Embed Size (px)
Citation preview

Ethology 105, 57-72 (1999) 8 1999 Blackwell Wissenschafts-Verlag. Berlin ISSN 0179-1613
Department of Ecology and Genetics, Institute of Biological Sciences, University of’ Aarhus, Aarhus
Local Mating, Dispersal and Sex Ratio in a Gregarious Parasitoid Wasp
Ian C. W. Hardy, Jesper B. Pedersen, Mikael K. Sejr & Ulla H. Linderoth
Hardy, 1. C. W., Pedersen, J. B., Sejr, M. K. & Linderoth, U H. 1999: Local mating, dispersal and sex ratio in a gregarious parasitoid wasp. Ethology 105, 57-72.
Abstract Parasitoid sex ratios can be greatly influenced by mating and dispersal behaviour. Many sex ratio
models assume that mating is strictly local (only mated females disperse from the natal patch) and that a single male is sufficient to inseminate all females in a brood. Bethylids (acukdte parasitoids) have been used to test predictions of these models, but less attention has been paid to testing their underlying assumptions. We investigated the timing of eclosion, mating and dispersal in mixed-sex and single-sex broods of the bethylid wasp Goniozus nephanridis. In mixed-sex broods, almost all females mate before dispersal and a single male is sufficient to inseminate virtually all females, even when brood sizes are large. Males disperse from both mixed-sex and all-male broods, but males in all-male broods disperse more slowly. Virgin females disperse from all-female broods, which are common. Virgin females can produce a brood, mate with their own sons and subsequently produce mixed-sex broods, but their Success rate is very low. Virgin females could potentially circumvent sex allocation constraints by superparasitizing mixed-sex broods, but when presented with hosts bearing mixed-sex broods they destroy all members of the initial brood before ovipositing. Because of the high prevalence of single- sex broods and dispersal of both sexes, the mating structure of C. nephantidk is unlikely to conform to the assumption of strict local mating.
Corresponding author: Ian C. W. Hardy, Department of Ecology and Genetics, Institute of Biological Sciences, University of Aarhus, DK-8000 Aarhus C, Denmark. E-mail: lan.Hardy(0 btology .aau.dk
Introduction
Bethylids (Hymenoptera: Bethylidae) are aculeate parasitoids of lepidopteran or cojeopteran larvae. Their life histories have stimulated the development and testing of a number of models which predict optimal sex allocation in response to local mate competition, clutch size and developmental mortality (Hamilton 1967; Green eta]. 1982; Griffiths & Godfray 1988; Heimpel 1994; Morgan & Cook 1994; Nagelkerke & Hardy 1994; Hardy & Cook 1995; Hardy & Mayhew 1998; Hardy etal. 1998). All of the models cited above, and most tests, have a number of restrictive assumptions. Offspring are assumed to develop and mate within a discrete and ephemeral group. One male is assumed sufficient to inseminate all
U. S. Copyright Clearance Center Code Statement: 0179-161 3/99/1051-0057$14,00/0

58 1. C. W. Hardy, J. B. Pedersen, M. K. Sejr & U. H. Linderoth
females. Females, but not males, then disperse. Individuals developing in single- sex broods have no mating opportunities and are assumed to have zero fitness.
The mating structure assumption is important because its relaxation (i.e. assuming partial local mating or panmixis) is predicted to influence sex ratios considerably: as nonlocal mating increases, sex ratio optima become less female biased (Fisher 1930; Nunney & Luck 1988; Hardy 1994a). Furthermore, this influence is predicted to be especially apparent when offspring groups are produced by a single foundress female (Nunney & Luck 1988). Single-foundress broods are the norm in bethylids because adult females commonly attempt to exclude com- peting conspecifics from the vicinity of their hosts and females finding previously parasitized hosts may destroy the initial brood before ovipositing or reject the host completely (Griffiths & Godfray 1988; Hardy & Blackburn 1991; Petersen & Hardy 1996; Mayhew 1997).
While local mating in bethylids is almost certainly predominant (there are numerous reports of protandry, sibling mating, a lack of inbreeding depression and short-lived and apterous males (literature cited in Griffiths & Godfray 1988; Hardy & Mayhew 1998), there is also evidence to suggest that some nonlocal mating occurs. For example, in all species of Goniozus described (the bethylid genus most used in tests of sex ratio models), males are winged, suggesting potential dispersal from the natal site. Field observations on Cephalonomia stepliunoderis Betrem report high proportions of male dispersal and also the dispersal of virgin females (Koch 1973; Barrerd et al. 1993). While in some species virgin females may be reluctant to mate post dispersal, in others they may not (van Emden 1931; Gordh et al. 1983; Cook 1993). Furthermore, comparison across 19 bethylid species has shown a positive relationship between relative male size (a likely correlate of male dispersal ability) and sex ratio (proportion of offspring which are males), again suggestive of some nonlocal mating (Hardy & Mayhew 1998).
One factor likely to promote dispersal and nonlocal mating is a lack of sib- mating opportunities. Sib-mating rates may be restricted by limited male sperm storage capacity and rates of spermatogenesis. Sib-mating is also not possible in single sex broods. Broods containing only females at eclosion (termed ‘virgin broods’) occur commonly in bethylids (7-50% of broods), chiefly due to devel- opmental mortality of males but also due to single-egg clutches and non-zero sex ratio variance at oviposition (Hardy & Cook 1995; Mayhew & Godfray 1997; Hardy et al. 1998). Because of haplo-diploid sex determination, virgin females can produce male offspring but these males will have zero fitness unless they disperse to mate nonlocally, mate with their mothers (in many species mothers remain with their broods for prolonged periods after completing oviposition and thus virgin mothers potentially have the opportunity to mate with their sons; van Emden 193 1 ) or are added to broods containing females produced by mated conspecifics (superparasi tism).
In this paper we test assumptions of sex ratio models by investigating the mating structure of the bethylid wasp Goniozus nrphantiih (Muesebeck). G . nephanridis occurs in the Indian subcontinent and is a gregarious ectoparasitoid of Opisina arrnosella (Lepidoptera: Oecophoridae) larvae which feed within galleries

