Embed Size (px)
Citation preview

Structure and function of phage p2 ORF34p2, a new type ofsingle-stranded DNA binding proteinmmi_6844 1156..1170
Erika Scaltriti,1,2 Mariella Tegoni,1 Claudio Rivetti,3
Hélène Launay,4 Jean-Yves Masson,4
Alfonso H. Magadan,5 Denise Tremblay,5
Sylvain Moineau,5 Roberto Ramoni,2 Julie Lichière,1
Valérie Campanacci,1 Christian Cambillau1* andMiguel Ortiz-Lombardía1**1Architecture et Fonction des MacromoléculesBiologiques, UMR 6098 CNRS and Universitésd’Aix-Marseille I and II, Campus de Luminy, case 932,13288 Marseille cedex 09, France.2Istituto di Biochimica Veterinaria, Facoltà di MedicinaVeterinaria, Università di Parma, Via del Taglio 8, 43100Parma, Italy.3Istituto di Scienze Biochimiche, Università di Parma,43100, Parma, Italy.4Genome Stability Laboratory Laval University CancerResearch Center, Hôtel-Dieu de Québec, 9 McMahon,Québec City, Québec, Canada G1R 2J6.5Département de Biochimie et de Microbiologie, Facultédes Sciences et de Génie, Groupe de Recherche enÉcologie Buccale, Faculté de Médecine Dentaire, Félixd’Hérelle Reference Center for Bacterial Viruses,Université Laval, Québec, Canada G1V 0A6.
Summary
Lactococcus lactis, a Gram-positive bacteriumwidely used by the dairy industry, is subject to infec-tion by a diverse population of virulent phages, pre-dominantly by those of the 936 group, including thesiphovirus phage p2. Confronted with the negativeimpact of phage infection on milk fermentation, thestudy of the biology of lactococcal provides insightfrom applied and fundamental perspectives. Wedecided to characterize the product of the orf34gene from lactococcus phage p2, which was consid-ered as a candidate single-stranded DNA bindingprotein (SSB) due to its localization downstream of agene coding for a single-strand annealing protein.Two-dimensional gel electrophoresis showed that
ORF34p2 is expressed in large amounts during theearly phases of phage infection, suggesting animportant role in this process. Gel-shift assays,surface plasmon resonance and atomic forcemicroscopy demonstrated that ORF34p2 interactswith single-strand DNA with nanomolar affinity. Wealso determined the crystal structure of ORF34p2 andshowed that it bears a variation of the typicaloligonucleotide/oligosaccharide binding-fold ofSSBs. Finally, we found that ORF34p2 is able tostimulate Escherichia coli RecA-mediated homolo-gous recombination. The specific structural and bio-chemical properties that distinguish ORF34p2 fromother SSB proteins are discussed.
Introduction
Single-stranded DNA binding (SSB) proteins are presentin all organisms and play an essential role in theirgrowth and metabolic activities. They are involved inmultiple pathways of DNA metabolism, including replica-tion, recombination and repair in Bacteria, Archaea,Eukarya and viruses (Lohman and Ferrari, 1994; Wold,1997). In particular, they coat and protect single-stranded DNA (ssDNA) intermediates and facilitatepairing of homologous strands by pre-empting DNA sec-ondary structures.
The SSB proteins can bind ssDNA in a non-specific andcooperative manner and form filaments that can saturatelong stretches of ssDNA (Lohman and Ferrari, 1994).Besides, SSBs can interact with a large number of heter-ologous proteins and these interactions both regulate andcan be regulated by the interaction with ssDNA (Wold,1997). Under particular conditions, SSBs can stimulatethe assembly of recombinases on ssDNA. Moreover, thefilaments formed from SSB–recombinase interactions arein general more efficient in strand-exchange reactionsthan filaments formed by recombinase alone (Sung,1997). Indeed, the presence of EcoSSB permits the for-mation of a continuous filament of RecA protein by desta-bilizing the secondary structure of ssDNA (Meyer andLaine, 1990). In Saccharomyces cerevisiae, the SSBprotein RPA (Replication Protein A) stimulates the forma-tion of nucleoprotein filaments including the recombinaseRad51 and, in combination with Rad52, stimulates
Accepted 31 July, 2009. For correspondence. *E-mail [email protected]; Tel. (+34) 491 82 55 90; Fax (+34) 491 26 67 20 or**E-mail [email protected]; Tel. (+34) 491 82 55 93; Fax(+34) 491 26 67 20.
Molecular Microbiology (2009) 73(6), 1156–1170 � doi:10.1111/j.1365-2958.2009.06844.xFirst published online 28 August 2009
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd

Rad51-mediated DNA strand exchange (Sung, 1997;Shinohara et al., 1998).
From a structural viewpoint SSBs exist as monomericor multimeric proteins and, with few exceptions,they share a structural domain named OB-fold(oligonucleotide/oligosaccharide binding-fold) involved innucleic acid recognition. The OB-fold domains are vari-able in length and functions, mainly because of differ-ences in the length of the loops forming the nucleic acidinteraction surfaces. EcoSSB is the paradigm of SSBproteins within the OB-fold structural family (Theobaldet al., 2003). The structure of EcoSSB consists of onea-helix and two three-stranded anti-parallel b-sheets thatform a semi-open barrel from where two long loops pro-trude, one of them involved in ssDNA interaction (Murzin,1993). The crystal structures of EcoSSB with (Raghu-nathan et al., 2000) and without ssDNA (Raghunathanet al., 1997; Matsumoto et al., 2000) confirmed that thisprotein forms homotetramers. Each monomer of theEcoSSB homotetramer consists of an N-terminal DNAbinding domain, the OB-fold domain, followed by aC-terminal region, including a glycine-rich loop and anamphipathic tail. The C-terminal region is proposed to beinvolved in the interaction with enzymes of the DNAmetabolism such as polymerases and recombinases(Genschel et al., 2000; Reddy et al., 2001; Jackson et al.,2002; He et al., 2003; Sun and Shamoo, 2003).
In bacteriophages, SSBs proteins play critical roles inDNA replication and recombination. Both mechanisms areoften closely coupled throughout much of the phage lyticcycle, particularly during recombination-dependent repli-cation processes (Bleuit et al., 2001). In some coliphages,like T4 and T7, phage-encoded SSBs are essential forDNA replication and cannot be replaced by their hostcounterparts with which they share no homology (Kim andRichardson, 1993; Miller et al., 2003). In contrast, coliph-age P1 (Hay and Cohen, 1983) and Bacillus phage SPP1(Ayora et al., 1998) encode proteins highly homologous tothe bacterial-host SSB. For these reasons, some phageSSBs have been identified by their sequence similarity tobacterial SSBs whereas others were recognized on thebasis of their activity. The crystal structures of phageSSBs, such as the monomeric T4 gp32 (Shamoo et al.,1995), the dimeric T7 gp2.5 (Hollis et al., 2001) and GN5from Escherichia coli filamentous phage f1 (Skinner et al.,1994; Su et al., 1997), confirmed their OB-fold structuraland functional motif (Murzin, 1993).
Virulent phages of Lactococcus lactis have a negativeeffect on milk fermentations, which explains the continu-ing interest on the biology of these phages, including theirstructure (Ricagno et al., 2006; Spinelli et al., 2006;Sciara et al., 2008), phage DNA replication and homolo-gous recombination processes, as well as their interactionwith their hosts (Moineau, 1999; Bouchard and Moineau,
2004; Ploquin et al., 2008). However, there are no struc-tures of lactococcal SSBs available yet.
In this work, we present the crystal structure of theORF34p2 protein, encoded by the virulent L. lactis phagep2 (936 group). The orf34 gene (GenBank: GQ253898) islocated downstream a putative single-strand annealingprotein named Sak3 (Bouchard and Moineau, 2004).ORF34p2 probably belongs to a set of genes involvedin phage-promoted DNA recombination, as postulatedfor the SSB protein of the lactococcal phage bIL67ORF14bIL67 (Szczepanska et al., 2007). Indeed, ORF34p2
bears 33% sequence identity with ORF14bIL67 (Fig. 1). Ofnote, virulent phages bIL67 and p2 belong to geneticallydistinct groups of lactococcal siphoviridae phages,namely c2 and 936 respectively (Deveau et al., 2006).Despite the lack of homology with SSB proteins present indatabases, ORF34p2 shares some characteristics gener-ally observed in this protein family, notably the frequentlyfound C-terminal phenylalanine (He et al., 2003), as wellas features conserved among SSB proteins from lacto-coccal phages, such as the consensus sequence DLFGGnear the C-terminus and three terminal residues LPF(Szczepanska et al., 2007). The presence of these fea-tures prompted us to explore the hypothesis that ORF34p2
is an SSB protein by studying its biochemical and struc-tural properties.
Results and discussion
ORF34p2 is expressed early and abundantly duringphage p2 infection
To investigate the expression course of ORF34p2 duringinfection, we infected the host strain L. lactis ssp. cre-moris MG1363 with the virulent phage p2. Infection wasallowed to proceed for 20 min, the minimum time reportedbefore new p2 virions are released (Haaber et al., 2008).Intracellular proteins were extracted and run on 2D polyacrylamide gels. Proteomic profiles were comparedbetween infected and non-infected cells in a time-coursemanner (Fig. 2). Based on the theoretical molecularweight (13 kDa) and pI (4.92) of ORF34p2, a spot corre-sponding to an early-expressed phage protein with appar-ent mass 15.4 kDa and pI 5.1 (Fig. 2) was picked out ofthe gels and analysed by liquid chromatography-tandemmass spectrometry. The mass spectrometry matchedORF34p2 protein with 97% of coverage and more than95% of confidence. The analysis of the time-courseexperiment revealed that ORF34p2 was detected as earlyas 5 min post infection and that its concentrationincreased significantly over time (Fig. 2). In fact, ORF34p2
was one of the highest expressed phage p2 proteins (datanot shown). Early and abundant expression likely corre-late with important roles during the phage infection
Structure & function of lactococcus phage p2 ORF34 1157
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

process. It should be noted that SSBs in general areessential proteins, which cannot be deleted or replaced.
