Embed Size (px)
Citation preview

Journal of Experimental Botany, Vol. 57, No. 14, pp. 3781–3791, 2006
doi:10.1093/jxb/erl191 Advance Access publication 17 October, 2006
RESEARCH PAPER
Respiratory Burst Oxidase Homologue A of barleycontributes to penetration by the powdery mildew fungusBlumeria graminis f. sp. hordei
Marco Trujillo1,*, Lothar Altschmied2, Patrick Schweizer2, Karl-Heinz Kogel1 and Ralph Huckelhoven1,y
1 Research Centre for BioSystems, Land Use and Nutrition, Institute of Phytopathology and Applied Zoology,Justus-Liebig-University of Giessen, Heinrich-Buff Ring 26–32, D-35392 Giessen, Germany2 The Institute of Plant Genetics and Crop Plant Research, D-06466 Gatersleben, Germany
Received 24 May 2006; Accepted 25 July 2006
Abstract
Reactive oxygen intermediates (ROI) are closely related
to defence reactions of plants against pathogens.
A prominent role in the production of ROI has been
attributed to the plant respiratory burst oxidase homo-
logues (RBOH) of the human phagocyte GP91(phox). A
barley RBOH, which encodes a putative super-
oxide (O��2 ) producing NADPH oxidase, is described
here. Histochemical analysis of the barley-Blumeria
graminis f. sp. hordei (Bgh) interaction showed that O��2
is produced locally at the site of penetration. In
contrast, hydrogen peroxide (H2O2) is produced in
non-penetrated cell wall appositions. A barley RBOHA
cDNA was isolated and a minor induction of expression
of RBOHA was observed during the interactions of
barley with Bgh. Transient RNA interference-mediated
gene silencing of HvRBOHA during the penetration
process of Bgh led to an increase of basal penetration
resistance. The results support a potential role of
HvRBOHA in cellular accessibility to Blumeria graminis.
Key words: Blumeria graminis, cell wall softening, hydrogen
peroxide, HvRBOH, penetration, resistance, superoxide (O��2 ),
susceptibility.
Introduction
Since Doke (1983) demonstrated that superoxide (O��2 ) was
generated during an incompatible plant–microbe interac-tion, a plethora of roles have been attributed to reactiveoxygen intermediates (ROI). The NADPH oxidizing
phagocyte respiratory burst oxidase homologues (RBOHs)were proposed to be a source of O��
2 and indirectly ofproximate ROI such as hydrogen peroxide (H2O2). In recentyears, several studies have provided ample genetic evidencesupporting RBOHs as a source for ROI production (Torreset al., 2002, 2005; Foreman et al., 2003; Kwak et al., 2003;Sagi et al., 2004).
Several RBOH genes have been isolated from differentplant species (Groom et al., 1996; Torres et al., 1998;Amicucci et al., 1999; Simon-Plas et al., 2002; Yoshiokaet al., 2003). RBOHs are partially similar to theGP91(phox) subunit of the mammalian respiratory burstNADPH oxidase found in phagocytes. GP91(phox) isresponsible for the production of O��
2 , which acts as asignal leading to protease activation (Reeves et al., 2002).Mutations in the catalytic subunit lead to chronicgranulomatous disease in humans, a disorder of the im-mune system. By contrast with the mammalian respiratoryburst oxidase, plant RBOHs possess an N-terminal extensionof approximately 300 amino acids. This additional domainincludes in most RBOHs two EF-hand motifs for potentialCa2+ regulation (Keller et al., 1998).
Both superoxide and hydrogen peroxide are producedduring pathogenesis. Superoxide can be converted intoH2O2, either spontaneously or by the superoxide dismu-tase. Although both oxygen species are moderately reactive,in the presence of transition metal ions as Fe2+ and Cu+,H2O2 is converted into the extremely reactive free hydroxylradical (OH�) via the Fenton reaction.
O��2 and H2O2 have been proposed to act as signals. Kwak
et al. (2003) showed that Arabidopsis thaliana AtRBOHDand AtRBOHF are necessary for H2O2 production, which is
* Present address: Sainsbury Laboratory, John Innes Centre, Norwich NR4 7UH, UK.y To whom correspondence should be addressed. E-mail: [email protected]
ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

a second messenger in the abscisic acid signal transductionpathway in stomatal closure. AtRBOHD is also responsiblefor ROI production to maintain elevated levels of Ca2+ atthe tip of root hairs during root hair growth (Foreman et al.,2003; Carol et al., 2005). Concomitantly, RBOH knock-downs in tomato display growth anomalies accompaniedby loss of tissue-specific regulation of genes related todevelopment (Sagi et al., 2004). Liszkay and colleagues(Liszkay et al., 2004) have proposed a more direct role of thehydroxyl radical in cell wall softening during cell expansion.Downstream responses induced by ROI were also shownfor activation of OXI1, an Arabidopsis serine/threoninekinase (Rentel et al., 2004) and a MAPK-induced cell deathin Nicotiana benthamiana (Yoshioka et al., 2003).
In barley (Hordeum vulgare), O��2 and H2O2 are produced
in different spatiotemporal patterns during host as well asnon-host interaction with the grass powdery mildew fungus(Blumeria graminis ff. spp.). O��
2 is generated at sitesof fungal penetration, during establishment of the fungalhaustorium, and in cells neighbouring hypersensitive celldeath reactions (HR). By contrast, H2O2 is produced in cellwall appositions during penetration resistance and through-out the cell during HR (Huckelhoven and Kogel, 2003;Trujillo et al., 2004).
In Arabidopsis, atrbohf mutants show increased HRduring the interaction with Hyaloperonospora parasitica,while mutation of atrbohd eliminates the majority of H2O2
(Torres et al., 2002). Interestingly, no difference wasobserved in the resistance of atrbohf or atrobohd toPseudomonas syringa pv. tomato. Knock-down of twoNADPH oxidase genes known in potato during theincompatible interaction with Phytophthora infestans ledto partially induced susceptibility (Yoshioka et al., 2003).
Regulation of plant RBOHs seem to involve both pre-and post-transcriptional elements. RBOH activity isstimulated by Ca2+ (Sagi and Fluhr, 2001) and also seemsto be triggered by fungal elicitors like cryptogein inNicotiana tabacum (Allan and Fluhr, 1997), hyphal wall com-ponents, arachidonic acid, and salicylic acid in potato tubers(Yoshioka et al., 2003). The catalytic subunit GP91(phox)of the multicomplex enzyme in mammals is regulated inpart by the small GTP-binding protein RAC (Babior,1999). In plants, there is evidence which suggests thata regulatory function of RAC is conserved (Kawasaki etal., 1999; Morel et al., 2004). Plant homologues of theregulatory subunits of mammalian oxidase p22, p47 or p67have not been found and oxidase activity can beindependent of activating proteins (Sagi and Fluhr, 2001).
The aim of this work was to investigate the role of aRBOH in grasses by analysing the barley–Blumeriagraminis f. sp. hordei (Bgh) interaction. A barley NADPHoxidase homologue, designated HvRBOHA (Hordeumvulgare respiratory burst oxidase homologue A) wasisolated. Transient knock-down via RNA interferenceusing dsRNA was performed to assess its role in plant
resistance. In addition, the expression profile of HvRBOHAwas analysed during the interaction with Bgh and O��
2
production using histochemical detection.
