Embed Size (px)
Citation preview

Scaling and Adaptations of Incisors and Cheek Teethin Caviomorph Rodents (Rodentia, Hystricognathi)
Federico Becerra,1,2* Aldo I. Vassallo,1,2 Alejandra I. Echeverrıa1 and Adria Casinos3
1Departamento de Biologıa, Facultad de Ciencias Exactas y Naturales (FCEyN), Universidad Nacionalde Mar del Plata (UNMdP), Mar del Plata, Argentina2Instituto de Investigaciones Marinas y Costeras (IIMyC), Facultad de Ciencias Exactas y Naturales (FCEyN),Universidad Nacional de Mar del Plata (UNMdP), Consejo Nacional de Investigaciones Cientıficas yTecnicas (CONICET), Mar del Plata, Argentina3Departament de Biologia Animal, Facultat de Biologia, Universitat de Barcelona (UB), Barcelona, Spain
ABSTRACT The South American hystricognathrodents are one of the most diverse mammalian cladesconsidering their occupied habitats, locomotor modesand body sizes. This might have been partly evolved bydiversification of their masticatory apparatus’ structureand its ecological commitment, for example, chisel-toothdigging. In this phylogeny-based comparative study, wetest the relationship between ecological behavior andmechanical features of their incisors and molariforms.In 33 species of nine families of caviomorph rodents, weanalyze incisor attributes related to structural stress re-sistance and molar features related with grindingcapacity, for example, second moment of inertia andenamel index (EI) (enamel band length/occlusal surfacearea), respectively. Most of these variables scaled isomet-rically to body mass, with a strong phylogenetic effect. Aprincipal component analysis discrimination on the EIclustered the species according to their geographic dis-tribution. We presume that selective pressures inAndean–Patagonian regions, on particular feeding hab-its and chisel-tooth digging behaviors, have modeled themorphological characteristics of the teeth. Subterranean/burrower ctenomyids, coruros, and plains viscachasshowed the highest bending/torsion strength and an-chorage values for incisors; a simplified enamel patternin molariforms would be associated with a better grind-ing of the more abrasive vegetation present in moreopen and drier biomes. J. Morphol. 273:1150–1162,2012. � 2012 Wiley Periodicals, Inc.
KEY WORDS: behavioral ecology; cheek-teeth;functional morphology; incisors; South Americansubregions
INTRODUCTION
Vertebrates present a huge diversity in the mor-phology of their feeding apparatus which relates toa wide range of alimentary items and a remarkableecological diversity. In particular, the large mono-phyletic group of South American hystricognathrodents, that is, Caviomorpha, achieved an explo-sive radiation resulting in four extinct and 12extant families (Wyss et al., 1993; Vucetich et al.,1999), becoming one of the most diverse mammalianclades taking into account their occupied habitats,
locomotor modes and body sizes (Mares and Ojeda,1982; Nowak, 1999; Huchon and Douzery, 2001).
Previous studies showed the importance of thephylogeny in caviomorph rodents and ecology onthe morphological variation finding that phyloge-netically close species present more similaritiesthan expected by chance. Ecological heterogeneityis usually associated with high taxonomic ranks(Alvarez et al., 2011).
For feeding behavior and activities such aschisel-tooth digging, biting performance might be akey factor for very different mechanical demands atincisor tips (Stein, 2000; Verzi, 2002; Verzi et al.,2010). Rodent incisors are subjected to bending andshearing stresses during feeding and digging func-tion (Currey, 2002), and thus, the analysis of theirmechanical properties is highly important for char-acterizing the masticatory apparatus structure inrodent taxa with specialized adaptations or modesof life. It has been proposed that most echimyids,for example, evolved from savannah-like environ-ments to Brazilian tropical forests, developingground dwelling and arboreal habits (Mares andOjeda, 1982; Vucetich and Verzi, 1991; Verzi, 2002).Ctenomyids and octodontids (fossorial and evensubterranean rodents), for example, evolved anddiversified in the relatively drier and more openedbiomes of the South-Western South America, with amostly abrasive diet and a chisel-tooth digging
Contract grant sponsor: Ministerio de Ciencia e Innovacion deEspana (2009–2011); Grant number: CGL2008-0832/BOS; Contractgrant sponsors: PIP-CONICET 1380 and PICT 2121 (2011–2013).
*Correspondence to: Federico Becerra, Departamento Biologıa,FCEyN-UNMdP, Dean Funes 3250, 3er. Piso, (B7602AYJ) Mar delPlata, Buenos Aires, Argentina.E-mail: [email protected]
Received 11 February 2012; Revised 10 April 2012;Accepted 18 May 2012
Published online 22 June 2012 inWiley Online Library (wileyonlinelibrary.com)DOI: 10.1002/jmor.20051
JOURNAL OF MORPHOLOGY 273:1150–1162 (2012)
� 2012 WILEY PERIODICALS, INC.

behavior for some genera (Vassallo and Verzi, 2001;Verzi, 2002; Lessa et al., 2008). These diets andbehaviors probably became a key selective factor forthe evolution of a more robust zygomatic arch andlower jaw with powerful adductor muscles. None-theless, some representatives within Cavioideahave gracile skulls and incisors (Vassallo and Verzi,2001), for example, the genera Cavia and Galea.Within this superfamily, which also includes thebiggest live representative, Hydrochoerus, fossorialhabits are barely common (with Microcavia as anexception). On the other hand, most of cheek-toothproperties, particularly occlusal surface area (OSA),are more strongly associated with food quality,alimentary strategy (e.g., monogastric animals vs.ruminants) and basal metabolic rate, than withbody size (Redford and Eisenberg, 1992; Nowak,1999), hence, important functional implicationsmight be assessed.
Because of their ecomorphological diversity, cavio-morph rodents represent an excellent model toanalyze how different ecological and functionaldemands affect biomechanical features and bodyproportions (Vassallo, 1998, 2000; Vassallo and Verzi,2001; Morgan, 2009; Alvarez et al., 2011). Themarked dietary and body size differences are likelyto be reflected by traits of the incisors and cheekteeth. Although there are many excellent anatomicaldescriptions of the feeding apparatus in the orderRodentia, even in caviomorph rodents (e.g., Woods,1972; Koenigswald et al., 1994; Vassallo, 1998, 2000;Vassallo and Verzi, 2001; Alvarez et al., 2011), thereis still a lack of a comprehensive morphofunctionalanalyses of rodent teeth. The aim of this study wasto assess the relationship among ecological behaviorand mechanical features of the teeth throughoutthe caviomorph linage, taking into account theirphylogeny background. We expected to find a corre-lation between: 1) tooth-digging behavior andharder soils with higher bending and torsionresistance values on incisors (e.g., second momentof inertia and polar moment of inertia, geometricalfeatures proportional to structural strength); and 2)feeding on hard vegetation with grinding capacityof cheek-teeth, measured by the occlusal surface ofarea and the enamel band pattern.
MATERIALS AND METHODS
Based on specimens corresponding to 33 species of nine differ-ent families of caviomorph rodents (see Table 1, Appendix A),housed in the Museo de Ciencias Naturales de Mar del Plata‘‘Lorenzo Scaglia,’’ specific functional properties of the incisorsand cheek-teeth were estimated by means of digital photo-graphs (Fig. 1). Because of the high functional symmetrybetween upper and lower hypsodont and rootless dentitionwithin species (Koenigswald et al., 1994), morphological attrib-utes of incisors and cheek teeth were obtained from the upperjaw to predict their mechanical behavior. Particularly, for theprocumbency angle (PA), the upper incisors were analyzedbecause of their proposed linkage to the ecological habits(Vassallo, 1998; De Santis and Moreira, 2000; Mora et al.,
2003). Also, to avoid ontogenetic variation, only adult specimenswere analyzed, defined by the presence of a functional thirdmolar. Even though we did not focus on sexual differences inthis work, specimens of both sexes were included whenever itwas possible.
The sampled species were selected following two differentcriteria: 1) a wide representation of the variation of body sizewithin and between families; 2) a wide representation of behav-ioral diversity considering diet (abrasive vs. nonabrasive foods)and burrowing habits (ground dwelling, fossorial, and burrow-ing/subterranean species).