Recombinant ORF34p2 forms dimers and tetramersin solution
An N-terminally His-tagged ORF34p2 protein wasexpressed in E. coli and purified as a soluble protein thatcould be concentrated to 10 mg ml-1. RecombinantORF34p2 was prone to time-dependent proteolysis, asSDS-PAGE gels showed signs of cleavage after keepingthe protein for 5 days at 4°C (data not shown). SEC-MALS/UV/RI (Wyatt Technology) experiments suggestthat ORF34p2 is a dimer (not shown), like coliphage T7 gp2.5 (Hollis et al., 2001) or phage Pf3 SSB (Folmer et al.,
1994). However, size-exclusion chromatography showedthat the protein would also form tetramers (not shown),like EcoSSB (Raghunathan et al., 1997; 2000) and thelactococcal phage bIL67 SSB protein ORF14bIL67 (Szc-zepanska et al., 2007). The (SeMet) M2B-ORF34p2
mutant prepared for structural studies (see below and inExperimental procedures) showed characteristics similarto the native protein, but with a faster time-dependentdegradation.
ORF34p2 binds ssDNA in vitro
Electrophoretic mobility shift assays (EMSA) were used toinvestigate the ssDNA- and double-stranded DNA(dsDNA) binding properties of ORF34p2 (Fig. 3). Quantifi-
Fig. 1. Sequence alignment of lactococcal phage closest relatives of ORF34p2. Except for ORF34p2, sequences are labelled with the name ofthe corresponding Lactococcus phage. Residue backgrounds are greyed out by similarity starting from a low-similarity cut-off of 70%.Numbers and secondary-structure elements above the sequences correspond to ORF34p2. b-Strands are numbered according to the text.Triangles and stars under the sequences indicate the conserved DLFGG and LPF motifs respectively.
Fig. 2. Expression of ORF34p2 during thephage p2 infection of L. lactis ssp. cremorisMG1363.A. Two-dimensional gel electrophoresis gel ofintracellular proteins of L. lactis MG1363extracted from a sample taken 10 min afterphage p2 infection. The box represents thezoom area of (B).B. Zoom areas from two-dimensional gelelectrophoresis gels obtained with intracellularproteins extracted from non-infected cells (NI)and after 5, 10, 15 and 20 min following thebeginning of the phage infection. Arrowsindicate the position of the SSB protein.
1158 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

cation of the DNA–protein complexes at 1 mM ORF34p2
concentration indicated that 40% of the ssDNA substratewas stably bound compared with 1% of the dsDNA(Fig. 3A, lanes 5 and 11). ORF34p2 binding to ssDNA wasalso monitored by EMSA using circular M13 ssDNA. Asshown in Fig. 3B, titration of M13 ssDNA with increasingamounts of protein resulted in a band shift that reaches aplateau at a molar ratio of 400 ORF34p2 : M13 ssDNA(Fig. 3B, lane 6). Given the length of the M13 ssDNA [7249nucleotides (nt)] and assuming a tetrameric organization ofORF34p2, this result implies a site size of ~72 nt, similar tothe 65 nt wrapped around the EcoSSB homotetramer inone of its binding modes (Lohman and Overman, 1985).
These results, based on ssDNAs with differentsequences, also prove that, consistently with its pre-sumed SSB function, ORF34p2 non-specifically bindsssDNA and not to dsDNA. This property is necessary tocoat and protect ssDNA intermediates during DNA repli-cation, repair and recombination (Gasior et al., 2001).
To confirm the qualitative results of EMSA and tomeasure a dissociation constant for the ORF34p2–ssDNAcomplexes, we performed surface plasmon resonanceexperiments. This method has already been used toevaluate SSB–ssDNA interactions in real time (Wanget al., 2000). We coated a streptavidin sensor chip withssDNA and recombinant ORF34p2 was injected over thechip surface at increasing concentrations, between 23and 375 nM (Fig. S1). The mean dissociation constant(KD) value, based on three independent experiments, was2.5 � 0.6 ¥ 10-8 M with an association (kon) and adissociation (koff) rates of 1 � 0.2 ¥ 105 M-1 s-1 and2.5 � 0.1 ¥ 10-3 s-1 respectively. These results confirmsthat ORF34p2 has a high affinity for ssDNA, although lowerthan other SSB proteins like EcoSSB (KD = 2.5 ¥ 10-9 Mwith faster kon and koff, 1.4 ¥ 105 M-1 s-1 and 3.4 ¥ 10-4 s-1
respectively) and Mycobacterium tuberculosis SSB(KD = 3.6 ¥ 10-9 M with a faster kon and koff, 6.3 ¥104 M-1 s-1 and 2.3 ¥ 10-4 s-1 respectively) (Reddy et al.,2001).
The crystal structure of ORF34p2 shows overall similaritywith bacterial SSBs but with significant differences
We determined the crystal structure of ORF34p2 to gainfurther insight into its possible role as an SSB protein. Ifthat hypothesis could be confirmed, the structure ofORF34p2 would have the bonus of giving the first informa-tion at atomic detail of a lactococcal phage SSB protein, afamily that shares no significant homology with any SSBsof known structure.
The recombinant (native) ORF34p2 protein was crys-tallized with space group symmetry P212121 (Table 1). Aslow sequence identity with possible models precludedthe possibility of solving the structure of the protein bythe molecular replacement method and due to theabsence of methionine residues in its sequence, wemutated leucines at positions 49 and 55 into methion-ines, thus generating the L49M-L55M double mutantM2B-ORF34p2. Seleno-methionyl (SeMet) M2B-ORF34p2
crystallizes under conditions and with different spacegroup symmetry (P6322, Table 1) from the native protein.The structure of (SeMet) M2B-ORF34p2 was solved bythe single-wavelength anomalous dispersion method.The structure of native ORF34p2 was subsequentlysolved by molecular replacement using the structure of(SeMet) M2B-ORF34p2 as search model. There is oneORF34p2 molecule per asymmetric unit in the (SeMet)M2B-ORF34p2 hexagonal crystals and four in the orthor-hombic crystals of native ORF34p2. Thus, we had at ourdisposal five independent monomers of ORF34p2 forstructural analysis.
As expected for an SSB protein, the N-terminal DNAbinding domain of ORF34p2 has the overall structure ofan OB-fold (Fig. 4A). Two secondary-structure elementsin the structure of ORF34p2 differ significantly from thestructures of other SSB proteins. Namely, the 1′ strandis unusually long and, remarkably, the capping a-helix isabsent. Instead, strands 3 and 4 are connected by aneight residues-long loop (P50–D57). These features are
Fig. 3. Electrophoretic mobility shift assays.A. DNA retardation gel assay of the indicatedconcentrations of ORF34p2 on single- (lanes2–5) and double-strand DNA (lanes 8–11).Lanes 1 and 7, controls without protein; lanes6 and 12, binding of EcoSSB (0.5 mM) onsingle- and double-strand DNA.B. Agarose gel electrophoresis of M13ssDNA-ORF34p2 complexes at increasingDNA : ORF34p2 monomer molar ratios (R, atthe bottom). Lane 1 corresponds toprotein-free ssDNA.
Structure & function of lactococcus phage p2 ORF34 1159
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

to some extent present in the structures of SSB proteinsfrom filamentous phages f1 (Su et al., 1997) and Pf3(Folmer et al., 1995). Notwithstanding, ORF34p2 stronglydiffers from the f1SSB and Pf3SSB proteins with respectto the orientation of the 4′-5′ b-hairpin, which in ORF34p2
closely matches that of EcoSSB. Such difference impliesthat ORF34p2 builds dimers analogous to those found forEcoSSB and other bacterial SSB proteins, which aretopologically different from the dimers formed by phagef1SSB and Pf3SSB (see below).
In comparing the structure of the ORF34p2 monomerwith that of EcoSSB, it is also noteworthy that the ori-entations of the 1′, 2 and 3 strands in the former deviatefrom their orientation in EcoSSB. In ORF34p2 the 1′-2b-hairpin is farther and the L23 loop closer to the 4′-5′b-hairpin of the same monomer respectively (Fig. 4A).As a result, the structure of the ORF34p2 monomer ismore compact than that of the monomer of EcoSSB.Furthermore, these three structural motifs are also thoseexhibiting a higher variability within the five independentmonomers presented in this study. Globally, the fiveindependent ORF34p2 monomers can be superposedwith a main-chain root mean square deviation (r.m.s.d.)of 0.8 Å for the alpha carbons of the 88 residuesaligned. In comparison, superposition of the fourindependent monomers of the EcoSSB structure(PDB : 1QVC) onto monomer A from the native ORF34p2
structure, gives a main-chain r.m.s.d. of 3.9 Å for thealpha carbons of the 74 residues aligned. In both casesa 2.0 Å lower limit for structural similarity was enforced.
If the most flexible motifs are excluded and the rigidparts of the five ORF34p2 monomers are aligned sepa-rately, the main-chain alpha carbon r.m.s.d. falls to0.55 Å for 81 residues aligned, as determined by theRAPIDO server (Mosca and Schneider, 2008). When thesame approach is used with the EcoSSB monomers,their r.m.s.d. with respect to ORF34p2 falls to 1.3 Å for 51residues aligned.