Mating. Dispersal and Sex Ratio in a Parasitoid WdSp 59
of silk and frass on the undersurface of coconut leaves. Host density is generally low, with occasional high density during outbreaks (Cock & Perera 1987). In Common with many bethylids, G. nrphanridis has a female biased and precise sex ratio and previous authors have observed protandry, sibling mating, polygyny and a lack of inbreeding depression (literature cited in Hardy &.Cook 1995). Both sexes possess functional wings but males have been reported to be ‘sluggish’, shorter- lived and smaller, suggesting poor dispersal ability relative to females. Despite this apparent fit to both the assumptions and predictions of theory, a laboratory study estimated that 32% of broods produced by mated mothers contain only females at adult eclosion (Hardy & Cook 1995). Some virgin broods are expected under strict local mating as a consequence of male developmental mortality (estimated at 28% by Hardy & Cook 1995) but 32% virginity is substantially higher than the predicted proportion, assuming perfect control of sex allocation (Heimpel 1994; Hardy & Cook 1995).
Our experiments addressed the following questions 1 . In mixed sex broods, do males inseminate all of their sisters and do males and any unmated females disperse from the natal site? 2. Do individuals disperse from all-female and all- male broods? 3. Do virgin mothers mate with their sons? 4. Do virgin females superparasitize broods containing developing females without entirely destroying the initial brood? In addition, we used our data to explore the possibility that previously reported percentages of developmental mortality and virginity (Hardy & Cook 1995) were aberrant.
Materials and Methods
The G. nepphanfidis used in this study were originally obtained from T. M. Manjunath, Biocontrol Research Laboratories, Bangalore, India. To our knowl- edge, this strain is separate from the Sri Lankan strain used by Hardy & Cook (1995). As in previous work (Hardy & Blackburn 1991; Hardy et al. 1992; Hardy & Cook 1995; Petersen & Hardy 1996), we reared G . nephantidis on the facultative host Corc-yru cephalonicu (Stainton) (Lepidopterd: Pyrdlidae), on which it had been maintained for more than 3 yr prior to experiments. G. nrphuntidis broods were obtained for experiments by placing adult females individually in 2.5 x 7.5cm glass vials stoppered with nylon gauze and cotton wool and containing one C . cepf1aIonic.u larva. Mated mothers produce broods containing female offspring while unmated mothers (e.g. those from all-female broods) produce broods of sons only. All cultures and experiments were maintained in a laboratory at 25 ‘C, ~ 7 0 % relative humidity (r.h.) and constant, though indirect and dim, illumination.
Eclosion and Dispersal
Pardlysed host larvae bearing clutches of eggs were placed in the central of three chambers in a plastic block decked with Plexiglas (Fig. I ) . The chambers were connected to each other and to the outside by a slot through which adult G. nep/?antic/i.s could pass with ease. Thus, as in nature, broods developed in an

60 I . C . W. Hardy, J. B. Pedersen, M. K. Sejr & U. H. Linderoth
F(q. I : Apparatus for local mating and dispersal experiments. An opaque plastic block ( I 5 x 4 x 1 cm) containing three chambers (diameter = 1.8 cm; depth = 0.6cm) and a slot (width 0.1 cm) was decked with Plexiglas and placed within a sealed transparent plastic box ( I7 x 1 1 x 4cm). A one-chambered block (7.5 x 4 x I cm) decked with Plexiglas was also
present to provide structure for dispersed adults
enclosed space which could be exited via a tunnel. The three-chambered block was placed inside a sealed transparent plastic box. The box also contained a one- chambered block, with a slot and decked with Plexiglas (Fig. 1). We included this because pilot observations suggested that G. nepphantidis spend much time in the slot of the three-chambered block. The slot in the one-chambered block provided a structure for dispersed adults to occupy similar to, but separate from, the three- chambered box: this reduced the probability that dispersed adults would return to the three-chambered block where they could not be distinguished from non- dispersed adults. However, during the experiments, individuals were seldom observed in the one-chambered block.
Once the brood members reached the late pupal stage we inspected the brood every 3h from 0600 h until 2400 h each day. Wasps did not appear to be disturbed from their normal behaviour (mostly fairly inactive) by these observations. We recorded the time that adults eclosed and their sex. Any adults that had dispersed were removed from the apparatus and their sex and the time of dispersal recorded. We defined wasps as having dispersed from the natal site if they were observed outside the three-chambered block. There were 42 replicates in total: 19 mixed-sex broods, nine all-female broods and 14 all-male broods.
Local Mating and Insemination Capacity
During the above inspections, we also noted any mating behaviour in mixed sex broods. Each female dispersing from a mixed-sex brood was presented with a host in a vial and her progeny reared and sexed to determine whether the dispersed female had mated. This allowed us to determine how many females the males in the brood had inseminated.

