochondrial and nuclear fractions catalysing 6shy -hydro-xylation of MF were 04 43 and 16 of that of pooledrat liver microsomes respectively
Transformation in everted rat intestine
Following the incubation of MF (C0 ˆ 19 middotM) with evertedintestinal tissue in K-H buffer MF disappeared rapidlyfrom the incubation buffer with considerable formation of6shy -hydroxy-MF as the major metabolite (Figure 4)Meanwhile most of the parent drug and the metaboliteformed were taken up by the intestinal tissue during incu-bation
Inhibition of MF 6b-hydroxylation by potentialchemical inhibitors
A non-specific CYP inhibitor 1-aminobenzotriazole(Mathews et al 1985 Carcillo et al 1998) inhibited MF6shy -hydroxylation in pooled rat liver microsomes in a
Rat metabolism and degradation kinetics of mometasone furoate 621
concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

contained MF and inhibitors together with rat liver micro-somes without pre-incubation of inhibitors and thereactions were initiated by the addition of the NADPH-generating system and carried out as for rat liver micro-somes Potential inhibitors were dissolved in a mixture ofmethanol and phosphate buffer except for ketoconazolewhich was dissolved in acetone and added at a volume of1 in the final incubation mixtures Parallel controlincubations were conducted with 1 of the correspondingsolvent with no inhibitor present Each potential inhibitorwas also incubated with microsomes under the NADPH-generating system to examine if they caused any chro-matographic interference in the measurement of6shy -hydroxy-MF
Sample extraction and analysis
For tissue and sub-cellular fractions on terminating thereaction with 40 mL ice-cold dichloromethane 05 mL ofinternal standard (10 middotg mLiexcl1 of ethanolic testosterone17-acetate) was added to the sample (05 mL) The mixturewas extracted for 2 min on a vibrating mixer followed bycentrifugation at 1328 g (20 macrC) for 10 min The organiclayer was then collected evaporated to dryness undernitrogen at 35 macrC stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
To each sample (05 g) of tissue homogenate was added05 mL of internal standard (10 middotg mLiexcl1 ethanolic testo-sterone 17-acetate) The mixture was extracted twice with4 mL dichloromethane by vigorous shaking for 2 min ona vibrating mixer followed by centrifugation at 2602 g(20 macrC) for 10 min The dichloromethane phase was col-lected evaporated stored at iexcl20 macrC and reconstituted inmobile phase before HPLC analysis
Data analysis
Data are expressed as the mean sect sd of replicate deter-minations (n 5 5) Statistical analyses were performedusing Microsoft Excel (version 97) or SPSS (Spss IncIL USA) Significance of differences in means of pairedsamples was assessed using a paired-sample t-testAnalysis of variance of two or more groups of data wasperformed using analysis of variance and Tukeyrsquos post-hoc analysis All values of P were based on two-tailed testsand P values of less than 005 were considered statisticallysignificant
The observed rate constants (kobs) of parent drug dis-appearance were estimated from the initial slope of thelog-linear phase of declining concentration versus timeplots The initial half-lives were calculated using the fol-lowing equation t1curren2 ˆ 0693kobs The rate constant ofeach metabolic reaction was estimated according to thecorresponding model developed based on the experi-mental data to limit underestimation of the parametersdue to possible sequential transformation The rate con-stant of parent drug disappearance was correlated to thesum of those of its direct transformation reactions
Substrate concentrations ranging from 02 to 100 middotM
were employed for MF in view of its unknown in-vitro
metabolic kinetics in rat liver microsomes Kinetic analysisof the parent drug disappearance and product formationdata was undertaken by the least-square regression programusing SCIENTIST version 20 (Micromath ScientificSoftware Salt Lake City UT USA) Apparent enzymekinetics was determined by fitting reaction velocity (v) versussubstrate concentration [MF] Initially the data were exam-ined on Eadie-Hofstee Lineweaver-Burk and Hanes plots toassess the linearity of enzyme kinetics The kinetic data werethen fitted to expressions containing one or more Michaelis-Menten terms The correlation randomness of the residualsthe standard deviation of the parameter estimates (lt10)and the model selection were used as indicators of goodness-of-fit of the model to the experimental data
The nature of inhibition of the chemical inhibitors forP450 was initially assessed by plots from transformed datafrom the kinetic studies including Dixon plotLineweaver-Burk plot Eadie-Hofstee plot and Cornish-Bowden plot For competitive enzyme inhibitors theapparent inhibitory constant (Ki) was given by the inter-section point of linear regression lines in the Dixon plotwhere data sets of 1v were plotted against the initialconcentration of the inhibitor [I] Results were sub-sequently subjected to Equation 1 (Ito et al 1998) byleast-square regression using SCIENTIST
cedil ˆ Vmax cent permilMFŠ
Km cent 1 Dagger permilIŠKi
8gt
9gtDagger permilMFŠ
hellip1dagger
where Km represents the Michaelis constant of the enzy-matic reaction (cf Equation 5) [MF] is the initial drugconcentration and Vmax indicates the maximum velocityof the reaction
Kinetic parameters for mechanism-based enzyme inac-tivation were obtained as described by Ito et al (1998) Thelogarithm of the remaining enzymatic activity (formationrate of 6shy -hydroxy-MF) was plotted against the pre-incu-bation time in the presence of the inhibitor at variousconcentrations The apparent inactivation rate constant(kiobs) was determined from the slope of the initial linearphase at each inhibitor concentration The apparent kiobs
values were compared with those of control incubationscontaining the solvent for each inhibitor as appropriateThe values of kiobs were plotted against the inhibitorconcentrations and the parameters (kimax and K0
i) wereestimated from Equation 2 by a non-linear least squaremethod using SCIENTIST
kiobs ˆ kimax cent permilIŠK0
i Dagger permilIŠhellip2dagger
where kiobs is the apparent inactivation rate constant ofthe enzyme at initial inhibitor concentration [I] kimax
represents the maximum inactivation rate constantand K0
i represents the apparent dissociation constantbetween the enzyme and the inhibitor respectively(Ito et al 1998)
620 X W Teng et al
The IC50 (the concentration inhibiting 50 of enzymeactivity) was determined by linear regression (percentage ofcontrol) against the logarithmic inhibitor concentration
Results
Degradation in rat serum
