Embed Size (px)
Citation preview

doi:10.1016/j.jmb.2005.03.072 J. Mol. Biol. (2005) 349, 814–824
Llama Single Domain Antibodies as a Tool for MolecularMimicry
Laura M. Zarebski, Mariela Urrutia and Fernando A. Goldbaum*
Fundacion Instituto LeloirConsejo Nacional deInvestigaciones Cientıficas yTecnicas and Facultad deCiencias Exactas y NaturalesUniversidad de Buenos AiresBuenos Aires, Argentina
0022-2836/$ - see front matter q 2005 E
Abbreviations used: CDR, compledetermining region; mAb, monocloenzyme-linked immunosorbent assabinding fragment; VHH, hcIgG varnormal mouse serum; Id, idiotype osurface plasmon resonance.E-mail address of the correspond
In camelids, a subset of the immunoglobulins consists of heavy-chainhomodimers devoid of light chains, and are thus called heavy-chain IgGs(hcIgGs). Their variable region (VHH) is the smallest antigen-bindingfragment possible, and being just one polypeptide chain it is especiallysuitable for engineering. In particular, camelid single domain antibodiesmight be very useful for molecular mimicry and anti-idiotypic vaccination.In the present work, we show that llamas immunized with an anti-DNAmouse mAb develop an important anti-Id response. Selection of VHHs byphage display, with specific elution of bound phages with the externalantigenic DNA, shows that selected private anti-Id VHHs compete forbinding to the external antigen and bear a functional mimicry of the DNA.These results indicate that llama anti-Id single domain antibodies would bean excellent tool for molecular mimicry studies.
q 2005 Elsevier Ltd. All rights reserved.
Keywords: anti-idiotypic antibodies; camelid IgGs; phage display; DNAmimicry; anti-DNA immunity
*Corresponding authorIntroduction
The complementarity-determining regions (CDR)of antibodies constitute its antigen combining sites.The combining sites of antibodies are large exposedareas on the surface of the variable domains anddisplay great structural diversity. As a consequence,under appropriate experimental conditions, thesehighly variable determinants give rise to humoralimmune responses in singeneic or xenogeneicsystems. The antibodies elicited against the com-bining site are called anti-idiotypic (anti-Id) andreact with antigenic determinants, the idiotopes.1,2
The sum of idiotopes of an antibody constitutes itsidiotype (Id).
Given the fact that external antigens and anti-Idantibodies compete for binding to their specificantibody, idiotopes are thought to share partiallyor completely its reactive surface with the com-bining site.3 Also, as the external antigen and theanti-Id both bind to the same surface, they are
lsevier Ltd. All rights reserve
mentarity-nal antibody; ELISA,y; Fab, antigen-iable region; NMS,r idiotypic; SPR,
ing author:
thought to share structural similarity. This idea hasled to proposals that anti-Id antibodies mayresemble the external antigen, thus mimicking itsstructure.2
This “popular” idea has been the base of anenormous amount of work from different groups.4
For example, functional mimicry of ligands ofbiological receptors by anti-Id has been describedin several systems.5 Besides, functional mimicry ofexternal antigens has led to proposals to use anti-Idas surrogate antigens.6
Anti-idiotypic antibodies have been character-ized structurally.7–15 As any conventional antibody,their recognition surfaces are large flat areascomposed of all six CDRs from heavy and light-chain variable domains. This fact seriously limitstheir use for mimicry of small ligands, carbo-hydrates and nucleic acids that interact withcavities or crevices of their target proteins.7 Incamelids, a subset of the immunoglobulins consistsof heavy-chain homodimers devoid of light chains,and are thus called heavy-chain IgGs (hcIgGs).Their variable region (VHH) is the smallest antigen-binding fragment possible, and being just onepolypeptide chain it is especially suitable forengineering. In particular, their different strategyof binding, longer CDR3 loops protruding from thebinding site and the deviation of CDR confor-mations from the equivalent human or mouse loopstructures, suggest that camelid single domain
d.

Figure 2. Sera reactivity is specific for ED84 idiotopes.Reactivity of llama serum, with or without absorption tonormal mouse IgG, was determined by ELISA against apanel of Fab fragments. A 1:100 (v/v) dilution of serumafter second immunization was used.
DNA Mimicry by Single Domain Antibodies 815
antibodies might be very useful for molecularmimicry and anti-idiotypic vaccination.16
We have recently described two monoclonalantibodies with unusual high affinity and specificityagainst a double-stranded oligonucleotide of 18 bp,Site35.17,18 This novel protein–DNA interactionconstitutes a challenging system for molecularmimicry using anti-Id strategy. In the presentwork we show that llamas immunized withmouse IgG1 ED84, one of the anti-DNAmonoclonalantibodies (mAbs), develop an important anti-Idresponse using both conventional and hcIgGs.Selection of VHHs by phage display using specificelution strategies19 rendered two types of anti-Idsingle domain antibodies, directed against public orprivate idiotopes. Furthermore, a selected anti-IdVHH competes for binding to the external antigenicoligonucleotide and should bear a functionalmimicry of the DNA. These results indicate thatllama anti-Id single domain antibodies would be anexcellent tool for molecular mimicry studies.
Results
Lama glama anti-Id response is present inconventional and hcIgG antibodies
In order to prove the efficacy of the immunizationprotocol, sera from two llamas prior to immuniz-ation and after two and three boosts with ED8417,18
were analyzed by ELISA. When assayed against thewhole ED84 molecule, sera from immunizedanimals showed a strong IgG response, with titersreaching 1/30,000 after three immunizations, asshown in Figure 1 for llama #8338.
