Embed Size (px)
Citation preview

27
Vox Sanguinis
(2007)
93
, 27–36
ORIGINAL PAPER
©
2007 The Author(s)Journal compilation
©
2007 Blackwell Publishing Ltd.DOI: 10.1111/j.1423-0410.2007.00912.x
Blackwell Publishing Ltd
Storage of buffy-coat-derived platelets in additive solutions at 4
°°°°
C and 22
°°°°
C: flow cytometry analysis of platelet glycoprotein expression
P. Sandgren, M. Hansson, H. Gulliksson & A. Shanwell
Clinical Immunology and Transfusion Medicine, Karolinska University Hospital, Stockholm, Sweden
Background
The aim of our
in vitro
study is to compare the effects on platelet membraneglycoproteins that play an important role in the main functions of platelets, whenplatelets are stored for a period of 21 days at 4
°
C or 22
°
C.
Study Design and Methods
Platelet concentrates (PC) were prepared from pooledbuffy-coats (BC) for paired studies (total eight pools from 80 BCs) by using theOrbiSac system. We divided each pool into two PCs and stored them at 4
°
C or 22
°
C.
Results
The activation marker CD62 remained almost unchanged during storage inall units. The expression of CD63 was higher in PCs stored at 22
°
C than in thosestored at 4
°
C
.
No significant difference in CD41 expression was detected over time.The expression of CD42b declined during storage and even more in PCs stored at 4
°
Cuntil day 21 [day 14: mean flourscence intensity: 32·5 ± 13·1 vs. 46·5 ± 19·1], but thepercentage of platelets expressing CD42b remained high in platelets stored at 4
°
C,but gradually decreased at 22
°
C (day 14: 95·0 ± 1·5 vs. 59·0 ± 9·9). Storage at 4
°
Creduced the rate of glycolysis and maintained the pH better after day 10 than in PCsstored at 22
°
C (day 14: 7·009 ± 0·067 vs. 7·233 ± 0·125). The concentration ofregulated upon activation of normal T-cells expressed and secreted was higher in PCsstored at 22
°
C than at 4
°
C (day 7: 414·7 ± 32·3 vs. 49·6 ± 19·0). No response toextent of shape change and no swirling were detected at 4
°
C.
Conclusion
Platelets stored at 4
°
C retain their
in vitro
characteristics better thanthose stored at 22
°
C, except for parameters that reflect changes in shape. Storage at4
°
C is not associated with an increased expression of glycoprotein (GpIb, GpIIb/IIIa)and platelet activation markers (CD62p and CD63) as compared with storage at 22
°
C.
Key words:
buffy coat, flow cytometry,
in vitro
, platelet glycoprotein expression,
platelet storage, temperature.
Introduction
Platelets are traditionally stored at 22
°
C, but this situationmay be reconsidered. Storage of platelets at room temperatureaccommodates bacterial growth, and bacterial sepsis is nowregarded as the main risk of transfusion-transmitted diseases
[1]. Storage at 4
°
C would reduce not only the risk of bacterialgrowth, but may also delay the impairment of platelets.Changes induced by processing platelet concentrates (PC)and during storage are frequently referred to as the plateletstorage lesion [2–4].
Recent studies suggest that platelets stored in additivesolution (T-Sol) at 4
°
C without agitation maintain acceptablemetabolic and cellular
in vitro
characteristics during 21 daysof storage [5]. However, several vital questions need to beassessed about platelet storage at 4
°
C, especially whetherchanges in morphology (a loss of discoid shape) at 4
°
C are
Correspondence
: Per Sandgren, Transfusion Medicine, C2 58, Karolinska University Hospital, 141 86 Stockholm, SwedenE-mail: [email protected]
Received: 3 October 2006, revised 14 February 2007, accepted 15 February 2007

28
P. Sandgren
et al.
©
2007 The Author(s)Journal compilation
©
2007 Blackwell Publishing Ltd.,
Vox Sanguinis
(2007)
93
, 27–36
related to increased expression of platelet membrane proteinsand activation markers during long-term storage. Previouswork has shown that refrigeration of platelets increasescytosolic calcium, actin polymerization and shape change[6,7]. As inside-out activation is Ca
2+
dependent and involveschanges in the conformations of both the ligand-bindingextracellular region and the cytoplasmic tails, we suggestthat inside-out signalling and cytoskeletal rearrangementseventually result in increased expression of receptors andactivation markers during prolonged storage at 4
°
C.Flow cytometry analysis has been used to study platelet
glycoproteins which play an important role in adhesion,aggregation and to evaluate activation of stored platelets [8].P-selectin (CD62p), a constituent of
α
-granules in restingplatelets, can be detected on the activated platelet surfaceafter
α
-granule secretion [9]. Anti-CD63 reacts with lysosomalmembrane-linked glycoprotein 3, subsequently identified asgranulophysin, a platelet dense granule glycoprotein suggestedbeing a marker for granule release [10]. CD41 (GPIIb), whichis usually linked with CD61 (GPIIIa), forms the GPIIb/IIIacomplex. This complex seems to be involved in plateletaggregation, serves as a receptor for soluble fibrinogen,fibrinonectin, thrombospondin, von Willebrand factor (vWF)and vitronectin [11] and plays a main role in the regulation ofplatelet aggregation and adhesion throughout haemostasis[12].
During platelet activation, the expression level of CD61/CD41 increases on the platelet surface, a conformational changethen occurs, which creates the active binding site for fibrinogenand vWF [13].
Recently, the importance of vWF-dependent adhesionreceptor (GPIb
α
) in maintaining platelet viability
in vivo
hasbeen clearly shown in murine models [14]. When platelets arestored in a cold environment, the vWF receptor (vWFR)(Gp1b
α
or CD42b) rearranges irreversibly to form a clusteron the surface of the platelets. After transfusion of theseplatelet units, the integrin receptor of hepatic macrophages,recognizes these clusters and removes the platelets from thecirculation [15]. The reason for storing platelets at 4
°
C is toprevent bacterial contamination during storage for more than5 days. To date, there is some evidence that Hoffmeister
et al
.’sapproach, which suggests that galactosylation with uridine51-diphosphogalactose of platelets can inhibit lectin bindingbefore or after cooling [14], allows for storage of humanplatelets in the cold and provides the protective effect on coldstorage of human platelets, as well as prolonging the survivaltime of refrigerated human platelets in the rabbit model [16].