MF (C0 ˆ 19 middotM) degraded in rat serum (pH 736) with ahalf-life of 80 sect 01 h (n ˆ 4) Four products namely AB C and D (Figure 1) were formed in rank order and arethe same as observed previously in human plasma and insimulated lung fluid (Teng et al 2001 products A B Cand D reported here correspond to products D1 D3D2 and D4 reported in Teng et al 2001) They were iden-tified as A 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione 17-(2-furoate) B 9not21shy -dichloro-11shy 21not-dihydroxy-16not-methylpregna-1417 20-tetraen-3-one 21-(2-furoate) C 21shy -chloro-21 not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-141720-tetraen-3-one 21-(2-furoate) and D 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione (Teng 2002)
Degradation in rat urine
MF (C0 ˆ 19 middotM) degraded in rat urine (pH 783) with a half-life of 111 sect 03 h (n ˆ 4) The degradation profile of MF inrat urine was qualitatively similar to that in rat serum
Metabolism of MF in S-9 fractions of rat tissues
MF (C0 ˆ 19 middotM) decomposed quickly with a half-life of233 sect 107 min (n ˆ 6) and rapid formation of 6shy -hydroxy-MF in the rat liver tissue S-9 fractions of proteincontent 4 mg mLiexcl1 The formation of 6shy -hydroxy-MFwas not observed in control samples in which the parentdrug or the S-9 fractions were absent In rat liver tissue S-9fraction the apparent rate of MF 6shy -hydroxylation in thepresence of cofactors was over 1000-times greater than thatin the absence of cofactors indicating that this reaction waslargely catalysed by enzymes dependent on cofactors
The apparent rate of MF (C0 ˆ 19 middotM) disappearanceand formation of 6shy -hydroxy-MF increased with anincrease in total protein concentration of rat liver tissueS-9 fractions from 1 to 8 mg mLiexcl1 On the other hand theapparent rates of MF disappearance and 6shy -hydroxy-MFformation were both enhanced with an increase in the initialconcentration of MF from 1 to 50 middotM but there was nofurther increase at 100 middotM when the total protein of rat livertissue S-9 fraction was kept constant at 4 mg mLiexcl1
In addition the relative activities of the S-9 fractions ofthe tested extrahepatic tissues catalysing MF 6shy -hydroxyla-tion were less than 1 of that of the rat liver tissue In theseS-9 fractions the apparent rates of MF 6shy -hydroxylationwere in the order of intestine 5 stomach gt spleen gt kid-ney gt lung In the rat lung tissue S-9 fraction 6shy -hydroxy-MF was detectable but not quantifiable in about half of thesamples using the HPLCUV assay
In the presence of cofactors the rate of degradation ofMF through the formation of products A B C and D wasmuch slower than in the corresponding tissue S-9 fractionswithout cofactors (P lt 001 paired-sample t-test) In factonly product A (not B C or D) was detectable duringincubation up to 72 h in all S-9 fractions of tissues testedwith cofactors
Metabolism in rat liver subcellular fractions
MF was metabolized rapidly in rat liver microsomes with6shy -hydroxy-MF as a primary metabolite (Figure 2) underan NADPH-generating system No significant transform-ation of MF to 6shy -hydroxy-MF was detected in thecontrols As in the S-9 fraction of rat liver tissue noneof the other potential metabolites listed by Affrime et al(2000a) were observed except for 6shy -hydroxy-MF
In pooled liver microsomes from six rats the apparentrate of both parent drug disappearance and 6shy -hydroxy-MF appearance increased with an increase in proteinconcentration over the range of 01 to 5 mg mLiexcl1 whenan initial concentration of MF was kept at 4 middotM
When total protein of rat liver microsomes was keptconstant at 02 mg mLiexcl1 the apparent rate of parent drugdisappearance (Figure 3) increased with an increase in theinitial drug concentration from 02 to 65 middotM The declineof MF followed first-order degradation kinetics in rat livermicrosomes of protein content 02 mg mLiexcl1 The rate of6shy -hydroxy-MF formation increased correspondinglywith increasing initial drug concentration from 02 to8 middotM However the formation was saturated at higherconcentrations reaching a plateau An optimal incubationtime of 8 min was selected as a convenient common incu-bation time for studying the kinetics of MF metabolism inrat liver microsomes of protein content 02 mg mLiexcl1
Little formation of 6shy -hydroxy-MF and disappearanceof the parent drug were observed following the incubationof MF (C0 ˆ 4 middotM) in pooled rat liver cytosolic mitochon-drial and nuclear fractions of protein content 4 mg mLiexcl1
at 37 macrC (Table 1) The enzyme activities of this cytosolicmitochondrial and nuclear fractions catalysing 6shy -hydro-xylation of MF were 04 43 and 16 of that of pooledrat liver microsomes respectively
Transformation in everted rat intestine
Following the incubation of MF (C0 ˆ 19 middotM) with evertedintestinal tissue in K-H buffer MF disappeared rapidlyfrom the incubation buffer with considerable formation of6shy -hydroxy-MF as the major metabolite (Figure 4)Meanwhile most of the parent drug and the metaboliteformed were taken up by the intestinal tissue during incu-bation
Inhibition of MF 6b-hydroxylation by potentialchemical inhibitors
A non-specific CYP inhibitor 1-aminobenzotriazole(Mathews et al 1985 Carcillo et al 1998) inhibited MF6shy -hydroxylation in pooled rat liver microsomes in a
Rat metabolism and degradation kinetics of mometasone furoate 621
concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

The IC50 (the concentration inhibiting 50 of enzymeactivity) was determined by linear regression (percentage ofcontrol) against the logarithmic inhibitor concentration
Results
Degradation in rat serum
MF (C0 ˆ 19 middotM) degraded in rat serum (pH 736) with ahalf-life of 80 sect 01 h (n ˆ 4) Four products namely AB C and D (Figure 1) were formed in rank order and arethe same as observed previously in human plasma and insimulated lung fluid (Teng et al 2001 products A B Cand D reported here correspond to products D1 D3D2 and D4 reported in Teng et al 2001) They were iden-tified as A 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione 17-(2-furoate) B 9not21shy -dichloro-11shy 21not-dihydroxy-16not-methylpregna-1417 20-tetraen-3-one 21-(2-furoate) C 21shy -chloro-21 not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-141720-tetraen-3-one 21-(2-furoate) and D 21-chloro-17not-hydroxy-16not-methyl-9shy 11shy -oxidopregna-14-diene-320-dione (Teng 2002)
Degradation in rat urine
MF (C0 ˆ 19 middotM) degraded in rat urine (pH 783) with a half-life of 111 sect 03 h (n ˆ 4) The degradation profile of MF inrat urine was qualitatively similar to that in rat serum
Metabolism of MF in S-9 fractions of rat tissues
