Embed Size (px)
Citation preview

Accepted by L. Page: 22 Jun. 2007; published: 19 Oct. 2007 1
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2007 · Magnolia Press
Zootaxa 1618: 1–50 (2007) www.mapress.com/zootaxa/
Phylogeny of species formerly assigned to the genus Archocentrus (Perciformes: Cichlidae)
JUAN J. SCHMITTER-SOTOEl Colegio de la Frontera Sur (ECOSUR), A.P. 424, MX-77000 Chetumal, QR, [email protected]
Table of contents
Abstract . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1Resumen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2Material and methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3Character description and analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 45Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
Abstract
A phylogeny for 28 cichlid species that have been included in or near the same clade as Archocentrus centrarchus (typespecies of the genus) in previous phylogenetic analyses is presented, based on 98 morphological characters (osteology, gut-coiling pattern, pigmentation, squamation, meristics, and others), plus one cytogenetic character. Monophyly is supported forArchocentrus sensu stricto, Cryptoheros, Hypsophrys, Amatitlania, and Rocio; the relationships among these genera are notresolved. The three subgenera of Cryptoheros are also supported; Cr. panamensis is the sister group of the rest of the speciesin the genus. Within Cryptoheros (Cryptoheros), Cr. chetumalensis is the sister group of the clade Cr. spilurus + Cr.cutteri. Within Amatitlania the pattern is: (Am. coatepeque (Am. nigrofasciata (Am. siquia + Am. kanna))).
Key words: phylogeny, cichlids, Middle America
Resumen
Se presenta una filogenia para 28 especies de cíclidos que han aparecido dentro o cerca del mismo clado que Archocentrus
centrarchus (especie tipo del género) en hipótesis filogenéticas previas, con base en 98 caracteres morfológicos (osteología,
patrones de convolución intestinal, coloración, escamación, merística y otros), además de un carácter citogenético. Se
demuestra monofilia para Archocentrus sensu stricto, Cryptoheros, Hypsophrys, Amatitlania y Rocio; las interrelaciones de
estos géneros no quedan resueltas. Se apoya también la monofilia de los tres subgéneros de Cryptoheros; Cr. panamensis es
la especie hermana de las demás en ese género. Dentro de Cryptoheros (Cryptoheros), Cr. chetumalensis es hermana del
clado Cr. spilurus + Cr. cutteri. Dentro de Amatitlania, el patrón es: (Am. coatepeque (Am. nigrofasciata (Am. siquia +
Am. kanna))).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO2 · Zootaxa 1618 © 2007 Magnolia Press
Introduction
The cichlid fish genus Archocentrus Gill has been redefined by Schmitter-Soto (2007) to include only Ar.centrarchus Gill in Gill & Bransford, 1877 (type species), Ar. spinosissimus (Vaillant & Pellegrin, 1902), andAr. multispinosus [formerly Herotilapia multispinosa (Günther, 1867)]. Other species that have sometimes beenassigned to Archocentrus were reassigned in that paper to the new genera Amatitlania [Am. nigrofasciata(Günther, 1867) and three new species] and Rocio [R. octofasciata (Regan, 1903) and two new species], as wellas Cryptoheros Allgayer [Cr. panamensis (Meek & Hildebrand, 1913), Cr. spilurus (Günther, 1862), theresurrected Cr. cutteri (Fowler, 1932), Cr. septemfasciatus (Regan, 1908), Cr. altoflavus Allgayer, 2001, Cr.nanoluteus (Allgayer, 1994), Cr. myrnae (Loiselle, 1997), Cr. sajica (Bussing, 1974), and one new species] andHypsophrys Agassiz [H. nicaraguensis Günther, 1864 and H. nematopus (Günther, 1867)]. The present articleprovides the cladistic support for these decisions.
Attempts to solve the ex-Cichlasoma problem have had a less-than-moderate success. Cichocki’s (1976)dissertation supported the hypothesis that all species then assigned to Cichlasoma Swainson formed amonophyletic group, and proposed several lineages within it; however, he chose to work out the relationships bymeans of a clique analysis, thus rendering his results hard to compare with subsequent parsimony-basedhypotheses (Kullander 1996). Stiassny (1991) presented a review of cichlid relationships; she demonstrated monophyly for the New Worldcichlids and established two main clades, one of which, her “cichlasomine group A,” included all MiddleAmerican cichlids along with a few taxa from South America (e.g. Caquetaia Fowler).
Kullander (1998) examined South American cichlids and established the tribe Heroini, which includes,among others, the species formerly in Cichlasoma; he did not offer resolved relationships within the Heroini (andhe did not include species from Middle America), but nevertheless he concluded that “the morphological basis forphylogeny reconstruction is practically available.”
Roe et al. (1997) performed the first molecular analysis of Middle American cichlids, using completecytochrome b sequences of 19 species. These authors found that Cr. spilurus and Nandopsis (= Parachromis)dovii (Günther) were part of the same clade as the true Amphilophus Agassiz [restricted by these authors to A.citrinellus (Günther) and A. labiatus (Günther)]. They found this clade to be basal to the “cichlasomine group A”clade, i.e. the Heroini (Kullander 1998).
The same gene allowed Martin and Bermingham (1998) to produce a partial phylogeny of Archocentrus, inthe context of an analysis of 21 Middle American (almost exclusively Costa Rican) cichlids. In the present work,some of Regan’s (1905) “sections” were recognised, including a restricted version of Archocentrus. However, theauthors felt that the “relationships among the genera remain elusive,” except for the monophyly of Hypsophrys +Neetroplus Günther (a clade that apparently contradicted morphological phylogenetic analyses, although it issupported in the present paper).
Farias et al. (1999, 2000, 2001) essentially focused on South American taxa, but they did include additionaltaxa from Middle America, among them ‘Cichlasoma’ octofasciatum, Archocentrus citrinellus (sic), and Ar.nigrofasciatum, which showed up together in one clade with Petenia splendida Günther and Caquetaiaspectabilis (Steindachner); hence the choice of additional material for the present study.
The phylogenetic reconstruction here presented establishes the monophyly and species composition ofArchocentrus, along with a redefinition of Cryptoheros and Hypsophrys, and supports the diagnosis of two newlydescribed genera (Amatitlania and Rocio—Schmitter-Soto, 2007). The cladistic analysis treats all nominalspecies ever assigned to the genus Archocentrus and six newly described species (Schmitter-Soto, 2007), as wellas the species that have been included in the same clade as (or as sister group to) Ar. centrarchus in relevantpublished phylogenetic analyses (see above).
Diagnoses and descriptions are given in Schmitter-Soto (2007), where an identification key is alsoavailable.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 3PHYLOGENY OF THE GENUS ARCHOCENTRUS
Material and methods
The specimens examined include representatives from all nominal species that have at various times beenassigned to the genus Archocentrus in scientific publications (Schmitter-Soto, 2007: table 1), as well ascomparative material of species that appeared in the same clades as species assigned to Archocentrus in Roeet al. (1997), Martin and Bermingham (1998), and Farias et al. (2000) (Table 1). Material examined is thesame listed by Schmitter-Soto (2007: appendix 1).
TABLE 1. List of species included in this phylogenetic analysis. Schmitter-Soto (2007) provides details on materialexamined; names in brackets refer to the name more frequently used in recent literature (e.g. Kullander 2003). Petenia,Tomocichla, and Oreochromis were found to lie outside the ingroup.
Pigmentation patterns were assessed primarily from preserved specimens. Osteological methodology (i.e.,clearing and staining) follows Taylor and van Dyke (1985, as modified by W.L. Fink, pers. comm.).Radiographs and skeletonised specimens were also used. Bone measurements were taken with an ocularmicrometer.
Because the choice of a sister group was not evident, the comparative material was considered part of theingroup, as a test of monophyly for Archocentrus. An African cichlid, Oreochromis mossambicus (Peters),was used as outgroup to root the tree (same choice of Roe et al. 1997). Cladistic analyses were based on
Amatitlania nigrofasciata [=Archocentrus nigrofasciatus]Am. coatepeque Am. kannaAm. siquiaAmphilophus citrinellus Archocentrus centrarchus Ar. multispinosus [=Herotilapia multispinosa]Ar. spinosissimusCaquetaia spectabilisCryptoheros (Panamius) panamensis
[=Archocentrus panamensis]Cr. (Bussingius) altoflavus [=Ar. altoflavus]Cr. (B.) myrnae [=Ar. myrnae]Cr. (B.) nanoluteus [=Ar. nanoluteus]Cr. (B.) sajica [=Ar. sajica]Cr. (B.) septemfasciatus [=Ar. septemfasciatus]Cr. (Cryptoheros) spilurus [=Ar. spilurus]Cr. (Cr.) chetumalensisCr. (Cr.) cutteri Hypsophrys nicaraguensisH. nematopus [=Neetroplus nematopus]Parachromis doviiP. loiselleiRocio octofasciata [=‘Cichlasoma’ octofasciatum]R. gemmataR. ocotalPetenia splendidaTomocichla sieboldii [=Paraneetroplus sieboldii]Oreochromis mossambicus
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO4 · Zootaxa 1618 © 2007 Magnolia Press
maximum parsimony, and reconstructions were performed with PAUP* 4.0 (Swofford 1998). I used heuristicsearch and strict consensus; character states were considered unordered, multistate, or binary; all received thesame weight. Meristic characters were used only if satisfactorily amenable to statistical coding: their stateswere determined by means of an ANOVA followed by a Duncan a posteriori test (homogeneous subsetcoding, Simon 1983); where differences were significant at α < 0.05, homogeneous groups with no overlapwere chosen as different character states. Optimisation followed the ACCTRAN option in order not to lose anyputative synapomorphy (Kitching et al. 1998).
Homoplasy level in the most parsimonious cladogram was evaluated with consistency (CI) and retention (RI)indices. Homoplasy (hi) and retention (ri) indices were also computed for every character, and synapomorphieswere considered relevant only when strict, or when the concerned character had a hi 0.50, or a hi > 0.50 but com-pensated by a relatively high ri (Kitching et al. 1998). Robustness of internal nodes was evaluated with a 1000-replicate bootstrap analysis (Felsenstein 1985). Branches with bootstrap support below 50% were collapsed,unless they displayed at least one strict synapomorphy.
ResultsCharacter description and analysis
The list treats 98 characters (see matrix: Table 2), selected mostly because they were amenable to reasonablyobjective coding. Many of them have been used before, but all relevant anatomical material was reevaluated;I compare my interpretations mainly against those of Cichocki (1976) and Kullander (1998). Many characters(43) are binary, but most are multistate, non-additive, unordered. I did not exclude terminal autapomorphies(12, which leaves 86 informative characters) because they provide special support for species diagnoses(Schmitter-Soto, 2007). I did not leave out highly homoplasious traits either: “Dismissing conflictingcharacters […] might be defended for small sets of taxa and characters. For more complex situations,however, selection of characters not only introduces a bias but also leaves out potentially useful charactersets” (Kullander 1998: 471). Homoplasy (hi: Archie 1989) and retention (ri: Farris 1989) indices are indicatedwhere relevant. Characters for species outside Archocentrus, Cryptoheros, Amatitlania, Rocio, andHypsophrys are usually not illustrated.
Gill arches, characters 1–111. Angle between arms of first epibranchial bone[0] arms divergent (Fig. 1a); [1] arms parallel (Fig. 1b).
Homoplasious (hi=0.89, ri=0.20). Fixed for state 1 in Rocio, but occurring in several other taxa.Cichocki’s (1976) character 30, “relative direction of the posterior arm [uncinate process] of the firstepibranchial bone,” distinguished three degrees of divergence: an angle of 35° or more, 15–30°, or less than15°; however, the polarity is the same. He found Am. nigrofasciata to have my state 0, but in my view thatspecies has parallel arms, although the rest of the genus Amatitlania has divergent arms.
This is also Kullander’s (1998) character 3, “angle of the uncinate process of epibranchial 1 relative to themain axis of the bone;” same polarity as here, although Kullander united Cichocki’s first two states andincluded a new state for those taxa where both limbs are parallel but angled relative to the bone axis.
2. Relative width of arms of first epibranchial [0] arms subequal; [1] posterior arm distinctly wider.
Homoplasious (hi=0.88, ri=0.25); fixed for state 1 in Archocentrus, polymorphic in Cr. chetumalensis.Polarity as in Kullander’s (1998, character 4), who observed that the “derived condition may have beendeveloped by increased relative width of the uncinate process or decreased width of the anterior arm.”
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 5PHYLOGENY OF THE GENUS ARCHOCENTRUS
TABLE 2. Character matrix used for parsimony analysis of character distribution in Cichlidae once or presently assigned
to the genus Archocentrus. See cladogram in Fig. 26.
...... continued on the next page
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
mossambicus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
sieboldii 0 0 1 4 1 0,1 0 0 2 0 1 0 2 2 0 0 1 0 1 0
splendida 0 1 4 1 0 0 1 0 1 1 0 1 0 1 1 0 1 1 1 1
dovii 0 0 1 3 0 1 1 0 2 0 1 2 1 0 0 0 1 0 1 1
loisellei 0 1 1 3 0 1 1 0 2 0 1 2 1 0 0 0 1 0 1 1
spectabilis 0 1 1 4 2 1 1 0 2 0 1 1 1 5 0 0 1 0 1 1
citrinellus 0 0 2 1 1,5 0 0 2 4 0 1 4 1 4 0 0 1 0 1 2
nicaraguensis 1 1 3 3 1 0 1 0 3 1 1 1 1 1 0 0 1 0 0 2
nematopus 0 0 1 3 1 0 1 0 2 0 1 3 3 3 0 1 1 0 1 0
panamensis 0 0 1 4 1 1 1 ? ? ? ? ? 2 1 0 0 1 0 1 0
chetumalensis 1 0,1 1 4 1 0 1 1 2 0,1 1 5 2 1 2 0 1 0 1 0
cutteri 0 0 1 4 1 0 1 1 2 1 1 5 2 4 2 0 1 0 1 0
spilurus 1 0 1 4 1 0 1 1 2 0,1 1 5 2 1 2 0 1 0 1 0
altoflavus 1 0 1 4 1 1 1 1 2 1 1 5 2 4 2 0 1 0 1 0
nanoluteus ? ? 1 4 1 0 0 1 ? 1 ? ? 2 4 2 0 1 0 1 0
septemfasciatus 0 0 1 4 1 0 0 1 2 1 1 6 4 1 0,2 0 1 0 1 0
myrnae 0 0 1 4 0 0 1 1 2 1 1 6 4 4 0 0 1 0 1 0
sajica 1 0 1 4 1 0 0 1 2 1 1 5 4 4 2 0 1 0 1 2
spinosissimus 1 1 1 4 1 0 1 0 2 0 1 5 2 2 0 0 1 0 1 2
centrarchus 1 1 0 2 3 0 1 0 2 1 1 5 2 5 0 0 1 0 1 2
multispinosus 0 1 1 4 1 0 1 0 2 0 1 3 2 5 2 0 1 0 1 0
gemmata 1 0 1 4 4 1 1 0 2 0 1 6 5 6 0 0 1 0 1 2
octofasciata 1 0 1 4 4 0 0,1 0 2 0 1 6 5 6 0 0 1 0 1 2
ocotal 1 ? 1 4 4 1 0 0 2 0 ? 6 5 6 0 0 1 0 1 2
kanna 0 1 1 4 1,5 1 0 0 2 0 1 3 2 1 0 0 1 0 1 2
siquia 0 0 1 4 1,5 1 0 0 2 0 1 3 4 1 0 0 1 0 1 2
nigrofasciata 1 1 1 4 1 1 0 0 2 0 1 6 2 1 0 0 1 0 1 2
coatepeque 0 0 1 4 1 1 1 0 2 1 1 6 2 1 0 0 1 0 1 2
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO6 · Zootaxa 1618 © 2007 Magnolia Press
TABLE 2 (cont.).
...... continued on the next page
21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40
mossambicus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
sieboldii 0 0 1 0 0 2 0 2 0 1 0 1 2 0 0 0 1 0 0 0
splendida 3 1 2 0 1 2 1 1 0 0 0 1 1 0 0 0 1 1 0 1
dovii 2 0 1 0 1 2 0 1 2 0 0 0 2 0 0 1 0 1 0 2
loisellei 2 0 1 1 1 2 0 1 2 0 0 0 2 0 0 1 0 1 0 2
spectabilis 2 2 3 1 1 2 1 0 0 0 0 1 1 0 0 0 0 1 0 2
citrinellus 1 0 1 1 1 2 0 0 1 0 0 1 2 0 0 0 1 0 1 2
nicaraguensis 0 0 1 0 1 2 0 2 1 0 0 1 2 0 0 1 1 0 0 0
nematopus 0 0 0 0 1 2 2 2 2 0 0 1 2 2 0 2 1 0 0 1
panamensis 0 0 1 1 0 2 0 0 ? 1 0 1 2 0 0 2 1 0 1 ?
chetumalensis 0 0 1 1 1 2 0 0 0 1 0 1 4 0 1 1 1 0 1 1
cutteri 0 0 1 1 1 2 0 0 1 1 0 1 4 0 1 2 0 0 1 1
spilurus 0 0 1 1 1 2 0 0 1 1 0 1 4 0 1 1 1 0 1 1
altoflavus 0 0 1 0 1 2 0 2 1 1 0 1 4 0 1 2 0 0 1 ?
nanoluteus 0 0 1 0 1 2 0 2 1 1 0 1 ? 0 1 2 0 0 1 ?
septemfasciatus 0 0 1 0 1 2 0 2 1 1 0 1 2 0 1 2 0 0 1 1
myrnae 0 0 1 0 1 2 0 2 1 1 0 1 4 0 1 2 0 0 0,1 1
sajica 0 0 1 0 1 2 0 2 1 1 0 1 2 0 1 1 1 0 0 1
spinosissimus 0 0 1 0 1 2 0 0 0 0 1 1 2 0 0 1 1 0 0 2
centrarchus 0 0 1 1 1 2 0 0 0 0 0 1 2 0 0 1 1 0 0 2
multispinosus 0 0 1 1 0 2 0 0 0 0 1 1 2 1 0 1 1 0 0 2
gemmata 1 0 1 0 0 1 0 1 1 0 0 1 4 0 0 1 1 0 1 4
octofasciata 1 0 1 0 0 2 0 0 0 0 0 1 4 0 0 0 1 0 1 3
ocotal 1 0 1 0 0 2 0 0 0 0 0 1 4 0 0 1 1 0 1 3
kanna 1 0 1 0 1 2 0 0 0 1 0 1 4 0 0 2 1 0 1 1
siquia 1 0 1 1 1 2 0 0 1 1 0 1 4 0 0 2 1 0 0,1 3
nigrofasciata 1 0 1 1 1 2 0 0 1 1 0 1 2 0 0 1 0 0 1 3
coatepeque 1 0 1 0 1 2 0 0 1 1 0 1 3 0 0 1 1 0 1 3
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 7PHYLOGENY OF THE GENUS ARCHOCENTRUS
TABLE 2 (cont.).