Materials and methods
Plants, pathogens and inoculation
Barley line Ingrid was grown in a growth chamber at 18 �C with60% humidity and a photoperiod of 16 h (60 lmol m�2 s�1 photonflux density). Inoculation was performed by air current dispersionin an inoculation tower. Primary leaves were inoculated 7 d aftergermination with Blumeria graminis f. sp. hordei isolate A6(BghA6). Inoculation density for RNA extraction was 50 conidiamm�2 and for the RNA interference experiments 100 conidia mm�2.Fungi were produced under the same conditions. BghA6 was grownon barley line Golden Promise. Primary leaves were harvested at 15,24, and 48 h after inoculation (hai) and shock frozen with liquidnitrogen. Mock inoculation and harvesting was carried out in parallelfor each time point.
Cytological analysis
The solution used for in situ detection of O��2 contained 1 mg NBT
ml�1 in 10 nM NaN3 and 10 mM potassium phosphate buffer pH7.8. For O��
2 production analysis, 1 ml of the solution was injectedinto primary leaves 25 min before fixation as described byHuckelhoven and Kogel (1998). Leaves were bleached and fungalstructures were subsequently stained by acidic ink (Huckelhoven andKogel, 1998). Only short epidermal cells were utilized for analysis(type A and B; Koga et al., 1990).
Leaves used for H2O2 detection were transiently transformed withDsRed using a protocol from Schweizer et al. (2000) and modifiedaccording to Schultheiss et al. (2002). The detection of H2O2 wascarried out 20 hai by immersing leaf segments in 10 ml of a loadingbuffer (KCl 30 mM, MES-KOH 10 mM, pH 6.15) with 4 ll of a 100mM 2#,7#-dichlorofluorescein diacetate stock in DMSO (H2-DCF-DA; Invitrogen, Karlsruhe, Germany). After 15 min of staining,leaves were washed with fresh loading buffer and fluorescenceintensity was evaluated in a time frame of 30 min by confocal laserscanning microscopy (Leica TCS SP2, Leica Microsystems,Bensheim Germany). Values represent the average relative fluores-cence on an arbitrary scale of a minimum of 30 interactions in fourindependent experiments. H2-DCF-DA was excited at 488 nm laserline and detected at 500–535 nm; DsRed was excited at 543 nm laserline and detected at 580–650 nm.
RNA extraction and reverse transcription
Tissue was ground into a fine powder in liquid nitrogen. Total RNAwas then extracted and integrity was assessed by examining rRNAbands after electrophoresis on a standard gel. DNA contaminationwas removed using RNase-free (Fermentas, St Leon-Rot, Germany)with 1 U DNase I lg�1 sample RNA, incubated at 37 �C for 30 minand inactivated by adding 1 ll EDTA (25 mM) and incubating10 min at 70 �C.
One lg of RNA was reverse transcribed using the BioRad iScriptcDNA synthesis kit (BioRad Laboratories Inc., California, USA)according to the manufacturer’s instructions. The cDNA was diluted5-fold and used as the template for PCR amplifications.
Isolation of HvRBOHA partial clones and fragments used for
dsRNA synthesis
cDNA was used to obtain partial clones of 5# and 3#-fragments ofHvRBOHA and for dsRNA synthesis of a 5#-fragment (nucleotide
3782 Trujillo et al.
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

position 49–251, GenBank accession number AJ871131), a predictedtransmembrane regions 3 to 4 fragment (nucleotide position 1452–1819,GenBank accession number AJ871131) and a 3#-fragment (nucleotideposition 1135–1434, GenBank accession number AJ251717).
Primers flanking the HvRBOHA gene were designed usingsequence information obtained by sequencing bacterial artificialchromosome (BAC)-clones with the following primers: fwd 5#-GAATTCCATAAGGAGCATTTC-3#, rev1 5#-TGAGCTCCTGC-GAGAACT-3# and rev2 5#-GACAATGGTGTCGTCCTCCT-3#.Flanking primers were employed in combination with primersderived from OsRBOHA (GenBank accession number AF015302)for amplification of the 5# and 3# overlapping fragments containingthe complete coding sequence ofHvRBOHA. Primers for 5# fragment:fwd 5#-AAATCGTCAACCCAGTTTGG-3#, rev 5#-CAGAAGGCA-TTGAAGCCAGT-3#. Primers for 3# fragment: fwd 5#-AACACCAT-TACTTGGTTGCG-3# and rev 5#-ACATACAGCGCAATCCTGTG-3#. PCR products were isolated after electrophoresis, cloned intopCR 4-TOPO (Invitrogen, Karlsruhe, Germany) and sequenced(GenBank accession numbers AJ871131, AJ251717).
Fragments used for dsRNA synthesis were obtained using thefollowing primers. 5#-fragment: fwd 5#-ACTGCTCTCCCGAA-CAAGG-3#, rev 5#-GACAATGGTGTCGTCCTCCT-3#. TMR 3–4:fwd 5#-AACACCATTACTTGGTTGCG-3#, rev 5#-CAGAAGG-CATTGAAGCCAGT-3#. 3#-fragment: fwd 5#-AGCAAGGCT-CTTTTGATTG-3#, rev 5#-GAAATGCTCCTTATGGAATTC-3#.PCR products were isolated after electrophoresis, cloned intopGEM-T (Promega, Mannhein, Germany) and sequenced.
Screening of BAC-libraries
PCR screening was performed with the primers, fwd 5#-CCGAT-CAGATGTATGCTCCA-3# and rev 5#-CAGAAGGCATTGAAGC-CAGT-3#, to isolate a BAC containing genomic HvRBOHA from theBAC-library Hv_MBa DNA pools (Clemson University, http://www.genome.clemson.edu/orders/HV_MBa_Clone.html).
Quantitative real-time PCR
Quantitative real-time PCR (qRT-PCR) was performed using thePlatinum SYBR Green qPCR SuperMix UDG Kit (Invitrogen,Karlsruhe, Germany) in a Mx3000P QPCR System from Stratagene(Stratagene, California, USA). Reactions contained 10 ll of reactionmixture, 2.0 ll cDNA, and 0.5 pmol of each primer in a total volumeof 20 ll. All reactions were carried out in triplicate for each sample.Primers were designed from the target gene sequence using Primer3(Rozen and Skaletsky, 2000) software and optimized to anneal at anequal temperature of 60 �C. The following primers were generatedfor gene expression analysis (MWG Biotech, Ebersberg, Germany).Target gene 1, NADPH oxidase (GenBank accession number:AJ251717), forward 5#-CCGATCAGATGTATGCTCCA-3# andreverse 5#-CAGAAGGCATTGAAGCCAGT-3#; target gene 2,PR-1b (GenBank accession number: X74940) forward 5#-GGAC-TACGACTACGGCTCCA-3#, reverse 5#-GGCTCGTAGTTG-CAGGTGAT-3#; reference gene Ubiquitin (GenBank accessionnumber: M60175), forward 5#-ACCCTCGCCGACTACAACAT-3#, reverse 5#-CAGTAGTGGCGGTCGAAGTG-3#; reference gene18s (GenBank accession number: AY552749), forward 5#-CGGCGGAGTCCTATAAGC-3#, reverse 5#-GCGAGCACC-GACCTACTC-3#.
PCR for quantification started with 2 min incubation at 50 �C and3 min denaturation at 95 �C followed by 50 cycles of 95 �C for 30 s,60 �C for 30 s, 72 �C for 20 s (1. fluorescence acquiring) and 82 �Cfor 15 s (2. fluorescence acquiring). Subsequently, a melting curvewas performed in which temperature was incremented from 50 �Cto 99 �C in one degree steps each held for 10 s.