MF (C0 ˆ 19 middotM) decomposed quickly with a half-life of233 sect 107 min (n ˆ 6) and rapid formation of 6shy -hydroxy-MF in the rat liver tissue S-9 fractions of proteincontent 4 mg mLiexcl1 The formation of 6shy -hydroxy-MFwas not observed in control samples in which the parentdrug or the S-9 fractions were absent In rat liver tissue S-9fraction the apparent rate of MF 6shy -hydroxylation in thepresence of cofactors was over 1000-times greater than thatin the absence of cofactors indicating that this reaction waslargely catalysed by enzymes dependent on cofactors
The apparent rate of MF (C0 ˆ 19 middotM) disappearanceand formation of 6shy -hydroxy-MF increased with anincrease in total protein concentration of rat liver tissueS-9 fractions from 1 to 8 mg mLiexcl1 On the other hand theapparent rates of MF disappearance and 6shy -hydroxy-MFformation were both enhanced with an increase in the initialconcentration of MF from 1 to 50 middotM but there was nofurther increase at 100 middotM when the total protein of rat livertissue S-9 fraction was kept constant at 4 mg mLiexcl1
In addition the relative activities of the S-9 fractions ofthe tested extrahepatic tissues catalysing MF 6shy -hydroxyla-tion were less than 1 of that of the rat liver tissue In theseS-9 fractions the apparent rates of MF 6shy -hydroxylationwere in the order of intestine 5 stomach gt spleen gt kid-ney gt lung In the rat lung tissue S-9 fraction 6shy -hydroxy-MF was detectable but not quantifiable in about half of thesamples using the HPLCUV assay
In the presence of cofactors the rate of degradation ofMF through the formation of products A B C and D wasmuch slower than in the corresponding tissue S-9 fractionswithout cofactors (P lt 001 paired-sample t-test) In factonly product A (not B C or D) was detectable duringincubation up to 72 h in all S-9 fractions of tissues testedwith cofactors
Metabolism in rat liver subcellular fractions
MF was metabolized rapidly in rat liver microsomes with6shy -hydroxy-MF as a primary metabolite (Figure 2) underan NADPH-generating system No significant transform-ation of MF to 6shy -hydroxy-MF was detected in thecontrols As in the S-9 fraction of rat liver tissue noneof the other potential metabolites listed by Affrime et al(2000a) were observed except for 6shy -hydroxy-MF
In pooled liver microsomes from six rats the apparentrate of both parent drug disappearance and 6shy -hydroxy-MF appearance increased with an increase in proteinconcentration over the range of 01 to 5 mg mLiexcl1 whenan initial concentration of MF was kept at 4 middotM
When total protein of rat liver microsomes was keptconstant at 02 mg mLiexcl1 the apparent rate of parent drugdisappearance (Figure 3) increased with an increase in theinitial drug concentration from 02 to 65 middotM The declineof MF followed first-order degradation kinetics in rat livermicrosomes of protein content 02 mg mLiexcl1 The rate of6shy -hydroxy-MF formation increased correspondinglywith increasing initial drug concentration from 02 to8 middotM However the formation was saturated at higherconcentrations reaching a plateau An optimal incubationtime of 8 min was selected as a convenient common incu-bation time for studying the kinetics of MF metabolism inrat liver microsomes of protein content 02 mg mLiexcl1
Little formation of 6shy -hydroxy-MF and disappearanceof the parent drug were observed following the incubationof MF (C0 ˆ 4 middotM) in pooled rat liver cytosolic mitochon-drial and nuclear fractions of protein content 4 mg mLiexcl1
at 37 macrC (Table 1) The enzyme activities of this cytosolicmitochondrial and nuclear fractions catalysing 6shy -hydro-xylation of MF were 04 43 and 16 of that of pooledrat liver microsomes respectively
Transformation in everted rat intestine
Following the incubation of MF (C0 ˆ 19 middotM) with evertedintestinal tissue in K-H buffer MF disappeared rapidlyfrom the incubation buffer with considerable formation of6shy -hydroxy-MF as the major metabolite (Figure 4)Meanwhile most of the parent drug and the metaboliteformed were taken up by the intestinal tissue during incu-bation
Inhibition of MF 6b-hydroxylation by potentialchemical inhibitors
A non-specific CYP inhibitor 1-aminobenzotriazole(Mathews et al 1985 Carcillo et al 1998) inhibited MF6shy -hydroxylation in pooled rat liver microsomes in a
Rat metabolism and degradation kinetics of mometasone furoate 621
concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

concentration-dependent manner Strong inhibition wasobtained at low concentrations of ketoconazole with anapparent Ki of 142 middotM determined by Equation 1 repre-senting Dixon plots (Figure 5A) Troleandomycin dis-played concentration and pre-incubation time-dependent
inhibition (Figure 6A) with an apparent K0i of approxi-
mately 177 middotM Diazepam also exhibited considerablecompetitive inhibition of MF 6shy -hydroxylation in ratliver with an apparent Ki of approximately 88 middotMMoreover co-incubation with high concentrations
C O
CH Cl2
CHO
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH39
Cl
OO
O23
24
25 26
27
22
MF
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
9 22
C
C
20
21
Cl
HO
Cl
H
B
CO
O23
24
25 26
27
O
ndash H O2
(and rearrangement)
C O
CH2Cl
C
O
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
20
21
5
2
1CH3
9
OO
O23
24
25 26
27
22
H
A
ndash HCl(epoxide formation)
O
CH3
3
4 6
7
810
11
12
13
14
15
1617
18
19
5
2
1CH3
922
O
C
C
20
21
Cl
H
C
CO
O23
24
25 26
27
O
C O
CH2Cl
O
CH3
CH3
3
4 6
7
810
11
12
13
14
15
16
17
18
19
20
21
5
2
1CH3
9
OH
22
O
H
D
(and rearrangement)
Hydrolysis
H
ndash HCl(epoxide formation)
ndash H O2
CH3
CH3
Figure 1 Structures of degradation products of mometasone furoate (MF) with proposed degradation pathways and 21-configuration of
products B and C Thick arrows indicate major pathways
622 X W Teng et al
(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

(300 and 600 middotM) of 4-methylpyrazole caused somedecrease in the reaction rate with an apparent Ki ofaround 700 middotM provided its inhibition was competitiveFurthermore diethyldithiocarbamate reduced the reac-tion rate with an IC50 of approximately 84 middotM In con-trast furafylline and sulfaphenazole did not exhibitsignificant inhibition of the reaction at concentrationshigher than its effective concentration
Analysis of kinetics of the in-vitro metabolismof MF in rat liver microsomes