...... continued on the next page
41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60
mossambicus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
sieboldii 6 1 2 0 0 2 0 3 1 0 1 1 0 1 0 1 0 0 0 2
splendida 1 1 1 1 1 0 1 0 0 3 1 1 0 1 0 1 1 1 0 1
dovii 1 2 3 2 0 0 1 4 0 0 1 1 0 0 0 1 0 1 0 1
loisellei 1 2 3 2 0 0 1 1 0 1 1 1 0 0 0 0 0 1 0 1
spectabilis 1 2 3 2 0 0 1 2 1 2 1 1 0 1 0 0 0 1 0 1
citrinellus 1 1 1 0 0 0 1 1 0 0 1 1 1 1 0 0 0 1 1 2
nicaraguensis 1 1 3 0 0 2 0 1 0 0 1 1 1 0 0 0 0 1 1 2
nematopus 2 1 3 0 0 2 0 1 0 2 1 1 1 0 0 0 0 1 1 2
panamensis 5 1 3 1 0 1 0 1 0 0 1 1 0 0 0 0 0 1 0 3
chetumalensis 1 1 5 0 0 1 0 1 1 0 1 1 1 1 1 0 0 1 0 2
cutteri 1 1 5 0 0 1 0 1 0,1 0 1 1 0 0 0 0 0 1 0 2
spilurus 1 1 5 0 0 1 0 1 1 0 1 1 0 0 0 0 0 1 0 2
altoflavus 4 1 5 0 0 1 0 0 1 0 ? 1 1 1 0 0 0 1 0 2
nanoluteus 4 1 3 0 0 1 0 1 0 0 ? 1 1 1 0 0 0 1 0 2
septemfasciatus 1 1 5 0 0 1 0 1 0 0 1 1 1 1 0 0 0 1 0 2
myrnae 4 2 3 0 0 1 0 0 0 0 1 1 1 1 0 0 0 1 0 2
sajica 4 1 3 0 0 1 0 0 1 0 1 1 1 1 0 0 0 1 0 2
spinosissimus 1 1 3 0 0 0 0 1 1 0 1 1 1 1 0 0 0 2 0 2
centrarchus 1 1 3 0 0 0 1 1 0 1 1 1 0 0 0 0 0 2 0 2
multispinosus 3 1 4 0 0 1 0 1 1 1 1 1 0 1 0 0 0 2 0 2
gemmata 1 2 5 1 0 0 0 1 0 0 1 1 0 1 0 0 0 1 0 1
octofasciata 1 2 5 1 0 0 0 1 0 0 1 1 1 0,1 0 0 0 1 0 2
ocotal 1 2 1 1 0 0 0 1 0 0 1 1 0 1 0 0 0 1 0 2
kanna 1 2 3 1 0 1 0 1 0,1 0 1 1 1 1 1 0 0 1 0 2
siquia 1 2 3 1 0 1 0 1 0 0 1 1 1 1 1 0 0 1 0 2
nigrofasciata 1 2 3 1 0 1 0 1 0,1 0 1 0,1 1 1 0 0 0 1 0 2
coatepeque 1 2 3 1 0 1 0 1 0 0 1 1 0,1 1 0 0 0 1 0 2
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO8 · Zootaxa 1618 © 2007 Magnolia Press
TABLE 2 (cont.).
...... continued on the next page
61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80
mossambicus 0 0 - 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 -
sieboldii 0 1 0 1 2 1 0 2 0 0 0 1 1 0 0 0,1 1 0 2 0
splendida 0 0 - 0 ? 1 1 1 1 0 1 1 1 0 1 1 1 0 1 0
dovii 0 1 0 1 0 0 1 0 0 0 0 2 1 0 0 0 1 1 1 0
loisellei 0 1 0 1 0 2 1 0 0 1 1 2 1 1 0 0 1 1 1 1
spectabilis 0 2 0 2 2 2 0 1 2 0 1 2 1 1 0 0 3 0 1 1
citrinellus 0 1 2 1 0 2 0 1 0 0 1 2 1 1 0 0 1 0 1 1
nicaraguensis 0 1 0 1 0 2 0 2 0 1 2 2 1 1 0 0 0 0 1 1
nematopus 0 1 2 1 0 0 0 2 0 1 0 2 1 1 2 0 0 0 7 1
panamensis 0 1 0 1 0 2 0 0 0 1 1 2 1 1 2 1 1 0 5 ?
chetumalensis 0 1 2 1 2 2 0 1 0 1 1 2 1 1 0 0,1 1 0 5 1
cutteri 0 1 2 1 2 2 0 2 0 1 1 2 1 1 0 2 1 0 5 1
spilurus 0 1 2 1 2 2 0 2 0 1 1 2 1 1 0 0,1 1 0 5 1
altoflavus 0 1 3 1 2 2 0 1 0 1 1 2 1 1 0 1 1 2 5 1
nanoluteus 0 1 3 1 2 2 0 1 0 1 1 2 1 1 0 1 1 2 5 ?
septemfasciatus 0 1 0 1 0 2 0 0 0 1 1 2 1 1 0 1 1 2 5 1
myrnae 0 1 3 1 0 2 0 0 0 1 1 2 1 1 0 1 1 2 1 1
sajica 0 1 0 1 0 2 0 1 0 1 1 2 1 1 0 1 1 2 1 1
spinosissimus 1 1 2 1 3 2 1 0 0 1 1 3 1 1 0 2 1 3 1 0
centrarchus 0 1 0 1 2 2 0 2 0 1 1 3 1 1 0 2 2 3 1 1
multispinosus 0 1 1 1 1 2 1 0 0 1 1 3 1 1 0 2 2 3 4 -
gemmata 1 1 0 1 0 2 1 0 0 1 0 2 1 0 0 2 1 0 1 0
octofasciata 1 1 2 1 0 2 1 0 0 1 0 2 1 1 0 2 1 0 1 1
ocotal 1 1 2 1 0 2 1 1 0 1 0 2 1 1 0 1,2 1 0 1 1
kanna 0 1 2 1 0 2 0 0 0 1 1 2 1 1 0 1 1 0 1 1
siquia 0 1 0 1 0 2 0 1 0 1 1 2 1 1 0 1 1 0 3 1
nigrofasciata 0 1 2 1 2 2 0 1 0 1 1 2 1 1 0 1 1 0 1 1
coatepeque 0 1 0 1 0 2 0 2 0 1 1 2 1 1 0 1 1 0 6 1
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 9PHYLOGENY OF THE GENUS ARCHOCENTRUS
TABLE 2 (cont.).
81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98
mossambicus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 - -
sieboldii 0 0 0 1 1 1 1 0 1 0 0 1 1 4 0 1 0 0
splendida 1 0 0 1 1 1 0 2 - 0 0 1 2 0 0 1 0 0
dovii 4 0 1 1 1 1 1 1 0 0 0 1 2 1 0 1 0 0
loisellei 3 0 0 1 0 5 1 1 0 0 0 1 3 1 0 1 0 0
spectabilis 2 2 0 0 0 2 1 0 0 0 0 1 0 1 0 1 1 0
citrinellus 4 2 1 0 0 3 0,1 1 0 0 0 1 0 0,1 0 1 1 1
nicaraguensis 4 0 0 0 0 1 1 2 - 0 0 1 0 1 0 1 1 0
nematopus 4 2 0 0 0 2 0 1 0 0 0 1 0 0 0 0 - -
panamensis ? 0 1 0 0 1 1 1 0 0 1 1 0 2 0 3 0 0
chetumalensis 1 0 0 1 0 3 0 1 0 1 0 3 0 0 0 3 0 0
cutteri 1 0 0 1 0 3 0 1 0 1 1 3 0 3 0 3 0 0
spilurus 1 0 0 1 0 3 0 1 2 1 1 3 0 0,1 0 3 0 0
altoflavus 1 0 0 0 0 1 1 1 0 0 1 3 1 1 1 2 0 0
nanoluteus 1 0 0 0 0 4 1 1 4 1 1 3 1 1 1 1 0 0
septemfasciatus 1 0 0 0 0 3 1 1 1 0 1 3 0 2 1 3 0 0
myrnae 1 0 0 0 0 3 1 1 0 0 1 3 2 1 1 2 0 0
sajica 1 0 0 0 0 3 1 1 1 1 0 4 0 2 0 - - -
spinosissimus 1 1 1 0 1 1 1 1 1 0 0 3 3 2 0 1 0 0
centrarchus 1 2 1 0 0 6 1 1 0 1 1 4 0 4 0 1 0 1
multispinosus 1 2 1 0 0 1 1 1 0 0 0 1 0 1 0 1 0 0
gemmata 4 2 1 2 1 1 2 0 1 0 0 1 3 2 0 1 1 1
octofasciata 4 2 1 0 1 1 1 0 1 0 0 1 3 0 0 1 1 1
ocotal 4 2 1 0 0 1 0 0 1 0 0 1 3 1 0 1 1 1
kanna 1 2 0 1 0 2 0 1 3 2 0 2 0 1 0 1 0 0
siquia 5 2 0 1 0 2 0 1 3 2 0 2 0 1 0 3 0 0
nigrofasciata 0 2 0 0 0 2 1 1 3 2 0 2 0 0 0 1 0 0
coatepeque 2 2 0 0 0 2 1 1 3 2 0 2 0 ? 0 1 0 0
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO10 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 1. Angle between arms of first epibranchial bone: (a) divergent (Cryptoheros cutteri, UMMZ 173195); (b) par-allel (Cr. chetumalensis, UMMZ 197224). Scales: a, 40X; b, 66X.
3. Number of lower gill rakers on first arch [0] mode 13 or more; [1] mode 8 or less; [2] mode 11; [3] mode 9; [4] mode 10.
High numbers of lower gill rakers (state 0) occurred only in the tilapia and in Ar. centrarchus, a reversal.Fewer lower gill rakers (state 1) are a putative synapomorphy of most Neotropical cichlids examined (exceptfor the autapomorphic modes of Petenia, Amphilophus, and H. nicaraguensis). For this trait, hi=0.20, ri=0.50.
No seriality was proposed for meristic characters because no genetic reason prevents a priori “jumping”from a low to a high number of elements or viceversa without “going through” intermediate numbers ofelements. For coding procedure of meristic characters, see Materials and methods.
This is Kullander’s (1998) character 15 (in part, because he considered distribution as well as number ofgill rakers). His states were: about 8–10, shape normal; 10–60; 4–10, short; 0–2. I agree with him that there isno definite tendency to increase or decrease the number of gill rakers.
4. Total number of gill rakers on first arch [0] mode 16–17; [1] mode 13–14; [2] mode 19 or more; [3] mode 11; [4] mode 10 or fewer.
By “total number” I mean both those on the ceratobranchial and on the epi- and hypobranchials. (Usingthe number of epibranchial gill rakers instead of the total number of gill rakers on the entire first branchialarch would have resulted in just two states, one of them autapomorphic for Ar. centrarchus.)
This character is not quite linked to character 3, because it does not duplicate it: for character 4, state 4occurs in all Neotropical cichlids examined except Petenia and Amphilophus (state 1), and ParachromisAgassiz and Hypsophrys (state 3, synapomorphically). State 2 is autapomorphic for Ar. centrarchus; hi=0.33,ri=0.50.
5. Main gill-raker shape [0] digitiform, blunt (Fig. 2a); [1] trapezoidal or bifid (Fig. 2b); [2] triangular (Fig. 2c); [3] elongated, slender(Fig. 2d); [4] with a mediad projection at base (Fig. 2e).
This character refers to the largest gill rakers on the first ceratobranchial, near the angle with theepibranchial. State 1, where the gill-raker tip has a rather distinct “trapezoidal” end (i.e. with one acute andone obtuse angle) is a synapomorphy of the Neotropical cichlids examined except Petenia and Parachromis,which retain the plesiomorphic digitiform, blunt, shape. There are reversals (Cr. myrnae) and autapomorphies(state 3, Ar. centrarchus), and further transformations: to state 2 in Caquetaia, and to the strictlysynapomorphic state 4 in Rocio. Am. kanna and Am. siquia share a possible state 5, a polymorphism betweenstate 1 and a bifid tip of the gill rakers, paralleled in Amphilophus; hi=0.50, ri=0.50.
This is Kullander’s (1998) character 16. He found “slender, subtriangular” to be the plesiomorphiccondition, with three other states: slender, short, triangular; “fingerlike;” and “enormously elongated.”However, in my data set, not only the tilapia, but also the two following basal clades display my state 0, whichexplains the polarity found here. Moreover, Kullander’s (1998: fig. 11) cladogram shows Old World cichlidspredominantly “fingerlike,” including Heterochromis Regan, which he considered the sister group of a clade
a b
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 11PHYLOGENY OF THE GENUS ARCHOCENTRUS
that includes the Middle American species. Thus, I do not agree with the polarity that Kullander (1998) founda posteriori for this character.
FIGURE 2. Main gill-rakers on first arch: (a) digitiform, blunt, denticulated, with a basal process (Parachromis dovii,UMMZ 188256); (b) trapezoidal or bifid, no basal process (Amphilophus citrinellus, UMMZ 188309); (c) triangular(Amatitlania nigrofasciata, UMMZ 197392); (d) elongated, slender, denticulated (Archocentrus centrarchus, UMMZ224131); (e) with a mediad projection at base (Rocio octofasciata, ECOCH 5468). Scales: a, 40X; b, 32X; c, 40X; d,66X; e, 50X.
6. Basal process on main gill-rakers [0] absent; [1] present (Fig. 2a).
The character refers to the largest gill rakers on the first ceratobranchial. State 1 is an homoplasioussynapomorphy of Amatitlania and of Parachromis, with several parallelisms and a polymorphism inTomocichla Regan; hi=0.88, ri=0.40.
7. Main gill-rakers denticulation [0] no denticulation; [1] denticulation present (Figs. 2a,d).
Homoplasious (hi=0.89, ri=0.22), apparently fixed independently for state 1 in Archocentrus, thesubgenus Cryptoheros, Parachromis, Hypsophrys and elsewhere.
Kullander (1998, character 17) found denticulate gill-rakers to be the plesiomorphic condition. However,according to his cladogram (his fig. 11), not only Old World species have toothless gill-rakers, but also a largeSouth American clade sister to the clade that includes Middle American species does, which supports thepolarity found here.
8. Sutural connection between halves of lower pharyngeal jaw [0] not sutured (Fig. 3a) or very slightly so (Fig. 3b); [1] 3–5 acute interdigitations (Fig. 3c); [2] strongly,digitiformly sutured (Fig. 3d).
a b
c
e
d
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO12 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 3. Connection between halves of lower pharyngeal jaw: (a) not sutured (Archocentrus multispinosus, UMMZ199539); (b) very slightly sutured (Rocio octofasciata, UMMZ 176671); (c) 3-5 acute interdigitations (Cryptoherossajica, UMMZ 194239); (d) strongly, digitiformly sutured (Amphilophus citrinellus, UMMZ 188309). Scales: a, 32X; b,16X; c, 40X; d, 8X.
State 1 is a synapomorphy of Cryptoheros. It corresponds to Kullander’s (1998, character 20) “partlysutured.” State 2 is autapomorphic for A. citrinellus; hi=0.00, ri=1.00.
Kullander believed that a strong interdigitation between the two halves of the pharyngeal jaw was relatedto the presence of “hard items in the diet.” I find the same polarity.
9. Rows of teeth on lower pharyngeal jaw [0] 25 rows wide (one half of lower pharyngeal jaw), 18 rows long; [1] 9 rows wide, 11 rows long; [2] 9–14rows wide, 5–9 rows long; [3] 19 rows wide, 11 rows long; [4] 16 rows wide, 9 rows long.
b
c
a
d
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 13PHYLOGENY OF THE GENUS ARCHOCENTRUS
State 1 is autapomorphic for Petenia. State 2 is common to the rest of the Neotropical cichlids examined,with autapomorphic changes to state 3 in H. nicaraguensis and state 4 in Amphilophus.
10. Main tooth type on upper pharyngeal jaw [0] bicuspidate (Fig. 4a); [1] tricuspidate (Fig. 4b).
State 1 appears in several places in the cladogram; it is fixed in Cryptoheros (although polymorphic insome species). The bicuspidate condition might appear to be due to grinding of the two proximal cusps, whichare close together and not very pronounced; however, in polymorphic taxa the trait seems not to be associatedwith age. Large amount of homoplasy, hi=0.86, but informative to some degree, ri=0.56.
FIGURE 4. Main tooth-type on upper pharyngeal jaw: (a) bicuspidate (Rocio gemmata, ECOCH 1468); (b) tricuspidate(Cryptoheros myrnae, UMMZ 217739). Scale: 66X.
11. Width vs. length, upper pharyngeal jaw[0] narrower; [1] subequal or wider.
“Width” was measured from tip to tip across both halves of the jaw. State 1 is present in all Neotropicalcichlids examined, except Petenia; hi=0.50, but character uninformative, ri=0.00.
Hyoid series, character 1212. Posterior and dorsal edges of urohyal [0] posterior edge concave, dorsal edge with a spine (Fig. 5a); [1] concave, with a posterior spine, dorsallyconvex; [2] posteriorly sigmoid, with a slightly protruding dorsal convexity (Fig. 5b); [3] sigmoid; [4]dorsally sigmoid, posteriorly straight-slanted; [5] irregular (Fig. 5c); [6] concave, simple (no spines orconvexities) (Fig. 5d).