We assayed anti-idiotypic specificity for ED84 byimmunodepletion of antibodies against constantregion and other shared IgG epitopes. An ELISAassay was performed to compare the reactivityagainst different mouse Fab fragments of the
Figure 1. Lama glama IgG response against ED84. Serialdilutions of preimmune and post-immunization serumwere analyzed by ELISA against 1 mg of ED84 IgG.
absorbed and non-absorbed sera. As shown inFigure 2 (ED84 bars), recognition of ED84-specificepitopes is as efficient as recognition of the wholeFab molecule, suggesting that there is an importantresponse against ED84 idiotopes (i.e. anti-Id84antibodies).Recognition of ED10,17,18 an IgG1 identical with
ED84 in specificity and very similar in heavy chainsequence (see Materials and Methods), is almost asefficient and specific as that of ED84. This resultcould imply that both mouse IgGs share idiotypicdeterminants (or idiotopes) regardless of their lightchain sequence differences, which agrees with theiridentical specificity. An immunodominant heavychain variable region, a scenario seen sometimes inanti-idiotypic responses,20 would explain theseresults. In any case, the difference in reactivitybetween ED84 and ED10 is significant, indicatingthe presence of anti-Id84 antibodies, recognizingprivate ED84 idiotopes exclusively.Recognition of control non-related IgG1 Fabs
ED19 and F10 is about 50% of that with ED84 inintact serum and almost completely abolishedwhen absorbed to normal mouse IgG, indicatingthat a high proportion of anti-idiotypic ED84-specific IgGs were elicited during the immunizationprotocol. These results highlight the possibilityof obtaining anti-Id antibodies that recognizeconformational epitopes for molecular mimicrygeneration.Phage display selection of antibodies is facilitated
by the use of llama hcIgG variable regions (VHH)amplified with specific primers from total lympho-cyte cDNA. This approach avoids the need ofheavy and light chain polypeptides to be combinedin conventional IgG libraries. To asses the presenceof a-Id84 VHHs prior to library construction, weperformed a similar ELISA as above with purified

Figure 3. Specific reactivity against ED84 is present inconventional and heavy chain IgGs. IgGs were purifiedfrom sera after third immunization and assayed by ELISAagainst Fab ED84 and control Fab (ED19), adding 100 ngof llama IgG per well previously coated with 0.5 mg of thecorresponding Fab. Absorption to normal mouse IgG wasperformed as described above. The different reactivityscales in both graphs cannot be ascribed to higher affinityof the conventional fraction, since it could be the result ofa less efficient recognition of the heavy chain only IgG bythe anti-llama IgG rabbit serum. Inset: PAGE showingpurified conventional (C) and heavy chain (HC) IgGs; M,molecular mass marker.
Figure 4. Enrichment in binders after two rounds ofpanning is achieved with both specific and non-specificelution. The size of the output relative to the input isshown. An equal input of approximately 3!1013 phageparticles was used in every case. The output/input ratiofrom non-coated control wells is negligible (%10K10).
816 DNA Mimicry by Single Domain Antibodies
heavy chain only IgG2 and the conventional IgG1
isotypes.As shown in Figure 3, both fractions present
antibodies against constant region and ED84-specific idiotopes. There is not a clear bias againstthese two specificities in these two populations ofantibodies, allowing for the further use of the heavychain only antibodies for the generation of an anti-idiotypic library.
Phage display selection of anti-Id llamaantibodies is facilitated by antigen-specificelution procedures
Once established that anti-Id antibodies arepresent in the hcIgG fraction, we cloned theirvariable regions (VHH) in a phage display system.Out of ten sequenced clones, nine were clearlydifferent (Table 2A), showing a good variability inthe initial pool. No clone without insert or withnon-VHH sequences was among those randomlyselected clones, indicating a good quality of theinitial library. We panned the phages displayingindividual VHHs by its binding to ED84 IgGimmobilized to plastic wells. The elution of thebinders was performed by specific elution ofphages bound to the combining site of ED84, asproposed by Goletz19 for the isolation of anti-idiotypic antibodies, competing their binding withan excess of the cognate antigen for ED84, theoligonucleotide Site35. In a duplicate experiment,
bound phages were eluted by classical, acidic pH,non-specific elution of all the bound phages, tocompare the yield of anti-Id VHHs of both methods.
Two rounds of panning were performed for eachelution protocol. The number of clones eluted inrelation to the size of the input with each protocolafter the first and second round of panning is shownin Figure 4. A similar number is achieved with bothmethods after two rounds. This indicates that, atthis point, specific elution is as effective as non-specific elution and, to avoid loss of variability, wedecided to analyze individual clones for theirreactivity before proceeding to further rounds ofselection.