Based on knowledge of the chemical structure introduc-tion of a hydroxy group at C-6 of the MF molecule wouldnot be expected to cause a significant environmentalchange to the 14-diene-3-one chromophore and thefuran-2-carboxyl chromophore It was evident that 6shy -hydroxy-MF exhibited a nearly identical UV spectrum toMF (data not shown) Assuming the HPLCUV responseof 6shy -hydroxy-MF was equivalent to that of MF the highyield of 6shy -hydroxy-MF in rat liver tissue S-9 and micro-somal fractions indicates that 6shy -hydroxylation is the pri-mary metabolic pathway of MF in rat liver in-vitro Theapparent formation of 6shy -hydroxy-MF would contributeto approximately 43 of the total loss of the parent drugin rat liver tissue S-9 fraction estimated from the slopes ofthe linear regression lines of the apparent rate versusprotein concentration This suggests that MF also under-goes metabolism through other pathways which can beparallel or subsequent or both Except for 6shy -hydroxy-MF under the present experimental conditions none ofthe other metabolites postulated by Affrime et al (2000a)were observed in rat liver in-vitro In addition the C-17ester-bond hydoxylated product of MF was not observedin any media tested The possible reasons of this could be(i) mometasone was not resolved from other compoundsin the HPLC analysis and (ii) the apparent formation ofmometasone was too low Comparing the chromatogramsfrom Affrime et al (2000a) (in which mometasone theproposed product of hydrolysis was eluted after 6shy -hydroxy-MF) with those in the present study mometa-sone was expected to be eluted after the degradationproduct D (the hydrolysate of degradation product A) asin parallel relationship of A and MF through the reversed-phase HPLC system used in this study It was less likelythat mometasone fell into the front peaks in the HPLCchromatogram Therefore it was likely that the mometa-sone formed was too low to be detected andor mometa-sone was sequentially metabolized to other product(s) thatwere not resolved by the HPLC method Thus a kineticscheme was proposed for the in-vitro MF metabolism inrat liver (Figure 7) The disappearance of MF the forma-tion and disappearance of metabolite 6shy -hydroxy-MF(M) can be described by differential Equations 3 and 4As no data were available for N and O no differentialequations for these supposed products were included
permilMFŠ0 ˆ iexclhellipkFM Dagger kFNdaggerpermilMFŠ hellip3dagger
permilMŠ0 ˆ kFMpermilMFŠ iexcl kMOpermilMŠ hellip4dagger
0
05
1
15
2
25
0 10 20 30 40 50 60
Time (min)
MF remaining
M formed
Co
nce
ntr
atio
n (
)mM
Figure 2 Typical time course for mometasone furoate (MF)
(C0 ˆ 26 middotM) and its metabolite 6shy -hydroxy-MF (M) in rat liver
microsomes (protein 02 mg mLiexcl1) at 37 macrC The concentration of
M formed is expressed relative to the MF concentration assuming
equal HPLCUV response Symbols represent mean values and the
vertical bars indicate the standard deviation of duplicate determina-
tions The lines represent simulation using Equations 3 and 4 accord-
ing to the model for MF metabolism in-vitro (Figure 7)
0
1
2
3
4
5
0 10 20 30 40 50 60 70
MF disappearance
M formation
v (n
mo
lmin
(mg
pro
tein
)ndash1
1ndash
MF conc ( )mM
Figure 3 The rates of mometasone furoate (MF) (C0 ˆ 02 iexcl 65 middotM)
disappearance and 6shy -hydroxy-MF (M) formation following incuba-
tion in pooled rat liver microsomes (protein 02mg mLiexcl1) at 37 macrC
Symbols represent experimental data and the lines were fitted by a
biphasic Michaelis-Menten model (Equation 7 Vmax1 ˆ 109nmol
miniexcl1 (mg protein)iexcl1 Km1 ˆ 132middotM CL2 ˆ 0054) and simple
Michaelis-Menten equation (Vmax ˆ 124 Km ˆ 215middotM) for MF
and M respectively The concentration of M formed is expressed
relative to the MF concentration assuming equal HPLCUV
response
Rat metabolism and degradation kinetics of mometasone furoate 623
Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

Characters in brackets represent the concentrations ofparent drug or product at time t while [X]0 represents thederivates of X with respect to t
This model was tested by fitting the experimental dataof MF obtained from experiments in rat liver microsomeswith curve-fitting performed using SCIENTIST Goodcorrelation (r2 gt 099) was obtained between the observedand the predicted values of MF remaining A typical plotwith both experimental and theoretical data is shown inFigure 2
A better estimation of the 6shy -hydroxy-MF formationrate (kFM) was achieved using Equations 3 and 4 com-pared with the estimation using the apparent 6shy -hydroxy-MF concentration in the initial phase For example byfitting the observed data to these equations the relation-ship between the microsomal protein concentration andthe reaction rate constant for both total loss of MF(kFM Dagger kFN) and formation of 6shy -hydroxy-MF (kFM)
was closer to the real situation In comparison the appar-ent rates of 6shy -hydroxylation obtained from the slopes ofinitial phase of 6shy -hydroxy-MF formation in the concen-trationplusmntime course were underestimated by 29 sect 11The result was that the apparent contribution of 6shy -hydroxylation was averaged at only sup130 of the totalloss of MF given by the slopes of the linear regressionlines
As indicated by the kinetic data in Figure 3 in rat livermicrosomes the relationship between the rate of 6shy -hydroxylation and the initial concentration of MF wasclosely correlated with a monophasic Michaelis-Mentenkinetics (Equation 5) with Km of 124 middotM and Vmax of215 nmol miniexcl1 (mg protein)iexcl1 respectively (r2 ˆ 09943MSC ˆ 34 SCIENTIST) whereas the disappearance ofthe parent drug exhibited atypical Michaelis-Mentenkinetics
v ˆ hellipVmaxpermilMFŠdagger=hellipKm Dagger permilMFŠdagger hellip5dagger
The hyperbolic Michaelis-Menten kinetics for the 6shy -hydroxylation was verified by the corresponding Eadie-Hofstee plot which was nearly linear (Figure 8) Thissuggests that the reaction involved one enzyme or severalenzymes with similar values of Km in rat liver microsomesIn comparison the Eadie-Hofstee plot for the loss of theparent drug from the reaction media was clearly concavein nature which is the feature of allosteric-enzyme kineticsor multiple-enzyme kinetics The studies using enzymeinhibitors have demonstrated the possibility of involve-ment of more than one enzyme in MF metabolismAdditionally as displayed in Figure 3 after 6shy -hydroxy-lation was saturated at high drug concentrations MF wasstill metabolized via other routes that were not saturatedunder the experimental conditions This provided theinformation that the catalysing components towards dif-ferent pathways probably had distinct affinities and capa-cities Thus the multiple-enzyme model (Equation 6)rather than the Hill equation would be more appropriatefor describing these kinetic processes
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger Vmax2 cent permilMFŠ
Km2 Dagger permilMFŠ Dagger hellip6dagger
Table 1 Rate of 6shy -hydroxy-mometasone furoate (M) formation and the effect of
1-aminobenzotriazole (200middotM) following incubation of mometasone furoate (MF) (C0 ˆ 4 middotM
n ˆ 3) in subcellular fractions of rat liver
Subcellular fraction of rat liver Rate of M formation (pmol miniexcl1 (mg protein)iexcl1)
Original rate In the presence of 1-aminobenzotriazole