An attempt to tackle the practical problem of how torefrigerate platelets for transfusion can be divided into twoseparate problems, the ‘clearance problem’ and the ‘storageproblem’.
Both the clearance problem, which involves a shortercirculation time after transfusion, and the storage problem
have been well known since the 1970s, especially the detri-mental changes in terms of
in vivo
behaviour when the storagetime at 4
°
C was longer than 24 h [17,18]. Since the 1970s,considerable improvements have been made concerningprocessing and storage of platelets, resulting in longer survivaland better function [19–22], and providing an option forprolonged storage at 4
°
C. As uridine 51-diphosphogalactosemay provide a solution concerning the clearance problem,further evaluation of the ability to store platelets for a prolongedtime at 4
°
C under modern human blood banking conditionsis needed.
The aim of our
in vitro
study is to compare the effects ofstoring platelets at 4
°
C or 22
°
C on platelet glycoproteinsthat play an important role in the main functions of platelets,and to determine the metabolic and cellular differencesbetween those platelets.
Materials and methods
Preparation and storage of platelet concentrates
Platelet concentrates were prepared from pooled buffy-coats(BC) for paired studies (Fig. 1). A total of 450 ml whole blood wascollected from healthy blood donors who met standard donationcriteria and gave their written informed consent in accordancewith our institution’s guidelines. We used a quadruple-bagblood container system (Imuflex-CRC, Terumo, Tokyo, Japan)made of polyvinylchloride with di(2-ethylhexyl)phthalate as
Fig. 1 Paired study design: preparation of PCs from pooled buffy-coats by
the OrbiSac system and division of the primary pool into two equal parts.

©
2007 The Author(s) Journal compilation
©
2007 Blackwell Publishing Ltd.,
Vox Sanguinis
(2007)
93
, 27–36
Flow cytometry analysis of platelets stored at 4
°
C
29
plasticizer) containing 63 ml of standard citrate phosphatedextrose. The whole-blood containers were kept at roomtemperature (20–24
°
C) by cooling plates (Sebra, Tucson, AZ,USA) and were centrifuged at 2700
g
for 10 min at 22
°
Cwithin 8 h of collection. Centrifugation was immediatelyfollowed by separation into red blood cells, plasma and BCswith automated equipment, Terumo automatic componentextractor (T-ACE, Terumo). Buffy-coats were kept over nightwithout agitation at room temperature (20–24
°
C).An automated technique (OrbiSac System, Gambro BCT, Inc.,
Lakewood, NJ, USA) was used to prepare the platelets frompooled ABO-identical BCs. In the first sequence, pooling offive BCs and 300 ml of platelet additive solution (PAS) (T-Sol, Baxter), which consists of 115·5 mmol/l NaCl, 30 mmol/lacetate, 10 mmol/l citrate, pH 7·2 were pooled in a ring-shaped container, is performed. This step involves washingof the BC containers with PAS. In the second sequence, thecontents of the ring-shaped pooling container are mixed andcentrifuged, and the platelet-rich supernatant is then passedthrough a leucocyte-reduction filter (LRP 6, PALL, Portsmouth,UK) into a container in the centre of the centrifuge. This processis monitored by a system of pressure sensors and photocells [23].
Two ABO-identical PCs, prepared according to this procedure,were pooled in a primary pool. A total of eight primary poolswere obtained from 80 BCs. Each primary pool was againdivided into two equal subunits, which were stored underdifferent conditions. One PC was stored on a flat bed agitator(60 cycles a minute, model LPR-3, Melco, Glendale, CA, USA)in a temperature-controlled cabinet at 22 ± 2
°
C while theother was stored at 4 ± 2
°
C without agitation. The sampleswere drawn aseptically on days 1, 5, 7, 10, 14 and 21.
All sampling was done by sterile connection (TSCD-II,Terumo) of sampling bags to the respective containers (OrbiSacstandard BC set, storage bag, Gambro BCT, Inc.). The storagebag is made of polyvinylchloride plastic treated with citrateand intended to contain up to 5
×
10
11
platelets in 400 ml ofplatelet storage medium.
Monoclonal antibody labelling
Platelet concentrate samples, fixed by adding an equal volumeof 1% paraformaldehyde (PFA), PFA-phosphate-bufferedsaline (pH 7·2–7·4) at 22
°
C for 10 min, were then stained for20 min at the same temperature in the dark by incubatingwith 20
µ
l of fluorochrome-labelled monoclonal antibodies(mAb) per 1
×
10
6
platelets. The following mAbs were usedfor single-colour staining: fluorescein isothiocyanate (FITC)-conjugated (IgG1 isotype) CD41 (clone P2), CD42b (cloneSZ2), CD61 (clone SZ21), phycoerythrin (PE)-conjugated(IgG1 isotype) CD63 (clone CLB-gran12) and CD62p (P-selectin/GMP-140/platelet activation dependent granule-externalmembrane protein; clone CLB Thromb/6), all purchased fromImmunotech (Beckman Coulter, Marseilles, France). Control
specimens were processed as above, but incubated with aFITC- or PE-conjugated mAb (IgG1 isotype) with irrelevantspecificity, purchased from Immunotech (Beckman Coulter).After incubation with fluorochrome-conjugated antibodies,the samples were washed twice by adding 2·0 ml filteredPBS-ethylenediaminetetraacetic acid (PBS-EDTA) with 0·1%Na-azid, and were centrifuged at 2760
g
(Eppendorf 5810R)for 10 min at 18
°
C.