Identity of qRT-PCR products was confirmed by the observationof a single melting peak. In addition, products were separated byagarose-gel electrophoresis to verify product size, and sequenced.Standard curves and threshold values were calculated with a 5-folddilution series (total of five dilutions) of template pools using theStratagene software (Stratagene, California, USA). Relative quanti-fication of fold expression changes were calculated according toPfaffl (2001).
Transient transformation assay and RNAi
Transient transformation protocol to assess gene function in thebarley–powdery mildew interaction via RNA interference (RNAi)through a biolistic approach was taken from Schweizer et al. (2000)and modified according to Schultheiss et al. (2002). Plants weregrown under the same conditions as previously described (Schultheisset al., 2002).
For RNAi, dsRNA of the three different HvRBOHA fragmentswere synthesized and leaf segments bombarded with the coatedparticles 4 h before inoculation with Bgh race A6. Inoculationdensity was 100 conidia mm�2. Interaction outcome (i.e. penetrationor effective cell wall apposition) was analysed 40 h after inoculationby fluorescence and light microscopy. Transformed GFP expressingcells were identified under blue light excitation. Surface structures ofBgh were detected by light microscopy or by fluorescence staining ofthe fungus with 0.3% calcofluor (w/v in 50 mM TRIS pH 9.0).
Relative penetration efficiency (PE), referring to average controlPE was used as a measure for susceptibility of cells that werebombarded with one of the three HvRBOHA-dsRNAs and comparedwith those bombarded with control TR-dsRNA (human thyroidreceptor-dsRNA). PE data were obtained from a set of a minimum offour experiments each consisting of at least 100 interaction sites. PEwas calculated for each experiment as the number of penetrated cellsdivided by the total number of attacked cells (PE) multiplied by 100.Relative PE was calculated using the formula: [(PE-HvRBOHA-dsRNA–PE-TR-dsRNA)/PE–TR-dsRNA]3100.
Results
Cytological analysis of ROI production
A microscopical analysis of the interaction between thehost barley (cv. Ingrid) and virulent Bgh (race A6) wasperformed.
Superoxide production was detected employing the in situnitroblue tetrazolium (NBT) reduction assay, considered asspecific for O��
2 (Doke, 1983; Fryer et al., 2002). The firstROI to be detected at sites of attempted penetration was O��
2 .At first, O��
2 was detected locally at the plasma membraneadjacent to successful penetration attempts in the early stageof the interaction, approximately 16 h after inoculation(hai; Fig. 1A). Importantly, no O��
2 was detected in non-penetrated cell wall appositions. By contrast, H2O2
accumulation detected with 2#,7#-dichlorofluorescein diac-etate (H2-DCF-DA), was primarily produced in non-penetrated cell wall appositions, starting at approximately18 hai (Fig. 1B). Staining patterns observed with H2-DCF-DA were very similar to those observed with theH2O2-specific dye 3,3-diaminobenzidine (Thordal-Christensen et al., 1997; Huckelhoven et al., 1999; Trujilloet al., 2004).
Barley respiratory burst oxidase homologue 3783
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Isolation of HvRBOHA
To isolate HvRBOHA, a reverse transcription (RT) step wasperformed using total RNA of barley (Hordeum vulgare)cv. Ingrid as the template followed by PCR. Degenerateprimers were designed based on the conserved C-terminalregion of the human GP91(phox) subunit (Royer-Pokoraet al., 1986) and of rice OsRBOHA (Groom et al., 1996;Huckelhoven et al., 2001). Additional primers for upstreamannealing were designed based on the rice OsRBOHAgenomic sequence (GenBank accession number AP003560).This allowed the isolation of overlapping cDNA fragmentscovering a total of 2215 base pairs of the 3# part of a barleyputative NADPH oxidase. However, the 5# end of thecDNA could not be isolated via RT-PCR with degenerateprimers. Therefore, the barley bacterial artificial chromo-some (BAC)-library HV_MBa of the Clemson University(http://www.genome.clemson.edu/orders/HV_MBa_Clone.html) was screened. PCR-based screening of DNA poolsand sub-pools was performed using a primer combinationthat generated an amplicon of 190 bp of a conserved regionof RBOH. Three independent BAC-clones were isolated(324E18, 450H05, 615H10) carrying the same RBOHgene. Primers were designed, based on BAC-sequencesobtained by primer walking to obtain the missing 5#-endportion. Based on the obtained sequence data, primers tocover the total open reading frame were designed. TwocDNA fragments were isolated, one comprising the 5#-endregion and the other the 3#-end region of the mRNA.Fragments share a 329 bp overlapping region with 100%identity, demonstrating that they correspond to the samegene. The complete coding sequence composed of bothfragments results in an open reading frame of 2889 bp.Extended 5# sequence possessed stop codons in all readingframes, confirming the predicted start codon. Hence, the
first full-length RBOH cDNA sequence information forbarley (GenBank accession number AM265370) is pro-vided here. The nucleotide sequence showed an estimated87% identity to a rice cDNA clone, which will be referredto as OsRBOH, and 84% identity to OsRBOHA usingBLASTN (GenBank accession numbers AK120905,AF015302 partial mRNA; Altschul et al., 1997; Kelleret al., 1998). The nearest Arabidopsis orthologue isAtRBOHF with a similarity of 81% (GenBank accessionnumber AB008111; Torres et al., 1998).
The deduced amino acid sequence of the RBOH ORF ispredicted to encode 963 amino acids and shows anextended N-terminal region not present in the humanGP91(phox) sequence (GenBank accession numberNM_000397). The C-terminal region of the deduced aminoacid sequence shares a 47% similarity to the GP91(phox)of human phagocytes, but domains relevant for the activityof the catalytic subunit are conserved (Fig. 2). Bindingsites for NADPH- and FAD of the human NADPH oxidase(Yoshida et al., 1998; Vignais, 2002) are conserved in thebarley homologue. Although plants lack clear P47phox
homologues, residues involved in the binding of P47phox
(De Leo et al., 1996) are partially conserved as shown forthe barley and the Arabidopsis sequences (Fig. 2). Sixhydrophobic stretches in a similar arrangement as intransmembrane regions of GP91(phox) were predictedusing the dense alignment surface program (Cserzo et al.,1997), implying analogous topological arrangement (datanot shown). In addition, haem-ligating histidines (Bibers-tine-Kinkade et al., 2001) of the human NADPH oxidaseare conserved in barley.
Two putative Ca2+-binding EF-hand motifs precedingthe first residue aligned with GP91(phox), are present inthe N-terminus in barley (Fig. 2). This is a typical feature
Fig. 1. Micrographs of interaction sites with histochemical detection of reactive oxygen intermediates. Primary leaves were treated with NBT or H2-DCF-DA and analysed for precipitation of dark-blue formazan representative of O��
2 production or fluorescence indicating H2O2 accumulation. (A) Penetratedbarley epidermal cell, in which Blumeria graminis f. sp. hordei (Bgh) established a haustorium initial (arrow), and localized formazan precipitation isvisible (arrowhead). Fungus is stained with blue ink (Huckelhoven and Kogel, 1998). (B) Overlay projection of a DsRed-transformed barley epidermal cell20 hai with Bgh and stained with H2-DCF-DA for H2O2 detection. The cell displays a non-penetrated papilla (arrow, green fluorescence of H2-DCF-DA)with cytoplasm aggregation (red) surrounding the non-penetrated papilla underneath the appressorial germ tube of the fungus (arrowheads). Insetsrepresent an overlay of the red and green fluorescence channels and of the transmission channel, respectively. Scale bar, 10 lm.