Mating, Dispersal and Sex Ratio in a Parasitoid Wasp
MotherSon Mating
Individual virgin females (n = 25) were obtained from all-female broods or by isolating wasp larvae prior to pupation. Virgin females were presented with a single host in a vial, allowed to oviposit and to remain with their brood. We checked daily whether the mother was still alive until male. offspring eclosed. 24 h after the first eclosed male was observed, any surviving mothers were removed and presented with a fresh host. The progeny were sexed at maturity to determine whether the mother had mated successfully with sons from the first brood.
61
Ovicide
Mated females (n = 15) were each allowed to paralyse and oviposit on a host in a vial. The mothers were then removed and the eggs were counted and marked with an aqueous solution of eosin stain. Marking does not influence egg hatching success or parasitoid behaviour (Hardy & Blackburn 1991). A virgin female, with no prior oviposition experience, was then introduced into the vial. On each following day, until all eggs had been destroyed or had hatched, we counted the numbers of marked (original) and unmarked (laid by virgin female) eggs on the host.
Sex Ratio, Mortality and Virginity
We estimated the probability of eggs surviving to adulthood and the adult sex ratio by counting the numbers of eggs on hosts (from the above experiments plus additional replicate broods) and the number and sexes of emerging adults. We obtained mortality estimates for 126 clutches and secondary sex ratio data from 287 broods. We analysed these data and calculated optimal numbers of sons and expected proportions of virginity as described in detail in Hardy & Cook (1995).
Results
Eclosion and Dispersal
Mi.ued s e . ~ broods
A total of 25 males and 170 females eclosed from the 19 mixed-sex replicates, There were either one or two males in each brood. Fig. 2 shows the cumulative Percentages of males and females eclosing as time progressed. Females within a brood eclosed over a 57-h period and, in broods with two males (n = 6), the second male eclosed up to 51 h after the first. Timing of eclosion was explored using cohort survival analysis in GLlM (Crawley 1993). This allows investigation of whether and how the probability of eclosion changes with time and comparison of male and female eclosion. 'Time until eclosion' of individuals was analysed by defining time zero for each brood as the last observation before any eclosion from a brood (Dijkstra 1986). Eclosion was best explained by an exponential model (fitting a Weibull model did not lead to a reduction in deviance), suggesting that the overall probability ofeclosion was constant with time. However, males eclosed significantly
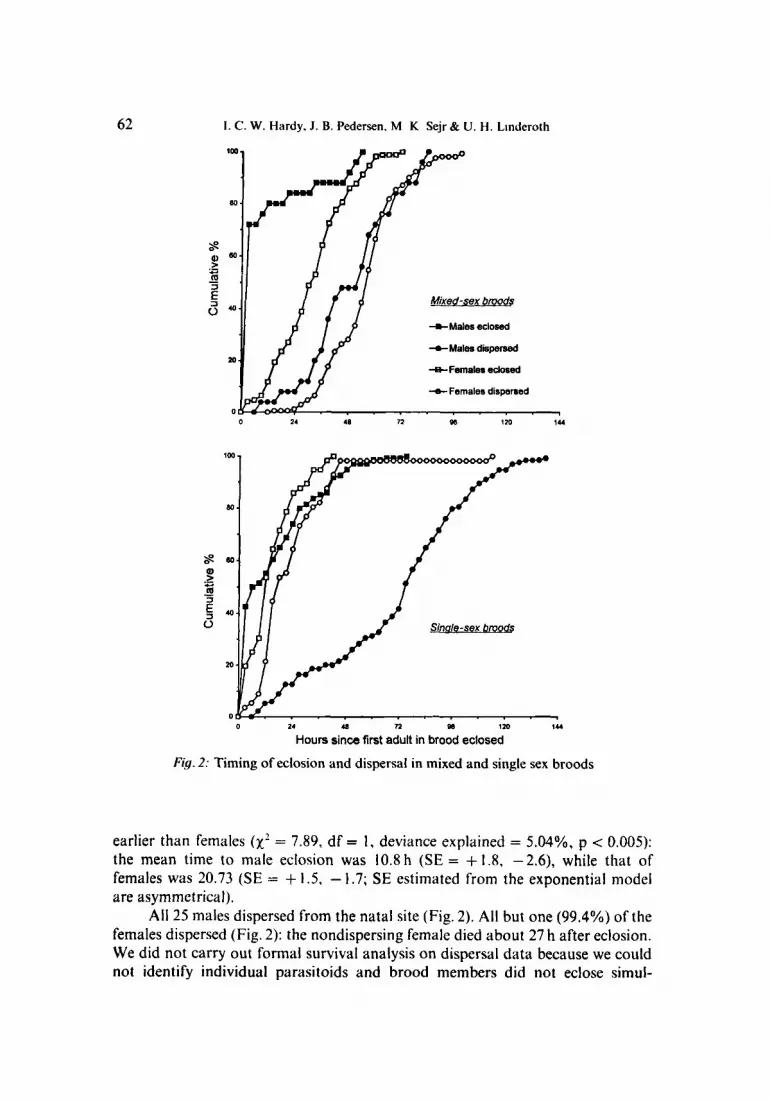
62 1. C. W. Hardy, J . B. Pedersen. M K Sejr & U. H. Linderoth
Mixed-sex bmd$
+Males eclosed
+Males dispersed
4 F m s l e s e c l d
-e- Females dispersed
0 24 40 72 m 120 I44
0 24 4a 72 Do im 144
Hours since first adult in brood eclosed
Fig. 2: Timing of eclosion and dispersal in mixed and single sex broods
earlier than females (x ’ = 7.89, df = 1, deviance explained = 5.04%. p -= 0.005): the mean time to male eclosion was 10.8 h (SE = + 1.8, -2.6), while that of females was 20.73 (SE = + 1.5, - 1.7; SE estimated from the exponential model are asymmetrical).
All 25 males dispersed from the natal site (Fig. 2). All but one (99.4%) of the females dispersed (Fig. 2): the nondispersing female died about 27 h after eclosion. We did not carry out formal survival analysis on dispersal data because we could not identify individual parasitoids and brood members did not eclose simul-