Flow cytometry analysis
A total of 100 000 platelet events were acquired on an FACSCalibur flow cytometer (Becton Dickinson, Franklin Lakes, NJ,USA) equipped with 15 mW argon ion lasers and CellQuestPro software. Daily controls of optics and fluorescence intensitywas performed using standardized beads (CaliBrite; BDBioscience, San Jose, CA, USA). The flow cytometer settingswere optimized for the acquisition of platelets by logarithmicsignal amplification in all four detectors (forward and sidescatter channels and fluorescence channels FL1 and FL2). Foranalysis, a gate was set around intact platelet populationas defined by forward and side scatter characteristics. Thepercentage of positive platelets of total platelet expressingactivation markers (CD62p and CD63) and surface membraneglycoproteins (CD41, CD42b and CD61) above that of background(negative control) as well as the mean flourescence intensity(MFI) was recorded.
Analysis of metabolic and cellular parameters
The metabolic and cellular differences between platelets storedat 4
°
C or 22
°
C were evaluated in an
in vitro
platelet-storagestudy, including measurements of platelet counts (10
9
perlitre and 10
9
per unit) and mean platelet volume (MPV) usingroutine methods (CA 620 Cellguard, Boule Medical, Stock-holm, Sweden). The volume (ml) was calculated by weighingthe contents of the storage bag, in grams, on a scale (MettlerPB 2000) and the result, in grams, was divided by 1·01(1·01 g/ml is the density of the storage medium composed ofapproximatly 70% PAS and 30% plasma). By use of routinemethods (ABL 705, Radiometer, Copenhagen, Denmark) wealso measured the pH, oxygen tension (pO
2
) (kPa at 37
°
C),partial pressure of carbon dioxide (pCO
2
) (kPa at 37
°
C), glucose(mmol/l) and lactate (mmol/l). Bicarbonate (mmol/l) wascalculated based on other measured variables.
The assessment of swirling was done by inspection andgrading with Bertolini’s method [24]. The white blood cellcount on day 1 was determined with a Nageotte chamber anda microscope (Zeiss, standard) [25]. Hypotonic shock response(HSR) reactivity and the extent of shape change (ESC) meas-urements were performed using a dedicated microprocessor-based instrument (SPA 2000, Chronolog, Havertown, PA,USA) with the modifications of these tests described by

30
P. Sandgren
et al.
©
2007 The Author(s)Journal compilation
©
2007 Blackwell Publishing Ltd.,
Vox Sanguinis
(2007)
93
, 27–36
VandenBroeke
et al
. [26]. The total adenosine triphosphate(ATP) concentration (
µ
mol per 10
11
) was determined with aLuminometer (Orion Microplate Luminometer, BertholdDetection Systems GmbH, Pforzheim, Germany) on the basisof principles described by Lundin [27]. Lactate dehydrogenase(LDH) activity, a marker for disintegration of platelets, wasmeasured with a spectrophotometric method (Sigma Aldrichkit 063K6003; Spectrophotometer DMS 100, Varian Techtron,Springvale, Vic., Australia) [28].
Cytokine analysis
Samples from the PCs were collected in pre-chilled citratetheophylline adenosine dipyridamole (CTAD) tubes. Thesamples from the collected PCs were centrifuged at 2500
g
per relative centrifugal force (Eppendorf 5810R) for 30 min at10
°
C and the supernatants were stored at
−
70 °C pendinganalysis. Regulated upon activation of normal T-cells expressedand secreted (RANTES) PCs were determined with commercialenzyme-linked immunosorbent assay (ELISA) kits in accordancewith the manufacturer’s recommendations (Quantikine R&DSystem, Abingdon, UK).
The standards for RANTES were those provided by themanufacturer. We used the following ranges for measurements inthe assay: 31·2–2000 pg/ml. Dilution and re-assay of specimenswere done as required. All samples and standards were assayedin duplicate. All results are given in pg per 106 platelets.
Bacterial cultures
Bacterial cultures were performed at day 21 with the routinebacterial culture methods in the Bacteriological Laboratoryat Karolinska University Hospital.
Statistical analyses
The mean values and standard deviations are usually given.Repeated measurement analysis of variance was performedon data. Two different groups were studied over time (days).
‘Days’ was the repeated factor and ‘group’ a between factor.The P value for the interaction term between ‘days’ and‘group’ is reported, as well as the 95% confidence interval (CI)as a measure of dispersion of the results. The analyses werecarried out in the Statistica software, version 14·0 (SPSS,Chicago, IL, USA).
Results
In this study, platelet units (n = 8) stored at 4 °C withoutagitation were compared with platelet units stored at 22 °Cwith agitation. Over a storage interval of 21 days we foundsignificant differences in a variety of flow cytometry, cellularand metabolic parameters between the various platelet-storageenvironments.
Flow cytometry analysis
The activation marker CD62p (Fig. 2) remained almostunchanged during storage in all units. The expression ofCD63 (Fig. 3) was higher in PCs stored at 22 °C than in thosestored at 4 °C. No significant difference in CD41 expressionwas detected (Fig. 4). The surface expression of CD61 (Fig. 5)was higher in PCs stored at 22 °C than at 4 °C. The expressionof CD42b (Fig. 6) decreased during storage in all units (MFI),but was higher in those stored at 22 °C than at 4 °C, with theexception of day 21. The percentage of platelets expressingCD42b remained high in PCs stored at 4 °C but graduallydecreased in PCs stored at 22 °C (Fig. 7).
Cellular assays
The platelet counts and contents on day 1 are given inTable 1. During storage the platelet content declined more inPCs stored at 22 °C than in those stored at 4 °C (508 × 109 ±146 per litre and 674 × 109 ± 80 per litre, respectively, on day21) (data not shown). Mean platelet volume and LDH activityincreased more when platelets were stored at 22 °C thanwhen they were stored at 4 °C (Table 2).
Fig. 2 CD62p expression (mean fluorescence intensity) of PCs stored in PAS II for 21 days during various storage conditions: mean ± 95% confidence interval
(CI); 22 °C (squares) and 4 °C (triangles) (n = 8). No significant difference from baseline (day 1) between groups (heterogenicity) during 21 days was detected.

© 2007 The Author(s) Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
Flow cytometry analysis of platelets stored at 4 °C 31
Fig. 3 CD63 expression (mean fluorescence intensity) of PCs stored in PAS II for 21 days during various storage conditions: mean ± 95% confidence interval
(CI); 22 °C (squares) and 4 °C (triangles) (n = 8). Significant difference from baseline (day 1) between groups (heterogenicity) during 21 days was detected
(P = 0·013).