To compare the number of a-Id84 VHHs eluted,we randomly picked 96 colonies from the secondround of panning of both methods of elution andproduced phages from each of them. Phage ELISAagainst ED84 IgG and the non-related IgG1 ED5were performed for every clone. The differencebetween the reading from both plates for each clone(DA) was calculated and clones showing a DAO0.1were considered as anti-Id clones. This criterion israther permissive, probably overestimating thenumber of real anti-Id clones present, but evenwith this low cut-off there is significant differencebetween both methods; with a more stringent one,the significance is even greater. When we comparedthe media of the DA obtained for all clones in eachgroup, there were also highly significant differ-ences, showing that ED84 specific recognition isachieved more effectively by phages eluted withSite35. Taken together, these results indicate that,with specific elution, we were able to effectivelyselect clones recognizing ED84 by its variableregion. The comparative results are shown inTable 1.
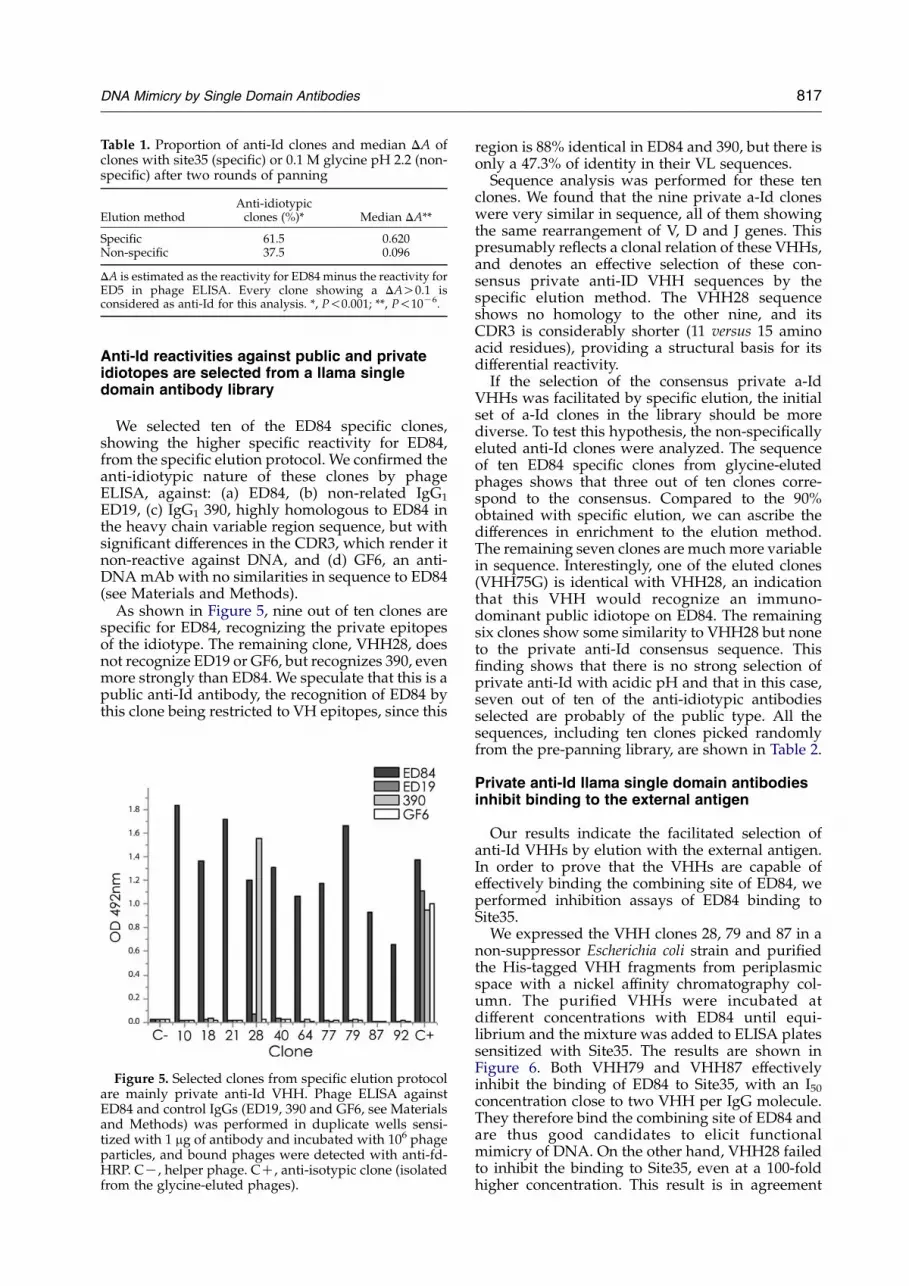
Table 1. Proportion of anti-Id clones and median DA ofclones with site35 (specific) or 0.1 M glycine pH 2.2 (non-specific) after two rounds of panning
Elution methodAnti-idiotypicclones (%)* Median DA**
Specific 61.5 0.620Non-specific 37.5 0.096
DA is estimated as the reactivity for ED84minus the reactivity forED5 in phage ELISA. Every clone showing a DAO0.1 isconsidered as anti-Id for this analysis. *, P!0.001; **, P!10K6.
DNA Mimicry by Single Domain Antibodies 817
Anti-Id reactivities against public and privateidiotopes are selected from a llama singledomain antibody library
We selected ten of the ED84 specific clones,showing the higher specific reactivity for ED84,from the specific elution protocol. We confirmed theanti-idiotypic nature of these clones by phageELISA, against: (a) ED84, (b) non-related IgG1
ED19, (c) IgG1 390, highly homologous to ED84 inthe heavy chain variable region sequence, but withsignificant differences in the CDR3, which render itnon-reactive against DNA, and (d) GF6, an anti-DNA mAb with no similarities in sequence to ED84(see Materials and Methods).
As shown in Figure 5, nine out of ten clones arespecific for ED84, recognizing the private epitopesof the idiotype. The remaining clone, VHH28, doesnot recognize ED19 or GF6, but recognizes 390, evenmore strongly than ED84. We speculate that this is apublic anti-Id antibody, the recognition of ED84 bythis clone being restricted to VH epitopes, since this
Figure 5. Selected clones from specific elution protocolare mainly private anti-Id VHH. Phage ELISA againstED84 and control IgGs (ED19, 390 and GF6, see Materialsand Methods) was performed in duplicate wells sensi-tized with 1 mg of antibody and incubated with 106 phageparticles, and bound phages were detected with anti-fd-HRP. CK, helper phage. CC, anti-isotypic clone (isolatedfrom the glycine-eluted phages).
region is 88% identical in ED84 and 390, but there isonly a 47.3% of identity in their VL sequences.Sequence analysis was performed for these ten
clones. We found that the nine private a-Id cloneswere very similar in sequence, all of them showingthe same rearrangement of V, D and J genes. Thispresumably reflects a clonal relation of these VHHs,and denotes an effective selection of these con-sensus private anti-ID VHH sequences by thespecific elution method. The VHH28 sequenceshows no homology to the other nine, and itsCDR3 is considerably shorter (11 versus 15 aminoacid residues), providing a structural basis for itsdifferential reactivity.If the selection of the consensus private a-Id
VHHs was facilitated by specific elution, the initialset of a-Id clones in the library should be morediverse. To test this hypothesis, the non-specificallyeluted anti-Id clones were analyzed. The sequenceof ten ED84 specific clones from glycine-elutedphages shows that three out of ten clones corre-spond to the consensus. Compared to the 90%obtained with specific elution, we can ascribe thedifferences in enrichment to the elution method.The remaining seven clones are much more variablein sequence. Interestingly, one of the eluted clones(VHH75G) is identical with VHH28, an indicationthat this VHH would recognize an immuno-dominant public idiotope on ED84. The remainingsix clones show some similarity to VHH28 but noneto the private anti-Id consensus sequence. Thisfinding shows that there is no strong selection ofprivate anti-Id with acidic pH and that in this case,seven out of ten of the anti-idiotypic antibodiesselected are probably of the public type. All thesequences, including ten clones picked randomlyfrom the pre-panning library, are shown in Table 2.