Mating, Dispersal and Sex Ratio in a Pdrasitoid Wasp 63
taneously (hence time zero for the cohort survival curve is indeterminate). Males dispersed about 48 h after eclosion. In 9/19 broods, the males had dispersed before female eclosion was complete. In broods with two males, the second male to disperse always dispersed within 6 h of the first. Females dispersed about 24 h after eclosion. There were up to 48 h between the dispersal of the first and last female within a brood, although 50% dispersed within 6 h of the first.
A I1 fivnalr brooch
A total of 56 females eclosed from the nine all-female replicates. Eclosion was best explained by an exponential model (a Weibull model did not reduce the deviance) and the mean time to eclosion was 15.2 h (SE = + 1.9, -2.6). The time until eclosion of females in all-female broods was not significantly different from that offemales in mixed-sex broods (x’ = 3.82, df = I , deviance explained = 2.6%, p > 0.05; in this analysis time zero was defined as the last observation before any female eclosed).
All 56 females dispersed from the natal site, usually about 12 h after eclosion (Fig. 2). There were up to 96 h between the dispersal of the first and last female within a brood, although this long period was due to an individual which dispersed 48 h after the penultimate female; 50% of females in a given brood dispersed within 3 h of the first.
AN male broods
A total of 134 males eclosed from the 14 all-male replicates. The probability of eclosion declined weakly over time (fitting a Weibull distribution reduced the deviance significantly compared with an exponential model: x’ = 6.67, df = I , p < 0.01, deviance explained = 4.3%). The mean time to eclosion was 16.2h (SE = + 1.35, - 1.5; SE estimated from the Weibull model are asymmetrical). Males were observed to examine the pupae of unemerged males and mount other males in an attempt to mate (such ‘copulation’ lasted 34min) . The time until eclosion of males in all-male broods was not significantly different from that of males in mixed-sex broods (xz = 2.33, df = I , deviance explained = 1.3%, p > 0.05).
All but one (99.25%) male dispersed from the natal site (Fig.2): the male that did not disperse had deformed wings and was unable to walk normally. In contrast to males in mixed-sexed broods, males in all-male broods dispersed about 72 h after eclosion with up to 114 h between the dispersal of the first and last male within a brood (50% of males dispersed within 45 h of the first). The slow dispersal of males in all-male broods is the most obvious difference in behaviour between single-sex and mixed sex-broods (Fig. 2).
Local Mating and Insemination Capacity
We observed males entering cocoons and mating with memerged females within. Mating was also observed after female eclosion (copulation duration was about 34min) . Seven of the 169 females which dispersed from mixed sex broods

64 I . C. W. Hardy, J . B. Pedersen. M. K. Sejr & U. H. Linderoth
died before ovipositing on the presented host, three escaped from their vials and 17 produced broods in which no offspring survived to maturity: we could not assess the mating status of these females. Of the remaining 142 females, 140 (98.6%) produced female offspring and two did not. These two females were from separate replicates and produced broods of six and eight sons, indicating that the lack of daughter production resulted from virginity rather than the developmental mor- tality of daughters in an initially mixed-sex brood (in mixed-sex broods the number of males rarely exceeds two; Hardy & Cook 1995).
The two uninseminated females were from large broods containing one male only. We explored the influence of the number of females in a brood on the probability of all females being inseminated, using data from broods containing one male at eclosion (n = 13). Insemination was fitted as a binary response variable ( 1 = all females mated, 0 = some females unmated), with females of unknown mating status assumed to be mated. The number of females at eclosion did not significantly influence the probability of complete insemination (logistic [binary] regression using GLIM: x’ = 3.68, df = I , deviance explained = 32.9%, 0.1 > p > 0.05). Since this regression fell only just short of significance (1’ critical value = 3.84, p = 0.05) and its form (Fig. 3) is suggestive of some male limitation, we carried out a logistic (proportional) regression (using GLIM) on data from all 19 replicates, which further showed that the proportion of females inseminated within a brood was unrelated to the number of males (x’ = 1.53, df = 1, deviance explained = 19.3%, p > 0.1) or the number of females ( x 2 = 2.98, df = 1, deviance explained = 39.6%, p > 0.05) in the brood. We conclude that male insemination capacity is sufficient for the optimal number of males a t eclosion to equal 1 in broods within the normal size range.
Mother-Son Mating
Of the 25 virgin mothers, five were still alive 24 h after their first son eclosed. We observed males mating with living mothers and also attempting to mate with
3 0.2 - 1 N o O + . . . . . . . . . r . = . - = ‘
0 2 4 6 8 10 12 14 16 Number of females
Fig.3: Probability that all females are inseminated by one male in relation to number of females eclosing. The data are binary ( 1 = all females inseminated, 0 = some females uninseminated) and the fitted curve is the estimated probability, p, ofcomplete insemination (p = I / [ I + (l/(exp (-0.4298~ + 6.334))}], where x is the number of females). The
regression falls just short of significance (see text)