Fig. 4 CD41 expression (mean fluorescence intensity) of PCs stored in PAS II for 21 days during various storage conditions: mean ± 95% confidence interval
(CI); 22 °C (squares) and 4 °C (triangles) (n = 8). No significant difference from baseline (day 1) between groups (heterogenicity) during 21 days was detected.
Fig. 5 CD61 expression (mean fluorescence intensity) of PCs stored in PAS II for 21 days during various storage conditions: mean ± 95% confidence interval
(CI); 22 °C (squares) and 4 °C (triangles) (n = 8). Significant difference from baseline (day 1) between groups (heterogenicity) during 21 days was detected
(P < 0·001).
Fig. 6 CD42b expression (mean fluorescence intensity) of PCs stored in PAS II for 21 days during various storage conditions: mean ± 95% confidence interval
(CI); 22 °C (squares) and 4 °C (triangles) (n = 8). Significant difference from baseline (day 1) between groups (heterogenicity) during 21 days was detected
(P < 0·001).

32 P. Sandgren et al.
© 2007 The Author(s)Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
Metabolic assays
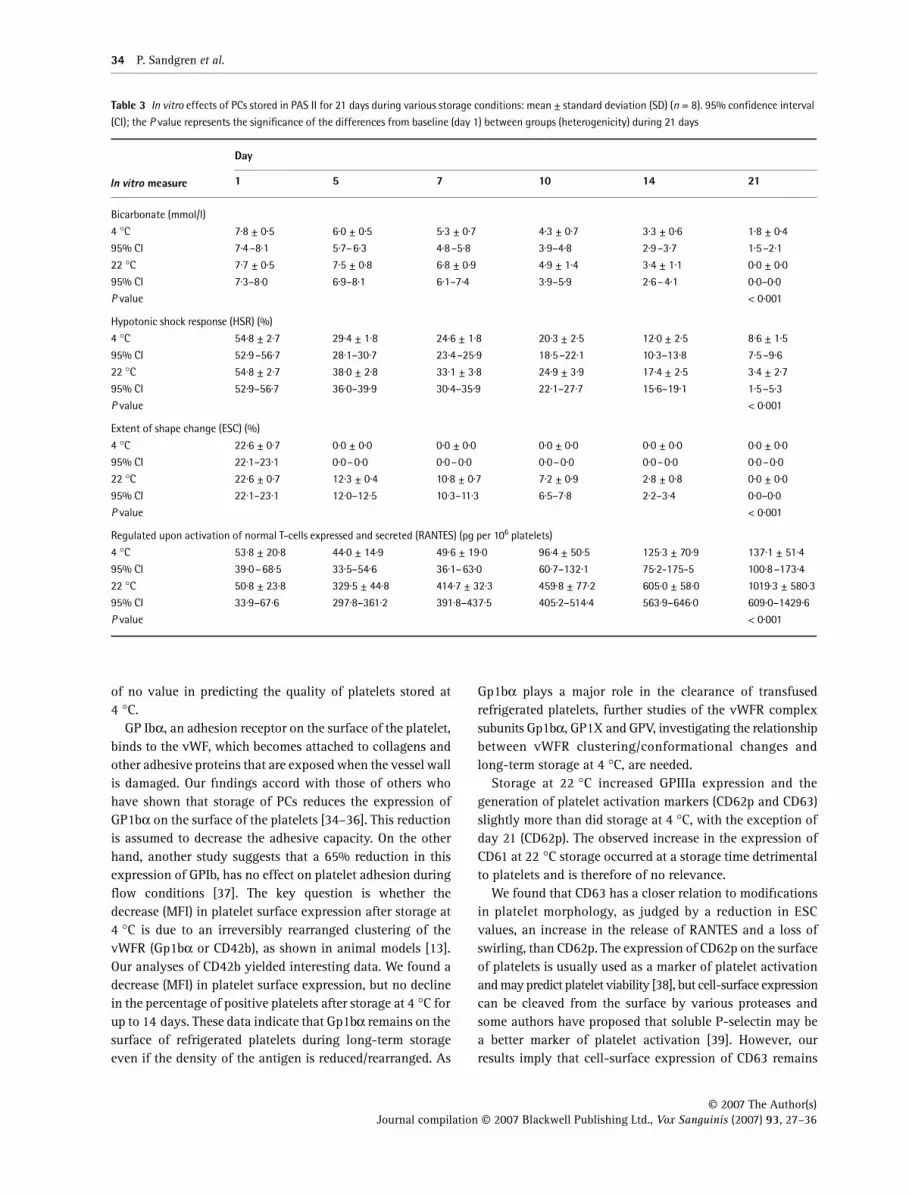
Glucose declined less and was present until day 21 at 4 °Cand lactate increased less in PCs stored at 4 °C than at 22 °C(Table 2). pH, ATP and pCO2 were better maintained after day10 in PCs stored at 4 °C than at 22 °C (Table 2). The oxygentension (pO2) was higher in PCs stored at 4 °C than at 22 °C(Table 2). Bicarbonate was better maintained from day 14 inPCs stored at 4 °C than at 22 °C (Table 3).
HSR, ESC and swirling
HSR was higher in PCs stored at 22 °C until day 21 and ESCwas higher in PCs stored at 22 °C than at 4 °C during theentire storage period (Table 3). Swirling was fully maintainedfor 7 days in PCs stored at 22 °C. No swirling was observedin those stored at 4 °C (data not shown).
Cytokine analysis
The concentrations of RANTES (Table 3) increased duringstorage in all units, but were higher in PCs stored at 22 °Cthan at 4 °C.
All bacterial cultures were negative.