Well-defined electronic density is lacking for theC-terminal region beyond residues 88–93 in all subunitsof both the (SeMet) M2B-ORF34p2 and the nativeORF34p2 structures. This result is in line with previousdata on EcoSSB (Savvides et al., 2004), M. tuberculosisSSB (Saikrishnan et al., 2003) and Thermotoga maritimaSSB (DiDonato et al., 2006). Savvides and co-workershave proposed that the C-terminus of EcoSSB mayprotrude away from the DNA binding domain asa disordered/unstructured region (Savvides et al., 2004).Indeed, the low sequence complexity of the C-terminaldomain of SSB proteins, heavily populated by proline,glycine and glutamine residues, had hinted at the pos-sibility of the intrinsic disorder of this region (Sancaret al., 1981). Electron density was also too weak tosupport a model in parts of the L23 loop for one of themonomers in the native structure (residues K36-G37,monomer D) and for the (SeMet) M2B-ORF34p2
monomer (residues K36-N39). Similarly, residues S70-E72 (monomer D, native structure) and A69-V74[(SeMet) M2B-ORF34p2 monomer] belonging to the 4′-5′b-hairpin could not be modelled.
Table 1. Data collection and refinement statistics.
Native ORF34p2 (SeMet) M2B-ORF34p2
Data collectionSpace group P212121 P6322Unit cell (Å, °) a = 48.28, b = 71.71, c = 109.73
a = b = g = 90a = 75.74, b = 75.74, c = 88.04a = b = 90, g = 120
Resolutiona (Å) 20.0–2.60 (2.74–2.60) 88.04–2.10 (2.21–2.10)Wavelengtha (Å) 0.8726 0.9800Completenessa (%) 98.8 (99.8) 99.8 (100.0)Multiplicitya 3.5 (3.5) 10.2 (10.5)Unique reflectionsa 12049 (1755) 9205 (1291)<I/s(I)> 11.7 (3.1) 25.8 (4.8)Rmeasa,b (%) 12.9 (47.7) 7.4 (49.2)Wilson B factor (Å2) 36.0 40.0
RefinementRfactor/Rfreec (%, #) 19.2/25.1 (974) 23.9/25.6 (918)r.m.s.d.bonds/anglesd (Å, °)
0.008 (0.022)/1.032 (1.951) 0.011 (0.022)/1.186 (1.949)
Average B (Å2) 44.2 55.7
a. Values in parenthesis are for the highest-resolution shell.b. The value of the multiplicity-weighted merging R factor between equivalent measurements of the same reflection, R = ∑hklv{nhkl/(nhkl - 1}∑j
|Ihkl,j - ⟨Ihkl⟩|/∑hkl∑j Ihkl,j.c. Crystallographic Rfactor, Rfree = ∑ ||Fo| - k|Fc||/∑ |Fo|. The size of the test set used for the calculation of Rfree is in parentheses.d. Root mean square deviations from the standard values are given with target values in parentheses.
1160 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

Fig. 4. Crystal structure ORF34p2.A. Stereo view of the ORF34p2 monomer. A ribbon representation of the main-chain is coloured from blue (N-terminus) to red (C-terminus).b-Strands are numbered. Side-chain atoms of positively charged and aromatic residues present in the possible ssDNA binding site arerepresented as sticks. Those among them that are conserved in lactococcal SSB genes are labelled.B. Structural comparison of the ORF34p2 dimer and the archetypical EcoSSB dimer. Dimers are shown in the same orientation after beingsuperposed. The 4′-5′ b-hairpin and L23 loop of one of the monomers are represented in each case for reference.C. Pseudo-D2 symmetry of the ORF34p2 tetramer. The dyad axes are represented as green bars capped with black spheres. Monomers arelabelled a–d.D. Electrostatic potential of the ORF34p2 tetramer mapped onto its solvent-accessible surface from negative (-52 mV, red) to positive (52 mV,blue). The tetramer is represented in the same orientation as in (C). This figure was prepared with the program CHIMERA. Electrostaticcalculations were performed with APBS using the PARSE force-field in water with an implicit solvent model including 150 mM NaCl.
Structure & function of lactococcus phage p2 ORF34 1161
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

Structure of the ORF34p2 dimer
The SSB proteins can be found in a variety of oligomericforms: monomeric [e.g. coliphage T4 gp32 (Shamooet al., 1995)], dimeric [e.g. coliphage T7 gp2.5 (Holliset al., 2001)], heterotrimeric [e.g. human RPA(Iftode et al., 1999)] and homotetrameric, as in bacterialSSB proteins such as EcoSSB. ORF34p2 protein formsdimers bearing the same structural disposition in both itsnative and the (SeMet) M2B-ORF34p2 crystal forms. Theasymmetric unit of the native ORF34p2 crystal includestwo dimers while in the (SeMet) M2B-ORF34p2 crystal onedimer is formed by the interaction between two symmetry-related monomers (Fig. 4B). The fact that these crystallineforms belong to two different crystallographic spacegroups strongly supports the notion that the observeddimer has a physiological entity.
In the ORF34p2 dimer, the extended, anti-parallelb-sheet formed by the one, four and five strands fromeach monomer observed in EcoSSB is also present andcontributes eight hydrogen bonds to the dimer interface.Therefore, the ORF34p2 dimer is globally similar to that ofEcoSSB and different from those formed by other phageSSB proteins such as f1SSB (Su et al., 1997), Pf3SSB(Folmer et al., 1995) or gp2.5 from coliphage T7 (Holliset al., 2001). However, beyond the overall similarity, thedifferences observed between the ORF34p2 and theEcoSSB monomers translate into different arrangementsof their respective dimers. The most salient differencebetween the ORF34p2 dimer and the dimers of bacterialSSB proteins is the close interaction between the L23 loopof each monomer and the 4′-5′ b-hairpin of the othermonomer (Fig. 4B). This unique interaction implies thatthe central prominence that in other SSB proteins is madeby the 4′-5′ b-hairpins opens up, creating a continuousgroove along the dimer longest dimension (Fig. 4B andD). This shallow groove communicates the two transver-sal channels where ssDNA binds in EcoSSB (Raghu-nathan et al., 2000). Although the interface between L23and the 4′-5′ b-hairpin is mainly hydrophobic in nature,this interaction contributes, on average, eight furtherhydrogen bonds to the dimer, including possible ionicinteractions between residue pairs E33-R64 and E35-K68. The actual bonding scheme varies in each casereflecting the flexibility of the structural motifs involved inthe interaction, and consistently with the fact that theseregions were not completely modelled in some of themonomers (see above). This variability is also observedwhen we compare the interface surfaces of the dimers. Inthe three independent dimers found [two in the nativestructure and one in that of (SeMet) M2B-ORF34p2] eachmonomer buries a surface ranging from 945 Å2 in the(SeMet) M2B-ORF34p2 structure to 1085 Å2 in one of thedimers in the native structure, as reported by the PISA
server at the EBI (http://www.ebi.ac.uk/msd-srv/prot_int/pistart.html) (Krissinel and Henrick, 2004).
Architecture of the ORF34p2 tetramer
The ambiguity observed in solution with respect to theoligomeric state of ORF34p2 is echoed by its crystal forms.Whereas the native structure unequivocally displays ahead-to-head tetrameric layout, the (SeMet) M2B-ORF34p2 structure clearly appears as a dimer.
Similarly to other SSB proteins, the ORF34p2 tetramerexhibits pseudo-D2 symmetry (Fig. 4C). However, theprincipal axes of the two dimers form an angle of ~85°,substantially bigger than the angles found in EcoSSB(~70°) or MtbSSB (~35°) tetramers. This almost orthogo-nal disposition of the dimers allows them to pack moreclosely than in other homotetrameric SSB proteins,making the ORF34p2 tetramer the most compact homotet-rameric SSB structure described until now.
The ORF34p2 tetramer is held together mainly by hydro-phobic interactions ensuing from the further 3425 Å2
buried upon its formation. The tetramerization interface,which involves strands 1, 4 and 5 of each monomer,includes only six hydrogen bonds, mostly involving resi-dues at both ends of the well-defined structure, namelyT1, T6, Q8, E84 and T89. Contrary to EcoSSB andHmtSSB, there are no ionic interactions to strengthen thetetramer. Therefore, we conclude that under the condi-tions tested, ORF34p2 can form strong dimers and weaktetramers.
Furthermore, a higher-order organization is suggestedby the native structure of ORF34p2, where a crystallo-graphic interaction is found between monomers A and Cof adjacent tetramers. Each monomer buries ~980 Å2 inthis interface, which includes more than 10 hydrogenbonds and a possible salt bridge between E57 inmonomer A and R15 in monomer C. This inter-tetramericinterface occludes the putative ssDNA binding groove ofmonomer C but leaves monomer A free to bind to ssDNA.If, as known for EcoSSB (Lohman and Overman, 1985),ORF34p2 supports different ssDNA binding modes, thistetramer–tetramer interaction could be the basis for one ofthose putative modes. However, this hypothesis must betempered by the fact that the (SeMet) M2B-ORF34p2
structure does not show this type of interaction.