Mating, Dispersal and Sex Ratio in a Parasitoid Wasp 65
each other and the bodies of their dead mothers. Of the five females presented with a second host only one produced female offspring. We conclude that although virgin females can circumvent sex allocation constraints by mating with their sons, the probability of success is low.
Ovicide
In 14 out of 15 replicates, the virgin female destroyed all marked eggs before laying her own clutch. In the remaining replicate, the virgin female neither com- mitted ovicide nor oviposited. We conclude that virgin females do not refrain from full ovicide as a means of providing their sons with local mating opportunities.
Sex Ratio, Mortality and Virginity
The mean secondary sex ratio of 259 broods produced by mated mothers was highly female biased (proportion males = 0.0896, SE = 0.006). Sex ratio variance was less than binomial (heterogeneity factor calculated in GLIM was 0.77; Meelis test, R = 0.585, u = -4.72, p < 0.001; details of the Meelis test are given in Nagelkerke & Sabelis 1991; Hardy & Cook 1995), but not all broods had similar compositions: 41.7% were all-female, 47.5% contained one male and 10.8% con- tained two males. No brood contained more than two males. Some (1 2/32) broods without developmental mortality contained no males. There was no trend in sec- ondary sex ratio as brood size (range 1-17) increased (logistic regression in GLIM: x 2 = 0.66, df = I , deviance explained = 0.3%, p > 0.1). This was because the number of males per brood also increased (log-linear regression in GLIM: x2 = 25.44, df = I , deviance explained = 12.1 YO, p < 0.OOl) .
The mean probability of egg to adult survival was 0.752 (SE = 0.026) (data from 126 broods produced by mated and unmated mothers). The eggs within a given brood tended to survive or die collectively (mortality had greater than binomial variance: heterogeneity factor = 5.2; Meelis test, R = 5.22, U = 27.78, P < 0.001). Mortality was unrelated to clutch size (range 3-18 eggs) (logistic regression, rescaled for overdispersion: F1.124 = 1.8, deviance explained = 0.9%, p > 0.05) and did not differ between broods produced by unmated mothers (con- taining males only) and those produced by mated mothers (containing mainly females) (logistic regression, rescaled for overdispersion: F1.1 I I = 1.6, deviance explained = I .4%, p > 0.05) suggesting an absence of sexually differential mortality.
The predicted optimal numbers of male eggs (calculated using the mean mortality, m = 0.248, see Heimpel 1994; Hardy dk Cook 1995) were 1 for clutches of up to six eggs and 2 for clutches of seven to 18 eggs. The predicted proportions O f all-female broods for these clutch size classes are, respectively, 0.248 (=m) and 0.061 5 (=m’). The observed overall proportion of all-female (virgin) broods (0.41) Was higher than predicted (G-test: x’ = 34.95, df = 1, p < 0.001) but, as predicted by Heimpel ( 1994). the proportion of all-female broods decreased as clutch size increased (logistic regression: x’ = 9.476, df = I , deviance explained = 37%. P < 0.005, Fig. 4).

66
0 8 - .- 2 . C
:@ 0 6 -
c . .- 0 4 - ::
2 . Q
0 2 -
I. C. W. Hardy, J. B. Pedersen, M. K. Sejr & U. H. Linderoth I . 7
o ~ . . . . . . . . . . . . . . . = , ; . . 2 4 6 8 10 12 14 16 18 20
Clutch sue Fig. 4: Proportion of all-female (virgin) broods in relation to the clutch size from which the broods developed. The stepped line is the expected prevalence of virginity (assuming binomial mortality with the observed mean). Data points are means per clutch size class and the thick curve is the fitted logistic regression. The thin curve describes data presented
in Fig. 2. of Hardy & Cook (1995). The curves are not significantly different (see text)
Discussion
Several authors have stressed the importance of mating and dispersal behav- iour to the understanding of bethylid sex ratios but the evidence has been largely incidental, indirect or absent (e.g. Griffiths & Godfray 1988; Hardy 1994a; Hardy &Cook 1995; Mayhew & Godfray 1997; Hardy & Mayhew 1998; see Introduction). In G. neplzantidis in particular, the high proportion of all-female broods reported by Hardy & Cook (1995) fits the theoretical predictions poorly and presented a conundrum. We first discuss new evidence for mortality and virginity in G. n c y h n - tidis and then consider its probable mating structure.
Consistency of Sex Ratio, Mortality and Virginity Estimates
Experimental artefacts offer a potential explanation for the high prevalence of virginity in G. nephantidis (Hardy & Cook 1995). For example, if male devel- opmental mortality were abnormally high, the proportion of virgin broods would be inflated. The repetition of the analysis of Hardy & Cook (1995) on our new data has however, produced consistent results. The mean sex ratios estimated in our study and by Hardy & Cook (1995) (0.0896 and 0.093, respectively) are almost identical (estimates from previous studies are summarized in Hardy &Cook 1995). Both studies found significantly underdispersed sex ratios and no trend in sex ratio (and an increase in number of males) as brood size increases. Mortality estimates were also similar: Hardy & Cook (1995) estimated the mean probability of mor- tality as 0.28 and in this study the estimate is 0.25. In both studies mortality was