Discussion
In this study, platelets stored at 4 °C seemed to be moremetabolically stable than platelets stored at 22 °C duringlong-term storage, with the exception of parameters associatedwith changes in shape. As mentioned in the literature, platelets
become activated over the storage time when stored at 22 °C[29,30], and platelet activation is usually associated with anincrease in the metabolic rate due to glucose consumption,lactate production and release of α-granule content and mayoccur together with a substantial loss of platelet viability [31].This is confirmed by our data. In contrast, at 4 °C storage weshow that platelets are less metabolically active. Also, thereis less lactate accumulation, no decline in pH and a reductionin the release of α-granule content. This study resembles ourprevious results [5] with regard to various aspects of metabolismand cellular quality after storage at 4 °C. Our main findingssuggest that the lower temperature is not associated withincreased expression of glycoprotein (e.g. GpIb, GpIIb/IIIa)and generation of platelet activation markers (CD62p andCD63) as compared with storage at 22 °C.
Platelets stored at 4 °C show characteristic morphologicalchanges related to activation, from a discoid to a spherocyticshape, so-called cold-induced activation [14,7]. Is thischange in shape after storage at 4 °C of the same importanceas in platelets stored at 22 °C? For platelets stored at roomtemperature, the ESC assay and the HSR assay are regardedas fairly good predictors of platelet recovery and survival[32]. However, no in vitro test can predict the recovery andsurvival in vivo [7]. In the present and previous study [5], weshowed that platelets stored at 4 °C perform poorly in thesetests as compared with those stored at room temperature.However, Hoffmeister et al. (2003) in animal models foundthat a spherocytic shape alone is insufficient to cause a rapidclearance of platelets [13]. Some authors claim that non-discoidplatelets may survive after transfusion and even revert to adiscoid shape [33]. Therefore the ESC and HSR assays may be
Fig. 7 CD42b expression (the percentage of positive platelets of total platelet expression CD42b) of PCs stored in PAS II for 21 days during various storage
conditions: mean ± 95% confidence interval (CI); 22 °C (squares) and 4 °C (triangles) (n = 8). Significant difference from baseline (day 1) between groups
(heterogenicity) during 21 days was detected (P < 0·001).
Table 1 Comparison of various storage conditions for platelet concentrates on day 1. Results are expressed as mean ± standard deviation, n = 8
Storage condition Volume (ml) Platelets (109 per l) Platelets (109 per units) Leucocytes (106 per units)
4 °C without agitation 381 ± 15 756 ± 79 287 ± 25 < 0·2
22 °C with agitation 381 ± 13 741 ± 75 281 ± 24 < 0·2

© 2007 The Author(s) Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
Flow cytometry analysis of platelets stored at 4 °C 33
Table 2 In vitro effects of PCs stored in PAS II for 21 days during various storage conditions: mean ± standard deviation (SD) (n = 8). 95% confidence interval
(CI); the P value represents the significance of the differences from baseline (day 1) between groups (heterogenicity) during 21 days
In vitro measure
Day
1 5 7 10 14 21
Mean platelet volume (MPV) (fl)
4 °C 8·6 ± 0·4 9·3 ± 0·6 9·4 ± 0·7 9·0 ± 0·7 8·8 ± 0·5 8·8 ± 0·5
95% CI 8·3–8·9 8·9–9·8 8·9–9·9 8·5–9·5 8·4–9·2 8·5–9·2
22 °C 8·4 ± 0·5 8·0 ± 0·4 8·4 ± 0·5 9·1 ± 0·5 10·6 ± 0·7 12·6 ± 1·7
95% CI 8·1–8·8 7·7–8·3 8·1–8·8 8·7–9·4 10·1–11·1 11·4–13·8
P value < 0·001
Lactate dehydrogenase activity (LDH) (%)
4 °C 5·7 ± 0·5 6·0 ± 0·8 6·1 ± 1·7 7·5 ± 1·7 9·1 ± 1·1 10·6 ± 1·1
95% CI 5·3–6·0 5·5–6·6 4·9–7·3 6·3–8·7 8·3–9·9 9·8–11·3
22 °C 5·7 ± 0·5 6·4 ± 1·1 7·1 ± 1·5 7·8 ± 1·4 9·0 ± 1·0 29·3 ± 15·3
95% CI 5·3–6·0 5·6–7·2 6·0–8·1 6·8–8·8 8·2–9·7 18·5–40·1
P value < 0·001
Glucose (mmol/l)
4 °C 6·3 ± 0·4 5·6 ± 0·5 5·3 ± 0·6 4·6 ± 0·7 4·0 ± 0·7 2·9 ± 0·7
95% CI 6·0–6·6 5·3–6·0 4·8–5·7 4·1–5·1 3·5–4·5 2·4–3·4
22 °C 6·1 ± 0·8 4·9 ± 0·7 3·5 ± 0·9 1·6 ± 1·4 0·0 ± 0·0 0·0 ± 0·0
95% CI 5·5–6·7 4·4–5·4 2·8–4·1 0·6–2·6 0·0–0·0 0·0–0·0
P value < 0·001
Lactate (mmol/l)
4 °C 4·8 ± 0·4 6·4 ± 0·4 7·2 ± 0·5 8·1 ± 0·7 9·4 ± 0·6 11·0 ± 0·8
95% CI 4·5–5·0 6·0–6·7 6·9–7·6 7·6–8·6 9·0–9·8 10·5–11·6
22 °C 5·2 ± 1·0 7·4 ± 0·7 9·9 ± 0·7 13·2 ± 1·8 16·6 ± 1·5 16·7 ± 2·0
95% CI 4·5–5·9 6·9–7·8 9·4–10·4 12·0–14·5 15·5–17·7 15·2–18·1
P value < 0·001
pH (37 °C)
4 °C 6·960 ± 0·017 7·030 ± 0·044 7·038 ± 0·046 7·016 ± 0·046 7·009 ± 0·067 6·881 ± 0·101
95% CI 6·948–6·972 6·999–7·061 7·005–7·071 6·983–7·048 6·962–7·056 6·809–6·952
22 °C 6·982 ± 0·082 7·155 ± 0·089 7·146 ± 0·061 7·100 ± 0·074 7·233 ± 0·125 7·566 ± 0·178
95% CI 6·924–7·040 7·092–7·217 7·103–7·189 7·047–7·152 7·144–7·321 7·442–7·694
P value < 0·001
Adenosine triphosphate (ATP) (umol per 1011)
4 °C 7·01 ± 0·60 5·37 ± 0·32 5·19 ± 0·25 5·01 ± 0·26 4·64 ± 0·38 3·50 ± 1·04
95% CI 6·59–7·44 5·15–5·60 5·01–5·37 4·83–5·20 4·37–4·91 2·76–4·23
22 °C 7·13 ± 0·57 6·68 ± 0·36 6·16 ± 0·40 4·70 ± 0·72 1·92 ± 0·63 0·48 ± 0·21