Microsomes 65835sect 2880 14478sect 096
Cytosol 269sect 041 094sect 021
Mitochondria 2839sect 211 1315sect 008
Nuclei 1069sect 041 374sect 019
M formed is expressed relative to the concentration of MF assuming equal HPLCUV response
Rate of M formation was calculated from the slope of the initial linear phase of M formed
0
20
40
60
80
100
0 1 2 3 4 5Time (h)
MF in buffer MF in tissue MF in sac M in buffer M in tissue M in sac
a
s in
itia
l MF
in b
uff
er
Figure 4 Typical time course for mometasone furoate (MF) and its
major metabolite 6shy -hydroxy-MF (M) in the K-H buffer incubated
with everted rat intestinal tissue Everted intestinal tissue of rat no 6
was incubated with MF (C0 ˆ 19 middotM ˆ 10 middotg mLiexcl1) in 25 mL K-H
buffer at 37 macrC Estimation was made from the peak area ratio of
MF and M in samples remaining in the incubation media at each
sampling point relative to the initial value of MF assuming equal
HPLCUV absorbance between MF and M and not taking account
of the loss as a result of sampling Symbols represent the mean values
of duplicate experimental data
624 X W Teng et al
When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

When multiple enzymes are involved in the reaction andthe low affinity enzymes possess high capacity that is notsaturable under the experimental conditions Equation 6can be simplified to Equation 7
cedil ˆ Vmax1 cent permilMFŠKml Dagger permilMFŠ Dagger CLi cent permilMFŠ hellip7dagger
where CLi represents VmaxKm the intrinsic clearance of adrug due to the enzymes having low affinity but highcapacity Equation 7 has one parameter less thanEquation 6 thus estimation of parameters for the high-affinity enzyme(s) can be more reliable Since in-vivo drugconcentrations can rarely exceed the Km of the low-affin-ity enzyme(s) the high-affinity process usually accounts
0
2
4
6
8
10
12
ndash4 ndash2 0 2 4 6 8 10 12
Ketoconazole (m )M
ndash Ki
96
32
16
A
0
2
4
6
8
10
12
ndash06 ndash04 ndash02 0 02 04 06
0
01
05
210
[I] (m )MB
ndash1Km
ndash1Kp
1V 1Vmax p=
[MF] (m )M
1[MF](m )M
1v
(mg
min
nm
olndash1
)
1v
(m
g m
in n
mo
lndash1)
Figure 5 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by ketoconazole illustrated by (A) Dixon and (B) Lineweaver-
Burk plots MF (16 32 and 96 middotM) was incubated for 8 min at 37 macrC in pooled rat liver microsomes (protein 02 mg mLiexcl1) in the presence of
01 05 2 or 10 middotM ketoconazole Symbols representing parameters Ki inhibition constant Km Michaelis constant Kp Michaelis constant in
the presence of enzyme inhibitor Vp maximum velocity in the presence of enzyme inhibitor
10
100
0 10 20 30
Pre-incubation time (min)
0
20
100
500
[I] ( )mM
slope = ndashkiobs
A
0 100 200 300 400 500 600
[I] (mM)
0
001
002
003
004
005
Kicent
05kimax
B
o
f co
ntr
ol
acti
vity
k(m
in)
lo
bs
ndash1
Figure 6 Inhibitory effect of mometasone furoate (MF) 6shy -hydroxylation by troleandomycin shown by (A) the catalysing activity of rat
liver microsomes to 6shy -hydroxylation of MF in the presence of troleandomycin at various concentrations versus pre-incubation time relative
to those of controls and (B) the values of the apparent inactivation rate constant (kiobs) versus the initial concentrations of troleandomycin
The parameters kimax and K0i were estimated using Equation 2 by the least-square method Incubation of MF (C0 ˆ 16 middotM) was carried out for
8 min at 37 macrC after pre-incubation of troleandomycin (20 100 and 500middotM) in pooled rat liver microsomes (protein 02 mg mLiexcl1) for 0 5 15
and 30 min
Rat metabolism and degradation kinetics of mometasone furoate 625
for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

for the majority of net intrinsic clearance Thereforecharacterization of the high-affinity process is generallyof most clinical importance
When the kinetic data set of the parent drug metabo-lism was tested with Equation 7 a good correlation withthe simulated data was obtained (Figure 3 r2 ˆ 09927MSC ˆ 36 SCIENTIST) Moreover the kinetic para-meters for the high-affinity catalyst (Vmax1 and Km1)were comparable with those for 6shy -hydroxylation (Table2) The intrinsic clearance of MF via the low-affinityenzymatic reaction was approximately 65 of thatthrough the high-affinity metabolic pathway 6shy -hydro-xylation
In addition to the Eadie-Hofstee plots the biphasic andmonophasic Michaelis-Menten models were assessedusing plots from transformed kinetic data for the totalloss of MF and the specific loss due to 6shy -hydroxylationrespectively The Lineweaver-Burk plot illustrating sub-strateplusmnactivity relationships (1v vs 1[MF]) became a
curvilinear line for MF disappearance but formed analmost straight line for 6-hydroxy-MF formation Thekinetic parameters (Table 2) for the catalysing enzymeswere estimated from the intercepts shown on the graphsThese data suggest the presence of multiple enzymes ormultiple isoforms of the enzyme involved in the in-vitrometabolism of MF in rat liver microsomes in which asingle enzyme or enzymes of similar affinity are responsi-ble for its 6shy -hydroxylation with the remainder for otherreaction(s) Furthermore the Hanes plot ([MF]v ratioversus [MF]) for the total loss of MF also displayedbiphasic nature whereas that for the specific loss via6-hydroxylation was again apparently monophasic in nat-ure The kinetic parameters for the enzyme systems dis-playing different roles (Table 2) were estimated from theslopes and intercepts on the plots From the nature of theplots it was also evident that the in-vitro metabolism ofMF was mediated by multiple enzymes of distinct affinityand capacity while the 6shy -hydroxylation was apparentlycatalysed by a single enzyme or enzymes of similar affinity
The values of kinetic parameters estimated for MF 6shy -hydroxylation from the three plots were rather close tothose generated using the simple Michaelis-Menten equa-tion based on the experimental data (Table 2) For thedisappearance of the parent drug the parameters derivedfrom the lines arbitrated to two-enzyme kinetics were alsosimilar among the three plots However there was a dis-crepancy between the values obtained from these plotsand those from the biphasic kinetics (Equation 7) exceptfor Vmax of the high-affinity process It is known thatmonophasic kinetics does not mean that a reaction is
MF
M
N
OkFM kMO
kFN
Figure 7 The scheme for the in-vitro biotransformation of mome-
tasone furoate (MF) in rat liver microsomes kFM kFN and kM O are
the rate constants of corresponding reactions
(A) MF disappearance
0
1
2
3
4
5
6
7
ndash05 0 05 1 15 2
Vmax2
slope = ndashKm2
Vmax1
(B) M formation
0
02
04
06
08
1
12
14
0 01 02 03 04
Vmax
slope = ndashKm
slope = ndashKm1
v[MF] (nmol min (mg protein) MF)ndash1 ndash1 mM
v (n
mo
l min
(mg
pro
tein
))
ndash1ndash1