Private anti-Id llama single domain antibodiesinhibit binding to the external antigen
Our results indicate the facilitated selection ofanti-Id VHHs by elution with the external antigen.In order to prove that the VHHs are capable ofeffectively binding the combining site of ED84, weperformed inhibition assays of ED84 binding toSite35.We expressed the VHH clones 28, 79 and 87 in a
non-suppressor Escherichia coli strain and purifiedthe His-tagged VHH fragments from periplasmicspace with a nickel affinity chromatography col-umn. The purified VHHs were incubated atdifferent concentrations with ED84 until equi-librium and the mixture was added to ELISA platessensitized with Site35. The results are shown inFigure 6. Both VHH79 and VHH87 effectivelyinhibit the binding of ED84 to Site35, with an I50concentration close to two VHH per IgG molecule.They therefore bind the combining site of ED84 andare thus good candidates to elicit functionalmimicry of DNA. On the other hand, VHH28 failedto inhibit the binding to Site35, even at a 100-foldhigher concentration. This result is in agreement
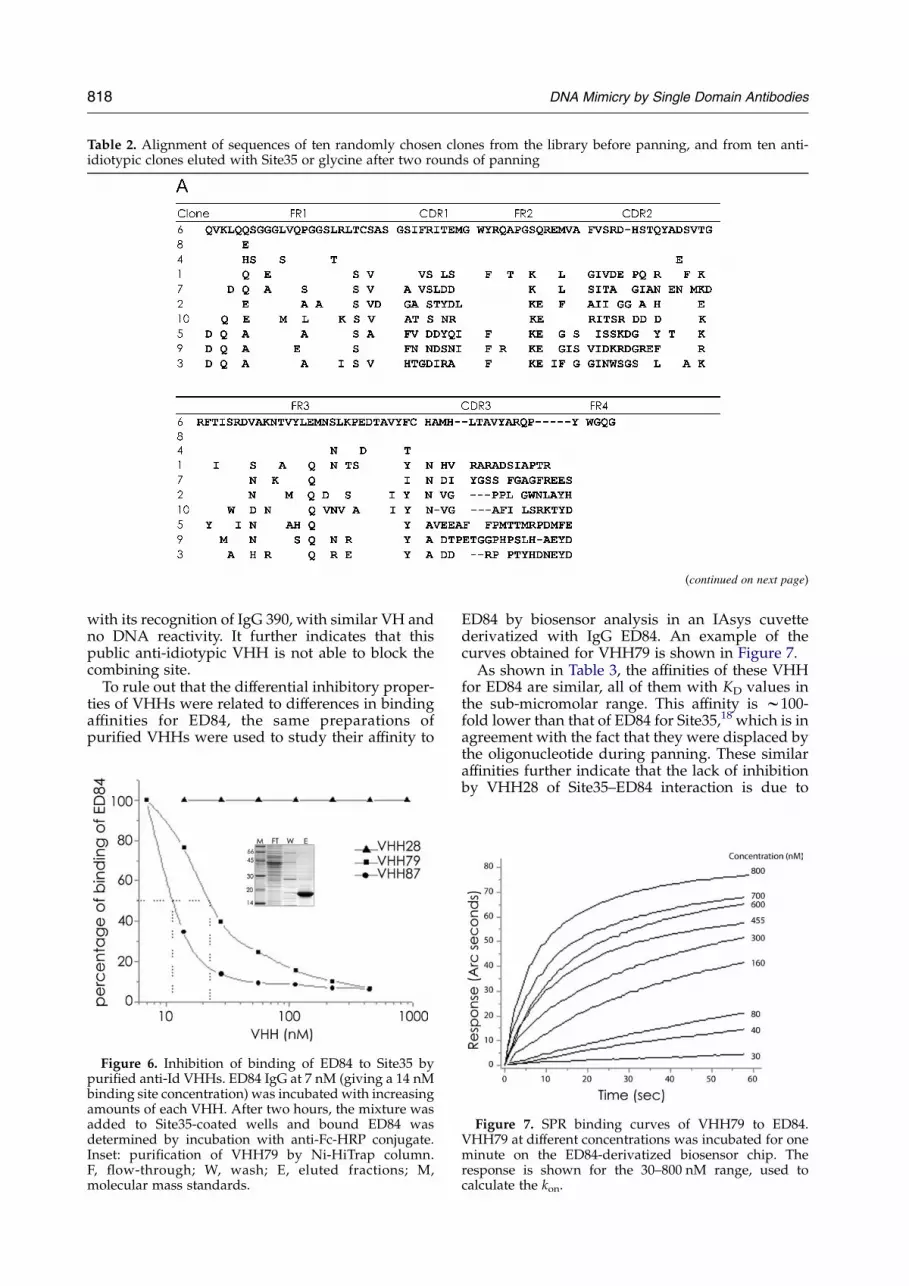
Table 2. Alignment of sequences of ten randomly chosen clones from the library before panning, and from ten anti-idiotypic clones eluted with Site35 or glycine after two rounds of panning
(continued on next page)
818 DNA Mimicry by Single Domain Antibodies
with its recognition of IgG 390, with similar VH andno DNA reactivity. It further indicates that thispublic anti-idiotypic VHH is not able to block thecombining site.
To rule out that the differential inhibitory proper-ties of VHHs were related to differences in bindingaffinities for ED84, the same preparations ofpurified VHHs were used to study their affinity to
Figure 6. Inhibition of binding of ED84 to Site35 bypurified anti-Id VHHs. ED84 IgG at 7 nM (giving a 14 nMbinding site concentration) was incubated with increasingamounts of each VHH. After two hours, the mixture wasadded to Site35-coated wells and bound ED84 wasdetermined by incubation with anti-Fc-HRP conjugate.Inset: purification of VHH79 by Ni-HiTrap column.F, flow-through; W, wash; E, eluted fractions; M,molecular mass standards.
ED84 by biosensor analysis in an IAsys cuvettederivatized with IgG ED84. An example of thecurves obtained for VHH79 is shown in Figure 7.
As shown in Table 3, the affinities of these VHHfor ED84 are similar, all of them with KD values inthe sub-micromolar range. This affinity is w100-fold lower than that of ED84 for Site35,18 which is inagreement with the fact that they were displaced bythe oligonucleotide during panning. These similaraffinities further indicate that the lack of inhibitionby VHH28 of Site35–ED84 interaction is due to
Figure 7. SPR binding curves of VHH79 to ED84.VHH79 at different concentrations was incubated for oneminute on the ED84-derivatized biosensor chip. Theresponse is shown for the 30–800 nM range, used tocalculate the kon.

Table 2 (continued)
(continued on next page)
A, Pre-panning clones. B, Anti-idiotypic clones from specific elution are labeled with an S; non-specific elution clones are labeled with aGC gray background.
DNA Mimicry by Single Domain Antibodies 819
differences in specificity, and not in affinity, with theprivate a-Id84 VHHs.