Incisor Stress Resistance and Anchorage
Cross-sectional area (CA) and the second moment of inertia(I) were used (see Verzi et al., 2010; Fig. 1a) as bendingstrength proxies for the incisors, whereas the polar moment ofinertia (J) was analyzed as a torsion resistance proxy (see
TABLE 1. Specimens of caviomorph rodents used in this study,and housed in the mammalogical (MMPMa) or paleontological
collection of the Museo Municipal de Ciencias Naturales‘‘Lorenzo Scaglia’’
Family ErethizontidaeE. dorsatum (N 5 1): MMPMa 1437S. spinosus (N 5 1): MMPMa 4097
Family ChinchillidaeL. maximus (N 5 6): MMPMa 1307, 2095, 2096, 2721, 2722,4220L. viscaccia (N 5 1): MMPMa 2088C. lanigera (N 5 1): MMPMa 332, 4212, 5010
Family MyocastoridaeM. coypus (N 5 4): MMPMa 24, 311, 4019, 4103
Family EchimyidaeT. dimidiatus (N 5 1): MMPMa 1474E. dasythrix (N 5 2): MMPMa 1261, 1262T. apereoides (N 5 3): MMPMa 150, 151, 1296T. cunicularius (N 5 3): MMPMa 1247, 1248, 1296P. albispinus (N 5 2): MMPMa 1259, 1260P. guainae (N 5 1): MMPMa 1914
Family CtenomyidaeActenomys sp. (N 5 6): 716S, 884M, 1224M, 1485M, 1565M,2075MC. magellanicus (N 5 4): MMPMa 7, 2500, 2501, 2502C. australis (N 5 7): MMPMa 1341, 1344, 3248, 3261, 3235,3287, 4219C. talarum talarum (N 5 4): 1345, 1346, 1352, 2999C. opimus opimus (N 5 1): MMPMa 1357C. leucodon (N 5 2): MMPMa 1358, 1359C. tuconax (N 5 5): MMPMa 2960, 3182, 3342, 3346, 3695
Family OctodontidaeO. gliroides (N 5 3): MMPMa 2532, 3557, 4151S. cyanus cyanus (N 5 4): MMPMa 3583, 3585, 3590, 3591O. mimax (N 5 1): MMPMa 388T. barrerae (N 5 1): MMPMa 3199
Family CaviidaeK. rupestris (N 5 2): MMPMa 1291, 1913G. musteloides (N 5 4): MMPMa 106, 109, 1065, 3295G. spixii (N 5 3): MMPMa 1249, 1250, 1388D. patagonum (N 5 6): MMPMa 23, 335, 336, 1353, 2143,4216C. aperea (N 5 5): MMPMa 29, 86, 605, 749, 2824C. porcellus (N 5 1): MMPMa 4202M. australis (N 5 5): MMPMa 289, 301, 304, 318, 3250
Family HydrochaeridaeH. hydrochaeris (N 5 5): MMPMa 47, 440,1354, 4022, 4023
Family DasyproctidaeA. paca (N 5 2): MMPMa 19, 21Dasyprocta sp. (N 5 2): MMPMa 25, 26
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1151
Journal of Morphology

Rocha-Barbosa and Casinos, 2011). The I is a geometrical prop-erty of the cross-section of structures, used as a maximum re-sistance proxy for bending, following the function: I 5
PdA 3
y2, where dA represents the differential of the CA, and y is thedistance to neutral axis. As the analysis considers two orthogo-nal axes, and because pressure from feeding and burrowing isexerted through the anteroposterior axis (sagittal axis), we ana-lyzed the bending resistance according to it. Then, J is calcu-lated as the sum of the maximum and minimum moments ofarea: J 5 Imax 1 Imin. The second moment of inertia and CAwere scaled and estimated by means of the left incisor’s depthand width, from the palatal view photographs (using MomentMacro application of ImageJ 1.41 by W. Rasband—NationalInstitute of Health, rsb.info.nih.gov/ij, and tpsDig by Rohlf,2006, respectively). Following Verzi et al. (2010), from lateralphotos of the left side of the skulls, the upper incisors’ PA wasestimated as the angle formed between the chord of the exposedincisor and a line parallel to the occlusal plane of the molari-form cheek-teeth (Thomas’ angle), using tpsDig, and latertransformed into radians for the analyses. Finally, using lateralimages of the rostrum, the left incisor root length (IRL) wasestimated by means of the Data Adjustment to Logarithmic Spi-rals (Aubone, 2008; Fig. 1b). This measure of anchorage to pre-maxillary and maxillary bones (Moxham and Berkovitz, 1995)was analyzed, because these rodents present the incisor rootalveolus protruded on either side of the rostrum, lateral to themolar roots, as previous researchers have mentioned (see Moraet al., 2003).
Functional Properties of Molar Rows
For the enamel band length (EL), that is, the total length ofthe enamel band from a palatal view, orthogonal to the surface(Fig. 1c), and the OSA, palatal view photos of the completemolar rows (fourth molariform premolar and three molars)were taken. The former parameter was considered as a measureof grinding capacity (Koenigswald et al., 1994). After contrast/brightness adjustment and enamel digital isolation (Fig. 1c),the occlusal area was determined using the Moment Macroapplication of ImageJ 1.41 (see above), whereas the EL wasmeasured by means of the ImageTool 3.0 (developed by theDDSDX-UTHSCSA, ddsdx.uthscsa.edu/dig/itdesc.html). Finally,the enamel index (EI) was calculated as the EL per unit ofOSA. Because of the horizontal component of masticatory direc-tion is most important for biomechanical interpretation of theflattened occlusal surfaces of hypsodont cheek teeth (Koenigs-
wald et al., 1994), this index reflects the amount of leading andtrailing enamel cutting edges through these surfaces.
Statistical Analyses
The original data were log10 transformed (see Peters, 1983)and, assuming a type-II regression model (Sokal and Rohlf,1995), were analyzed by means of allometric equations carriedout using the Reduced Major Axis software (Bohonak, 2004).This method was used because there was error associated tothe measurement of both x and y and it is the structural rela-tionship between the two variables that is required (Wartonet al., 2006; Smith, 2009). The equation y 5 a 3 xb wasassumed as the best descriptor for the allometric pattern: b wastermed the allometry index (Huxley, 1932). Incisor strength, in-cisor root length and procumbency values, and molar row’s pa-rameters were regressed against the body mass (BM) as a bodysize proxy. Then, preliminary tests were run out for parallelismand, depending on their results, ANCOVAs or separate-slopesmodels were performed for all the variables (i.e., CA, I, J, IRL,PA, EL, OSA, EI) to study the effect of body size and phyloge-netic affiliation by families. In these analyses, we used the BMas covariate and the family as categorical variable (i.e., Cteno-myidae vs. Octodontidae vs. Chinchillidae vs. Caviidae vs.Hydrochaeridae vs. Myocastoridae vs. Echimyidae vs. Erethi-zontidae vs. Dasyproctidae). Measurements of BM were takenfrom Eisenberg (1981), Redford and Eisenberg (1992), Nowak(1999), and Vassallo and Echeverrıa (2009).
The traditional allometric approach fails to account for possi-ble nonindependence of taxa (Harvey and Pagel, 1991), how-ever, our analysis of interspecific scaling relations for dentalmeasurements was performed taking into account the phyloge-netically independent contrasts (Felsenstein, 1985). Log10 datawere converted to phylogenetically independent standardizedcontrasts using the PDAP (Midford et al., 2003) package modulein the program MESQUITE (Maddison and Maddison, 2006).Standardization tests in PDTREE indicated that Grafen’sbranch length transformation (Grafen, 1992) was the appropri-ate method for assigning arbitrary branch lengths (Garlandet al., 1992). This method assumes a gradual Brownian motionmodel of evolution, and the height of each node is proportionalto the number of species derived from it. Slopes, confidenceintervals, and other regression statistics were obtained fromPDTREE, Statistica 6.0, Microsoft Excel and a program for run-ning Model II Regression (Reduced Major Axis, see above). Phy-logenetic relationships among taxa were taken from Opazo(2005) for caviomorph rodents (Mammalia, Rodentia, Hystricog-nathi). Different sources were consulted for families Echimyidae(Patterson and Velazco, 2006) and Chinchillidae (Spotornoet al., 2004), while for species within Ctenomys (Hystricognathi,Ctenomyidae) phylogenies from Castillo et al. (2005), D’Elıaet al. (1999), and Mascheretti et al. (2000) were considered.