Mating. Dispersal and Sex Ratio in a Parasitoid Wasp 67
overdispersed, with no difference between the sexes and no trend with clutch size. Virginity in both studies was more prevalent than predicted and the proportions of virginity in the two studies were not significantly different (G-test: x’ = 2.32, df = 1, p > 0.1). In both studies the proportion of virginity decreased with clutch size (in qualitative agreement with predictions) and differences between these decreases (Fig. 4) were not significant (x ’ = 0.43, df = I , deviance explained =
The experiments by Hardy & Cook (1995) and in this study were carried out under similar, but not identical, physical conditions, in different laboratories and using different strains of G. nepltuntidis. We conclude that, because results appear insensitive to minor details, high prevalences of virginity are unlikely to be due to experimental artefacts, nor are they likely to be due to the use of a facultative host species (Hardy & Cook 1995). Although we have not been able to assess mortality and virginity in the field, we consider that virginity in G. neppliuntidis is likely to be quite common in nature and that the behaviour of individuals developing in all- female and all-male broods warrants consideration.
0.7%, p > 0.1).
Local Mating and Insemination Capacity
Many sex ratio models assume that local mating occurs and that one male is able to inseminate all females developing in a given brood (Hamilton 1967; Green et al. 1982; Griffiths & Godfray 1988; Heimpel 1994; Nagelkerke & Hardy 1994). The assessment of male insemination capacity may be complicated by the occur- rence oftemporary sperm depletion if males are presented with mates in abnormally rapid succession (Nadel & Luck 1985; Ode etal. 1996; literature cited in Quicke 1997). Preliminary data suggested this may also apply in G. neplruntidis (Hardy 1991). In the present study, males were ‘presented’ with mating opportunities as their sisters matured, eclosed and dispersed naturally. Our finding that males inseminate virtually all females in their natal broods, even when brood sizes are large, supports the assumptions of the above models.
Our data also suggest that limited insemination ability can be rejected as an explanation (Hart1 197 1 ; Hardy et al. 1998) for the increasing number of males per brood as brood size increases. Under strict local mating, the remaining (non- mutually exclusive) alternatives are a primary sex ratio response to anticipated developmental mortality, and incomplete control of sex allocation (Green el a\. 1982; Nagelkerke & Hardy 1994; Hardy & Cook 1995; Hardy etal. 1998). An increased primary sex ratio in response to mortality is expected within the observed Clutch size range given the estimated mortality (Green et al. 1982) and provides a Strong candidate explanation. However, despite evidence for an underdispersed sex ratio in this and many other bethylid species (Hardy 1992; Hardy et al. 1998). there appears to remain a stochastic component to sex allocation: the available primary sex ratio data indicate that G. nephuntidis clutches commonly contain no males at oviposition (32-37’/0; Hardy & Cook 1995; this study), which may account for the observed increase.

68 I. C. W. Hardy, J. B. Pedersen, M. K. Sejr & U. H. Linderoth
Dispersal
The modelling assumption that only mated females disperse from the natal site is clearly not met in G. nephantidis since virtually every male and female dispersed, including unmated individuals. Complete dispersal has also been reported in the gregarious eulophid Co/poc/ypeus,florus (Walker), but only mixed- sex broods were examined (Dijkstra 1986). A potential explanation for why virgin female G. nephantidis appear to disperse from all-female broods sooner after eclosion than do mated females from mixed-sex broods is that both types of female disperse at similar ages after initial pupation, but mixed-sexed brood females eclose relatively early owing to males chewing open and entering their pupal cocoons. Alternatively, unmated females may disperse earlier in order to have more time to seek mates elsewhere. This is likely since mating status is expected to affect greatly the reproductive success of female gregarious parasitoids (Godfray 1994; Hardy 1994b). In Trichoyramma minufum Riley, unmated females tend to disperse later than mated females (Forsse et al. 1992), perhaps because the hosts of T. minutum are more aggregated than hosts of G. nephantidis and thus female T. minurum are more likely to encounter males if they remain at the natal site rather than dispersing.
In G. nephuntidis, our expectation was that, if males dispersed at all, males in all-male broods would have dispersed earlier than males in mixed-sexed broods because males developing in single-sex broods can only attain mating success by dispersal and nonlocal mating. Instead, males in all-male broods disperse later. Selection could have favoured dispersal primarily after mating if most males are the descendants of individuals in mixed-sex broods, but since all-male broods will be about as common as all-female broods (the mothers of all-male broods will mainly derive from all-female broods) i t seems surprising that males may not be able to discriminate the sexual composition of their natal brood and disperse earlier if females are not present.
Post-Dispersal Behaviour of Virgin Females
Dispersed males may search for immature all-female broods and mate with the females as these mature. However, many females may disperse as virgins and these will be constrained to produce male offspring only, unless mating also takes place away from the natal site. The dispersal of virtually all males in our experi- ments suggests that nonlocal mating opportunities are present, although we have no quantitative estimate.
Cook (1993) used mother-son mating to investigate the mechanism of sex determination in G. nephantidis, but since mothers were kept at low temperatures it was unknown whether mother-son mating might occur under more normal conditions, as reported in at least one other bethylid (Cephalonomia quadridentala Duchaussoy, van Emden 193 1 ; Hamilton 1967). We have shown that under normal conditions virgin female G. nephantidis can produce a brood of sons, mate with these and subsequently produce mixed-sex broods. However, in our experiments the majority of females died before their sons eclosed ( z 14 d after oviposition)

Mating, Dispersal and Sex Ratio in a Parasitoid Wasp 69
and only one of the five females that produced a second brood had mated with her sons. Virgin mothers survived for less time than the majority of mothers which were given subsequent hosts when the brood pupated (about 7 d after oviposition) rather than eclosed (Hardy et al. 1992), probably due to the more frequent host- feeding opportunities in the earlier experiments. However, virgin mothers also survived for less time than most females kept with neither hosts nor feeding opportunities (Hardy et al. 1992). We conclude that while mother-son mating may occur in nature, it is unlikely to be frequent owing to maternal mortality.
Hardy & Cook ( 1 995) suggested that females may circumvent the constraint of virginity by adding a few eggs to existing broods produced by mated females, thus providing their sons with local mating opportunities. However, virgin G. nc~phantidis females do not refrain from full ovicide when presented with a host bearing a mixed sex brood: the suggested strategy is clearly not adopted.
Criticism of the Experiment
The investigations reported here face the classic problem of extrapolation from the laboratory to the natural world. We have discussed potential artefacts from rearing G. nephuntidis on a facultative host elsewhere (Hardy & Cook 1995; see also Remadevi et al. 1994) and our results further indicate that mortality is not sensitive to small variations in other experimental conditions. Although laboratory mortality may differ from that on the natural host in the field, it is not clear whether field mortality is likely to be lower or higher than our estimate (see Kapadia & Mittal 1986; Hardy & Cook 1995) and only higher mortality could explain an abnormally high proportion of virgin broods. Although their exact frequency is Unknown, we expect single-sex broods to occur in the field because these have been found in several other bethylid species, and in field broods of other gregarious Pdrasitoids (Hardy et al. 1998).
As in nature, wasps eclosed in a confined place and had to pass through a tunnel to disperse. We performed intermittent rather than continuous observations because each replicate required observations and preobservation checking for more than 6d, because replicates were not fully synchronized and because adult G. ncydlanti&s of both sexes are generally inactive. Accordingly, we cannot exclude the possibility that individuals re-entered the chambers post dispersal, which would lead to an underestimate of dispersal rates. However, the openings in the blocks were small in relation to the general surface of the interior, SO re-entry prior to the following observation period was relatively unlikely (and re-entry is also possible in nature). We were also unable to identify wasps on an individual basis because individuals of each sex look alike and hence the statistical analysis of dispersal is hampered. Even with continuous observations, identification of individuals would have been highly prone to error. Marking individuals is possible (Hardy & Black- burn 1991; Petersen & Hardy 1996) but would have disturbed the experiment to the point of no value. The strengths of this methodology are: 1. whether and when individuals disperse following eclosion can be observed; and 2. the capacity of males to inseminate females is estimated with mating opportunities presented as