A llama anti-Id antibody elicits functionalmimicry of DNA
The specific elution protocol followed by a simpleELISA assay for a-Id identification, allowed us toisolate private and public a-Id llama antibodies. Weevaluated the ability of these antibodies to func-tionally mimic the DNA antigen by immunizingmice with the soluble forms of VHH79 and VHH28.We hypothesized that the private a-Id VHH79 could
bear mimicry of DNA. The results from the ELISAassays performed with sera from mice after fourimmunizations, against salmon sperm double-stranded DNA, Site35 double-stranded DNA oligo-nucleotide, and the immunizing VHH, are shown inFigure 8.The group immunized with VHH79 developed
an important reactivity against double-strandedDNA and a smaller but significant reactivity againstdouble-stranded Site35. This is a very interestingresult, showing that this private a-Id VHH shouldbe capable of bearing a mimic of DNA forpresentation to the immune system. The reactivity
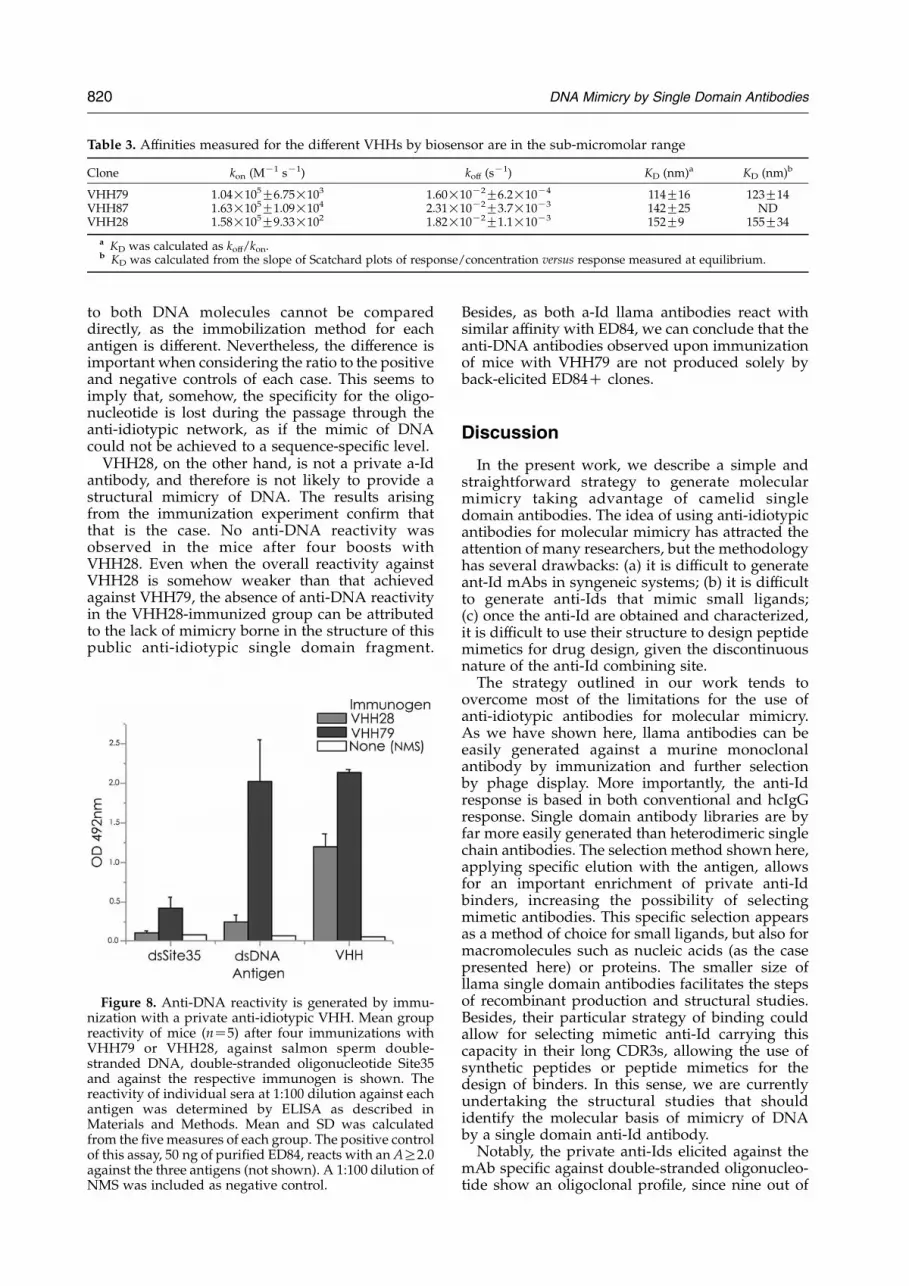
Table 3. Affinities measured for the different VHHs by biosensor are in the sub-micromolar range
Clone kon (MK1 sK1) koff (sK1) KD (nm)a KD (nm)b
VHH79 1.04!105G6.75!103 1.60!10K2G6.2!10K4 114G16 123G14VHH87 1.63!105G1.09!104 2.31!10K2G3.7!10K3 142G25 NDVHH28 1.58!105G9.33!102 1.82!10K2G1.1!10K3 152G9 155G34
a KD was calculated as koff/kon.b KD was calculated from the slope of Scatchard plots of response/concentration versus response measured at equilibrium.
820 DNA Mimicry by Single Domain Antibodies
to both DNA molecules cannot be compareddirectly, as the immobilization method for eachantigen is different. Nevertheless, the difference isimportant when considering the ratio to the positiveand negative controls of each case. This seems toimply that, somehow, the specificity for the oligo-nucleotide is lost during the passage through theanti-idiotypic network, as if the mimic of DNAcould not be achieved to a sequence-specific level.
VHH28, on the other hand, is not a private a-Idantibody, and therefore is not likely to provide astructural mimicry of DNA. The results arisingfrom the immunization experiment confirm thatthat is the case. No anti-DNA reactivity wasobserved in the mice after four boosts withVHH28. Even when the overall reactivity againstVHH28 is somehow weaker than that achievedagainst VHH79, the absence of anti-DNA reactivityin the VHH28-immunized group can be attributedto the lack of mimicry borne in the structure of thispublic anti-idiotypic single domain fragment.
Figure 8. Anti-DNA reactivity is generated by immu-nization with a private anti-idiotypic VHH. Mean groupreactivity of mice (nZ5) after four immunizations withVHH79 or VHH28, against salmon sperm double-stranded DNA, double-stranded oligonucleotide Site35and against the respective immunogen is shown. Thereactivity of individual sera at 1:100 dilution against eachantigen was determined by ELISA as described inMaterials and Methods. Mean and SD was calculatedfrom the fivemeasures of each group. The positive controlof this assay, 50 ng of purified ED84, reacts with an AR2.0against the three antigens (not shown). A 1:100 dilution ofNMS was included as negative control.
Besides, as both a-Id llama antibodies react withsimilar affinity with ED84, we can conclude that theanti-DNA antibodies observed upon immunizationof mice with VHH79 are not produced solely byback-elicited ED84C clones.
Discussion
In the present work, we describe a simple andstraightforward strategy to generate molecularmimicry taking advantage of camelid singledomain antibodies. The idea of using anti-idiotypicantibodies for molecular mimicry has attracted theattention of many researchers, but the methodologyhas several drawbacks: (a) it is difficult to generateant-Id mAbs in syngeneic systems; (b) it is difficultto generate anti-Ids that mimic small ligands;(c) once the anti-Id are obtained and characterized,it is difficult to use their structure to design peptidemimetics for drug design, given the discontinuousnature of the anti-Id combining site.