Residual values of regressions (CA, I, J, IRL, EL, and OSAvs. BM), were examined to assess their variation beyond thebody size differences. The EI was explored by a principal com-ponent analysis [PCA; using the log10-BM, log10-OSA, log10-EL,and EI, with Family as the grouping variable, based on a corre-lation matrix, computed variances as SS/(N-1)]. Then, the EIwas reassessed by ANCOVA (with log10 OSA and the main clus-ters from the PCA as predictor variables).
RESULTSFoods Cutting and Ecological Behavior—Incisors
Incisors’ bending stress resistance. The cav-iomorph rodent species studied here show strikingdifferences at both, size and shape, of the cross-sectional area of the incisors (Fig. 2).
Fig. 1. Detailed views of the masticatory apparatus. (A) Pala-tal view of the upper incisors with the cross-sectional area (CA)of a single tooth. (B) Lateral view of the rostrum with the esti-mation of Thomas’ angle (PA). The extension of the incisor’s root(IRL) is outlined by black dots. (C) Occlusal surface area (OSA)of the upper molar row of C. australis (left) and T. dimidiatus(right). The enamel outline (EL) is enhanced by a solid line.
1152 F. BECERRA ET AL.
Journal of Morphology

By the overall regression analyses of CA, I andJ against the body size, and considering the geo-metrical similarity, we found that these variablesscale isometrically by power coefficients nonstatis-tically different from 0.66, 1.33, and 1.33, respec-tively, once they are phylogenetically weighted(Table 2). The coefficients did not vary substan-tially considering original data versus phylogeneti-cally independent contrasts. Even more, thepreliminary analysis for parallelism showed nosignificant difference between the family slopes inrelation to the BM (F[6,17] 5 2.01, P 5 0.121, F[6,17]
5 2.25, P 5 0.088 and F[6,17] 5 1.55, P 5 0.222 forCA, I and J, respectively). Noteworthy, the ana-lyzed caviomorph rodents showed a highly similardistribution values for all structural resistancevariables, with a strong effect of both body size(ANCOVA: F[1,23] 5 123.31, P � 0.001, F[1,23] 5109.22, P � 0.001 and F[1,23] 5 136.14, P � 0.001for CA, I and J, respectively) and phylogeny onthe bending variables (ANCOVA: F[1,23] 5 21.04,P � 0.001, F[1,23] 5 23.49, P � 0.001 and F[1,23] 520.20, P � 0.001 for CA, I and J, respectively;Figs. 3 and 4). Results for I and J were highly sim-ilar in pattern and, then, figures of the polarmoment were omitted. The inspection of residualvalues showed that although Hydrochaeridae, Cte-nomyidae, and Myocastoridae presented positive
relative values, Caviidae, Dasyproctidae, Chinchil-lidae, and Echimyidae had negative values. Erethi-zontidae and Octodontidae presented values thatwere close to those expected for their body sizes.The American porcupine (Erethizon dorsatum;Linnaeus, 1758), the coruro (Spalacopus cyanus;Molina, 1782) and the plains viscacha (Lagostomusmaximus; Desmarest, 1817) showed much highervalues than their relatives.
Incisor anchorage. The overall regressionanalysis indicated that the IRL scaled isometri-cally to BM by a power coefficient nonstatisticallydifferent from 0.33 (Table 2). Besides, the analysisfor parallelism did not show a significant differ-ence for family slopes (F[6,17] 5 0.55, P 5 0.763).By the inspection of residual values, we found thatthese species showed an overall qualitative similarpattern for root length values, to those correspond-ing to the bending stress resistance variables, witha strong effect of body size and family differentia-tion (ANCOVA: F[1,23] 5 46.65, P � 0.001 andF[8,23] 5 7.94, P � 0.001, respectively; Fig. 5;Table 2). The American porcupine, the coruro andthe plains viscacha maintained their higher thanrelatives condition. However, the values for Hydro-chaeridae decreased to a negative relative position,while the value for Dasyproctidae rose to a posi-tive position.
Fig. 2. Palatal view photographs of upper incisors’ diversity in caviomorph rodents, with the detailed shape of the left tooth’scross-sectional area. Species from left to right: first row: C. talarum (subterranean, Ctenomyidae), L. maximus (burrower, Chinchil-lidae), M. coypus (burrower-semiaquatic, Myocastoridae); second row: H. hydrochaeris (semiaquatic, Hydrochaeridae), A. paca(ground dweller, Dasyproctidae), S. spinossus (arboreal, Erethizontidae); third row: O. mimax (burrower, Octodontidae), E. dasy-thrix (ground dweller, Echimyidae), C. aperea (ground dweller, Caviidae). Scale bar: 5 mm.
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1153
Journal of Morphology

Incisor procumbency. The angle of attack ofthe incisors showed nonstatistically significantdifferences considering the body size (Parallelism:F[6,17] 5 1.46, P 5 0.251; ANCOVA: F[1,23] 5 0.53,P 5 0.47) but it did so by family differentiation(Parallelism: F[6,17] = 1.17, P 5 0.365; ANCOVA:F[1,23] 5 0.53, P 5 0.47 and F[8,23] 5 3.13, P 50.015; Fig. 6).
Food Processing—Cheek-TeethCheek-teeth measures. As was shown for
most of the incisor parameters, the analyzed cavio-morph rodents presented a strong effect of bothbody size and family differentiation over thecheek-teeth variables. Although the overall regres-
TABLE
2.Red
ucedmajoraxis(typ
eII)regressionvalues
oftheanalyzedincisors
andmolarrow’spara
metersversu
sthelog10BM
asabod
ysize
proxy
Originallog10
mea
suremen
tsCA
(mm
2)
I(m
m4)
J(m
m4)
IRL(m
m)
PA
(radians)
OSA
(mm
2)
EL
(mm)
EI
Allom
etric
coefficien
tb
0.60
(0.51–0.71)
1.26
(1.06–1.51)
1.17
(0.99–1.38)
0.31
(0.26–0.38)
20.36
(20.47–0.50)
0.74
(0.62–0.85)
0.48
(0.42–0.55)
25.93
(29.24–23.91)
Normaliza
tion
constant
20.89
(21.21to
20.61)
23.19
(23.88to
22.62)
22.43
(23.08to
21.94)
0.38
(0.18–0.55)
2.56
(0.24–2.82)
20.94
(21.24to
20.60)
0.43
(0.28–0.61)
22.36
(16.15–30.72)
R2
0.72
0.67
0.75
0.56
<0.01
0.83
0.83
0.20
Standard
ized
contrasts
Allom
etric
coefficien
tb
0.70
(0.57–0.74)
1.46
(1.21–1.55)
1.39
(1.14–1.47)
0.37
(0.33–0.49)
0.35
(20.59–0.66)
0.81
(0.68–0.89)
0.49
(0.32–0.56)
25.10
(29.31to
23.46)
R2
0.91
0.91
0.91
0.78
0.02
0.88
0.83
0.30
Mea
nvalues
ofthedifferentsp
eciesanalyzed(n
533)wereusedfortheallom
etricanalyses;
whilecontrastsbetwee
nthosesp
ecies(n
532)wereconsidered
forphylogen
eti-
callyindep
enden
tcontrastsanalyses,
Incisors:CA:Cross-section
alarea;I:
Secon
dmom
entof
inertia;J:polarmom
entof
inertia;IR
L:In
cisor’sroot
length;PA;Procu
mben
cyangle.Molarrows:
OSA:Occlusa
lsu
rface
area;EL:Enamel
barlength;EI:
Enamel
index
.Con
fiden
ceintervals
(95%)are
betwee
nbrack
ets.
Fig. 3. Regression of incisor cross-sectional area versus BMin caviomorph rodents, grouped by families. (A) Bivariate scat-ter-plot with the overall regression line; and (B) residual valuesorganized by their phylogenetic position. Chinchillidae: opensquares; Ctenomyidae: open triangles; Hydrochaeridae: opencircle; Erethizontidae: solid squares; Caviidae: solid triangles;Myocastoridae: solid circle; Octodontidae: solid diamond; Echi-myidae: asterisks; Dasyproctidae: vertical crosses.