State 1 might support Burgess’ (2000, based on Miller in litt., 1996) view of a relationship betweenCaquetaia and Petenia; the state appears in H. nicaraguensis, too. Although it is here interpreted as anhomoplasy, for the sake of testing that hypothesis the character was not deleted. State 2 is a strictsynapomorphy of Parachromis. State 3 occurs in several clades, synapomorphically in Am. kanna + siquia.State 4 is an autapomorphy of A. citrinellus. State 5 is most characteristic in Cryptoheros and Archocentrus;state 6 is a synapomorphy of Rocio, but occurs also in Amatitlania and Cryptoheros; hi=0.50, ri=0.54.
a b
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO14 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 5. Posterior and dorsal edges of urohyal: (a) posterior edge concave, dorsal edge with a spine (Tomocichla sie-boldii, UMMZ 194240); (b) posteriorly sigmoid, with a slight protruding dorsal convexity (Parachromis loisellei,UMMZ 203897); (c) irregular (Cryptoheros cutteri, UMMZ 199678); (d) concave, simple, no spines or convexities(Rocio gemmata, ECOCH 1468). Scale: a, 32X; b, 8X; c, 25X; d, 20X.
FIGURE 6. Posterior edge of mesethmoid: (a) with indentation (Amphilophus citrinellus, UMMZ 188309); (b) rounded(Amatitlania kanna, UMMZ 145716); (c) nematopus-type (Hypsophrys nematopus, UMMZ 197507); (d) straight-irregu-lar (Amatitlania siquia, UMMZ 196948); (e) with an indentation and a posteriad spine (Rocio gemmata, ECOCH 1468).Scales: a, 12X; b,d, 50X; c,e, 32X.
a b
c
d
a
e
b
d
c
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 15PHYLOGENY OF THE GENUS ARCHOCENTRUS
FIGURE 7. Occipital region: (a) caudal edge of supraoccipital crest straight, vertical, dorsal tip angled, predorsal withrostrad process, first neural spine antrorse, three dorsal elements between first two first neural spines; anterodorsal spineon first dorsal pterygiophore (Amatitlania siquia, UMMZ 196948); (b) caudal edge straight, inclined rostrad, dorsal tiprounded (Hypsophrys nematopus, UMMZ 197507); (c) caudal edge undulating, no deep concavity, first neural spine ret-rorse, two dorsal elements between first two first neural spines (Cryptoheros cutteri, UMMZ 173195); (d) caudal edgeconvex, first neural spine retrorse (Archocentrus centrarchus, UMMZ 224131); (e) undulating, with a deep concavity,first neural spine retrorse, three dorsal elements between first two first neural spines (Rocio gemmata, ECOCH 1468).Scales: a,d,e, 16X; b, 20X; c, 12X.
e
d
c
ab
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO16 · Zootaxa 1618 © 2007 Magnolia Press
Neurocranium, characters 13–1613. Mesethmoid posterior edge [0] straight-concave, covered by lateral ethmoid; [1] with a double indentation (Fig. 6a); [2] rounded (Fig.6b); [3] nematopus-type (Fig. 6c); [4] straight-irregular (Fig. 6d); [5] with an indentation and a posteriad spine(Fig. 6e).
State 1 is an homoplasious synapomorphy for Parachromis; state 2 occurs in several clades, such asArchocentrus and most of Amatitlania and Cryptoheros. State 3 is autapomorphic for H. nematopus. State 4 iscommon in the subgenus Bussingius; state 5 is a strict synapomorphy for Rocio; hi=0.58, ri=0.30.
14. Supraoccipital crest, caudal edge [0] irregularly undulating; [1] straight, vertical, dorsal tip angled (Fig. 7a); [2] concave; [3] straight, inclinedrostrad, dorsal tip rounded (Fig. 7b); [4] undulating, no deep concavity (Fig. 7c); [5] convex (Fig. 7d); [6]undulating, with a deep concavity (Fig. 7e).
States 1 and 2 are homoplasious, although state 1 is fixed in Amatitlania. State 3 is autapomorphic for H.nematopus. State 4 may be synapomorphic for Cryptoheros, but reversed several times. State 5 might be asynapomorphy of Ar. centrarchus + multifasciatus, with a parallelism in Caquetaia. State 6 is a strictsynapomorphy of Rocio; hi=0.54, ri=0.42.
FIGURE 8. Coronal pore triple, tubes T-shaped (Cryptoheros cutteri, UMMZ 188136). Semidiagrammatic. Scale: 6X.
15. Coronal pore [0] single; [1] none; [2] triple, tubes T-shaped (Fig. 8).
Cichocki (1976, character 3) defined this character in a more complex manner, as the “configuration ofthe coronal frontals lateralis canals and associated frontal ridges” and established eleven character states.However, my coding coincides with Cichocki’s only in distinguishing Petenia, with a coronal pore“obliterated by development of a median trough running the full length of the skull roof” (my state 1: no
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 17PHYLOGENY OF THE GENUS ARCHOCENTRUS
coronal pore). He combined my states 0 and 2 as “united medially, raised on medial frontal ridge as commonanterodorsally directed coronal tube with single coronal pore.” However, I find that state 2, a triple coronalpore, with the three pores joined by tubes shaped like a T, is synapomorphic for Cryptoheros, with a reversalin Cr. myrnae, a polymorphism in Cr. septemfasciatus (states 0 and 2), and a parallelism in Ar. multispinosus(hi=0.50, ri=0.71).
On the other hand, Cichocki (1976) coded a Parachromis, P. friedrichsthalii (Heckel), as having adifferent character state, “coronal canal directed straight dorsally and moderately elevated in the medianfrontal crest…,” with a figure (his fig. 1.6 D) depicting a single coronal pore. As I construct the character, Ifail to see a difference in either P. loisellei (Bussing, 1989) or P. dovii, both of them being single-pored, state0.
16. Supraoccipital crest height on postethmoid skull roof length [0] 0.33 or more; [1] 0.27 or less.
State 1 is autapomorphic for H. nematopus. Cichocki (1976, character 2) recognised five character states, with not quite the same polarity found here:
0.26–0.50, 0.18–0.20, 0.10 or less, 0.52–0.55, and 0.80 or more; within his state 1 (which he applies to H.nematopus and the other taxa which occur in both studies), I find two different states, separated by a well-defined gap.
Infraorbitals, characters 17–1917. Sensory canals on lachrymal [0] five marginal-directed canals; [1] four marginal-directed canals.
State 1 is a synapomorphy of all Neotropical cichlids examined, hi=0.00.This character and its states are in part Cichocki’s (1976, character 45) “configuration of the infraorbital
ossicles associated with the anterior four neuromasts,” with eight states, and Kullander’s (1998, character 38)“configuration of the lateralis canal in the lachrymal,” which in addition to the number of openings (marginal-directed canals) considered the number of lachrymal bones, 1 or 2. All the species considered here have justone lachrymal. I find the same polarity as Kullander (1998).
I refrained from using the shape of the lachrymal (Kullander’s [1998] character 40, Cichocki’s [1976]character 46) because it is strongly affected by ontogeny (juveniles of all species tend to have widerlachrymals). It is also a very homoplasious character: a wide lachrymal (in adults) is parallel betweenCaquetaia, Petenia, Amphilophus, and Ar. centrarchus, and a deep one is convergent between H. nematopusand Tomocichla.
I would have coded this second part of the character (shape of the lachrymal) as Cichocki (1976) did. Herecognised that there are two trends in its evolution: one towards wider, the other one towards deeperlachrymals. Kullander (1998), however, noted that heroines (that is, all my study taxa except the tilapia) andcichlasomines have all an “approximately square form.”
18. Overlap between lachrymal and second infraorbital [0] some overlap; [1] no overlap.
An autapomorphy of Petenia. Cichocki’s (1976) character 47 and its states are the same; however,Kullander’s (1998, character 44) distinguished two degrees of overlap (distal connection vs. extensiveoverlap) and found the opposite polarity, although both the Malagasy-Indian Etroplus Cuvier+Ptychochromis Steindachner and Heterochromis, sister group of the clade that includes Middle Americancichlids (Kullander 1998), show his state 2, which supports my view (and Cichocki’s, 1976) that the overlaprepresents the plesiomorphic condition.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO18 · Zootaxa 1618 © 2007 Magnolia Press
19. Number of infraorbitals posterior to lachrymal [0] five; [1] four.
State 1 is synapomorphic for all Neotropical cichlids examined, with the sole autapomorphic reversal ofH. nicaraguensis; hi=0.50, but ri=0.00.
Same interpretation as Kullander’s (1998) character 41, although he needed two more states, for specieswith three or two infraorbitals.
Mouth and oral jaws, characters 20–4720. Anterior vs. posterior palatine arms [0] anterior longer (Fig. 9a); [1] posterior longer; [2] subequal (Fig. 9b).
State 1 seems to be a convergent adaptation (hi=0.75) for the piscivore habit, occurring in Parachromis,Caquetaia, and Petenia. State 2 is synapomorphic (ri=0.57) for Amatitlania, Rocio, and Archocentrus,although reversed in H. nematopus and Ar. multispinosus and parallel in Cr. sajica and Amphilophus.
21. Ascending premaxillary process on orbit [0] process not reaching eye rim; [1] process to anterior eye rim; [2] process to mid-eye; [3] process beyondposterior eye rim.
The elongation of the dorsal process (ascending arm) of the premaxillary bone is an adaptation to makethe mouth protrusible. State 1 is synapomorphic for Rocio and Amatitlania; state 2 appears to have evolvedseparately in Caquetaia and Parachromis. State 3 is autapomorphic for Petenia. State 0, once used todiagnose Archocentrus (e.g. Allgayer 2001), is just a symplesiomorphy of these small-mouthed Heroini;hi=0.50, ri=0.67.
FIGURE 9. Anterior vs. posterior palatine arms: (a) anterior longer (Cryptoheros chetumalensis, UMMZ 197224); (b)subequal (Archocentrus centrarchus, UMMZ 224131). Scales: a, 32X; b, 40X.
22. Ascending premaxillary process on skull length [0] ratio 0.20–0.66; [1] ca. 0.76; [2] 0.87 or more.
States 1 and 2 are autapomorphic for Petenia and Caquetaia, respectively. An obvious coding alternativewould be to combine them into a putative synapomorphy, which would support the view of a closerelationship between those taxa; however, the difference between them is as large as, or larger, than the onebetween Petenia and the rest of the examined species.
Cichocki (1976, character 12) recognised 10 character states for this trait, based on several combinationsof the ascending premaxillary process relative to the prefrontal and standard skull lengths. He conferredCaquetaia kraussi Steindachner and Petenia the same state, “ascending process extremely long,” both withrespect to prefrontal skull and standard skull lengths. H. nematopus and Amatitlania also had one same state,“ascending process moderate with respect to prefrontal skull, long with respect to standard skull length,” butAmphilophus had a different state, “ascending process long with respect to prefrontal skull, long with respectto standard skull length.”
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 19PHYLOGENY OF THE GENUS ARCHOCENTRUS
23. Ascending vs. dentigerous arms of premaxilla [0] ascending arm shorter; [1] ascending arm longer (ascending / dentigerous = 1.18–2.03); [2] ascending armmuch longer (ca. 2.37); [3] ascending arm longer still (2.44–2.47).
State 1 is a synapomorphy for the Neotropical cichlids examined, apart of Petenia (and the autapomorphicreversal in H. nematopus); hi=0.25, but ri=0.00. A much longer dorsal process hints at a relationship betweenPetenia and Caquetaia, but the still longer ascending arm of Caquetaia is interpreted here as a differentcharacter state. Kullander (1998, character 64) followed Casciotta and Arratia (1993) in uniting my states 2and 3, but the polarity that he found is the same as mine.
I was not able to code Cichocki’s (1976) character 14, the length of the articulating process relative to thedorsal process. He distinguished two states, 0.40–0.69 and 0.70–0.90 (ratio articulating / ascending), but theyoverlap in my material, not allowing a statistical definition of states. Also Kullander (1998) did not considerthe character to be “general” enough to be useful.
24. Dorsal convexity on dentigerous arm of premaxilla [0] not distinct (Fig. 10a); [1] distinct (Fig. 10b).
Quite homoplasious (hi=0.89), but state 1 appears to be synapomorphic for Cryptoheros (ri=0.20), with areversal for the five species in the subgenus Bussingius, and also for Ar. centrarchus + multispinosus withinArchocentrus, with parallelisms elsewhere over the tree.
25. Narrowest point of dentigerous arm of premaxilla [0] at caudal tip (Fig. 10c); [1] before caudal tip (Figs. 10a,b).
State 1 is synapomorphic for the Neotropical cichlids examined, although with several reversals, mostimportantly in Rocio; hi=0.75, ri=0.50.
FIGURE 10. Dentigerous arm of premaxilla: (a) dorsal convexity not distinct, arm narrowest before caudal tip(Amatitlania coatepeque, UMMZ 181823); (b) dorsal convexity distinct, arm narrowest before caudal tip (Am. siquia,UMMZ 196948); (c) arm narrowest at caudal tip (Rocio octofasciata, ECOCH 5468). Scales: a,b, 40X; c, 25X.
missing among submitted files
ba
c
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO20 · Zootaxa 1618 © 2007 Magnolia Press
26. Toothed length in premaxilla [0] ca. 0.97 total length of dentigerous arm; [1] ca. 0.84; [2] 0.30–0.80.
State 2 is synapomorphic for the Neotropical cichlids examined; state 1 is autapomorphic for R. gemmata;hi=0.00.
Cichocki (1976, character 18) called this character “outer premaxillary tooth distribution,” with the samepolarity as here, but states defined as 0.75–1.00, 0.50–0.67, and 0.25 or less, thus not only not quite the sameas mine, but also somewhat conflicting (apparently he did not observe specimens with toothed length between0.26 and 0.50). I defined states by looking for non-overlapping value distributions.
27. Width vs. length, posterior premaxillary expansion [0] 0.21–0.40; [1] ca. 0.18; [2] 0.41 or more.
State 1 is another possible synapomorphy (here considered a convergence) for Petenia and Caquetaia;state 2 is autapomorphic for H. nematopus; hi=0.33, but ri=0.00.
Cichocki (1976, character 27) recognised four states: 0.20–0.33, 0.34–0.36, 0.38–0.43, and 0.14–0.17,delimited by breaks in the range of variation; his polarity is the same as mine. The species here examinedshow too extensive an overlap within the range 0.21–0.40 to warrant recognition of Cichocki’s first threestates.
28. Maxillary on orbit [0] maxillary reaches only to horizontal level of orbit; [1] maxillary reaches orbit both horizontally andvertically; [2] maxillary falls short of orbit.
Homoplasious (hi=0.71); fixed for state 1 in Parachromis and for state 2 in Hypsophrys, the subgenusBussingius of Cryptoheros, etc. (ri=0.50).
FIGURE 11. Maxillary: (a) rostral end convex, no notch, caudal end entire (Rocio octofasciata, UMMZ 187779); (b)rostral end notched or at least concave, dorsal edge not serrated (R. gemmata, ECOCH 1468); (c) no notch at rostral end,but a dorsad protuberance (Hypsophrys nematopus, UMMZ 197507); (d) dorsal edge serrated (Cryptoheros altoflavus,UMMZ 145722); (e) caudal end with a concavity (Archocentrus multispinosus, UMMZ 199539). Scales: a, 16X; b, 25X;c, 32X; d,e, 40X.
a b
c
d
e
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 21PHYLOGENY OF THE GENUS ARCHOCENTRUS
29. Rostral end of maxillary [0] convex, no notch (Fig. 11a); [1] notched or at least concave (Fig. 11b); [2] no notch, but a dorsad spine(Fig. 11c).
Homoplasious (hi=0.78) but somewhat informative (ri=0.42). State 1 is fixed in Cryptoheros (reversal inCr. chetumalensis); state 2 fixed in Parachromis.
30. Dorsal edge of maxillary [0] not serrated (Fig. 11a,b); [1] serrated (Fig. 11d).
State 1 is fixed in Amatitlania and Cryptoheros, parallel in Tomocichla; hi=0.67, but a high retentionindex, ri=0.85.
31. Posterior end of maxillary [0] entire (Figs. 11a–d); [1] with a concavity (Fig. 11e).
State 1 appears in Ar. spinosissimus and Ar. multispinosus, either as a parallelism or as a synapomorphyfor Archocentrus with a reversal in Ar. centrarchus; hi=0.50, but ri=0.00.
FIGURE 12. Posterior end of dentigerous arm of dentary: (a) rounded or squarish (Amatitlania nigrofasciata, UMMZ188245); (b) triple-spined (Am. coatepeque, UMMZ 181823); (c) bluntly pointed (Am. siquia, UMMZ 196948). Scales:a, 40X; b, 32X; c, 50X.
32. Arms of dentary [0] subequal, or dentigerous longer than ventral (dentigerous / ventral = 0.93–1.62); [1] ventral longer thandentigerous (dentigerous / ventral, ca. 0.88) (Fig. 12).
State 1 is synapomorphic for the Neotropical cichlids examined, with a reversal in Parachromis (hi=0.50,ri=0.50). Among the rest, only H. nematopus and Tomocichla occasionally have a dentigerous / ventral lengthratio slightly less than 1.
a b
c
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO22 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 13. Articular and retroarticular: (a) dorsal edge straight, ventral angle right, no rostrad pronounced convexityon ventral process (Amatitlania nigrofasciata, UMMZ 188245; retroarticular not shown); (b) ventral angle acute (Rociooctofasciata, UMMZ 187779); (c) dorsal edge convex, ventral angle obtuse (Cryptoheros altoflavus, UMMZ 145722);(d) dorsal edge straight, ventral angle obtuse (Am. siquia, UMMZ 190199; retroarticular not shown); (e) arms of articularequal, no posteriad projection on retroarticular (Hypsophrys nematopus, UMMZ 197507); (f) dorsal edge straight,ventral angle right, a rostrad pronounced convexity on ventral process (R. gemmata, ECOCH 1468); (g) retroarticularmuch elongated anterocaudally (Petenia splendida, UMMZ 196461). Scales: a,c. 40X; b,e, 20X; d, 50X; f, 32X; g, 10X.
a b
f
e
c
d
g
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 23PHYLOGENY OF THE GENUS ARCHOCENTRUS
33. Posterior end of dentigerous arm of dentary [0] tapering; [1] sharply pointed; [2] rounded or squarish (Fig. 12a); [3] triple-spined (Fig. 12b); [4] bluntlypointed (Fig. 12c).