70 I. C. W. Hardy. J. B. Pedersen, M. K. Sejr & U H. Linderoth
individuals mature and disperse over a natural time scale. This provides an estimate of more relevance to the assumptions of sex ratio theory than presenting males with a succession of virgin females at a fixed rate.
Conclusions
We have shown that, in agreement with the assumptions of local mate com- petition theory, individual male G . nephuntidis are capable of inseminating (vir- tually) all of their sisters before these disperse to forage for fresh hosts. The larger number of males in larger clutches cannot thus be explained as a sex allocation response to limited mating capacity. Sex ratio models which assume strict local mating may not be fully applicable to G . nephantidk because single-sex broods are common and both males and virgin females disperse from the natal patch. As for other bethylids (Hardy & Mayhew 1998), quantitative estimates of host distribution and the prevalence of post-dispersal (nonlocal) mating in G . nc1phuntidi.s are clearly needed. Future investigations will employ genetic techniques (e.g. Kazmer & Luck 1991; Molbo & Parker 1996) to complement these behavioural observations.
Acknowledgements
We are grateful to P. J. Mayhew, P. J. Ode, S. Stokkebo, E. D. Parker and an anonymous referee for comments on the manuscript. I.C.W.H. was funded by European Commission postdoctoral research training grant ERBFMBICT 961025 and additionally supported by grants from the Carlsberg Foundation and the Danish National research Council to E. D. Parker. Equal contributions to this workweremadebyJ .B .P . ,M.K.S .andU.H.L .
Literature Cited
Barrera, J. F., Infant. F., Alauzet. C., Gomez, J., De La Rosa. W. & Castillo. A. 1993: Biologie de Cepku/ononriu .s/c~p/iurroderis Betrem (Hymenoptera: Bethylidae) en laboratoire. 11. Duree. de developpement, sex-ratio. longevite et esperance, de vie des adultes. Cafe Cacao The 37,205-214.
Cock, M. J. W. & Perera, P. A. C. R. 1987: Biological control of Opisinu urenoselku (Lepidoptera. Oecophoridae). Biocontrol. News Info. 8, 2 0 - 3 10.
Cook, J. M. 1993: Experimental tests of sex determination in G o n i o x s nc~pkon~ic/i.~ (Hymenoptera: Bethylidae) Heredity 71, 130 - 137.
Crawley, M. J . 1993: GLlM for Ecologists. Blackwell Sci. Publ.. Oxford. Dljkstra, L. J . 19x6. Optimal selection and exploitation of hosts in the parasitic wasp Co/poc./ypeu.\
van Emden, F. 1931: Zur Kenntnis der Morphologie und des Brotklfer-Parasiten Cep/iei/cJnofnicr
Fisher, R. A. 1930 The Genetical Theory of Natural Selection. Clarendon Press, Oxford. Forsse, L.. Smith. S. M. & Bourchier. R. S. 1992: Flight initiation in the egg parasitoid Trit.ho.q?qrumnm
niitiuluni Riley: effects of ambient temperature. mates. food, and host eggs. Entomol. Exp. Appl.
Godfray, H. C. J . 1994: Parasitoids. Behavioral and Evolutionary Ecology. Princeton Univ. Press. Princeton.
Gordh, G., Woolley, J. B. & Medeved, R. A. 1983: Biological studies on Goniozus k:qnciri Gordh (Hymenoptera: Bethylidae) primary external parasite of the navel orangeworm Anfjtchis /run.si/i~//e~ and pink bollworm Pac./mop/ioru g ~ ~ . ~ j . p i ~ . l k i (Lepidoptera: Pyralidae: Gelechiidiie). Contrib. Am. Entomol. Inst. 20,433-468.
f/urus (Hym., Eulopidae). Neth. J. 2001. 36, 177-301.
qiiudri&n/uiu Duchaussoy. 2. Morphoi. Okol. Tiere. 23,425-547.
62, 147 ~ 154.