1154 F. BECERRA ET AL.
Journal of Morphology

sion analysis showed that OSA scaled isometricallyto BM by a power coefficient nonstatistically differ-ent than 0.66 (Fig. 7; Table 2), the analysis forparallelism exhibited a significant difference in thefamily slopes (F[5,17] 5 7.63, P < 0.001). However,the separate-slope model revealed an importantsize effect (F[6,17] 5 7.11, P < 0.001). On the otherhand, EL was positively allometric by a powercoefficient statistically higher than 0.33 (Fig. 8;Table 2), with no significant parallelism betweenfamily slopes (F[5,17] 5 3.58, P 5 0.022) and animportant size effect as well (Separate-slopemodel: F[6,17] 5 3.53, P 5 0.019). The residualvalues did not show affiliation strictly familialpattern for either OSA or EL. However, some over-all familial patterns may be seen (separate-slopemodel: F[5,17] 5 7.73, P < 0.001 and F[5,17] 5 4.54,P 5 0.008; Figs. 7 and 8 for OSA and EL, respec-tively). In general terms, families Dasyproctidae,
Fig. 4. Regression of the second moment of inertia of theincisors versus BM in caviomorph rodents, grouped by families.(A) Bivariate scatter-plot with the overall regression line; and(B) residual values organized by their phylogenetic position.Family references as in Figure 3.
Fig. 5. Regression of the incisor root length versus BM incaviomorph rodents, grouped by families. (A) Bivariate scatter-plot with the overall regression line; and (B) residual valuesorganized by phylogenetic position. Family references as inFigure 3.
Fig. 6. Scatter-plot of the regression of the incisor PA versusBM in caviomorph rodents, grouped by families. Dashed linecorresponds to the orthodont condition. Family references as inFigure 3.
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1155
Journal of Morphology
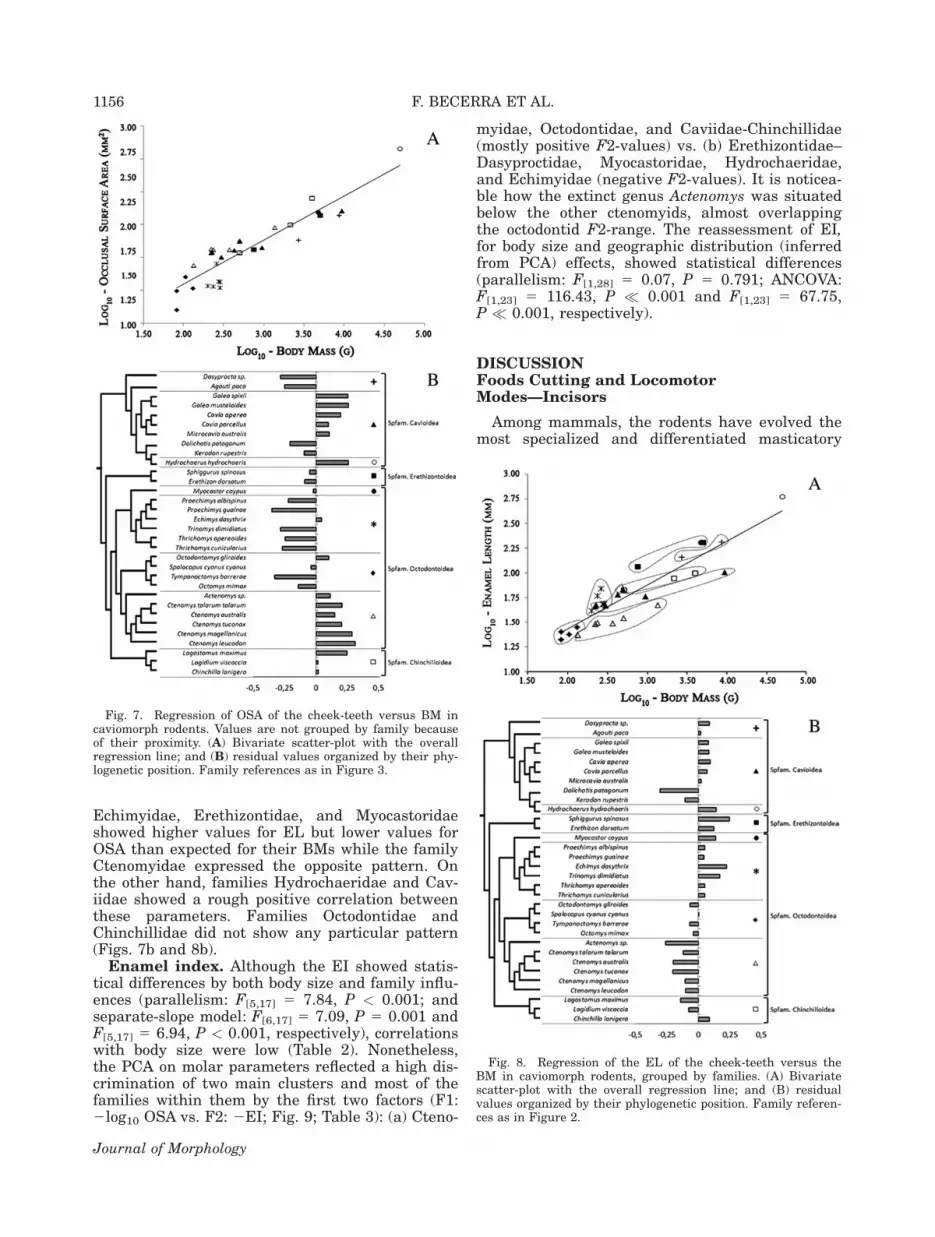
Echimyidae, Erethizontidae, and Myocastoridaeshowed higher values for EL but lower values forOSA than expected for their BMs while the familyCtenomyidae expressed the opposite pattern. Onthe other hand, families Hydrochaeridae and Cav-iidae showed a rough positive correlation betweenthese parameters. Families Octodontidae andChinchillidae did not show any particular pattern(Figs. 7b and 8b).
Enamel index. Although the EI showed statis-tical differences by both body size and family influ-ences (parallelism: F[5,17] 5 7.84, P < 0.001; andseparate-slope model: F[6,17] 5 7.09, P 5 0.001 andF[5,17] 5 6.94, P < 0.001, respectively), correlationswith body size were low (Table 2). Nonetheless,the PCA on molar parameters reflected a high dis-crimination of two main clusters and most of thefamilies within them by the first two factors (F1:2log10 OSA vs. F2: 2EI; Fig. 9; Table 3): (a) Cteno-
myidae, Octodontidae, and Caviidae-Chinchillidae(mostly positive F2-values) vs. (b) Erethizontidae–Dasyproctidae, Myocastoridae, Hydrochaeridae,and Echimyidae (negative F2-values). It is noticea-ble how the extinct genus Actenomys was situatedbelow the other ctenomyids, almost overlappingthe octodontid F2-range. The reassessment of EI,for body size and geographic distribution (inferredfrom PCA) effects, showed statistical differences(parallelism: F[1,28] 5 0.07, P 5 0.791; ANCOVA:F[1,23] 5 116.43, P � 0.001 and F[1,23] 5 67.75,P � 0.001, respectively).
DISCUSSIONFoods Cutting and LocomotorModes—Incisors
Among mammals, the rodents have evolved themost specialized and differentiated masticatory
Fig. 7. Regression of OSA of the cheek-teeth versus BM incaviomorph rodents. Values are not grouped by family becauseof their proximity. (A) Bivariate scatter-plot with the overallregression line; and (B) residual values organized by their phy-logenetic position. Family references as in Figure 3.
Fig. 8. Regression of the EL of the cheek-teeth versus theBM in caviomorph rodents, grouped by families. (A) Bivariatescatter-plot with the overall regression line; and (B) residualvalues organized by their phylogenetic position. Family referen-ces as in Figure 2.