3784 Trujillo et al.
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

of plant RBOHs and is also described for rice andArabidopsis (Keller et al., 1998; Torres et al., 1998).
The deduced amino acid sequence of the isolated RBOHgene was compared with divergent RBOHs to constructa phylogenetic tree using ClustalW series program (Chennaet al., 2003). The deduced amino acid sequence of a riceRBOH (GenBank accession number AK120905) ispredicted to be related closest to HvRBOHA and displays91% similarity (Fig. 3). OsRBOHA is the second closesthomologue in the same subgroup. The barley NADPHoxidase will be referred to as HvRBOHA. AtRBOHF is theclosest Arabidopsis homologue with 79% similarity.
Expression analysis of HvRBOHA
A check was made for epidermal expression of HvRBOHA,because this is the only tissue with direct contact to Bgh.Similar to that described previously (Schultheiss et al.,2003), expression was compared in epidermal peels of barleyinoculated with Bgh and the rest of the leaves at 24 h afterinoculation (hai). Expression of HvRBOHA was comparedwith that of actin, epidermis-expressed oxalate oxidase-likeprotein (HvGLP4), and ribulose bisphosphate carboxylasefor mesophyll contamination by semi-quantitative RT-PCR.When compared to strong signals of germin-like protein 4and ribulose bisphosphate carboxylase in the epidermis andthe rest of the leaves, respectively, HvRBOHA appearedequally expressed in both epidermis and the rest of leaves. Itwas further examined if HvRBOHA expression was inducedby the interaction of barley with Bgh. For this purpose, totalRNA was isolated from epidermal peels at 0, 15, and 24 haiand after mock inoculation and subjected to first strandcDNA synthesis. No HvRBOHA signal could be detected bynorthern-blot analysis. Therefore, relative gene expressionwas determined by quantitative real-time PCR (qRT-PCR).Low abundance of transcripts was confirmed by the thresholdvalues obtained by qRT-PCR. On average, nine additionalcycles of amplification were necessary to obtain the thresholdvalue of HvRBOHA (target) compared to Ubiquitin used asa constitutively expressed reference (Schultheiss et al.,2002). HvRBOHA expression increased by a maximum of2.7-fold during the interaction of barley with Bgh at 15 hai.To verify the induction of defence responses, expression ofthe pathogenesis-related-1b (PR-1b) gene was also analysed.A strong increase of 36-fold of transcript abundance wasdetected after inoculation of barley with Bgh at 15 hai (Table1). In independent experiments, a similar expression patternof HvRBOHA was detected. In addition, expression level ofHvRBOHA decreased at 48 hai to the same levels as thecontrol. Semi-quantitative reverse transcription-PCR resultedin similar expression patterns as those obtained by qRT-PCRfor HvRBOHA and PR-1b. Expression patterns during thenon-host interaction of barley with Blumeria graminis f. sp.tritici were similar. Epidermal expression of HvRBOHA isthus slightly and transiently induced by Bgh.
Functional analysis of HvRBOHA
To analyse the function of HvRBOHA during the interactionof barley with Bgh, a transiently induced gene-silencingassay was performed via RNAi in the barley cultivar Ingrid(Schweizer et al., 2000; Schultheiss et al., 2002). Sub-sequently, dsRNA of the target gene together with GFPwas introduced in an expression vector as reporter gene intoepidermal cells via a biolistic approach. Interaction out-come was evaluated 40 h after inoculation and comparedwith control cells containing heterologous human thyroidreceptor (TR) dsRNA. To determine the effect of HvRBOHAon penetration efficiency, dsRNA of three different frag-ments including a 5#-fragment (nucleotide position 49–251, GenBank accession number AJ871131), a fragmentcontaining the predicted transmembrane stretches 3 and 4(nucleotide position 1452–1819, GenBank accessionnumber AJ871131), and a 3#-fragment (GenBank acces-sion number AJ251717) was used. The average penetrationefficiency (PE) of the fungus in the controls (cellstransformed with TR-dsRNA and GFP) was 48% over allexperiments. Fragments corresponding to the highly con-served sequences of the TMR 3–4 and 3#-region induceda similar PE decrease of 36% of Bgh on a susceptiblebarley cultivar (Fig. 4). The 5#-region is the most variablebetween RBOHs, thus the most specific of all fragmentsused for RNAi. Cells transformed with dsRNA from the5#-stretch, showed 28% penetration on average, while 59%of interaction sites of control cells were penetrated. Thistranslates in a highly significant (Student’s t-test, P <0.001)decrease of 52% of the relative penetration efficiency ofthe fungus (Fig. 4). Similar results were obtained wheninstead of Ingrid, the also susceptible barley cultivar Pallaswas transformed with dsRNA from the 3#-stretch.Together, knock-down of HvRBOHA reduced susceptibil-ity to penetration by Bgh.
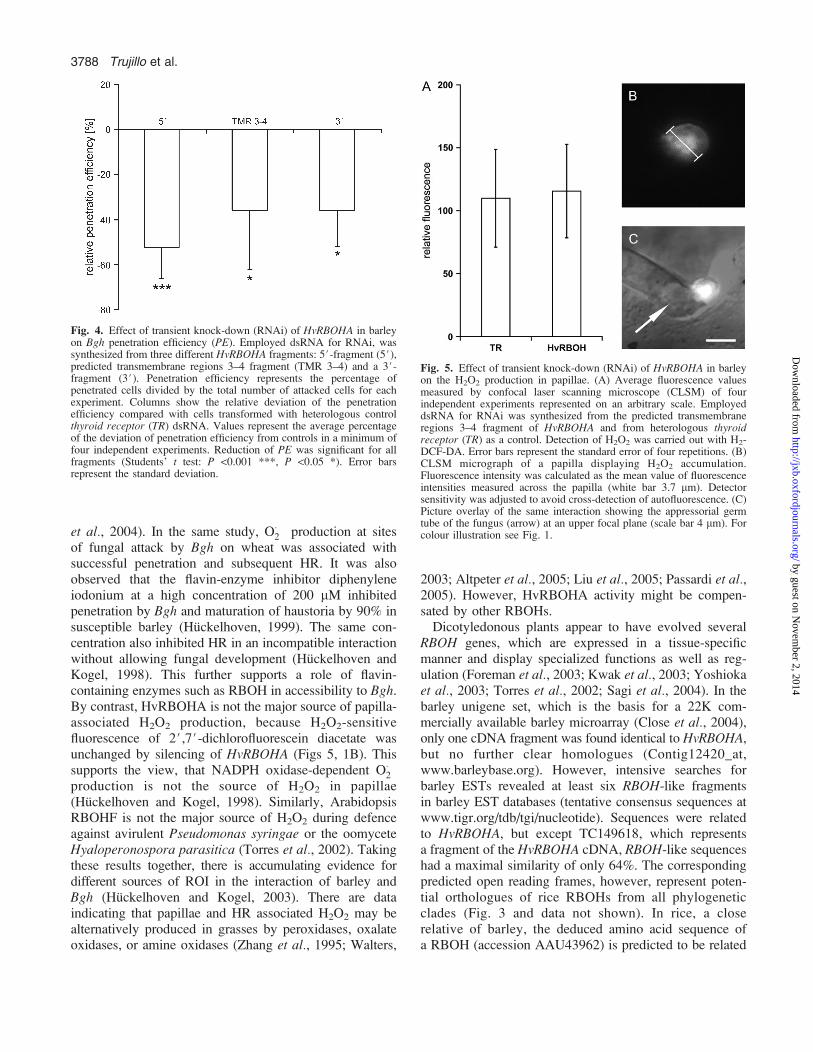
H2O2 accumulation was quantified after transient knock-down of HvRBOHA by measuring H2-DCF-DA fluores-cence intensities of papillae at 20 hai by confocal laserscanning microscopy. However, H2-DCF-DA fluorescencewas unchanged in Bgh-attacked cells in which HvRBOHAwas knocked down (Fig. 5). Hence, the abundance ofHvRBOHA does not seem to limit H2O2 in cell wallappositions produced in response to Bgh.