State 1 is a convergence of Caquetaia and Petenia (see Cichocki 1976: fig. 1.9E). State 2 seemssynapomorphic for the Neotropical cichlids examined, with an autapomorphic progression to state 3 in Am.coatepeque and to state 4 in Am. kanna + siquia and Cryptoheros (some reversals to state 2); hi=0.56, ri=0.50.
34. Arms of articular [0] rostral clearly longer than dorsal (dorsal / rostral = 0.41–0.77) (Figs. 13a–d); [1] dorsal / rostral, ca. 0.87;[2] rostral and dorsal equal (Fig. 13e).
State 1 is autapomorphic for Ar. multispinosus. State 2 is autapomorphic for H. nematopus.
35. Dorsal edge of articular [0] straight (Figs. 13a,d); [1] convex (Fig. 13c).
State 1 is a strict synapomorphy of Cryptoheros (without the basal Cr. panamensis); hi=0.00, ri=1.00.
36. Ventral angle of articular [0] acute (Fig. 13b); [1] right (Fig. 13a); [2] obtuse (Fig. 13d).
State 1 occurs in most Neotropical cichlids examined; state 2 is most common within Cryptoheros, and issynapomorphic for the clade Am. kanna + siquia; hi=0.82, ri=0.31.
37. Anteriorly directed pronounced convexity on ventral process of articular [0] absent (Figs. 13a,c,d); [1] present (Fig. 13f).
State 1 occurs in most Neotropical cichlids examined, with many reversals, notably in most Cryptoheros
and Parachromis. Fixed for Rocio, Archocentrus, and Hypsophrys; hi=0.88, ri=0.22.
38. Shape of retroarticular [0] slightly elongated (Figs. 13a–f); [1] much elongated anterocaudally (Fig. 13g).
State 1 is probably a convergence of piscivores: Petenia, Caquetaia, and Parachromis (hi=0.67, ri=0.33).The same is the case for many of these mouth and oral jaw characters; this is the reason why Kullander (1998)shunned most of Casciotta and Arratia’s (1993) characters, whose emphasis on trophic-related traits led tolumping in one clade all piscivores in their matrix. I feel, nevertheless, that the convergence hypothesis,however plausible, should be tested by inclusion of these characters in a larger matrix.
39. Posteriad projection on ventroposterior angle of retroarticular [0] absent (Fig. 13g); [1] present (Figs. 13b–d,f).
State 1 is homoplastically synapomorphic for Cryptoheros (with reversal in Cr. sajica), Rocio, andAmatitlania (hi=0.86, but ri=0.64). Cr. myrnae and Am. siquia are polymorphic for the trait.
40. Quadrate [0] no indentation on posterior edge, longer than wide (Fig. 14a); [1] no indentation on posterior edge, widerthan long (Fig. 14b); [2] a dorsoposterior concavity (Fig. 14c); [3] shaped like a quarter-circle (Fig. 14d); [4]with a spine (Fig. 14e).
State 1 is a synapomorphy of Cryptoheros, although occurring homoplastically also in Petenia, H.nematopus, and Am. kanna. State 2 is a synapomorphy of Archocentrus, albeit it is present also in Caquetaiaand Amphilophus. State 3 is common in Amatitlania and Rocio; state 4 is an autapomorphy of R. gemmata;hi=0.56, but ri=0.58.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO24 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 14. Quadrate: (a) no indentation on posterior edge, bone longer than wide (Oreochromis mossambicus, UMMZ199401); (b) no indentation on posterior edge, bone wider than long (Petenia splendida, UMMZ 196461); (c) adorsoposterior concavity (Amphilophus citrinellus, UMMZ 188309); (d) shaped like a quarter-circle (Amatitlanianigrofasciata, UMMZ 202707); (e) with a spine on rostral margin (Rocio gemmata, ECOCH 1468). Scales: a,b, 16X; c,10X; d, 32X; e, 25X.
a b
cd
e
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
Zootaxa 1618 © 2007 Magnolia Press · 25PHYLOGENY OF THE GENUS ARCHOCENTRUS
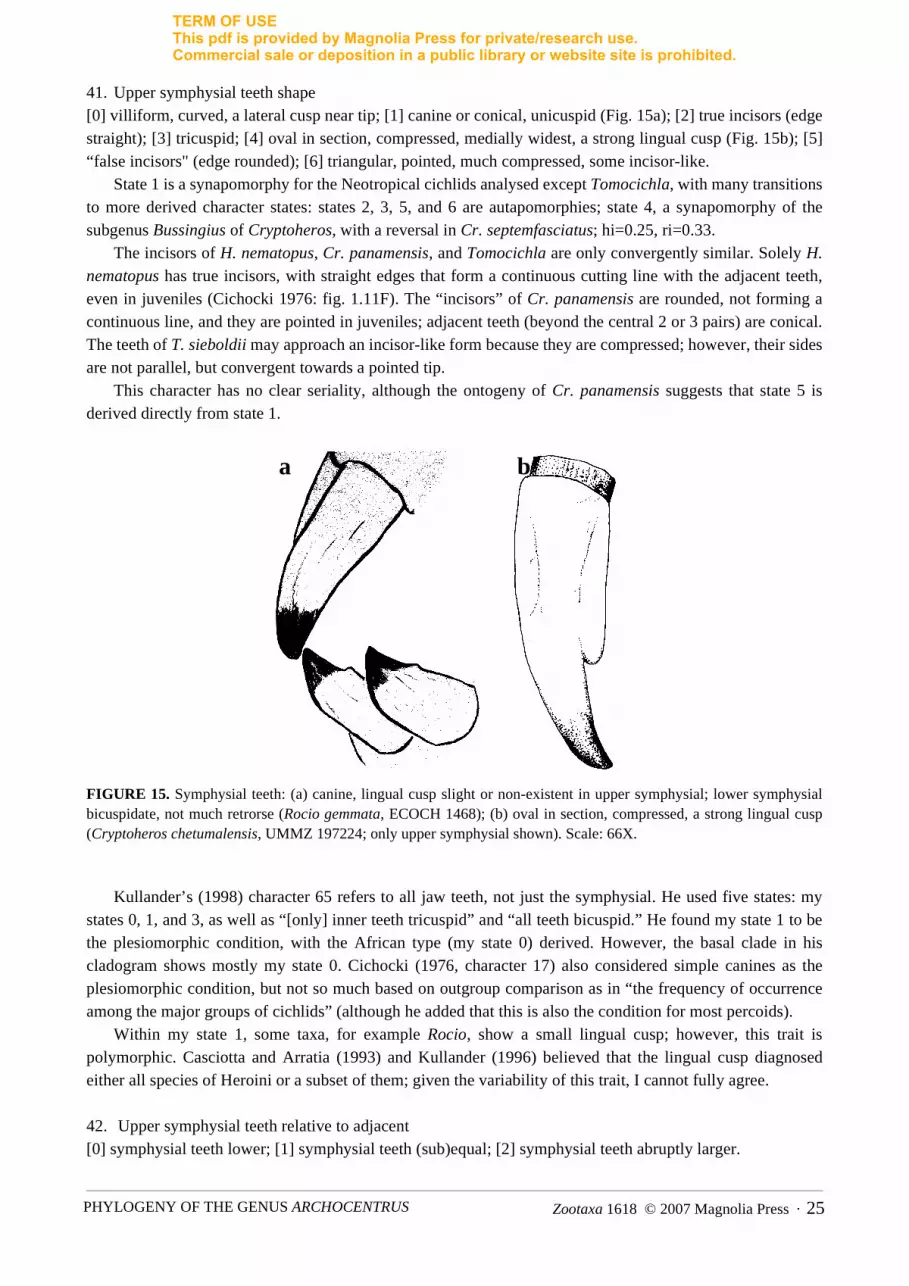
41. Upper symphysial teeth shape [0] villiform, curved, a lateral cusp near tip; [1] canine or conical, unicuspid (Fig. 15a); [2] true incisors (edgestraight); [3] tricuspid; [4] oval in section, compressed, medially widest, a strong lingual cusp (Fig. 15b); [5]“false incisors" (edge rounded); [6] triangular, pointed, much compressed, some incisor-like.
State 1 is a synapomorphy for the Neotropical cichlids analysed except Tomocichla, with many transitionsto more derived character states: states 2, 3, 5, and 6 are autapomorphies; state 4, a synapomorphy of thesubgenus Bussingius of Cryptoheros, with a reversal in Cr. septemfasciatus; hi=0.25, ri=0.33.
The incisors of H. nematopus, Cr. panamensis, and Tomocichla are only convergently similar. Solely H.nematopus has true incisors, with straight edges that form a continuous cutting line with the adjacent teeth,even in juveniles (Cichocki 1976: fig. 1.11F). The “incisors” of Cr. panamensis are rounded, not forming acontinuous line, and they are pointed in juveniles; adjacent teeth (beyond the central 2 or 3 pairs) are conical.The teeth of T. sieboldii may approach an incisor-like form because they are compressed; however, their sidesare not parallel, but convergent towards a pointed tip.
This character has no clear seriality, although the ontogeny of Cr. panamensis suggests that state 5 isderived directly from state 1.
FIGURE 15. Symphysial teeth: (a) canine, lingual cusp slight or non-existent in upper symphysial; lower symphysialbicuspidate, not much retrorse (Rocio gemmata, ECOCH 1468); (b) oval in section, compressed, a strong lingual cusp(Cryptoheros chetumalensis, UMMZ 197224; only upper symphysial shown). Scale: 66X.
Kullander’s (1998) character 65 refers to all jaw teeth, not just the symphysial. He used five states: mystates 0, 1, and 3, as well as “[only] inner teeth tricuspid” and “all teeth bicuspid.” He found my state 1 to bethe plesiomorphic condition, with the African type (my state 0) derived. However, the basal clade in hiscladogram shows mostly my state 0. Cichocki (1976, character 17) also considered simple canines as theplesiomorphic condition, but not so much based on outgroup comparison as in “the frequency of occurrenceamong the major groups of cichlids” (although he added that this is also the condition for most percoids).
Within my state 1, some taxa, for example Rocio, show a small lingual cusp; however, this trait ispolymorphic. Casciotta and Arratia (1993) and Kullander (1996) believed that the lingual cusp diagnosedeither all species of Heroini or a subset of them; given the variability of this trait, I cannot fully agree.
42. Upper symphysial teeth relative to adjacent [0] symphysial teeth lower; [1] symphysial teeth (sub)equal; [2] symphysial teeth abruptly larger.
a b
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO26 · Zootaxa 1618 © 2007 Magnolia Press
State 0 occurs only in the tilapia. State 2 is homoplastically synapomorphic for Parachromis, Rocio, andAmatitlania, with parallelisms in Cr. myrnae and Caquetaia; hi=0.67, but ri=0.60.
43. Lower symphysial teeth shape[0] multicuspidate; [1] canine retrorse; [2] oval in section, medially widest; [3] conical, not much retrorse; [4]tricuspidate; [5] bicuspidate (Fig. 15a).
State 2 is autapomorphic for Tomocichla. State 3 is most common within the Neotropical cichlidsexamined, fixed in Amatitlania, Hypsophrys, and Parachromis. State 4 is an autapomorphy for Ar.multispinosus, once diagnostic for the monotypic genus Herotilapia. State 5 appears in Rocio (without R.ocotal) and Cryptoheros; hi=0.50, ri=0.38.
44. Lower symphysial teeth relative to adjacent [0] symphysial teeth (sub)equal; [1] symphysial teeth lower; [2] symphysial teeth much lower, almostcompletely embedded.
State 1 is homoplastically synapomorphic for Rocio and Amatitlania, and occurs also in Cr. panamensisand Petenia. State 2 is synapomorphic for Parachromis, convergent with Caquetaia; hi=0.67, but ri=0.60.
45. Number of lower mandibular external teeth [0] modally less than 20; [1] modally more than 24.
State 1 is autapomorphic for Petenia.
46. Protruding lower jaw [0] yes (prognathous), at least slightly; [1] no (mouth terminal); [2] lower jaw receding (mouth inferior).
State 1 is homoplastically synapomorphic for Amatitlania and for Cryptoheros, and parallel also in Ar.multispinosus. State 2 is a synapomorphy of Hypsophrys, convergent in Tomocichla; hi=0.60, but a high ri,0.75.
47. Frenum [0] present (Fig. 16); [1] absent.
Once considered diagnostic of genera (Heros Heckel vs. Cichlasoma, according to Jordan & Evermann1898), this character (a medial interruption of the lower lip fold) has long been considered very variableamong species (e.g. Pellegrin 1904). Nevertheless, state 1 is fixed in Parachromis; hi=0.80, ri=0.20.
FIGURE 16. Frenum and dentary pores (Rocio ocotal, UMMZ 171140). Note fifth dentary pore on right dentary. Scale:16X.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 27PHYLOGENY OF THE GENUS ARCHOCENTRUS
Preopercle, opercle, and pectoral girdle, characters 48–5148. Preopercular posterior edge [0] convex to straight; [1] slightly, roundly concave (Fig. 17); [2] irregular, no lobes; [3] squarely concave(“bracket-shaped”); [4] irregular, with lobes.
State 1 occurs in all Neotropical cichlids examined outside Tomocichla and Petenia, with reversals withinCryptoheros. States 2–4 are autapomorphies; hi=0.50, but ri=0.00.
FIGURE 17. Posterior edge of preopercle: slightly, roundly concave (Amatitlania coatepeque, UMMZ 181823). Scale:10X.
49. Spine on dorsal margin of opercle [0] absent (Fig. 18a); [1] present (Fig. 18b).
Highly homoplasious (hi=0.90). Some species are polymorphic for this trait; however, it supports somestructure (e.g., it is a synapomorphy of the subgenus Cryptoheros, with a polymorphism in Cr. cutteri);ri=0.14.
50. Shape of dorsal margin of opercle [0] evenly convex, no protuberances (Fig. 18a); [1] evenly convex, with a dorsoanterior protuberance (Fig.18c); [2] extremely convex, almost semicircular (Fig. 18d); [3] almost straight.
State 1 is a putative synapomorphy of Ar. centrarchus + multispinosus (but ri=0.00), with a parallelism inP. loisellei. State 2 occurs in H. nematopus and Caquetaia. State 3 is autapomorphic for Petenia; hi=0.50.
51. Foramen in scapula [0] slanted (Fig. 19a); [1] parallel to anteroposterior axis of bone (Fig. 19b).
State 1 is a synapomorphy of all Neotropical cichlids examined; hi=0.00.
Axial skeleton, characters 52–5952. Rostrad process on first predorsal element [0] absent; [1] present (Fig. 7).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.
SCHMITTER-SOTO28 · Zootaxa 1618 © 2007 Magnolia Press
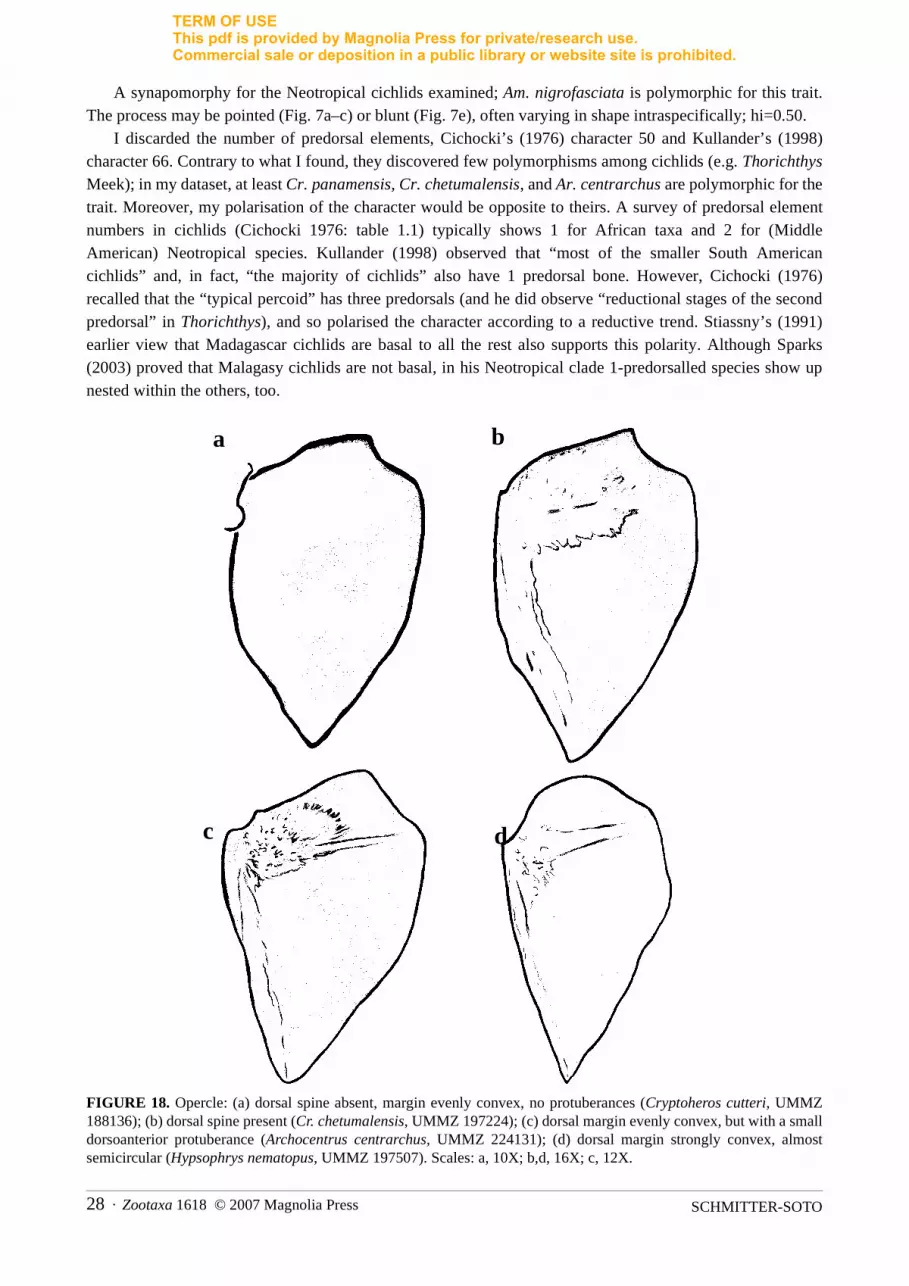
A synapomorphy for the Neotropical cichlids examined; Am. nigrofasciata is polymorphic for this trait.The process may be pointed (Fig. 7a–c) or blunt (Fig. 7e), often varying in shape intraspecifically; hi=0.50.