Figure 8 Eadie-Hofstee plots for (A) mometasone furoate (MF) disappearance and (B) 6shy -hydroxy-MF (M) formation following the
incubation of MF of initial concentration ranging from 02 to 65 middotM in pooled rat liver microsomes (protein 02 mg mLiexcl1) at 37 macrC The lines in
graph (1) were fitted by y ˆ iexcl 0288xDagger 08614 (r2 ˆ 07905) and y ˆ iexcl 30245xDagger 60795 (r2 ˆ 09275) respectively and the straight line in graph
(2) was y ˆ iexcl 28436xDagger 11838 (r2 ˆ 09279) The concentration of M formed is expressed relative to the MF concentration assuming equal
HPLCUV response
626 X W Teng et al
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al
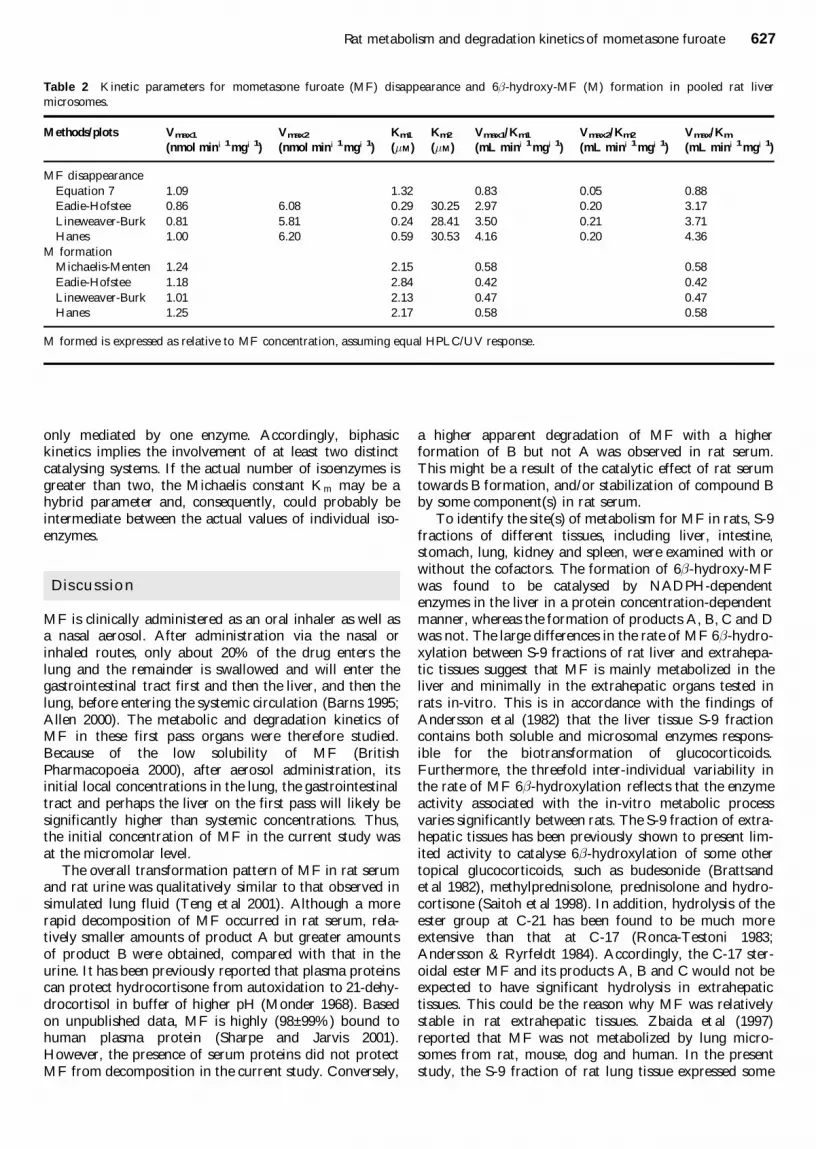
only mediated by one enzyme Accordingly biphasickinetics implies the involvement of at least two distinctcatalysing systems If the actual number of isoenzymes isgreater than two the Michaelis constant Km may be ahybrid parameter and consequently could probably beintermediate between the actual values of individual iso-enzymes
Discussion
MF is clinically administered as an oral inhaler as well asa nasal aerosol After administration via the nasal orinhaled routes only about 20 of the drug enters thelung and the remainder is swallowed and will enter thegastrointestinal tract first and then the liver and then thelung before entering the systemic circulation (Barns 1995Allen 2000) The metabolic and degradation kinetics ofMF in these first pass organs were therefore studiedBecause of the low solubility of MF (BritishPharmacopoeia 2000) after aerosol administration itsinitial local concentrations in the lung the gastrointestinaltract and perhaps the liver on the first pass will likely besignificantly higher than systemic concentrations Thusthe initial concentration of MF in the current study wasat the micromolar level
The overall transformation pattern of MF in rat serumand rat urine was qualitatively similar to that observed insimulated lung fluid (Teng et al 2001) Although a morerapid decomposition of MF occurred in rat serum rela-tively smaller amounts of product A but greater amountsof product B were obtained compared with that in theurine It has been previously reported that plasma proteinscan protect hydrocortisone from autoxidation to 21-dehy-drocortisol in buffer of higher pH (Monder 1968) Basedon unpublished data MF is highly (98plusmn99) bound tohuman plasma protein (Sharpe and Jarvis 2001)However the presence of serum proteins did not protectMF from decomposition in the current study Conversely
a higher apparent degradation of MF with a higherformation of B but not A was observed in rat serumThis might be a result of the catalytic effect of rat serumtowards B formation andor stabilization of compound Bby some component(s) in rat serum
To identify the site(s) of metabolism for MF in rats S-9fractions of different tissues including liver intestinestomach lung kidney and spleen were examined with orwithout the cofactors The formation of 6shy -hydroxy-MFwas found to be catalysed by NADPH-dependentenzymes in the liver in a protein concentration-dependentmanner whereas the formation of products A B C and Dwas not The large differences in the rate of MF 6shy -hydro-xylation between S-9 fractions of rat liver and extrahepa-tic tissues suggest that MF is mainly metabolized in theliver and minimally in the extrahepatic organs tested inrats in-vitro This is in accordance with the findings ofAndersson et al (1982) that the liver tissue S-9 fractioncontains both soluble and microsomal enzymes respons-ible for the biotransformation of glucocorticoidsFurthermore the threefold inter-individual variability inthe rate of MF 6shy -hydroxylation reflects that the enzymeactivity associated with the in-vitro metabolic processvaries significantly between rats The S-9 fraction of extra-hepatic tissues has been previously shown to present lim-ited activity to catalyse 6shy -hydroxylation of some othertopical glucocorticoids such as budesonide (Brattsandet al 1982) methylprednisolone prednisolone and hydro-cortisone (Saitoh et al 1998) In addition hydrolysis of theester group at C-21 has been found to be much moreextensive than that at C-17 (Ronca-Testoni 1983Andersson amp Ryrfeldt 1984) Accordingly the C-17 ster-oidal ester MF and its products A B and C would not beexpected to have significant hydrolysis in extrahepatictissues This could be the reason why MF was relativelystable in rat extrahepatic tissues Zbaida et al (1997)reported that MF was not metabolized by lung micro-somes from rat mouse dog and human In the presentstudy the S-9 fraction of rat lung tissue expressed some
Table 2 Kinetic parameters for mometasone furoate (MF) disappearance and 6shy -hydroxy-MF (M) formation in pooled rat liver
microsomes
Methodsplots Vmax1 Vmax2 Km1 Km2 Vmax1Km1 Vmax2Km2 VmaxKm
(nmol miniexcl1mgiexcl1) (nmol miniexcl1mgiexcl1) (middotM) (middotM) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1) (mL miniexcl1mgiexcl1)
MF disappearance
Equation 7 109 132 083 005 088
Eadie-Hofstee 086 608 029 3025 297 020 317
Lineweaver-Burk 081 581 024 2841 350 021 371
Hanes 100 620 059 3053 416 020 436
M formation
Michaelis-Menten 124 215 058 058
Eadie-Hofstee 118 284 042 042
Lineweaver-Burk 101 213 047 047
Hanes 125 217 058 058
M formed is expressed as relative to MF concentration assuming equal HPLCUV response
Rat metabolism and degradation kinetics of mometasone furoate 627
ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

ability in catalysing metabolism of MF The discrepancymay have resulted from differences in experimental con-ditions and assay methods
To elucidate the intracellular site of MF metabolism inrat liver incubations of MF were further performed innuclear mitochondrial cytosolic and microsomal frac-tions of liver tissue The results indicated that rat livermicrosomes were the primary host of enzymes responsiblefor MF 6shy -hydroxylation To further investigate the con-tribution of individual CYP isoenzymes to MF 6shy -hydro-xylation in rat liver microsomes a number of potent andor selective chemical inhibitors andor substrates used inprevious studies were employed The competitive inhibi-tory effect by ketoconazole was clearly indicated by thecharacteristic intersecting points of the fitted lines in theDixon Lineweaver-Burk and Eadie-Hofstee plots as wellas the parallel lines in the Cornish-Bowden plots In thepresent investigation the apparent Ki is roughly equival-ent to the degree of potency of ketoconazole in the inhibi-tion of testosterone 6shy -hydroxylation in rat livermicrosomes (IC50 ˆ 029 middotM Eagling et al 1998) In addi-tion the apparent Ki here showed little dependence on the100-fold ketoconazole concentration as indicated by thelow standard deviation This implied that the inhibitoryeffect present at low concentrations