The structure of ORF34p2 suggests putative ssDNAbinding sites
The OB-fold proteins tend to use a common ligandbinding interface centred on b-strands 2 and 3, with con-tributions from the L12, L3a and L45 loops (Murzin, 1993). Inthe case of EcoSSB, this interface is further extended sothat the ssDNA binds along the groove defined in each
1162 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

monomer by the 1′-2 and the 4′-5′ b-hairpins, with furtherinteractions involving the L23 loop and the a-helix. Theparticular architecture of the ORF34p2 tetramer makesthat the clefts expected to direct ssDNA binding form analmost continuous trench that has a positive electrostaticpotential (Fig. 4D). Lining this trench, several positivelycharged and aromatic residues are interspaced that mightplay a role in ssDNA binding: K12, R15, K21, K24, K34,K36, R64, K68, K85 and Y44, F48, F52, Y73, F80 and F87respectively. Residues marked in bold italics are con-served, or replaced by an amino-acid of the same type,among the homologues of ORF34p2 in other lactococcalphages (Fig. 1) and thus constitute potential candidates tobe involved in ssDNA binding. With a complementaryapproach, site-directed mutagenesis and subsequentelectrophoretic mobility-shift assays performed by Szc-zepanska and co-workers on Orf14bIL67 have shown thatresidues V17, K65 and Y70, especially the latter two, areinvolved in ssDNA binding (Szczepanska et al., 2007). Ofthese residues, only V17 is conserved in ORF34p2.However, the other two residues would indeed localize inthe ssDNA binding groove. Further experiments will berequired to determine the actual residues involved indirect interaction with ssDNA.
In summary, whereas the crystal structure of ORF34p2
clearly corresponds to that of an SSB protein, this phageprotein displays unique traits, notably the compactnessand reorganization of the L23 loop and the 4′-5′ b-hairpin inthe monomer and the contacts established between thesestructural elements in the dimer. These unique featuressupport the notion that lactococcal SSB proteins consti-tute a new family within the SSB protein superfamily, ashas been proposed elsewhere (Szczepanska et al.,2007).
Imaging of SSB–ssDNA complexes by atomic forcemicroscopy
Atomic force microscopy (AFM) has been previouslyexploited to confirm the capacity of SSB proteins to bindssDNA, e.g. for coliphage T7 gp2.5 (He et al., 2003) andfor EcoSSB (Hamon et al., 2007). Compared with dsDNA,ssDNA is considerably more difficult to image by AFMbecause it is thinner and forms secondary structures thatfold the molecules into complex shapes, or even intoblobs (Fig. 5A). To curtail this inconvenience we replacedmagnesium with the trivalent cation spermidine in thedeposition buffer (Hamon et al., 2007). Spermidine wasalso efficient in spreading the SSB–ssDNA complexes onthe surface at all the protein concentrations used in theexperiments presented herein.
We titrated the circular coliphage M13 genomic ssDNAwith ORF34p2 (Fig. 5). Under our experimental conditionsand in the absence of ORF34p2, M13 ssDNA displayed
several regions with secondary structure organization,spaced by extended stretches (Fig. 5B). We did notobserve fully extended M13 ssDNA molecules.
Incubation of M13 ssDNA with increasing concentra-tions of ORF34p2 (Fig. 5C–F) resulted in the formation ofnucleoprotein complexes that reached saturation at anORF34p2 : M13 ssDNA molar ratio (R) of 400:1, in fullagreement with the EMSA data obtained under verysimilar experimental conditions (Fig. 3B). In AFM experi-ments, saturation was established by the appearance ofmany unbound ORF34P2 particles spread over the micasurface and by the observation of fully coated M13 mol-ecules (Fig. 5F). Under such conditions, the ORF34p2–ssDNA complexes formed packed ‘beads-on-a-string’structures that were more extended than the M13 ssDNAalone. However, from inspection of the images, it wasclear that the overall contour length of the ORF34p2–ssDNA nucleoprotein complexes was significantly lessthan what would be expected for the fully extendedssDNA (Fig. 5E and F). This result strongly indicated thatthe DNA binding mode of ORF34p2 involves wrapping ofthe ssDNA around the protein, as anticipated for an SSBprotein. The analysis of 30 molecules of M13 ssDNA in thepresence of saturating amounts of ORF34p2 (Fig. 5G)showed that, on average, there were about 63 ORF34p2
particles, possibly tetramers, bound to a single M13ssDNA molecule. Given the size of M13 ssDNA (7249 nt),these data suggest that binding of an ORF34p2 particle tossDNA involves ~115 nt, either wrapped around theprotein or constituting the DNA linker between adjacentparticles. Unfortunately, the compact structure of the com-plexes and the broadening effect of the tip did not permitdirect visualization of the DNA linker. On a similar study onEcoSSB (Griffith and Shores, 1985), stretches of about30 nt protein-free DNA were found.
In order to better quantify the length of ssDNA occupiedby a single ORF34p2 particle and to determine the degreeof DNA compaction consequent to the binding, we per-formed experiments with a linear ssDNA fragment(1204 nt) obtained by asymmetric PCR. This procedureresults in a mixture of single-stranded and double-stranded DNA fragments that can easily be discernedfrom AFM images because dsDNA is thicker and moreextended than ssDNA (data not shown). Figure 5H showsa set of images obtained with this DNA substrate undersaturating concentrations of ORF34p2 (R = 400:1). On theone hand, these images show that ORF34p2 does not binddsDNA, confirming the EMSA experiments. On the otherhand, ssDNA fragments were completely covered byORF34p2 and formed more extended conformations thanthose observed with M13 ssDNA. In this case, the limitedsize of the ssDNA fragment allowed a more accuratedetermination of the nucleoprotein filament contour lengthand a more precise evaluation of the number of ORF34p2
Structure & function of lactococcus phage p2 ORF34 1163
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

particles bound to a single ssDNA fragment (Fig. 5I). Inparticular, in a sample of 170 ORF34p2–ssDNA complexesthe average number of SSB tetramers bound to the 1204-nt-long ssDNA was 9 � 0.3, which implies an averageSSB nucleo-particle spacing of 134 � 25 nt. Upon com-parison with site sizes for EcoSSB [up to 65 nt/
homotetramer (Lohman and Overman, 1985)] and withour own results from EMSA (72 nt/homotetramer), thisresult might indicate that the observed ORF34p2 nucleo-particles could correspond to ssDNA bound to ORF34p2
octamers. Indeed, octamers, observed as ‘beads’ by elec-tron microscopy (Chrysogelos and Griffith, 1982), play a
Fig. 5. AFM images of ORF34p2-DNA complexes. Image of M13 ssDNA without ORF34p2: deposition with magnesium (A) and spermidine(B-a: typical view, B-b: case of a partially extended M13 ssDNA – image size: 250 nm). From (C) to (F), increasing concentrations of ORF34p2
were used to titrate M13 ssDNA (0.5 nM): 30 nM (C), 50 nM (D), 100 nM (E) and 200 nM (F).G. Representative 3D image of an ORF34p2-saturated M13 ssDNA (Image size: 300 nm).H. Collage of AFM images showing representative examples of the 170 ORF34p2–ssDNA(linear) complexes analysed. Contour lengthdistribution (I) from these ORF34p2–ssDNA(linear) complexes.
1164 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170
role in the context of the EcoSSB65 binding mode(Lohman and Ferrari, 1994). Further work addressing thessDNA binding modes of ORF34p2 should elucidate thefunctions of the different oligomeric states of the protein.
Contour length measurements of these 170 ORF34p2–ssDNA complexes (Fig. 5I) resulted in an average contourlength value of 164 � 4.6 nm. Assuming that the nucle-otide step in ssDNA is about 0.6 nm (Murphy et al., 2004),the expected contour length of the 1204-nt-long ssDNAfragment would be 722 nm. Therefore, under in vitro con-ditions of protein saturation, binding of ORF34p2 results ina ~77% compaction of the ssDNA.