1156 F. BECERRA ET AL.
Journal of Morphology

apparatuses (Hildebrand, 1985; Walker and Liem,1994). Particularly, caviomorph rodents present ahigh morphological diversity of the rostral region(Vassallo, 2000) with various species using theirincisors for cutting stems/roots or digging throughcompact soils (Vassallo, 1998; Lessa et al., 2008).We presume that selection on particular feedinghabits and digging behaviors (Biknevicius et al.,1996; Bacigalupe et al., 2002) have evolutionarilymodeled the morphological characteristics of theforeteeth (incisors and canines) in certain mamma-lian groups as precise engineering pieces resistantto the bending stress exerted on their tips. Thesecharacteristics of the incisors, studied here in cav-iomorph rodents by means of CA, I and J, dependon the locomotor mode and/or the food cuttingefforts, in accordance with that was found by Croftet al. (2011). Taking into account that chisel-toothdigging in subterranean species implies muchhigher forces than ground dwelling or arborealbehaviors, we found that species previously identi-fied as subterranean or fossorial (Mares andOjeda, 1982) presented higher bending strengthresistance values, specifically tuco-tucos (genusCtenomys), coruros (S. cyanus), and plains visca-chas (L. maximus). Besides the coypu (Myocastorcoypus; Molina, 1782) forages primarily on hygro-philic and terrestrial monocotyledons (Guichonet al., 2003), some authors have stated that it alsoincorporates some shelled molluscs in its diet(Mares and Ojeda, 1982; El-Kouba et al., 2009)which, besides its digging behavior, might involvestrong forces at the incisor tips. It is notable thatthe American porcupine, a mainly arboreal rodent,also shows high stress resistance values.
Because teeth are nearly inelastic (Freeman andLemen, 2007), pressures exerted at the incisor tipswould propagate directly to the rostral region, pos-sibly posing a fracture risk to the alveolar bone(Boyle, 1938; Moxham and Berkovitz, 1995). None-theless, the non-enameled surface of the insicors’embedded portion is covered by the periodontalligament that provides an elastic matrix that dissi-pates the external loads on the incisors (Zajicek,1976; Sloan and Carter, 1995). These loads gener-ate an anteroposterior pressure that acts mainlyalong the incisors’ root length. Therefore, the rela-tively deeper root present in the digging species(see Fig. 5) would offer notable ecological advan-tages against either too hard soils or obstacles tobe mechanically overcome by the forelimbs.
Verzi (2002), Tomkins et al. (2005), and Loewe(2009) among others, suggested that selective pres-sures would force body structures to adapt to com-pete for very different needs and demands. Thiswould constrain the development of morphologicaladaptations explaining many ‘‘suboptimizations’’as compromises to produce the best overall condi-
Fig. 9. Scores on Factor 1 and Factor 2 axes from the PCA of molar variables in caviomorph rodents, grouped by families. Sil-houettes, BM (g) and molar rows with highlighted enamel bands correspond to the highest and lowest Factor 1 values (T. barreraeand H. hydrochaeris, respectively), the two highest and the lowest Factor 2 values (T. dimidiatus, Dasyprocta sp, and C. australis,respectively), and the central value for both factors (C. lanigera). Family references as in Figure 3.
TABLE 3. Molar rows’ parameters by the different factors ofthe PCA, and total variance explained
Factor 1 Factor 2 Factor 3
OSA10 0.4519a 0.0004 0.5477EL10 0.2933 0.4458 0.2609EI 0.2548 0.5538a 0.1914Total variance (%) 72.85 26.42 0.74
aHighest representation by factor.OSA10, Log10-occlusal surface of area; EL10, Log10-Enamelband length; EI, Enamel index.
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1157
Journal of Morphology

tion. The opposite relative values for incisors’bending stress resistance and anchorage observedin the capybaras (Hydrochaeridae) and the com-mon agoutis–acouchis (Dasyproctidae) may thus beinterpreted as a suboptimal compromise betweenthe feeding/digging demands of the masticatoryapparatus versus other demands for rostral devel-opment, for example, aggressive intra/interspecificinteractions (Macdonald, 1981; Herrera and Mac-donald, 1993).
Extending the work of Vassallo (1998) and Moraet al. (2003) on the genus Ctenomys, we found thatthe PA of the incisors is not significantly influencedby the body size in Caviomorpha, although it isassociated with digging behavior, as proposed forcoruros by Bacigalupe et al. (2002). Hence, tuco-tucos, coypus, coruros, and plains viscachas showproodont upper incisors, allowing more directly con-frontation of obstacles and soils during excavation(Fig. 6; Vassallo, 1998; Verzi, 2002; Mora et al.,2003). Taking into account, the high procumbencyin the genus Actenomys (family basal genus) and itsmorphotype as an ancestral ctenomyid (De Santisand Moreira, 2000; Lessa et al., 2008), the highproodont condition shared by the extant ctenomyidrodents could be considered a plesiomorphic charac-ter developed early in the family Ctenomyidae. Par-ticularly, the white-toothed tuco-tuco (Ctenomysleucodon; Waterhouse, 1848) showed some of thehighest relative values for every analyzed variable(Figs. 3b, 4b, 5b, 6) supporting its description as achisel-tooth ctenomyid digger. Coruros’ incisors arehighly proodont but slim, as in the other octodon-tids (Lessa et al., 2008). In this case, the high pro-cumbency could be associated to a phylogeneticallyconstrained CA working against the selective pres-sures of the chisel-tooth digging behavior. Signifi-cantly high procumbency was also found in twonondigger species, the cui (Cavia aperea; Erxleben,1777) and the Guinea pig (Cavia porcellus; Lin-naeus, 1758) but without the deep incisor rootsneeded for chisel-tooth digging (Verzi, 2002; Fig.5b). As seen for other parameters of the incisors,the American porcupine shows a deep root, but ouranalyses cannot explain this fact.
Food Processing—Cheek-Teeth
In the caviomorph rodents, the relationshipbetween basal metabolic rates and ecological fea-tures (e.g., substrate, climate, distribution) and spe-cific characteristics (such as BM and family affilia-tion) is highly complex (Arends and McNab, 2001).Nevertheless, our findings showed that withinthese hystricognath rodents the occlusal area cer-tainly scale isometrically to BM. This might be pos-sible because of the dietary homogeneity within thegroup, compared with the high variability presentwithin mammals generally (Ungar, 2010). Beyondthis influence, nearly every species that evolved
linked to the more open and drier biomes of the Pat-agonian Subregion exhibits larger and more speci-alized grinding areas than those species linked tothe Brazilian Subregion of the Neotropical Region(sensu; Hershkovitz, 1958; Vucetich and Verzi,1995; Verzi, 2002). It suggests that these grindingocclusal areas have been a key feature to use theseless nutritious and more abrasive food sources inthe Patagonian Subregion. Thus, dietary qualitydifferences might have influenced different dietarygroups and food-processing strategies (e.g., grazersvs. browsers; see Perez-Barberıa and Gordon,1998). Some researchers have proposed another hy-pothesis for the differences on grinding areas,which would be linked to diverse physiologicalmechanisms of digestion (Janis and Fortelius, 1988;Janis, 1995; Vizcaıno et al., 2006). A deeper investi-gation on these mechanisms in caviomorph rodentswould be necessary to test the later hypothesis.
Many caviomorph species evolved throughout therelatively arid South-Western portion of SouthAmerica (i.e., Andean-Patagonian regions), develop-ing special adaptations for a fossorial, burrowing oreven subterranean life, such as skull shortening,wider mandibular crests, more robust zygomaticarches and masseter muscles, and orientatedenamel bands for better grinding of food (Greaves,1982; Wyss et al., 1993; Vucetich and Verzi, 1995;Vucetich et al., 1999; De Santis and Moreira, 2000;Olivares et al. 2004; Verzi et al., 2010). Further-more, some caviomorph species present an evolu-tionary simplification of the enamel contour lines(Verzi, 1999, 2001, 2002; Candela, 2002, Verzi et al.,2004). A simplification of the EL in Patagonianrodents would lead to a lower EI, as shown by thePCA discrimination (higher values for Factor 2, Fig.9). This reduction is functionally important becauseit implies fewer leading and/or trailing enamel cut-ting edges. Because of the orthogonal disposition ofthe enamel bands with respect to lower jaw move-ment during chewing (Koenigswald et al., 1994;Verzi, 2002; Olivares et al., 2004; Verzi et al., 2004),we suggest that taxa such as Ctenomyidae, Octo-dontidae, and some Chinchillidae, concentratechewing pressure onto fewer edges (Fig. 10). There-fore, the association of this reduction with theenamel reinforcement, by the repeated secondaryacquisition of radial enamel in the superfamiliesOctodontoidea, Cavioidea, and Chinchilloidea, espe-cially in ctenomyid species (Vieytes, 2003), wouldimprove grinding of the more abrasive vegetationpresent in the South-West of South America. Evenmore, the grinding performance could be enhancedby an oblique chewing mode, as suggested by theconcordance of 1) the overall pattern of the PCA F2-values of octodontoid rodents (Ctenomyidae 1 Octo-dontidae), and 2) the clustering of Ctenomys-Octodont-omys/Spalacopus/Octomys-Tympanoctomys (oblique topropalinal chewing mode) found by Vassallo and Verzi(2001) and Olivares et al. (2004).