Discussion
Plant RBOHs have been shown to be a source for ROIproduction and play a pivotal role in manifold processes.Our purpose was to investigate a possible role of a barleyRBOH in the interaction of barley with the powderymildew fungus.
An RNAi method was employed for transient silencing ofthe barley NADPH oxidase homologue HvRBOHA.Knock-down of HvRBOHA led to a decrease of fungal
Barley respiratory burst oxidase homologue 3785
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

3786 Trujillo et al.
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

penetration, which was independent of the cDNA fragmentused for the generation of dsRNA. Nevertheless, the degreeof resistance achieved varied dependent on the dsRNA used.Fragments spanning the predicted transmembrane regions3–4 and the 3# stretch are highly conserved in other RBOHsand induced an increase in penetration resistance of 36%.Sequence conservation implies homology to any otherRBOH possibly present in barley and their concomitantdown-regulation. Maximum resistance induction wasachieved with the 5# stretch (52%). This region showsthe highest sequence variability between RBOH indifferent species. Thus, the 5# stretch might be the mostspecific for silencing of HvRBOHA. Accordingly, specificsilencing of highly similar RAC-gene family members inrice was achieved by using gene-specific inverted repeatconstructs (Miki et al., 2005). On the other hand, it is alsopossible that different dsRNA fragments had differentsuitability as substrate for RNAi machinery (Yiu et al.,2004). The latter explanation is supported by the fact thatno barley EST could be detected out of more than 300 000public sequences, showing more than 64% sequence identityto HvRBOHA (tentative consensus sequence: TC147482,
www.tigr.org/tdb/tgi/nucleotide). Together, this suggeststhat RNAi was specific for HvRBOHA.
In the barley–Bgh interaction, H2O2 has been related toresistance (Thordal-Christensen et al., 1997; Huckelhovenet al., 1999). Nevertheless, O��
2 production occurs con-comitant to penetration during the early interaction stages(Fig. 1A) and around growing fungal haustoria (Huck-elhoven and Kogel, 1998; Trujillo et al., 2004). This hintsat the possibility that O��
2 either contributes to penetrationor acts as a signal supporting biotrophy. In addition,distinct spatiotemporal production patterns of H2O2 andO��
2 , respectively, suggest different roles as well asdifferent sources of ROI (Allan and Fluhr, 1997; Huck-elhoven and Kogel, 2003; Trujillo et al., 2004). Superox-ide, produced by RBOH and the hydroxyl radicals that canbe generated by the Fenton reaction, have been shown tobe involved in cell wall polymer breakdown during cellgrowth (Foreman et al., 2003; Liszkay et al., 2004). Thesame process leading to polymer breakdown may provokelocal cell wall softening. This could be induced by theinvading fungus to promote penetration by triggeringRBOH activity. Hence, knock-down of HvRBOHA mightlead to decreased levels of hydroxyl radicals and enhancepenetration resistance by preventing host cell wallsoftening. In this context it is interesting that an extra-cellular germin-like protein GLP4 with superoxide dismutaseactivity is involved in background penetration resistance ofbarley to Bgh (Christensen et al., 2004). Simultaneoushistochemical detection of O��
2 and H2O2 during theinteraction of barley with Blumeria graminis f. sp. triticishowed that non-penetrated papillae contained H2O2
whereas O��2 could almost never be detected (Trujillo
Fig. 2. Alignment of deduced amino acid sequences of the barley (HvRBOHA), Arabidopsis (AtRBOHF), and human NADPH oxidase subunitGP91(phox) homologues. Conserved amino acid residues are shaded in grey. EF-hand motifs are encased and highlighted by bars. (A–C) Residuesinvolved in p47phox binding in mammals. Sites identified for FAD- and NADPH-binding in GP91(phox) FAD-binding I and II, NADPH units IIIpyrophosphate, IV ribose, V adenine, and VI nicotinamide. GP91(phox) haem-ligating histidines, asparagines involved in N-glycosylation as well asproline415 and aspartate500, involved in chronic granulomatous disease are denoted by white letters with black background. Dashes represent gapsintroduced to optimize alignment.
Fig. 3. Tentative phylogenetic tree of plant respiratory burst oxidasehomologues (RBOH). Tree was constructed by alignment of deducedamino acid sequences using CLUSTALW (http://www.ebi.ac.uk/clustalw/, Chenna et al., 2003). Estimated branch lengths given afterabbreviation are proportional to the amount of inferred evolutionarychange. Sequences were obtained from GenBank. Abbreviations: barley(Hv), rice (Os), Arabidopsis (At) GenBank accession numbers:AtRBOHA (At5g07390), AtRBOHB (At1g09090), AtRBOHC(At5g51060), AtRBOHD (At5g47910), AtRBOHE (At1g19230), AtR-BOHF (At1g64060), AtRBOHG (At4g25090), AtRBOHH(At5g60010), AtRBOHI (At4g11230), AtRBOHJ (At3g45810) asreported by Torres and Dangl (2005). For rice accession numbers, seefigure.
Table 1. Relative expression levels of barley RBOHA andPR1-b in response to inoculation with virulent Bgh
Gene Fold inductiona
15 hai 24 hai
HvRBOHA 2.7 2.5HvPR1-b 36.5 16.2
a Total RNA was extracted from 8-d-old barley plants of cv. Ingridafter inoculation with Bgh and mock inoculation. RNA was subsequentlysubjected to reverse-transcription, followed by real-time PCR analysis.Expression of the target genes HvRBOHA and PR1-b is displayedrelative to the expression of the reference gene Ubiquitin. Relativechange of expression was calculated according to Pfaffl (2001). Bothtechnical and biological repetitions of the experiment led to similarresults.