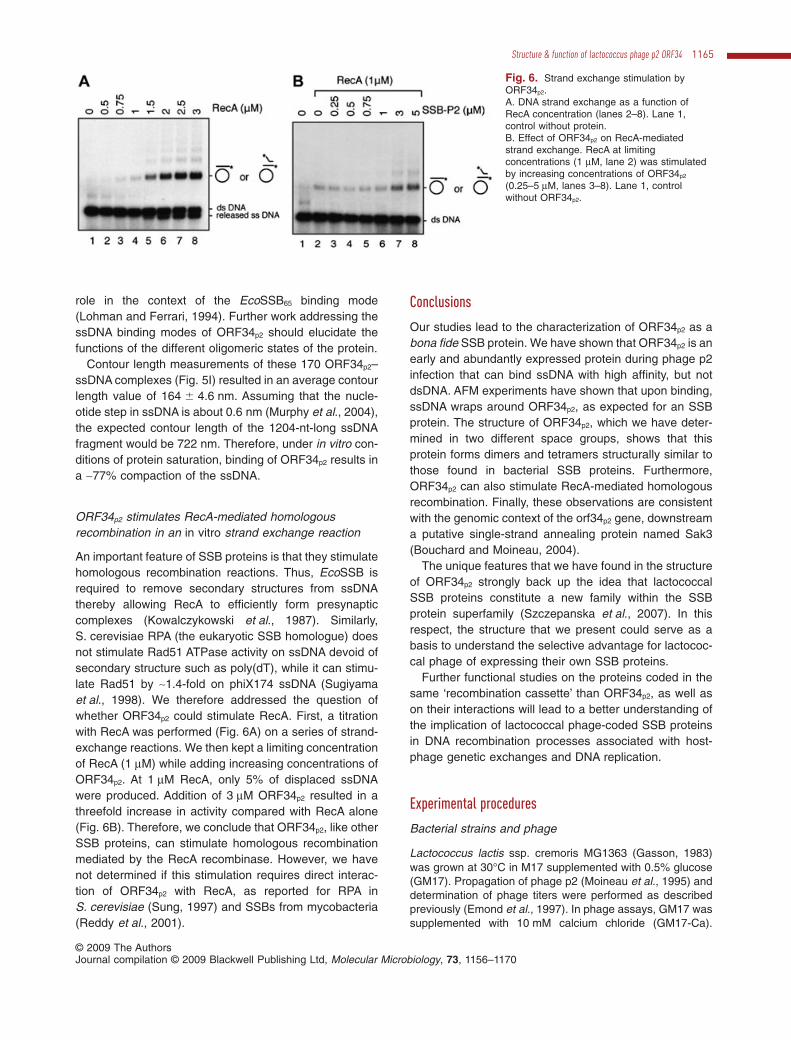
ORF34p2 stimulates RecA-mediated homologousrecombination in an in vitro strand exchange reaction
An important feature of SSB proteins is that they stimulatehomologous recombination reactions. Thus, EcoSSB isrequired to remove secondary structures from ssDNAthereby allowing RecA to efficiently form presynapticcomplexes (Kowalczykowski et al., 1987). Similarly,S. cerevisiae RPA (the eukaryotic SSB homologue) doesnot stimulate Rad51 ATPase activity on ssDNA devoid ofsecondary structure such as poly(dT), while it can stimu-late Rad51 by ~1.4-fold on phiX174 ssDNA (Sugiyamaet al., 1998). We therefore addressed the question ofwhether ORF34p2 could stimulate RecA. First, a titrationwith RecA was performed (Fig. 6A) on a series of strand-exchange reactions. We then kept a limiting concentrationof RecA (1 mM) while adding increasing concentrations ofORF34p2. At 1 mM RecA, only 5% of displaced ssDNAwere produced. Addition of 3 mM ORF34p2 resulted in athreefold increase in activity compared with RecA alone(Fig. 6B). Therefore, we conclude that ORF34p2, like otherSSB proteins, can stimulate homologous recombinationmediated by the RecA recombinase. However, we havenot determined if this stimulation requires direct interac-tion of ORF34p2 with RecA, as reported for RPA inS. cerevisiae (Sung, 1997) and SSBs from mycobacteria(Reddy et al., 2001).
Conclusions
Our studies lead to the characterization of ORF34p2 as abona fide SSB protein. We have shown that ORF34p2 is anearly and abundantly expressed protein during phage p2infection that can bind ssDNA with high affinity, but notdsDNA. AFM experiments have shown that upon binding,ssDNA wraps around ORF34p2, as expected for an SSBprotein. The structure of ORF34p2, which we have deter-mined in two different space groups, shows that thisprotein forms dimers and tetramers structurally similar tothose found in bacterial SSB proteins. Furthermore,ORF34p2 can also stimulate RecA-mediated homologousrecombination. Finally, these observations are consistentwith the genomic context of the orf34p2 gene, downstreama putative single-strand annealing protein named Sak3(Bouchard and Moineau, 2004).
The unique features that we have found in the structureof ORF34p2 strongly back up the idea that lactococcalSSB proteins constitute a new family within the SSBprotein superfamily (Szczepanska et al., 2007). In thisrespect, the structure that we present could serve as abasis to understand the selective advantage for lactococ-cal phage of expressing their own SSB proteins.
Further functional studies on the proteins coded in thesame ‘recombination cassette’ than ORF34p2, as well ason their interactions will lead to a better understanding ofthe implication of lactococcal phage-coded SSB proteinsin DNA recombination processes associated with host-phage genetic exchanges and DNA replication.
Experimental procedures
Bacterial strains and phage
Lactococcus lactis ssp. cremoris MG1363 (Gasson, 1983)was grown at 30°C in M17 supplemented with 0.5% glucose(GM17). Propagation of phage p2 (Moineau et al., 1995) anddetermination of phage titers were performed as describedpreviously (Emond et al., 1997). In phage assays, GM17 wassupplemented with 10 mM calcium chloride (GM17-Ca).
Fig. 6. Strand exchange stimulation byORF34p2.A. DNA strand exchange as a function ofRecA concentration (lanes 2–8). Lane 1,control without protein.B. Effect of ORF34p2 on RecA-mediatedstrand exchange. RecA at limitingconcentrations (1 mM, lane 2) was stimulatedby increasing concentrations of ORF34p2
(0.25–5 mM, lanes 3–8). Lane 1, controlwithout ORF34p2.
Structure & function of lactococcus phage p2 ORF34 1165
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

Phage DNA was isolated as reported elsewhere (Deveauet al., 2006).
Cloning, expression and purification of ORF34p2
Phage p2 orf34 gene was amplified by PCR from phage p2genomic DNA using specific Gateway primers containing attBsequences at both ends, a ribosome binding site and anN-terminal His6-tag coding sequence, and was cloned byrecombination in a Gateway pETG-20 A vector kindly pro-vided by Arie Geerlof (EMBL). Due to the lack of methioninesin ORF34p2 sequence and in order to introduce selenium fordiffraction studies, the orf34 gene was recloned introducingtwo leucine-to-methionine substitutions: L49M, L55M (M2B-ORF34p2) using the QuickChange Site-Directed MutagenesisKit (Stratagene).
Protein expression experiments were carried out withthe Rosetta (DE3)pLyS (Novagen) strain. After an over-night induction with 1 mM isopropyl 1-thio-beta-D-galactopyranoside at 25°C, cells were harvested by centrifu-gation for 10 min at 4000 g. Mutant M2B-ORF34p2 wasexpressed in minimal conditions in presence of seleno-methionine.
Bacterial pellets were resuspended in 40 ml l-1 of culture oflysis buffer (Tris 50 mM, NaCl 300 mM, imidazole 10 mM,pH 8.0) supplemented with 0.25 mg ml-1 lysozyme, 1 mg ml-1
DNAse, 20 mM MgS04 and antiproteases (complete EDTA-free antiproteases, Roche) and frozen at -80°C. Afterthawing and sonication, lysates were cleared by a 30 mincentrifugation at 12 000 g. Overexpressed proteins werepurified on a Pharmacia Äkta FPLC by nickel-affinity chroma-tography (His-Trap 5 ml column, GE Healthcare) using a stepgradient of imidazole (at 25 and 250 mM). The N-terminalHis6-tag was removed by digestion with TEV protease fol-lowed by Ni-NTA affinity chromatography, to exclude both thetag peptide and the protease. A final purification step wasconducted on a preparative size-exclusion column (Super-dex200 HR26/60) equilibrated in 10 mM Tris, 300 mM NaClpH 8.0. In the case of the selenomethionyl (SeMet) M2B-ORF34p2, 1 mM Tris(2-carboxyethyl)phosphine hydrochloridewas added to the gel filtration buffer to stabilize the redoxstate of the protein. Purified proteins were concentrated usingAmicon Ultra-15 ml filtration systems (Millipore) and charac-terized by SDS-PAGE and trypsin peptide mass fingerprint bymatrix-assisted laser desorption ionization time-of-flight massspectrometry (Bruker Autoflex). E. coli RecA was purified asdescribed previously (Eggleston et al., 1997), while E. coliSSB was purchased from Promega.
Size-exclusion chromatography and light scattering
SEC-MALS/UV/RI experiments were carried out on an Alli-ance 2695 HPLC system (Waters) using a silica-basedKW402.5 column (Shodex) equilibrated in Tris 10 mM,300 mM NaCl, pH 8.0 at a flow of 0.35 ml min-1. The proteinwas injected at a concentration of 3 mg ml-1. Detection wasachieved by using a triple-angle light-scattering detector(Mini-DAWN TREOS, Wyatt Technology), a quasi-elastic lightscattering instrument (Dynapro, Wyatt Technology) and adifferential refractometer (Optilab rEX, Wyatt Technology).
Molecular weight and hydrodynamic radius determinationwas performed by ASTRA V software (Wyatt Technology)using a dn/dc value of 0.185 ml g-1.
Phage infection experiments
Lactococcus lactis ssp. cremoris MG1363 was grown inGM17-Ca until the optical density at 600 nm reached 0.2,then the virulent phage p2 was added at a multiplicity ofinfection of 5. Samples were taken at 5 min intervals andflash-frozen (-80°C). Cell pellets were resuspended in10 mM Tris-Cl, pH 8.0, 1 mM EDTA, 0.3% SDS, 60 mM DTT,protease inhibitors (Roche Diagnostics) and glass beads(106 mm, Sigma-Aldrich). The mixture was vortexed with aMini-Beadbeater-8 cell (BioSpec Products) five times for1 min each. Between treatments, cell suspensions werechilled on ice for 1 min. The lysed culture was centrifuged andthe cytoplasmic extracts were dosed by RC DC protein assay(Bio-Rad). Then, 350 mg of total protein was precipitated bythe methanol/chloroform/water method of Wessel and Flügge(Wessel and Flügge, 1984). Pellets were dried and reconsti-tuted overnight with 450 ml of rehydratation buffer (7 M urea,2 M thiourea, 50 mM DTT, 4% CHAPS and traces of bro-mophenol blue).