95% CI 6·73–7·53 6·43–6·94 5·88–6·44 4·19–5·20 1·48–2·37 0·33–0·63
P value < 0·001
pCO2 (37 °C)
4 °C 4·84 ± 0·34 3·18 ± 0·21 2·74 ± 0·10 2·36 ± 0·25 1·82 ± 0·10 1·30 ± 0·06
95% CI 4·59–5·08 3·03–3·32 2·67–2·81 2·18–2·53 1·75–1·89 1·26–1·34
22 °C 4·60 ± 0·81 2·97 ± 0·31 2·71 ± 0·09 2·14 ± 0·46 1·07 ± 0·17 0·0 ± 0·0
95% CI 4·03–5·17 2·76–3·19 2·65–2·77 1·81–2·46 0·95–1·18 0·0–0·0
P value < 0·001
pO2 (37 °C)
4 °C 19·8 ± 1·2 23·6 ± 0·9 23·9 ± 0·5 23·5 ± 1·6 24·9 ± 1·1 24·5 ± 0·9
95% CI 18·9–20·7 23·0–24·2 23·6–24·3 22·3–24·6 24·1–25·7 23·9–25·1
22 °C 19·9 ± 0·7 21·3 ± 1·3 20·9 ± 0·8 21·6 ± 0·8 23·5 ± 0·4 24·1 ± 0·3
95% CI 19·3–20·4 20·4–22·2 20·4–21·5 21·0–22·1 23·2–23·8 23·9–24·3
P value < 0·001
34 P. Sandgren et al.
© 2007 The Author(s)Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
of no value in predicting the quality of platelets stored at4 °C.
GP Ibα, an adhesion receptor on the surface of the platelet,binds to the vWF, which becomes attached to collagens andother adhesive proteins that are exposed when the vessel wallis damaged. Our findings accord with those of others whohave shown that storage of PCs reduces the expression ofGP1bα on the surface of the platelets [34–36]. This reductionis assumed to decrease the adhesive capacity. On the otherhand, another study suggests that a 65% reduction in thisexpression of GPIb, has no effect on platelet adhesion duringflow conditions [37]. The key question is whether thedecrease (MFI) in platelet surface expression after storage at4 °C is due to an irreversibly rearranged clustering of thevWFR (Gp1bα or CD42b), as shown in animal models [13].Our analyses of CD42b yielded interesting data. We found adecrease (MFI) in platelet surface expression, but no declinein the percentage of positive platelets after storage at 4 °C forup to 14 days. These data indicate that Gp1bα remains on thesurface of refrigerated platelets during long-term storageeven if the density of the antigen is reduced/rearranged. As
Gp1bα plays a major role in the clearance of transfusedrefrigerated platelets, further studies of the vWFR complexsubunits Gp1bα, GP1X and GPV, investigating the relationshipbetween vWFR clustering/conformational changes andlong-term storage at 4 °C, are needed.
Storage at 22 °C increased GPIIIa expression and thegeneration of platelet activation markers (CD62p and CD63)slightly more than did storage at 4 °C, with the exception ofday 21 (CD62p). The observed increase in the expression ofCD61 at 22 °C storage occurred at a storage time detrimentalto platelets and is therefore of no relevance.
We found that CD63 has a closer relation to modificationsin platelet morphology, as judged by a reduction in ESCvalues, an increase in the release of RANTES and a loss ofswirling, than CD62p. The expression of CD62p on the surfaceof platelets is usually used as a marker of platelet activationand may predict platelet viability [38], but cell-surface expressioncan be cleaved from the surface by various proteases andsome authors have proposed that soluble P-selectin may bea better marker of platelet activation [39]. However, ourresults imply that cell-surface expression of CD63 remains
Table 3 In vitro effects of PCs stored in PAS II for 21 days during various storage conditions: mean ± standard deviation (SD) (n = 8). 95% confidence interval
(CI); the P value represents the significance of the differences from baseline (day 1) between groups (heterogenicity) during 21 days
In vitro measure
Day
1 5 7 10 14 21
Bicarbonate (mmol/l)
4 °C 7·8 ± 0·5 6·0 ± 0·5 5·3 ± 0·7 4·3 ± 0·7 3·3 ± 0·6 1·8 ± 0·4
95% CI 7·4–8·1 5·7–6·3 4·8–5·8 3·9–4·8 2·9–3·7 1·5–2·1
22 °C 7·7 ± 0·5 7·5 ± 0·8 6·8 ± 0·9 4·9 ± 1·4 3·4 ± 1·1 0·0 ± 0·0
95% CI 7·3–8·0 6·9–8·1 6·1–7·4 3·9–5·9 2·6–4·1 0·0–0·0
P value < 0·001
Hypotonic shock response (HSR) (%)
4 °C 54·8 ± 2·7 29·4 ± 1·8 24·6 ± 1·8 20·3 ± 2·5 12·0 ± 2·5 8·6 ± 1·5
95% CI 52·9–56·7 28·1–30·7 23·4–25·9 18·5–22·1 10·3–13·8 7·5–9·6
22 °C 54·8 ± 2·7 38·0 ± 2·8 33·1 ± 3·8 24·9 ± 3·9 17·4 ± 2·5 3·4 ± 2·7
95% CI 52·9–56·7 36·0–39·9 30·4–35·9 22·1–27·7 15·6–19·1 1·5–5·3
P value < 0·001
Extent of shape change (ESC) (%)
4 °C 22·6 ± 0·7 0·0 ± 0·0 0·0 ± 0·0 0·0 ± 0·0 0·0 ± 0·0 0·0 ± 0·0
95% CI 22·1–23·1 0·0–0·0 0·0–0·0 0·0–0·0 0·0–0·0 0·0–0·0
22 °C 22·6 ± 0·7 12·3 ± 0·4 10·8 ± 0·7 7·2 ± 0·9 2·8 ± 0·8 0·0 ± 0·0
95% CI 22·1–23·1 12·0–12·5 10·3–11·3 6·5–7·8 2·2–3·4 0·0–0·0
P value < 0·001
Regulated upon activation of normal T-cells expressed and secreted (RANTES) (pg per 106 platelets)
4 °C 53·8 ± 20·8 44·0 ± 14·9 49·6 ± 19·0 96·4 ± 50·5 125·3 ± 70·9 137·1 ± 51·4
95% CI 39·0–68·5 33·5–54·6 36·1–63·0 60·7–132·1 75·2-175-5 100·8–173·4
22 °C 50·8 ± 23·8 329·5 ± 44·8 414·7 ± 32·3 459·8 ± 77·2 605·0 ± 58·0 1019·3 ± 580·3
95% CI 33·9–67·6 297·8–361·2 391·8–437·5 405·2–514·4 563·9–646·0 609·0–1429·6
P value < 0·001

© 2007 The Author(s) Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
Flow cytometry analysis of platelets stored at 4 °C 35
during the entire storage period. Therefore CD63 may bea better marker of platelet activation than CD62p.