Barley respiratory burst oxidase homologue 3787
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from
et al., 2004). In the same study, O��2 production at sites
of fungal attack by Bgh on wheat was associated withsuccessful penetration and subsequent HR. It was alsoobserved that the flavin-enzyme inhibitor diphenyleneiodonium at a high concentration of 200 lM inhibitedpenetration by Bgh and maturation of haustoria by 90% insusceptible barley (Huckelhoven, 1999). The same con-centration also inhibited HR in an incompatible interactionwithout allowing fungal development (Huckelhoven andKogel, 1998). This further supports a role of flavin-containing enzymes such as RBOH in accessibility to Bgh.By contrast, HvRBOHA is not the major source of papilla-associated H2O2 production, because H2O2-sensitivefluorescence of 2#,7#-dichlorofluorescein diacetate wasunchanged by silencing of HvRBOHA (Figs 5, 1B). Thissupports the view, that NADPH oxidase-dependent O��
2
production is not the source of H2O2 in papillae(Huckelhoven and Kogel, 1998). Similarly, ArabidopsisRBOHF is not the major source of H2O2 during defenceagainst avirulent Pseudomonas syringae or the oomyceteHyaloperonospora parasitica (Torres et al., 2002). Takingthese results together, there is accumulating evidence fordifferent sources of ROI in the interaction of barley andBgh (Huckelhoven and Kogel, 2003). There are dataindicating that papillae and HR associated H2O2 may bealternatively produced in grasses by peroxidases, oxalateoxidases, or amine oxidases (Zhang et al., 1995; Walters,
2003; Altpeter et al., 2005; Liu et al., 2005; Passardi et al.,2005). However, HvRBOHA activity might be compen-sated by other RBOHs.
Dicotyledonous plants appear to have evolved severalRBOH genes, which are expressed in a tissue-specificmanner and display specialized functions as well as reg-ulation (Foreman et al., 2003; Kwak et al., 2003; Yoshiokaet al., 2003; Torres et al., 2002; Sagi et al., 2004). In thebarley unigene set, which is the basis for a 22K com-mercially available barley microarray (Close et al., 2004),only one cDNA fragment was found identical to HvRBOHA,but no further clear homologues (Contig12420_at,www.barleybase.org). However, intensive searches forbarley ESTs revealed at least six RBOH-like fragmentsin barley EST databases (tentative consensus sequences atwww.tigr.org/tdb/tgi/nucleotide). Sequences were relatedto HvRBOHA, but except TC149618, which representsa fragment of the HvRBOHA cDNA, RBOH-like sequenceshad a maximal similarity of only 64%. The correspondingpredicted open reading frames, however, represent poten-tial orthologues of rice RBOHs from all phylogeneticclades (Fig. 3 and data not shown). In rice, a closerelative of barley, the deduced amino acid sequence ofa RBOH (accession AAU43962) is predicted to be related
Fig. 5. Effect of transient knock-down (RNAi) of HvRBOHA in barleyon the H2O2 production in papillae. (A) Average fluorescence valuesmeasured by confocal laser scanning microscope (CLSM) of fourindependent experiments represented on an arbitrary scale. EmployeddsRNA for RNAi was synthesized from the predicted transmembraneregions 3–4 fragment of HvRBOHA and from heterologous thyroidreceptor (TR) as a control. Detection of H2O2 was carried out with H2-DCF-DA. Error bars represent the standard error of four repetitions. (B)CLSM micrograph of a papilla displaying H2O2 accumulation.Fluorescence intensity was calculated as the mean value of fluorescenceintensities measured across the papilla (white bar 3.7 lm). Detectorsensitivity was adjusted to avoid cross-detection of autofluorescence. (C)Picture overlay of the same interaction showing the appressorial germtube of the fungus (arrow) at an upper focal plane (scale bar 4 lm). Forcolour illustration see Fig. 1.
Fig. 4. Effect of transient knock-down (RNAi) of HvRBOHA in barleyon Bgh penetration efficiency (PE). Employed dsRNA for RNAi, wassynthesized from three different HvRBOHA fragments: 5#-fragment (5#),predicted transmembrane regions 3–4 fragment (TMR 3–4) and a 3#-fragment (3#). Penetration efficiency represents the percentage ofpenetrated cells divided by the total number of attacked cells for eachexperiment. Columns show the relative deviation of the penetrationefficiency compared with cells transformed with heterologous controlthyroid receptor (TR) dsRNA. Values represent the average percentageof the deviation of penetration efficiency from controls in a minimum offour independent experiments. Reduction of PE was significant for allfragments (Students’ t test: P <0.001 ***, P <0.05 *). Error barsrepresent the standard deviation.
3788 Trujillo et al.
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

closest to HvRBOHA. HvRBOHA and OsRBOH_AAU43962 constitute a subgroup of the phylogenetic treetogether with OsRBOHA (Fig. 3). Except HvRBOHA,none of the predicted barley RBOHs grouped to the sameclade, supporting its annotation as a potential orthologue ofOsRBOHA. Together, diversification of RBOHs observedin dicotyledonous Arabidopsis, seems to have also takenplace in monocotyledonous plants such as rice and probablyalso in barley.
Mutation of AtRBOHF and/or AtRBOHD leads to anenhanced cell death after inoculation with H. parasitica.This effect relies on enhanced HR (Torres et al., 2002). Ithas been shown previously, that O��
2 is produced adjacentto HR and RBOHs have an HR-restricting function inwild-type plants (Jabs et al., 1996; Trujillo et al., 2004;Torres et al., 2005). By contrast, silencing of Nicotianabenthamiana RBOHA and RBOHB led to inducedsusceptibility to Phytophthora infestans and less HR inan incompatible interaction (Yoshioka et al., 2003).Therefore, suppression of RBOH-dependent O��
2 pro-duction may result in distinct modulations of resistance,probably depending on fungus-specific infection strategyand/or signalling pathways. In this investigation effects onHR could not be detected, because transiently inducedgene silencing was performed on a single cell level withGFP as a marker to detect transformed cells. Nevertheless,the effect on reduced penetration may well rely ona missing signal necessary for the establishment ofbiotrophy, which would impede fungal growth in theinitial stage. Genetic evidence indicates that RBOH isinvolved in restriction of pathogen-induced cell death inArabidopsis (Torres et al., 2005), which is in accordancewith the assumption that HvRBOHA may have a functionin the survival of barley cells in a compatible interactionwith Bgh. Alternatively, lack of HvRBOHA and itspotential role in defence might be over-compensated byalternative host defence strategies.
There are also data supporting a function of RBOH in theinduction of cell death. Barley plants expressing a constitu-tively activated RAC1 GTPase (Schultheiss et al., 2003), thepresumptive rice orthologue of which was shown to induceRBOH dependent cell death (Kawasaki et al., 1999), aremore susceptible to penetration by Bgh and, at the sametime, show more hypersensitive cell death reactions thanwild type (I Priyadarshini, G Hensel, J Kumlehn, and RHuckelhoven, unpublished results). During barley race-specific response to penetration by Bgh a long-lasting O��
2
production is detected that may facilitate cell death(Huckelhoven and Kogel, 1998). Therefore, O��
2 may alsoreduce the threshold of cell death by sensitizing penetratedcells. It is plausible that O��
2 can support cell death initially,whereas after cell death has been triggered, it is required forthe restriction of lesions (Fig. 6). This suggests theinvolvement of additional specific components to determinethe outcome of O��
2 production during defence responses.
Because expression of HvRBOHA is only weaklyinduced by Bgh (this study; Huckelhoven et al., 2001;Jain et al., 2004), local O��
2 generation may be regulatedrather at the activity level, potentially after Ca2+ influxsensed by the EF-hand motifs of RBOH (Sagi and Fluhr,2001). There is indirect evidence for Ca2+ influx intosusceptible barley cells attacked by Bgh because apoplastic[Ca2+] decreases when Bgh attempts to penetrate (Felle et al.,2004). Moreover, Ca2+ is linked to susceptibility becausethe major susceptibility factor MLO binds calmodulin ina Ca2+-dependent manner to fulfil its role in accessibilityto Bgh (Kim et al., 2002). Astonishingly, little O��
2 isobserved in resistant mlo5-mutant lines at the time whenO��
2 is produced in susceptible near-isogenic MLOgenotypes during fungal penetration (Huckelhoven andKogel, 1998). Together, HvRBOHA may support fungalpenetration and/or be indispensable for the establishmentof haustoria.