I discarded the number of predorsal elements, Cichocki’s (1976) character 50 and Kullander’s (1998)character 66. Contrary to what I found, they discovered few polymorphisms among cichlids (e.g. ThorichthysMeek); in my dataset, at least Cr. panamensis, Cr. chetumalensis, and Ar. centrarchus are polymorphic for thetrait. Moreover, my polarisation of the character would be opposite to theirs. A survey of predorsal elementnumbers in cichlids (Cichocki 1976: table 1.1) typically shows 1 for African taxa and 2 for (MiddleAmerican) Neotropical species. Kullander (1998) observed that “most of the smaller South Americancichlids” and, in fact, “the majority of cichlids” also have 1 predorsal bone. However, Cichocki (1976)recalled that the “typical percoid” has three predorsals (and he did observe “reductional stages of the secondpredorsal” in Thorichthys), and so polarised the character according to a reductive trend. Stiassny’s (1991)earlier view that Madagascar cichlids are basal to all the rest also supports this polarity. Although Sparks(2003) proved that Malagasy cichlids are not basal, in his Neotropical clade 1-predorsalled species show upnested within the others, too.
FIGURE 18. Opercle: (a) dorsal spine absent, margin evenly convex, no protuberances (Cryptoheros cutteri, UMMZ188136); (b) dorsal spine present (Cr. chetumalensis, UMMZ 197224); (c) dorsal margin evenly convex, but with a smalldorsoanterior protuberance (Archocentrus centrarchus, UMMZ 224131); (d) dorsal margin strongly convex, almostsemicircular (Hypsophrys nematopus, UMMZ 197507). Scales: a, 10X; b,d, 16X; c, 12X.
a b
dc
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 29PHYLOGENY OF THE GENUS ARCHOCENTRUS
FIGURE 19. Foramen in scapula: (a) slanted (Oreochromis mossambicus, UMMZ 199401); (b) parallel toanteroposterior axis of bone (Amphilophus citrinellus, UMMZ 188309). Scales: a, 25X; b, 8X.
53. Orientation of first neural spine [0] retrorse (Fig. 7c,d); [1] antrorse (Fig. 7a,b).
Very homoplasious (hi=0.88), sometimes polymorphic, but nevertheless informative (ri=0.50). State 1 issynapomorphic in Hypsophrys and in Cryptoheros (without the basal Cr. panamensis, and with a reversal inthe clade Cr. spilurus + cutteri); it is also fixed in Amatitlania (polymorphic in Am. coatepeque).
54. Number of dorsal elements between first two epineural spines [0] two (Fig. 7c); [1] three (Fig. 7a,e).
By “dorsal elements” I mean both the predorsal bones and the anteriormost pterygiophores of the dorsalfin. State 1 is fixed in Amatitlania, Rocio, and in Cryptoheros without the basal Cr. panamensis, and with areversal in the clade C. spilurus+ cutteri, but it homoplastically occurs in many other species (hi=0.86,ri=0.38). It is polymorphic in R. octofasciata.
55. Spinous anterodorsal process on first dorsal pterygiophore [0] absent; [1] present (Fig. 7a).
Cichocki (1976, character 51) found the derived state to be rare among Cichlidae. I find a minuteanterodorsal spine to be an homoplasious synapomorphy of Am. kanna + siquia, parallel with Cr.chetumalensis; hi=0.50, ri=0.50.
56. Number of total vertebrae [0] modally fewer than 31; [1] modally 31 or more.
State 1 occurs convergently in Petenia, Tomocichla, and P. dovii; hi=0.67, ri=0.00. Cichocki (1976,character 57) coded for more character states based on “breaks in the frequency distribution,” but he assumedthe range 26–30 to be primitive based on his extensive outgroup survey, a polarity that I concur with. On theother hand, he coded H. nematopus as having 31–34 vertebrae, but all specimens I have seen show 30vertebrae.
Kullander (1998, character 67) also coded for more states, considering not just the total number but alsothe number of abdominal vertebrae, absolute and relative to caudal vertebrae. He found “abdominal vertebrae
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO30 · Zootaxa 1618 © 2007 Magnolia Press
14–15, the number of caudal vertebrae equal or higher” to be the plesiomorphic condition, acknowledgingthat “there is no obvious polarity in the outgroup criterion.” In my taxa, the number of caudal vertebrae isalways equal or higher, except in Cr. panamensis.
57. Number of caudal vertebrae [0] modally fewer than 18; [1] modally 18 or more.
State 1 is an autapomorphy of Petenia. See preceding character. Caudal vertebrae are taken to be those that present haemal spines, either forming an arch or not.
58. Anal pterygiophores in contact with first haemal spine [0] one or two; [1] three or four; [2] usually five.
State 1 is synapomorphic for the Neotropical cichlids examined, without Tomocichla; state 2 is a strictsynapomorphy of Archocentrus (hi=0.00, ri=1.00). The character is associated with the number of anal spines(Kullander 1998).
This is Cichocki’s (1976, character 53) “number of anal-fin pterygiophores associated with and supportedby the first haemal vertebral spine,” with the same polarity as here. He coded Petenia as having my state 0;that is not what I see in the material at hand. He needed a fourth state, seven pterygiophores, for SymphysodonHeckel.
Kullander (1998, character 72) coded this character in the same way and found the same polarity, exceptthat he did not use a fourth state, but defined it as 5–7 pterygiophores articulating with the firsthaemapophysis.
59. Parhypurapophysis [0] present (Fig. 20); [1] absent.
State 1 is a synapomorphy of Hypsophrys, parallel with Amphilophus; hi=0.50, ri=0.50.Character and polarity as Cichocki’s (1976, character 60), who nevertheless distinguished two states
within my state 0, based on the relative length of the parhypurapophysis, and considered it present in H.nematopus, contrary to what I observe.
FIGURE 20. Parhypurapophysis present in caudal skeleton (Caquetaia spectabilis, UMMZ 215564). Scale:66X.
Squamation, characters 60–6860. Number of scale rows in cheek [0] modally 2 rows; [1] modally 7–8 rows; [2] modally 4–6 rows; [3] modally 3 rows.
No Neotropical cichlid examined had state 0. State 2 seems synapomorphic for all of them, except for thepiscivores, whose large jaws may be responsible for an expansion of the cheek and hence more scale rows on
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 31PHYLOGENY OF THE GENUS ARCHOCENTRUS
it (state 1). An autapomorphic reduction (state 3) towards (but not reaching) the plesiomorphic state occurs inCr. panamensis; hi=0.50, but ri=0.25.
61. Number of scale rows between lateral line and base of first dorsal ray [0] modally 3 or fewer; [1] modally 3.5 or more.
A “half-scale,” as in “3.5 scales,” is a scale distinctly smaller than those in the adjacent row on the bodyside, but not forming part of the scaly sheath at the base of the dorsal fin (Fig. 21). State 1 is synapomorphicfor Rocio (with a convergence in Ar. spinosissimus); hi=0.50, ri=0.67.
FIGURE 21. Scale rows between lateral line and base of first dorsal ray: 1.5 rows (Amatitlania siquia, UMMZ 196948).Scale: 32X.
62. Longest interradial scale row on dorsal fin [0] no scale rows; [1] longest row modally 2–12 scales long (Fig. 22); [2] longest row up to 23 scales long.
State 1 is a synapomorphy of all Neotropical cichlids examined except Petenia (convergence inTomocichla). An extremely long interradial scale row is autapomorphic for Caquetaia; hi=0.33, ri=0.00.
63. Number of rows of interradial scales on dorsal fin [0] one row; [1] two rows at base; [2] distally two rows (Fig. 22); [3] one row with sporadic supplementaryscales.
State 1 is autapomorphic for Ar. multispinosus. State 2 is a putative synapomorphy for the Neotropicalcichlids examined, except Tomocichla and Parachromis (not applicable in Petenia), but with many reversals(hi=0.75, ri=0.18). State 3 occurs only in the subgenus Bussingius of Cryptoheros, although not in Cr. sajicaand Cr. septemfasciatus. Most species show state 0 as juveniles, an ontogenetic criterion that reinforces thepolarity found.
Kullander (1998: 483) believed that “fin squamation has considerable potential as a source ofphylogenetic information, but requires more careful study,” a view I concur with.
64. Longest interradial row of scales on anal fin [0] 0–1 scales; [1] modally 3–10 scales; [2] up to 47 scales.
State 1 is synapomorphic for all Neotropical cichlids examined, except Petenia. State 2 is anautapomorphy of Caquetaia; hi=0.33, ri=0.00.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO32 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 22. Interradial scales on dorsal fin: longest row 12-scales long, distally two rows (Amatitlania kanna, FMNH59243). Scale: 16X.
65. Number of rows of interradial scales on anal fin[0] one row; [1] basally two rows; [2] distally two rows; [3] distally three rows.
States 1 and 3 are autapomorphies of Ar. multispinosus and Ar. spinosissimus, respectively. State 2 isfixed only in the subgenus Cryptoheros; hi=0.67, ri=0.25.
66. Number of scales between interpelvic scale and vent[0] modally 16 scales; [1] modally 19 scales; [2] modally 13 scales or fewer.
The interpelvic scale is an enlarged scale (sometimes a group of scales) between the bases of theinnermost pelvic-fin rays, overlapping the rays. States coded through the statistical procedure outlined above.State 1 homoplastically occurs in Petenia and Tomocichla, perhaps as a by-product of their relatively longerbodies. State 2 is synapomorphic for the rest of the Neotropical cichlids examined, with reversals in P.loisellei and H. nematopus; hi=0.60, ri=0.00.
67. Number of circumpeduncular scales [0] modally 18 or fewer; [1] modally 19 or more.
Homoplastically synapomorphic for Rocio, Parachromis, and Archocentrus (reversal in Ar. centrarchus);hi=0.80, but ri=0.43, not very low.
68. Secondary caudal pores [0] none or sporadic; [1] yes, isolated; [2] yes, forming rows
By “secondary” caudal pores I mean those pored scales that do not continue the lateral line, but appearbetween other caudal fin rays. The character is highly homoplasious (hi=0.86), but it is synapomorphicallyfixed for state 2 in Cr. spilurus + cutteri and in Hypsophrys (ri=0.20).
Fins, characters 69–7769. Number of dorsal fin spines [0] modally 16 or more; [1] modally 15; [2] modally 14 or fewer.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 33PHYLOGENY OF THE GENUS ARCHOCENTRUS
States 1 and 2 are autapomorphic for Caquetaia and Petenia, respectively. A tendency within state 0 toincrease the number of dorsal spines (a formerly diagnostic character for Archocentrus sensu lato, that is, formost of the Neotropical cichlids examined here) was not detected by the statistical coding procedure.
70. Number of dorsal fin rays [0] modally 11 or more; [1] modally 10 or fewer.
State 1 occurs in most Neotropical cichlids examined, without Petenia and Tomocichla, with reversals inCaquetaia, Amphilophus, and P. loisellei; hi=0.80, ri=0.20.
71. First dorsal fin ray [0] not divided; [1] divided; [2] spiniform.
State 1 is synapomorphic for the Neotropical cichlids examined without Petenia and Tomocichla, withreversals to State 0 in Rocio, H. nematopus, and P. dovii. State 2 is autapomorphic for H. nicaraguensis, and itis probably nested within state 0; considering this, the reversal for H. nematopus may actually be interpretedas an additional synapomorphy for Hypsophrys; hi=0.60, ri=0.50.
72. Number of anal fin spines [0] modally 3; [1] modally 5; [2] modally 6–10; [3] modally 11–12.
The states may show seriality. State 1 is homoplasious between Petenia and Tomocichla. State 2 is asynapomorphy of the rest of the Neotropical cichlids examined. State 3 is a strict synapomorphy ofArchocentrus; it was considered a diagnostic character since the description of the genus, which confirmsRegan’s (1906) intuitions (Kullander 1998: 464); hi=0.25, ri=0.67.
Cichocki (1976, character 52) “arbitrarily partitioned” the character into three states: 3, 4–6, and 7 ormore anal spines. My statistical coding does not fully coincide with his decision, but the polarity is the same.All his non-cichlid outgroups had 2 or 3 spines.
Kullander (1998, character 82) determined only two states, 4 or fewer vs. 5 or more anal spines; he foundthe same polarity as here. He observed a correlation between number of anal fin elements and body depth.
73. Number of anal fin rays [0] modally 10 or more; [1] modally 9 or fewer.
State 1 is synapomorphic for all Neotropical cichlids examined; hi=0.00. This character and my character36 are part of Kullander’s (1998) character 31, total anal fin count (spines + rays).
74. Dorsal and anal fins bearing filaments[0] no; [1] yes.
Filaments trailing caudad from the last rays of the dorsal and anal fins are putatively synapomorphic forall Neotropical cichlids examined, without Petenia and Tomocichla, and with reversals in R. gemmata and P.loisellei; hi=0.75, ri=0.25.
75. Relative position of pelvic vs. dorsal fins [0] pelvic behind dorsal; [1] pelvic below dorsal; [2] pelvic much behind dorsal.
State 1 is present only in Petenia. State 2, which is easily distinguishable from state 1 (origin of pelvicmuch more than 1 or 2 scale rows behind dorsal origin), is one of the few characters supporting the hypothesisthat Cr. panamensis and H. nematopus belong in a same genus, Neetroplus; here it is interpreted as anhomoplasy: hi=0.33, relatively low, but ri=0.00.
I discussed above the supposed incisors of Cr. panamensis and the status of Neetroplus, synonymised bySchmitter-Soto (2007) with the formerly monotypic Hypsophrys.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO34 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 23. Shape of genital papilla and pore (fish head to the left): (a) papilla oval, feminine opening oval, not (much)crenulated (Cryptoheros chetumalensis, UMMZ 197224); (b) same state, medially-constricted variation (Amatitlaniasiquia, UMMZ 196948); (c) same state, vase-shaped variation (Am. kanna, UMMZ 145716); (d) same state, circularvariation (Hypsophrys nicaraguensis, FMNH 5996); (e) same state, longer papilla (Am. siquia, UMMZ 213938); (f)papilla with mostly parallel contours, rounded terminal notch (Parachromis dovii, UMMZ 199651); (g) papilla oval,feminine opening proximally (rostrally) V-shaped (Cr. altoflavus,. MNHN 2001-1164); (h) same state, V-shape lesspronounced (Cr. myrnae, UMMZ 217739); (i) papilla oval, feminine opening very crenulated (Archocentrusspinosissimus, UMMZ 197249); (j) same state, opening deeply and not roundly notched (Ar. centrarchus, FMNH 5982).[Anal creases or folds were counted from the outside: for example, the count in (f) is 14.] Scales: a,b, 16X; c,h, 20X; d,12X; e, 66X; f,g, 32X; i, 40X; j, 25X.
a b
c d
e f
g h
i j
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 35PHYLOGENY OF THE GENUS ARCHOCENTRUS
76. Number of procurrent caudal rays, dorsal side [0] four or more; [1] three; [2] two.
Often polymorphic. State 1 is frequent in Cryptoheros and fixed in Amatitlania; state 2 is homoplasticallysynapomorphic for Archocentrus and Rocio (ri=0.64). The reductive trend has been followed several times(hi=0.80).
Kullander (1998, character 84) found the same polarity, but distinguished only two character states: morethan three vs. three procurrent rays.
77. Caudal fin profile [0] emarginate; [1] rounded-truncate; [2] definitely truncate; [3] definitely rounded.
State 1 is common to all Neotropical cichlids examined. Hypsophrys has synapomorphically reverted tostate 0. State 2 is a strict synapomorphy of Ar. centrarchus + multispinosus. State 3 is an autapomorphy ofCaquetaia; hi=0.40, ri=0.33.
Kullander (1998, character 83) found an emarginate caudal fin to be the plesiomorphic condition too, butdistinguished only two other states, namely truncate-subtruncate (my states 1 and 2) and rounded.
Genitals and internal organs, characters 78–8178. Shape of genital papilla and pore[0] papilla oval, opening oval, not (much) crenulated (Figs. 23a–e); [1] papilla with mostly parallel contours,rounded terminal notch (Fig. 23f); [2] papilla oval, opening proximally (rostrally) V-shaped (Figs. 23g,h); [3]papilla oval, opening very crenulated (Figs. 23h,i).
Oval papillae with the pore and distal end not much crenulated (state 0) occur in Oreochromis, Petenia,and most species in the study group. There are minor variations, especially in the “oval” profile of the rostralside of the papilla, which may in some individuals be peanut-shaped (medially constricted) in Am. siquia (Fig.23b), vase-shaped in R. octofasciata and Am. kanna (Fig. 23c), or approximately circular in H. nicaraguensis(Fig. 23d). Long papillae (twice as long as wide) may occur in Am. siquia (Fig. 23e), Cr. chetumalensis (Fig.23a), or Cr. altoflavus (Fig. 23g), among others, but never with such parallel contours as in the two examinedParachromis, whose rounded terminal notch is also characteristic (state 1), a strict synapomorphy.
The opening, in its proximal-rostral end, is V-shaped (not always markedly, e.g. Fig. 23h) in the subgenusBussingius, a strict synapomorphy (state 2).
The edge of the pore varies from very crenulated (Fig. 23i) to deeply and not roundly notched (Fig. 23j) inArchocentrus; a strict synapomorphy (state 3).
There is no obvious seriality in the four states; hi=0.25, ri=0.88.
79. Gut-coiling pattern [0] IP=13 in adults (Fig. 24a); [1] simple, S-shaped in adults (Figs. 24b,c); [2] double medial loop in adults,Tomocichla-type (Fig. 24d); [3] “S” folded ventrorostrally in adults, anal loop and medial loop not touching(Fig. 24e); [4] IP=6 in adults (Fig. 24f); [5] median loop twisted in adults (Fig. 24g); [6] double medial loop inadults, coatepeque-type (Fig. 24h); [7] “S” folded ventrorostrally in adults, anal loop and medial loop nearlytouching (Fig. 24i).