After day 14, the increase in the expression of CD62p andCD63 in platelets stored at 4 °C was associated with anincrease in the release of RANTES, reflecting increased plateletactivation.
A higher expression of CD63 is suggested to reflect anincreased dense granula release, which is associated withplatelet activation. Platelets tend to be activated and tochange from a discoid to a spherocytic shape, which goestogether with a decreased response to the ESC assay, indicatinga loss of discoid shape. At 22 °C storage this condition isassociated with an increased release of RANTES from plateletα-granules as well as a higher expression of CD63. This isneither a bad nor a good relation; it merely reflects thebehaviour of platelets when activated. On the other hand,from a 22 °C storage point of view, it is a bad relation, whichis not desirable during storage. However, at 4 °C storageplatelets lose their discoid shape (lower ESC, higher MPV) butunder these storage conditions, loss of the discoid shape isnot associated with a higher expression of CD63 or anincreased release of RANTES from platelet α-granules. Fromday 1, our findings in cold-stored platelets are in line withthose in cold-treated mouse platelets , which suggest thatsuch treatment does not cause surface expression of theα-granule activation marker, P-selectin, or up-regulation ofGPIIb/IIIa [13].
In summary, cold-induced activation is not associatedwith an increased expression of human platelet membraneproteins and activation markers during long-term storage.Current reports suggest that if the reduction in CD42b expressionduring 4 °C storage reflects the irreversible rearrangementin clustering of the vWFR (Gp1bα or CD42b) as shown inanimal models, glycan modification [14] may prolong thecirculation of 4 °C platelets in humans. Therefore, theseeffects of a cold temperature on human platelet glycoproteinexpression during long-term storage may prove to be ofpractical value.
However, the biological impact of these findings needs tobe examined performing in vivo studies of recovery and survival,probably with addition of any protecting substance againstclearance, as well as increments in thrombocytopenic patientsto confirm the haemostatic effect of the platelets stored at4 °C. In addition, inclusion of supplementary components inthe platelet-storage medium, such as magnesium and potassiumions [40,41] are some of the tools that could be used to furtheroptimize the storage conditions at 4 °C.
Acknowledgements
The authors thank Elisabeth Ishizaki, Ingeborg Ohlsson andAnette Jonsson, Department of Clinical Immunology andTransfusion Medicine for valuable technical advice.
References1 Blajchman MA: Transfusion-associated bacterial sepsis: the
most common current transfusion-transmitted entity. TransfusToday 1994; 21:5–6
2 Bode AP: Platelet activation may explain the storage lesion inplatelet concentrates. Blood Cells 1990; 16:109–125
3 Chernoff A, Snyder EL: The cellular and molecular basis of theplatelet storage lesion: a symposium summary. Transfusion1992; 32:386–390
4 Seghatchian J, Krailadsiri P: The platelet storage lesion. TransfusMed Rev 1997; 11:130–144
5 Sandgren P, Shanwell A, Gulliksson H: Storage of buffy-coat-derived platelets in additive solutions: in vitro effects of storageat 4 °C. Transfusion 2006; 46:824–834
6 Oliver AE, Tablin F, Walker NJ, Crowe JH: The internal calciumconcentration of human platelets increases during chilling.Biochim Biophys Acta 1999; 1416:349–360
7 Hoffmeister K, Falet H, Toker A, Barkalow KL, Stossel TP,Hartwig JH: Mechanisms of cold-induced platelet actin assembly.J Biol Chem 2001; 276:24751–24759
8 Cardigan R, Turner C, Harrison P: Current methods of assessingplatelet function: relevance to transfusion medicine. Vox Sang2005; 88:153–163
9 de Bruijne-Admiral LG, Modderman PW, Von dem Borne AEG,Sonnenberg A: P-selectin mediates Ca++-dependent adhesion ofactivated platelets to many different types of leukocytes: detectionby flow cytometry. Blood 1992; 80:132–142
10 Metzelaar MJ, Korteweg J, Sixma JJ, Nieuwenhuis HK: Comparisonof platelet membrane markers for detection of platelet activationin vitro and during platelet storage and cardiopulmonary bypasssurgery. J Lab Clin Med 1993; 121:576–587
11 McGregor JL, Brochier J, Wild F, Follea G, Trzeciak MC, James E,Dechavanne M, McGregor L, Clementson KJ: Monoclonal antibodiesagainst platelet membrane glycoproteins. Characterization andeffect on platelet function. Eur J Biochem 1983; 131:427–436
12 Fullard JF: The role of the platelet glycoprotein IIb/IIIa in thrombosisand haemostasis. Curr Pharm Des 2004; 10:1567–1576
13 Shattil SJ: Function and regulation of the beta 3 integrinsin hemostasis and vascular biology. Thromb Haemost 1995;74:149–155
14 Hoffmeister KM, Felbinger TW, Falet H, Denis CV, Bergmeister W,Mayadas TN, von Andrian UH, Wagner DD, Stossel TP,Hartwig JH: The clearance mechanism of chilled blood platelets.Cell 2003; 112:87–97
15 Hoffmeister KM, Josefsson EC, Isaac NA, Clausen H, Hartwig JH,Stossel TP: Glycosylation restores survival of chilled bloodplatelets. Science 2003; 301:1531–1534
16 Tang RC, Huang CY, Pei XP, Chen BA, Li CP, Shi GY: Evaluationof the effects of glycosylation on in vivo survival of cold – storagehuman platelets by using rabbit model. Zhong Guo Shi Yan XueYe Xue Zhi 2005; 13:1116. (in Chinese)
17 Murphy S, Gardner FH: Effect of storage temperature onmaintenance of platelet viability – deleterious effect of refrigeratedstorage. N Engl J Med 1969; 280:1094–1098
18 Becker GA, Tucelli M, Kunicki T, Chalos MK, Aster RH: Storageof platelet concentrates stored at 22 °C and 4 °C. Transfusion1973; 13:61–68