1158 F. BECERRA ET AL.
Journal of Morphology

The families Hydrochaeridae and Myocastoridaehave evolved under other conditions. These twoclades have a large geographic distribution (Maresand Ojeda, 1982) including both Patagonian andBrazilian subregions, but their semiaquatic habitsconfine them to wetter biomes and mostly semia-quatic and softer food sources, for example, sprout-ing grass (Guichon et al., 2003; Desbiez et al.,2011; Leggieri et al., 2011). Although they have ahighly oblique chewing direction and powerful inci-sors, the semiaquatic coypu have not really speci-alized to an abrasive diet and still possess a highEI value (Martin, 1993; Vassallo and Verzi, 2001).Interestingly, Vucetich and Verzi (1995), and Verzi(2002) established that the extinct genus Acten-omys could represent the plesiomorphic conditionfor living ctenomyid rodents and may be close tosome generalized octodontid genera. Its F2-valuein the PCA relatively low for a ctenomyid rodentbut was close to the more generalized octodontidOctodontomys gliroides; Gervais and d’Orbigny,1844, which has a wetter habitat with less abra-sive vegetation (De Santis and Moreira, 2000).This overall clustering fits quite well with the eco-logical and geographical distribution of these taxa,even though it does not show as strong a phyloge-netic signal as other caviomorph cranio-dentalmorphological features (Alvarez et al., 2011).
In conclusion, certain components of the feedingapparatus studied in caviomorph rodents evidencedconstancy to the rodent Bauplan (see also Vassallo,2000), but different ecological demands appear tohave shaped several adaptive properties of theteeth. Digging species from the South-West ofSouth America, for example, tuco-tucos and plainsviscachas, have incisors that are relatively resistant
to bending stresses, deeper roots and higher pro-cumbency, what agrees with higher biting forces(Becerra et al., 2011). In addition, their molars areadapted to abrasive foods by enlargement of theOSA and simplification of the enamel pattern.
ACKNOWLEDGMENTS
The authors thank the Mammalogical SectionCurator of the Museo Municipal de Ciencias Natu-rales ‘‘Lorenzo Scaglia’’, Lic. Damian Romero, forthe access to the collection; and Prof. Adrian P.Cisilino, Ph.D. (INTEMA-UNMdP) for mechanicalcounseling. Also, they specially thank Prof. SusanW. Herring (University of Washington) and twoanonymous reviewers for their valuable sugges-tions and corrections.
LITERATURE CITED
Alvarez A, Perez SI, Verzi DH. 2011. Ecological and phyloge-netic influence on mandible shape variation of South Ameri-can caviomorph rodents (Rodentia: Hystricomorpha). Biol JLinn Soc 102:828–837.
Arends A, McNab BK. 2001. The comparative energetics of‘‘caviomorph" rodents. Comp Biochem Physiol A 130:105–122.
Aubone A. 2008. Data adjustment on logarithmic spirals,v:020608. UNMdP-INIDEP, Dpto. de Matematica. Availableupon request: [email protected].
Bacigalupe LD, Iriarte-Dıaz J, Bozinovic F. 2002. Functionalmorphology and geographic variation in the digging appara-tus of coruros (Octodontidae: Spalacopus cyanys). J Mammal83:145–152.
Becerra F, Echeverrıa AI, Vassallo AI, Casinos A. 2011. Biteforce and jaw biomechanics in the subterranean rodent Talastuco-tuco (Ctenomys talarum) (Caviomorpha: Octodontoidea).Can J Zool 89:334–342.
Biknevicius AR, Van Valkenburgh B, Walker J. 1996. Incisorsize and shape: implications for feeding behaviours in saber-toothed ‘‘cats.’’ J Vertebr Paleontol 16:510–521.
Bohonak A. 2004. RMA: software for reduced major axis regres-sion. Biotropica 38:581–591.
Boyle PE. 1938 Tooth suspension: A comparative study of theparadental tissues of Man and of the Guinea pig. J Dent Res17:37–46.
Candela AM. 2002. Lower deciduous tooth homologies in Ere-thizontidae (Rodentia, Hystricognathi): Evolutionary signifi-cance. Acta Paleontol Pol 47:717–723.
Castillo AH, Cortinas MN, Lessa EP. 2005. Rapid diversificationof South American tuco-tucos (Ctenomys; Rodentia, Ctenomyi-dae): Contrasting mitochondrial and nuclear intron sequen-ces. J Mammal 86:170–179.
Croft DA, Niemi K, Franco A. 2011. Incisor morphology reflectsdiet in caviomorph rodents. J Mammal 92:871–879.
Currey JD. 2002. Bone: Structure and Mechanic, 2nd ed. NewJersey, USA: Princeton University Press.
D’Elıa G, Lessa EP, Cook JA. 1999. Molecular phylogeny oftuco-tucos, genus Ctenomys (Rodentia, Octodontidae): Evalua-tion of the mendocinus species group and the evolution ofasymmetric sperm. J Mammal Evol 6:19–38.
De Santis LJM, Moreira GJ. 2000. El aparato masticatorio delgenero extinto Actenomys Burmeister, 1888 (Rodentia, Cteno-myidae): Inferencias sobre su modo de vida. Estudios Geologi-cos 56:63–72.
Desbiez ALJ, Santos SA, Magalhaes Alvarez J, Moraes TomasW. 2011. Forage use in domestic cattle (Bos indicus), capibara(Hydrochoerus hydrochaeris) and pampas deer (Ozotoceros
Fig. 10. OSA cut following the chewing direction, with anhypothetic parasagittal profile through the upper molar row ofthe subterranean species C. leucodon (A) and the ground dwell-ing species T. dimidiatus (B). Dark gray blocks represent den-tine bulks; while light gray bars, the enamel edges. Note thestricking difference in the number of enamel leading and trail-ing edges (sensu Koenigswald et al., 1994).
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1159
Journal of Morphology

bezoarticus) ina seasonal Neotropical wetland. Mammal Biol76:351–357.
Eisenberg JF. 1981. The Mammalian Radiation. Chicago, USA:The University of Chicago Press.
El-Kouba, MMAN, Marques SMT, Pilati C, Hamann W. 2009.Presence of Faciola hepatica in feral nutria (Myocastorcoypus) living in a public park in Brazil. J Zoo Wildl Med40:103–106.
Felsenstein J. 1985. Phylogenies and the comparative method.Am Nat 125:1–15.
Freeman PW, Lemen CA. 2007. The trade-off tooth strengthand tooth penetration: Predicting optional shape of canineteeth. J Zool 273:273–280.
Garland T Jr, Harvey PH, Ives AR. 1992. Procedures for theanalysis of comparative data using phylogenetically independ-ent contrasts. Syst Biol 41:18–32.
Grafen A. 1992. The uniqueness of phylogenetic regression.J Theor Biol 156:405–423.
Greaves WS. 1982. A mechanical limitation on the position ofthe jaw muscle of mammals: The one-third rule. J Mammal63:261–266.
Guichon ML, Benıtez VB, Abba A, Borgnia M, Cassini MH.2003. Foraging behaviour of coypus Myocastor coypus: Whydo coypus consume aquatic plants? Acta Oecol 24:241–246.