Two-dimensional gel electrophoresis
Two-dimensional gel electrophoresis was performed using theImmobiline/polyacrylamide system. Isoelectric focusing (IEF)was performed with 24 cm IPG strips pH 4–7 (GE Healthcare).The reconstituted protein sample (see above) was applied onIPG DryStrips (IPGphor II, Isoelectric Focusing Unit, GEHealthcare) using the in-gel sample rehydration techniqueaccording to the manufacturer’s instructions. After rehydrationat 20°C for 12 h, IEF (50 mA/strip) was conducted at 500 V for1 h, 1000 V for 1 h, at gradient steps from 1000 to 8000 V for3 h and then at 8000 V for 6 h 30 min, to reach a total of60–70 kVh. The temperature was maintained at 20°C. AfterIEF, each strip was equilibrated for 15 min in 10 ml of equili-bration buffer 1 (7 M urea, 1% DTT, 36% glycerol, 2% SDS,50 mM Tris-Cl, pH 8.8) and then in 10 ml of equilibration buffer2 (7 M urea, 2.5% iodoacetamide, 36% glycerol, 2% SDS,50 mM Tris-Cl, pH 8.8) for 15 min. For the second dimension,vertical gradient 10–20% SDS-PAGE was run for about 4 h(17 W gel-1; Ettan DALTsix, GE Healthcare). The gels werestained using Colloidal CBB (0.1% CBB G-250, 20% metha-nol, 10% ammonium sulphate, 2% ortho-phosphoric acid).Scanning was carried out with an ImageScanner (GE Health-care) and image analysis was performed using ImageMaster2D Elite software. Spots were cut out of the gel, digested withtrypsin and identified by liquid chromatography-tandem massspectrometry at the Centre Protéomique de l’Est du Québec(Quebec city, Quebec, Canada).
Gel retardation assays
Electrophoretic mobility shift assay experiments with 100 ntssDNA or 100 bp dsDNA fragments were performed asfollows: in a volume of 10 ml binding buffer (25 mM MOPSpH 7.0, 60 mM NaCl, 1% Tween, 2 mM DTT) 32P-labelled
1166 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

DNA was added to a final concentration of 100 nM and thesolution was incubated for 5 min at 37°C. Either ORF34p2 orE. coli SSB was added to final concentrations of 100, 250,500 and 1000 nM and the reactions were incubated for other10 min at 37°C. Complexes were fixed by the addition of0.2% glutaraldehyde followed by a 15 min incubation at 37°C.The reactions were run onto a 4.2% non-denaturing polyacrylamide gel on 1¥ Tris-Glycine buffer (50 mM Tris-Cl,50 mM glycine, pH 8.8). Gels were dried on DE81 filter paperfollowed by autoradiography. The dsDNA substrate was pre-pared by annealing two complementary oligonucleotides100 nt long and by purifying the annealed product on a 10%non-denaturing poly acrylamide gel. The sequence ofthe 100-mer was 5′-GGGCGAATTGGGCCCGACGTCGCATGCTCCTCTAGACTCGAGGAATTCGGTACCCCGGGTTCGAAATCGATAAGCTTACAGTCTCCATTTAAAGGACAAG-3′.
Electrophoretic mobility shift assay experiments with circu-lar M13 ssDNA (7249 nt long) (Sigma Aldrich) wereperformed under the same conditions used for AFMexperiments. Namely, ORF34p2–ssDNA complexes wereassembled using 100 ng of M13 ssDNA (0.5 nM final concen-tration) and increasing amounts of protein (15, 30, 50, 100,200, 400 nM) in 20 mM Tris-HCl pH 7.5, 20 mM NaCl, 50 mMSpermidine. The reactions were incubated for 10 min at 37°Cand loaded in a 1% agarose gel after addition of a gel loadingbuffer containing glycerol. Electrophoresis was run at 4°C inTBE buffer at 3 V cm-1. After a 4 h run, the gel was stainedwith ethidium bromide (0.5 mg ml-1) for 60 min.
Surface plasmon resonance
Experiments were performed using a Biacore 1000 instru-ment (Biacore) at 25°C. The chip streptavidin was firstwashed with 10 mM NaOH, 1 M NaCl (three times 100 ml at40 ml min-1) to eliminate loosely bound streptavidin.Biotinylated-DNA 40-mer (ssDNA; 45 mg ml-1) in 10 mM Trisbuffer, 150 mM NaCl, pH 8.0, 0.005% (v/v) P20 were fixed asligand at 550 resonance units). ORF34p2 at concentrationfrom 23 to 375 nM in 10 mM Tris, 300 mM NaCl, pH 8.0,0.005% (v/v) P20, was used as analyte (80 ml at 10 ml min-1).Regeneration was achieved by injection of 4 M MgCl2 (5 ml at40 ml min-1). The resonance unit signal at different analyteconcentrations was corrected for the buffer contribution in thesame flow-cell and for aspecific interactions on the referenceflow-cell. The KD values were estimated assuming a 1:1Langmuir model (BIAevaluation Software).
Crystallization, data collection, structure determinationand refinement
The chromatographic peaks of native ORF34p2 and (SeMet)M2B-ORF34p2 matching the theoretical molecular weight of atetramer were concentrated in each case to 7 mg ml-1 andsubjected to crystallization screening with a Cartesiannanodrop-dispensing robot (Sulzenbacher et al., 2002).Native ORF34p2 and (SeMet) M2B-ORF34p2 crystals wereobtained in 0.1 M Sodium Cacodylate, pH 6.2, 45% MMEPEG 2000 and 0.1 M HEPES, pH 7.2, 60% MME PEG 550respectively. In both cases crystals appeared after 3 daysand were fished using a nylon loop and, as the crystallization
condition was found to be cryoprotectant, directly flash-cooled in liquid nitrogen. Native diffraction data were col-lected to 2.6 Å resolution at the European SynchrotronRadiation Facility (Grenoble, France) on beamline ID23-2.Data were integrated and reduced using MOSFLM and thenscaled with the program SCALA from CCP4 suite (Collabora-tive Computational Project Number 4, 1994).
Diffraction data from a (SeMet) M2B-ORF34p2 crystal werecollected to 2.1 Å resolution at Soleil (Saint-Aubin, Essonne,France) on beamline Proxima 1. These data were integratedusing XDS (Kabsch, 1993) and scaled and reduced with theprogram XSCALE (Kabsch, 1993). The program SHELXD (Usonand Sheldrick, 1999) was used to locate the Se atoms andSHELXE (Sheldrick, 2002) to produce the initial phases. Thefirst model of the (SeMet) M2B-ORF34p2 protein was builtusing the program ARP-WARP (Cohen et al., 2004). This modelwas improved by successive cycles of manual rebuilding withCOOT (Emsley and Cowtan, 2004) and refinement withREFMAC5 (Murshudov et al., 1997).
The native ORF34p2 structure was solved by molecularreplacement with MOLREP (Vagin and Teplyakov, 1997) usingthe best defined parts of the (SeMet) M2B-ORF34p2 structureas the search model. The native model was completed usingRESOLVE (Terwilliger, 2002), manually rebuilt using COOT andrefined with REFMAC5. TLS (Traslation/Libration/Screw)segments (Winn et al., 2001), defined with the help of the TLSMotion Determination server (http://skuld.bmsc.washington.edu/~tlsmd/), were used during refinement. In the nativestructure, non-crystallographic symmetry restraints wereapplied to all protein backbone at the initial stages ofrefinement. These restraints were loosened in further refine-ment cycles and the most flexible loops excluding from them.
Analysis of the stereochemical quality of the native modelwas performed using Molprobity (Lovell et al., 2003). Datacollection and refinement statistics are summarized inTable 1. All structural figures were prepared with CHIMERA
(Pettersen et al., 2004). Atomic co-ordinates and experimen-tal structure factors of native ORF34p2 and (SeMet) M2B-ORF34p2 have been deposited within the PDB and areaccessible under the codes 2WKC and 2WKD respectively.
Atomic force microscopy
ORF34p2–M13 ssDNA complexes were prepared by mixingM13 ssDNA at a final concentration of 0.5 nM in buffer con-taining 20 mM Tris pH 8.0 and 20 mM NaCl with increasingconcentrations of ORF34p2 protein, from 30 up to 200 nM.Reactions were incubated at 25°C for 5 min and then sper-midine (Fluka) was added to a final concentration of 50 mM.Spermidine was used as deposition agent instead of magne-sium because it allows a better spreading of the nucleopro-tein filaments onto mica (Hamon et al., 2007). A 5 ml droplet ofthe reaction was deposited onto freshly cleaved ruby mica for1 min, followed by rinsing with water milliQ (Millipore) anddrying with a weak flux of nitrogen.
Linear ssDNA fragment (1204 nt long) were obtained byasymmetric PCR using pNEB193 plasmid as a template andan unequal primer concentration. The PCR mixture con-tained: 20 ng dsDNA template, 25 pmol pNEB-For2 primer,0.25 pmol pNEB-Rev primer, 50 mM dNTPs (Fermentas),1.5 mM MgCl2, 10¥ PCR buffer and 2.5 U Taq DNA Poly-
Structure & function of lactococcus phage p2 ORF34 1167
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

merase (Fermentas) was thermally cycled for 35 times asfollows: 95°C for 60 s, 58°C for 30 s, 72°C for 2 min. TheDNA product was purified using PCR Clean-up NucleospinExtract II (Macherey-Nagel).
The AFM images were obtained in air with a Nanoscope IIImicroscope (Digital Instruments) operating in tapping mode.All operations were done at room temperature. Commercialdiving board silicon cantilevers (mmasch) were used. Themicroscope was equipped with a type E scanner(12 mm ¥ 12 mm). Images (512 ¥ 512 pixels) were collectedwith a scan size of 2 or 4 mm at a scan rate varying betweentwo and four scan lines per second.
The AFM images were analysed using software written inMatlab (MathWorks). Contour length measurements of thenucleoprotein filaments were performed as described else-where (Rivetti and Codeluppi, 2001).