The strategy outlined in our work tends toovercome most of the limitations for the use ofanti-idiotypic antibodies for molecular mimicry.As we have shown here, llama antibodies can beeasily generated against a murine monoclonalantibody by immunization and further selectionby phage display. More importantly, the anti-Idresponse is based in both conventional and hcIgGresponse. Single domain antibody libraries are byfar more easily generated than heterodimeric singlechain antibodies. The selection method shown here,applying specific elution with the antigen, allowsfor an important enrichment of private anti-Idbinders, increasing the possibility of selectingmimetic antibodies. This specific selection appearsas a method of choice for small ligands, but also formacromolecules such as nucleic acids (as the casepresented here) or proteins. The smaller size ofllama single domain antibodies facilitates the stepsof recombinant production and structural studies.Besides, their particular strategy of binding couldallow for selecting mimetic anti-Id carrying thiscapacity in their long CDR3s, allowing the use ofsynthetic peptides or peptide mimetics for thedesign of binders. In this sense, we are currentlyundertaking the structural studies that shouldidentify the molecular basis of mimicry of DNAby a single domain anti-Id antibody.
Notably, the private anti-Ids elicited against themAb specific against double-stranded oligonucleo-tide show an oligoclonal profile, since nine out of

DNA Mimicry by Single Domain Antibodies 821
the ten clones selected share almost the samesequence. These results suggest that there are notmany different structural solutions to create aspecific binder against a given idiotope, as shownpreviously in other systems.21,22 Thus, the fact thatwe have been able to select a family of anti-Idssingle domain antibodies showing functionalmimicry strongly suggests that the combinationof the generation of single domain binders plusspecific selection would be a straightforwardstrategy for the generation of molecular mimicry.
Materials and Methods
Immunization of llamas
Two llamas (Lama glama) were immunized every 21days with 1 mg of purified mouse IgG ED8417,18 inStimune (Cedi Diagnostics) at a 0.8:1 protein to adjuvantratio, for a total of three immunizations. Prior to eachboost, blood samples were collected and the sera wereused to ascertain the immune response from the lastimmunization. Twenty-one days after the third immu-
Table 4. Variable region sequences of mouse IgGs used to as
The sequence of ED84 is compared to the sequence of IgG1Ed1018,19 (
anti-DNA reactivity (GenBank accession nos AY816717 (VH) and AYsequence similarities to ED84 (GenBank accession nos AY816715 (VHas negative controls, are not shown. They are all IgG1 against protpresented here.
nization, 150 ml of heparinized blood was obtained fromthe better responsive animal and used for mononuclearcell isolation.
Polyclonal anti-Id response
Sera from llamas after each immunization were assayedby standard ELISA procedures. For titer determination,microtiter plates (Maxisorp, NUNC) were sensitized with50 ml of 20 ng/ml of ED84 in PBS for one hour at roomtemperature. After blocking with 200 ml of blocking buffer(3% (w/v) skim milk, 0.05% (v/v) Tween-20 in PBS) forone hour and washing each well three times with PBST,the sera were serially diluted in blocking buffer, added tothe plate in duplicate wells (50 ml/well) and incubated forone hour at room temperature. Following washing, theassay was revealed with polyclonal rabbit anti-llama IgGs(1:800 dilution in blocking buffer, 50 ml, one hour at roomtemperature), washed, incubated for one hour at roomtemperature with goat anti-rabbit IgGs conjugated toHRP (Sigma) at 1:500 dilution, and washed again.Substrate (OPD, Sigma) was added, reaction was stoppedwith 2M H2SO4 and A492 nm was measured on an ELISAreader (S960 Metertech Inc.). The titer was considered asthe dilution at which the reactivity is 50% of the 1:100dilution reactivity.
ses the specificity of anti-idiotypic VHHs
with identical anti-DNA reactivity as ED84), to IgG1 390, with no816716 (VL)] and to IgG2a GF6, an anti-DNA MAb with no CDR) and AY816714 (VL)]. The sequences of ED5, ED19 and F10, usedein antigens, with no variable region similarities with the IgGs

822 DNA Mimicry by Single Domain Antibodies
For analysis of the reactivity against different mouseIgGs, similar procedures were followed, in which sera at1:100 dilution, with or without absorption to normalmouse IgG-agarose (Sigma), was added to plates sensi-tized with 0.5 mg/well of the different Fab fragments.(Relevant mouse IgG variable region sequences are listedin Table 4.) Absorption was performed by incubating withgentle agitation for three hours at room temperature, a 5:1mixture of the diluted sera and IgG-agarose. Followingcentrifugation, supernatant was added directly to theplate wells.For conventional and hcIgG fractionation, serum was
passed through HiTrap Protein A and HiTrap Protein Gcolumns (Pharmacia) following described procedures.23
Purity of the different fractions was analyzed by SDS-PAGE. Reactivity against Fab ED84 and control mouseIgG (ED19) of both fractions at 100 ng of llama IgG/well,with or without absorption to normal mouse IgGs, wasdetermined by ELISA as described above.