of ketoconazole wasnot confounded by its increased concentration Thereforethe reaction could be mainly catalysed by CYP3A Inaddition the concentration and pre-incubation time-dependent inhibition by troleandomycin (Figure 6A) aselective inhibitor of rat liver CYP3A (Delaforge et al1988) further suggested the contribution of CYP3A Theconcentration-dependent inhibition by diazepam might beowing to its competitive effect on CYP3A which contri-butes to diazepam C3-hydroxylase in rat liver (Neville et al1993) However the possible involvement of CYP2C11could not be simply discounted since diazepam is also asubstrate of CYP2C11 for diazepam N-demethylation inrat liver microsomes (Neville et al 1993) The inhibitoryeffect by 4-methylpyrazole at very high concentrationsimplied possible involvement of CYP2E1 (Yamazakiet al 1992) andor 2D1 (Clarke 1998) the equivalentenzyme in the rat to human CYP2D6 (Umehara et al1997) The involvement of CYP2D1 however was notsupported by the test with quinidine The latter is a select-ive and potent CYP2D1 inhibitor with an effective con-centration of 5 middotM (Umehara et al 1997) but did notdisplay marked inhibition at concentrations as high as100 middotM Owing to the low selectivity of diethyldithiocar-bamate in rat liver microsomes (Eagling et al 1998) it wasnot able to distinguish the role of individual CYP isoforms(CYP1A2 2C 2E1 and 3A) in this reaction although itcaused marked inhibition of MF 6shy -hydroxylationAmong the related CYP families of this inhibitorCYP1A2 appeared not to be involved because no signifi-cant inhibition was obtained with high concentrations offurafylline an inhibitor of CYP1A2 in rat liver micro-somes (Hu amp Bunce 1999) In addition CYP2A appearednot to be responsible for the in-vitro MF 6shy -hydroxyla-tion in rat liver since coumarin a substrate of rat CYP2A3(Honkakoski amp Negishi 1997) did not affect the reaction
The results of the chemical inhibition study suggest thatCYP3A can be the major isoenzyme mediating MF 6shy -hydroxylation in rat liver while CYP1A2 2A3 2C112D1 were not likely to be involved The contribution ofother CYP isoforms such as CYP2B 2E1 and 2C11 wasnot clearly defined Further studies are needed to verifythese findings and to further clarify the responsibility ofindividual CYP isoforms through reaction phenotypingusing antibodies against individual rat CYP isoformspurified or cDNA expressed enzymes and through corre-lation studies using inter-individual variability in expres-sion of the responsible CYP isoforms Although there aresome species differences in the CYP isoform responsiblefor xenobiotic metabolism (Correia 1995 Eagling et al1998) the involvement of CYP3A in 6shy -hydroxylationof steroids has been observed in both rats and humans(Clarke 1998 Eagling et al 1998 Kenworthy et al 1999Obach et al 2001) The preliminary results here may throwsome light on the subsequent study and identification ofhuman CYP isoforms contributing to MF metabolism
Whereas limited metabolism of MF was observed inS-9 and microsomal fractions of rat intestinal tissue con-siderable metabolism of MF was observed with everted ratintestine This agrees with the previous finding that MF ismetabolized moderately in everted intestines of rat mouseand dog with formation of 6shy -hydroxy-MF (Zbaida et al1997) These results are also consistent with the knowledgethat everted sacs of mouse small intestine exhibited a highcatalytic activity in the metabolism of another steroidtestosterone (Emoto et al 2000) These findings indicatethat the small intestine could also be a site of pre-systemicmetabolism for MF Additionally an important finding isthat the rapid disappearance of the MF from the incuba-tion buffer was not solely due to metabolism of MF by theintestinal tissue In fact while MF was metabolized by theeverted intestine the parent drug and the metaboliteformed were taken up rapidly into the intestinal tissueduring the incubation At the end of the incubation (5 h)the majority of MF (sup188) and 6shy -hydroxy-MF (sup180)remained in the intestinal tissue and small amounts ofboth the parent drug (sup13) and the metabolite (sup14)were detected in buffer in the sac Thus the loss of MFfrom the incubation buffer was primarily a result ofuptake by the intestinal tissue and secondly metabolismThe rapid drug uptake by tissue could be due to bindingto tissue proteins partitioning into the adipose portion ofthe tissue and membrane lipid bilayer because of thelipophilic nature of both MF and the metabolite
In summary the present in-vitro kinetic studies revealthe qualitative and quantitative differences in the in-vitrobiotransformation profiles of MF in different rat tissuesIn rat urine and serum MF degrades to A B C and Dfollowing the chemical degradation pathways of MF thatalso observed in buffer solutions In-vitro MF is metabol-ized rapidly and extensively in rat liver minimally in ratextrahepatic tissues tested Under the experimental condi-tions 6shy -hydroxy-MF is the primary metabolite of MFFurthermore in rat liver microsomes are the majorhost of enzymes mediating this biotransformation Inparticular CYP3A is the major CYP 450 enzyme respons-
628 X W Teng et al
ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

ible for this in-vitro biotransformation of MF in rat livermicrosomes Studies on enzyme kinetics of the in-vitrometabolism of MF indicate that the overall metabolicprocess in rat liver microsomes follows biphasicMichaelis-Menten kinetics while 6shy -hydroxylation obeysmonophasic Michaelis-Menten kinetics The kinetic para-meters derived from the kinetic models along with theenzyme inhibition studies suggest that MF is mainly meta-bolized by 6shy -hydroxylation mediated by CYP3A primar-ily and also biotransformed via other pathway(s)catalysed by other enzymes in rat liver in-vitro In theex-vivo studies with rat tissues MF is moderately meta-bolized in the intestine with 6shy -hydroxy-MF as theprimary metabolite In addition tissue uptake was foundto account for the major loss of this drug from the incuba-tion media
In conclusion this pre-clinical knowledge may clarifyhow MF is metabolized in humans given the potentialsuitability of rat as a model for studying the metabolickinetics of MF Furthermore the ability to optimize andgenerate metabolite and degradation products of MF hasbeen established Further studies on MF metabolismkinetics in human and on the pharmacological aspects ofthe degradation and metabolic products are ongoing
References
Affrime M B Cuss F Padhi D Wirth M Pai S ClementR P Lim J Kantesaria B Alton K Cayen M N (2000a)Bioavailability and metabolism of MF following administra-tion by metered-dose and dry-powder inhalers in healthyhuman volunteers J Clin Pharmacol 40 1227plusmn1236