Harvey PH, Pagel MD. 1991. The Comparative Method in Evo-lutionary Biology. New York, USA: Oxford University Press.
Herrera EA, Macdonald DW. 1993. Aggression, dominance, andmating success among capybara males (Hydrochaeris hydro-chaeris). Behav Ecol 4:114–119.
Hershkovitz P. 1958. A geographic classification of Neotropicalmammals. Fieldiana: Zoology, Vol. 36. Chicago, USA: ChicagoNatural History Museum Press; Num. 6.
Hildebrand MD. 1985. Digging of quadrupeds (pp 89-109). In:Hildebrand MD, Bramble M, Liem KF and Wake DB, Edi-tors. Functional Vertebrate Morphology. Cambridge, USA:Belknap Press.
Huchon D, Douzery EJP. 2001. From the old world to the newworld: A molecular chronicle of the phylogeny and biogeogra-phy of hystricognath rodents. Mol Phylogenet Evol 20:238–251.
Huxley J. 1932. Problems of Relative Growth. London:Methuen.
Janis CM. 1995. Correlations between craniodental morphologyand feeding behavior in ungulates: reciprocal illuminationbetween living and fossil taxa. In: Thomason J, editor. Func-tional Morphology in Vertebrate Palaeontology. Cambridge,USA: Cambrigde University Press.
Janis CM, Fortelius M. 1988. On the means whereby mammalsachieve increased functional durability of their dentitions,with special reference to limiting factors. Biol Rev 63:197–230.
Koenigswald Wv, Sanders PM, Leite MB, Mors T, Santel W.1994. Functional symmetries in the schmelzmuster and mor-phology of rootless rodent molars. Zool J Linn Soc 110:141–179.
Leggieri LR, Guichon ML, Cassini MH. 2011. Landscape corre-lates of the distribution of coypu Myocastor coypus (Rodentia,Mammalia) in Argentinean Pampas. Ital J Zool 78:124–129.
Lessa EP, Vassallo AI, Verzi DH, Mora MS. 2008. Evolution ofthe morphological adaptation for digging in living and extinctctenomyid and octodontid rodents (Rodentia: Caviomorpha:Octodontoidea). Biol J Linn Soc 95:267–283.
Loewe L. 2009. A framework for evolutionary systems biology.BMC Syst Biol 3:27–60.
Macdonald DW. 1981. Dwindling resources and the socialbehaviour of Capybara (Hydrochoerus hydrochaeris) (Mam-malia). J Zool 194:371–391.
Maddison WP, Maddison DR. 2005. Mesquite: A modularsystem for evolutionary analysis. Evolution, 62:1103–1118.
Mares MA, Ojeda RA. 1982. Patterns of diversity and adapta-tion in South American hystricognath rodents. (pp. 393–432).In: Mares MA, Genoways HH, editors. Mammalian Biologyin South America. Pymatuning Laboratory of Ecology, Special
Publication Series, 6. Pittsburgh, USA: University of Pitts-burgh.
Martin RA. 1993. Patterns of variation and speciation in Qua-ternary rodents (Chapter 11, pp. 226–280). In: Martin RA,Barnosky AD, editors. Morphological Change in Quaternarymammals of North America. Cambridge, USA: CambridgeUniversity Press.
Mascheretti S, Mirol PM, Gimenez MD, Bidau JC, ContrerasJR, Searle JB. 2000. Phylogenetics of the speciose and chro-mosomally variable genus Ctenomys (Ctenomyidae, Octodonti-dea) based on mitochondrial cytochrome b sequence. Biol JLinn Soc 70:361–376.
Midford PE, Garland T Jr, Maddison WP. 2003. PDAP:PDTREEpackage for Mesquite, version 1.00.
Mora MS, Olivares AI, Vassallo AI. 2003. Size, shape and struc-tural versatility of the skull of the subterranean rodent Cten-omys (Rodentia, Caviomorpha): functional and morphologicalanalysis. Biol J Linn Society, 78:85–96.
Morgan CC. 2009. Geometrics morphometrics of the scapula ofthe South American caviomorph rodents (Rodentia: Hystri-cognathi): form, function and phylogeny. Mammal Biol74:497–506.
Moxham BJ, Berkovitz BKB. 1995. The effects of externalforces on the periodontal ligament. In Berkovitz KB, MoxhamBJ, Newman HN, editors. The Periodontal Ligament inHealth and Disease, 2nd ed. London: Mosby-Wolf.
Nowak RM. 1999. Walker’s Mammals of the World, 6th ed. Bal-timore, USA: Johns Hopkins University Press.
Olivares AI, Verzi DH, Vassallo AI. 2004. Masticatory morpho-logical diversity and chewing modes in South American cavio-morph rodents (Family Octodontidae). J Zool 263:167–177.
Opazo JC. 2005. A molecular timescale for caviomorph rodents(Mammalia, Hystricognathi). Mol Phylogenet Evol 37:932–937.
Patterson BD, Velazco PM. 2006. A distinctive new cloud-forestrodent (Hystricognathi: Echimyidae) from the Manu Bio-sphere Reserve, Peru. Mastozoologıa Neotropical 13:175–191.
Perez-Barberıa FJ, Gordon IJ. 1998. Factors affecting food com-minution during chewing in ruminants: A review. Biol J LinnSoc 63:233–256.
Peters RH. 1983. The Ecological Implications of Body Size.Cambridge, USA: Cambridge University Press.
Redford KH, Eisenberg JF. 1992. Mammals of the Neotropics.Volume 2: The Southern Cone: Chile, Argentina, Uruguay,Paraguay. Chicago, USA: The University of Chicago Press.
Rocha-Barbosa O, Casinos A. 2011. Geometry and evolutionaryparallelism in the long bones of cavioid rodents and smallartiodactyls. J Biosci, 36:1–9.
Rohlf FJ. 2006. TpsDig, Version 2.1. SUNY, Department of Ecol-ogy and Evolution, State University of New York, StonyBrook, NY.
Sloan P, Carter DH. 1995. Structural organization of the fibersof the periodontal ligament. In: Berkovitz KB, Moxham BJ,Newman HN, editors. The Periodontal Ligament in Healthand Disease, 2nd ed. London: Mosby-Wolf.
Smith RJ. 2009. Use and misuse of the reduced major axis forline-fitting. Am J Phys Anthropol 140:476–486.
Sokal RR, Rohlf FJ. 1995. Biometry: The Principles and Prac-tice of Statistics in Biological Research, 3rd ed. San Fran-cisco, USA: WH Freeman and Co.
Spotorno AE, Valladares JP, Marin JC, Palma E, Zuleta RC.2004. Molecular divergence and phylogenetic relationships ofchinchillids (Rodentia: Chinchillidae). J Mammal 85:384–388.
Stein BR. 2000. Morhology of subterranean rodents. In: LaceyEA, Patton JL, Cameron GN, editors. Life Underground: TheBiology of Subterranean Rodents. Chicago, USA: The Univer-sity of Chicago Press, pp.19–61.
Tomkins JL, Kotiaho JS, LeBas NR. 2005. Phenotypic plasticityin the developmental integration of morphological trade-offsand secondary sexual trait compensation. Proc R Soc B272:543–551.
Ungar P. 2010. Mammal Teeth: Origin, Evolution, and Diver-sity. Baltimore, USA: The Johns Hopkins University Press.
1160 F. BECERRA ET AL.
Journal of Morphology

Vassallo AI. 1998. Functional morphology, comparative behav-iour, and adaptation in two sympatric subterranean rodentsgenus Ctenomys (Caviomorpha: Octodontidae). J Zool (Lond)244:415–427.
Vassallo AI. 2000. Alometrıa e isometrıa en varias especies deroedores caviomorfos con comentarios sobre la estructura delaparato trofico en el Orden Rodentia. Mastozoologıa Neotropi-cal 7:37–46.
Vassallo AI, Echeverrıa AI. 2009. Evolution of brain size in ahighly diversifying linaje of subterranean rodent genus Cten-omys (Caviomorpha: Ctenomyidae). Brain Behav Evol73:138–149.
Vassallo AI, Verzi DH. 2001. Patrones craneanos y modalidadesde masticacion en roedores caviomorfos. Boletın de la Socie-dad de Biologıa de Concepcion, 72:139–145.