Acknowledgements
This work was supported by the German Academic ExchangeService and the BASF AG.
References
Allan AC, Fluhr R. 1997. Two distinct sources of elicited reactiveoxygen species in tobacco epidermal cells. The Plant Cell 9,1559–1572.
Altpeter F, Varshney A, Abderhalden O, Douchkov D, Sautter C,Kumlehn J, Dudler R, Schweizer P. 2005. Stable expression ofa defense-related gene in wheat epidermis under transcriptionalcontrol of a novel promoter confers pathogen resistance. PlantMolecular Biology 57, 271–283.
Fig. 6. Schematic representation of the potential involvement of RBOHin pathogenesis-related O��
2 generation in the interaction of barley withBlumeria graminis f. sp. hordei. Possible functions of O��
2 produced byRBOH may include cell wall softening and support of haustoriaestablishment in compatible interactions and sensitizing cells for celldeath in incompatible interactions. Conversely, O��
2 in conjunction withHR-like cell death derived signals may be necessary for the restriction ofcell death. The model suggests the involvement of additional specificcomponents to determine the outcome of O��
2 production duringinteraction with Bgh.
Barley respiratory burst oxidase homologue 3789
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Altschul SF, Madden TL, Schaffer AA, Zhang J, Zhang Z,Miller W, Lipman DJ. 1997. Gapped BLAST and PSI-BLAST:a new generation of protein database search programs. NucleicAcids Research 25, 3389–3402.
Amicucci E, Gaschler K, Ward JM. 1999. NADPH oxidase genesfrom tomato (Lycopersicon esculentum) and curly leaf pondweed(Potamogeton crispus). Plant Biology 1, 524–528.
Babior BM. 1999. NADPH oxidase: an update. Blood 93,1464–1476.
Biberstine-Kinkade KJ, DeLeo FR, Epstein RI, LeRoy BA,Nauseef WM, Dinauer MC. 2001. Heme-ligating histidines inflavocytochrome b(558): identification of specific histidines ingp91(phox). Journal of Biological Chemistry 276, 31105–31112.
Carol RJ, Takeda S, Linstead P, Durrant MC, Kakesova H,Derbyshire P, Drea S, Zarsky V, Dolan L. 2005. A RhoGDPdissociation inhibitor spatially regulates growth in root hair cells.Nature 438, 1013–1016.
Chenna R, Sugawara H, Koike T, Lopez R, Gibson TJ,Higgins DG, Thompson JD. 2003. Multiple sequence alignmentwith the Clustal series of programs. Nucleic Acids Research 31,3497–3500.
Christensen AB, Thordal-Christensen H, Zimmermann G,Gjetting T, Lyngkjaer MF, Dudler R, Schweizer P. 2004. Thegerminlike protein GLP4 exhibits superoxide dismutase activityand is an important component of quantitative resistance in wheatand barley. Molecular Plant–Microbe Interactions 17, 109–117.
Close TJ, Wanamaker SI, Caldo RA, Turner SM, Ashlock DA,Dickerson JA, Wing RA, Muehlbauer GJ, Kleinhofs A,Wise RP. 2002. A new resource for cereal genomics: 22K BarleyGenechip comes of age. Plant Physiology 134, 960–968.
Cserzo M, Wallin E, Simon I, von Heijne G, Elofsson A. 1997.Prediction of transmembrane alpha-helices in procaryotic mem-brane proteins: the Dense Alignment Surface method. ProteinEngineering Design and Selection 10, 673–676.
De Leo FR, Ulman KV, Davis AR, Jutila KL, Quinn MT. 1996.Assembly of the human neutrophil NADPH oxidase involvesbinding of p67phox and flavocytochrome b to a commonfunctional domain in p47phox. Journal of Biological Chemistry271, 17013–17020.
Doke N. 1983. Involvement of superoxide anion generation in thehypersensitive response of potato tuber tissues to infection with anincompatible race of Phytophthora infestans and to the hyphalwall components. Physiological Plant Pathology 23, 345–357.
Felle HH, Herrmann A, Hanstein S, Huckelhoven R, Kogel KH.2004. Apoplastic pH signaling in barley leaves attacked by thepowdery mildew fungus Blumeria graminis f. sp. hordei.Molecular Plant–Microbe Interactions 17, 118–123.
Foreman J, Demidchik V, Bothwell JH, et al. 2003. Reactiveoxygen species produced by NADPH oxidase regulate plant cellgrowth. Nature 422, 442–446.
Fryer MJ, Oxborough K, Mullineaux PM, Baker NR. 2002.Imaging of photo-oxidative stress responses in leaves. Journal ofExperimental Botany 53, 1249–1254.
Groom QJ, Torres MA, Fordham-Skelton AP, Hammond-Kosack KE, Robinson NJ, Jones JD. 1996. rbohA, a ricehomologue of the mammalian gp91phox respiratory burst oxidasegene. The Plant Journal 10, 515–522.
Huckelhoven R. 1999. Untersuchungen zur Rolle Reaktiver Sauer-stoffintermediate in der Resistenz der Gerste gegenuber demEchten Gerstenmehltaupilz. PhD thesis, University of Giessen.
Huckelhoven R, Dechert C, Trujillo M, Kogel K-H. 2001.Differential expression of putative cell death regulator genes innear-isogenic, resistant and susceptible barley lines inoculatedwith the powdery mildew fungus. Plant Molecular Biology 47,739–748.
Huckelhoven R, Fodor J, Preis C, Kogel K-H. 1999.Hypersensitive cell death and papilla formation in barley attackedby the powdery mildew fungus are associated with H2O2 butnot with salicylic acid accumulation. Plant Physiology 119,1251–1260.
Huckelhoven R, Kogel K-H. 1998. Tissue-specific superoxidegeneration at interaction sites in resistant and susceptible near-isogenic barley lines attacked by the powdery mildew fungus (Erysiphegraminis f. sp. hordei). Molecular Plant–Microbe Interactions11, 292–300.
Huckelhoven R, Kogel K-H. 2003. Reactive oxygen intermediatesin plant–microbe interactions: who is who in powdery mildewresistance? Planta 216, 891–902.
Jabs T, Dietrich RA, Dangl JL. 1996. Initiation of runaway celldeath in anArabidopsis mutant by extracellular superoxide. Science273, 1853–1856.
Jain SK, Langen G, Borner T, Hess W, Huckelhoven R,Kogel K-H. 2004. The barley albino mutant Albostrians showsenhanced resistance to the biotroph Blumeria graminis f. sp.hordei. Molecular Plant–Microbe Interactions 17, 374–382.
Kawasaki T, Henmi K, Ono E, Hatakeyama S, Iwano M, Satoh H,Shimamoto K. 1999. The small GTP-binding protein rac is aregulator of cell death in plants. Proceedings of the NationalAcademy of Sciences, USA 96, 10922–10926.
Keller T, Damude HG, Werner D, Doerner P, Dixon RA, Lamb C.1998. A plant homolog of the neutrophil NADPH oxidasegp91phox subunit gene encodes a plasma membrane protein withCa2+ binding motifs. The Plant Cell 10, 255–266.