The Intersecting Point (IP) is a complexity index defined by Yamaoka (1982) as the number ofintersections of the gut with an imaginary left-right line (here modified as a dorsoventral line). No MiddleAmerican cichlid approximates the IP of Oreochromis (state 0). In the study group, closest in complexity isAr. multispinosus (state 4), although its autapomorphic spiral coiling pattern is quite different. IP is probablycorrelated with gut length, which is greater in herbivores.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO36 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 24. Gut type (view from right side of fish): (a) IP=13 in adults (Oreochromis mossambicus, UMMZ 213374);(b) simple, S-shaped in adults (young Cryptoheros septemfasciatus, UMMZ 190367); (c) same state, “S” shortened inadults (Caquetaia spectabilis, UMMZ 190822); (d) double medial loop in adults (secondary loop projecting away fromobserver, shown underneath gut), Tomocichla-type (T. sieboldii, UMMZ 194240); (e) “S” folded ventrorostrally inadults, anal loop and medial loop not touching (Amatitlania siquia, UMMZ 190191); (f) IP=6 in adults (Archocentrusmultispinosus, UMMZ 199539); (g) median loop twisted in adults (Cr. cutteri, UMMZ 199678); (h) double medial loopin adults (secondary loop projecting towards observer), coatepeque-type (Am. coatepeque, UMMZ 181823); (i) “S”folded ventrorostrally in adults, anal loop and medial loop nearly touching (Hypsophrys nematopus, UMMZ 197507).Scales: a,c,d,e,f,g,h,i, 6X; b, 16X.
a b
i
hg
fe
dc
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 37PHYLOGENY OF THE GENUS ARCHOCENTRUS
Most widespread within the examined species is the pattern here termed “simple,” or “S-shaped” (state 1),which is retained from juveniles to adults in most cichlids: when viewed from the right side (in order to keepstomach and liver, which are on the left side, out of view), the gut depicts an S, forming first (from the vent tothe mouth) an “anal loop” rostrally, then turning caudad to form a “medial loop,” then again rostrad to a“rostral esophageal loop,” then once again caudad to a “caudal esophageal loop,” and finally to the esophagus,which it joins at approximately the same point as the stomach. The rostral esophageal loop and the caudalesophageal loop are less abruptly folded than the anal and median loops, which are rather evenly rounded.
The “simple” type has several variations. In T. sieboldii, the median loop is doubled, with a secondaryloop projected to the left of the fish. The secondary loop occurs between the anal and median loops; this is anautapomorphy (state 2). In Am. coatepeque there is also a secondary medial loop, but this is not a parallelismwith Tomocichla, but another autapomorphy (state 6), because the secondary loop occurs between the medianand rostral esophageal loops, and it is projected to the right of the fish.
Another apparent parallelism here interpreted as separate autapomorphies occurs between H. nematopusand Am. siquia. In H. nematopus (state 7), the basic S-shape becomes ventrorostrally folded with growth, andthe anal loop is displaced caudad until it almost touches the median loop. In Am. siquia (state 3), the fold is notso abrupt, and the anal loop is displaced more dorsad than caudad, so that it does not come in contact with themedian loop; it may even come under the rostral esophageal loop.
In Cryptoheros there is a synapomorphic pattern similar to that of H. nematopus, but the median loop andsometimes also the anal loop are usually twisted clockwise (state 5). The presence of state 1 in Cr. myrnae andCr. sajica is a reversal.
In some piscivores, the S-shape is retained throughout ontogeny, but the distance between the anal loopand the median loop becomes shorter (Fig. 24c), as gut length itself becomes shorter (relative to body length).This occurs in the carnivores P. dovii (although apparently not in P. loisellei), Petenia, and Caquetaia, and isprobably an adaptive convergence. It is coded as state 1 (but see character 80, below).
The fact that most species show the “simple” pattern as juveniles supports considering this stateplesiomorphic (following an ontogenetic criterion), but, asides from that, there seems to be no clear serialityin the states of this character; hi=0.22, ri=0.67.
80. Contact of anal loop and rostral esophageal loop [0] loops not adjacent (Fig. 24c); [1] loops adjacent (Figs. 24b,d,e,g–i).
In the “simple” gut-coiling pattern, the anal and rostral esophageal loops may or may not be adjacent (intouch). This seems not to depend on the size of the liver or the repletion of the stomach (for states 2,3,5–7 incharacter 79, the anal loop and the rostral esophageal loop are always adjacent).
This is a possible synapomorphy for all examined Neotropical cichlids except Petenia and Tomocichla,but it is highly homoplasious (hi=0.80), with reversals in at least three species in different clades.
81. Peritoneum[0] uniformly dark; [1] moderate pigmentation dorsolaterally; [2] pigmentation uniformly sparse; [3] onlyroof of cavity pigmented; [4] only anterodorsally pigmented; [5] only rostrally pigmented.
State 0 differs from state 2 because the latter shows distinct melanophore concentrations, although sparseand uniformly distributed over the lining of the body cavity. State 1 is common in several clades, fixed inArchocentrus and Cryptoheros, and may be plesiomorphic to states 3, 4, and 5 (although here the trait is codedunordered). State 4 is an homoplasious synapomorphy of Rocio and Hypsophrys. States 3 and 5 areautapomorphies; hi=0.50, ri=0.44.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO38 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 25. Head and anterior-body coloration (semidiagrammatic): (a) a vertical bar just extending across nape,opercular spots part of bar (Archocentrus spinosissimus, MNHN A-0352); (b) a vertical bar fully across opercle;anteriormost bar on body sides Y-shaped, marked, its caudal arm discontinuous (Amatitlania siquia, UMMZ 245585); (c)interorbital bands present; spot on opercle part of longitudinal stripe on body sides; anteriormost bar on body sidesapproximately Y-shaped, diffuse (Rocio octofasciata, UMMZ 196387, spmn. 1); (d) full snout-eye stripe (Cryptoheroschetumalensis, ECOCH 5467); (e) interrupted snout-eye stripe (live Rocio gemmata); (f) one opercular spot, not locatedat angle of opercle; anteriormost bar on body sides Y-shaped (or V-shaped), marked, both arms continuous, rostral armcurved rostrad (Cr. nanoluteus, MNHN 1993-0260); (g) two opercular spots, aligned vertically (live Ar. centrarchus); (h)anteriormost bar on body sides I-shaped (Cr. cutteri, ANSP 53930); (i) anteriormost bar on body sides Y-shaped, marked,arms continuous, rostral arm not curved rostrad (Cr. spilurus, BMNH 1864.1.26.52).
Pigmentation pattern, characters 82–9882. Vertical bar on head [0] none or faint; [1] just extending across nape (Fig. 25a); [2] full bar across opercle (Fig. 25b).
State 1 is autapomorphic for Ar. spinosissimus. State 2 is homoplastically synapomorphic forArchocentrus, Rocio, and Amatitlania (albeit bar much stronger in the latter genus); hi=0.67, but ri=0.64.
a b c
d e f
g h i
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 39PHYLOGENY OF THE GENUS ARCHOCENTRUS
I concur with Kullander (1998: 483) that “there is more phylogenetic information in pigmentation patternthan currently recognized.” He used only four colour-pattern characters; as for Cichocki (1976), not one of his64 characters was based on pigmentation. Colour-pattern characters are usually easily observable, even in oldmuseum specimens, and quite constant (except in such polymorphic species as A. citrinellus).
83. Interocular bands [0] absent; [1] present (Fig. 25c).
State 1 is fixed in Rocio and Archocentrus; hi=0.80, ri=0.50. I do not consider the “mask” of T. sieboldiito be homologous, because its position is different.
84. Snout-eye stripe [0] absent or diffuse; [1] present, full (Fig. 25d); [2] interrupted (Fig. 25e).
State 1, homoplasious (hi=0.67), but synapomorphic (ri=0.50) for the subgenus Cryptoheros, the cladeAm. kanna + siquia, and Parachromis. State 2 is autapomorphic for R. gemmata; the “interruption” of thedark snout-eye stripe is a dorsorostrad extension of the white-bluish subocular streak.
85. Speckles on cheek [0] absent; [1] present.
State 1 hints at a synapomorphy for R. octofasciata + gemmata, with convergences in Ar. spinosissimusand P. loisellei (and Petenia). However, hi=0.83 and ri=0.00.
86. Spots on opercle [0] one, at angle; [1] as part of longitudinal stripe (Fig. 25c); [2] as part of bar on head (Fig. 25b); [3] none; [4]one, not at angle (Fig. 25f); [5] two, aligned diagonally from eye; [6] two, aligned vertically (Fig. 25g).
State 1 is homoplasious, plesiomorphically fixed in Rocio. State 2 is synapomorphic for Amatitlania, withparallelisms in Caquetaia and H. nematopus. State 3 is synapomorphic for Cryptoheros (without the basalpanamensis), with a convergence in Amphilophus and a reversal in Cr. altoflavus. States 4–6 areautapomorphic; hi=0.40, ri=0.64.
This is not Kullander’s (1998) character 90, a “strongly pigmented opercular spot,” which is “one of thestronger synapomorphies of African cichlids.” The also strong, usually ocellated opercular spot ofThorichthys would have been coded here as a different character state.
87. Dots on scales[0] none; [1] smaller than scales; [2] larger than scales.
State 1 shows up independently in several clades; although homoplasious (hi=0.71), it supports somestructure (ri=0.38). State 2 is an autapomorphy of R. gemmata. Some species, like A. citrinellus, arepolymorphic for these pigmentation features.
88. Number of bars on sides of body[0] 8 bars; [1] 7 bars; [2] 6 bars.
State 1 is probably a synapomorphy for all examined Neotropical cichlids, except Petenia andTomocichla, although with several changes to states 0 or 2 (hi=0.60, ri=0.50). The reversal to state 0 in Rociois a synapomorphy for the new genus (the bars can be hard to discern, but they are present, in R. ocotal, called“entirely black” by Miller 1957).
Although number of bars in itself is not an homologous feature, it seems that state 1 is accounted for bythe same mechanism in all examined species, i.e. the loss of the peduncular bar (bar number 2, countingposterorostrally—Kullander & Silfvergrip 1991).
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO40 · Zootaxa 1618 © 2007 Magnolia Press
89. Shape of anteriormost bar on sides of body [0] I-shaped (Fig. 25h); [1] Y-shaped, diffuse (Fig. 25c); [2] Y-shaped, marked, arms continuous, rostral armnot curved rostrad (Fig. 25i); [3] Y-shaped, marked, caudal arm discontinuous (Fig. 25b); [4] Y-shaped (or V-shaped), marked, arms continuous, rostral arm curved rostrad (Fig. 25f).
This character was interpreted as not applicable in species with only 6 bars (state 2 in character 88). States1–3 were defined separately because their differences are more conspicuous than their general Y-shape;perhaps more “Y-shaped” states might be identifiable, especially within state 1, the diffusely Y-shaped bar ofT. sieboldii, Rocio, some Archocentrus, and some Cryptoheros. State 3, a Y-shaped bar with the caudal limbdiscontinuous (or at least not uniformly intense), is a strict synapomorphy of Amatitlania. States 2 and 4 areautapomorphies; hi=0.50, ri=0.56.
90. Extension into dorsal and anal fins by bars on sides of body [0] no extension; [1] partial extension, only at fin bases; [2] full extension, to edge of fins.
State 1 is very common within Cryptoheros: a probable synapomorphy, although with some reversals anda parallelism in Ar. centrarchus. State 2 is a strict synapomorphy of Amatitlania; hi=0.60, ri=0.63.
91. Ocellus on dorsal fin of mature females [0] absent; [1] present.
State 1 is synapomorphic for Cryptoheros (ri=0.57), although with reversals in Cr. sajica and Cr.chetumalensis, and a parallelism in Ar. centrarchus (hi=0.75). The extension to the dorsal fin of a bar inAmatitlania was not considered a true ocellus (except in Am. nigrofasciata, where an ocellated spot wasobserved in 2 specimens out of 559 examined), and it is apparently not sexually dimorphic in that genus.
This ocellus occurs between the spines; not to be confused with the “tilapia spot” (Kullander 1998,character 91), which appears on the base of the soft dorsal.
92. Medial intensification of bars on sides of body[0] on bars 3, 6, 7; [1] on bar 5, sometimes also on 6; [2] on bars 6, 7, sometimes also on 8; [3] only on bar 6;[4] none.
Numbering of bars follows Kullander and Silfvergrip (1991). These medial intensifications of bars onsides of body can look like a series of blotches. State 0 does not occur in the Neotropical cichlids examined.State 1 is most widespread. State 2 is a strict synapomorphy of Amatitlania. State 3 is a synapomorphy ofCryptoheros without the basal Cr. panamensis; Cr. sajica and Ar. centrarchus convergently have state 4(hi=0.33, ri=0.82).
93. Longitudinal stripe on sides [0] none; [1] from opercle to pectoral origin; [2] full, to caudal origin; [3] from eye to lateral blotch.
State 1 occurs in Cr. nanoluteus and Cr. altoflavus, with a parallelism in Tomocichla. State 2 is aconvergence of P. dovii and Cr. myrnae. State 3 is a synapomorphy for Rocio, with parallelisms in Ar.spinosissimus and P. loisellei; hi=0.67, ri=0.25.
94. Predominant abdominal colour in life [0] whitish or greyish; [1] yellowish-reddish; [2] blue or green; [3] wine-coloured; [4] yellow green.
This is not properly a colour-pattern character, but a true colour trait. States were taken from Mayland(1995) and Werner and Stawikowski (1998), from personal observation in the field (for Petenia, Cr.chetumalensis, and R. gemmata), and from FishBase (Froese & Pauly 2004).
The character is quite homoplasious and uninformative (hi=0.75, ri=0.00). Allgayer’s (2001) assertionthat Cryptoheros may be divided into “yellow” and “grey” species is only partially sustained: C. spilurus may
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 41PHYLOGENY OF THE GENUS ARCHOCENTRUS
occasionally be yellowish, whereas Cr. septemfasciatus and Cr. sajica (but not Cr. myrnae) are greyish. Awine-coloured abdomen (and fins) is autapomorphic for C. cutteri, the so-called “weinrotflossigerBuntbarsch” of German aquarists (Velasco 2001).
95. Abdominal black blotch in mature females [0] absent; [1] present.
State 1 occurs only in the subgenus Bussingius (not in Cr. sajica); hi=0.67, ri=0.33. It is only partlycorrelated with character 91, the other trait exclusive to mature females.
96. Horizontal position of caudal spot [0] on fin; [1] on peduncle; [2] on both.
A majority of the Neotropical species examined have a caudal spot (actually most Heroini andCichlasomini: Kullander 1998, his character 88), usually on the base of the caudal fin rays (state 1). State 2, aspot exclusively on the caudal peduncle, occurs only in Cr. altoflavus and Cr. myrnae; state 3, a spot abouttwo-thirds on the peduncle and one-third on the fin, is synapomorphic for Cryptoheros (with reversals, and aparallelism with Am. siquia); hi=0.63, ri=0.38.
It can be questioned whether these states are homologous or not. However, it seems more parsimonious topostulate, for example, that the totally peduncular spot of most Amatitlania is homologous to the two-thirds-peduncular, one-third-pterygial, spot of Am. siquia, than to postulate for this latter species a second spot onthe fin, complementing the oval shape of the first, peduncular spot. Another example of a caudal spot whoseposition varies between the peduncle and the fin can be found in the ‘Cichlasoma’ urophthalmus complex(Barrientos-Medina 2005).
97. Vertical position of caudal spot [0] crossing lateral line; [1] completely above lateral line.
State 1 is homoplastically synapomorphic for Rocio; hi=0.75, ri=0.40.
98. Margin of caudal spot [0] not ocellated; [1] ocellated.
A true caudal ocellus, with a whitish-bluish margin, is synapomorphic for Rocio, but homoplasious withA. citrinellus (where it is polymorphic), Ar. centrarchus, and many other taxa not examined here, e.g. “C.”urophthalmus, Cichla Bloch & Schneider, and Astronotus Swainson; hi=0.67, ri=0.50.
Cytotaxonomy
Thompson’s (1979) cytogenetic data (corroborated for some species by Salas & Boza 1991) were tried as a
99th character, with three states: [0] 48 chromosomes, 8 (sub)metacentric (sm) / 40 (sub)telocentric (st); [1] 48,6 sm / 42 st; [2] 48, 4 sm / 44 st. For simplicity, Oreochromis, with a fundamental number different from 48,was considered not applicable. Polarity was argued by Thompson (1979) himself. Most of the speciesincluded here had to be coded as unknown. The character was not particularly homoplasious (hi=0.50): A.citrinellus, H. nematopus and P. dovii showed state 0; Archocentrus (Ar. centrarchus and Ar. multispinosus),Cr. septemfasciatus and R. octofasciata had state 1, thus homoplastically synapomorphic for Archocentrus;Amatitlania displayed state 2.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO42 · Zootaxa 1618 © 2007 Magnolia Press
FIGURE 26. Strict consensus of the three minimum-length trees under ACCTRAN optimization in PAUP (556 steps,CI=0.42, RI=0.45, HI=0.61). Clades with less than 50% bootstrap support are shown collapsed, except for those thathave a strict synapomorphy with a character whose hi = 0.50, even when not recovered by the bootstrap procedure.Numbers above lines are synapomorphies, with the exponent indicating the character state and an asterisk markinghomoplasious synapomorphies. Numbers below lines are boostrap support. For character numbering, see Characterdescription and analysis.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 43PHYLOGENY OF THE GENUS ARCHOCENTRUS
Phylogeny
An exhaustive search was not feasible; three most-parsimonious trees were found by a heuristic search. Figure26 shows the strict consensus of those minimum-length trees (556 steps, CI=0.42, RI=0.45, HI=0.61).Excluding uninformative characters (12 autapomorphies), the hypothesis does not substantially weaken:CI=0.40, HI=0.60.
The consensus cladogram shows no evidence in support of Archocentrus as commonly used in theliterature (e.g. Bussing 1998). Archocentrus as traditionally used actually consists of three clades(Archocentrus, Cryptoheros, and Amatitlania), one of which contains the former monotypic genusHerotilapia in a derived, not basal, position.