Verzi DH. 1999. The dental evidence on the differentiation ofthe ctenomyine rodent. Acta Theriol 44:263–282.
Verzi DH. 2001. Phylogenetic position of Abalosia and the evo-lution of the extant Octodontinae (Rodentia, Caviomorpha,Octodontidae). Acta Theriol 46:243–268.
Verzi DH. 2002. Patrones de evolucion morfologica en Cteno-myinae (Rodentia, Octodontidae). Mastozoologıa Neotropical9:309–328.
Verzi DH, Alvarez A, Olivares AI, Morgan CC, Vassallo AI.2010. Ontogenetic trajectories of key morphofunctional cra-nial traits in South American subterranean ctenomyidrodents. J Mammal 91:1508–1516.
Verzi DH, Vieytes EC, Montalvo CI. 2004. Dental evolution inXenodontomys and first notice on secondary acquisition of ra-dial enamel in rodents (Rodentia, Caviomorpha, Octodonti-dae). Geobios 37:795–806.
Vieytes EC. 2003. Microestructura del esmalte de roedores hys-tricognathi sudamericanos fosiles y vivientes. Significadomorfofuncional y filogenetico. Unpublished D. Phil. Thesis.Facultad de Ciencias Naturales y Museo, Universidad Nacio-nal de La Plata, Argentina.
Vizcaıno SF, Bargo MS, Cassini GH. 2006. Dental occlusal sur-face area in relation to BM, food habits and other biologicalfeatures in fossil xenarthrans. Ameghiniana 43:11–26.
Vucetich MG, Verzi DH. 1991. Un nuevo Echimyidae (Rodentia,Hystricognathi) de la Edad Colhuehuapense de Patagonia yconsideraciones sobre la sistematica de la familia. Ameghini-ana 28:67–74.
Vucetich MG, Verzi DH. 1995. Los roedores caviomorfos. In:Alberdi MT, Leone G, Tonni EP, editors. Evolucion Biologicay Climatica de la Region Pampeana Durante los UltimosCinco Millones de Anos. Un Ensayo de Correlacion con elMediterraneo Occidental. Madrid, Spain: Monografıas CSCIC12.213–225.
Vucetich MG, Verzi DH, Hartenberger JL. 1999. Review andanalysis of the radiation of the South American Hystricogna-thi (Mammalia, Rodentia). Comptes Rendus de l’Academiedes Sciences IIA 329:763–769.
Walker WF, Liem KF. 1994. Functional Anatomy of the Verte-brates: An Evolutionary Perspective. Philadelphia: SaundersCollege Publishing. 788 pp.
Warton DI, Wright IJ, Falster DS, Westoby M. 2006. Bivariateline-fitting methods for allometry. Biol Rev Camb Philos Soc81:259–291.
Woods CA. 1972. Comparative myology of jaw, hyoid, and pecto-ral appendicular regions of the new and old world hystrico-morph rodents. Bull Am Mus Nat Hist 147:115–198.
Wyss AR, Flynn JJ, Norell MA, Swisher CC, Charrier R, Nova-cek MJ, McKenna MC. 1993. South America’s earliest rodentand recognition of a new interval of mammalian evolution.Nature 365:434–437.
Zajicek G. 1976. The rodent incisor tooth proliferon. Cell TissueKinet 9:207–214.
APPENDIX A
Raw values for all the variables used in thisstudy. (*) Molar rows without the M3 (fallen pi-ece).
Variables
Body mass (BM). Upper incisors’ variables:Cross-sectional area (CA); second moment of iner-tia (I); polar moment of inertia (J); root length(IRL); and procumbency angle (PA). Upper molars’variables: Occlusal surface area (OSA); enamelband length (EL); and enamel index (EI).
Family SpeciesBM(g)
CA(mm2)
I(mm4)
J(mm4)
IRL(mm)
PA(radians)
OSA(mm2)
EL(mm) EI
Erethizontidae E. dorsatum 5,000.0 23.522 32.919 98.506 34.796 1.663 46.928 208.889 4.646Sphiggurus spinosus 750.0 7.054 2.473 9.499 19.851 1.672 14.941 121.165 8.717
Chinchillidae L. maximus 3,950.0 19.400 27.639 74.266 33.807 1.829 85.070 103.959 1.231Lagidium viscaccia 2,150.0 8.233 4.706 11.301 17.140 1.465 34.434 92.885 2.805Chinchilla lanigera 500.0 4.153 1.133 3.731 11.616 1.506 13.497 71.357 5.628
Myocastoridae M. coypus 4,700.0 35.867 87.803 219.944 42.425 1.884 52.105 210.727 4.189Echimyidae Trinomys dimidiatus 235.0 3.441 0.549 2.275 16.529 1.281 3.467 62.224 16.185
Echimys dasythrix 260.0 2.440 0.329 1.120 10.278 1.208 9.231 73.419 8.470Thrichomys apereoides 275.0 3.659 0.698 2.517 15.793 1.211 4.465 51.078 10.954Thrichomys
cunicularius285.0 2.622 0.326 1.313 13.815 1.300 4.326 51.920 11.436
Proechimys albispinus 200.0 3.120 0.387 2.034 11.933 1.597 3.539 44.374 11.011Proechimys guainae 285.0 2.916 0.420 1.592 14.214 1.296 3.385 51.168 13.602
Ctenomyidae Actenomys sp. 1,360.0 9.659 6.070 20.822 28.203 1.998 31.799 50.564 1.623Ctenomys magellanicus 234.8 8.479 4.850 12.482 20.705 1.743 15.202 34.100 2.341Ctenomys australis 371.1 7.896 4.734 10.993 18.289 1.672 15.081 33.664 2.328Ctenomys talarum 134.6 5.056 1.948 4.362 14.199 1.632 8.727 26.043 3.087Ctenomys opimus 218.3 6.370 3.350 6.712 17.464 1.675 20.085(*) 34.680(*) (*)C. leucodon 220.0 6.678 3.585 7.434 22.248 1.967 15.339 33.396 2.268Ctenomys tuconax 500.0 10.180 9.205 14.769 19.421 1.756 20.591 37.804 1.898
Octodontidae O. gliroides 105.0 3.663 0.682 2.557 11.751 1.570 5.659 26.383 4.697S. cyanus 83.0 3.268 0.586 2.062 18.103 1.999 2.879 27.813 8.071Octomys mimax 131.0 3.181 0.469 1.972 13.083 1.375 3.240 30.714 8.269
SCALING OF DENTAL TRAITS IN CAVIOMORPH RODENTS 1161
Journal of Morphology

Family SpeciesBM(g)
CA(mm2)
I(mm4)
J(mm4)
IRL(mm)
PA(radians)
OSA(mm2)
EL(mm) EI
Tympanoctomysbarrerae
82.0 2.399 0.391 0.943 10.588 1.405 0.536 23.554 13.305
Caviidae Kerodon rupestris 950.0 3.855 0.793 2.792 12.791 1.399 15.684 61.245 4.055Galea musteloides 225.0 1.709 0.166 0.564 7.876 1.505 13.525 50.182 3.861Galea spixii 220.0 2.140 0.246 0.894 11.292 1.407 13.096 49.240 3.888Dolichotis patagonum 9,400.0 14.215 11.933 36.985 25.750 1.199 54.271 104.890 1.971C. aperea 496.0 2.854 0.414 2.154 11.753 1.793 19.886 72.341 3.835C. porcellus 425.0 1.858 0.160 1.038 8.416 1.887 14.391 63.826 4.675Microcavia australis 296.0 1.976 0.191 0.780 10.764 1.564 11.250 49.452 4.519
Hydrochaeridae Hydrochoerushydrochaeris
50,000.0 79.541 403.784 1,113.666 48.502 1.506 462.557 598.590 1.297
Dasyproctidae Agouti paca 8,700.0 20.928 20.694 82.629 44.382 1.232 47.136 209.730 4.635Dasyprocta sp. 2,700.0 11.451 7.204 23.373 30.358 1.390 20.657 147.952 7.679
1162 F. BECERRA ET AL.
Journal of Morphology





