36 P. Sandgren et al.
© 2007 The Author(s)Journal compilation © 2007 Blackwell Publishing Ltd., Vox Sanguinis (2007) 93, 27–36
19 Högman CF: Aspects of platelets storage. Transfus Sci 1994;15:351–355
20 Murphy S: The efficacy of synthetic media in the storage ofhuman platelets for transfusion. Transfus Med Rev 1999;13:153–163
21 Gulliksson H: Additive solutions for the storage of platelets fortransfusion. Transfus Med 2000; 10:257–264
22 Holme S: Platelet storage in a liquid environment. Transfus Sci1994; 15:117–130
23 Larsson S, Sandgren P, Sjödin A, Vesterinen M, Gulliksson H:Automated preparation of platelet concentrates from pooledbuffy coats: in vitro studies and experiences with the OrbiSacsystem. Transfusion 2005; 45:743–751
24 Bertolini F, Murphy S: A multicenter evaluation of reproducibilityof swirling in platelet concentrates. Transfusion 1994; 34:802–810
25 Moroff G, Eich J, Dabay M: Validation of use of the Nageottehemocytometer to count low levels of white cells in whitecell-reduced platelet components. Transfusion 1994; 34:35–38
26 VandenBroeke T, Dumont LJ, Hunter S, Nixon J, Murphy S,Roger J, Herschel L, AuBuchon JP, Gulliksson H, Dengler T,Hornsey V, Prowse C; Biomedical Excellence for Safer TransfusionWorking Party of the International Society of Blood Transfusion:Platelet storage solution effects on the accuracy of laboratorytests for platelet function: a multilaboratory study. Vox Sang2004; 86:183–188
27 Lundin A: Use of firefly luciferase in ATP-related assays ofbiomass, enzymes and metabolites. Meth Enzymol 2000;305:346–370
28 King J: A routine method for the estimation of lactic dehydrogenaseactivity. J Med Lab Tech 1959; 16:265–272
29 Snyder EL: Activation during preparation and storage of plateletconcentrates. Transfusion 1992; 32:500–502
30 Rinder HM, Snyder EL, Bonan JL, Napychank PA, Malkus H,Smith BR: Activation in stored platelet concentrates: correlationbetween membrane expression of P-selectin. GlycoproteinIIb/IIIa, and β-thromboglobulin release. Transfusion 1993;33:25–29
31 Gulliksson H: Defining the optimal storage conditions for thelong-term storage of platelets. Transfus Med Rev 2003; 17:209–215
32 Holme S, Moroff G, Murphy S: A multi-laboratory evaluation ofin vitro platelet assays: the tests for extent of shape change andresponse to hypotonic shock. Transfusion 1998; 38:31–40
33 Mintz PD, Anderson G, Avery N, Clarl P, Bonner RF: Assessmentof the correlation of platelet morphology with in vivo recoveryand survival. Transfusion 2005; 45:72–80
34 Bolin RB, Medina F, Cheney BA: Glycoprotein changes in freshvs room temperature-stirred platelets and their buoyant densitycohorts. J Lab Clin Med 1981; 98:500–510
35 Lozano ML, Rivera J, Gonzales-Conejero R, Moraleda JM, Vicente V:Loss of high-affinity thrombin receptors during platelet concentratestorage impairs the reactivity of platelets to thrombin. Transfusion1997; 37:368–375
36 Turner CP, Sutherland J, Wadhwa M, Dilger P, Cardigan R:In vitro function of platelet concentrates prepared after filtrationof whole blood or buffy coat pools. Vox Sang 2005; 88:164–171
37 Henrita van Zanten G, Saelman EU, Schut-Hese KM, Wu YP,Slootweg PJ, Nieuwenhius HK, de Groot PG, Sixma JJ: Plateletadhesion to collagen type IV under flow conditions. Blood 1996;88:3862–3871
38 Holme S, Sweeney JD, Sawyer S, Elfath MD: The expression ofp-selectin during collection, processing and storage of plateletconcentrates: relationship to loss of in vivo viability. Transfusion1997; 37:12–17
39 Kostelijk EH, Fijnheer R, Nieuwenhuis HK, Gouwerok CW,de Korte D: Soluble P-selectin as parameter for platelet activationduring storage. Thromb Haemost 1996; 76:1086–1089
40 Gulliksson H, AuBuchon JP, Vesterinen M, Sandgren P, Larsson S,Pickard CA, Herschel I, Roger J, Tracy JE, Langweiler M: Storageof platelets in Additive solutions: a pilot study of the effects ofpotassium and magnesium. Vox Sang 2002; 82:131–136
41 Shanwell A, Falker C, Gulliksson H: Storage of platelets in additivesolutions: the effects of magnesium and potassium on therelease of RANTES, beta-thromboglobulin, platelet factor 4 andinterleukin-7 during storage. Vox Sang 2003; 85:206–212





