Counting of ORF34p2 particles coating ssDNA was donemanually. Only molecules that were completely visible in theimage and did not have an ambiguous shape were consid-ered. Data were plotted using Sigma Plot (Systat Software).
Strand exchange
Reactions (10 ml) contained purified circular single-strandedpPB4.3 DNA (15 mM) with the indicated concentrations ofE. coli RecA and ORF34p2 in TD buffer (25 mM Tris-AcetatepH 7.5, 8 mM MgCl2, 1 mM DTT, 1 mM ATP, 20 mM creatinephosphate, 5 U ml-1 phosphocreatine kinase). ORF34p2 wasadded first to the reaction.After 5 min at 37°C, 32P-end labelledpPB4.3 DNA (400 bp fragment, 1.38 mM) was added andincubation was continued for 90 min. Reaction products weredeproteinized by addition of one-fifth volume of stop buffer(0.1 M Tris-HCl, pH 7.5, 0.1 M MgCl2, 3% SDS, 5 mg ml-1
ethidium bromide and 10 mg ml-1 proteinase K) followed by45 min incubation at 37°C. Labelled DNAproducts were analy-sed by electrophoresis through 0.8% TAE agarose gels con-taining 1 mg ml-1 ethidium bromide, run at 4.3 V cm-1, driedonto DE81 filter paper and visualized by autoradiography.
Acknowledgements
This work was supported in part by Marseille-Nice Géno-pole, by the Natural Sciences and Engineering ResearchCouncil of Canada (Strategic Grant to S.M. and J.Y.M.), bya grant of Università Italo-francese (Bando Vinci 2007-capII)to E.S. and by a grant from Fondazione Cariparma to C.R.A.H.M. is the recipient of a Clarín postdoctoral scholarship(FICYT) from the Gobierno del Principado de Asturias(PCTI 2006-09). We thank the Centro InterdipartimentaleMisure of the University of Parma for access to the AFMfacility. We also thank the staff from the ID23-2 beamline atthe European Synchrotron Radiation Facility (Grenoble,France) and from the Proxima 2 beamline at the Soleil Syn-chrotron (Saint-Aubin, Essonne, France) for their help andsupport.
References
Ayora, S., Langer, U., and Alonso, J.C. (1998) Bacillus sub-tilis DnaG primase stabilises the bacteriophage SPP1G40P helicase-ssDNA complex. FEBS Lett 439: 59–62.
Bleuit, J.S., Xu, H., Ma, Y., Wang, T., Liu, J., and Morrical,S.W. (2001) Mediator proteins orchestrate enzyme-ssDNAassembly during T4 recombination-dependent DNA repli-cation and repair. Proc Natl Acad Sci USA 98: 8298–8305.
Bouchard, J.D., and Moineau, S. (2004) Lactococcal phagegenes involved in sensitivity to AbiK and their relation tosingle-strand annealing proteins. J Bacteriol 186: 3649–3652.
Chrysogelos, S., and Griffith, J. (1982) Escherichia colisingle-strand binding protein organizes single-strandedDNA in nucleosome-like units. Proc Natl Acad Sci USA 79:5803–5807.
Cohen, S.X., Morris, R.J., Fernandez, F.J., Ben Jelloul, M.,Kakaris, M., Parthasarathy, V, et al. (2004) Towards com-plete validated models in the next generation of ARP/wARP. Acta Crystallogr D Biol Crystallogr 60: 2222–2229.
Collaborative Computational Project Number 4 (1994) TheCCP4 suite: programs for protein crystallography. ActaCryst D 50: 760–763.
Deveau, H., Labrie, S.J., Chopin, M., and Moineau, S. (2006)Biodiversity and classification of lactococcal phages. ApplEnviron Microbiol 72: 4338–4346.
DiDonato, M., Krishna, S.S., Schwarzenbacher, R., McMul-lan, D., Jaroszewski, L., Miller, M.D, et al. (2006) Crystalstructure of a single-stranded DNA-binding protein(TM0604) from Thermotoga maritima at 2.60 A resolution.Proteins 63: 256–260.
Eggleston, A.K., Mitchell, A.H., and West, S.C. (1997) In vitroreconstitution of the late steps of genetic recombination inE. coli. Cell 89: 607–617.
Emond, E., Holler, B.J., Boucher, I., Vandenbergh, P.A.,Vedamuthu, E.R., Kondo, J.K., and Moineau, S. (1997)Phenotypic and genetic characterization of the bacterioph-age abortive infection mechanism AbiK from Lactococcuslactis. Appl Environ Microbiol 63: 1274–1283.
Emsley, P., and Cowtan, K. (2004) Coot: model-building toolsfor molecular graphics. Acta Crystallogr D Biol Crystallogr60: 2126–2132.
Folmer, R.H., Folkers, P.J., Kaan, A., Jonker, A.J., Aelen,J.M., Konings, R.N., and Hilbers, C.W. (1994) Secondarystructure of the single-stranded DNA binding proteinencoded by filamentous phage Pf3 as determined by NMR.Eur J Biochem 224: 663–676.
Folmer, R.H., Nilges, M., Konings, R.N., and Hilbers, C.W.(1995) Solution structure of the single-stranded DNAbinding protein of the filamentous Pseudomonas phagePf3: similarity to other proteins binding to single-strandednucleic acids. EMBO J 14: 4132–4142.
Gasior, S.L., Olivares, H., Ear, U., Hari, D.M., Weichselbaum,R., and Bishop, D.K. (2001) Assembly of RecA-like recom-binases: distinct roles for mediator proteins in mitosis andmeiosis. Proc Natl Acad Sci USA 98: 8411–8418.
Gasson, M.J. (1983) Plasmid complements of Streptococcuslactis NCDO 712 and other lactic streptococci afterprotoplast-induced curing. J Bacteriol 154: 1–9.
Genschel, J., Curth, U., and Urbanke, C. (2000) Interaction ofE. coli single-stranded DNA binding protein (SSB) withexonuclease I. The carboxy-terminus of SSB is the recog-nition site for the nuclease. Biol Chem 381: 183–192.
Griffith, J., and Shores, C.G. (1985) RecA protein rapidlycrystallizes in the presence of spermidine: a valuable step
1168 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

in its purification and physical characterization. Biochem-istry 24: 158–162.
Haaber, J., Moineau, S., Fortier, L., and Hammer, K. (2008)AbiV, a novel antiphage abortive infection mechanism onthe chromosome of Lactococcus lactis subsp. cremorisMG1363. Appl Environ Microbiol 74: 6528–6537.
Hamon, L., Pastre, D., Dupaigne, P., Le Breton, C., Le Cam,E., and Pietrement, O. (2007) High-resolution AFM imagingof single-stranded DNA-binding (SSB) protein – DNAcomplexes. Nucleic Acids Res 35: e58.
Hay, N., and Cohen, G. (1983) Requirement of E. coli DNAsynthesis functions for the lytic replication of bacteriophageP1. Virology 131: 193–206.
He, Z., Rezende, L.F., Willcox, S., Griffith, J.D., and Richard-son, C.C. (2003) The carboxyl-terminal domain of bacte-riophage T7 single-stranded DNA-binding proteinmodulates DNA binding and interaction with T7 DNApolymerase. J Biol Chem 278: 29538–29545.
Hollis, T., Stattel, J.M., Walther, D.S., Richardson, C.C., andEllenberger, T. (2001) Structure of the gene 2.5 protein, asingle-stranded DNA binding protein encoded by bacte-riophage T7. Proc Natl Acad Sci USA 98: 9557–9562.
Iftode, C., Daniely, Y., and Borowiec, J.A. (1999) Replicationprotein A (RPA): the eukaryotic SSB. Crit Rev Biochem MolBiol 34: 141–180.
Jackson, D., Dhar, K., Wahl, J.K., Wold, M.S., and Borgstahl,G.E.O. (2002) Analysis of the human replication protein A:Rad52 complex: evidence for crosstalk between RPA32,RPA70, Rad52 and DNA. J Mol Biol 321: 133–148.
Kabsch, W. (1993) Automatic processing of rotation diffrac-tion data from crystals of initially unknown symmetry andcell constants. J Appl Crystallogr 26: 795–800.
Kim, Y.T., and Richardson, C.C. (1993) Bacteriophage T7gene 2.5 protein: an essential protein for DNA replication.Proc Natl Acad Sci USA 90: 10173–10177.
Kowalczykowski, S.C., Clow, J., and Krupp, R.A. (1987)Properties of the duplex DNA-dependent ATPase activity ofEscherichia coli RecA protein and its role in branchmigration. Proc Natl Acad Sci USA 84: 3127–3131.
Krissinel, E., and Henrick, K. (2004) Secondary-structurematching (SSM), a new tool for fast protein structure align-ment in three dimensions. Acta Crystallogr D Biol Crystal-logr 60: 2256–2268.
Lohman, T.M., and Ferrari, M.E. (1994) Escherichia colisingle-stranded DNA-binding protein: multiple DNA-binding modes and cooperativities. Annu Rev Biochem 63:527–570.
Lohman, T.M., and Overman, L.B. (1985) Two binding modesin Escherichia coli single strand binding protein-single stranded DNA complexes. Modulation by NaClconcentration. J Biol Chem 260: 3594–3603.
Lovell, S.C., Davis, I.W., Arendall, W.B., 3rd, de Bakker,P.I.W., Word, J.M., Prisant, M.G., et al. (2003) Structurevalidation by Calpha geometry: phi, psi and Cb deviation.Proteins 50: 437–450.