Kim MC, Panstruga R, Elliott C, Muller J, Devoto A, Yoon HW,Park HC, Cho MJ, Schulze-Lefert P. 2002. Calmodulininteracts with MLO protein to regulate defence against mildewin barley. Nature 28, 447–451.
Koga H, Bushnell WR, Zeyen RJ. 1990. Specificity of cell typeand timing of events associated with papilla formation and thehypersensitive reaction in leaves of Hordeum vulgare attacked byErysiphe graminis f. sp. hordei. Canadian Journal of Botany 68,2344–2352.
Kwak JM, Mori IC, Pei ZM, Leonhardt N, Torres MA,Dangl JL, Bloom RE, Bodde S, Jones JD, Schroeder JI.2003. NADPH oxidase AtrbohD and AtrbohF genes function inROS-dependent ABA signalling in Arabidopsis. EMBO Journal22, 2623–2633.
Liszkay A, van der Zalm E, Schopfer P. 2004. Production ofreactive oxygen intermediates (O��
2 ), H2O2, and �OH by maizeroots and their role in wall loosening and elongation growth. PlantPhysiology 136, 3114–3123.
Liu G, Sheng X, Greenshields DL, Ogieglo A, Kaminskyj S,Selvaraj G, Wei Y. 2005. Profiling of wheat class III peroxidasegenes derived from powdery mildew-attacked epidermis revealsdistinct sequence-associated expression patterns. MolecularPlant–Microbe Interactions 18, 730–741.
Miki D, Itoh R, Shimamoto K. 2005. RNA silencing of single andmultiple members in a gene family of rice. Plant Physiology 138,1903–1913.
Morel J, Fromentin J, Blein JP, Simon-Plas F, Elmayan T. 2004.Rac regulation of NtrbohD, the oxidase responsible for theoxidative burst in elicited tobacco cell. The Plant Journal 37,282–293.
Passardi F, Cosio C, Penel C, Dunand C. 2005. Peroxidases havemore functions than a Swiss army knife. Plant Cell Report 5,255–265.
Pfaffl MW. 2001. A new mathematical model for relativequantification in real-time RT-PCR. Nucleic Acids Research 29, e45.
Reeves EP, Lu H, Jacobs HL, Messina CG, Bolsover S,Gabella G, Potma EO, Warley A, Roes J, Segal AW. 2002.
3790 Trujillo et al.
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from

Killing activity of neutrophils is mediated through activation ofproteases by K+ flux. Nature 21, 291–297.
Rentel MC, Lecourieux D, Ouaked F, et al. 2004. OXI1 kinase isnecessary for oxidative burst-mediated signalling in Arabidopsis.Nature 427, 858–861.
Royer-Pokora B, Kunkel LM, Monaco AP, Goff SC,Newburger PE, Baehner RL, Cole FS, Curnutte JT,Orkin SH. 1986. Cloning the gene for an inherited humandisorder chronic granulomatous disease on the basis of itschromosomal location. Nature 322, 32–38.
Rozen S, Skaletsky HJ. 2000. Primer3 on the WWW for generalusers and for biologist programmers. In: Krawetz S, Misener S,eds. Bioinformatics methods and protocols. Methods in MolecularBiology. Totowa, NJ: Humana Press, 365–386.
Sagi M, Davydov O, Orazova S, Yesbergenova Z, Ophir R,Stratmann JW, Fluhr R. 2004. Plant respiratory burst oxidasehomologs impinge on wound responsiveness and development inLycopersicon esculentum. The Plant Cell 16, 616–628.
Sagi M, Fluhr R. 2001. Superoxide production by plant homologuesof the gp91(phox) NADPH oxidase. Modulation of activity bycalcium and by tobacco mosaic virus infection. Plant Physiology126, 1281–1290.
Schultheiss H, Dechert C, Kogel KH, Huckelhoven R. 2002. Asmall GTP-binding host protein is required for entry of powderymildew fungus into epidermal cells of barley. Plant Physiology128, 1447–1454.
Schultheiss H, Dechert C, Kogel K-H, Huckelhoven R. 2003.Functional analysis of barley RAC/ROP G-protein familymembers in susceptibility to the powdery mildew fungus. ThePlant Journal 36, 589–601.
Schweizer P, Pokorny J, Schulze-Lefert P, Dudler R. 2000.Double-stranded RNA interferes with gene function at the single-cell level in cereals. The Plant Journal 24, 895–903.
Simon-Plas F, Elmayan T, Blein JP. 2002. The plasma membraneoxidase NtrbohD is responsible for AOS production in elicitedtobacco cells. The Plant Journal 31, 137–147.
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB. 1997.Subcellular localization of H2O2 in plants. H2O2 accumulation inpapillae and hypersensitive response during the barley-powderymildew interaction. The Plant Journal 11, 1187–1194.
Torres MA, Dangl JL. 2005. Functions of the respiratory burstoxidase in biotic interactions, abiotic stress and development.Current Opinion in Plant Biology 8, 397–403.
Torres MA, Dangl JL, Jones JD. 2002. Arabidopsis gp91phoxhomologues AtrbohD and AtrbohF are required for accumulationof reactive oxygen intermediates in the plant defense response.Proceedings of the National Academy of Sciences, USA 99, 517–522.
Torres MA, Jones JD, Dangl JL. 2005. Pathogen-induced,NADPH oxidase-derived reactive oxygen intermediates suppressspread of cell death in Arabidopsis thaliana. Nature Genetics 37,1130–1134.
Torres MA, Onouchi H, Hamada S, Machida C, Hammond-Kosack KE, Jones JD. 1998. Six Arabidopsis thalianahomologues of the human respiratory burst oxidase (gp91phox).The Plant Journal 14, 365–370.
Trujillo M, Kogel K-H, Huckelhoven R. 2004. Superoxide andhydrogen peroxide play different roles in the nonhost interactionof barley and wheat with inappropriate formae speciales ofBlumeriagraminis. Molecular Plant–Microbe Interactions 17, 304–312.
Vignais PV. 2002. The superoxide-generating NADPH oxidase:structural aspects and activation mechanism. Cellular andMolecular Life Sciences 59, 1428–1459.
Walters DR. 2003. Polyamines and plant disease. Phytochemistry64, 97–107.
Yiu SM, Wong PW, Lam TW, Mui YC, Kung HF, Lin M,Cheung YT. 2004. Filtering of ineffective siRNAs and improvedsiRNA design tool. Bioinformatics PMID, 15333460.
Yoshida LS, Saruta F, Yoshikawa K, Tatsuzawa O, Tsunawaki S.1998. Mutation at histidine 338 of gp91(phox) depletes FAD andaffects expression of cytochrome b558 of the human NADPHoxidase. Journal of Biological Chemistry 273, 27879–27886.
Yoshioka H, Numata N, Nakajima K, Katou S, Kawakita K,Rowland O, Jones JD, Doke N. 2003. Nicotiana benthamianagp91phox homologs NbrbohA and NbrbohB participate in H2O2
accumulation and resistance to Phytophthora infestans. The PlantCell 15, 706–718.
Zhang Z, Collinge DB, Thordal-Christensen H. 1995. Germin-like oxalate oxidase a H2O2-producing enzyme accumulates inbarley attacked by the powdery mildew fungus. The Plant Journal8, 139–145.
Barley respiratory burst oxidase homologue 3791
by guest on Novem
ber 2, 2014http://jxb.oxfordjournals.org/
Dow
nloaded from





