Construction of the library and phage displayselection of anti-Id llama antibodies
Mononuclear cells were isolated from llama #8338heparinized blood by Ficoll-Hypaque (Pharmacia) gra-dient centrifugation.Total RNA from 107 cells was purified by TRIZOL
reagent (Pharmacia) and subjected to first strand syn-thesis. VHH specific primers:23
Lam07f-NotI(5 0gatggtgatgatgatgtgcggccgcgctggggtcttcgctgtggtgcg3 0)
VH1b-SfiI(5 0gctggattgttattactcgcggcccagccggccatggcccaggtsmarctgcagsagtcwgg3 0) VH6b-SfiI(5 0cgtggattgttattatctgcggcccagccggccatggccgatgtgcagctgcaggcgtctggrggagg3 0)were used in PCR to amplify VHH cDNAs. PCR products(w500 bp) were purified from agarose gels (GFX PCRDNA & Gel band purification kit, Pharmacia), digestedwith SfiI and NotI sequentially and repurified.For phage-display library construction, 1 mg of SfiI-
NotI-digested plasmid pHEN24 and the digested frag-ments were ligated in a 1:3 ratio. The ligation reaction waspurified with phenol/chloroform, precipitated and resus-pended in sterile MilliQ water.Electrocompetent XL1-Blue cells were transformed and
library size was calculated by plating aliquots on LB Ampagar, yielding a total of 1.1!108 clones. Ten clones wereused for plasmid DNA preparation (Quiaprep SpinMiniprep Kit, Quiagen) and sequenced for diversityanalysis.Transformants were grown in SB mediumC1.5% (w/v)
GlcCAmpCTet for two hours at 37 8C, and infected with20-fold excess of VCS helper phage (Promega) for onehour at 37 8C. Infected cells were harvested by centrifu-gation, resuspended in 700 ml of SBCAmpCTetCKanand incubated overnight at 30 8C with agitation. Phageparticles were precipited form culture supernatant with4% PEG 8000, 0.5 MNaCl, resuspended in sterile PBS andused for panning.Panning was performed on 24-well culture plates. Four
wells were sensitized with 2 mg of IgG ED84 and twowells were left uncoated. After blocking with 2% skimmilk in PBS, 2.8!1013 phage particles in 1% skimmilk/PBS were added to each well, incubated for
two hours at room temperature, and washed tentimes with PBST. Bound phages were eluted eitherwith 0.1 M glycine (pH 2.2) (non-specific elution) orwith a solution of 2.5 ng/ml of dsSite35 in PBS(specific elution) in duplicate wells. The two non-coatedwells were subjected to the same procedures and wereused as the negative control of panning for each elutionmethod.Eluted phages were titrated and subjected to a second
round of panning, following the same procedure.Phage titers of input and output at all steps wereestimated by infection of XL1-Blue cells and plating onLB Amp plates.For estimation of anti-Id VHH presence, of XL1-Blue
cells were infected with the phages eluted in eachmethod. Ninety-six individual colonies from eachmethod were grown in culture plates in 2!TYCAmpCGlc, three hours at 37 8Cwith agitation, and superinfectedwith 2!109 pfu of VCS helper phage for 30 minutes. Cellswere pelleted, resupended in 2!TYCAmpCKan andincubated for 16 hours at 30 8C. Culture supernatantscontaining the phages were diluted twofold in 2% skimmilk in PBS and added to ELISA plates sensitized with1 mg of ED84 or control IgG1 (ED5) and blocked with2% skim milk in PBS. After washing with PBST,bound phages were revealed with 1/2000 anti-fd rabbitIgG conjugated to HRP (Sigma). Readings from bothED84 and control wells were done and those clones witha DAO0.1 were considered as presumptive a-Id clones.Statistical comparison by c2 test of the proportion ofanti-Id clones eluted with each method and one-factorANOVA of DA values from both populationswere performed with Origin 6.0 professional (MicrocalSoftware, Inc.).From the strongest anti-Id clones of both methods,
showing only background recognition of ED5, ten cloneswere selected, their plasmidic DNA isolated andsequenced.The selected phage clones from the specific elution
method were expressed in higher scale and re-evaluatedin their specificity for ED84, using 106 particles/well induplicate wells, in a similar ELISA. Reactivity wasmeasured against ED84, against IgG1 3-90, highlyhomologous to ED84 but with no anti-DNA activity,and against GF6, an anti-DNA IgG2a with no sequencesimilarity to ED84 (Table 4). This time IgG1 ED19 wasused as control.
Inhibition assays
The selected clones were expressed as soluble peri-plasmic proteins in E. coli non-suppressor strain Top10(Invitrogen) for ED84 inhibition studies. Two of the VHHclones with specificity for ED84 (VHH79 and VHH87)and the clone with clearly different sequence andrecognition pattern, VHH28, were expressed. Periplasmicspace proteins were isolated following established tech-niques25,26 and purified by Hi-Trap Ni column (Pharma-cia). Their purity was assessed by SDS-15% PAGEelectrophoresis and their concentration by A280 nm read-ing (average yield, 2 mg/l of pure VHH). Their theoreticalconcentration factor was obtained from the amino acidsequence with ProtParam at ExPASy.27
ELISA plates were sensitized for one hour with 0.5 mgof streptavidine (Sigma), blocked and incubated with25 ng of biotinilated Site35 for 30 minutes. ED84 (7 nM)was preincubated with the VHHs at different concen-tration ratios for two hours and added to the plate for one

DNA Mimicry by Single Domain Antibodies 823
hour. Bound ED84 was determined with aFC-HRP asdescribed above.
Affinity measurements
The affinities of purified VHHs for ED84 weredetermined using an IAsys biosensor. A CMD cuvettewas sensitized with 100 ng/ml of IgG ED84 following themanufacturer’s instructions. VHHs were incubated atdifferent concentrations on the cuvette, either for oneminute (kon determination) or until equilibrium (KD
determination). The dissociation was measured for thehigher concentrations of VHH, after washing with PBS.All parameters were calculated using GraFit 5.0.3(Erithacus Software Ltd.) and Origin 6.0 professional(Microcal Software, Inc).
Immunization of mice with VHH fragments and anti-DNA response
Groups of five female BALB/c mice of six to eightweeks of age were immunized every 15 days with 50 mg ofprotein per boost with either VHH79 or VHH28 inStimune. After four immunizations, blood was takenfrom the retro-orbital sinus and the individual sera wereassayed at ELISA at 1:100 dilution in 1% (w/v) BSA. Theplate was sensitized in duplicate wells with: 50 ng ofbiotinilated Site35 (as described above), 0.5 mg of salmonsperm DNA (dried overnight at 37 8C) or 0.5 mg of VHH(one hour at room temperature; mice immunized witheach VHHwere assayed with the corresponding protein).As a negative control, normal mouse serum (NMS) wasused and ED84 was used as a positive control at 50 ng ofIgG per well. The ELISA was performed essentially asdescribed above for ED84.
Data Bank accession codes
Sequences of the ten clones have been deposited withGenBank, with accession numbers from AY816718 toAY816744.
Acknowledgements
We are grateful to Vanina Alzogaray for help withthe immunizations of llamas, to Gonzalo PerezZabala and Guillermo Vila Melo for providing thellamas, farm site and veterinarian care, and toMariaLaura Cerutti for her helpful comments about thiswork.
References
1. Oudin, J. & Michel, M. (1963). A new allotype form ofrabbit serum gammaglobulins, apparently associatedwith antibody function and specificity. C. R. Hebd.Seances Acad. Sci. 257, 805–808. French.
2. Jerne, N. K. (1974). Towards a network theory of theimmune system. Ann. Immunol. (Paris), 25C, 373–389.
3. Nisonoff, A. (1991). Idiotypes: concepts and appli-cations. J. Immunol. 147, 2429–2438.
4. Gaulton, G. N. & Greene, M. I. (1986). Idiotypicmimicry of biological receptors. Annu. Rev. Immunol.4, 253–280.
5. Greenspan, N. S. (1992). Antigen mimicry withantiidiotypic antibodies. In Structure of Antigens (VanRegenmortel, M., ed.), pp. 55–79, CRC Press, BocaRaton, FL.