Mating. Dispersal and Sex Ratio in a Parasitoid Wasp 71
Green. R. F., Gordh. G. & Hawkins, B. A. 1982: Precise sex ratios in highly inbred parasitic wasps.
Griffiths. N. T. & Godfray, H. C. J. 1988: Local mate competition sex ratio and clutch size in bethylid
Hamilton. W. D. 1967: Extraordinary sex ratios. Science 156,477488. Hardy, 1. C. W. 1991: Reproductive strategies in parasitic wasps. PhD thesis, Univ. of London. London. Hardy. 1. C. W. 1992 Non-binomial sex allocation and brood sex ratio variances in the pdrdsitoid
Hardy. 1. C. W. 1994a: Sex ratio and mating structure in the parasitoid Hymenoptera. Oikos 69, 3-
Hardy. 1. C. W. 1994b Polyandrous parasitoids: multiple-mating for variety's sake? Trends. Ecol. Evol.
Hardy, 1, C. W. & Blackburn. T. M. 1991: Brood guarding in a bethylid wasp. Ecol. Entomol. 16.55- 62.
Hardy. 1. C. W. & Cook, J. M. 1995: Brood sex ratio variance developmental mortality and virginity in a gregarious parasitoid wasp. Oecologia 103, 162-169.
Hardy, I . C. W. & Mayhew, P. J. 1998: Sex ratio. sexual dimorphism and mating structure in bethylid wasps. Behav. Ecol. Sociobiol. 42,383--395.
Hardy. 1. C. W.. Grifiths, N . T. & Godfray. H. C. J. 1992: Clutch size in a parasitoid wasp: a manipulation experiment. J . Anim. Ecol. 61, 121-129.
Hardy. I . C. W , Dijkstra., L. J.. Gillis. J. E. M. & Luft, P. A. 1998: Patterns of sex ratio virginity and developmental mortality in gregarious parasitoids. Biol. J. Linn. SOC. 64, 239---270.
Hartl. D. L. 1971: Some aspects of natural selection in arrhenotokous populations. Am. 2001. 11,309- 325.
Heimpel. G . E. 1994: Virginity and the cost of insurance in highly inbred Hymenoptera. Ecol. Entomol. 19,299-302.
Kapadia. M. N. & Mittal. V. P. 1986: Biology of Prrrciscwolcr ric,plifrn/rdi,s Muesebeck and its importance in the control of Opisiiici urerto.w/lu Walker under Mahuva (Gujurat State) conditions. GAU Res.
Kazmer. D. J. & Luck. R. F. 1991: The genetic-mating structure of natural and agricultural populations of T,.ic./toJ~,.N/ri~,ll~. In: Proc. 3rd Int. Symp. Tric,/rc),~rcrninra and Other Egg Pdrasitoids (Wajnberg. E. & Vinson. S. 6.. eds). INRA. Paris. pp. 107-1 10.
Koch. V. J. M. 1973: Abondance de Hypat/icw.riiu.s /runrpivY Ferr. Scolyte des graines de caf6 en fonction de sa plante-hbte et de son parasite C'cyhdoriomrcr stc~p/iunoderi.s Bertrem en Cote Dlvoire. PhD thesis. Agric. Univ. of Wageningen, Wageningen
Mayhew. P. J. 1997: Fitness consequences of ovicide in a pamitoid wasp. Entomol. Exp. Appl. 84,
Mayhew. P. J. & Godfray, H. C. J. 1997: Mixed sex allocation strategies in a parasitoid wasp. OKOlOgid
Molbo. D. & Parker Jr. E. D. 1996: Mating structure and sex ratio variation in a natural population of Nu.wtrru ritripwrii.$. Proc. R. SOC. Lond. B. 263, 1703- - 1709.
Morgan, D. J. W. & Cook. J. M. 1994: Extremely precise sex ratios in small clutches of a bethylid wasp. Oikos 71,423430.
Nadel, H. & Luck. R. F. 1985: Span of female emergence and male sperm depletion in the female- biased quasi-gregarious parasitoid Pcrc/i,~c~rc,poic/c.u.s firidcwftiiue ( Hymenoptera: Pteromalidae). Ann. Entomol. Soc. Am. 78,410 414.
Nagelkerke. C. J. & Hardy, I . C. W 1994: The influence of developmental mortality on optimal sex allocation under local mate competition. Behav. Ecol. 5 , 4 0 1 4 1 I .
Nagelkerke, C. J. & Sabelis, M. W. 1991: Precise sex ratio control in the pseudo-arrhenotokous pliytoseiid mite ~~p/i/oc/rortiits occrc/cvifeilis Nesbitt. In: The Acari: Reproduction. Developnient. and Life-History Strategies (Schuster. R. & Murphy P. W.. eds). Chapman & Hall, London. pp. 193 707.
Nunncy. L. & Luck. R. F. IYXX: Factors influencing the optimum sex ratio in a structured population. Theor. Pop. Biol. 33, 1-30.
Ode, P. J.. Antolin. M. F. & Strand. M. R. 1996: Sex allocation and sexual asymmetries in intra-brood competition in the parasitic wasp Brcrcwrr l r i h ~ t ~ r . J. Anim. Ecol. 65, 690--700.
Am. Nat. 120,653-665.
wasps. Behav. Ecol. Sociobiol. 22,211-217.
Hymenoptera. Oikos 65,143-1 58.
20.
9,202-203.
J. 12, 29-34.
115-126.
110,218-221.

72 I. C. W. Hardy, J. B. Pedersen, M. K. Sejr & U. H. Linderoth
Petersen, G . & Hardy, I. C. W. 1996: The importance of being larger: pdrasitoid intruder-owner
Quicke. D. L. J. 1997: Parasitic Wasps. Chapman and Hall London. Remadevi, 0. K. Sasidharan, T. 0. & Abdurahiman, U. C. 1994: Ovisite preference of Goniozus
contests and their implications for clutch size. Anim. Behav. 51, 1363-1373.
nepiluntirlis Mues. (Hymenoptera: Bethylidae). Insect Sci. Applic. 15,439-443.
Rrcciivd: Nnwmher 19. I997
Initial acceptance: April 22.1998
Final acccpiunce: August I . 1998 (K. Le~scll.~)





