Affrime M B Kosoglou T Thonoor C M Flannery B EHerron J M (2000b) Mometasone furoate has minimaleffects on the hypothalamic-pituitary-adrenal axis when deliv-ered at high doses Chest 118 1538plusmn1546
Allen D B (2000) Systemic effects of intranasal steroids anendocrinologistrsquos perspective J Allergy Clin Immunol 106S179plusmnS190
Andersson P Ryrfeldt A (1984) Biotransformation of thetopical glucocorticoids budesonide and beclomethasone17not21-dipropionate in human liver and lung homogenateJ Pharm Pharmacol 36 763plusmn765
Andersson P Edsbacker S Ryrfeldt A von Bahr C (1982)In vitro biotransformation of glucocorticoids in liver and skinhomogenate fraction from man rat and hairless mouse JSteroid Biochem 16 787plusmn795
Barnes P J (1995) Drug therapy inhaled glucocorticoids forasthma N Engl J Med 332 868plusmn875
Brattsand R Kallstrom L Nilsson E Ryrfeldt A (1982)The lung disposition of budesonide in guinea pig and ratEur J Respir Dis (Suppl) 122 263plusmn265
British Pharmacopoeia (2000) Mometasone furoate Medicinaland pharmaceutical substances In British PharmacopoeiaVol 1 General Medical Council Constable amp Co Londonpp 1065plusmn1066
CarcilloJ A KorzekwaK R JonesG S Parise R A GillespieD G Whalen M J Kochanek P M Branch R A KostC K Jr (1998) The cytochrome P450 suicide inhibitor1-aminobenzotriazole sensitizes rats to zymosan-induced toxicityRes Commun Mol Pathol Pharmacol 102 57plusmn68
Clarke S E (1998) In vitro assessment of human cytochromeP450 Xenobiotica 28 1167plusmn1202
Correia M A (1995) Rat and human liver cytochromes P450In Ortiz de Montellano P R (ed) Cytochrome P450 struc-ture mechanism and biochemistry 2nd edn Plenum Press NewYork pp 607plusmn630
Delaforge M Sartori E Mansuy D (1988) In vivo and invitro effects of a new macrolide antibiotic roxithromycin onrat liver cytochrome P-450 comparison with troleandomycinand erythromycin Chem Biol Interact 68 179plusmn188
Doumas B T Bayse D D Carter R J Peters T JSchaffer R (1981) A candidate reference method for determin-ation of total protein in serum I Development and validationClin Chem 27 1642plusmn1650
Eagling V A Tjia J F Back D J (1998) Differential selec-tivity of cytochrome P450 inhibitors against probe substratesin human and rat liver microsomes Br J Clin Pharmacol45 107plusmn114
Ekins S MaEgrave enpaEgrave aEgrave J Wrighton S A (1999) In vitro metabo-lism subcellular fractions In Woolf T F (ed) Handbook ofdrug metabolism Marcel Dekker New York pp 363plusmn399
Emoto C Yamazaki H Yamasaki S Shimada NNakajima M T Y (2000) Use of everted sacs of mousesmall intestine as enzyme sources for the study of drug oxida-tion activities in vitro Xenobiotica 30 971plusmn982
Esposito G Csaky T Z (1974) Extracellular space in theepithelium of rats small intestine Am J Physiol 226 50plusmn55
Honkakoski P Negishi M (1997) The structure function andregulation of cytochrome P450 2A enzymes Drug Metab Rev29 977plusmn996
Hu K Bunce N J (1999) Metabolism of polychlorinateddibenzo-p-dioxins by rat liver microsomes J Biochem MolToxicol 13 307plusmn315
Ito K Iwatsubo T Kanamitsu S Ueda K Suzuki HSugiyama Y (1998) Prediction of pharmacokinetic alterationscaused by drugplusmndrug interactions metabolic interaction in theliver Pharmacol Rev 50 387plusmn412
Kenworthy K E Bloomer J C Clarke S E Houston J B(1999) CYP3A4 drug interactions correlation of 10 in vitroprobe substrates Br J Clin Pharmacol 48 716plusmn727
Mathews J M Dostal L A Bend J R (1985) Inactivationof rabbit pulmonary cytochrome P-450 in microsomes andisolated perfused lungs by the suicide substrate 1-amino-benzotriazole J Pharmacol Exp Ther 235 186plusmn190
Monder C (1968) Stability of corticosteroids in aqueous solu-tions Endocrinology 82 318plusmn326
Neville C F Ninomiya S Shimada N Kamataki TImaoka S Funae Y (1993) Characterization of specificcytochrome P450 enzymes responsible for the metabolism ofdiazepam in hepatic microsomes of adult male rats BiochemPharmacol 45 59plusmn65
Obach R S Zhang Q-Y Dunbar D Kaminsky L S (2001)Metabolic characterization of the major human small intest-inal cytochrome P450s Drug Metab Dispos 29 347plusmn352
Ocuma T Sato R (1964) The carbon monoxide binding pig-ment of liver microsomes I Evidence for its hemoproteinnature J Biol Chem 23 2370plusmn2378
Onrust S V Lamb H M (1998) Mometasone furoateA review of its intranasal use in allergic rhinitis Drugs 56725plusmn745
Pedersen P L Greenawalt J W Reynafarje B Hullihen JDecker G L Soper J W Bustamente E (1978)Preparation and characterization of mitochondria andsubmitochondrial particles of rat liver and liver-derivedtissues Methods Cell Biol 20 411plusmn481
Rat metabolism and degradation kinetics of mometasone furoate 629
Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al

Pickering H Pitcairn G R Hirst P H Bacon P RNewman S P Affrime M B Marino M (2000) Regionallung deposition of a technetium 99m-labeled formulation ofmometasone furoate administered by hydrofluoroalkane 227metered-dose inhaler Clin Ther 22 1483plusmn1493
Prakash A Benfield P (1998) Topical mometasone A reviewof its pharmacological properties and therapeutic use in thetreatment of dermatological disorders Drugs 55 145plusmn163
Ronca-Testoni S (1983) Hydrolysis of cyclomethasone by thehuman lung Int J Clin Pharmacol Res 3 17plusmn20
Saitoh H Hatakeyama M Eguchi O Oda M Takada M(1998) Involvement of intestinal P-glycoprotein in therestricted absorption of methylprednisolone from rat smallintestine J Pharm Sci 87 73plusmn75
Sharpe M and Jarvis B (2001) Inhaled mometasone furoate areview of its use in adults and adolescents with persistentasthma Drugs 61 1325plusmn1350
Teng X W (2002) Metabolism and degradation of mometasonefuroate PhD Thesis University of Sydney
Teng X W Foe K Brown K F Cutler D J Davies N M(2001) High-performance liquid chromatographic analysis of
mometasone furoate and its decomposition products applica-tion to in vitro degradation studies J Pharm Biomed Anal26 313plusmn331
Umehara K Kudo S Odomi M (1997) Involvement ofCYP2D1 in the metabolism of carteolol by male rat livermicrosomes Xenobiotica 27 1121plusmn1129
Wang C J Tian Z Byrnes K Lin C C (1992) A competi-tive enzyme immunoassay for the direct determination ofmometasone furoate (SCH 32088) in human plasma JPharm Biomed Anal 10 473plusmn479
Yamazaki H Oda Y Funae Y Imaoka S Inui YGuengerich F P Shimada T (1992) Participation ofrat liver cytochrome P450 2E1 in the activation ofN-nitro sodimethylamine and N-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressingSalmonella typhimurium strain (NM2009) Carcinogenesis 13979plusmn985
Zbaida S Shannon D Du Y Lu X Ng K Chowdhury SBlumenkrantz N Patrick J Cayen M N (1997) In vitrometabolism of mometasone furoate (Abstract no P339)FASEB 11 (8 Suppl S) A829
630 X W Teng et al





