Other taxa at various times associated with Archocentrus are also probably distinct, contained in theclades Rocio and Hypsophrys (which includes the former monotypic genus Neetroplus). Beyond that level,the tree collapses (Fig. 26; synapomorphies discussed above). According to this hypothesis, Tomocichla is thesister taxon to the rest of the American cichlids examined; Petenia is basal to the rest, and then there is apolytomy, including Amphilophus, Caquetaia, and the following resolved groups:
• Archocentrus sensu stricto consists of Ar. centrarchus, Ar. multispinosus, and Ar. spinosissimus, the lat-ter being basal to the others.
• Amatitlania consists of Am. nigrofasciata, Am. coatepeque, Am. kanna, and Am. siquia, the Salvadoranspecies being basal to the rest, and the southernmost species-pair being most derived.
• Rocio consists of R. octofasciata, R. ocotal, and R. gemmata. Internal structure of this clade was notresolved.
• Cryptoheros consists of Cr. spilurus, Cr. chetumalensis, Cr. cutteri (the three of them integrating thesubgenus Cryptoheros, with Cr. chetumalensis in the basal position), and the five species in the newsubgenus Bussingius (no internal structure resolved), as well as Cr. panamensis, which is the sistergroup of the other two subgenera.
• Hypsophrys nicaraguensis and H. nematopus show up together in one clade.• The two examined species of Parachromis appear alone in their own clade.Including the cytogenetic data did not alter the topology of the tree (now with 561 steps, CI=0.43, same
RI and HI).
Discussion
The present interpretation of Archocentrus (synonymizing Herotilapia—Schmitter-Soto 2007) hinges on threestrict synapomorphies: five pterygiophores in contact with the first haemal spine, eleven or more anal-finspines, and genital papilla oval, its opening very crenulated to deeply and not roundly notched. The first twocharacters are part of the traditional diagnosis of Archocentrus; in spite of their recurrent use in the literature,and in spite of their being probably correlated between themselves (Kullander 1998), I do not think that theyhave been overused, since they constitute a strict, robust, synapomorphy in the context of the present matrix.
Other many-spined cichlids, like Symphysodon, have been considered to have a different character-state(Cichocki 1976): 7 or more, rather than 5 or more spines. The peritoneal pigmentation pattern, althoughapparently acquired independently by Archocentrus, Cryptoheros, and other cichlids, is anothersynapomorphy for the genus as here construed.
Martin and Bermingham’s (1998: fig. 4) cladogram implied restricting Archocentrus to Cr.septemfasciatus, Am. nigrofasciatus, and Ar. centrarchus. These authors suggested that Cr. myrnae and Cr.sajica, which appeared in a different clade, might belong in their own distinct genera. Cr. myrnae and Cr.sajica clustered together in their cladogram with species of Parachromis and Hypsophrys (with Neetroplus),and this clade had a sister-group relationship to the purely-Archocentrus clade mentioned above;Paraneetroplus (= Tomocichla) sieboldii was the sister group of these clades. On the other hand, although
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO44 · Zootaxa 1618 © 2007 Magnolia Press
Archocentrus resulted paraphyletic in Martin and Bermingham’s (1998) work, cladograms constraining thegenus to be monophyletic were not significantly different in terms of changes per site.
Previously (Roe et al. 1997: fig. 2A), Cr. spilurus showed up in the same clade as representatives ofAmphilophus and Parachromis. Typically, Cryptoheros and Parachromis were sister groups in the cladogramsof Roe et al. (1997); evidence for such a relationship is lacking in my study.
Kullander (1998) found a suite of characters which “express the deep body” (high number of anal-finspines, presence of laminar expansions on anterior haemapophyses, lachrymal deeper than wide, etc.) in theclade that consists of the heroine genera Symphysodon, Heros, Uaru Heckel, Mesonauta Günther, andPterophyllum Heckel. It seems that these characters are not associated together in the same way inArchocentrus; for example, the lachrymal is wider than deep in adult Ar. centrarchus.
Further characters that could a priori be considered part of a deep-body gestalt, such as a low number ofvertebrae, do not unequivocally characterise either Archocentrus or the Symphysodon clade; still others, like ahigh number of circumpeduncular scales, are (rather paradoxically) convergent between the deep-bodiedArchocentrus, the slender Parachromis, and Rocio, whose body depth tends to decrease ontogenetically.
Other deep-bodied American cichlids include Heroina Kullander, some Theraps Günther, and Heros,among others, but none of these has ever been considered close to Archocentrus by traditional standards orfound near Archocentrus in phylogenetic analyses. On the contrary, most of them have appeared quite apartfrom Archocentrus in published cladograms. Hence, they were not included in the present analysis, and itseems unlikely that future work will change this, given that they lack the synapomorphic characters of thegenus (see also Kullander 1998).
The placement of Ar. multispinosus, the former Herotilapia, constitutes a conflict between morphologyand molecules. Several workers (e.g. Farias et al. 2001; P. Chakrabarty, in litt., 2003; Concheiro Pérez et al.,2007) have found Herotilapia to lie far outside the clade that includes the type species of Archocentrus.However, in addition to the synapomorphies for the genus, Ar. multispinosus shares characters with the typespecies, Ar. centrarchus: the caudal fin profile, definitely truncate, is a uniquely derived character within thespecies suite examined, whereas the evenly-convex dorsal margin of the opercle (with a dorsoanteriorprotuberance) is homoplastically synapomorphic, same as the combination of 6 (sub)metacentric and 42(sub)telocentric chromosomes (Thompson 1979). Thus, Ar. multispinosus occurs well within theArchocentrus clade, on morphological grounds.
On the other hand, the conflicting cytochrome-b data may possibly suffer from the general problem withsuch information as applied to Middle American cichlids (Schmitter-Soto 2007; see below). In addition, thelarge number of autapomorphies of Ar. multispinosus might be a confounding factor (the “long-branchattraction” phenomenon—cf. Wiens & Hollingsworth 2000). Hulsey et al. (2004) were unable to place Ar.multispinosus unambiguously in their cladogram. Data based on genes other than cytochrome b should helpclarify this problem.
Notwithstanding the large number of “trophic characters” in my study, piscivores (Parachromis, Petenia, andCaquetaia) did not cluster together. Considering this, their resemblance is most likely due to adaptiveconvergence, as Kullander (1998) believed, and contra Miller (1993, Miller et al. 2005), who suggested thatCaquetaia should be classified as a subgenus of Petenia. Hulsey et al. (2004) coincided in differentiatingCaquetaia from Petenia; Martin and Bermingham (1998) even suggested that Caquetaia was polyphyletic.
On the other hand, a nuclear-gene analysis by Farias et al. (2000) did find Caquetaia in the same clade asPetenia and Rocio, a “piscivore clade.” Farias et al. (1999) found Caquetaia and Petenia lumped together with A.citrinellus and Cr. nigrofasciatus, and the clade persisted in a later hypothesis (Farias et al. 2001), whereParachromis, Paraneetroplus, and “Nandopsis” were added. My cladogram speaks against including eitherCaquetaia, Parachromis, or Rocio in a clade with Petenia, but it cannot rule out a clade consisting of the formerthree genera.
Rocio is the most strongly supported clade in my tree. According to Miller (1993, Miller et al. 2005), it is anArchocentrus. Regan (1908) put R.. octofasciata in section Archocentrus, but he acknowledged that its
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 45PHYLOGENY OF THE GENUS ARCHOCENTRUS
relationships were “rather doubtful,” and shortly later (Regan 1909) he considered it a Parapetenia, mirroring thefact that Rocio species are deep-bodied (Archocentrus-like) when young and as they grow they become slender-bodied (Parachromis-like). Farias et al. (2000) located the species within a “piscivore clade” with Petenia andCaquetaia; Kullander (2003) listed R. octofasciata as incertae sedis. The present hypothesis cannot rule out aRocio+Archocentrus clade, but it doesn’t forbid a possible Rocio + Parachromis clade either, or indeed anycombination of Rocio, Parachromis, Amphilophus, Cryptoheros, Archocentrus, Amatitlania, and Hypsophrys.
The strict synapomorphies supporting Rocio have to do with gill rakers and neurocranium. A hint at aputative Rocio-Parachromis relationship like the one found by the molecular studies cited above might be, forexample, the state 2 of character 42, symphysial teeth abruptly larger than adjacent teeth: another “piscivoretrait,” which occurs in Rocio, Parachromis, and Caquetaia (also in Amatitlania and elsewhere, but not inPetenia). These characters tend to be highly homoplasious, as might be expected of adaptive structures;moreover, in the present hypothesis “piscivore characters” tend to be apomorphic and convergently evolved indifferent clades, which reinforces the idea that they constitute adaptations (Coddington 1988).
Schmitter-Soto’s (2007) decision to exclude Amatitlania from Archocentrus (or Cryptoheros) will not remainuncontroversial. Previously, Martin and Bermingham (1998) and Hulsey et al. (2004) found Am. nigrofasciata ina clade with Cr. septemfasciatus. However, Amatitlania is fairly well supported in the present cladogram,exhibiting three strict synapomorphies: the shape of the anteriormost bar on sides, the degree of extension of barson sides onto dorsal and anal fins, and the type of medial intensification of these bars.
The synapomorphies of Amatitlania are all related to pigmentation. Available published information oncolour (also live colour) and relation to behaviour could be used more deeply for phylogenetic analyses. Loiselle(pers. comm. 2005) suggested exploring as cladistic characters the presence/absence of sexual colour dimorphismand its polarity; presence/absence of yellow or red pigment in quiescent, courting and parental fish; enhancementor suppression of vertical elements in the melanophore pattern of parental individuals; presence/absence ofreverse countershading in parental individuals; etc.
Although homologisation might remain a problem for many live colour and behavioral characters, it isinteresting that Archocentrus is (almost) not sexually dimorphic, contrary to Amatitlania and Cryptoheros,whose sexual dimorphism is strong. Moreover, Archocentrus species are open-substrate breeders, whereasAmatitlania and Cryptoheros lay their eggs under stones (Stawikowski & Werner 1998). Predominantabdominal colour in life was used, but it was not a particularly informative character, as yellow or red pigmentis present in many clades. A blue iris occurs in Cr. myrnae and Cr. septemfasciatus, but such a clade was notsupported in this hypothesis. The reverse countershading of mature H. nematopus (Bussing 1998) seems to beanother autapomorphy of that species.
Cryptoheros including Cr. panamensis is not a very robust clade: its bootstrap support is only 59%,compared to 77% for Cryptoheros without Cr. panamensis, or 86% for the subgenus Cryptoheros. The placementof Cr. panamensis is controversial: it was described as a Neetroplus (Meek & Hildebrand 1913), Bussing (1998)considered it a Hypsophrys, and Kullander (2003) felt that it belonged in Archocentrus (including Cryptoheros).
Cr. panamensis is clearly not related to Hypsophrys (including Neetroplus), the supposedly incisor-like teethhaving a different ontogeny from the true incisors of H. nematopus; other similarities, for example the posterior-end shape of the dentigerous arm of the dentary bone, are symplesiomorphies. It is not an Archocentrus, either,the shared characters being primitive. It lacks important synapomorphies of the rest of the genus Cryptoheros,such as the convex dorsal edge of the articular bone; however, it shows the strict synapomorphy of having thevery characteristic three to five acute interdigitations between the halves of the lower pharyngeal jaw.
There are similarities in breeding coloration between Cr. panamensis and species of Tomocichla (Loiselle,pers. comm., 2005). However, T. sieboldii lies quite far from the clade that includes Cryptoheros, both in thepresent cladogram and in previous hypotheses (e.g. Martin & Bermingham 1998). Hulsey et al. (2004) foundTomocichla to be polyphyletic, but T. underwoodi Regan was even farther away from Cryptoheros in theircladogram.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO46 · Zootaxa 1618 © 2007 Magnolia Press
The subgenera of Cryptoheros are biogeographically disjunct: Cryptoheros in the northwest, Bussingius inthe southeast. The subgenus Cryptoheros shows no strict synapomorphy, but a bootstrap support of 86%; itshomoplasious synapomorphies concern the number of scale rows on the anal fin and the presence of a spineon the dorsal margin of the opercle, among others. Bussingius has a very low bootstrap support (below 50%),but one strict synapomorphy, a characteristically V-shaped opening of the genital papilla.
The subtribe Archocentrina Allgayer (2001) is not recovered in my hypothesis: no character unequivocallyunites Archocentrus, Cryptoheros, and Amatitlania but excluding Cr. panamensis. An irregular posterior edge ofthe urohyal bone characterises Cryptoheros and Archocentrus, but not Amatitlania; the limbs of the palatinebones are subequally long in Amatitlania and Archocentrus, but not in Cryptoheros (except Cr. sajica); theposterior end of the dentigerous arm of the dentary bone is bluntly pointed in Amatitlania and Cryptoheros, butnot in Archocentrus; etc.
The monophyletic pair Hypsophrys + Neetroplus has been sustained by several workers on moleculargrounds (Martin & Bermingham 1998, Hulsey et al. 2004, Concheiro Pérez et al., 2007). However, the verystriking autapomorphies of both species seem to have discouraged any decision to synonymise theirmonotypic genera.
As represented in this study, Parachromis is monophyletic, a fact which supports recognition of this taxon,for which there has been no study including all species assignable to the genus.
The tilapia, a rather derived African cichlid, was not always applicable as an outgroup. However, replicatingthe analysis using Tomocichla and Petenia as outgroups yields the same result. I saw no need to include otherSouth American or non-American cichlids, because the monophyly of Heroini is already well-established(Kullander 1998; Farias et al. 1999, 2000, 2001).
As discussed above, the results of the available phylogenies are partly at variance with mine, but they varyamong themselves as well. Martin and Bermingham (1998) rather gloomily observed that the Heroini “…areunfortunately refractory to phylogenetic inference using cytochrome b gene sequences…” Meyer (1994)commented also that “…cytochrome b may not be ideally suited for inferring the evolutionary history ofCentral American cichlids,” and Doyle (1992) pungently characterised many (most?) molecular studies,especially those based on the non-recombinant mtDNA, as “one-character phylogenies.” Moore (1995) added:“…single-gene reconstructions may lead to inconclusive discrimination of species trees among gene trees.” Ithink it is consilience, the congruence between independent analyses and datasets, either molecular ormorphological, that gives strength to evolutionary hypotheses (cf. Farias et al. 2000).
On the other hand, consistency for the phylogenetic hypothesis presented here may seem low (0.42), butthe figure is not atypical, compared to other works on cichlids: 0.57 for Roe et al. (1997) and 0.48 forKullander (1998), but 0.37 for the total-evidence cladogram of Farias et al. (2000), 0.35 for Sparks (2003) and0.22 for Farias et al. (2001). The large number of taxa and the inclusion of many homoplasious (althoughinformative to greater or lesser extent) characters is the cause of this, and the phenomenon seems hard toavoid in (heroine) cichlid phylogenies, whether morphological or molecular.
The use of statistics for coding quantitative characters has been criticised (e.g. Pimentel & Riggins 1987)for using attributes of populations, not of individuals. It remains true, however, that overlapping variabilitymay exist also in “qualitative” characters (Kitching et al. 1998), and a statistical coding approach allows for amore objective decision on what constitutes a “representative” individual (i.e., one displaying the mostrepresentative character states) for a taxon. “It is expected that taxa that inherit a particular character state fora metric attribute from a common ancestor will inherit both the central tendency and distribution of the stateunchanged” (Rae 1998, his italics).
Kullander (1998) found that meristic scale characters were too variable to be useful for cladisticinvestigations of cichlids. In the present study, for example, the number of predorsal scales was statisticallyamenable to the distinction of two states, modally less than 16 vs.16 or more, but the difference failed to besignificant; moreover, the distribution of state 1 was highly homoplasious (although this was not in itself a
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 47PHYLOGENY OF THE GENUS ARCHOCENTRUS
reason for discarding the information). Other meristic characters, however, did provide useful data, withretention index values up to 0.67.
The use of gut-coiling data is not new in fish systematics (Yamaoka 1982). One synapomorphy of familyCichlidae lies in the position of the intestine relative to the stomach (Stiassny 1991). Takahashi (2003)provides a recent example with cichlids from Lake Tanganyika; although the gut of these African cichlids ismuch more complex than that of the Mesoamerican Heroini, the latter often also require some ontogeneticinformation to define character states. In any case, the consistence of the patterns must be considered, but thisis true of almost any taxonomic character.
Two main evolutionary trends are apparent in gut-coiling patterns. One is associated with the relativereduction of intestine length in carnivores (as seen in Parachromis, Petenia, and Caquetaia). The other one isthe elongation of the gut in herbivores, an adaptation for a better digestion of vegetal matter. The problem ofaccomodating an elongated gut within the limited space of the coelom provides more informative patterns;Shen and Wu (1995) were able to construct a phylogeny for Balistidae and Monacanthidae using onlyintestinal-coiling pattern data. Middle American Heroini show much simpler patterns than those found inthese families, or than the patterns of tilapiines; the rather subtle differences found (minute variations on theS-shape theme) are mostly autapomorphic, except for the interesting intestinal synapomorphy of Cryptoheros(including Cr. panamensis).
The external morphological diversity of genital papillae in cichlids is probably not adaptive. Genitalpapillae provided strict synapomorphies for Archocentrus, Bussingius, and Parachromis, which showcharacter states that are independent modifications of the basic oval-shaped papilla with a simple, not muchcrenulated opening: in Parachromis the papilla elongates, in Bussingius and Archocentrus the opening showsdiverse degrees of divergence from the plesiomorphic evenly-rounded pore.
Functional differences of the genital apparatus, for example in the timing and intensity of spawning, arecontrolled by internal structures such as the presence and type of sphincter muscles (Rasotto & Shapiro 1998),and relative differences in size may be mediated by differences in androgen levels, which in cichlids can besocially modulated (Oliveira et al. 2002). The internal musculature of the genital apparatus in cichlids mayhold promise to the discovery of further cladistic characters.