Matsumoto, T., Morimoto, Y., Shibata, N., Kinebuchi, T.,Shimamoto, N., Tsukihara, T., and Yasuoka, N. (2000)Roles of functional loops and the C-terminal segment of asingle-stranded DNA binding protein elucidated by X-Raystructure analysis. J Biochem 127: 329–335.
Meyer, R.R., and Laine, P.S. (1990) The single-stranded
DNA-binding protein of Escherichia coli. Microbiol Rev 54:342–380.
Miller, E.S., Heidelberg, J.F., Eisen, J.A., Nelson, W.C.,Durkin, A.S., Ciecko, A, et al. (2003) Complete genomesequence of the broad-host-range vibriophage KVP40:comparative genomics of a T4-related bacteriophage.J Bacteriol 185: 5220–5233.
Moineau, S. (1999) Applications of phage resistance in lacticacid bacteria. Antonie Van Leeuwenhoek 76: 377–382.
Moineau, S., Walker, S.A., Vedamuthu, E.R., and Vanden-bergh, P.A. (1995) Cloning and sequencing of LlaDCHI[corrected] restriction/modification genes from Lactococ-cus lactis and relatedness of this system to the Streptococ-cus pneumoniae DpnII system. Appl Environ Microbiol 61:2193–2202.
Mosca, R., and Schneider, T.R. (2008) RAPIDO: a webserver for the alignment of protein structures in the pres-ence of conformational changes. Nucleic Acids Res 36:W42–W46.
Murphy, M.C., Rasnik, I., Cheng, W., Lohman, T.M., and Ha,T. (2004) Probing single-stranded DNA conformationalflexibility using fluorescence spectroscopy. Biophys J 86:2530–2537.
Murshudov, G.N., Vagin, A.A., and Dodson, E.J. (1997)Refinement of macromolecular structures by themaximum-likelihood method. Acta Crystallogr D Biol Crys-tallogr 53: 240–255.
Murzin, A.G. (1993) OB(oligonucleotide/oligosaccharidebinding)-fold: common structural and functional solution fornon-homologous sequences. EMBO J 12: 861–867.
Pettersen, E.F., Goddard, T.D., Huang, C.C., Couch, G.S.,Greenblatt, D.M., Meng, E.C., and Ferrin, T.E. (2004)UCSF Chimera – a visualization system for exploratoryresearch and analysis. J Comput Chem 25: 1605–1612.
Ploquin, M., Bransi, A., Paquet, E.R., Stasiak, A.Z., Stasiak,A., Yu, X., et al. (2008) Functional and structural basis fora bacteriophage homolog of human RAD52. Curr Biol 18:1142–1146.
Raghunathan, S., Ricard, C.S., Lohman, T.M., and Waksman,G. (1997) Crystal structure of the homo-tetrameric DNAbinding domain of Escherichia coli single-stranded DNA-binding protein determined by multiwavelength x-ray diffrac-tion on the selenomethionyl protein at 2.9-A resolution. ProcNatl Acad Sci USA 94: 6652–6657.
Raghunathan, S., Kozlov, A.G., Lohman, T.M., andWaksman, G. (2000) Structure of the DNA bindingdomain of E. coli SSB bound to ssDNA. Nat Struct Biol 7:648–652.
Reddy, M.S., Guhan, N., and Muniyappa, K. (2001) Charac-terization of single-stranded DNA-binding proteins fromMycobacteria. The carboxyl-terminal of domain of SSB isessential for stable association with its cognate RecAprotein. J Biol Chem 276: 45959–45968.
Ricagno, S., Campanacci, V., Blangy, S., Spinelli, S., Trem-blay, D., Moineau, S, et al. (2006) Crystal structure of thereceptor-binding protein head domain from Lactococcuslactis phage bIL170. J Virol 80: 9331–9335.
Rivetti, C., and Codeluppi, S. (2001) Accurate length deter-mination of DNA molecules visualized by atomic forcemicroscopy: evidence for a partial B- to A-form transition onmica. Ultramicroscopy 87: 55–66.
Structure & function of lactococcus phage p2 ORF34 1169
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170

Saikrishnan, K., Jeyakanthan, J., Venkatesh, J., Acharya, N.,Sekar, K., Varshney, U., and Vijayan, M. (2003) Structureof Mycobacterium tuberculosis single-stranded DNA-binding protein. Variability in quaternary structure and itsimplications. J Mol Biol 331: 385–393.
Sancar, A., Williams, K.R., Chase, J.W., and Rupp, W.D.(1981) Sequences of the ssb gene and protein. Proc NatlAcad Sci USA 78: 4274–4278.
Savvides, S.N., Raghunathan, S., Futterer, K., Kozlov, A.G.,Lohman, T.M., and Waksman, G. (2004) The C-terminaldomain of full-length E. coli SSB is disordered even whenbound to DNA. Protein Sci 13: 1942–1947.
Sciara, G., Blangy, S., Siponen, M., McGrath, S., van Sin-deren, D., Tegoni, M, et al. (2008) A topological model ofthe baseplate of lactococcal phage Tuc2009. J Biol Chem283: 2716–2723.
Shamoo, Y., Friedman, A.M., Parsons, M.R., Konigsberg,W.H., and Steitz, T.A. (1995) Crystal structure of a replica-tion fork single-stranded DNA binding protein (T4 gp32)complexed to DNA. Nature 376: 362–366.
Sheldrick, G.M. (2002) Macromolecular phasing withSHELXE. Z Kristallogr 217: 644–650.
Shinohara, A., Shinohara, M., Ohta, T., Matsuda, S., andOgawa, T. (1998) Rad52 forms ring structures and coop-erates with RPA in single-strand DNA annealing. GenesCells 3: 145–156.
Skinner, M.M., Zhang, H., Leschnitzer, D.H., Guan, Y.,Bellamy, H., Sweet, R.M, et al. (1994) Structure of thegene V protein of bacteriophage f1 determined by multi-wavelength x-ray diffraction on the selenomethionylprotein. Proc Natl Acad Sci USA 91: 2071–2075.
Spinelli, S., Desmyter, A., Verrips, C.T., de Haard, H.J.W.,Moineau, S., and Cambillau, C. (2006) Lactococcal bacte-riophage p2 receptor-binding protein structure suggests acommon ancestor gene with bacterial and mammalianviruses. Nat Struct Mol Biol 13: 85–89.
Su, S., Gao, Y.G., Zhang, H., Terwilliger, T.C., and Wang,A.H. (1997) Analyses of the stability and function of threesurface mutants (R82C, K69H, and L32R) of the gene Vprotein from Ff phage by X-ray crystallography. Protein Sci6: 771–780.
Sugiyama, T., New, J.H., and Kowalczykowski, S.C. (1998)DNA annealing by RAD52 protein is stimulated by specificinteraction with the complex of replication protein A andsingle-stranded DNA. Proc Natl Acad Sci USA 95: 6049–6054.
Sulzenbacher, G., Gruez, A., Roig-Zamboni, V., Spinelli, S.,Valencia, C., Pagot, F, et al. (2002) A medium-throughput
crystallization approach. Acta Crystallogr D Biol Crystallogr58: 2109–2115.
Sun, S., and Shamoo, Y. (2003) Biochemical characterizationof interactions between DNA polymerase and single-stranded DNA-binding protein in bacteriophage RB69.J Biol Chem 278: 3876–3881.
Sung, P. (1997) Function of yeast Rad52 protein as a media-tor between replication protein A and the Rad51recombinase. J Biol Chem 272: 28194–28197.
Szczepanska, A.K., Bidnenko, E., Plochocka, D., McGovern,S., Ehrlich, S.D., Bardowski, J, et al. (2007) Adistinct single-stranded DNA-binding protein encoded by the Lactococcuslactis bacteriophage bIL67. Virology 363: 104–112.
Terwilliger, T.C. (2002) Automated structure solution, densitymodification and model building. Acta Crystallogr D BiolCrystallogr 58: 1937–1940.
Theobald, D.L., Mitton-Fry, R.M., and Wuttke, D.S. (2003)Nucleic acid recognition by OB-fold proteins. Annu RevBiophys Biomol Struct 32: 115–133.
Uson, I., and Sheldrick, G.M. (1999) Advances in directmethods for protein crystallography. Curr Opin Struct Biol9: 643–648.
Vagin, A., and Teplyakov, A. (1997) MOLREP: an AutomatedProgram for Molecular Replacement. J Appl Crystallogr 30:1022–1025.
Wang, M., Mahrenholz, A., and Lee, S.H. (2000) RPA stabi-lizes the XPA-damaged DNA complex through protein–protein interaction. Biochemistry 39: 6433–6439.
Wessel, D., and Flügge, U.I. (1984) A method for the quan-titative recovery of protein in dilute solution in the presenceof detergents and lipids. Anal Biochem 138: 141–143.
Winn, M.D., Isupov, M.N., and Murshudov, G.N. (2001) Useof TLS parameters to model anisotropic displacements inmacromolecular refinement. Acta Crystallogr D Biol Crys-tallogr 57: 122–133.
Wold, M.S. (1997) Replication protein A: a heterotrimeric,single-stranded DNA-binding protein required for eukary-otic DNA metabolism. Annu Rev Biochem 66: 61–92.
Supporting information
Additional supporting information may be found in the onlineversion of this article.
Please note: Wiley-Blackwell are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
1170 E. Scaltriti et al. �
© 2009 The AuthorsJournal compilation © 2009 Blackwell Publishing Ltd, Molecular Microbiology, 73, 1156–1170





