6. Nisonoff, A. & Lamoyi, E. (1981). Implications ofthe presence of an internal image of the antigenin anti-idiotypic antibodies: possible application tovaccine production. Clin. Immunol. Immunopathol. 21,397–406.
7. Evans, S. V., Rose, D. R., To, R., Young, N. M. &Bundle, D. R. (1994). Exploring the mimicry ofpolysaccharide antigens by anti-idiotypic antibodies.The crystallization, molecular replacement, andrefinement to 2.8 A resolution of an idiotope-anti-idiotope Fab complex and of the unliganded anti-idiotope Fab. J. Mol. Biol. 241, 691–705.
8. Bentley, G. A., Boulot, G., Riottot, M. M. & Poljak, R. J.(1990). Three-dimensional structure of an idiotope–anti-idiotope complex. Nature, 348, 254–257.
9. Garcıa, K. C., Ronco, P. M., Verroust, P. J., Brunger,A. T. & Amzel, L. M. (1992). Three-dimensionalstructure of an angiotensin II-Fab complex at 3 A:hormone recognition by an anti-idiotypic antibody.Science, 257, 502–507.
10. Garcıa, K. C., Desiderio, S. V., Ronco, P. M., Verroust,P. J. & Amzel, L. M. (1992). Recognition of angio-tensin II: antibodies at different levels of an idio-typic network are superimposable. Science, 257,528–531.
11. Ban, N., Escobar, C., Garcıa, R., Hasel, K., Day, J.,Greenwood, A. & McPherson, A. (1994). Crystalstructure of an idiotype–anti-idiotype Fab complex.Proc. Natl Acad. Sci. USA, 91, 1604–1608.
12. Fields, B. A., Goldbaum, F. A., Ysern, X., Poljak,R. J. & Mariuzza, R. A. (1995). Molecular basis ofantigen mimicry by an anti-idiotope. Nature, 374,739–742.
13. Ban, N., Day, J., Wang, X., Ferrone, S. &McPherson, A.(1996). Crystal structure of an anti-anti-idiotypeshows it to be self-complementary. J. Mol. Biol. 255,617–627.
14. Braden, B. C., Fields, B. A., Ysern, X., Dall’Acqua, W.,Goldbaum, F. A., Poljak, R. J. & Mariuzza, R. A.(1996). Crystal structure of an Fv-Fv idiotope–anti-idiotope complex at 1.9 A resolution. J. Mol. Biol. 264,137–151.
15. Ban, N., Escobar, C., Hasel, K. W., Day, J., Greenwood,A. & McPherson, A. (1995). Structure of an anti-idiotypic Fab against feline peritonitis virus-neutra-lizing antibody and a comparison with the complexedfab. FASEB J. 9, 107–114.
16. Muyldermans, S. & Lauwereys, M. (1999). Uniquesingle-domain antigen binding fragments derivedfrom naturally occurring camel heavy-chain anti-bodies. J. Mol. Recogn. 12, 1–10.
17. Cerutti, M. L., Centeno, J. M., Goldbaum, F. A. & dePrat-Gay, G. (2001). Generation of sequence-specific,high affinity anti-DNA antibodies. J. Biol. Chem. 276,12769–12773.
18. Di Pietro, S. M., Centeno, J. M., Cerutti, M. L., Lodeiro,M. F., Ferreiro, D. U., Alonso, L. G. et al. (2003).Specific antibody-DNA interaction: a novel strategyfor tight DNA recognition. Biochemistry, 42,6218–6227.
19. Goletz, S., Christensen, P. A., Kristensen, P., Blohm,D., Tomlinson, I., Winter, G. & Karsten, U. (2002).

824 DNA Mimicry by Single Domain Antibodies
Selection of large diversities of antiidiotypic antibodyfragments by phage display. J. Mol. Biol. 315,1087–1097.
20. George, A. J., McBride, M. H., Glennie, M. J., Smith,L. J. & Stevenson, F. K. (1991). Monoclonal antibodiesraised against the idiotype of the murine B-celllymphoma Bcl1 act primarily with heavy chaindeterminants. Hybridoma, 10, 219–227.
21. Goldbaum, F. A., Braden, B. C. & Poljak, R. J. (1998).Antigen mimicry by anti-idiotopic antibodies. Immu-nologist, 6, 13–18.
22. Goldbaum, F. A., Velikovsky, C. A., Dall’Acqua, W.,Fossati, C. A., Fields, B. A., Braden, B. C. et al. (1997).Characterization of anti-anti-idiotypic antibodies thatbind antigen and an anti-idiotype. Proc. Natl Acad. Sci.USA, 94, 8697–8701.
23. van der Linden, R., de Geus, B., Stok, W., Bos, W., vanWassenaar, D., Verrips, T. & Frenken, L. (2000).Induction of immune responses and molecularcloning of the heavy chain antibody repertoire ofLama glama. J. Immunol. Methods, 240, 185–195.
24. Griffiths, A. D., Williams, S. C., Hartley, O.,Tomlinson, I. M., Waterhouse, P., Crosby, W. L. et al.(1994). Isolation of high affinity human antibodiesdirectly from large synthetic repertoires. EMBO J. 13,3245–3260.
25. Wingfield, P. T. (1995). Overview of the purificationof recombinant proteins produced in Escherichia coli.In Current Protocols in Protein Science (Coligan, J. E.,Dunn, B. M., Ploegh, H. L., Speicher, D. W. &Wingfield, P. T., eds), vol. 1, pp. 6.1.1–6.1.22, Wiley,New York.
26. Wingfield, P. T. (1995). Preparation of soluble proteinsfrom Escherichia coli. In Current Protocols in ProteinScience (Coligan, J. E., Dunn, B. M., Ploegh, H. L.,Speicher, D. W. & Wingfield, P. T., eds), vol. 1, pp.6.2.1–6.2.15, Wiley, New York.
27. Gasteiger, E., Gattiker, A., Hoogland, C., Ivanyi, I.,Appel, R. D. & Bairoch, A. (2003). Expasy: theproteomics server for in-depth protein knowledgeand analysis. Nucl. Acids Res. 31, 3784–3788.
Edited by I. Wilson
(Received 30 November 2004; received in revised form 19 February 2005; accepted 25 March 2005)





