A formal biogeographic analysis of the present phylogenetic hypothesis remains to be performed, butexamination of the distributional patterns within each clade suggests interesting generalities:
Basal species apparently tend to be inland isolated endemics: such is the case within Amatitlania, wherethe crater-lake species Am. coatepeque is the sister group of the other three species in the genus. Within Rociothere was no cladistic resolution, but one could speculate that the highland R. ocotal would be basal to theother two species; indeed, R. octofasciata and R. gemmata do exhibit putative synapomorphies (e.g.bicuspidate lower symphysial teeth, speckled cheeks), although a larger sample of characters will be neededto elucidate this issue.
Basal species apparently tend to be native to northern-western Central America: this is the pattern withinCryptoheros (Cryptoheros), with the Mexican-Belizean Cr. chetumalensis basal to the other two species, onedwelling in the Izabal drainage, the other in Honduran waters. The pattern appears also in Amatitlania(without Am. coatepeque), where Am. nigrofasciata of Guatemala-Honduras is basal to the other two species,which occur from Nicaragua to Panama. This is seen again in Archocentrus, with Ar. spinosissimus ofGuatemala basal to the Honduran-Nicaraguan-Costa Rican species.
Northern Central America or “Nuclear Central America” (Kesler 1971) corresponds roughly to thehighlands of Chiapas and Guatemala. It is a geologically complex area, which remained available tofreshwater fishes while Lower Central America did not fully emerge to connect the region to South America.The biogeographic patterns outlined above seem consistent with a recolonisation of Lower Central Americafrom Nuclear Central America by several cichlid lineages, after the initial northwards colonisation by theirancestors (the first colonisation of Nuclear Central America by cichlids most likely occurred before a fully
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO48 · Zootaxa 1618 © 2007 Magnolia Press
freshwater connection from South America was established—Bussing 1976, Álvarez del Villar 1977). Thisscenario might hold for other fishes as well, for example Rhamdia laticauda Heckel (Perdices et al. 2002:185).
Deeper in the cladogram, the pattern is coherent with the invasion of Middle America from SouthAmerica, because the Panamanian Cr. panamensis is basal to Costa Rican-Nicaraguan and Guatemalan-Mexican clades. However, the South American Caquetaia lies within the ingroup; virtually all previousworkers have also found Caquetaia nested within the Mesoamerican cichlids (Martin & Bermingham 1998;Farias et al. 1999, 2000, 2001; Chakrabarty in litt., 2003; Hulsey et al. 2004), which speaks againstmonophyly of the strictly Middle American cichlids and underscores the probably great complexity of theirbiogeographical history.
Acknowledgments
Sven Kullander, Paul Loiselle, and Carter Gilbert performed very detailed and constructive reviews of thispaper.
This work was done while on sabbatical stay at the University of Michigan Museum of Zoology, Divisionof Fishes, under a Fulbright grant. I thank most warmly my hosts, Bill Fink and Jerry Smith. Doug Nelsonhelped with loan management, and Prosanta Chakrabarty with radiographs.
Not only for loan of material under their care (including permission to C&S some specimens), but also forhospitality and enlightening exchange of ideas, I wholeheartedly thank Mary Anne Rogers and MarkWestneat (FMNH), Guy Duhamel and Javier Gregorio (MNHN), Oliver Crimmen and Patrick Campbell(BMNH), Sonia Fisch-Müller and Claude Weber (MHNG), John Lundberg and Kyle Luckenbill (ANSP),
Rocío Rodiles-Hernández and Alfonso González-Díaz (ECOSC), J.T. Williams (USNM), Oldřich Říčan and
Sven Kullander (NRM), Salvador Contreras-Balderas (UANL, co-discoverer of R. gemmata), Reeve Bailey,Jean Huber, Darrin Hulsey, Ron Oldfield, Melanie Stiassny, and Martha Valdez. Prosanta and Oldřich freelyand generously discussed with me their unpublished phylogenetic analyses.
References
Agassiz, L. (1859) Remarks on new fishes from Lake Nicaragua. Proceedings of the Boston Society of Natural History,6, 407–408.
Allgayer, R. (1994) Description d’une espèce nouvelle du genre Archocentrus. Revue Française des Cichlidophiles, 135,6–24.
Allgayer, R. (2001) Description d'un genre nouveau, Cryptoheros, d'Amérique Centrale et d'une espèce nouvelle du Pan-ama (Pisces: Cichlidae). L’An Cichlidé, 1, 13–20.
Álvarez del Villar, J. (1977) Relación entre la geomorfología mesoamericana y la distribución actual de los peces. In :Ferrusquía-Villafranca, I. (Ed.) Conexiones Terrestres entre Norte y Sudamérica: Simposio Interdisciplinario sobrePaleogeografía Mesoamericana. Instituto de Geología, Universidad Nacional Autónoma de México, Mexico City,pp. 182-192.
Archie, J. (1989) Homoplasy excess ratios: new indices for measuring levels of homoplasy in phylogenetic systematicsand a critique of the consistency index. Systematic Zoology, 38, 253–269.
Barrientos-Medina, R.C. (2005) Estado Taxonómico de la Mojarra Rayada ‘Cichlasoma’ urophthalmus Günther, 1862(Teleostei: Cichlidae). Unpubl. M.Sc. Thesis, El Colegio de la Frontera Sur, Chetumal, 108 pp.
Burgess, W.E. (2000) The Cichlasoma story. Herichthys, the break-up. Tropical Fish Hobbyist, 48, 44–54.Bussing, W.A. (1974) Two new species of cichlid fishes, Cichlasoma sajica and C. diquis, from southeastern Costa Rica.
Revista de Biología Tropical, 22, 29–49.Bussing, W.A. (1976) Geographic distribution of the San Juan ichthyofauna of Central America with remarks on its ori-
gin and ecology. In: Thorson, T.B. (Ed.), Investigations of the Ichthyofauna of Nicaraguan Lakes. University ofNebraska, Lincoln, pp.157–175.
Bussing, W.A. (1989) Cichlasoma loisellei, a new Nandopsis group cichlid fish from Central America. Revista de
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

Zootaxa 1618 © 2007 Magnolia Press · 49PHYLOGENY OF THE GENUS ARCHOCENTRUS
Biología Tropical, 37, 153–161. Bussing, W.A. (1998) Peces de las aguas continentales de Costa Rica. Revista de Biología Tropical, 46, 1–468. Casciotta, J. & Arratia, G. (1993) Jaws and teeth of American cichlids (Pisces: Labroidei). Journal of Morphology, 217,
1–36.Cichocki, F.P. (1976) Cladistic History of Cichlid Fishes and Reproductive Strategies of the American Genera Acarich-
thys, Biotodoma, and Geophagus. Vol. 1. Unpubl. Ph.D. Thesis, University of Michigan, Ann Arbor, 356 pp.Coddington, J.A. (1988) Cladistic tests of adaptational hypotheses. Cladistics, 4, 3-22.Concheiro Pérez, G.A, Řičan, O., Ortí, G., Bermingham, E., Doadrio, I. & Zardoya, R. (2007) Phylogeny and biogeogra-
phy of 91 species of heroine cichlids (Teleostei: Cichlidae) based on sequences of the cytochrome b gene. MolecularPhylogenetics and Evolution, 43, 91–110.
Doyle, J.J. (1992) Gene trees and species trees: Molecular systematics as one-character taxonomy. Systematic Botany,17, 144–163.
Farias, I.P., Ortí, G. & Meyer, A. (2000) Total evidence: molecules, morphology, and the phylogenetics of cichlid fishes.Journal of Experimental Zoology, 288, 76–92.
Farias, I.P., Ortí, G., Sampaio, I., Schneider, H. & Meyer, A. (1999) Mitochondrial DNA phylogeny of the family Cichl-idae: monophyly and fast evolution of the Neotropical assemblage. Journal of Molecular Evolution, 48, 703–711.
Farias, I.P., Ortí, G., Sampaio, I., Schneider, H. & Meyer, A. (2001) The cytochrome b gene as a phylogenetic marker: thelimits of resolution for analyzing relationships among cichlid fishes. Journal of Molecular Evolution, 53, 89–103.
Farris, J.S. (1989) The retention index and the rescaled consistency index. Cladistics, 5, 417–419.Felsenstein, J. (1985) Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39, 783–791.Fowler, H.W. (1932) Notes on the freshwater fishes from Central America. Proceedings of the Academy of Natural Sci-
ences of Philadelphia, 64, 379–385.Froese, R. & Pauly, D. (Eds.) (2004) FishBase. Available from www.fishbase.org (accessed 15 December 2004)Gill, T. & Bransford, J.F. (1877) Synopsis of the fishes of Lake Nicaragua. Proceedings of the Academy of Natural Sci-
ences of Philadelphia, 29, 175–218.Günther, A. (1862) Catalogue of the fishes in the British Museum. Vol. IV. British Museum, London, 534 pp.Günther, A. (1864) Report on a collection of fishes made by Messrs. Dow, Godman and Salvin in Guatemala. Proceed-
ings of the Zoological Society of London, 1864, 114–154.Günther, A. (1867) Memoir on the Fishes of the States of Central America, founded upon specimens collected in the
fresh and marine waters of various parts of that country by Messrs. Salvin and Godman and Capt. J.M. Dow. Pro-ceedings of the Zoological Society of London, 1866, 600–609.
Hulsey, C.D., García de León, F.J., Sánchez Johnson, Y., Hendrickson, D.A. & Near, T.J. (2004) Temporal diversificationof Mesoamerican cichlid fishes across a major biogeographic boundary. Molecular Phylogenetics and Evolution, 31,754–764.
Jordan, D.S. & Evermann, B.W. (1898) The fishes of North and Middle America: A descriptive catalogue of the speciesof fishes and fish-like vertebrates found in the waters of North America, north of the Isthmus of Panama. UnitedStates National Museum Bulletin, 47, 955–1936.
Kesler, S.E. (1971) Nature of ancestral orogenic zone in Nuclear Central America. American Association of PetroleumGeologists Bulletin, 55, 2116–2129.
Kitching, I.J., Forey, P.L., Humphries, C.J. & Williams, D.M. (1998) Cladistics. The Theory and Practice of ParsimonyAnalysis. 2nd Ed. Oxford University Press, Oxford, 228 pp.
Kullander, S.O. (1996) Heroina isonycterina, a new genus and species of cichlid fish from Western Amazonia, with com-ments on cichlasomine systematics. Ichthyological Exploration of Freshwaters, 7, 149–172.
Kullander, S.O. (1998) A phylogeny and classification of the South American Cichlidae (Teleostei: Perciformes). In:Malabarba, L.R., Reis, R.E., Vari, R.P., Lucena, Z.M.S. & Lucena, C.A.S. (Eds.), Phylogeny and classification ofNeotropical fishes. Edipucrs, Porto Alegre, pp. 461–498.
Kullander, S.O. (2003) Family Cichlidae (Cichlids). In: R.E. Reis, S.O. Kullander & C.J. Ferraris, Jr., Jr. (eds.). CheckList of the Freshwater Fishes of South and Central America. Edipucrs, Porto Alegre, pp. 605–654.
Kullander, S.O. & Silfvergrip, A. (1991) Review of the South American cichlid genus Mesonauta Günther (Teleostei,Cichlidae), with descriptions of two new species. Revue Suisse de Zoologie, 98: 407–448.
Loiselle, P.V. (1997) Diagnosis of two new cichlids from the Río Sixaola drainage, Costa Rica. Buntbarsche Bulletin,180, 1–8.
Martin, P.A. & Bermingham, E. (1998) Systematics and evolution of lower Central American cichlids inferred fromanalysis of cytochrome b gene sequences. Molecular Phylogenetics and Evolution, 9, 192–203.
Mayland, H.J. (1995) Cichliden. Landbuch, Hannover, 596 pp.Meek, S.E. & Hildebrand, S.F. (1913) New species of fishes from Panama. Field Columbian Museum of Natural History
Publications, Zoological Series, 166, 77–91.Meyer, A. (1994) Shortcomings of the cytochrome b gene as a molecular marker. Trends in Ecology and Evolution, 9,
278–280.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.

SCHMITTER-SOTO50 · Zootaxa 1618 © 2007 Magnolia Press
Miller, R.R. (1957) Biological investigations in the Selva Lacandona, Chiapas. Fishes from Laguna Ocotal. Bulletin ofthe Museum of Comparative Zoology, 116, 238–242.
Miller, R.R. (1993) [Review of] North and Central American Cichlids, by Donald Conkel. Buntbarsche Bulletin, 157,18–19.
Miller, R.R., Minckley, W.L. & Norris, S.M. (2005) Freshwater Fishes of México. Museum of Zoology, University ofMichigan, & University of Chicago Press, Chicago, 652 pp.
Moore, W.S. (1995) Inferring phylogenies from mtDNA variation: mitochondrial-gene trees versus nuclear-gene trees.Evolution, 49, 718–726.
Oliveira, R.F., Hirschenhauser, K., Carneiro, L.A. & Canario, A.V.M. (2002). Social modulation of androgen levels inmale teleost fish. Comparative Biochemistry and Physiology, B 132, 203–215.
Pellegrin, J. (1904) Contribution à l’étude anatomique, biologique et taxinomique des poissons de la famille desCichlidés. Mémoires de la Societé Zoologique de la France, 16, 41–399.
Perdices, A., Bermingham, E., Montilla, A. & Doadrio, I. (2002) Evolutionary history of the genus Rhamdia (Teleostei:Pimelodidae) in Central America. Molecular Phylogenetics and Evolution, 25, 172–189.
Pimentel, R.A. & Riggins, R. (1987) The nature of cladistic data. Cladistics, 3, 201–209.Rae, T.C. (1998) The logical basis for the use of continuous characters in phylogenetic systematics. Cladistics, 14, 221–
228.Rasotto, M.B. & Shapiro, D.Y. (1998). Morphology of gonoducts and male genital papilla, in the bluehead wrasse: impli-
cations and correlates on the control of gamete release. Journal of Fish Biology, 52, 716-725.Regan, C.T. (1903) Descriptions de poissons nouveaux faisant partie de la collection du Musée d'Histoire Naturelle de
Genève. Revue Suisse de Zoologie, 11, 413–418.Regan, C.T. (1905) A revision of the fishes of the American cichlid genus Cichlosoma and of the allied genera. Annual
Magazine of Natural History, 7, 60–77, 225–243, 316–340, 433–445.Regan, C.T. (1906) Pisces. In: Biologia Centrali-Americana. Vol. 8, 203 pp.Regan, C.T. (1908) A collection of freshwater fishes made by Mr. C.F. Underwood in Costa Rica. Annual Magazine of
Natural History, 8, 84–88.Regan, C.T. (1909) Descriptions of three new freshwater fishes from South America, presented to the British Museum by
Herr J. Paul Arnold. Annual Magazine of Natural History, 3, 234–235. Roe, K.J., Conkel, D. & Lydeard, C. (1997) Molecular systematics of Middle American cichlid fishes and the evolution
of trophic-types in ‘Cichlasoma (Amphilophus)’ and ‘C. (Thorichthys)’. Molecular Phylogenetics and Evolution, 7,366–376.
Salas, E. & J. Boza (1991) Citotaxonomía comparativa de tres especies de Cichlasoma (Pisces: Cichlidae) nativas deCosta Rica. Revista de Biología Tropical, 39, 219–224.
Schmitter-Soto, J.J. (2007) A systematic revision of the genus Archocentrus (Perciformes: Cichlidae), with the descrip-tion of two new genera and six new species. Zootaxa 1603: 1–76.
Shen S.-C. & Wu K.-Y. (1995) Phylogenetic study of Balistoid fishes based on intestinal coiling pattern. Acta ZoologicaTaiwanica, 6, 41–56.
Simon, C. (1983) A new coding procedure for morphometric data with an example from periodical cicada wing veins. In:Felsenstein, J. (Ed.), Numerical Taxonomy. Springer, Berlin, 378–383.
Sparks, J.S. (2003) Molecular phylogeny and biogeography of the Malagasy and South Asian cichlids (Teleostei: Perci-formes: Cichlidae). Molecular Phylogenetics and Evolution, 30, 599–614.
Stawikowski, R. & Werner, U. (1998) Die Buntbarsche Amerikas. Bd. I. Eugen Ulmer, Stuttgart, 540 pp.Stiassny, M.L.J. (1991) Phylogenetic intrarelationships of the family Cichlidae. In: Keenleyside, M.H.A. (Ed.), Cichlid
Fishes — Behavior, Ecology and Evolution. Chapman & Hall, London, 1–31.Swofford, D.L. (1998) PAUP*, Phylogenetic Analysis Using Parsimony (*and other methods). Version 4. Sinauer, Sun-
derland.Takahashi T. (2003) Systematics of Xenotilapia Boulenger, 1899 (Perciformes, Cichlidae) from Lake Tanganyika,
Africa. Ichthyological Research, 50, 36–47.Taylor, W.R. & van Dyke, G.C. (1985) Revised procedures for staining and clearing small fishes and other vertebrates for
bone and cartilage study. Cybium, 9, 107–119. Thompson, K.W. (1979) Cytotaxonomy of 41 species of Neotropical Cichlidae. Copeia, 1979, 679–691.Vaillant, L. & Pellegrin, J. (1902) Cichlidés nouveaux de l’Amérique centrale. Bulletin du Muséum Nationa d’Histoire
Naturelle, 8, 84–88.Velasco, F.T. (2001) Cutter’s cichlid, not! A review of the literature concerning a colourful and possibly undescribed
Archocentrus from northern Honduras. Buntbarsche Bulletin, 2001, 206.Wiens, J.J. & Hollingsworth, B.D. (2000) War of the Iguanas: conflicting molecular and morphological phylogenies and
long-branch attraction in iguanid lizards. Systematic Biology, 49, 143–159.Yamaoka K. (1982) Morphology and feeding behavior of five species of genus Petrochromis (Teleostei, Cichlidae).
Physiology and Ecology, Japan, 19, 57–75.
TERM OF USEThis pdf is provided by Magnolia Press for private/research use. Commercial sale or deposition in a public library or website site is prohibited.














![[Citogenetic characterization of the tropical freshwater fish Parachromis managuensis (Pisces: Cichlidae)]](https://img.pdfslide.net/doc/110x75/635a1f110f31679e830c63f7/citogenetic-characterization-of-the-tropical-freshwater-fish-parachromis-managuensis.jpg)














